Embed Size (px)
Citation preview

515
Biochimica et Biophysica Acta, 587 (1979) 515--521 © Elsevier/North-Holland Biomedical Press
B B A 2 9 0 6 2
INHIBITION OF UREA SYNTHESIS BY PENT-4-ENOATE ASSOCIATED WITH DECREASE IN N-ACETYL-L-GLUTAMATE CONCENTRATION IN ISOLATED RAT HEPATOCYTES
KAZUMASA AOYAGI *, MASATAKA MORI and MASAMITI TATIBANA **
Department of Biochemistry, Chiba University School of Medicine, 1-8-1 Inohana, Chiba 280 (Japan)
(Received March 6th, 1979)
Key words: Urea synthesis; Pent-4~noate; N-Acetyl-L-glutamate; Hyperammonemia; (Hepa tocy te)
Summary
Pent-4~noate at 0.1 to 1.0 mM strongly inhibited urea synthesis in isolated rat hepatocytes. Pent-4~noate at the same concentrations markedly decreased concentrations of N-acetyl-L-glutamate, an essential activator of carbamoyl- phosphate synthase-I (EC 2.7.2.5), and the decrease was well in parallel with the inhibition of urea synthesis by pent~-enoate. This compound also lowered cellular concentrations of acetyl-CoA, a substrate of acetylglutamate synthase (EC 2.3.1.1). Pent-4~noate in a dose of 1 mM did not significantly affect cel- lular concentrations of ATP, and had no direct effect on acetylglutamate syn- thase activity. These results suggest that the inhibition of urea synthesis by pent4-enoate is due to decrease in N-acetyl-L-glutamate concentration and that the decrease is probably brought about by decreased rate of its synthesis due to the lowered concentration of cellular acetyl-CoA.
Introduction
Pent-4~noate is a chemical analog of hypoglycin, the compound which causes Jamaican vomiting sickness [ 1,2]. Pent-4~anoate inhibits long chain fatty acid oxidation [3,4] and gluconeogenesis [5] and produces profound hypo- glycemia in vivo in laboratory animals. Recently, Glasgow and Chase [6] reported that pent-4~noate produces many of the essential features of Reye's
* P r e s e n t add re s s : I n s t i t u t e o f Cl in ica l M e d i c i n e , The Universi ty o f T s u k u b a , N i iha r l - gun , I b a r a k i - k e n 3 0 0 - 3 I , J a p a n .
** T o w h o m c o r r e s p o n d e n c e s h o u l d b e a d d r e s s e d .

516
syndrome [7] including hyperammonemia. These workers found that pent-4- enoate strongly inhibited urea synthesis in rat liver slices and indicated that inhibition of urea synthesis led to an elevated plasma ammonia in rats injected with the compound [ 8]. However, the mechanism of the inhibition of urea syn- thesis is poorly understood.
We have been studying the regulation of urea biosynthesis and have pro- posed that the hepatic level of acetylglutamate, an allosteric activator of mito- chondrial carbamoyl-phosphate synthase (ammonia) (EC 2.7.2.5), plays a criti- cal role in the regulation of urea synthesis in mammalian liver [9--12]. Our proposal has been supported by recent work from other laboratories [13--15]. In the present paper, we report that pent-4~enoate strongly inhibits urea syn- thesis in isolated rat hepatocytes and that the inhibition is closely associated with decrease in acetylglutamate concentration.
Materials and Methods
Preparation of isolated hepatocytes. Male Wistar rats, weighing 200 to 250 g, were fed a commercial laboratory chow (crude protein content, 24%) ad libi- turn and fasted for 24 h before experiments. Hepatocytes were isolated by a modification [16] of the method of Berry and Friend [17].
Incubation of isolated hepatocytes. Immediately after preparation of hepato- cytes, or after preincubation as described in the tables, the cells were incubated at 37°C with shaking at 90 oscillations/min under O2/CO2 (95:5) in 1.5 or 4.5 ml of Ca2+-free Krebs-Henseleit bicarbonate buffer (pH 7.4 at 37°C) containing 10 mM NH4C1, 10 mM sodium lactate, 10 mM sodium aspartate, 5 mM L-orni- thine and 1.5% (w/v) gelatin in a siliconized glass scintillation vial. Urea plus citrulline synthesis was determined after 10 min of incubation and aceiylglu- tamate, ATP and acetyl-CoA after 5 min of incubation.
