Embed Size (px)
Citation preview

Instructions for use
Title Restriction fragment length polymorphism of nuclear rDNA in Sorex caecutiens/shinto group (Eulipotyphla, Soricidae)
Author(s) Naitoh, Yukako; Iwasa, Masahiro A.; Ohdachi, Satoshi D.; Han, Sang-Hoon; Suzuki, Hitoshi
Citation Mammal Study, 30(2): 101-107
Issue Date 2005
Doc URL http://hdl.handle.net/2115/44410
Type article
File Information MS30-2_101-107.pdf
Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP

MammalStudソ30:101-107(2005)
◎theMammalogicalSocietyofJapan
Restrictionfragmentlengthpolymorphismofnuclearrl)NAin
Sorexcaecutiens/shintogroup(Eulipotyphla,Soricidae)
YukakoNaitohi,MasahiroA.Iwasa2,SatoshiD.Ohdachii・de,Sang-HoonHan3
andHitoshiSuzuki`
11nstitute(~ プ五〇wTemperatz〃eScience,HokkaidoUniγersity,SapporoO60-0819,.ノOPan
2五ab・rat・ry・fWildlifeScience,DepartmentofAnimalScien・e・ndRes・urces,C・llege・fBi・res・urceS・iences,嶺 ・n
しiniversiリ ノ,.Ka〃leino1866,Fzグisa「 レレo,.Kanaga「wa252-8510,.ノO卯an
3∠tsiatic、Bla(]k.Bear一 ル1αnage〃zent1セa〃z,ノ 〉∂tionalParks∠{uthoriり ノ,511-1〃7hangleon-ri,ハ4asan-〃1アeon,G〃 ぞアe-gun,
泥o〃a-1>∂md()542-853,」Republic(~ プKorea
4五aboratorP・ofEcologyandGenetics,GraduateSchool()fEnvironmentalEarthScience,HokkaidoUnivez吻,Saρporo
O60-0810,.ノOPan
Abstract.WeestimatedphylogeneticrelationshipsamongshrewsoftheSorexcaecu舵 η誘 伽'o
group(Eulipotyphla,Soricidae)fヒomvariouslocationsthroughitsrange,basedonrestriction
丘agmentlengthpolymorphism(RFLP)analysisofthenuclearribosomalRNAgene(rDNA)spacer
region.SevenrDNA-RFLPrepetitivetypes(repetypes)wererecognizedamongl5shrewsexam-
ined.RestrictionpatternsofSoreac60θ6〃 舵 η5Laxmann,1788and3.sh枷oThomas,1905were
distinguishablefromeachother,buttheseparationwasnotstatisticallysupportedinthemaximum
parsimonyanalysis.TheRFLPrepetypefヒomCh(加Islandwasclosetothatof3.6α ε6〃舵 η8fヒomthe
Eurasiancontinent,indicatingthattheshrewofCh句ushouldbeclassi∬edas3.oα θ磁'θ η5.Within3.
caecutiens,thereweretwoaltemativephylogenetichypotheses.Accordingtoaparsimonioustree
andasimplenetwork,theHokkaidopopulationwasregardedtobederived仕omtheSakhalin
population,whichintumwasderivedfヒomthecontinentalpopulation.Altematively,itwasinferred
thatthecontinentandHokkaidopopulationswerefirstlyseparatedfromtheancestralpopulation,
andthenshrewsfrombothpopulationsimmigratedintoSakhalinandhybridizationoccurredthere.
Thelatterhypothesisseemstobemoreplausiblebecauseitismorecongruentwithaprevious
mitochondrialphylogeny.
Keywords:ChejOuIsland,nuclearrDNA,RFLP,Soreaccaecutiens.
TaxonomicranksofSorexcaecutiensLaxmann,1788
and3.shintoThomas,1905hadbeendebatedbyvarious
authorsbecauseinterpretationoftheirmorphologyis
difficult(seeDokuchaevetal.1999)andlittlephyloge-
neticinfbrmationwasavailablefbrtheseshrews.How-
ever,afterphylogeneticinvestigationsofsomespeciesof
Sorex(Georgel988;Ohdachietal.1997;Fumagalliet
al.1999),itisnowwidelyacceptedthatthesetwospe-
ciesaredistinct.Further,Ohdachietal.(1997,2001,
2003)revealedbasedonmitochondrialcytochromeわ
gene(mtDNAcyt一 わ)sequencesthat3.caecutiensandS.
s伽o鉤 ㎜amonophyleticgroup(3.caecu伽s/s伽o
group),andthat3.caecutiensoccursintheEurasian
continent,Sakhalin,Hokkaido,andneighboringsmall
islandswhile3.shintoisdistributedinthesouthernparts
oftheJapaneseIslands(Honshu,Shikoku,andSado).
FromCh(加Island,SouthKorea,aSorexshrewspecies
wasrecentlydiscovered(H.S.Oh,personalobserva-
tion).Ithasnotyetbeenfbrmallydescribedbutinitial
observationsuggestsitbelongstothe3.caecut'ens/
訪'η'ogroup.Ohdachietal.(2003)showedthatthe
shrewonCh〔 加Islandshouldbeincludedin3.caecu-
'蜘s,basedonmtDNAcyt一 わsequences.
AccordingtothephylogenetictreebasedonmtDNA
cyt一 わ,therearetwomainclustersin3.caecutiens:the
HokkaidoclusterandtheEurasianContinent-Sakhalin一
*7b納0〃icorrespondとnceshOU〃beadと かessed .E-〃iai1'ohd@ρ(4P.IOivte〃1.hokudai.αC.塑
Mamlllal SlIIciy 30: 10 1- 107 (2005) C the Mamma logical Society of Japan
Restriction fragment length polymorphism of nuclear rDNA in Sorex caecutienslshinto group (Eulipotyphla, Soricidae)
Yukako Naitoh', Masahiro A. Iwasa', Satoshi D. Ohdachi" ', Sang-Hoon Han.1 and Hitoshi Suzuki4
/ Instilllte of Low Temperatllre Science, Hokkaido University, Sapporo 060-0819, Japan ] Laboratory of Wildlife Science, Department of Animal Science and Resources, College of Bioresollrce Sciences, Nihon University, Kameino 1866, Fujisawa, Konagawa 252-8510, Japan J Asialic Black Bear Management Team, Nalionol Parks Aulhorify, 51 I- I Whangjeon-ri, JV!ason-myeon, Gwye-gun, Jeolla-Namdo 542-853. Republic of Korea 4 LaboralOry a/Ecology and Genetics, Graduate School oj Environmenlal Earth Science, Hokkaido University. Sapporo 060-08/0, Japan
Ahstract. We est imated phylogenetic re lationsh ips among shrews o f the SOI"ex caecl/fiens/shimo
group (Eulipotyphla, Soricidae) fro l11 va rioll s locat ions through its range, based on restriction fragment length polymorphism (RFLP) ana lys is of the nuclear ribosomal RNA gene (rONA) spacer region . Seven rDNA-RF LP repet itive types (repelypes) were recognized among 15 shrews examined. Restriction patterns of Sorex caeClifiens Laxmann, 1788 and S. shinto Thomas, 1905 were disting uishab le from each other, but the separation was not stat istica ll y supported in the max imum pars imony analysis . The R FLP repetype from Cheju Island was close to that or S. caeClifiens from the Euras ian continent, ind icati ng that the shre\v ofCheju should be class itied as S. caeClifiens. Within S. caeclIfiens, there were two a lternative phylogenetic hypotheses. According to a parsimoniolls tree and a simple netwo rk , the Hokkaido populati on was regarded to be derived from the Sakhalin population, which in turn was deri ved from the continenta l populatio n. Alternati ve ly, it was in ferred ihat the conti nent and Hokkaido populat ions were firs tl y separated from the ancestra l populat ion, and then shrews from hoth populations immigrated into Sakhalin and hyhridization occurred there. The latter hypothesis seems to be more plausible because it is more congruent with a previolls mi tochondrial phylogeny.
Key words: Cheju Island, nuclear rONA, RFLP, Sorex caeclifiens.
Taxonomic ranks of Sorex caecutiens Laxmann, 1788 and S. shil1/o Thomas, 19U5 had been debated by various
authors beca use in te rpretation of their morphology is
difficult (see Dokuchaev et a l. 1999) and little phylogenetic information was ava ilable for these shrews. How
ever, afte r phylogenetic invest iga tions of some species of
Sorex (George 1988; Ohdachi et a l. 1997; Fumagalli et
a l. 1999). it is now widely accepted that these two spe
c ies are di stinct. Further, Ohdac hi et al. ( 1997, 2001, 2003) revealed based on mitochondria l cytochrome b
gene (mtDNA cyt-b) sequences that S. caeclitiens and S.
shinto form a monophy leti c group (s. caecliliens/shinlo
group), and that S. caeclitiens occurs in the Euras ian
continent, Sakhali n, Hokkaido, and neighboring small islands while S. shinto is distributed in the southern parts
of the Japanese Islands (Honshu, Shikoku, and Sado). From Cheju Island, South Korea, a Sorex shrew spec ies
was recentl y discovered (H. S. Oh, personal observa
tion). It has not yet been formally described but initia l observation sugges ts it belongs to the S. caecutiens/
shinto group. Ohdachi et al. (2003) showed that the shrew on Cheju Is land should be included in S. caecll
liens, based on mtDNA cyt-b sequences.
