Embed Size (px)
Citation preview

Journal of Bioscience and BioengineeringVOL. 108 No. 5, 446–449, 2009
www.elsevier.com/locate/jbiosc
TECHNICAL NOTE
Insertional chromatin immunoprecipitation: A method for isolatingspecific genomic regions
Akemi Hoshino,1 and Hodaka Fujii1,2,⁎
⁎ CorrespondResearch InstitOsaka 565-087
E-mail add
1389-1723/$doi:10.1016/j
Department of Pathology, New York University School of Medicine, New York, NY 10016, USA1 and Combined Program on Microbiology and Immunology,Research Institute for Microbial Diseases, Osaka University, 3-1 Yamadaoka, Suita-shi, Osaka 565-0871, Japan2
Received 20 April 2009; accepted 11 May 2009
We established a novel method, insertional chromatin immunoprecipitation (iChIP), for isolation of specific genomicregions. In iChIP, specific genomic domains are immunoprecipitated with antibody against a tag, which is fused to theDNA-binding domain of an exogenous DNA-binding protein, whose recognition sequence is inserted into the genomic domainsof interest. The iChIP method will be a useful tool for dissecting chromatin structure of genomic region of interest.
© 2009, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Chromatin structure; Purification; iChIP; Epigenetics; Transcriptional regulation]
Detailed biochemical and molecular biological analysis of chro-matin domains is critical for understanding mechanisms of geneticand epigenetic regulation of gene expression, hetero- and euchroma-tinization, X-chromosome inactivation, genomic imprinting, and otherimportant biological phenomena (1). However, biochemical nature ofchromatin domains is poorly understood. This is mainly becausemethods for performing biochemical and molecular biologicalanalysis of chromatin structure are limited (2–8).
There are several existingmethodologies to analyzemolecules thatinteract with specific genomic regions. For example, if interactingproteins of a genomic region of interest are known, it is possible toisolate the region by the ChIPmethod. However, if interacting proteinsare not known, ChIP cannot be used. In addition, since a DNA-bindingprotein generally binds to many different sites of genomic DNA,complexes immunoprecipitated with Ab against the DNA-bindingprotein are mixtures of many different genomic regions, causingbiochemical analysis to be problematic. For identification of interact-ing genomic regions, chromosome conformation capture (3C) and itsderivatives can be used (9–11). Although non-bias screening ispossible with these methods, they include enzymatic reactions suchas digestion with restriction enzymes and ligation, which areperformed in non-optimal conditions such as under crosslinking,causing artifactual results. In addition, since digestion with restrictionenzymes in non-optimal conditions gives rise to incomplete digestion,PCR amplification of neighboring regions of the target genomic regionoccurs, inhibiting amplification of regions that interact with the targetregion. Fluorescence microscopy including fluorescence in situhybridization (FISH) can be used to show colocalization of specific
ing author. Combined Program on Microbiology and Immunology,ute for Microbial Diseases, Osaka University, 3-1 Yamadaoka, Suita-shi,1, Japan. Tel./fax: +81 6 6879 8358.ress: [email protected] (H. Fujii).
- see front matter © 2009, The Society for Biotechnology, Japan. All.jbiosc.2009.05.005
genomic regions and proteins or RNA. However, this technique cannotbe used for non-bias screening.
Recently, a novel method, proteomics of isolated chromatinsegments (PICh), was developed to purify proteins associated withspecific genomic loci (8). This method utilizes a specific nucleic acidprobe to isolate genomic DNA with its associated proteins, and it wasshown that PICh can successfully isolate telomeres that exist in eachchromosome and have multiple repeats corresponding to the probe. Itis of interest if PICh can be applied to isolation of specific genomicregions in the low copy number genes that contain a single or a fewrepeats corresponding to the probe.
To perform biochemical and molecular biological analysis ofspecific genomic regions, it is essential to purify those regions. Toachieve this goal, we developed the iChIP technology to purify thegenomic regions of interest. The scheme of this system is as follows(Fig. 1):
(i) A repeat of the recognition sequence of LexA is inserted into thegenomic region of interest in the cell to be analyzed (Fig. 1A).This can be achieved by knock-in of the LexA elements into thegenomic region of interest. Alternatively, transgenes containingthe LexA elements in the genomic region of interest can betransfected into the cell to be analyzed.
