Embed Size (px)
Citation preview

Insect External Anatomy © Houseman – page 1
Insect External Anatomy
by Jon G. Houseman
In this lab we continue or observations of the external anatomy we started previously. This time we look at insects and use the grasshopper and cockroach to complete the comparative grid started in the last lab where we examined panarthopods and other insect relatives. We’ll finish up our investigations of external anatomy with a comprehensive look at insect mouthparts and some different types of insect legs. An autapomorphy of the kingdom Animalia is being heterotrophs and a symplesiomorphy that Insects share with their sister groups is that they manipulate their food with their appendages. Insect cuticle can be molded into any one of a variety of structures and this applies to the appendages used for feeding. Is it a surprise that insects have shaped their feeding appendages into the variety of structures you will be looking at later? And, the more different ways you can successfully feed on unique foods is certainly a formula for diversification. The same diversification applies to the different types of insect legs.
Grasshopper: Romalea guttata Orthoptera, the insect order that includes the grasshopper, provides excellent specimens for studying many ancestral features of insects, such as mouthparts and the architecture of the tagmata that form the body. The Lubber grasshopper, Romalea guttata, is a large species and therefore an ideal hopper for lab observations. Unlike most hoppers, however, this species cannot fly because it has small wings; so it disperses by hopping. It’s found in the southeastern United States.
External anatomy The basic arthropod body plan is metameric, with the ancestral segments, metameres, fused to form larger tagmata. Identify the three major insect tagmata in your specimen: head, thorax, and abdomen (Figure 1). What are the functions associated with each of the different insect tagmata?
The arthropods, including insects, are covered with an exoskeleton (cuticle) produced by an underlying epidermis. In the most primitive arthropods, either two or three
Figure 1 Major anatomical features of an insect.

Insect External Anatomy © Houseman – page 2
cuticular plates, the sclerites, surround each segment. A sclerite on the dorsal surface is called a tergite, the plate on the ventral surface is called a sternite, and the cuticular plates on the sides are called pleurites. In the abdomen, there are only two sclerites, the tergite and sternite with and pleural membranes, instead of solid pleurites, connecting the two. Membranes between the plates allowed adjacent sclerites to move relative to each other. For example, the pleural membrane allows the abdomen to expand during feeding, or during egg production in the female. As metameres became fused into tagma, the articulation between the various sclerites often disappeared, and what remains of the junction between the two sclerites is a groove or indentation on the surface of the cuticle and is referred to as a suture. Often, a suture is more than a line on the surface of the cuticle, and internally the cuticle folds to create an internal ridge. What is the advantage of these areas of infolded cuticle for muscle attachment? Not all sutures reflect ancestral segmentation of the tagma, and other indentations of cuticle, apodemes for example, also provide internal sites for muscle attachment.
Head The insect head is a fusion of six segments, three of which are preoral (pregnathal), and three are postoral (postgnathal). The appendages on the preoral segments include the labrum and antennae; there are no appendages on the third head segment. In the closely related subphylum of mandibulate crustaceans, this third head segment had the second pair of antennae. In Atelocerata the second pair of antennae have been lost and this is the origin of the subphylum's taxonomic name. The postoral appendages from front to back are as follows: mandible, maxillae , and fused labium. The labrum and labium form the top and bottom of the preoral cavity respectively; the mandibles and maxilla form the sides.
The head capsule (Figure 2 and Figure 3) is almost a solid piece of hardened cuticle, and most of the sutures and lines visible on its surface are the apodemes of internal cuticular ridges that prevent the head capsule from collapsing under the force of the mouthpart musculature. Only the posterior sutures near the neck reflect the original segmentation of the head. You will look at these posterior sutures in a moment; for now begin with the front of the head. Before you start your observation of the external anatomy, be sure that the surface of the specimen is dry; many of the smaller details
Figure 2 Lateral view of the grasshopper head.