According to the phylogenetic tree based on on tDNA
cyt-b, there are two mai n c lusters in S. caeculiens: the Hokkaido c luster and the Eurasian Cont inent-Sakhalin-
· To whol1l corrcspolldclIcesholilci be addressed. E-mail: [email protected]
Mamlllal SlIIciy 30: 10 1- 107 (2005) C the Mamma logical Society of Japan
Restriction fragment length polymorphism of nuclear rDNA in Sorex caecutienslshinto group (Eulipotyphla, Soricidae)
Yukako Naitoh', Masahiro A. Iwasa', Satoshi D. Ohdachi" ', Sang-Hoon Han.1 and Hitoshi Suzuki4
/ Instilllte of Low Temperatllre Science, Hokkaido University, Sapporo 060-0819, Japan ] Laboratory of Wildlife Science, Department of Animal Science and Resources, College of Bioresollrce Sciences, Nihon University, Kameino 1866, Fujisawa, Konagawa 252-8510, Japan J Asialic Black Bear Management Team, Nalionol Parks Aulhorify, 51 I- I Whangjeon-ri, JV!ason-myeon, Gwye-gun, Jeolla-Namdo 542-853. Republic of Korea 4 LaboralOry a/Ecology and Genetics, Graduate School oj Environmenlal Earth Science, Hokkaido University. Sapporo 060-08/0, Japan
Ahstract. We est imated phylogenetic re lationsh ips among shrews o f the SOI"ex caecl/fiens/shimo
group (Eulipotyphla, Soricidae) fro l11 va rioll s locat ions through its range, based on restriction fragment length polymorphism (RFLP) ana lys is of the nuclear ribosomal RNA gene (rONA) spacer region . Seven rDNA-RF LP repet itive types (repelypes) were recognized among 15 shrews examined. Restriction patterns of Sorex caeClifiens Laxmann, 1788 and S. shinto Thomas, 1905 were disting uishab le from each other, but the separation was not stat istica ll y supported in the max imum pars imony analysis . The R FLP repetype from Cheju Island was close to that or S. caeClifiens from the Euras ian continent, ind icati ng that the shre\v ofCheju should be class itied as S. caeClifiens. Within S. caeclIfiens, there were two a lternative phylogenetic hypotheses. According to a parsimoniolls tree and a simple netwo rk , the Hokkaido populati on was regarded to be derived from the Sakhalin population, which in turn was deri ved from the continenta l populatio n. Alternati ve ly, it was in ferred ihat the conti nent and Hokkaido populat ions were firs tl y separated from the ancestra l populat ion, and then shrews from hoth populations immigrated into Sakhalin and hyhridization occurred there. The latter hypothesis seems to be more plausible because it is more congruent with a previolls mi tochondrial phylogeny.
Key words: Cheju Island, nuclear rONA, RFLP, Sorex caeclifiens.
Taxonomic ranks of Sorex caecutiens Laxmann, 1788 and S. shil1/o Thomas, 19U5 had been debated by various
authors beca use in te rpretation of their morphology is
difficult (see Dokuchaev et a l. 1999) and little phylogenetic information was ava ilable for these shrews. How
ever, afte r phylogenetic invest iga tions of some species of
Sorex (George 1988; Ohdachi et a l. 1997; Fumagalli et
a l. 1999). it is now widely accepted that these two spe
c ies are di stinct. Further, Ohdac hi et al. ( 1997, 2001, 2003) revealed based on mitochondria l cytochrome b
gene (mtDNA cyt-b) sequences that S. caeclitiens and S.
shinto form a monophy leti c group (s. caecliliens/shinlo
group), and that S. caeclitiens occurs in the Euras ian
continent, Sakhali n, Hokkaido, and neighboring small islands while S. shinto is distributed in the southern parts
of the Japanese Islands (Honshu, Shikoku, and Sado). From Cheju Island, South Korea, a Sorex shrew spec ies
was recentl y discovered (H. S. Oh, personal observa
tion). It has not yet been formally described but initia l observation sugges ts it belongs to the S. caecutiens/
shinto group. Ohdachi et al. (2003) showed that the shrew on Cheju Is land should be included in S. caecll
liens, based on mtDNA cyt-b sequences.
According to the phylogenetic tree based on on tDNA
cyt-b, there are two mai n c lusters in S. caeculiens: the Hokkaido c luster and the Eurasian Cont inent-Sakhalin-
· To whol1l corrcspolldclIcesholilci be addressed. E-mail: [email protected]

102 ル勿〃1〃ialStu4ソ30(2005)
Ch()jucluster(Ohdachietal.2003;Ohdachi2005).Fur-
thermore,itwasdemonstratedthatgeneticvariationwas
lowandtherewasalmostnolocaldivergenceofhaplo-
typeamongindividualsof3.caec〃 舵 η8fヒomtheEura-
siancontinentandSakhalin(Ohdachietal.2001,2003).
Basedontheseresults,Ohdachietal.(2001,2003)sug-
gestedthattheancestralpopulationof3.eoeeutie〃8were
firstdividedintotwosubpopulations,oneinHokkaido
andtheotherintheeastempartoftheEurasiancon-
tinent.Then,thelattersubpopulationspreadrapidly
throughoutthecontinentandSakhalinafterthelast
glacialperiod.
PhylogenetictreesinferredfrommtDNA,however,
sometimescontradictthoseinferredfromnuclearDNA
(e.g.Mukai2001;SotaandVogler2001;Iwasaand
Suzuki2002).Insuchcases,taxonomyandevolutionary
eventsinferredfrommtDNAalonewillleadtoerroneous
conclusions.Inthe3.cα θα4〃(~ns/sh'ntogroup,phylo-
geneticinvestigationsbasedonnuclearDNAhavenot
beenconducted.Thus,itisneededtoin色rthephylog-
enyfヒomtheinfbrmationofnuclearDNA.
Inthepresentpaper,weinvestigaterestrictionfrag-
mentlengthpolymorphism(RFLP)ofnuclearriboso-
malRNAgenes(rDNA)andestimatethephylogenetic
relationshipsbetweenspeciesorpopulationsinthe3.
caecutノ θη3/タ痂 η'ogroup.Inaddition,phylogeneticstatus
oftheSorexshrewf士omChq.uIslandisinvestigated.
Herein,weshortlysummarizethenatureofnuclear
rDNA.TherDNAgeneunitmultipliesseveralhundred
timesinanucleargenomeandformsamultigenefamily.
Thus,itisusefUltoestimateanoverallgeneticvariation
ofanucleargenomeofanindividualinapopulation.
TherDNAgeneunitiscomposedofthecodingand
spacerregions.Theformerregions,whichencodehighly
conservedrDNA(18S,5.8S,and28S),aresuitableas
probe-anchoringregions(Arheimetal.1980).Thespacerregions,incontrast,areknowntoevolverapidlyandare
usefultodetectvariationbetweenandwithinspeciesin
phylogeneticinferences(e.g.Arheimetal.1980;Suzukietal.1990;Suzukil994).
Materialsandmethods
Specimens
EightindividualsofS.caecutiensandfiveS.shinto
weresampledfroml210calitiesofnorthemEurasia
(TablelandFig.1).TwoindividualsofSorexshrewspecies(Sorexsp.oftheS.caecutiens/shintogroup)from
ChejuIsland,SouthKorea,werealsoexamined.
S伽hernhybridizationand〃 即 卿9(ゾrestrictionsites
TotalgenomicDNAwasextractedfromlivertissue
preservedin90%ethanolbythephenol/proteinaseK/
SDSmethod(Sambrooketal.1989).ThegenomicDNA
wasdigestedwithsevenrestrictionenzymes,Aatl(A),
Bglll(B),Dral(D),-EcoRI(E),五 伽dIII(H),-Pstl(P)
Table1.Sorexshrewsamplesandtheircollectionlocalitiesexaminedinthisstudy.
Species Repetype LocalityNo, Locality SpecimenCode
S.shinto
Shi-A
Shi.B
Shi-B
Shi.B
Shi-C
1
(∠
弓」
3
4.
Mt,Kamegamori,Shikoku,Japan
NaganoPref.,Honshu,Japan
Ryotsu,SadoIs.,Japan
Ryotsu,SadoIs。,Japan
Mt。Hayachine,Honshu,Japan
SO97/5/25-l
SO96misc-13
HA5961
HA5962
SO97/8/5-1
s.caecutiens
Cae-A
Cae-A
Cae.B
Cae-C
Cae.C
Cae-C
Cae-C
Cae-C
5
6
7
8
9
0
2
3
1
1
1
Kaminokuni,Hokkaido,Japan
Sarobetsumoor,Hokkaido,Japan
Trudovoe,Sakhalin,Russia
Kedrovaya,Primorye,Russia
Mt.Gaya,SouthKorea
Mt.Odae,SouthKorea
Milkovo,Kamchatka,Russia
Pallasj註rvi,Lapland,Finland
SO97/6/7-l
SO94scl
SO94sak5
SO96/7/16-2
SO99misc.9
SO99/10/17-l
SO97misc-82
VH365
Sorexsp.
Sp-Cheju
Sp-Cheju
l
l
l
l
Mt。Halla,ChejuIs.,SouthKorea
Mt.Halla,Ch()juIs.,SouthKorea
SO99/10/12-l
SO99/10/13-1
102
Cheju cluster (Ohdachi et a J. 2003; Ohdachi 2005). Furthermore, it was demonstrated that genetic variation was low and there was almost no local divergence of haplotype among ind ividuals of S. eoeeliliens from the Eurasian continent and Sakhalin (Ohdachi et a J. 200 I, 2003). Based on these results, Ohdachi et a J. (2001 , 2003) suggested that the ancestral populat ion of S. eoeell/iens were first div ided into two subpopu lations, one in Hokkaido and the other in the eastern part of the Eurasian continent. Then, the latter subpopu lat ion spread rapid ly
throughout the continent and Sakha lin after the last glacial period.
Phylogenetic trees inferred from mtDNA, however, sometimes contradict those inferred from nuclear DNA (e.g. Mukai 2001; Sota and Vogler 2001 ; Iwasa and Suzuki 2002). In such cases, taxonomy and evolutionary events inferred from mtDNA alone wil l lead to erroneous conclusions. In the S. caecliliens/shinlo group, phylogenetic investigations based on nuclear DNA have not
been conducted. Thus, it is needed to infer the phy logeny from the information of nuclear DNA.