Determinations. Urea plus citrulline was determined colorimetrically [18] on 0.5 M perchloric acid extracts of reaction mixtures, based on the fact that citrulline and urea give practically the same color intensities. Urea was deter- mined colorimetrically [19]. Acetylglutamate was determined on 10% (w/v) trichloroacetic acid extract of the cells as described previously [9,12] except for the following modifications; the recovery of acetylglutamate during its purification was corrected, this correction being based on the recovery of [3H]- acetylglutamate which was added at the step of extraction and carbamoyl- phosphate synthase (ammonia) assay was performed radiochemically using ['4C]bicarbonate as a substrate. ATP was determined enzymatically [20] as modified [ 21] on neutralized perchloric acid extracts. For the determination of acetyl-CoA, cells were rapidly separated from the medium by the method of Hems et al. [22]. Acetyl-CoA was determined enzymatically [23] on neu- tralized extracts. Protein was determined by the biuret method.
Materials. Pent-4-enoic acid and pent-2enoic acid were obtained from the Tokyo Chemical Industry Co., Tokyo. Acetylglutamate was obtained from Sigma, and gelatin, was from Difco Laboratories. Acetyl[3H]glutamate (313 Ci/mol) was synthesized from [3H]glutamate and acetic anhydride [24]. Rat liver carbamoyl-phosphate synthase (ammonia) and bovine liver ornithine car- bamoyltransferase (EC 2.1.3.3) were purified as described [25,26].

517
Results and Discussion
Urea and glucose synthesis in isolated rat hepatocytes Ureogenesis from ammonia in isolated hepatocytes was determined in the
presence of 5 mM ornithine, 10 mM aspartate and 10 mM lactate; lactate is required for maximal urea product ion [ 27 ] and aspartate was added to ensure that its supply was sufficient for urea synthesis. (In the course of our studies, aspartate was found not to be required for maximal urea synthesis by isolated hepatocytes .) Urea plus citruUine synthesis proceeded linearly with time up to 60 min at a rate of 0.75 pmol/h per mg protein. This value is similar to or somewhat lower than values reported by other groups [ 27,28]. Citrulline accu- mulated significantly in 20 min of incubation (38% of urea plus citrulline) and the absolute amount remained almost unchanged with time, resulting in a smaller proport ion in 60 min of incubation (15% of urea plus citrulline). Addi- tion of 1.3 mM Ca 2÷ to the incubation mixture had no appreciable effect on ureogenesis under the condit ions used. The hepatocytes actively synthesized glucose from lactate in the presence of 1.5 mM NH4C1 at a rate of 0.15 ~mol/h per mg protein, when assayed according to the method of Zahlten et al. [29]. The glucose synthetic rate was enhanced 1.8-fold by 10 ~M glucagon (data not shown). Thus, the isolated hepatocytes used here had a high cellular integrity of liver parenchymal cells.
Effect o f pent-4-enoate on urea synthesis and acetylglutamate level in isolated hepatocytes
Urea plus citrulline synthesis in isolated hepatocytes was strongly inhibited by pent-4-enoate at concentrat ions of 1 • 10 -4 to 1 • 10 -3 M (Table I). The ratio of urea to citrulline was essentially the same, regardless of whether or not urea synthesis was inhibited by pent-4-enoate, indicating that either one or both of the initial steps leading to citrulline synthesis is subject to inhibition by pent-4- enoate. Pent-4-enoate at 1 mM did not directly inhibit the activities of car- bamoyl-phosphate synthase (ammonia) and ornithine carbamoyltransferase par- tially purified from rat liver. Pent-2-enoate, which has no hypoglycemic action [30], did not affect urea synthesis at a dose of 0.4 mM.