(ii) The DNA-binding domain (DNA DB) of bacterial DNA-bindingprotein, LexA, was fused with FLAG tag, tobacco etch virus (TEV)protease cleavage site, calmodulin-binding peptide, and thenuclear localization signal (NLS) of SV40 T-antigen (FCNLD)(Fig. 1B), and expressed into the cell to be analyzed. The TEVcleavage site and calmodulin-binding peptide allow us toperform tandem immunoprecipitation.
(iii) The resultant cell is stimulated, if necessary, and crosslinkedwith formaldehyde. This process crosslinks proteins, RNA, DNA
rights reserved.
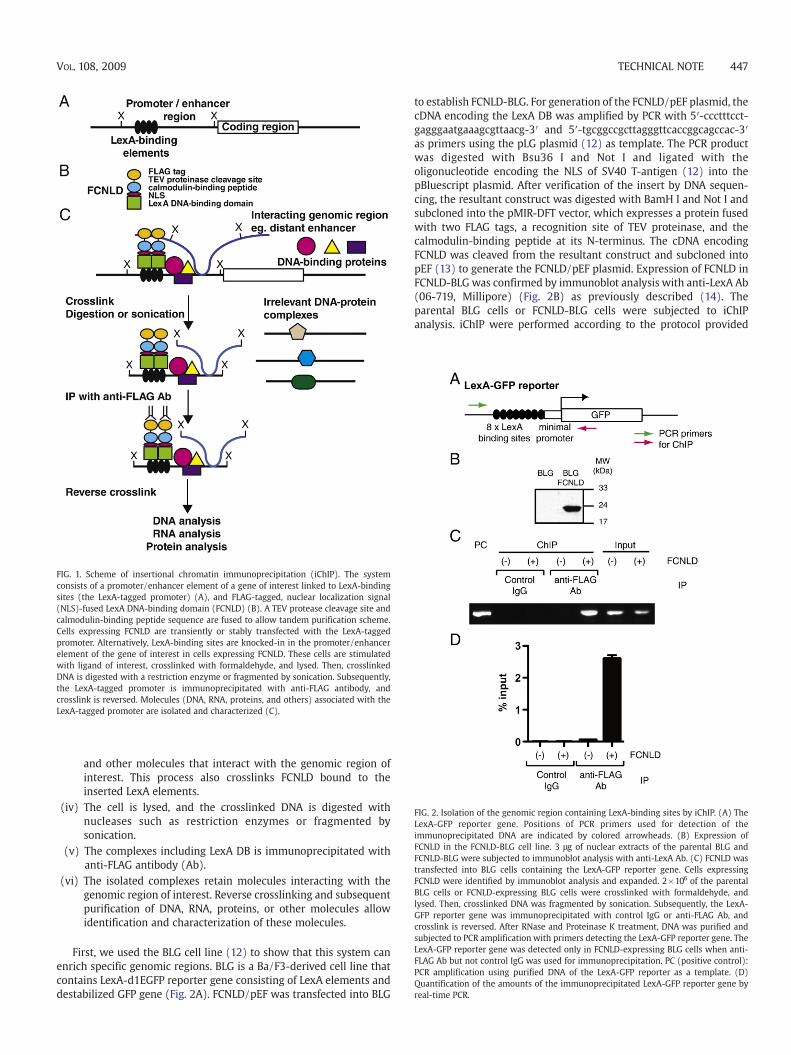
FIG. 1. Scheme of insertional chromatin immunoprecipitation (iChIP). The systemconsists of a promoter/enhancer element of a gene of interest linked to LexA-bindingsites (the LexA-tagged promoter) (A), and FLAG-tagged, nuclear localization signal(NLS)-fused LexA DNA-binding domain (FCNLD) (B). A TEV protease cleavage site andcalmodulin-binding peptide sequence are fused to allow tandem purification scheme.Cells expressing FCNLD are transiently or stably transfected with the LexA-taggedpromoter. Alternatively, LexA-binding sites are knocked-in in the promoter/enhancerelement of the gene of interest in cells expressing FCNLD. These cells are stimulatedwith ligand of interest, crosslinked with formaldehyde, and lysed. Then, crosslinkedDNA is digested with a restriction enzyme or fragmented by sonication. Subsequently,the LexA-tagged promoter is immunoprecipitated with anti-FLAG antibody, andcrosslink is reversed. Molecules (DNA, RNA, proteins, and others) associated with theLexA-tagged promoter are isolated and characterized (C).