In the present paper, we investigate restriction fragment length po lymorphism (RFLP) of nuclear ribosomal RNA genes (rDNA) and est imate the phylogenetic relationships behveen species or popu lations in the S.
caeclil iens/shinta group. fn addition, phylogenetic status of the Sorex shrew from Cheju Island is investigated. Herein, we shortly summarize the nature of nuclear
Mammol Study 30 (2005)
rDNA. The rDNA gene unit mUltip lies several hundred times in a nuclear genome and form s a ITI ultigene fam ily. Thus, it is useful to estimate an overall genetic variation of a nuclear genome of an individual in a popu lation. The rDNA gene unit is composed of the coding and spacer regions. The fanner regions, which encode highly conserved rDNA ( 18S, 5.8S, and 28S), are suitable as probe-anchoring regions (Arheim et aJ. 1980). The spacer regions, in contrast, are known to evo lve rapidly and are usefu l to detect variat ion between and withi n species in phylogenetic inferences (e.g. Arhe im et aJ. 1980; Suzuk i et aJ. 1990; Suzuki 1994).
Materials and methods
Specimens
Eight individuals of S. caecllliens and five S. shinJO
were samp led from 12 locali ties of northern Eurasia (Table I and Fig. I). Two individuals of Sorex shrew
species (Sorex sp. of the S. eoeell/iens/shin/o group) from Cheju Island, South Korea, were also examined.
SOUlhern hybridization and mapping afrestriction siles Tota l genomic DNA was extracted from liver tissue
preserved in 90% ethanol by the phenol/proteinase K/ SDS method (Sambrook et a J. 1989). The genomic DNA was digested with seven restriction enzymes, Aa/I (A),
8gnJ (B), Dral (D), EeoRl (E), HindllJ (H), Psll (P)
Ta ble 1. Sorer shrew samples and thei r co llection loca liti es examined in this study.
Species Repcrype Loca lity No. Loca lity Specimen Code
S. shima
Shi·A Mt. Kamegamori, Shikoku, Japan 5097/5/25-1
Shi-B 2 Nagano Pref.. Honshu. Japan S096misc-13
Shi-O J Ryotsu, Sado \s. , Japan l lA5961
Shi-B 3 Ryotsu, Sado Is. , Japan HA5962
Shi -C 4 ML Hayachin e, Honshu, Japan 5097/8/5- 1
S. caecllfiens
Cae-A 5 Kmninokuni, Il okkaido, Japan S097/6/7-1
Cae-A 6 Sarobetsu moor, Hokka ido. Japan S094sc l
Cae- B 7 Trudovoe, Sakha lin, Russia S094sak5
Cae-C 8 Kedrovaya, Primorye, Russ ia S096/7/ 16-2
Cae-C 9 Mt. Gaya, South Korea S099misc-9
Cae-C 10 Me Odae, South Korea S099/ IO/l7- 1
Cae-C 12 Milkovo, Kamchatka, Russia S097misc-82
Cae-C 13 Pa llasjarvi, Lapland, Fin land VH365
Sorex sp.
Sp-C hcj u II Mt. I lall a, Chcj u Is., Soulh Korea S099/ I 0112-1
Sp-Cheju II Mt. Halla, Chej u Is., Soulh Korea S099/ I 0/13- 1
102
Cheju cluster (Ohdachi et a J. 2003; Ohdachi 2005). Furthermore, it was demonstrated that genetic variation was low and there was almost no local divergence of haplotype among ind ividuals of S. eoeeliliens from the Eurasian continent and Sakhalin (Ohdachi et a J. 200 I, 2003). Based on these results, Ohdachi et a J. (2001 , 2003) suggested that the ancestral populat ion of S. eoeell/iens were first div ided into two subpopu lations, one in Hokkaido and the other in the eastern part of the Eurasian continent. Then, the latter subpopu lat ion spread rapid ly
throughout the continent and Sakha lin after the last glacial period.
Phylogenetic trees inferred from mtDNA, however, sometimes contradict those inferred from nuclear DNA (e.g. Mukai 2001; Sota and Vogler 2001 ; Iwasa and Suzuki 2002). In such cases, taxonomy and evolutionary events inferred from mtDNA alone wil l lead to erroneous conclusions. In the S. caecliliens/shinlo group, phylogenetic investigations based on nuclear DNA have not
been conducted. Thus, it is needed to infer the phy logeny from the information of nuclear DNA.
In the present paper, we investigate restriction fragment length po lymorphism (RFLP) of nuclear ribosomal RNA genes (rDNA) and est imate the phylogenetic relationships behveen species or popu lations in the S.
caeclil iens/shinta group. fn addition, phylogenetic status of the Sorex shrew from Cheju Island is investigated. Herein, we shortly summarize the nature of nuclear
Mammol Study 30 (2005)
rDNA. The rDNA gene unit mUltip lies several hundred times in a nuclear genome and form s a ITI ultigene fam ily. Thus, it is useful to estimate an overall genetic variation of a nuclear genome of an individual in a popu lation. The rDNA gene unit is composed of the coding and spacer regions. The fanner regions, which encode highly conserved rDNA ( 18S, 5.8S, and 28S), are suitable as probe-anchoring regions (Arheim et aJ. 1980). The spacer regions, in contrast, are known to evo lve rapidly and are usefu l to detect variat ion between and withi n species in phylogenetic inferences (e.g. Arhe im et aJ. 1980; Suzuk i et aJ. 1990; Suzuki 1994).
Materials and methods
Specimens
Eight individuals of S. caecllliens and five S. shinJO
were samp led from 12 locali ties of northern Eurasia (Table I and Fig. I). Two individuals of Sorex shrew
species (Sorex sp. of the S. eoeell/iens/shin/o group) from Cheju Island, South Korea, were also examined.
SOUlhern hybridization and mapping afrestriction siles Tota l genomic DNA was extracted from liver tissue
preserved in 90% ethanol by the phenol/proteinase K/ SDS method (Sambrook et a J. 1989). The genomic DNA was digested with seven restriction enzymes, Aa/I (A),
8gnJ (B), Dral (D), EeoRl (E), HindllJ (H), Psll (P)
Ta ble 1. Sorer shrew samples and thei r co llection loca liti es examined in this study.
Species Repcrype Loca lity No. Loca lity Specimen Code
S. shima
Shi·A Mt. Kamegamori, Shikoku, Japan 5097/5/25-1
Shi-B 2 Nagano Pref.. Honshu. Japan S096misc-13
Shi-O J Ryotsu, Sado \s. , Japan l lA5961
Shi-B 3 Ryotsu, Sado Is. , Japan HA5962
Shi -C 4 ML Hayachin e, Honshu, Japan 5097/8/5- 1
S. caecllfiens
Cae-A 5 Kmninokuni, Il okkaido, Japan S097/6/7-1
Cae-A 6 Sarobetsu moor, Hokka ido. Japan S094sc l
Cae- B 7 Trudovoe, Sakha lin, Russia S094sak5
Cae-C 8 Kedrovaya, Primorye, Russ ia S096/7/ 16-2
Cae-C 9 Mt. Gaya, South Korea S099misc-9
Cae-C 10 Me Odae, South Korea S099/ IO/l7- 1
Cae-C 12 Milkovo, Kamchatka, Russia S097misc-82
Cae-C 13 Pa llasjarvi, Lapland, Fin land VH365
Sorex sp.
Sp-C hcj u II Mt. I lall a, Chcj u Is., Soulh Korea S099/ I 0112-1
Sp-Cheju II Mt. Halla, Chej u Is., Soulh Korea S099/ I 0/13- 1

Naitohetal.,RF乙PqプnuclearD八 「Ainshrei・vs 103
andPvull(V).AfterelectrophoresiswithO.7%agarose
gelfbrl7hoursat30V,thedigestedDNAwasimmo-
bilizedonnylonmembrane(Hybond-N,Amersham)and
hybridizedwithdigoxigenin-labeled(DIGDNALabel-
ingKitandDetectionKit,RocheDiagnostics)28Sprobe
(0.7μg/ml)frommouse(Suzukietal.1994).Detection
oftheprobeonthemembranefollowedthesupplier's
protocol.Fragmentlengthsweredeterminedbycom-
parisonwithasizemarker(DNAMolecularWeight
MarkerIIdigoxigenin-labeledlambdaHindlll).Restric-
tionmapsofthe28Sdownstreamspacerregionwere
constructedbyrefeningtothestandardrestrictionmap
ofthel8S,5.8S,and28SrRNAgenesthatareconser-
vativeamongmammalianspecies(Suzukietal.1994;
Iwasaetal.2001).
Phylogeneticona!yses
Allrestrictionsiteswerenumberedfromltol3(Fig.
60。N 80。N
.ノ
、
ぷ避
ぐ
詳
ノ
ギ
/
/
≠
ゲ
縁
溝
Cheju7
話11
Kyushu
や
締 、
Hokkaido
XtTsug・ ・uSt・ait
⊂=⊃S.caecutiens
-s.shinto
9〈'12
Fig.1.Collectionlocalitiesofshrewsamples.LocalitynumberscorrespondwiththoseofTablel.
Naifoh el al.. RFLP oillllclear DNA ill shrews
and PvurJ (V). After electrophoresis with 0.7% agarose gel for 17 hours at 30 Y, the digested DNA was immobilized on nylon membrane (Hybond-N, Amersham) and hybridized with digoxigenin-Iabeled (DIG DNA Labeling Kit and Detection Kit, Roche Diagnostics) 28S probe
(0.7 ,"g/ml) from mouse (Suzuki et al. 1994). Detection of the probe on the membrane followed the supplier' s protocol. Fragment lengths were determined by comparison with a size marker (DNA Molecular Weight
SO ON BO ' N
0 1000 km
1:'>0 0
q,~.§'
t1
103
Marker J] digoxigenin-labeled lambda HindlJl). Restric
tion maps of the 28S downstream spacer region were constructed by referring to the standard restriction map of the 18S, S.8S, and 28S rRNA genes that are conservative among mammalian species (Suzuki et al. 1994;
Iwasa et al. 200 I).
Phylogenetic analyses All restriction sites were numbered from I to 13 (Fig.
Baika lake
7
'* ·r
<'f , §
~O ; 0 0'" Cj t~
Pacific Ocean
2
c=J S. caecutiens _ So shinto
Fig. I. Collect ion loca lities o f shrew sampl es. Locality numbers correspond with those of Table I.
