Embed Size (px)
Citation preview

0 1992 MUNKSGAARD
Inhibitory influence of late afternoon melatonin injections and the counter-inhibitory action of melatonin-containing pellets on thyroid growth process in male Wistar rats: Comparison with effects of other indole substances
Wajs E, Lewinski A. Inhibitory influence of late afternoon melatonin injections and the counter-inhibitory action of melatonin-containing pellets on thyroid growth processes in male Wistar rats: Comparison with effects of other indole substances. J . Pineal Res. 1992:13:158-166.
Abstract: Previous studies performed in our laboratory indicated that melatonin, when adminstered in late afternoon (1600-1800) as S.C.
injections for 10 days, exerted an inhibitory effect on thyroid growth in mice and Sprague-Dawley rats. The goal of the present study was to examine the effects of melatonin and other indole substances (5-methoxytryptophol, N-acetylserotonin, 6-methoxytryptamine), administered for 10 days, for either 4, 8, or 10 weeks, on thyroid growth processes in adult male Wistar rats. We have also compared the action of melatonin administered in late afternoon as S.C. injections with effects of the melatonin-released chronically from S.C. implanted beeswax pellets containing I mg of the hormone. Additionally, the effects of melatonin injections in animals with the pineal gland removed and in thyroid stimulating hormone (TSH)-injected rats have been studied. We examined the mean mitotic activity rates (MMARs) of thyroid follicular cells (TFC) and the wet thyroid weights. We concluded that melatonin, of the examined substances, is the most potent inhibitor of thyroid growth; the hormone reduced the MMAR of TFC and the thyroid weight at all time-points. 5-Methoxytryptophol, like melatonin, exerted the inhibitory influence on the mitotic activity; however, it did not affect thyroid weight. The influence of other indole substances was much less pronounced. It was shown that melatonin- pellets prevented the inhibitory effect of late afternoon melatonin injections on thyroid growth processes. This observation is consistent with results of other authors concerning the counter-antithyroid effect of melatonin on thyroid hormone secretion. Pinealectomy revealed the stimulatory effect on thyroid growth processes, while melatonin treatment reversed the effect of the surgery. Melatonin suppressed the
I stimulatory effect of TSH on the thyroid growth.
Ewa Wajs and Andrzej Lewinski Laboratory of Thyrology, Institute of Endocrinol- ogy, The University School of Medicine at-C6dz, t o d i , Poland
Key words: melatonin-N-acetyiserotonin- 5-methoxytryptophol-the rat-mitotic activity-thyroid follicular cells-thyroid weight
Address reprint requests to Ewa Wajs, Laboratoly of Thyrology, Institute of Endocrinology, The University School of Medicine at k o d i , Xodi, Poland
Received March 10, 1992; accepted September 1, 1992.
Introduction
Melatonin, the main pineal hormone, exerts a well-documented inhibitory effect on thyroid secre- tory processes; late afternoon subcutaneous (s.c.) injections of melatonin into Syrian hamsters de- creased the serum thyroxine (T,) concentration and free thyroxine index (FT,I) in both male [Vriend et al., 1977; Vriend and Reiter, 19771 and female [Vriend et al., 19821 animals. Pinealectomy (PX)
158
prevented the depression of plasma T, and FTJ obtained in blinded hamsters [Vriend et al., 19771. On the other hand, chronic release of melatonin from S.C. implants [Vaughan et al., 1984al or the administration of the pineal hormone in drinking water [Vriend and Gibbs, 19841 were not effective in reducing T, levels and even prevented constant darkness or blinding from inhibiting the thyroid secretion, thus producing the so called counter- antithyroid effect. In experiments performed on rats

Melatonin and other indoles affect thyroid
in our laboratory, we found that administration of melatonin released from S.C. implanted pellets in- creased thyroid hormone concentrations after 10 days and 10 weeks. This effect may be called the “prothyroid” action of melatonin. On the other hand, the joint effect of late afternoon melatonin injections and melatonin implants brought about no changes in thyroid hormone concentrations [Krotewicz et al., in press].
Thyroid growth processes, as well as thyroid secretion, are under the inhibitory influence exerted by the pineal gland via melatonin. Early experi- ments showed that elimination of the main source of that hormone by PX resulted in an increase of thyroid weight in rats [Miline, 1963; Pazo et al., 1968; DeFronzo and Roth, 19721, mice [Houssay et al., 19661, and cats [Davis and Martin, 19401. Previous studies from our laboratory demonstrated that the administration of melatonin for 10 days at doses of 10 p,g/daily and 100 pg/daily, reduced the mean mitotic activity rate (MMAR) of murine thyrocytes [Lewinski and Sewerynek, 19861; at a daily dose of 25 pg, melatonin decreased the MMAR of thyroid follicular cells (TFC) in Sprague- Dawley rats [Sewerynek et al., 19881. We also succeeded in showing that PX resulted in the increase of TFC mitotic activity in female Wistar rats and melatonin treatment partially reversed the effect of the surgery [Wajs et al., 19891.