Insect External Anatomy © Houseman – page 3
will be hard to see if the specimen is wet. If necessary, place the specimen on a piece of paper towel and wait for it to dry off.
A pair of compound eyes is located on the sides and near the top of the head (Figure 2). Take a close look under the dissecting microscope, and you will see the complex pattern of ommatidia, the fundamental repeating optic units of the arthropod eye. The part of the ommatidia you see is the cornea (lens). In addition to the pair of compound eyes, there are three simple eyes called the ocelli. A pair of these is situated between the base of the antennae, and the compound eyes. A third medial ocellus is located on the anterior surface of the head, below the groove that runs between the antennae and the compound eyes. Cross sections through an insect head are available that show the compound and simple eyes. Be sure to look at these specimens and compare them to the figures in you lecture notes. The number and position of the ocelli varies in
different insects, but this arrangement of three is the ancestral pattern for the ocelli. Can ocelli form an image; what is their function?
A single pair of antennae is characteristic of the Atelocerata, and each antenna is composed of three basic parts: the scape, which is most proximal, the pedicel, and the flagellum, which is most distal. The flagellum is a single segment, but there is a series of annulations that give it a segmented appearance; the annulations help it bend. How does the arrangement of the internal musculature differ between annulations and a segment in the antennae? Examine the antennae closely using the dissecting microscope to see the sensory setal hairs that cover its surface. Although some setal hairs are mechanoreceptive, they primarily have a chemosensory role. During flight, the antennae bend and move, providing important sensory information on wind speed, direction, and resistance. This twisting and bending of the flagellum generates forces in the cuticle of the pedicel, and these pressures are detected by a special mechanoreceptive structure known as Johnston’s organ. All flying insects have a Johnston’s organ that provides information that enables the insect to adjust its flight to compensate for wind currents.
The other appendages on the head are mouthparts. The fleshy lip is the labrum, which is attached to the head by the clypeus. Look at the side of the head to see the powerful and hardened mandibles (Figure 2).Behind the mandibles, there are the maxillae with their sensory maxillary palps. The last set of appendages on the head is the fused
Figure 3 Anterior view of the grasshopper head

Insect External Anatomy © Houseman – page 4
appendages of the labium, which also have sensory labial palps. You will take a closer look at prepared slides of the hopper mouthparts later in the lab when we compare the ancestral pattern with mouthparts that are specialized for different types of food.
The head capsule is divided into a number of regions, or plates, by sutures and lines. The top of the head is the vertex (Figure 3). From the vertex a set of lines appears as an inverted Y; this is the epicranial suture and when the hopper molts, these lines break open, and the insect emerges from the old cuticle. The cuticle on the sides of the head and behind the epicranial suture is the gena, and the part of the cuticle between the branched base of the epicranial sutures is the frons. Behind the gena there are two more saddle-like plates that cover the dorsal and lateral surfaces of the head: the occiput and postocciput, separated from each other by the postoccipital suture (Figure 3). The postoccipital suture is the only suture on the head that identifies the original segments of the head, the fusion of the fifth and sixth segments. The posterior opening to the head is the foramen.
Two connected sutures run horizontally across the head, above to the attachment of the mouthparts. Inside the head and beneath the two sutures is a ridge of cuticle that encircles the head. The mouthparts articulate with the head in this region, and this ridge of cuticle reinforces the head against the forces generated by the musculature of the mouthparts. The small pits at the lateral edges of the suture are the apodemes that form the endoskeletal tentorium of the head. The other apodemes of the tentorium are located at the base of the postoccipital suture. The tentorium creates internal “flying buttresses” from the back and the front to the top of the head, ensuring that the cranium doesn’t collapse under the forces created by the mouthparts. Cleared specimens of the hopper head are available. Please handle them carefully and look inside the head through the foramen to see components of the tentorium.
Thorax Insects have two pairs of wings, and in the grasshoppers the two wings have a different appearance. The forewings, attached to the mesothorax, are leathery and
Figure 4 Major sclerites and sutures of the prothoracic segment of an insect

Insect External Anatomy © Houseman – page 5
protect the hindwings underneath. This is often the case among the insects, and beetles perfect this strategy by having hardened forewings called elytra that are not used for flying, but as protective wing covers. You can see the difference between the wings by unfolding the hindwing and comparing it to the forewing. The wings are supported by a series of veins, but don’t let the term vein confuse you; in insects the word has nothing to do with the circulatory system of an insect. The veins are ridges of cuticle that strengthen the wing. In the Lubber grasshopper, both sets of wings are small and not large enough for flight.
The thorax is composed of three segments: the prothorax, mesothorax, and metathorax. The thoracic segments can be divided into three major sclerites that have their own special names: the dorsal notum (tergite), lateral pleurites, and the sternum (sternite). Each of these thoracic segments bears a pair of appendages and wings located on the mesothoracic and metathoracic segments, which together form the pterothorax.
The wingless prothorax is formed from the fusion of the four sclerites : a notum, sternum and two lateral pleurites . The dorsal notum, the pronotum, is enlarged and saddle-like, and extends down the sides of the segment. Ancestrally, each of the thoracic and abdominal segments had spiracles. Locate the spiracle on the prothorax. Articulation of the prothoracic leg with the thorax follows the same pattern as the pterothorax (Figure 4). On each segment of the pterothorax, lateral pleurites are fused with the ventral sternites, forming a three-sided thoracic box. The notum is divided into the anterior alinotum and behind it, the postnotum (Figure 5). The postnotum is continuous with the posterior edges of left and right pleurites, and it forms the upper surface of the rigid thoracic box.
Figure 5 Major sclerites and sutures of the insect metathorax.