Acetylglutamate concentrat ion which was measured at the midpoint of urea synthesis assay decreased markedly with addition of pent-4-enoate and the rate of urea synthesis was in good parallel with levels of acetylglutamate (Table I). A similar result was reported in a previous paper [12] and the relationship between acetylglutamate levels and ureogenesis was shown. Preincubation of hepatocytes with pen t -4~noate for more than 20 min was required to deplete cellular acetylglutamate and to produce maximal inhibition of the urea syn- thesis (data not shown).
The concentrat ion of acetylglutamate in isolated hepatocytes ( 0 . 0 3 - 0.43 nmol/mg cell protein) corresponds to 4 -- 60 nmol/g wet wt. of hepatocytes and is in the same range as that observed in intact mouse liver [9,12] and rat liver [14]. Since acetylglutamate is localized in the mitochondria [9] , the mito- chondrial acetylglutamate concentrat ion is calculated to be 0.02--0.30 mM, assuming that the mitochondrial space occupies about 20% of the cellular volume. These values appear to be adequate for the function of acetylglutamate
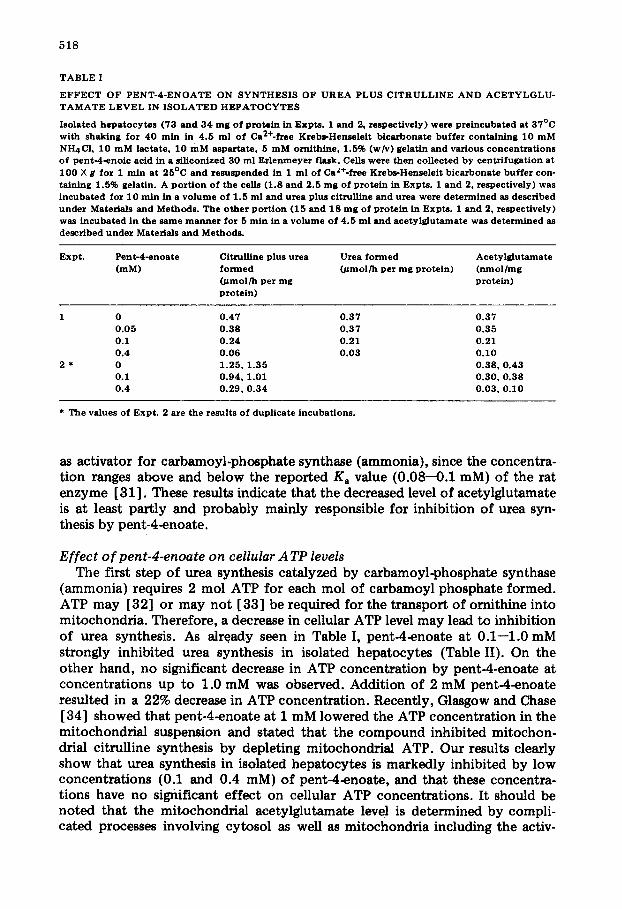
518
T A B L E I
E F F E C T O F P E N T - 4 - E N O A T E O N S Y N T H E S I S O F U R E A P L U S C I T R U L L I N E A N D A C E T Y L G L U - T A M A T E L E V E L IN I S O L A T E D H E P A T O C Y T E S
I so la ted h e p a t o c y t e s ( 73 and 3 4 m g o f p r o te in in E x p t s . 1 a n d 2, r e s p e c t i v e l y ) w e r e pre ineubated at 3 7 ° C w i t h shaking for 4 0 rain in 4 . 5 m l o f Ca2+-free Kreb~-Hensele i t b i ca rbo na te buf fer c o n t a i n i n g 1 0 m M NH4CI , 1 0 m M l a c t a t e , 1 0 m M a s p a r t a t e , 5 m M o r n i t h i n e , 1 ,5% (w/v) gelat in and various c o n c e n t r a t i o n s o f p e n t - 4 - e n o i c acid in a s i l i eon ized 3 0 m l E r l e n m e y e r f lask . Cells w e r e then c o l l e c t e d by cen tr i fuga t ion at 1 0 0 X g f o r 1 m i n at 2 5 ° C and r e s u s P e n d e d in 1 m l o f CaZ*-frec Krebs-Hensc le i t b i ca rbo na te buf fer con- taining 1 .5% gelat in . A p o r t i o n o f the cel ls ( 1 . 8 and 2 .5 m g of prote in in E x p t s . 1 a n d 2, re spec t iv e ly ) was i n c u b a t e d for 1 0 r a i n in a v o l u m e o f 1 .5 m l and urea plus c i trul l ine and urea w e r e d e t e r m i n e d as descr ibed under Materials and Methods . Th e o t h e r p o r t i o n ( 1 5 and 1 8 m g o f prote in in E x p t s . 1 a n d 2, re spec t iv e ly ) was i n c u b a t e d in the s a m e m a n n e r for 5 m l n in a v o l u m e o f 4 . 5 m l and a c e t y l g l u t a m a t e wa s d e t e r m i n e d as descr ibed under Materials and M e t h o d s .