Thus far, we have not studied the long-term effects of melatonin on the thyroid growth. The goal of the present experiment was to determine whether melatonin alters the MMAR of TFC and the thyroid weight after 10 days, and after either 4, 8, or 10 weeks of administration. We have also examined the action of other indole compounds; two of them existing in the mammalian pineal gland [N-acetyl- serotonin (NAc-5HT) and 5-methoxytryptophol (5- MTF)] and one synthetic [6-methoxytryptamine (6-MT)I. The thyroid growth response was also investigated after PX and after melatonin treatment in PX animals. Additionally, we compared the effect of melatonin administered in late afternoon as S.C. injections and as S.C. implanted pellets (con- tinuous availability of the hormone in question). Finally, we have studied the influence of melatonin on thyroid stimulating hormone (TSH)-stimulated thyroid growth.
Materials and methods
Animals
The study was performed on 3 17 adult male Wistar rats, weighing 260 2 20 g each. The animals were maintained in a room with controlled illumination (14:lO 1ight:dark cycle, lights on at 0600, for 3
weeks before experiment) and temperature (22 3t 2”C), and had constant access to granulated rat LSM food and to tap water.
Experiment I
This study examined the influence of indole com- pounds on the thyroid weight and mitotic activity of thyroid follicular cells (TFC).
All substances were administered in the late afternoon (between 1600 and 1800) as S.C. injec- tions once daily (0.2 ml volume). Fresh solutions of indole compounds were prepared ex tempore; in- doles were dissolved in absolute alcohol and diluted with 0.9% NaCl (l:lO, V:V).
Rats were divided into 24 groups. Group 0 served as the control group; the animals of that group (n = 20) were killed at the beginning of the exper- iment prior to injections. Groups I-VI were divided into four subgroups (a, b, c, d), depending on the duration of treatment, as follows: a, 10 days; b, 4 weeks; c, 8 weeks; d, 10 weeks; n is the number of animals in each subgroup. Group I received injec- tions of solvent-diluent for indoles specified above as follows: n = 9 (a), n = 9 (b), n = 8 (c), n = 9 (d). group I1 received injections of melatonin (25 p,g/daily; n = 10 [a], n = 10 [b], n = 8 [c], n = 10 [d]; group I11 was treated with melatonin (50 p,g/daily; n = 9 [a], n = 10 [b], n = 7 [c], n = 8 [d]); group IV animals were injected with 5-methoxytryptophol (25 Fgldaily; n = 10 [a], n = 10 [b], n = 8 [c], n = 10 [d]; group V was administered 6-methoxytryptamine (25 p,g/daily; n = 10 [a], n = 10 [b], n = 9 [c], n = 10 [d]); and the rats of group VI were injected with N-acetyl- serotonin (25 pg/daily; n = 10 [a], n = 10 [b], n = 10 [c], n = 11 [d]). All indoles were purchased from Sigma Chemical Co., St. Louis, Mo.
Stathmokinetic method. In order to evaluate thyroid mitotic activity, the metaphase-arrest technique was employed. On the day after the last injection series, 2 hr prior to the thyroid collection, the animals were injected with colchicine (Fluka AG, 1 mg/kg BW, intraperitoneally - i.p.). All the rats were sacrificed by decapitation. Thyroid lobes, immediately after collecting, were weighed on a torsion balance and then fixed in Bouin’s fluid. Paraffin sections (6 km thick) were stained with haematoxylin and eosin.
In the microscopic preparations, the mitotic ac- tivity rates (number of metaphases per 4,000 sub- sequently scored TFC, 2,000 cells from each lobe, in the randomly selected sections of thyroid lobes) were evaluated. Relative wet thyroid weights were calculated per 100 g of animal body weight.
159

Wajs and Lewinski
Experiment II
This experiment examined the influence of melato- nin, administered in late afternoon as S.C. injections and/or released from S.C. implanted pellets, on certain indices of thyroid growth.
The animals were divided into 12 groups; in six groups the experiment was continued for 10 days (subgroup a) and in six corresponding groups the study was continued for 10 weeks (subgroup b). Numbers of animals (n), examined in particular groups at these two time-points, are given in parentheses, The rats of group I (controls) received injections of solvent-diluent for melatonin (n = 9 [a], n = 9 [b]). The animals of group I1 were administered S.C. injections of melatonin (25 pg/ daily) in late afternoon (1600-1800) (n = 10 [a], n = 10 [b]). Group I11 received also late afternoon S.C. injections of melatonin (50 pg/daily), (n = 9 [a], n = 8 [b]). The animals of group IV received S.C. implanted pellets containing 25 mg of beeswax (Pel C) and served as controls (n = 9 [a], n = 11 [b]). Melatonin-containing beeswax pellets (Pel Mel) were implanted under the skin of the back in animals of groups V (n = 10 [a], n = 7 [b]) and VI (n = 7 [a], n = 10 [b]); the latter group also re- ceived late afternoon melatonin injections (25 pg/ daily). The procedure of pellet implantation was performed once in groups in which the experiment lasted 10 days (a), and was repeated every 2 weeks for rats used in the 10-week study (b). Each melatonin pellet consisted of 24 mg of beeswax and of 1 mg of melatonin.
The subsequent procedures were identical to those in Experiment 1.
Experiment I I I
The study examined the influence of pinealectomy and melatonin treatment on thyroid weight and MMAR of TFC.