Insect External Anatomy © Houseman – page 6
The alinotum is connected to the membranous part of the pleura that lies below, and this allows the alinotum to move up and down relative to the box underneath. Look closely at the pleural plates of the mesothorax and metathorax. A central pleural suture divides the pleurite in half and the pleural process is an extension of cuticle at the dorsal end of the pleural suture. It provides the central point of articulation for the wings, which pivot on the upper edges of the pleural process when pushed up and down by the flight muscles. As the notum rises, the wings are lowered; as it falls, the
wings are raised. Similar to the structure of the head, the pleural suture has a ridge of cuticle folded inside, and this cuticle braces the thoracic box against the muscular forces of the flight muscles. Like the pleurites, the sternite is also divided into anterior and posterior portions separated by a suture that functions as an internal reinforcing apodeme. Take a close look at the suture and locate the external apodemes, apophyseal pits, which, along with the plural suture, maintain the rigidity of the thoracic box when the flight muscles contract.
The three pairs of legs are attached on the ventral side of the pleurites and articulate with the ventral end of the pleural suture and with the pleurite. Each leg has six tubular segments, which means that each joint can only move in one direction (Figure 6). As you flex each of the segments, do they all move in the same direction relative to each other? The segment closest to the body is the coxa, and this is followed by the trochanter, the large femur, and the b. In the hopper, the trochanter is fused with the femur; so be sure to find the suture between the two. The fifth segment is the tarsus, and in insects it is divided by a series of up to five articulations called tarsomeres with the pretarsal segment and claws at the tip of the last tarsomere. The prothoracic and mesothoracic legs are very similar to each other; the metathoracic leg is modified for jumping and inside are the muscles that elongate the leg for the jump.
Abdomen Ancestrally, the insect abdomen had eleven segments, but it’s often difficult to see all these because some have become fused together, or reduced in size, or modified because of the genitalia associated with the posterior segments. The first seven abdominal segments have the usual sclerotized tergite and sternite, and membranous pleura and spiracular opening. The terminal abdominal segments bear appendages involved in copulation and egg laying.
Identify the tergite, sternite, pleural membrane, and spiracle on any of the first seven pregenital segments of your specimen (Figure 7). With a few exceptions, these
Figure 6 Anatomy of a typical insect leg.

Insect External Anatomy © Houseman – page 7
segments will look identical. The exceptions include the tympanum on the first abdominal tergite, and the extension of the metathoracic sternite into a groove on the first abdominal sternite. The whole first abdominal segment is fused to the thorax, and
this modification of the sternite helps to lock the abdomen to the thorax. The air sac behind the tympanal membrane is connected to the rest of the tracheal system and it opens to the outside through the spiracle beside the tympanum. Why is it important to have an air sac behind the tympanum? The remaining segments are the genital segments; these retain ancestral appendages that have been modified into the genitalia of the male and female. Look at both male and female specimens.
In the female, the genital plate is the eighth sternite, the last one visible on the ventral surface of the body. Although the eighth tergite is unmodified, tergites nine and ten are partially fused. The eleventh tergite, an enlarged triangular plate, is the epiproct, which has the anus underneath it. Identify the short sensory circus, a modified eleventh appendage, located below the eleventh tergite. In the general plan of the female genitalia, the paired appendages of segments eight and nine form the ovipositor. The valves surround a central egg canal inside, and the gonopore opens at the base of the egg canal. The first paired valves of the ovipositor are on segment eight, and the second and third valves are on segment ten. In the hopper, valves one and three are large and heavily sclerotized; the second valve is reduced in size. The two valves in each pair interlock and are used to dig into the soil before the eggs are laid.
In the male, the ancestral appendages at the tip of the abdomen are involved in transferring sperm, usually inside a spermatophore, to the female. It’s not easy to define a generalized plan for the male, and a variety of different morphologies and terms are used for the male genitalia in insects. Instead of being constructed from the appendages of the eighth and nine segments, male genitalia are formed from the ninth. In your specimen, both the eighth and ninth sternites are visible, and the ninth forms both the genital plate and the ancestral appendage that surrounds the male aedeagus that passes sperm to the female. On the dorsal surface, the tergites resemble those of the female, although the ninth and tenth are not fused.
Figure 7 External anatomy of the grasshopper abdomen