Naifoh el al.. RFLP oillllclear DNA ill shrews
and PvurJ (V). After electrophoresis with 0.7% agarose gel for 17 hours at 30 Y, the digested DNA was immobilized on nylon membrane (Hybond-N, Amersham) and hybridized with digoxigenin-Iabeled (DIG DNA Labeling Kit and Detection Kit, Roche Diagnostics) 28S probe
(0.7 ,"g/ml) from mouse (Suzuki et al. 1994). Detection of the probe on the membrane followed the supplier' s protocol. Fragment lengths were determined by comparison with a size marker (DNA Molecular Weight
SO ON BO ' N
0 1000 km
1:'>0 0
q,~.§'
t1
103
Marker J] digoxigenin-labeled lambda HindlJl). Restric
tion maps of the 28S downstream spacer region were constructed by referring to the standard restriction map of the 18S, S.8S, and 28S rRNA genes that are conservative among mammalian species (Suzuki et al. 1994;
Iwasa et al. 200 I).
Phylogenetic analyses All restriction sites were numbered from I to 13 (Fig.
Baika lake
7
'* ·r
<'f , §
~O ; 0 0'" Cj t~
Pacific Ocean
2
c=J S. caecutiens _ So shinto
Fig. I. Collect ion loca lities o f shrew sampl es. Locality numbers correspond with those of Table I.

104 ル勿〃1〃ialStu4ソ30(2005)
-spacerregionatdownstreamof28Sgene
k--28S・RNAg・ ・e-一 明Il PHEGDA V
1kb一一
PH EGD AG V E
.畠=
PH D AG V E
PH D AGG V E一V PH ADG E
▲
V PH D DADG V E一V PH DAG V E一1 23 456 789101112 13
restrictionsitenumber
Fig.2.RestrictionmapsofthemajorrepeatingunittypesoftheribosomalRNAgenedownstreamofthe28ScodingregionintheSorex
caecutiens/shintogroupfromvariouslocalitiesofEurasia.Abbreviationsofrestrictionenzymes:A,Aatl;B,Bglll;D,Dral;E,EcoRI;H,Hindlll;
P,Pstl;V,1)vull.Amongl5individualsexamined,somehadidenticalrepetitivetypes(repetypes).SeeTablelf()rindividualsexaminedandtheir
RFLPrepetyes.Opentrianglesindicatepolymorphicsitesassizevariationwithinagenomeofanindividual。
2)andconvertedintoaO/ldatamatrix,whereaOindi-
catesabsenceofthebandandalindicatespresence.
Phylogeneticanalyseswereconductedusingthemaxi-
mumparsimonymethod,appliedusingPAUPver.
4.OblO(Swofford2000)withtheexhaustivesearch
methodandDollo'scriterion.Thecostfbrgainofa
restrictionsitewassixtimeshigherthanfbrlossofasite
becausetherestrictionenzymesusedallhadasixbase
recognitionsequence(SwoffordandOlsenl990).Nodal
supportwasevaluatedbylO,000bootstrapreplicateswith
50%majorityruleconsensus.Aparsimoniousnetwork
wasalsoconstructedaccordingtoBandelt(1994).This
methodindicateseverychangeofstateinanetwork.
Thus,whenthenumberofinformativesitesissmall,this
methodismorestraightfbrwardthanothermethodsof
phylogenyreconstruction.
Results
Restrictionmap
SevenrDNA-RFLPrepetitivetypes(repetypes)were
recognizedamongl5shrewsexamined(Fig.2).The
restrictionpattemsofS.shintoandS.caecutienswere
distinguishedbysitel(Fig.2).FourindividualsofS.
shintoshowedthreerDNA-RFLPrepetitivetype:Shi-A,
Shi-BandShi-C(Tablel,Fig.2).Incontrast,fivein-
dividualsofS.caecutiensfromseverallocalitiesonthe
EurasiancontinentincludingtheKoreaPeninsulashared
onlyonerepetype(Cae-C;Tablel,Fig.2).TwoS.
caecutiensindividualsfromHokkaidohadasingle
repetype(Cae-A;Tablel,Fig.2),andsites4and5were
notobservedinCae-Cwhilesitesllandl3shownin
Cae-CwereabsentinCae-A(Fig.2).Therepetypefrom
Sakhalin(Cae-B)hadsites4,5,ll,andl3(Fig.2),
demonstratingitcontainedcharacteristicsofrepetypes
bothfromthecontinent(Cae-C)andHokkaido(Cae-A).
TwoindividualsfromChej'uIslandhadasinglerepe-
type(Sp-Cheju),whichshowedauniquerestrictionsite
(sitelO)butothersiteswereidenticaltotherepetype
(Cae-C)fromthecontinent(Fig.2).
C1α ∂bgra〃2α 掘 雁wo鴻
Oneshortesttree(treelength=16)wasobtainedby
themaximumparsimonymethod(Fig.3).Sorexsh'η'o
and3.6α θ6〃'1θη8weredistinguishedinthecladogram
(Fig.3)buttheconfidenceofthisdivisionwasstatisti-
callylow(bootstrapvalues,<50%).Therepetypefヒom
Sp-Ch〔 加(Chq.uIsland)wasincludedinthe3.oα θo〃一
伽scladeandformedasubcladewithCae-C(Eurasian
continent).TherepetypesfbrCae-A(Hokkaido)and
Cae-B(Sakhalin)formedasecondsubcladewithinthe
3.caecutiensclade(Fig.3).
Inthephylogeneticnetwork(Fig.4A),3.caecutiens
and3.3痂 η'oweredistinguishedbyrestrictionsitel,and
104 Mammal Study 30 (2005)
spacer region at downstream of 28S gene 1kb
28S rR NA gene • PH E G O A V ---- I 1 I \1 1 1 PH EGO A G V E ---- I I I \1 1 1 I I
PH 0 A G V E ---- I I I 1 !. 1 I
PH 0 AG G V E ____ smHeI§i'_ I I I 11 !. I I
V PH AOG E
~ I I I 1(!. I
V PH 0 OAOG V E
~ I I I I 'if !. 1 I
V PH OA G V E
~ I I I ';j, !. 1 I
2 3 4 5 6 7 8 9 10 11 12 13 restriction site number I I I ~I '-'l l ~ ~ I I
Fig. 2. Restr ict ion maps of the major repeating unit types of the ri bosomal RNA gene dow nstream of the 28S codi ng region in the Sorex caeclItiells/shinfo group From various localit ies of Ellrasia. Abbrevi~Hions o f restriction enzymes: A, Aal l ; B, BgflI: 0 , Dral ; E. EcoRl ; H. Hilldll[ ~
P, Pstl ; V, PVII II . Among 15 ind ividua ls examined. some had identical repetitive types (repetypes) . See Table I for individuals exam ined and thei r RFLP repetyes. Open triang les indicate polymorphic s ites as size var iat ion within a genome of an ind ividual.
2) and converted in to a 0/ 1 data matrix, where a 0 indi
cates absence of the band and a 1 indicates presence. Phylogenetic ana lyses were conducted us ing the maxi
mum pars imony method, appl ied using PAUP ver. 4 .0b I 0 (Swofford 2000) with the exhausti ve search
method and Da lla ' s criterion. The cost for ga in of a
restriction site was six times higher than for loss of a site
because the restriction enzymes used a ll had a six base
recogniti on sequence (Swofford and O lsen 1990). Nodal
support was evaluated by 10,000 bootstrap replicates with 50% majority ru le consensus. A parsimoniolls nehvork
was a lso constructed according to Bandelt ( 1994). This
method indicates every change of state in a network. Thus, when the number of into nnative sites is small , th is
method is more stra ightforward than other methods of
phylogeny reconstruction.
R es u lts
Restriction map
Seven rDNA-RFLP repetiti ve types (repetypes) were recognized among IS shrews examined (Fig. 2). The
restriction patte rns of S. shinto and S. caeclIfiens were distinguished by site I (Fig. 2). Four individua ls of S.
shil/to showed three rDNA-RFLP repetitive type: Shi-A,
Shi-B and Shi-C (Table I , Fig. 2). In contrast, fi ve individua ls of S. caecutiens from severa l localities on the
Euras ian continent including the Korea Pen insula shared onl y one repetype (Cae-C; Table I, Fig. 2). Two S.
caecllfiens individuals from Hokkaido had a single repetype (Cae-A; Table I, Fig. 2), and sites 4 and 5 were
not observed in Cae-C whil e sites I I and 13 shown in
Cae-C were absent in Cae-A (Fig. 2). The repetype from
Sakhalin (Cae-B) had sites 4 , 5, I I, and 13 (Fig. 2), demonstrating it conta ined characterist ics of repetypes
both from the continent (Cae-C) and Hokkaido (Cae-A). Two individua ls from C heju Island had a sing le repe
type (Sp-Chej u), which showed a unique restriction si te
(s ite 10) but other sites were identical to the repetype (Cae-C) from the continent (F ig. 2).
Cladogram and network One shortest tree (tree length = 16) was obta ined by
the maximum parsimony method (Fig. 3). Sorex shinto and S. caeclftiens were distinguished in the c ladogram
(Fig. 3) but the confide nce of thi s d ivision was stat isti
ca lly low (bootstrap va lues, <50%). The repetype from Sp-C heju (Chej u Island) was included in the S. caec,,liens clade and formed a sube lade with Cae-C (Euras ian
continent). The repetypes for Cae-A (Hokkaido) and Cae-B (Sakhalin) formed a second subc lade within the
S. caeculiens clade (F ig. 3).
In the phylogenetic network (Fig. 4A), S. caeculiens and S. shinto were d istinguished by restr iction site I , and
104 Mammal Study 30 (2005)
spacer region at downstream of 28S gene 1kb
28S rR NA gene • PH E G O A V ---- I 1 I \1 1 1 PH EGO A G V E ---- I I I \1 1 1 I I
PH 0 A G V E ---- I I I 1 !. 1 I
PH 0 AG G V E ____ smHeI§i'_ I I I 11 !. I I
V PH AOG E
~ I I I 1(!. I
V PH 0 OAOG V E
~ I I I I 'if !. 1 I
V PH OA G V E
~ I I I ';j, !. 1 I
2 3 4 5 6 7 8 9 10 11 12 13 restriction site number I I I ~I '-'l l ~ ~ I I
Fig. 2. Restr ict ion maps of the major repeating unit types of the ri bosomal RNA gene dow nstream of the 28S codi ng region in the Sorex caeclItiells/shinfo group From various localit ies of Ellrasia. Abbrevi~Hions o f restriction enzymes: A, Aal l ; B, BgflI: 0 , Dral ; E. EcoRl ; H. Hilldll[ ~
P, Pstl ; V, PVII II . Among 15 ind ividua ls examined. some had identical repetitive types (repetypes) . See Table I for individuals exam ined and thei r RFLP repetyes. Open triang les indicate polymorphic s ites as size var iat ion within a genome of an ind ividual.