The animals were divided into eight groups, four groups in which the experiment lasted 10 days (a), and four groups in which the study was continued for 10 weeks (b). Group I served as controls and received injections of solvent-diluent for melatonin (n = 9 [a], n = 9 [b]). The animals of group I1 received late afternoon melatonin injections (25 pg/daily; n = 10 [a], n = 10 [b]. In groups 111 (n = 10 [a], n = 10 [b]) and IV (n = 8 [a], n = 12 [b]) pinealectomy (PX) was performed according to Kuszak and Rodin’s method [ 19771, the latter group additionally receiving late afternoon melatonin in- jections (25 pg/daily) beginning the day after PX. On the day after PX, injections of melatonin or its solvent-diluent begin in nonoperated animals
160
(groups I, 11). For the surgery, the animals were anesthetized with hexobarbital (Germed, 100 mg/kg BW, i.p.). Completeness of PX was checked during autopsy, at the termination of the experiment.
The remaining part of the experiment was iden- tical to that of Experiment I.
Experiment IV
This study tested the influence of melatonin, TSH, and their joint administration on the thyroid weight and the mitotic activity of TFC.
The animals were divided into four groups of eight rats each; the late afternoon S.C. injections of melatonin, TSH (Thyreostymulin, Organon, 2 IU, i.p.) or its solvent (0.9% NaCl), were continued for 10 days, according to the following scheme: group I - controls, 0.9% NaCI; group I1 - TSH; group 111 - TSH together with melatonin, 25 pg/daily; group IV - TSH with melatonin, 50 pg/daily.
Statistical analysis. The data from all the experi- ments were analyzed using a one-way analysis of variance (ANOVA). The statistical significance of differences among particular groups was subse- quently determined with the use of Newman-Keuls’ test [Hinkle et a]., 19791.
Results
The values of the MMAR of TFC and of the wet thyroid weight did not differ significantly in animals without treatment (group 0), in solvent-injected rats, and in animals implanted with beeswax pellets, i.e., in all control groups used in the study.
Experiment I
Melatonin, in both doses (25 pg/daily and 50 pgidaily), reduced the MMAR of TFC (Fig. 1 ) when it was injected for either 10 days or 4, 8, or 10 weeks. Exceptionally, after the administration of melatonin (25 pg/daily) for weeks, the effect did not attain statistical significance.
The thyroid mitotic activity after treatment with other indole compounds is shown in Figure 2. 5-Methoxytryptophol, like melatonin, exerted an inhibitory influence at all time-points. 6-Methoxy- tryptamine decreased significantly the MMAR of TFC after 4 and 10 weeks of injections, while after 10 days and 8 weeks the indole was ineffective. The inhibitory effect of NAc-5HT occurred after 10 weeks of treatment while shorter periods of injec- tions did not alter the MMAR.
At all time-points, melatonin injections caused a slight reduction of the thyroid weight; the decrease
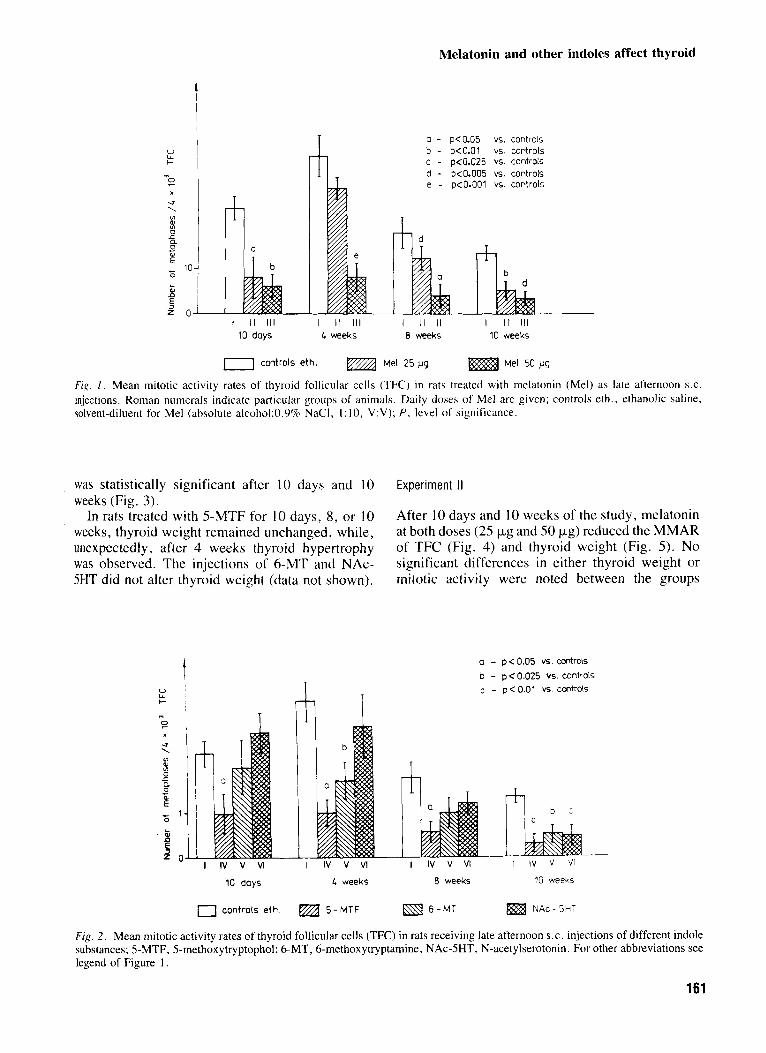
Melatonin and other indoles affect thyroid
C
I I I I l l 10 days
a - pc0.05 vs. controls b - ~(0.01 vs. controls c - p<0.025 vs. controls d - p<0.005 vs. controls
I I I Ill I I I I l l I I I I I I L weeks 8 weeks 10 weeks
0 controls eth. Me1 25 pg Me1 50 pg
Fig. 1 . Mean mitotic activity rates of thyroid follicular cells (TFC) in rats treated with melatonin (Mel) as late afternoon S.C.
injections. Roman numerals indicate particular groups o f animals. Daily doses of Me1 are given; controls eth., ethanolic saline, solvent-diluent for Me1 (absolute alcohol:0.9% NaCI, I : 10, V:V); P , level of significance.
was statistically significant after 10 days and 10 weeks (Fig. 3).