Insect External Anatomy © Houseman – page 8
Cockroach: Periplaneta americana Cockroaches, in the insect order Blattodea, represent some of the most ancient and successful insects, and the best example is the roach Periplaneta americana that you will study in this lab. This cosmopolitan species can be found wherever there are humans. Despite the word americana in their name, they originate in Africa and are believed to have first appeared in North America in the early 1600s, soon after the continent became colonized. Their universal distribution is the result of global shipping, from the age of sailing vessels to the cargo planes of today. Their success can be attributed to their generalized body plan, which also makes this an excellent species for examining the anatomical features of an insect.
External anatomy Use the preserved roaches for your observations of the external anatomy.
Head The head capsule of the roach is very small, and you will need to use the dissecting scope to see the various structures (Figure 8 and Figure 9). Two large, paired, compound eyes are visible on the sides of the head and like you did for the grasshopper look for the individual ommatidia that are the fundamental units of the compound eye. Most insects have an epicranial suture that starts at the top of the head and branches below the eyes; it splits open when the insect molts, but unfortunately, in the roach the suture is almost impossible to see as are many of the cranial sutures you saw in the grasshopper. In addition to the compound eyes, there are ocelli, known as simple eyes, but these will be hard to see in your specimen.
A single pair of antennae is characteristic of the Atelocerata, and each antenna is composed of three basic parts: the scape, which is most proximal, the pedicel, and the annulated flagellum, which is most distal.
Figure 8 Dorsal view of the main external features of a cockroach

Insect External Anatomy © Houseman – page 9
The mandibulate mouthparts of the roach resemble those of the generalized “grasshopper-like” or primitive mouthparts you will examine later in the lab when we compare the mouthparts of different insects. Because the head capsule of a roach is very small, it is almost impossible to remove the individual mouthparts. Try and differentiate between the labrum, and the labium with its labial palps. Without removing mouthparts the labium and maxilla are best seen as a set of palps. If you pull the set of labial and maxillary palps back you should also be able to see the outer surface of the mandible.
Thorax The two pairs of wings that are found in insects have a different appearance in cockroaches (Figure 8). The forewings, attached to the mesothorax, are leathery tegmen and protect the more delicate hindwings on the metathorax. You can see the difference between the wings by unfolding the hindwing and comparing it to the forewing.
The wings are supported by a series of veins, but don’t let the term vein confuse you; in insects the word has nothing to do with the circulatory system. The veins are ridges of cuticle that strengthen the wing, and, starting at the anterior edge, include the costa on the leading edge and the subcosta behind it. The radius, medial, and cubital veins are behind the subcosta, and are part of the remigium. Behind the remigium is the vannus region. This region in roaches is very large and includes the highly branched vannal and anal veins. Most of the folding of the wing involves this vannus region.
Identify the three thoracic segments (Figure 8): prothorax, mesothorax, and metathorax and how they are divided into the dorsal notum (tergite), lateral pleurites, and the sternum (sternite) and the two pairs of wings attached to the pterothorax. In the roaches the pronotum is enlarged into a pronotal shield that protects the underlying body; this is an autapomorphy for the family. Spiracles are
Figure 9 Ventral view of the main external features of a cockroach.

Insect External Anatomy © Houseman – page 10
located on the last two segments of the thorax but appear between the pro- and mesothorax and the meso- and metathorax.
Each leg has six tubular segments (Figure 9). Closest to the body is the large coxa, and this segment is followed by the trochanter, the large femur, and the tibia. The sixth segment is the tarsus, and in insects it is divided by a series of up to five articulations called tarsomeres. Like the antennal articulations observed on the flagellum, the tarsomeres are not segments. The claws at the tip are the nearly invisible pretarsus. Don’t let the name of the pretarsus confuse you; it is located after the tarsus, not before it as the name implies. A fleshy pad is often associated with the claw at the tip of the organism. This pad, when located between the claws, is referred to as an arolium; when located under and at the base of the claw, it is called a pulvillus.
Abdomen Identify the typical abdominal segment components including: sclerotized tergite, sternite, membranous pleura, and spiracular openings. On the terminal segments identify the appendages involved in copulation and egg laying. Most insects have a complex ovipositor formed from the ancestral appendages of the eighth and ninth segments. But this is missing in roaches, an autapomorphy for the order, because of their use of an ootheca. In the female the eighth and ninth segments are reduced and hidden by the seventh, which is enlarged into two genital plates on either side of the genital opening where the ootheca forms. Unlike in the male specimen you will observe next, only the cerci are present. In the male the eight and ninth segments of the abdomen resemble the ancestral form. The tergites of the tenth and eleventh segments have a central notch and extend towards the back of the roach. The cuticular plates on either side of the anal opening are formed from the eleventh sternite; the tenth sternite is reduced in size and not visible. Males have a pair of cerci and stylets at the tip of the abdomen.
The Insect Head Insect heads may have one of three different orientations based on the position of the mouthparts relative to the remainder of the head capsule. When the mouthparts are directed downward the head is considered hypognathous compared to prognathous, when the mouthparts are directed forward. In both these last two cases the oral appendages are located anteriorly but when positioned in the posterior region the head is opisthognathous.
There are some distinct advantages to these different orientations.Many larval insects have prognathous heads because they live in the same medium on which they feed. Forward directed mouthparts are an obvious advantage. Take a look at the mealworm larva as our example of a prognathous head. Others such as the cicada and a variety of plant sap feeding insects must have the mouthparts penetrate the vascular plant tissue. To achieve this requires considerable strength and the opisthognathous head allows the