FIG. 2. Isolation of the genomic region containing LexA-binding sites by iChIP. (A) TheLexA-GFP reporter gene. Positions of PCR primers used for detection of theimmunoprecipitated DNA are indicated by colored arrowheads. (B) Expression ofFCNLD in the FCNLD-BLG cell line. 3 μg of nuclear extracts of the parental BLG andFCNLD-BLG were subjected to immunoblot analysis with anti-LexA Ab. (C) FCNLD wastransfected into BLG cells containing the LexA-GFP reporter gene. Cells expressingFCNLD were identified by immunoblot analysis and expanded. 2×106 of the parentalBLG cells or FCNLD-expressing BLG cells were crosslinked with formaldehyde, andlysed. Then, crosslinked DNA was fragmented by sonication. Subsequently, the LexA-GFP reporter gene was immunoprecipitated with control IgG or anti-FLAG Ab, andcrosslink is reversed. After RNase and Proteinase K treatment, DNA was purified andsubjected to PCR amplification with primers detecting the LexA-GFP reporter gene. TheLexA-GFP reporter gene was detected only in FCNLD-expressing BLG cells when anti-FLAG Ab but not control IgG was used for immunoprecipitation. PC (positive control):PCR amplification using purified DNA of the LexA-GFP reporter as a template. (D)Quantification of the amounts of the immunoprecipitated LexA-GFP reporter gene byreal-time PCR.
TECHNICAL NOTE 447VOL. 108, 2009
and other molecules that interact with the genomic region ofinterest. This process also crosslinks FCNLD bound to theinserted LexA elements.
(iv) The cell is lysed, and the crosslinked DNA is digested withnucleases such as restriction enzymes or fragmented bysonication.
(v) The complexes including LexA DB is immunoprecipitated withanti-FLAG antibody (Ab).
(vi) The isolated complexes retain molecules interacting with thegenomic region of interest. Reverse crosslinking and subsequentpurification of DNA, RNA, proteins, or other molecules allowidentification and characterization of these molecules.
First, we used the BLG cell line (12) to show that this system canenrich specific genomic regions. BLG is a Ba/F3-derived cell line thatcontains LexA-d1EGFP reporter gene consisting of LexA elements anddestabilized GFP gene (Fig. 2A). FCNLD/pEF was transfected into BLG
to establish FCNLD-BLG. For generation of the FCNLD/pEF plasmid, thecDNA encoding the LexA DB was amplified by PCR with 5′-ccctttcct-gagggaatgaaagcgttaacg-3′ and 5′-tgcggccgcttagggttcaccggcagccac-3′as primers using the pLG plasmid (12) as template. The PCR productwas digested with Bsu36 I and Not I and ligated with theoligonucleotide encoding the NLS of SV40 T-antigen (12) into thepBluescript plasmid. After verification of the insert by DNA sequen-cing, the resultant construct was digested with BamH I and Not I andsubcloned into the pMIR-DFT vector, which expresses a protein fusedwith two FLAG tags, a recognition site of TEV proteinase, and thecalmodulin-binding peptide at its N-terminus. The cDNA encodingFCNLD was cleaved from the resultant construct and subcloned intopEF (13) to generate the FCNLD/pEF plasmid. Expression of FCNLD inFCNLD-BLG was confirmed by immunoblot analysis with anti-LexA Ab(06-719, Millipore) (Fig. 2B) as previously described (14). Theparental BLG cells or FCNLD-BLG cells were subjected to iChIPanalysis. iChIP were performed according to the protocol provided

FIG. 3. Detection of IRF-1 promoter in the chromatin complexes isolated by iChIP. (A)The LexA-element IRF-1 promoter GFP (LIPG) reporter. (B) IFNγ-induced GFPexpression in FCNLD-expressing Ba/F3 containing the LIPG reporter (FCNLD-BLIPG).(C) IFNγ-induced binding of Stat1 to LIPG. 2×107 of FCNLD-BLIPG cells were mock-stimulated or stimulated with IFNγ (10 ng/ml) for the indicated time intervals andsubjected to ChIP analysis using 5 μg of anti-Stat1 Ab. (D) Detection of Stat1-bindingsites adjacent to the LexA-binding elements by iChIP. (E) Detection of Stat1 in thechromatin complexes isolated by iChIP. 1×108 cells were subjected to iChIP using anti-FLAG beads. Chromatin complexes were washed and digested with AcTEV protease at30 °C for 3 h. Second immunoprecipitation was performed with normal mouse IgG oranti-Stat1 Ab and Protein G-Sepharose. Immunoprecipitants were washed extensively,and DNA purification and PCRwere performed as described in the text. Left panel: inputand TEV protease-treated sample. Right panel: results of second ChIP using control IgGand anti-Stat1 Ab.