In rats treated with 5-MTF for 10 days, 8, or 10 weeks, thyroid weight remained unchanged, while, unexpectedly, after 4 weeks thyroid hypertrophy was observed. The injections of 6-MT and NAc- 5HT did not alter thyroid weight (data not shown).
I U U t-
10 days
Experiment I I
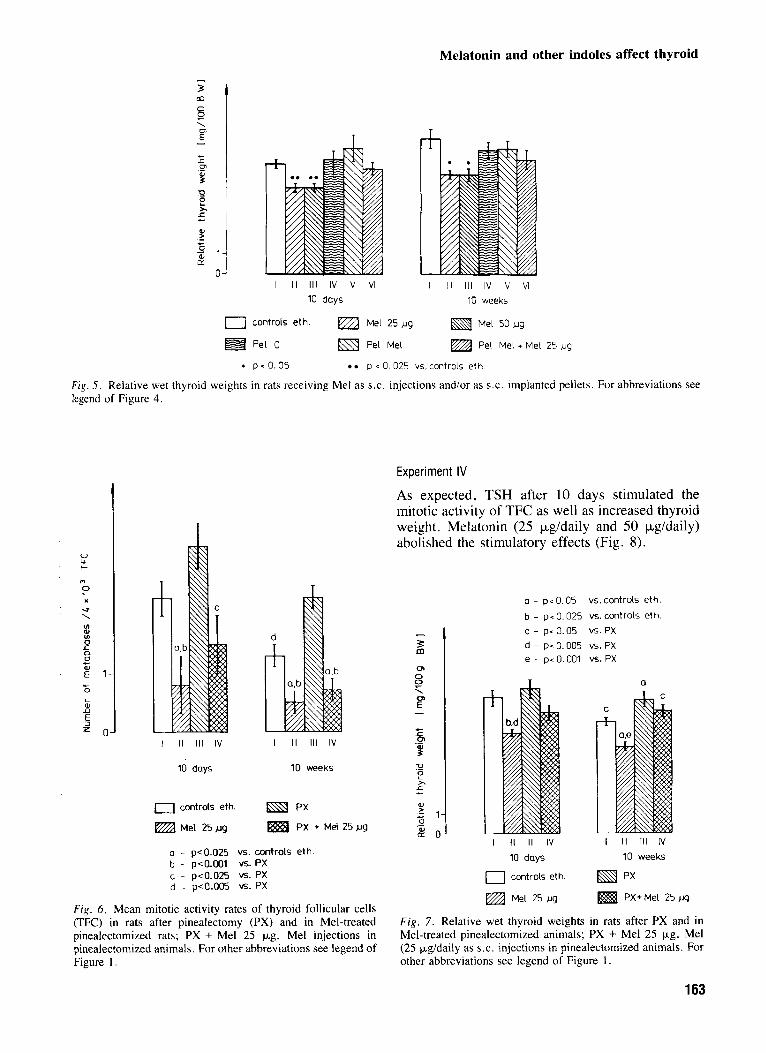
After 10 days and 10 weeks of the study, melatonin at both doses (25 pg and 50 pg) reduced the MMAR of TFC (Fig. 4) and thyroid weight (Fig. 5 ) . No significant differences in either thyroid weight or mitotic activity were noted between the groups
a - p < 0.05 vs. controls b - p<O.O25 vs. controls c - pc 0.01 vs. controls
I IV v VI I IV v VI I IV v V I
8 weeks 10 weeks 1 weeks
0 controls eth 5 - MTF 6 - M T NAG - 5 H T
Fig. 2 . Mean mitotic activity rates of thyroid follicular cells (TFC) in rats receiving late afternoon S.C. injections of different indole substances; 5-MTF, 5-methoxytryptophol; 6-MT, 6-methoxytryptamine, NAc-SHT, N-acetylserotonin. For other abbreviations see legend of Figure 1 .