Expt . P e n t - 4 - e n o a t e Citrul l ine plus urea Urea f o r m e d A c e t y l g l u t a m a t e ( m M ) f o r m e d 0 ~ m o l / h per m g pro te in ) ( n m o l / m g
( # m o l / h p e r m g prote in ) p r o t e i n )
1 0 0 . 4 7 0 . 3 7 0 . 3 7 0 . 0 5 0 . 3 8 0 . 3 7 0 . 3 5 0 .1 0 . 2 4 0 . 2 1 0 .21 0 .4 0 . 0 6 0 . 0 3 0 . 1 0
2 * 0 1 . 2 5 , 1 . 3 5 0 . 3 8 , 0 . 4 3 0 .1 0 . 9 4 , 1 .01 0 . 3 0 , 0 . 3 8 0 .4 0 . 2 9 , 0 . 3 4 0 . 0 3 , 0 . 1 0
* T h e va lues o f E x p t . 2 are the resul ts o f dupl ica te incubat ions .
as activator for carbamoyl-phosphate synthase (ammonia), since the concentra- tion ranges above and below the reported Ka value (0.08--0.1 mM) of the rat enzyme [31]. These results indicate that the decreased level of acetylglutamate is at least partly and probably mainly responsible for inhibition of urea syn- thesis by pent-4-enoate.
Effect o f pent-4-enoate on cellular A TP levels The first step of urea synthesis catalyzed by carbamoyl-phosphate synthase
(ammonia) requires 2 mol ATP for each mol of carbamoyl phosphate formed. ATP may [32] or may not [33] be required for the transport of ornithine into mitochondria. Therefore, a decrease in cellular ATP level may lead to inhibition of urea synthesis. As already seen in Table I, pent-4~enoate at 0.1--1.0 mM strongly inhibited urea synthesis in isolated hepatocytes (Table II). On the other hand, no significant decrease in ATP concentration by pent-4-enoate at concentrations up to 1.0 mM was observed. Addition of 2 mM pent~l~enoate resulted in a 22% decrease in ATP concentration. Recently, Glasgow and Chase [34] showed that pent-4~enoate at 1 mM lowered the ATP concentration in the mitochondrial suspension and stated that the compound inhibited mitochon- drial citruUine synthesis by depleting mitochondrial ATP. Our results clearly show that urea synthesis in isolated hepatocytes is markedly inhibited by low concentrations (0.1 and 0.4 mM) of pent-4~enoate, and that these concentra- tions have no significant effect on cellular ATP concentrations. It should be noted that the mitochondrial acetylglutamate level is determined by compli- cated processes involving cytosol as well as mitochondria including the activ-

519
T A B L E II
E F F E C T O F P E N T - 4 - E N O A T E O N U R E A P L U S C I T R U L L I N E S Y N T H E S I S A N D A T P L E V E L IN I S O L A T E D H E P A T O C Y T E S
I s o l a t e d h e p a t o c y t e s ( 3 4 a n d 2 2 m g o f p r o t e i n in E x p t s . 1 a n d 2, r e s p e c t i v e l y ) we re p r e i n c u b a t e d f o r 4 0 m i n a n d cel ls w e r e c o l l e c t e d as d e s c r i b e d in T a b l e I. A p o r t i o n o f t h e cel ls ( 2 .5 a n d 7 .2 m g o f p r o t e i n in ExiSts. 1 a n d 2, r e s p e c t i v e l y ) was i n c u b a t e d f o r 1 0 r a i n as d e s c r i b e d u n d e r Ma te r i a l s a n d M e t h o d s f o r d e t e r m i n a t i o n o f u r e a p lu s c i t ru l l ine . T h e o t h e r p o r t i o n (5 .6 a n d 1 4 rng o f p r o t e i n in E x p t s . 1 a n d 2, r e s p e c t i v e l y ) w a s i n c u b a t e d in t h e s a m e m a n n e r f o r 5 r a i n in a v o l u m e o f 4 . 5 m l a n d A T P was d e t e r m i n e d as d e s c r i b e d u n d e r Ma te r i a l s a n d M e t h o d s .