448 HOSHINO AND FUJII J. BIOSCI. BIOENG.,
by Upstate Biotechnology (EZ ChIP kit) with some modifications.2×106 cells in 10 ml of RPMI complete mediumwere crosslinked with270 μl of 37% formaldehyde for 10 min at room temperature (RT) andneutralized with 1 ml of 1.25 M glycine for 5 min at RT. Aftercentrifugation (1 krpm) for 5 min at RT, cell pellets were washed withice-cold PBS twice and suspended with ice-cold PBS containingprotease inhibitor cocktail (Complete, Mini, EDTA-free, Roche). Aftercentrifugation (700×g) for 5 min at 4 °C, the pellets were suspendedin 400 μl of SDS lysis buffer (50mMTris (pH 8.1),10mMEDTA,1% SDS,protease inhibitor cocktail). Crosslinked DNA was fragmented bysonication (sonicator: Cole Parmer, Ultra Sonic Processor, ModelCP130; probe: CV18, 4273, amplitude 30, 10 s×4 with 10 s intervals).After centrifugation (10 krpm) for 10 min at 4 °C, supernatant wasrecovered and diluted with dilution buffer 1 (0.01% SDS, 1.1% TritonX-100, 1.2 mM EDTA, 16.7 mM Tris–HCl (pH 8.0), 167 mM NaCl,Complete Mini, EDTA-free). Subsequently, the reporter gene wasimmunoprecipitated with control IgG+Protein G-Sepharose (GEHealthcare) or anti-FLAG M2 affinity gel (Sigma-Aldrich). Afterwashing, immunoprecipitated complexes were eluted with 200 μl of
elution buffer (100 mM NaHCO3, 1% SDS). Crosslink was reversed byadding 8 μl of 5 M NaCl and incubation at 65 °C overnight. After RNaseA and Proteinase K treatment, DNA was purified and subjected to PCRamplification. Primers used for detecting the LexA-GFP reporter geneare: 5′-ccccagtgcaagtgcaggtgcc-3′ and 5′-cgtcgccgtccagctcgaccag-3′.As shown in Fig. 2C, the LexA-GFP reporter gene was detected only inthe presence of FCNLD when anti-FLAG Ab but not control IgG wasused for immunoprecipitation. Real-time PCR analysis showed that2.5% of the input LexA-GFP reporter was recovered by iChIP with anti-FLAG Ab (Fig. 2D). These results showed that iChIP can enrich specificgenomic regions.
Next, we examined whether we can purify promoter regionsadjacent to the LexA-binding sites by iChIP. LexA-binding sites wereinserted into upstream of Stat-binding sites of human IRF-1 promoterfused to GFP gene to generate the LexA-element IRF-1 promoter GFPreporter (LIPG). Stat-binding sites of human IRF-1 promoter wereshown to be essential for interferon (IFN) γ-induced transcription ofIRF-1 gene (15). In addition, Stat1 was shown to be necessary forIFNγ-induced transcription of IRF-1 gene (16, 17). For construction ofLIPG, the 1.1 kbp Sac I–Sac II fragment of human IRF-1 promoter (15)was blunted and inserted into Kpn I-digested LexA-d1EGFP (12) afterthe ends were blunted. The resultant plasmid was digested with Xho I,and after its ends were blunted, the blunted 220 bp Sac I–Sac IIfragment of human IRF-1 promoter was inserted. LIPG was stablytransfected into Ba/F3 to generate the BLIPG cell line. Subsequently,FCNLD was stably transfected into BLIPG to generate the FCNLD-BLIPGcell line. Expression of FCNLD was confirmed by immunoblot analysiswith anti-LexA Ab (data not shown). IFNγ-induced expression of GFPreporter was confirmed by flowcytometry (Fig. 3B). Flowcytometricanalysis of GFP expression was performed as previously described(18). We also detected IFNγ-induced Stat1-binding to LIPG by ChIPusing anti-Stat1 Ab (06-501, Santa Cruz Biotechnology) (Fig. 3C).