2) and converted in to a 0/ 1 data matrix, where a 0 indi
cates absence of the band and a 1 indicates presence. Phylogenetic ana lyses were conducted us ing the maxi
mum pars imony method, appl ied using PAUP ver. 4 .0b I 0 (Swofford 2000) with the exhausti ve search
method and Da lla ' s criterion. The cost for ga in of a
restriction site was six times higher than for loss of a site
because the restriction enzymes used a ll had a six base
recogniti on sequence (Swofford and O lsen 1990). Nodal
support was evaluated by 10,000 bootstrap replicates with 50% majority ru le consensus. A parsimoniolls nehvork
was a lso constructed according to Bandelt ( 1994). This
method indicates every change of state in a network. Thus, when the number of into nnative sites is small , th is
method is more stra ightforward than other methods of
phylogeny reconstruction.
R es u lts
Restriction map
Seven rDNA-RFLP repetiti ve types (repetypes) were recognized among IS shrews examined (Fig. 2). The
restriction patte rns of S. shinto and S. caeclIfiens were distinguished by site I (Fig. 2). Four individua ls of S.
shil/to showed three rDNA-RFLP repetitive type: Shi-A,
Shi-B and Shi-C (Table I , Fig. 2). In contrast, fi ve individua ls of S. caecutiens from severa l localities on the
Euras ian continent including the Korea Pen insula shared onl y one repetype (Cae-C; Table I, Fig. 2). Two S.
caecllfiens individuals from Hokkaido had a single repetype (Cae-A; Table I, Fig. 2), and sites 4 and 5 were
not observed in Cae-C whil e sites I I and 13 shown in
Cae-C were absent in Cae-A (Fig. 2). The repetype from
Sakhalin (Cae-B) had sites 4 , 5, I I, and 13 (Fig. 2), demonstrating it conta ined characterist ics of repetypes
both from the continent (Cae-C) and Hokkaido (Cae-A). Two individua ls from C heju Island had a sing le repe
type (Sp-Chej u), which showed a unique restriction si te
(s ite 10) but other sites were identical to the repetype (Cae-C) from the continent (F ig. 2).
Cladogram and network One shortest tree (tree length = 16) was obta ined by
the maximum parsimony method (Fig. 3). Sorex shinto and S. caeclftiens were distinguished in the c ladogram
(Fig. 3) but the confide nce of thi s d ivision was stat isti
ca lly low (bootstrap va lues, <50%). The repetype from Sp-C heju (Chej u Island) was included in the S. caec,,liens clade and formed a sube lade with Cae-C (Euras ian
continent). The repetypes for Cae-A (Hokkaido) and Cae-B (Sakhalin) formed a second subc lade within the
S. caeculiens clade (F ig. 3).
In the phylogenetic network (Fig. 4A), S. caeculiens and S. shinto were d istinguished by restr iction site I , and
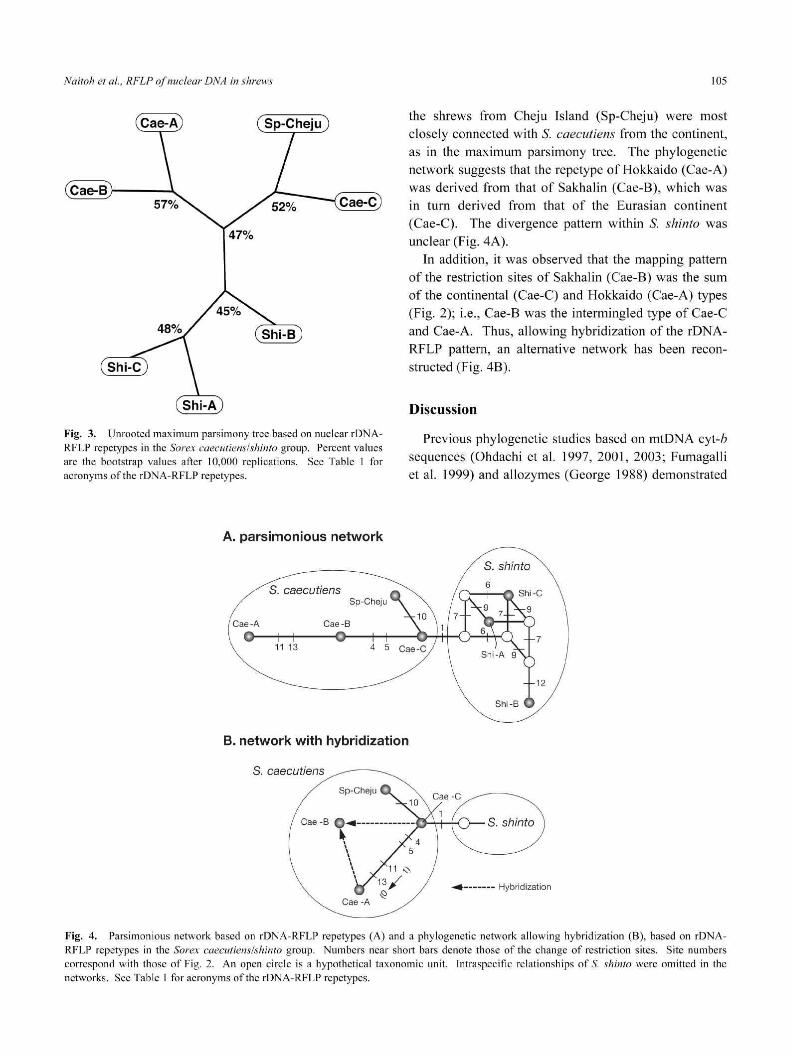
ハlaitohetal.,」RF乙 、PofηuclearDハIAinshrews 105
Sp・Cheju
Fig.3.UnrootedmaximumparsimonytreebasedonnuclearrDNA-
RFLPrepetypesintheSorexcaecutiens/shintogroup.Percentvalues
arethebootstrapvaluesafterlO,000replications.SeeTablelfbr
acronymsoftherDNA-RFLPrepetypes.
theshrewsfromCh(加Island(Sp-Ch(加)weremost
closelyconnectedwithS.eoeeutiensfromthecontinent,
asinthemaximumparsimonytree.Thephylogenetic
networksuggeststhattherepetypeofHokkaido(Cae-A)
wasderivedfromthatofSakhalin(Cae-B),whichwas
inturnderivedfromthatoftheEurasiancontinent
(Cae-C).ThedivergencepattemwithinS.shintowas
unclear(Fig.4A).
Inaddition,itwasobservedthatthemappingpattem
oftherestrictionsitesofSakhalin(Cae-B)wasthesum
ofthecontinental(Cae-C)andHokkaido(Cae-A)types
(Fig.2);i.e.,Cae-BwastheintermingledtypeofCae-C
andCae-A.Thus,allowinghybridizationoftherDNA-
RFLPpattem,analternativenetworkhasbeenrecon-
structed(Fig.4B).
Discussion
PreviousphylogeneticstudiesbasedonmtDNAcyt-b
sequences(Ohdachietal.1997,2001,2003;Fumagalli
etal.1999)andallozymes(Georgel988)demonstrated
A.parsimoniousnetwork
B.networkwithhybridization
S.caecu亡1eηs
11N)x
13/Q
Hybridization
Fig.4.ParsimoniousnetworkbasedonrDNA-RFLPrepetypes(A)andaphylogeneticnetworkallowinghybridization(B),basedonrDNA-
RFLPrepetypesinthe5「orexcaecutiens/shintogroup.Numbersnearshortbarsdenotethoseofthechangeofrestrictionsites。Sitenumbers
correspondwiththoseofFig。2.Anopencircleisahypotheticaltaxonomicunit.IntraspecificrelationshipsofS.shintowereomittedinthe
networks.SeeTablelfbracronymsoftherDNA-RFLPrepetypes.
Naifoh el al.. RFLP oillllclear DNA ill shrews
Cae-B 57% 52% Cae-C
Fig. 3. Unrooted maximum parsimony tree based on nuc lear rDNA RFLP repetypes in the Sorex caeclifiells/shillfO group. Perce nt va lues are the bootstrap va lu es after 10,000 repl ications. See Table I for acronyms oflhe rDNA-RFLP repetypes .
A. parsimonious network
S. caecutiens Sp-Cheju
Cae-A Cae-8
11 13
B. network with hybridization
S. caecutiens
Sp-Cheju
105
the shrews fro m Cheju Island (Sp-Cheju) were most closely connected with S. caecutiens from the continent,
as in the maximum parsimony tree. The phylogenetic network suggests that the repetype of Hokkaido (Cae-A) was derived from that of Sakhalin (Cae-S), which was in turn derived from that of the Eurasian continent
(Cae-C). The divergence pattern within S. shinlo was unclear (Fig. 4A).
In addition, it was observed that the mapping pattern of the restriction sites of Sakhal in (Cae-S) was the sum of the continenta l (Cae-C) and Hokkaido (Cae-A) types (Fig. 2); i.e. , Cae-S was the interm ingled type of Cae-C and Cae-A. Thus, allowing hybridi zation of the rDNARFLP pattern, an alternative network has been reconstructed (Fig. 4S).