E x p t . P e n t - 4 - e n o a t e Urea p lu s c i t r u l i i ne A T P level (mM) f o r m e d ( n m o l / m g p r o t e i n )
0 ~ m o l / h p e r m g p r o t e i n )
1 * 0 1 . 2 5 , 1 . 3 5 8 .9 , 9 .4 0 . I 0 . 9 4 , 1 .01 8 .3 , 8 . 5 0 . 4 0 . 2 9 , 0 . 3 4 8 .3 , 8 .3 1 .0 0 . 1 6 , 0 .31 8 .5 , 9 .2
2 0 1 . 6 0 4 .6 0 .4 0 . 3 8 4 .5 2 .0 0 . 2 9 3 .6
* The va lues o f E x p t . 1 a re t he r e su l t s o f d u p l i c a t e i n c u b a t i o n s .
ity of acetylglutamate synthase (EC 2.3.1.1), supply of the enzyme substrates and transport of acetylglutamate from mitochondria to cytosol where acetyl- glutamate is assumably degraded. Therefore, the effect of pent-4-enoate on the acetylglutamate level cannot be validly analyzed using isolated mitochondria. Actually, we found that pent-4-enoate (1 raM) had little effect on acetylglu- tamate concentration in isolated mitochondria {data not shown).
Effect of pent-4-enoate on cellular acetyl-CoA level Pent-4~noate is a potent inhibitor of fatty acid oxidation and is known to
decrease acetyl-CoA concentration in perfused rat liver [5,35]. Since acetyl- CoA is a substrate for acetylglutamate synthase reaction [9] and since cellular acetylglutamate concentration is determined mainly by the rate of its synthesis [12], it seems likely that depletion of cel lul~ acetylglutamate by pent-4~noate is due to decreased concentration of acetyl-CoA. In fact, pent-4~noate at con- centrations of 0.1 and 0.4 mM decreased acetyl-CoA concentration in isolated hepatocytes by 59 and 72%, respectively, and the decrease was closely asso- ciated with inhibition of urea synthesis (Expt. 1, Table III) and with a decrease in acetylglutamate concentration (Expt. 2, Table III). The acetyl-CoA concen- trations observed in isolated hepatocytes (0.03 - -0 .29 nmol /mg protein) cor- respond to 4 -- 41 nmol/g wet wt. of hepatocytes. Assuming that 56% of cellu- lar acetyl-CoA is located in mitochondria [36], the concentration in mito- chondria is calculated to be 11 - - 115 pM; these values are still lower than the apparent K m of rat liver acetylglutamate synthase for acetyl-CoA (0.7 raM) [11]. Thus, the rate of acetylglutamate synthesis in vivo may be approximately proportional to cellular acetyl-CoA concentration. Pent-4-enoate at 1 mM did not inhibit acetylglutamate synthase partially purified from rat liver.