161
Tyndale-Biscoe and Hinds
500
0
400 37
? a
2 300
W
a, v) a, 0
c
? Q 200
2 0 a a - 100
0 I I I -I
-4 0 5 10
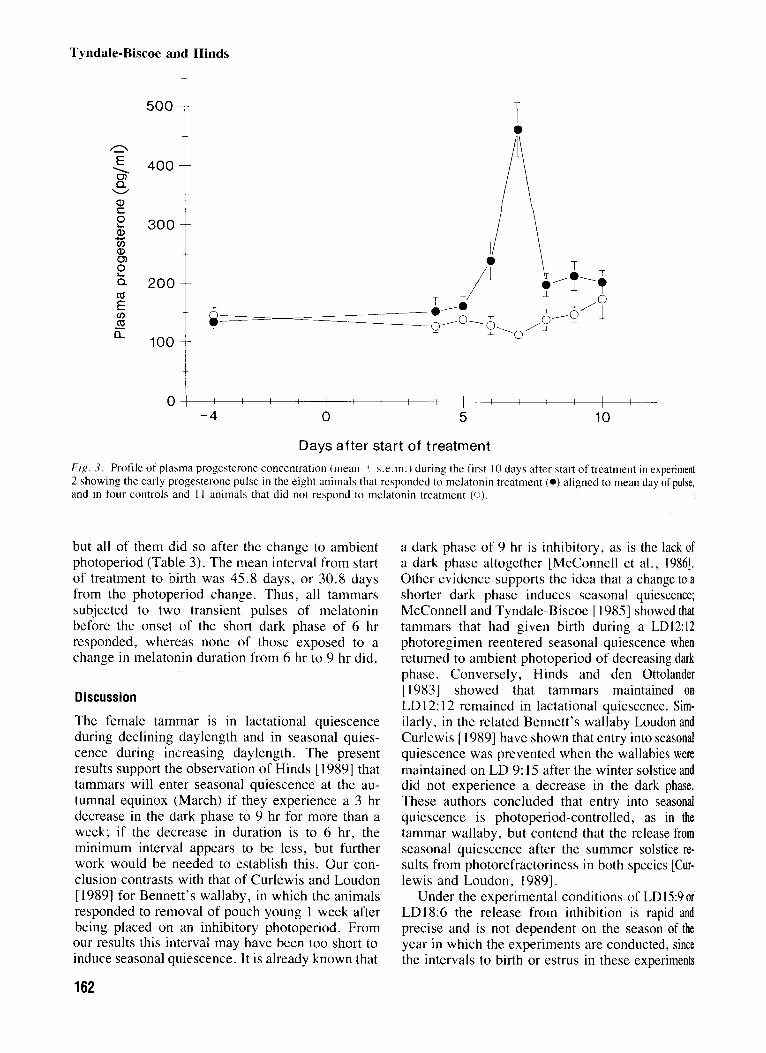
Days after start of treatment Fig . 3. Profile of plasma progesterone concentration (mean ? s.e.m.) during the first 10 days after start of trcatment in experiment 2 showing the early progesterone pulse in the eight animals that responded to mclatonin treatment ( 0 ) aligned to mean day of pulse, and in four controls and I I anirnals that did not respond to melatonin treatment (0).
but all of them did so after the change to ambient photoperiod (Table 3). The mean interval from start of treatment to birth was 45.8 days, or 30.8 days from the photoperiod change. Thus, all tammars subjected to two transient pulses of melatonin before the onset of the short dark phase of 6 hr responded, whereas none of those exposed to a change in melatonin duration from 6 hr to 9 hr did.
Discussion
The female tammar is in lactational quiescence during declining daylength and in seasonal quies- cence during increasing daylength. The present results support the observation of Hinds [ 19891 that tammars will enter seasonal quiescence at the au- tumnal equinox (March) if they experience a 3 hr decrease in the dark phase to 9 hr for more than a week; if the decrease in duration is to 6 hr, the minimum interval appears to be less, but further work would be needed to establish this. Our con- clusion contrasts with that of Curlewis and Loudon [ 19891 for Bennett’s wallaby, in which the animals responded to removal of pouch young 1 week after being placed on an inhibitory photoperiod. From our results this interval may have been too short to induce seasonal quiescence. It is already known that
162
a dark phase of 9 hr is inhibitory, as is the lack of a dark phase altogether [McConnell et al., 19861. Other evidence supports the idea that a change to a shorter dark phase induces seasonal quiescence; McConnell and Tyndale-Biscoe [ 19851 showed that tammars that had given birth during a LD12:12 photoregimen reentered seasonal quiescence when returned to ambient photoperiod of decreasing dark phase. Conversely, Hinds and den Ottolander [ 19831 showed that tammars maintained on LD 12: 12 remained in lactational quiescence. Sim- ilarly, in the related Bennett’s wallaby Loudon and Curlewis [ 19891 have shown that entry into seasonal quiescence was prevented when the wallabies were maintained on LD 9: 15 after the winter solstice and did not experience a decrease in the dark phase. These authors concluded that entry into seasonal quiescence is photoperiod-controlled, as in the tammar wallaby, but contend that the release from seasonal quiescence after the summer solstice re- sults from photorefractoriness in both species [Cur- lewis and Loudon, 19891.
Under the experimental conditions of LD15:9 or LD18:6 the release from inhibition is rapid and precise and is not dependent on the season of the year in which the experiments are conducted, since the intervals to birth or estrus in these experiments
Melatonin and other indoles affect thyroid
T
I I I 111 IV v VI 10 days
0 controls eth. Me1 25 )-1g
Pel c Pel Me1
4l r T
I I I 1 1 1 IV v VI
10 weeks
Me1 50 ug
Pel Mel + Met 25 ug
4 pcO.05 a 0.025 vs. controls eth
Fig. 5 . Relative wet thyroid weights in rats receiving Me1 as S.C. injections and/or as S.C. implanted pellets. For abbreviations see legend of Figure 4.
u U t-
el
0 U \
ul 0) ul 0 L
0 a L
E 1- L 0
aJ L
n 5 = o
I II 111 IV
10 days
a controls eth.