Discussion
Previous phylogenetic studies based on mtDNA cyt-b
sequences (Ohdach i et al. 1997, 200 1, 2003; Fumagalli et al. 1999) and allozymes (George 1988) demonstrated
S. shinto
Cae -8 O ... -------------I!:f+t-+o - S. shinto
~
\ .. ------- Hybridization
Cae-A
Fig. 4. Parsimonious net\vork based on rDNA -RFLP repetypes (A) and a phylogenetic network allow ing hybridization (8 ), based on rDNA RFLP repety pes in the Sorex caeclifiens/sh infO group. Numbers near ShOlt bars denote those of the change of restriction si tes. Site num bers correspond wi lh those of Fig. 2. An open c irc le is a hypolhet ical taxonomic unit. Inlraspecific relationships of S. shimo were omitted in the networks. See Table I fo r ac ronyms of the rDNA-RFLP repetypes.
Naifoh el al.. RFLP oillllclear DNA ill shrews
Cae-B 57% 52% Cae-C
Fig. 3. Unrooted maximum parsimony tree based on nuc lear rDNA RFLP repetypes in the Sorex caeclifiells/shillfO group. Perce nt va lues are the bootstrap va lu es after 10,000 repl ications. See Table I for acronyms oflhe rDNA-RFLP repetypes .
A. parsimonious network
S. caecutiens Sp-Cheju
Cae-A Cae-8
11 13
B. network with hybridization
S. caecutiens
Sp-Cheju
105
the shrews fro m Cheju Island (Sp-Cheju) were most closely connected with S. caecutiens from the continent,
as in the maximum parsimony tree. The phylogenetic network suggests that the repetype of Hokkaido (Cae-A) was derived from that of Sakhalin (Cae-S), which was in turn derived from that of the Eurasian continent
(Cae-C). The divergence pattern within S. shinlo was unclear (Fig. 4A).
In addition, it was observed that the mapping pattern of the restriction sites of Sakhal in (Cae-S) was the sum of the continenta l (Cae-C) and Hokkaido (Cae-A) types (Fig. 2); i.e. , Cae-S was the interm ingled type of Cae-C and Cae-A. Thus, allowing hybridi zation of the rDNARFLP pattern, an alternative network has been reconstructed (Fig. 4S).
Discussion
Previous phylogenetic studies based on mtDNA cyt-b
sequences (Ohdach i et al. 1997, 200 1, 2003; Fumagalli et al. 1999) and allozymes (George 1988) demonstrated
S. shinto
Cae -8 O ... -------------I!:f+t-+o - S. shinto
~
\ .. ------- Hybridization
Cae-A
Fig. 4. Parsimonious net\vork based on rDNA -RFLP repetypes (A) and a phylogenetic network allow ing hybridization (8 ), based on rDNA RFLP repety pes in the Sorex caeclifiens/sh infO group. Numbers near ShOlt bars denote those of the change of restriction si tes. Site num bers correspond wi lh those of Fig. 2. An open c irc le is a hypolhet ical taxonomic unit. Inlraspecific relationships of S. shimo were omitted in the networks. See Table I fo r ac ronyms of the rDNA-RFLP repetypes.

106 ル勿〃1〃ialStu4ソ30(2005)
A.hypotheticaIphylogeny(Tree1)
Eurasian
Continent
Sakhalin
S.caeα ノオleηs
B.hypotheticalphylogeny(Tree2)
S.caecutiens
SakhalinAr
o
Eurasian
Continent
Fig.5.TwohypotheticalphylogeniesofnuclearDNAintheSorexcaecutiens/shintogroup,basedontherDNA-RFLPrepetypes,Treel(A)
wasinferredfromtheparsimoniousnetwork(Fig.4A)andtree2(B)fromthenetworkallowinghybridization(Fig。4B)。Arrowswithbrokenlines
indicatehybridization.
thatS.caecutiensandS.shintoshouldbetreatedastwo
separatespecies.Inthepresentstudy,itwasnotstatis-
ticallysupported,althoughtopologyshowedthesetwo
wereseparated(Fig.3).However,theshrewfromCh〔 加
IslandwasmostclosetoS.caecutiensintheEurasian
continentanditshouldberecognizedasS.caecutiens
(Fig.3).Inaddition,althoughsamplinglocationsfrom
theEurasiancontinentweregreatlyscattered,therewas
novariationintherDNA-RFLPrepetypewithinS.cae-
cutiensoverthisrange(Tablel,Fig.2).Thesameten-
dencywasobservedinthegeneticvariationofmtDNA
cyt一わ(Ohdachietal.2001,2003;Ohdachi2005).These
findingsfピombothnuclearDNAandmtDNAsuggest
thatS.caecutienshasgonethrougharecentandrapid
rangeexpansionintheEurasiancontinent.Pattemsof
geneticvariationwerelargelydifferentbetweenHokkaido
andthecontinentbothinrDNA-RFLPrepetypes(Fig.4)
andmtDNAhaplotypes(Ohdachietal.2001,2003;
Ohdachi2005).Therefbre,therangeexpansionofS.
caecutiensinthecontinentmusthaveoccurredafter
Hokkaidoandthecontinentweregeologicallyseparated
afterthelastglacialperiod.ItisestimatedthatHokkaido
andSakhalin+Eurasiancontinentwereseparatedca.
12,000yearsago(Ohshimal990).Thus,thegreat
rangeexpansioninthecontinentpresumablybeganafter
Variouslocatities
intheEurasian
Continentand
Sakhalin
Fig.6.SchematictreeofthemitochondrialDNAphylogenyinthe
Sorexcaecutiens/shintogroup,basedonmtDNAcytochromebgenesequences(Ohdachietal.2003;Ohdachiinpress),Anopencircleisa
hypotheticaltaxonomicunit.
12,000yearsago.
Themaximumparsimonytree(Fig.3)andthephylo-
geneticnetwork(Fig.4A)wereusedtoconstructahypotheticalphylogenyofthenuclearrDNA(Treel,
Fig.5A).Ontheotherhand,allowinghybridization
betweentherDNA-RFLPs,analtemativenetworkwas
reconstructed(Fig.4B),andfromthisphylogeneticnet-
workwithhybridization,anotherhypotheticalphylogeny
wasinferred(Tree2,Fig.5B).
Wecannotconcludewhichphylogeny(Fig.5:treesl
and2)ismoreappropriateasanevolutionaryprocessof
106 Mammal Study 30 (2005)
A. hypothetical phylogeny (Tree 1)
S. shinto Cheju
Hokkaido Shikoku
Sakhalin Honshu
S. caecutiens
B. hypothetical phylogeny (Tree 2)
S. caecutiens S. shinto Cheju
Shikoku
Sakhalin ...... ••••• Honshu
o .... ~~.
Hokkaido
Fig. 5. Two hypothetical phylogenies of nuclear DNA in the Sorel' caeculiells/sitilllo group, based on the rDNA- RFLP rcpetypcs. Tree I (A) was inferred from the parsimonious network (Fig. 4A ) and tree 2 (B) from the network allowing hybridization (Fig. 4B). Arrows \vith broken lines indicate hybridi zation .
that S. caecutiens and S. shinto should be treated as t\vo separate species. In the present study, it was not stat is
tically supported, although topology showed these two were separated (Fig. 3). However, the shrew trom Cheju
Island was most close to S. caeculiens in the Eurasian
cont inent and it should be recognized as S. caecliliens
(Fig. 3). In addi tion, a lthough sampling locations from
the Eurasian continent were great ly scattered, there was
no variation in the rDNA-RFLP repetype within S. cae
cliliens over this range (Table I , Fig. 2). The same tendency was observed in the genetic variation of IlltDNA
cyt-b (Ohdachi et a l. 2001, 2003 ; Ohdachi 2005). These findings from both nuclear DNA and mtDNA suggest
that S. caecliliens has gone through a recent and rapid range expansion in the Eurasian cont inent. Patterns of
genetic variation were largely different between Hokkaido
and the continent both in rDNA-R FLP repetypes (Fig. 4)
and mtDNA haplotypes (Ohdachi et al. 200 1, 2003;
Ohdachi 2005). Therefore, the range expansion of S.
caeclfliens in the continent Illust have occurred after
Hokkaido and the continent were geologica lly separated
after the last glac ial period. It is estimated that Hokkaido and Sakha lin+ Eurasian continent were separated ca.
12 ,000 years ago (Ohshima 1990). Thus, the great
range expansion in the continent presumably began after
S. caecutiens
Various local ities in the Eurasian Continent and Sakhalin
S. shinto
Fig. 6. Schematic tree of the mitochondrial DNA phylogeny in the So/'ex caecutiens/shillto group, based on mtDNA cytochrom e b gene sequences (Ohdachi et al. 2003; Ohdachi in press). An open ci rcle is a hypothetical taxonomic unit.
12,000 years ago.
The maximum parsimony tree (Fig. 3) and the phylogenetic network (Fig. 4A) were used to construct a
hypothetical phylogeny of the nuclear rDNA (Tree I, Fig. SA). On the other hand, a llowing hybridization
between the rDNA-RFLPs, an a lternative network was
reconstructed (Fig. 4B), and from this phylogenetic net
work with hybridi zation, another hypothetical phylogeny
was inferred (Tree 2, Fig. 5B). We can not conclude which phylogeny (F ig. 5: trees I
and 2) is more appropriate as an evolutionary process of
106 Mammal Study 30 (2005)
A. hypothetical phylogeny (Tree 1)
S. shinto Cheju
Hokkaido Shikoku
Sakhalin Honshu
S. caecutiens
B. hypothetical phylogeny (Tree 2)
S. caecutiens S. shinto Cheju
Shikoku
Sakhalin ...... ••••• Honshu
o .... ~~.
Hokkaido
Fig. 5. Two hypothetical phylogenies of nuclear DNA in the Sorel' caeculiells/sitilllo group, based on the rDNA- RFLP rcpetypcs. Tree I (A) was inferred from the parsimonious network (Fig. 4A ) and tree 2 (B) from the network allowing hybridization (Fig. 4B). Arrows \vith broken lines indicate hybridi zation .
that S. caecutiens and S. shinto should be treated as t\vo separate species. In the present study, it was not stat is
tically supported, although topology showed these two were separated (Fig. 3). However, the shrew trom Cheju
Island was most close to S. caeculiens in the Eurasian
cont inent and it should be recognized as S. caecliliens
(Fig. 3). In addi tion, a lthough sampling locations from
the Eurasian continent were great ly scattered, there was
no variation in the rDNA-RFLP repetype within S. cae
cliliens over this range (Table I , Fig. 2). The same tendency was observed in the genetic variation of IlltDNA
cyt-b (Ohdachi et a l. 2001, 2003 ; Ohdachi 2005). These findings from both nuclear DNA and mtDNA suggest
that S. caecliliens has gone through a recent and rapid range expansion in the Eurasian cont inent. Patterns of
genetic variation were largely different between Hokkaido
and the continent both in rDNA-R FLP repetypes (Fig. 4)
and mtDNA haplotypes (Ohdachi et al. 200 1, 2003;
Ohdachi 2005). Therefore, the range expansion of S.
caeclfliens in the continent Illust have occurred after
Hokkaido and the continent were geologica lly separated
after the last glac ial period. It is estimated that Hokkaido and Sakha lin+ Eurasian continent were separated ca.