Insect External Anatomy © Houseman – page 11
insect to apply the pressure required to penetrate the plant. Identify the different mouth part types on the specimens when you examine their mouthparts.
Insect Mouthparts One of the autapomorphies defining the Kingdom Animalia is heterotrophy, and animals must consume high-energy chemical bonds in the form of food and then break these bonds to fuel their life’s processes. It should come as no surprise that animal diversity is often reflected in the varied diets of animals. Specialists feeding on one type of food, phytoplankton for example, have a very different morphology than burrowers living in the sediments of the bottom of the ocean. It’s the same with insects, and one of the reasons for their success is their ability to exploit a wide variety of food sources, from plant sap or blood to feathers, wood, or leaves. In order to consume these different types of food, insect mouthparts have become modified in various ways to create specialized tools for processing different types of food for consumption.
An insect’s mouth lies at the base of a preoral cavity formed by the feeding appendages. This arrangement of appendages is ideal for the manipulation and modification of food before ingestion. The preoral cavity is like an open box: the paired mandibles and maxillae form one pair of opposing sides, and the labrum and labium form the other pair of sides. The front of the preoral cavity is the opening to the box; at the bottom is the mouth and the opening of the salivary glands. The preoral cavity is divided in two by the tongue-like hypopharynx; this creates a dorsal cibarium that leads to the mouth and, below the hypopharynx, a ventral salivarium with the opening of the salivary glands. What appendages on an insect’s head are not involved in feeding?
One of the earliest innovations in insect mouthparts was a change from a mandible with one joint (condyle) connecting it with the head capsule to a double-jointed mandible articulating with the head capsule. The symplesiomorphy of the dicondylic
Figure 10 Three different types of insect heads From left to right, hypognathous, prognathous and opisthognathous

Insect External Anatomy © Houseman – page 12
mandible is used to separate the most primitive hexapods from the insects. Are there any other differences between insects and other more primitive hexapods? This chewing, mandibulate condition is primitive, and the grasshopper is the classic example of this morphology. Fluid-feeding insects have haustellate mouthparts. One of the reasons insects are so successful is that they are able to modify their body cuticle into an amazing array of shapes and forms; think of a dragonfly, a house fly, and a beetle, and you can see just how dramatic these changes can be. The same applies to their mouthparts. For example, a large grinding mandible can be modified into a delicate syringe-like structure that penetrates the circulatory vessels of plants and animals to suck up fluid plant sap or blood. Not only are the shapes of the original mouthparts changed, they often disappear completely. In this lab, you will get a chance to see some of these modifications.
Mandibulate: Chewing and biting The ancestral mouthparts were mandibulate, and teeth on the mandibles ripped the food into pieces that are ground up by the molar surface of the mandible before being swallowed. As was mentioned earlier, the chewing and biting mandible is the most primitive type of insect mouthpart, and the grasshopper is a good example for studying this type of mouthpart.
Grasshopper - mandibulate The mandibulate mouthparts of the grasshopper resemble those of the generalized or
primitive insect (Figure 11). Prepared slides are available of the hopper mouthparts and as you examine them, locate them on the head of the preserved specimen.
The labrum is suspended from the clypeus, and together the two pieces form a plate-like appendage on the roof of the preoral cavity; the underside of the labrum is referred to as the epipharynx. Although not all entomologists agree, it is generally accepted that the labrum is formed from the fused appendages of the first head segment. The mandibles, highly sclerotized and hardened, have a cutting (incisor) and a grinding (molar) surface; find these on your specimen. The mandible articulates
Figure 11 Mouthparts of the grasshopper.