Me1 25pg
I II Ill IV
10 weeks
a - pcO.025 vs. controls eth b - pc0.001 vs.Px c - p<0.025 6. PX d - p<0.005 v5. px
F i g . 6 . Mean mitotic activity rates of thyroid follicular cells (TFC) in rats after pinealectomy (PX) and in Mel-treated pinealectomized rats; PX + Me1 25 kg, Me1 injections in pinealectomized animals. For other abbreviations see legend of Figure I .
Experiment IV
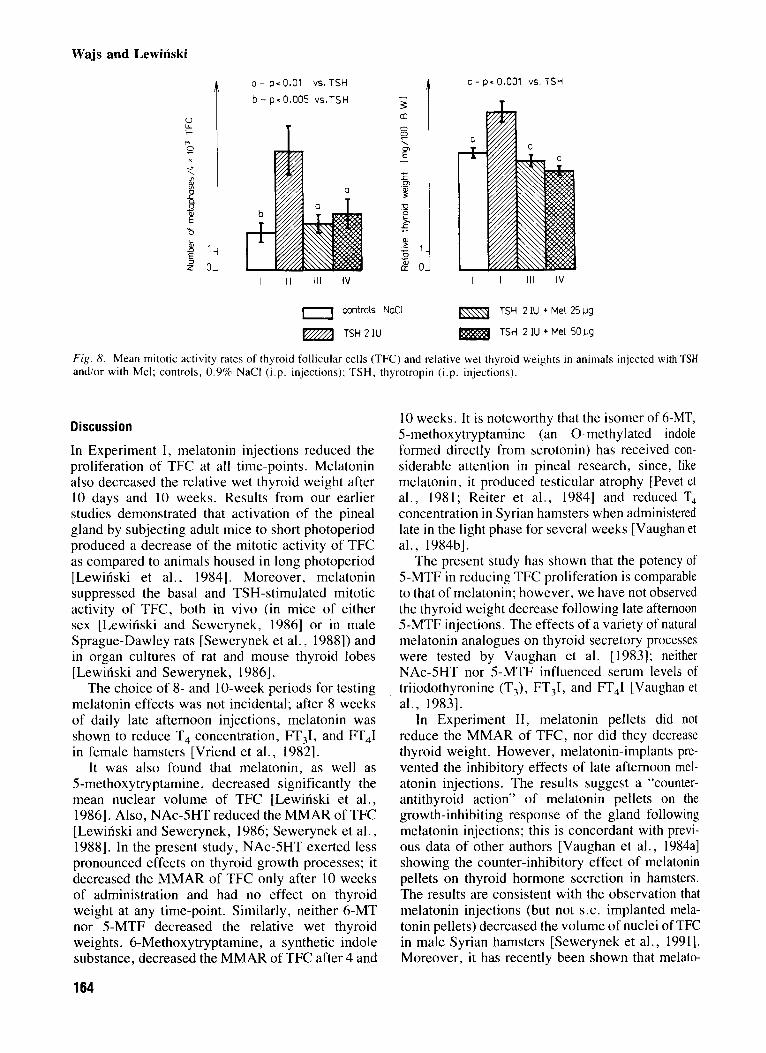
As expected, TSH after 10 days stimulated the mitotic activity of TFC as well as increased thyroid weight. Melatonin (25 pg/daily and 50 kg/daily) abolished the stimulatory effects (Fig. 8).
a - p< 0.05 vs. controls eth.
b - pc 0.025 vs. controls eth. c - p< 0.05 VS. PX d - p<0.005 VS. PX e - p< 0.001 vs. PX
I II 1 1 1 IV I I1 Ill IV
10 days 10 weeks
c] controk eth PX
Mel 25 pg PX+Mel 25 pg
F i g . 7. Relative wet thyroid weights in rats after PX and in Mel-treated pinealectomized animals; PX + Me1 25 kg, Me1 (25 Fgldaily as S.C. injections in pinealectomized animals. For other abbreviations see legend of Figure 1 .
163
Wajs and Lewinski
a - p< 0.01 VS. TSH c - PI 0.001 VS. TSH t b - p e 0.005 VS.TSH - 1 . 3
I I1 1 1 1 IV I I1 1 1 1 IV
a controls NaCl TSH 21U + Me1 25ug
TSH 2 IU + Me1 501.19 TSH21U
Fig . 8. Mean mitotic activity rates of thyroid follicular cells (TFC) and relative wet thyroid weights in animals injected with TSH and/or with Mel; controls, 0.9% NaCl (i.p. injections); TSH. thyrotropin (i.p. injections).
Discussion
In Experiment I, melatonin injections reduced the proliferation of TFC at all time-points. Melatonin also decreased the relative wet thyroid weight after 10 days and 10 weeks. Results from our earlier studies demonstrated that activation of the pineal gland by subjecting adult mice to short photoperiod produced a decrease of the mitotic activity of TFC as compared to animals housed in long photoperiod [Lewinski et al., 19841. Moreover, melatonin suppressed the basal and TSH-stimulated mitotic activity of TFC, both in vivo (in mice of either sex [Lewinski and Sewerynek, 19861 or in male Sprague-Dawley rats [Sewerynek et al., 19881) and in organ cultures of rat and mouse thyroid lobes [Lewinski and Sewerynek, 19861.