12 ,000 years ago (Ohshima 1990). Thus, the great
range expansion in the continent presumably began after
S. caecutiens
Various local ities in the Eurasian Continent and Sakhalin
S. shinto
Fig. 6. Schematic tree of the mitochondrial DNA phylogeny in the So/'ex caecutiens/shillto group, based on mtDNA cytochrom e b gene sequences (Ohdachi et al. 2003; Ohdachi in press). An open ci rcle is a hypothetical taxonomic unit.
12,000 years ago.
The maximum parsimony tree (Fig. 3) and the phylogenetic network (Fig. 4A) were used to construct a
hypothetical phylogeny of the nuclear rDNA (Tree I, Fig. SA). On the other hand, a llowing hybridization
between the rDNA-RFLPs, an a lternative network was
reconstructed (Fig. 4B), and from this phylogenetic net
work with hybridi zation, another hypothetical phylogeny
was inferred (Tree 2, Fig. 5B). We can not conclude which phylogeny (F ig. 5: trees I
and 2) is more appropriate as an evolutionary process of

Naitohetal.,RF乙PqプnuclearD八 「Ainshrei・vs
thenuclearrDNA.AmtDNAphylogenyoftheS.
eaecutiens/shintogroup,wasinferred(Fig.6)basedon
Ohdachietal.(2003)andOhdachi(2005),whereS.
eoeeutienswasdividedintoHokkaidoandContinent-
Sakhalin-Chej●ugenealogiesafterS.(3aecutiensandS.
shintodiverged.Hence,tree2ismorecongruentwith
themtDNAphylogenythantreelandfbrthisreason
seemsthemoreappropriatephylogenetictree(network)
ofthenuclearrDNA.
Acknowledgments:WewishtothankH.Abe,N.E.
Dokuchaev,V.Haukisalmi,M.Yamagishiforoffering
shrewsamples,G.Hintenforreviewingthemanuscript,
andK.Yoshizawafbrsupportinganalysisofthemaxi-
mumparsimonymethod.Wealsowouldliketoexpress
ourgratitudetoanonymousreviewersfbrcriticalcom-
ments.
References
Arheim,N.,Krystal,M,,Schmickel,R,,Wilson,G,,Ryder,0.and
Zimmer,E.1980.Molecularevidencefbrgeneticexchanges
amongribosomalgenesonnonhomologouschromosomesinman
andapes.ProceedingsoftheNationalAcademyofSciencesof
theUnitedStatesofAmerica73:7323-7327.
Bandelt,H.J.1994.Phylogeneticnetworks.Verhandlungendes
NaturwissenschaftlichenVereinsinHamburgN.F.34:51-71.
Dokuchaev,N,E,Ohdachi,S,andAbe,H,1999.Mo叩hometric
statusofshrewsoftheSorexcaecutiens/shintogroupinJapan.
MammalStudy24:67-78.
Fumagalli,L,Taberlet,P。,Stewart,D.T.,Gielly,LHausser,J.and
Vogel,P.1999.MolecularphylogenyandevolutionofSorex
shrews(Soricidae:Insectivora)inferredfrommitochondrialDNA
sequencedata,MolecularPhylogeneticsandEvolutionll:222-
235.
George,S。1988.Systematics,historicalbiogeography,andevolution
ofthegenusSorex。JournalofMammalogy69:443-461。
Iwasa,M。A。,Ohdachi,S.,Han,S.-H.,Oh,H.-S.,Abe,H。andSuzuki,
H.2001.KaryotypeandRFLPofthenuclearrDNAofthe
Croeidurasp,onChejuIsland,SouthKorea.Mammalia65:
451-459,
Iwasa,M.A.andSuzuki,H.2002.Evolutionarynetworksofmaternal
andpaternalgenelineagesinEothenomysvolesendemictoJapan.
JoumalofMammalogy83:852-865.
107
Mukai,T.2001.Hybridizationandintrogressioninthespeciation
processoffishes.JapaneseJoumalofIchthyology48:1-18(in
JapanesewithEnglishsummary).
Ohdachi,S.2005.Historyofcommunityorganizationofshrewsin
Hokkaido,inferredfromDNA.In(R.MasudaandH.Abe,eds。)
NaturalHistoryofAnimalBiogeography.HokkaidoUniversity
P「ess,SaPporo(inJapanese)(inpress).
Ohdachi,S.,Masuda,R.,Abe,H.,Adachi,J.,Dokuchaev,N.E.,
Haukisalmi,V,andYoshida,M,C,1997,PhylogenyofEurasian
soricineshrews(lnsectivora,Mammalia)inferredfromthemito-
chondrialcytochromebgenesequences.ZoologicalSciencel4:
527-532。
Ohdachi,S.,Dokuchaev,NE.,Hasegawa,M.andMasuda,R.2001.
Intraspecificphylogenyandgeographicvariationofsixspeciesof
northeasternAsiaticSorexshrewsbasedonthemitochondrial
cytochromebsequences.MolecularEcologylO:2199-2213.
Ohdachi,S。D.,Abe,H.andHan,S.-H。2003.Phylogeneticalposi-
tionsofSorexsp。(lnsectivora,Mammalia)fromCh(加Islandand
S.caecutiensfromtheKoreanPeninsula,inferredfrommitochon-
drialcytochromebgenesequences.ZoologicalScience20:91-
95.
Ohshima,K.1990.ThehistoryofstraitsaroundtheJapaneseIs-
landsinthelate-Quaternary.DaiyonkiKenkyu[TheQuatemary
Research]29:193-208(inJapanesewithEnglishabstract).
SambrookJ.,Fritsch,E.F.andManiatis,T.1989.MolecularCloning:
ALaboratoryManual,2nded.ColdSpringHarborLaboratory,
NewYork.
Sota,T.andVogler,A.P.2001.Incongruenceofmitochondrialand
nucleargenetreesinthecarabidbeetlesOhomoρterus.System-
aticBiology50:39-59。
Suzuki,H。1994.GeneticdiversityofribosomalDNA:phylogenetic
analysisofsmallmammals.HonyuruiKagaku[Mammalian
Science]34:67-79(inJapanesewithEnglishabstract),
Suzuki,H.,Tsuchiya,K.,Sakaizumi,M.,Wakana,S.,Gotoh,0.,
Saitou,N,Moriwaki,K.andSakurai,S.1990.Differentiation
ofrestrictionsitesinribosomalDNAinthegenusApodemus.
BiochemicalGenetics28:137-149.
Suzuki,H.,Kawamoto,Y.,Takenaka,0.,Munechika,1.,Hori,H。and
Sakurai,S.1994,PhylogeneticrelationshipsamongHomo
sapiensandrelatedspeciesbasedonrestrictionsitevariationin
rDNAspacers.BiochemicalGenetics32:257-269.
Swofford,D.L2000.PAUP*:phylogeneticanalysisusingparsi-
mony(*andothermethods).Version4.Sinauer,Sunderland,
Massachusetts.
Swofford,D,L.andOlsen,G,Ll990.Phylogenyreconstruction.In
(D.M.HillisandC.Moritz,eds.)MolecularSystematicsPp.
411-501.Sinauer,Sunderland,Massachusetts.
Received18January2005.Accepted4June2005.
Nai foh el al.. RFLP oillllclear DNA ill shrews
the nuclear rDNA. A mtDNA phylogeny of the S.
eaeel/liens/shinlo group, was inferred (Fig. 6) based on Ohdachi et a l. (2003) and Ohdachi (2005), where S.
caeclffiens was divided in to Hokkaido and ContinentSakhalin-Cheju genealogies after S. eaeel/liens and S.
shinto diverged. Hence, tree 2 is more congruent with the mtDNA phylogeny than tree I and for this reason seems the more appropriate phylogenetic tree (network) of the nuclear rDNA.
Acknowledgments : We wish to thank H. Abe, N. E. Dokuchaev, V. Haukisalmi , M. Yamagi shi for offering shrew samples, G. Hinten for reviewing the manuscript, and K. Yoshizawa for supporting ana lysis of the maximum parsimony method. We also would like to express our gratitude to anonymous reviewers for critical com
ments.
References
Arhcim, N., KrystaL M., Schmickcl , R .. Wil son, G., Ryder, O. and Zi mmer, E. 1980. Molecular ev idence for genetic exchanges among ribosomal genes on nonhomo logous chromosomes in man and apes. Proceedin gs of the National Academy of Sc iences of the United States o f America 73: 7323-7327.
Bandelt, H. J. 1994. Phy logenetic networks. Verhand lungen des Naturwissensehaftl ichen Vereins in Hamburg N. F. 34: 5 1- 7 1.
Dokudmcv, N. E., Ohdachi, S. amI Abe, H. 1999. Morphometric status of shrews of the Sorex caecllf;enslsiJinto group in Japan . Mammal SHldy 24: 67- 78.
FUl11agalli, L.. Taberlet. P .. Stewart. D. T .. Gidly. L. Hausser. J. and Voge l, P. 1999. Molecular phylogeny and evol ution of Sorex shrews (Soricidae: Insectivora) inferred from mitochondrial DNA sequence data . Molecular Phylogenetics and Evol ut ion II : 222-235 .
George, S. 1988. Systematics, hi storica l biogeography, and evo lution of tile genus Sorex. Journal of Mammalogy 69: 443-461.