Insect External Anatomy © Houseman – page 13
at the front and back of the gena and is dicondylic with cuticular extensions known as condyles that fit into grooves or slots. The presence of two joints means that the mandible swings in and out when the animal chews. Try to locate the articulations and the condyle.
The maxillae are also paired, and the basal segments include the cardo that articulates with the gena and the stipes, with its tooth-like lacinia, a galea, and a leg-like maxillary palp. The teeth on the lacinia help the mandible to tear up food, while sensory hairs and bristles on the galea and palp are used to taste and feel the food. The labium is the fused appendages of the sixth head segment, and it forms the floor to the preoral cavity. It attaches to the head by the gula, which articulates across the back of the head. The gula has a fleshy inner ligula and outer sensory palps. The hypopharynx is not an appendage but lies in the preoral cavity as a tongue-like structure that has an oral opening above and a salivary opening below: the cibarium and salivarium, respectively.
Haustellate: Fluid feeders Mouthpart morphologies in fluid-feeding insects are classified based on where the ingested fluid is found and how it is ingested. Piercing mouthparts are modified into long needle-like structures, called stylets, that pierce into a plant or animal and form a drinking tube to suck up its internal fluids. Insects can also feed on fluids by either sponging them off a surface or by using a drinking tube to feed on hidden or hard-to-reach liquid foods.
Piercing mouthparts are an autapomorphy that defines the Hemiptera who feed on a range of fluids from plant sap to vertebrate blood. The mosquito, horsefly, house fly, stable fly, and tsetse fly are all in the Order Diptera, which is divided into two groups: the lower and higher flies. One of the differences between the two dipteran groups is found in the morphology of their mouthparts. Lower flies have modified versions of all five mouthparts; higher flies have lost the mandible, and the palp is the only part of the maxilla that remains. Both piercing and sponging mouthparts are found in the Diptera. Adult Lepidoptera are all siphon feeders, using their reduced mouthparts to feed on the nectar of flowers. The fleshy mouthparts of a house fly, which extend as the fly crawls across a table are an example of the sponging type, and the long proboscis of a butterfly sipping nectar from within a flower is an example of the siphoning type of haustellate mouthpart.

Insect External Anatomy © Houseman – page 14
Mosquito - piercing When a female mosquito feeds, the labrum, maxillae, and mandibles (Figure 12) all penetrate the skin, and the labium folds out of the way as the mouthparts cut deeper in search of a capillary. Most of the cutting work is done by the maxilla and mandible. Once a capillary is located, the maxilla anchors the mouthparts, the mandibles cut through the capillary wall, and the tip of labrum is inserted into the blood vessel.
Saliva from the hypopharynx is added to the blood just before it is sucked up the food canal into the cibarium and then into the mouth. Only female mosquitoes are blood feeders, and their saliva contains an anticoagulant that is mixed with the blood so it doesn’t clot as it passes through the food canal. Like male mosquitoes, females also feed on nectar, and the sugar is used to fuel flight in both. The protein of the blood meal consumed by the female is used to produce eggs. Technically it’s not a piercing mouthpart when the mosquito feeds on nectar: Why and what type of a mouthpart is it?
Although whole mount slides of mosquitoes can be used for observation of the mouthparts, it will be difficult to see the individual stylets that are usually inside the labial sheath.
Figure 12 Mouthparts of the mosquito.