The FCNLD-BLIPG cells were mock-stimulated or stimulated withIFNγ for 30 min and subjected to iChIP analysis. 1×108 cells percondition were used. After immunoprecipitation with anti-FLAGbeads, chromatin complexes were washed and digested with1000 units of AcTEV protease (Invitrogen) at 30 °C for 3 h. Super-natants (ca. 3 ml) were collected by centrifugation and diluted byadding 12 ml of dilution buffer 2 (0.39% SDS, 1.0% Triton X-100,4.75 mM EDTA, 21.1 mM Tris–HCl (pH 8.0), 144.5 mM NaCl, CompleteMini, EDTA-free). Second immunoprecipitation was performed with5 μg of normal mouse IgG (Millipore) or anti-Stat1 Ab and 40 μl ofProtein G-Sepharose. Immunoprecipitants were washed extensively,and DNA purification and PCR were performed as described above.Primers used for detecting the LIPG are: 5′-tgtacttccccttcgccgctagct-3′and 5′-gcaatccaaacacttagcgggatt-3′. As shown in Fig. 3D, the Stat-binding sites were detected both in mock- and IFNγ-stimulated cellswhen anti-FLAG Ab but not control IgG was used for immunopreci-pitation. These results showed that iChIP system can be used to purifypromoter regions adjacent to the LexA-binding sites.
Next, to examine if the iChIP-isolated IRF-1 promoter regioncontains Stat1, we performed sequential ChIP assay. After immuno-precipitation with anti-FLAG beads, chromatin complexes werereleased by TEV cleavage. Then, second ChIP was performed withcontrol IgG or anti-Stat1 Ab. As shown in Fig. 3E, a band wasspecifically detected in the sample precipitated with anti-Stat1 Ab inan IFNγ-dependent manner. These results showed that the isolatedchromatin complexes by iChIP contain transcription factors activatedby extracellular signal and can be subjected to biochemical andmolecular biological analysis.
The iChIP system enables us to purify specific genomic DNAregions. Biochemical and molecular biological analysis of the purifiedcomplexes can identify their constituents including DNA, RNA,proteins, and others. In other words, this method enables us toidentify unknown interaction of specific genomic DNA regions with

TECHNICAL NOTE 449VOL. 108, 2009
DNA, RNA, proteins and others. Non-bias screening of interactingmolecules is possible using this system. In addition, unknownmodifications of chromatin constituents such as histone proteinscan be revealed. Interacting proteins can be detected by enzyme-linked immunosorbent assay (ELISA), immunoblot analysis, or massspectrometry. Interacting DNA can be detected by region-specific PCR,DNA microarray analysis, or sequencing. Interacting RNA can bedetected by microarray, Northern blot analysis, or RT-PCR. Otherdeveloping technologies for detection of protein, DNA, RNA, and othertypes of molecules can be flexibly combined with this technology.
In summary, we developed the iChIP technology, which can beapplied for (i) identification of transcription factors involved intranscriptional regulation of a gene of interest, (ii) identification ofdistant enhancers, (iii) identification of intra- and interchromosomalinteraction, (iv) biochemical analysis of chromatin structure of specificgenomic regions, (v) biochemistry of euchromatin/heterochromatin,and (vi) biochemical analysis of inactive X-chromosome. We usedtransgenes to show the feasibility of the iChIP technology.We recognizethe possibility that chromosomal condition of genomic transgene maybe different from that of endogenous wild-type locus. The knocking-inof the LexA-binding elements in the genome by gene targeting wouldenable us to utilize the iChIP technique in more physiological contexts.Knock-in technology can be applied to cell lines and primary cells fromany organisms to which the technology has been shown to beapplicable. Although cultured cell lines are much easier to handle,primary cells with targeted modification can be obtained fromindividuals derived from targeted embryonic stem cells. Recentdevelopment of induced pluripotent stem (iPS) cells (19) enables usto apply the iChIP technology to primary human cells. However,insertion of the LexA elements could change the physiologicalchromatin structure. Therefore, it is necessary to validate resultsobtained from the iChIP analysis using wild-type cells with conven-tional techniques. The iChIP techniquewill be a useful tool for dissectingchromatin structure.
ACKNOWLEDGMENTS
We thank Dr. L. Zhou for the pMIR-DFTC vector. We also thankDrs. S. Saint Fleur and T. Fujita for critical reading of the manuscript.
References
1. Kornberg, R. D. and Lorch, Y.: Chromatin rules, Nat. Struct. Mol. Biol., 14, 986–988(2007).
2. Zhang, X. Y. and Horz, W.: Analysis of highly purified satellite DNA containingchromatin from the mouse, Nucleic Acids Res., 10, 1481–1494 (1982).