The choice of 8- and 10-week periods for testing melatonin effects was not incidental; after 8 weeks of daily late afternoon injections, melatonin was shown to reduce T, concentration, FT,I, and FT,I in female hamsters [Vriend et al., 19821.
It was also found that melatonin, as well as 5-methoxytryptamine, decreased significantly the mean nuclear volume of TFC [Lewinski et al., 19861. Also, NAc-5HT reduced the MMAR of TFC [Lewinski and Sewerynek, 1986; Sewerynek et al., 19881. In the present study, NAc-5HT exerted less pronounced effects on thyroid growth processes; it decreased the MMAR of TFC only after 10 weeks of administration and had no effect on thyroid weight at any time-point. Similarly, neither 6-MT nor 5-MTF decreased the relative wet thyroid weights. 6-Methoxytryptamine, a synthetic indole substance, decreased the MMAR of TFC after 4 and
10 weeks. It is noteworthy that the isomer of 6-MT, 5-methoxytryptamine (an 0-methylated indole formed directly from serotonin) has received con- siderable attention in pineal research, since, like melatonin, it produced testicular atrophy [Pevet et at., 1981; Reiter et al., 19841 and reduced T, concentration in Syrian hamsters when administered late in the light phase for several weeks [Vaughan et al., 1984bl.
The present study has shown that the potency of 5-MTF in reducing TFC proliferation is comparable to that of melatonin; however, we have not observed the thyroid weight decrease following late afternoon 5-MTF injections. The effects of a variety of natural melatonin analogues on thyroid secretory processes were tested by Vaughan et al. [1983]; neither NAc-5HT nor 5-MTF influenced serum levels of triiodothyronine (T3), FTJ, and FT,I [Vaughan et at., 19831.
In Experiment 11, melatonin pellets did not reduce the MMAR of TFC, nor did they decrease thyroid weight. However, melatonin-implants pre- vented the inhibitory effects of late afternoon mel- atonin injections. The results suggest a “counter- antithyroid action” of melatonin pellets on the growth-inhibiting response of the gland following melatonin injections; this is concordant with previ- ous data of other authors [Vaughan et at., 1984al showing the counter-inhibitory effect of melatonin pellets on thyroid hormone secretion in hamsters. The results are consistent with the observation that melatonin injections (but not S.C. implanted mela- tonin pellets) decreased the volume of nuclei of TFC in male Syrian hamsters [Sewerynek et al., 19911. Moreover, it has recently been shown that melato-
164

Melatonin and other indoles affect thyroid
nin, administered to rats as S.C. implanted pellets for 10 days and 10 weeks, exerts an enhancing effect on T, and T, plasma concentrations [Krotewicz et a]., in press].
The existence of different melatonin effects on thyroid secretory processes, depending not only on the way of administration but also on timing [Vriend and Reiter, 19771 and on the dose of melatonin [Vriend et al., 19821, is well documented in ham- sters; high doses of melatonin (2.5 mg daily) prevent the reduction in T, caused by the late afternoon injections of 25 k g of the hormone in question. The hypothesis, which may account for the action of melatonin on the thyroid function and, possibly, growth, assumes that the chronic avail- ability of melatonin renders the presumed receptors insensitive to the acute administration of the indole, i.e., it keeps the receptors down-regulated.
The PX-induced increase of the MMAR of TFC and of thyroid weight, as well as melatonin-induced reversal of the PX effect, confirms our previous results obtained in female Wistar rats [Wajs et a]., 19891. In earlier studies, mild hypertrophy of the thyroid in hypophysectomized mice and rats after PX was found [Houssay et al, 1966; Losada, 19771. Houssay et al. [ 19661 demonstrated that the admin- istration of melatonin prevented the thyroid hyper- trophy obtained in mice after PX.
The observation that melatonin reduced the TSH- induced increase of the MMAR and elevation of thyroid weight is consistent with our previous results obtained after short-term ( 1 0-day) experi- ments on mice in vivo and in in vitro experiments, i.e., incubation of the mouse and rat thyroid lobes with both melatonin and TSH [Lewinski and Se- werynek, 19861.
Summarizing the most important observations from our present paper, it appears that the inhibitory effects of late afternoon melatonin injections and the counter-antithyroid influence of melatonin implants are characteristic phenomena not only for thyroid hormone secretion, but also for growth of the gland. Moreover, these phenomena can be observed not only in hamsters but also in rats.
Literature cited
DAVIS, L. , J. MARTIN (1940) Results of experimental removal of the pineal gland in young mammals. Arch. Neurol. Psychiatry 43:23-25.
DEFRONZO, R.A., W.D. ROTH (1972) Evidence for the exist- ence of a pineal-adrenal and pineal thyroid axis. Acta Endo- crinol. 70:3 1 4 2 .
HINKLE, D.E., W. WIERSMA, S.G. JURS (1979) Applied Statistics for the Behavioral Sciences. Houghton Mifflin Company, Boston.
HOUSSAY, A.B., J.H. PAZO, C.E. EPPER (1966) Effects of the
pineal gland upon their hair cycles in mice. J . Invest. Dermatol. 47:230-234.
KROTEWICZ, M., A. LEwrfisKi, E. WAlS The inhibitory effect of late-afternoon melatonin injections but not of melatonin- containing subcutaneous implants on thyroid hormone secre- tion in male Wistar rats. Neuroendocrinol. Lett. (in press).