Iwasa , M. A ., Ohdachi , S., Han, S.-H. , Oh , H.-S., Abe, H. and Suzu ki, H. 200 1. Karyotype and RFLP of the nu clear rONA of the Crocidum sp. on Cheju Is land. South Korea. Mammalia 65: 45 1-459.
Iwasa, M. A. and Suzuki , H. 2002. Evo lutionary networks of maternal and paternal gene lineages in Eorhenomys voles endemic to Japan. Journa l of Mamma logy 83: 852- 865.
107
Mukll i. T. 2001. Hybrid ization lind in trogression in the speciation process of fi shes . Japanese Journal of Ic hthyology 48 : \- 18 ( in Japanese with Eng lish summary).
Ohdachi , S. 2005 . Hi story of co mmunity organ ization o f shrews in Hokkaido, inferred from DNA. In (R. Masuda and H. Abe , eds.) Natural History of Anim al Biogeography. Hokkaido Uni versity Press, Sapporo (in JlIpanese) (in press).
Ohdachi, S., Masuda, R., Abe, 1-1 ., Adachi, J., Dokuchaev, N. E., Hauki sa lmi, V. and Yos hida, M. C. 1997. Phylogeny of Eurasian soric ine shrews ( Insecti vora, Mam malia) in fe rred from the mitochondrial cytochrome b gene sequences. Zoo log ical Sc ience 14: 527-532.
Ohdachi, S .. Dokuchaev, N . E. , Hasegawa, M. and Masuda, R. 2001. Intraspecific phy loge ny and geographi c variation of s ix spec ies o f northeastern Asiatic Sorex shrews based on the mitochondrial cytoc hrome h sequences . Molecular Ecology 10: 2 199-22 13.
Ohdachi. S. D., Abe, H. and Han, SAl. 2003. Phylogenetical positions of Sorex sp. (Insecti vora, Mammalia) from Chej u Is land and S. caeclIl;ellS from Ihe Korean Peni nsula. in ferred fro m mitochondria l cytoc hrome b gene seq uences. Zoo logical Sc ience 20: 9 1-95.
Ohsilima, K. 1990. The history of st rait s aro und th e Japanese Islands in the late·Quatemary. Daiyonki Kenkyu lThe Quaternary Research1 29: 193-208 (in Japanese with Eng lish abstract).
Sambrook J ., Fritsc h, E. F. and Man iati s, T. 1989. Molecul ar Clonin g: A Laboratory Manua l, 2nd ed . Co ld Spring Harbor Laboratory, New York.
Sota, T. and Vog ler, A. P. 2001. In congruence of mitochondri al and nuclear gene trees in th e carabid beetles Oholl/opferus. Systemat ic Bio logy 50: 39- 59,
Suzuki, H. 1994. Genetic diversity of ribosomal DNA: phylogenetic ana lysis of small mammals. Honyurui Kagaku [Mammali an Sc ience] 34: 67~79 (in Japanese with English abstract).
Suzuki, H., Tsuchiya, K., Sakai zumi , M., Wakana, S., Gotoh, 0., Saitou, N., Mori waki, K. and Sakurai , S. 1990. Differentiation of restri ction sites in r ibosoma l DNA in the gen us Apodellllls. Biochemica l Genetics 28: 137- 149.
Suzuki , H., Kawamoto, Y., Takenaka, 0 ., Mun echi ka, I., Hori, H. and Saku rai , S, 1994. Phyloge net ic relationships among Homo sapiens and reialeu spec ies based un rest ri ction site variation in rONA spacers. Biochemical Genetics 32: 257-269.
Swofford, D. L. 2000. PAUP*: phy logenetic analysis using parsimon y (* and olher met hods). Version 4, S inauer. S underl and. Massacllusetts.
Swofford, D. L. and Olsen , G. L. 1990. Phylogeny reconstruction. In (D. M. Hill is and C. Mori tz. eds.) Molecular Systematics Pp. 4 1\- 50 I. S inauer, Sunderland, Massachusetts .
Received 18 Jallt/lllY 2005. Accepted 4 JUlie 2005.
Nai foh el al.. RFLP oillllclear DNA ill shrews
the nuclear rDNA. A mtDNA phylogeny of the S.
eaeel/liens/shinlo group, was inferred (Fig. 6) based on Ohdachi et a l. (2003) and Ohdachi (2005), where S.
caeclffiens was divided in to Hokkaido and ContinentSakhalin-Cheju genealogies after S. eaeel/liens and S.
shinto diverged. Hence, tree 2 is more congruent with the mtDNA phylogeny than tree I and for this reason seems the more appropriate phylogenetic tree (network) of the nuclear rDNA.
Acknowledgments : We wish to thank H. Abe, N. E. Dokuchaev, V. Haukisalmi , M. Yamagi shi for offering shrew samples, G. Hinten for reviewing the manuscript, and K. Yoshizawa for supporting ana lysis of the maximum parsimony method. We also would like to express our gratitude to anonymous reviewers for critical com
ments.
References
Arhcim, N., KrystaL M., Schmickcl , R .. Wil son, G., Ryder, O. and Zi mmer, E. 1980. Molecular ev idence for genetic exchanges among ribosomal genes on nonhomo logous chromosomes in man and apes. Proceedin gs of the National Academy of Sc iences of the United States o f America 73: 7323-7327.
Bandelt, H. J. 1994. Phy logenetic networks. Verhand lungen des Naturwissensehaftl ichen Vereins in Hamburg N. F. 34: 5 1- 7 1.
Dokudmcv, N. E., Ohdachi, S. amI Abe, H. 1999. Morphometric status of shrews of the Sorex caecllf;enslsiJinto group in Japan . Mammal SHldy 24: 67- 78.
FUl11agalli, L.. Taberlet. P .. Stewart. D. T .. Gidly. L. Hausser. J. and Voge l, P. 1999. Molecular phylogeny and evol ution of Sorex shrews (Soricidae: Insectivora) inferred from mitochondrial DNA sequence data . Molecular Phylogenetics and Evol ut ion II : 222-235 .
George, S. 1988. Systematics, hi storica l biogeography, and evo lution of tile genus Sorex. Journal of Mammalogy 69: 443-461.
Iwasa , M. A ., Ohdachi , S., Han, S.-H. , Oh , H.-S., Abe, H. and Suzu ki, H. 200 1. Karyotype and RFLP of the nu clear rONA of the Crocidum sp. on Cheju Is land. South Korea. Mammalia 65: 45 1-459.
Iwasa, M. A. and Suzuki , H. 2002. Evo lutionary networks of maternal and paternal gene lineages in Eorhenomys voles endemic to Japan. Journa l of Mamma logy 83: 852- 865.
107
Mukll i. T. 2001. Hybrid ization lind in trogression in the speciation process of fi shes . Japanese Journal of Ic hthyology 48 : \- 18 ( in Japanese with Eng lish summary).
Ohdachi , S. 2005 . Hi story of co mmunity organ ization o f shrews in Hokkaido, inferred from DNA. In (R. Masuda and H. Abe , eds.) Natural History of Anim al Biogeography. Hokkaido Uni versity Press, Sapporo (in JlIpanese) (in press).
Ohdachi, S., Masuda, R., Abe, 1-1 ., Adachi, J., Dokuchaev, N. E., Hauki sa lmi, V. and Yos hida, M. C. 1997. Phylogeny of Eurasian soric ine shrews ( Insecti vora, Mam malia) in fe rred from the mitochondrial cytochrome b gene sequences. Zoo log ical Sc ience 14: 527-532.
Ohdachi, S .. Dokuchaev, N . E. , Hasegawa, M. and Masuda, R. 2001. Intraspecific phy loge ny and geographi c variation of s ix spec ies o f northeastern Asiatic Sorex shrews based on the mitochondrial cytoc hrome h sequences . Molecular Ecology 10: 2 199-22 13.
Ohdachi. S. D., Abe, H. and Han, SAl. 2003. Phylogenetical positions of Sorex sp. (Insecti vora, Mammalia) from Chej u Is land and S. caeclIl;ellS from Ihe Korean Peni nsula. in ferred fro m mitochondria l cytoc hrome b gene seq uences. Zoo logical Sc ience 20: 9 1-95.
Ohsilima, K. 1990. The history of st rait s aro und th e Japanese Islands in the late·Quatemary. Daiyonki Kenkyu lThe Quaternary Research1 29: 193-208 (in Japanese with Eng lish abstract).
Sambrook J ., Fritsc h, E. F. and Man iati s, T. 1989. Molecul ar Clonin g: A Laboratory Manua l, 2nd ed . Co ld Spring Harbor Laboratory, New York.
Sota, T. and Vog ler, A. P. 2001. In congruence of mitochondri al and nuclear gene trees in th e carabid beetles Oholl/opferus. Systemat ic Bio logy 50: 39- 59,
Suzuki, H. 1994. Genetic diversity of ribosomal DNA: phylogenetic ana lysis of small mammals. Honyurui Kagaku [Mammali an Sc ience] 34: 67~79 (in Japanese with English abstract).
Suzuki, H., Tsuchiya, K., Sakai zumi , M., Wakana, S., Gotoh, 0., Saitou, N., Mori waki, K. and Sakurai , S. 1990. Differentiation of restri ction sites in r ibosoma l DNA in the gen us Apodellllls. Biochemica l Genetics 28: 137- 149.
Suzuki , H., Kawamoto, Y., Takenaka, 0 ., Mun echi ka, I., Hori, H. and Saku rai , S, 1994. Phyloge net ic relationships among Homo sapiens and reialeu spec ies based un rest ri ction site variation in rONA spacers. Biochemical Genetics 32: 257-269.
Swofford, D. L. 2000. PAUP*: phy logenetic analysis using parsimon y (* and olher met hods). Version 4, S inauer. S underl and. Massacllusetts.
Swofford, D. L. and Olsen , G. L. 1990. Phylogeny reconstruction. In (D. M. Hill is and C. Mori tz. eds.) Molecular Systematics Pp. 4 1\- 50 I. S inauer, Sunderland, Massachusetts .
Received 18 Jallt/lllY 2005. Accepted 4 JUlie 2005.





