Insect External Anatomy © Houseman – page 15
Horse fly – Cutting and sponging Horse flies, Tabanus spp., and deer flies, Crysops spp., are also lower Diptera. All the ancestral mouthparts are present, but highly modified and divided in two main functional components (Figure 13). The cutting blades are formed from the mandibles
and the lacinea of the maxilla. They open and close much like a pair of scissors as they cut through the skin. It is the combined action of the paired mandibles and maxillae that makes the bite of these two flies so painful. The hypopharynx is elongated and has a central salivary canal that releases saliva into the blood that pools in a wound. The labellum is made up of the fleshy lobes of the labium, and each of these lobes is a modified labial palps. Take a close look at the surface of the labellum and you will see it is covered in small channels referred to as pseudotrachea. When the labellum contacts the blood in the wound, the pseudotrachea fill by capillary action. Partial rings of cuticle reinforce the pseudotrachea; this prevents the channels from collapsing when blood is sucked into the food canal that is formed from the inner surface of the labrum. Are maxillary palps present?
Figure 13 Cutting and sponging mouthparts of the horsefly or deer fly
Insect External Anatomy © Houseman – page 16
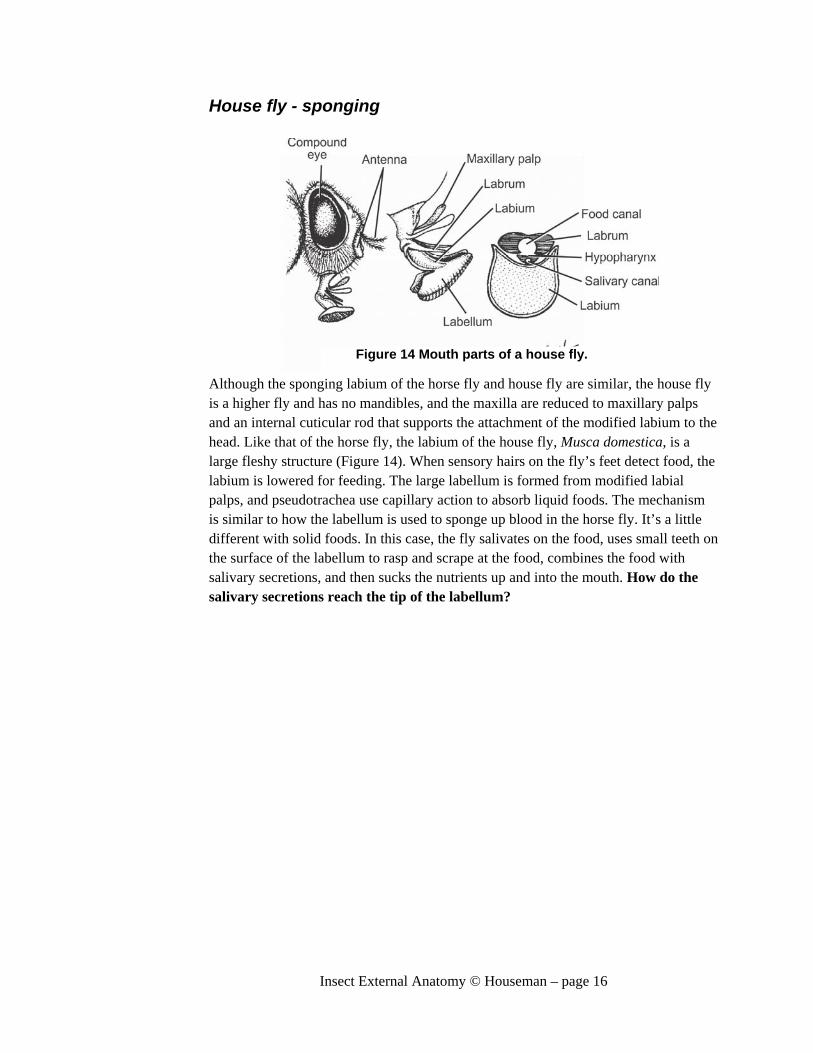
House fly - sponging
Although the sponging labium of the horse fly and house fly are similar, the house fly is a higher fly and has no mandibles, and the maxilla are reduced to maxillary palps and an internal cuticular rod that supports the attachment of the modified labium to the head. Like that of the horse fly, the labium of the house fly, Musca domestica, is a large fleshy structure (Figure 14). When sensory hairs on the fly’s feet detect food, the labium is lowered for feeding. The large labellum is formed from modified labial palps, and pseudotrachea use capillary action to absorb liquid foods. The mechanism is similar to how the labellum is used to sponge up blood in the horse fly. It’s a little different with solid foods. In this case, the fly salivates on the food, uses small teeth on the surface of the labellum to rasp and scrape at the food, combines the food with salivary secretions, and then sucks the nutrients up and into the mouth. How do the salivary secretions reach the tip of the labellum?
Figure 14 Mouth parts of a house fly.

Insect External Anatomy © Houseman – page 17
Stable fly and tsetse fly - piercing The stable fly, Stomoxys sp., and the tsetse fly, Glossina sp., are higher flies. Similarly to other members of the group, the labium is the most prominent mouthpart; the mandible is missing and the palps are all that remain of the maxillae (Figure 16). The
tsetse and the stable fly are both blood feeders; to be able to pierce through skin, the fleshy, large labium typical of the higher flies has been modified into a strong, sturdy shaft or rostrum. The hollow cavity of the rostrum forms the food canal, and prestomal teeth at the tip of the rostrum are used to pierce through the skin. The teeth, which are part of the modified labellum, move in and out of the opening to the buccal cavity as they rasp and dig away at the skin until it is pierced; in the process the mouthparts are pushed deeper into the wound (Figure 15). The hypopharynx extends to the tip of the
rostrum, and the hollow core forms the salivary canal, which delivers an anticoagulant-rich salivary secretion to the blood before it enters the food canal. Blood-feeding insects are often carriers, or vectors, of disease: Are either of these insects vectors, and if they are, what disease(s) do they carry?
Figure 16 With the exception of the labellar teeth the piercing mouthparts of the stable fly and the tsetse fly are similar.
Figure 15 Labellar teeth in the Stable fly (A) and the tsetse fly (B).