KUSZAK J. , M. RODIN (1977) A new technique of pinealec- tomy. Experientia 33:283-294.
LEWIfiSKI, A. , E. SEWERYNEK (1986) Melatonin inhibits the basal and TSH-stimulated mitotic activity of thyroid follicular cells in vivo and in organ culture. J. Pineal Res. 3:291-299.
ITER, N.K.R. SMITH (1984) Inhibitory action of the pineal gland on the nuclear volume of thyroid follicular cells in male mice with intact pineal. Experienta 40: 1284-128s.
M.K. VAUGHAN, R.J. RElTER (1986) Influence of melatonin and 5-methoxytryptamine on nuclear volume of thyroid folli- cular cells in the Syrian hamster (Mesocricetus uuratus). Neuroendocrinol. Lett. 8:63-68.
LOSADA, J. (1977) Effects of experimental pinealectomy. Ann. Anat. 26: 133- 153.
MILINE, R. (1963) La part du noyau paraventriculaire dans I’histophysiologie correlative de la glans thyroide et de la glande pineale. Ann. Endocr. 24:25S-269.
PAZO, J.H., A.B. HOUSSAY, P.A. DAVIDSON, R.J. CHAIT ( 1 968) On the mechanism of thyroid hypertrophy in pinealec- tomized rats. Acta Physiol. Pharmacol. Latinoam. 18:332- 340.
5-methoxytryptophan and 5-methoxytryptamine on the repro- ductive system of the male golden hamster. J. Neural Transm.
REITER, R.J. , A. HOLTORF, T.H. CHAMPNEY, M.K. VAUGHAN (1984) Relative efficacy of melatonin and 5-methox- ytryptamine in terms of their antigonadotrophic and counter- antigonadotrophic actions in male Syrian hamsters. J. Pineal Res. 19-98 .
M ~ N ( 1988) The effect of melatonin and N-acetylserotonin on mitotic activity of thyroid and adrenal cortex in the rat. Endokrynol. Pol. 39:269-275.
R.J. RElTER (1991) Karyometric studies of thyroid follicular cells and thyroid hormone secretion in the male Syrian hamsters following melatonin and diazepam administration. Neuroendocrinol. Lett. 13:393-399.
PETTERBORG, M.C. POWANDA, R.J. REITER, I. SMITH (1983) Natural and synthetic analogues of melatonin and related compounds 11. Effects on plasma thyroid hormones and cholesterol levels in male Syrian hamsters. J. Neural Transm.
VAUGHAN, M.K. , B.A. RICHARDSON, L.J. PETTERBORG, A.P. HOLTORF, G.M. VAUGHAN, T.H. CHAMPNEY, R.J. RElTER (1984a) Effects of injections and/or chronic implants of melatonin and 5-methoxytryptamine on plasma thyroid hor- mones in male and female Syrian hamsters. Neuroendocrinol- ogy 39:361-366.
VAUGHAN, M.K., B.A. RICHARDSON, L.J. PETTERBORG, G.M. VAUGHAN, R.J. REITER (1984b) Pineal indoles and ana- logues-effects on thyroxine in the Syrian hamster. In: The Pineal Gland Endocrine Aspects. G.M. Brown, S.D. Wain- wright, eds. Pergamon Press, New York pp. 251-256.
VRIEND, J . , F.P. GIBBS (1984) Coincidence of counter-anti- gonadal and counter-antithyroid action of melatonin adminis- tration via drinking water in male golden hamsters. Life Sci. 34:6 17-623.
LEWI&SKI, A, , M.K. VAUGHAN, T.H. CHAMPNEY, R.J. RE-
LEWlkKI, A. , S.M. WEBB, E. SEWERYNEK, T.H. CHAMPNEY,
PEVET, P . , c . HALDAR-MISRA, T. OCAL (1981) Effect Of
51:303-311.
SEWERYNEK, E., A. LEWffiSKl, M. SZKUDLiNSKI, G. AEREK-
SEWERYNEK, E., A. LEWIfiSKI, S.M. WEBB, M.K. VAUGHAN,
VAUGHAN, M.K., B.A. RICHARDSON, L.Y. JOHNSON, L.J.
56:279-29 I .
165

Wajs and Lewinski
VRIEND, J., R.J. REITER (1977) Free thyroxin index in normal, melatonin treated and blind hamsters. Hormone Metab. Res. 9:231-234. Endocrinol. 86:758-762.
JOHNSON, R.J. REITER (1982) Effects of melatonin on thyroid physiology of female hamsters. Neuroendocrinology 35:79- 85.
VRLEND, J., J.W. SACKMAN, R.J. REITER (1977) Effects of
blinding, pinealectomy and superior cervical ganglionectomy on free thyroxine index of male golden hamsters. Acta
VRIEND, J . , B . A . RICHARDSON, M.K. VAUGHAN, L.Y. WAJS, E., M. KROTEWICZ, J. FRYCZAK, J. KWAK, E. SE- WERYNEK, M. SZKUDLII~KI , A. LEWISSKI (1989) Melatonin suppresses the pinealectomy-induced increase of the mitotic incidence in the rat thyroid gland. Med. Sci. Res. 17:61-62.





