The results presented here indicate that pent-4~noate decreases cellular ace- tylglutamate concentration by lowering cellular acetyl-CoA concentration,

5 2 0
T A B L E III
E F F E C T O F P E N T - 4 - E N O A T E O N U R E A P L U S C I T R U L L I N E S Y N T H E S I S A N D L E V E L S O F ACE- T Y L G L U T A M A T E A N D A C E T Y L - C o A
I s o l a t e d h e p a t o c y t e s ( 1 4 4 a n d 1 3 3 r ag of prote in in E x p t s . 1 a n d 2, r e spec t i ve ly ) we re p r e i n c u b a t e d for 4 0 m i n a n d cel ls w e r e c o l l e c t e d as d e s c r i b e d in T a b l e I. In E x p t s . 1, a port ion o f the cells (1 .8 m g o f p ro - t e in ) was incubated for 10 rain as d e s c r i b e d u n d e r Mate r i a l s a n d M e t h o d s a n d u r e a p lu s c i t ru l l i ne was d e t e r m i n e d . In E x p t s . 2, ce i ls ( 1 9 m g o f prote in) were incubated for 5 min a n d a c e t y l g l u t a m a t e was de te r - m i n e d . The other por t ion (57 a n d 3 3 m g of prote in in Exp t s . 1 a n d 2, r e s p e c t i v e l y ) w a s incubated in the s a m e m a n n e r f o r 5 r a in a n d a c e t y l - C o A was d e t e r m i n e d as d e s c r i b e d u n d e r Mate r ia l s a n d M e t h o d s .
E x p t . P e n t - 4 - e n o a t e Urea p lus c i t r u l l i ne A c e t y l g l u t a m a t e level A c e t y l C o A level ( m M ) f o r m e d ( n m o l / m g p r o t e i n ) ( n m o l / r a g
( /~mol/h p e r m g protein) prote in)
1 0 0 . 5 8 0 . 2 0 0 . 0 5 0 . 5 8 0 . 1 5 0 .1 0 . 5 4 0 . 1 4 0 .4 0 . 3 7 0 . 0 8
2 * 0 1 . 0 4 , 1 .24 0 . 2 7 , 0 . 2 9 0 .1 0 . 9 7 , 1 .14 0 . 1 1 , 0 . 1 2 0 ,4 0 . 2 0 , 0 . 2 4 0 . 0 3 , 0 . 0 5
* T h e va lues o f E x p t . 2 a re the results o f dupl icate incubat ions .
which in turn reduces carbamoyl-phosphate synthase (ammonia) activity and inhibits urea synthesis. On the other hand, effects of pent-4-enoate on the ornithine transport, a possible limiting step in the urea synthesis [32,33], were not studied. Sinatra et al. [37] reported that incubation of rat liver homoge- nates with 1 mM pent-4-enoate for 15 rain resulted in about 50% inhibition of carbamoyl-phosphate synthase (ammonia), increased the apparent Km of orni- thine carbamoyltransferase for carbamoyl phosphate and decreased V of the carbamoyltransferase. The relationship between their findings and our results concerning the inhibition of ureogenesis is the subject of ongoing studies in our laboratory.
Acknowledgements
We are grateful to Dr. H. Ishida of our laboratory for conducting part of this work and to M. Ohara, Kyoto University, for assistance with preparation of the manuscript. This work was supported in part by the Scientific Research Fund (B 348119) from the Ministry of Education, Science and Culture of Japan.
References