Insect External Anatomy © Houseman – page 18
"Bugs": Hemiptera Hemipterans are specialists at piercing, and an autapomorphy of the order is their unique structural “beak” with its interlocking maxillary stylets inside (Figure 17). Like that of the mosquito, the labium forms a sheath surrounding the mouthparts that are modified into stylets. These are used to feed on a variety of foods ranging from seeds to plant fluids and blood. As a hemipteran feeds, the labium folds back out of the way,
and salivary secretions are important in preparing the food for ingestion. In seed feeders, salivary secretions liquefy the seed; in aphids, pectinases dissolve the glue between plant cells so the mouthparts can reach the vascular tissues in the plant. In blood feeders, salivary secretions include anticoagulents. Unlike the other insects you have examined, the Hemiptera don’t have a hypopharynx to deliver the salivary secretions to the tip of the mouthparts. Instead, the salivary canal is part of their unique interlocking maxillary. The paired maxillary stylets, linked to each other in a ziplock-like seal, create the food canal and the salivary canal. Mandibles are the main cutting structure and are found on either side the maxillary stylets. Labial palps have disappeared along with the maxillary palps.
Hemipteran mouthparts can be examined from prepared slides, for example, of the squash bug Anasa tristis or whole mounts of the bloodsucking bug Rhodnius prolixus. The mouthparts on the slides may still be surrounded by the labium.
Figure 17 Mouthparts of a hemipteran define the insect order because of the zipper mechanism that locks the maxillary stylets together.

Insect External Anatomy © Houseman – page 19
Honey bee - Combination The haustellate mouthparts of the honey bee are used to drink the nectar of flowers, a reward for the bee’s services as a pollinator. Nectar is a near-perfect food: an almost pure solution of sucrose and its monosaccharide components of glucose and fructose. Plants produce nectar for no other reason than to attract pollinators, and the honey bee’s mouthparts have been modified to ingest this sweet solution. However, honey bee mouthparts have a second function that isn’t related to feeding. Their mouthparts work the waxes that build the hive’s honeycomb, which stores pollen, honey, and the incubation cells of larval bees. Strictly speaking, honey bees have haustellate mouthparts, but this second function of manipulating wax classifies them as having combination mouthparts. Horse flies could also be placed in the same category: why?
The food canal in a honey bee is different from the simple food tube or “straw” in other insects (Figure 18). The difference is related to one of the properties of the insect cuticle. Insects are able to survive in a desiccating terrestrial environment because of their waxy, waterproof epicuticle. Although it hasn’t been mentioned previously, this is a problem for insects with haustellate mouthparts made of cuticle: the waxy, water-repelling epicuticle also lines the food canal! In most cases, the pumping action of the pharynx is sufficient to overcome the problem. Honey bees, however, have another way of solving the problem.
The donut-shaped food canal in a honey bee is made of a central, cuticular tongue, surrounded by a sheath formed by the other mouthparts; it is not like the simple tube of the other insects you have examined in the lab (Figure 9). The central tongue is formed from the fleshy glossa of the labium and is covered in setal hairs that trap liquid between the hairs. The tongue functions as cuticular sponge, whereby fluid beads up between the water repellent hairs like a droplet of water beads on a freshly waxed surface. When the honey bee feeds, its tongue is extended into the sugar solution and it becomes coated in nectar. When the tongue-like glossa is retracted, it
Figure 18 The mouthparts of a honey bee lap up nectar and then swing out of the way for mandible to manipulate wax.

Insect External Anatomy © Houseman – page 20
becomes surrounded by a sheath that is formed from the galea of the maxilla and modified labial palps. On the ventral surface of the glossa a groove extends up and into the cibarium and the mouth, and either capillary action or suction pulls the nectar up the donut-shaped food canal and into the mouth.
As already mentioned, insects are covered in a waxy cuticle, and the abdominal sclerites of the worker bees produce extra wax, beeswax. The worker bee scrapes the excess wax from the abdomen and chews it with the mandibles, molding it into the hexagonal honeycomb of the hive and the caps that cover the cells. When mandibles are used, the other mouthparts are folded back and out of the way. What type of mouthparts do larval honey bees have?
Butterflies and moths – Drinking tube Adult Lepidoptera, butterflies and moths, feed on the nectar of plants, and most of the ancestral mouthparts are either lost or highly modified. The coiled proboscis is formed from the galea of the maxilla; the maxillary palps have been lost (Figure 19). The galea is hollow and the space inside is an extension of the hemocoel. The proboscis is extended by hydrostatic pressure and recoiled by a combination of internal musculature and cuticular elasticity. The food canal is formed between galea, which is held together by hooks formed from cuticular hairs. Mandibles are missing, and the hypopharynx is part of the cibarial pump. Labial palps are all that remains of the labium.
Figure 19 The mouthparts of Lepidoptera are perfect drinking straws for nectar.

Insect External Anatomy © Houseman – page 21
DragonFly nymph Dragon fly nymphs are deadly predators in freshwater lakes and streams. The whole labium has been modified into a viscous prehensile structure with opposable labial palps at the tip that grasp onto the prey. The overlapping visual field of the large compound eyes is located precisely at the point of full extension of the labium. When a potential meal wanders into this field of view, the mouthparts shoot forward and immediately retract carrying the trapped prey to the mouth. We only have a few of these slides so there will be one placed on demonstration in the lab; be sure to take a look.
Figure 20 Mouthparts of a dragon fly nymph





