3. Workman, J. L. and Langmore, J. P.: Nucleoprotein hybridization: a method forisolating specific genes as high molecular weight chromatin, Biochemstry, 24,7486–7497 (1985).
4. Boffa, L. C., Carpaneto, E. M., and Allfrey, V. G.: Isolation of active genes containingCAG repeats by DNA strand invasion by a peptide nucleic acid, Proc. Natl. Acad. Sci.U. S. A., 92, 1901–1905 (1995).
5. Jaskinskas, A. and Hamkalo, B. A.: Purification and initial characterization ofprimate satellite chromatin, Chromosome Res., 7, 341–354 (1999).
6. Griesenbeck, J., Boeger, H., Strattan, J. S., and Kornburg, R. D.: Affinity purificationof specific chromatin segments from chromosomal loci in yeast, Mol. Cell Biol., 23,9275–9282 (2003).
7. Ghirlando, R. and Felsenfeld, G.: Hydrodynamic studies on defined heterochro-matin fragments support a 30-μm fiber having six nucleosomes per turn, J. Mol.Biol., 376, 1417–1425 (2008).
8. Déjardin, J. and Kingston, R. E.: Purification of proteins associated with specificgenomic loci, Cell, 136, 175–186 (2009).
9. Dekker, J., Rippe, K., Dekker, M., and Kleckner, N.: Capturing chromosomeconformation, Science, 295, 1306–1311 (2002).
10. Simonis, M., Klous, P., Splinter, E., Moshkin, Y., Willemsen, R., de Wit, E., vanSteensel, B., and de Laat,W.:Nuclear organization of active and inactive chromatindomains uncovered by chromosome conformation capture-on-chip (4C), Nat.Genet., 38, 1348–1354 (2006).
11. Simonis, M., Kooren, J., and de Laat, W.: An evaluation of 3C-based methods tocapture DNA interactions, Nat. Methods, 4, 895–901 (2007).
12. Hoshino, A., Matsumura, S., Kondo, K., Hirst, J. A., and Fujii, H.: Inducibletranslocation trap: a system for detecting inducible nuclear translocation, Mol. Cell,15, 153–159 (2004).
13. Fujii, H., Nakagawa, Y., Schindler, U., Kawahara, A., Mori, H., Gouilleux, F.,Groner, B., Ihle, J. N., Minami, Y., Miyazaki, T., and Taniguchi, T.: Activation ofStat5 by interleukin 2 requires a carboxyl-terminal region of the interleukin 2receptor β chain but is not essential for the proliferative signal transmission, Proc.Natl. Acad. Sci. U. S. A., 92, 5482–5486 (1995).
14. Hoshino, A. and Fujii, H.: Redundant promoter elements mediate IL-3-inducedexpression of a novel cytokine-inducible gene, cyclon, FEBS Lett., 581, 975–980(2007).
15. Sims, S. H., Cha, Y., Romine, M. F., Gao, P.-Q., Gottlieb, K., and Deisseroth, A. B.:A novel interferon-inducible domain: structural and functional analysis of thehuman interferon regulatory factor 1 gene promoter, Mol. Cell. Biol., 13, 690–702(1993).
16. Durbin, J. E., Hackenmiller, R., Simon,M. C., and Levy, D. E.: Targeted disruption ofthe mouse Stat1 gene results in compromised innate immunity to viral diseases,Cell, 84, 443–450 (1996).
17. Meraz, M. A., White, J. M., Sheehan, K. C. F., Bach, E. A., Rodig, S. J., Dighe, A. S.,Kaplan, D. H., Riley, J. K., Greenlund, A. C., Campbell, D., Carver-Moore, K.,DuBois, R. N., Clarck, R., Aguet, M., and Schreiber, R. D.: Targeted disruption of theStat1 gene in mice reveals unexpected physiologic specificity in the Jak–Statsignaling pathway, Cell, 84, 431–442 (1996).
18. Hoshino, A., Saint Fleur, S., and Fujii, H.: Regulation of Stat1 protein expression byphenylalanine 172 in the coiled-coil domain, Biochem. Biophys. Res. Commun., 346,1062–1066 (2006).
19. Yamanaka, S.: Pluripotency and nuclear reprogramming, Philos. Trans. R. Soc. Lond.B. Biol. Sci., 363, 2079–2087 (2008).





