1 S h e r r a t t , H . S . A . ( 1 9 6 9 ) Br. M e d . Bul l . 2 5 , 2 5 0 - - 2 5 5 2 Bress ler , R . , C o r z e d o r , C. a n d B r e n d e l , K . ( 1 9 6 9 ) P h a r m a e o l . Rev . 2 1 , 1 0 5 - - 1 3 0 3 C o r r e d o r , C. , B r e n d e l , K . a n d B r e u l e r , R . ( 1 9 6 7 ) P r o c . Na t l . A c a d . Sci . U .S . 58° 2 2 9 9 - - 2 3 0 6 4 H o l l a n d ° P C. a n d S h e r r a t t , H . S . A . ( 1 9 7 3 ) B i o c h e m . J . 1 3 6 , 1 5 7 - - 1 7 1 5 T o e w s , C . J . , L o w y , C. a n d R u d e r m a n , N.B. ( 1 9 7 0 ) J . Biol . C h e m . 2 4 5 , 8 1 8 - - 8 2 4 6 G l a s g o w , A.M. a n d C h a s e , H .P . ( 1 9 7 5 ) Ped i a t . Res . 9 , 1 3 3 - - 1 3 8 7 R e y e , R . D . K . , M o r g a n , G. a n d Bazal , J . ( 1 9 6 3 ) L a n c e t 2 , 7 4 9 - - 7 5 2 8 G l a s g o w , A .M. a n d C h a s e , H .P . ( 1 9 7 5 ) B i o e h e m . B i o p h y s . Res . C o m m u n . 6 2 , 3 6 2 - - 3 6 6 9 S h i g e s a d a , K . a n d T a t i b a n a , M. ( 1 9 7 1 ) J . B io l , C h e m . 2 4 6 , 5 5 8 8 - - 5 5 9 5

521
I0 Shigasada, K. and Tatibana, M. (1972) Biochem. Biophys. Res. Commun. 46, 491--497 11 Shigesada, K, and Tatibana, M. (1978) Eur. J. Biochem. 84,285--291 12 Sh/gesada, K., Aoyagi, K. and Tatibana, M. (1978) Eur. J. Biochem. 85, 385--391 13 McGivan, J,D., Bradford, N.M. and Mendes-Mour~'o, J. (1976) Bloc'hem. J. 154,415--421 14 Saheki, T., Katsunuma, T. and Sase, M. (1977) J. Biochem. (Tokyo) 82,551--558 15 Meijer, A.J. and Van Woerkom, G.M. (1978) FEBS Lett. 86,117--121 16 Zahlten, R.N. and Stratman, F.W. (1974) Arch. Biochem. Biophys. 163, 600--608 17 Berry, M.N. and Friend, D.S. (1969) J. Cell Biol. 43, 506--520 18 Prescott, L.M. and Jones, M.E. (1969) Anal. Biochem. 32,408--419 19 Archibald, R.M. (1945) J. Biol. Chem. 157, 507--518 20 Lamprecht, W, and Trautschold (1974) in Methods of Enzymatic Analysis (Bergmeyer, H.U. and
Gawehn, K., eds.), Vol. 4, pp, 2101--2110, Academic Press, New York 21 Lund, P., CorneIl, N.W. and Krebs, H.A. (1975) Biochem. J. 152,593--599 22 Hems, R., Lund, P. and Krebs, H.A. (1975) Biochem. J. 150, 47--50 23 Decker, K. (] 974) in Methods of Enzymatic Analys/s (Bergmeyer, H.U. and Gawehn, K., eds.), Vol. 4,
pp. 1988--1993, Academic Press, New York 24 Marshall, M., Metzenberg, R.L. and Cohen, P.P. (1961) J. Biol. Chem. 236, 2229--2237 25 Guth6hrlein, G. and Knappe, J. (1968) Eur. J. Biochem. 7,119--127 26 Marshall, M. and Cohen, P.P. (1972) J. Biol. Chem. 247, 1641--1653 27 Briggs, S. and Freedland, R.A. (1976) Biochem. J. 160,205--209 28 Krebs, H.A., Lund, P. and Stubbs, M. (1976) in Gluconeogenesis (Hanson, R.W. and Mehlman, M.A.,
eds.), pp. 269--291, John Wiley and Sons, New York 29 Zahlten, R.N., Kneer, N.M., Stratman, F.W. and Lardy, H.A. (1974) Arch. Biochem. Biophys. 161,
528--535 30 Senior, A.E. and Sherratt, H.S.A. (1967) Biochem. J. 104, 56P 31 Lusty, CJ . (1978) J. Biol. Chem. 253, 4270--4278 32 Gamble, J.G. and Lehninger, A.L. (1973) J. Biol. Chem. 248,610--618 33 Bryta, J. and Harris, E J . (1976) FEBS Lett. 72,331---336 34 Glasgow, A.M, and Chase, H.P. (1976) Biochem. J, 156,301--307 35 Williamson, J.R., Fukami, M.H., Peterson, M.J., Rostand, S.G. and Scholz, R. (1969) Biochem. Bio-
phys. Res, Commun. 36,407--413 36 Siess, E.A., Brocks, D.G., Lattke, H.K, and Wieland, O.H, (1977) Blochem. J. 166, 225--235 37 Sinatra, F., Yoshida, T., Applebaum, M., Mason, W., Hoogenraad, N.J. and Sunshine, P. (1975) Pediat.
Res. 9. 829--833





























