Embed Size (px)
Citation preview

ORIGINAL PAPER
Interactions among mutualism, competition, and predation fosterspecies coexistence in diverse communities
Benedicte Bachelot & María Uriarte & Krista McGuire
Received: 23 December 2014 /Accepted: 2 January 2015# Springer Science+Business Media Dordrecht 2015
Abstract In natural systems, organisms are simultaneouslyengaged in mutualistic, competitive, and predatory interac-tions. Theory predicts that species persistence and communitystability are feasible when the beneficial effects of mutualismsare balanced by density-dependent negative feedbacks.Enemy-mediated negative feedbacks can foster plant speciescoexistence in diverse communities, but empirical evidenceremains mixed. Disparity between theoretical expectationsand empirical results may arise from the effects of mutualisticmycorrhizal fungi. Here, we build a multiprey species/predator model combined with a bidirectional resource ex-change system, which simulates mutualistic interactions be-tween plants and fungi. To reach population persistence, (1)the per capita rate of increase of all plant population mustexceed the sum of the negative per capita effects of predation,interspecific competition, and costs of mycorrhizal associa-tion, and (2) the per capita numerical response of enemies tomycorrhizal plants must exceed the magnitude of the percapita enemy rate of mortality. These conditions reflect thebalance between regulation and facilitation in the system. In-teractions between plant natural enemies and mycorrhizal fun-gi lead to shifts in the strength and direction of net mycorrhizaleffects on plants over time, with common plant species
deriving greater benefits from mycorrhizal associations thanrare plant species.
Keywords Prey–predatormodel . Foodweb dynamics .
Mutualism . Janzen-Connell . Mycorrhizal associations
Introduction
Interspecific interactions play a key role in driving populationdynamics (Gause and Witt 1935). A large number of theoret-ical and empirical studies have focused on understanding theeffects of competition and predation on population dynamics(Holling 1959; Rosenzweig and MacArthur 1963; Tilman1982; Chesson 2000; Chesson and Kuang 2008). More re-cently, studies have also investigated the role of mutualismsin isolation (Bronstein 2001a; Holland et al. 2002; Bever2003; Bruno et al. 2003). Two generalizations can be drawnfrom mutualism studies (Holland et al. 2002): mutualismsinvolve costs and benefits for both partners that are likelydensity-dependent (Roughgarden 1975; Addicott 1979; Mo-rales 2000; Bronstein 2001b), and population stability re-quires that the positive feedbacks derived from mutualismare balanced by negative feedbacks (Gause and Witt 1935;Vandermeer and Boucher 1978; Chesson 2000; Bever 2003).
Stabilizing negative feedbacks can arise from multiplemechanisms such as resource limitation, competition, preda-tion, or change in the net effects of mutualism (i.e., the balancebetween its costs and benefits) as population size increases(Holland et al. 2002; Schmitt and Holbrook 2003; Hollandand DeAngelis 2010; Holland et al. 2013). Recent theoreticalstudies have enhanced our understanding of the joint role ofmutualisms and negative feedbacks in stabilizing species in-teractions; however, they often remain limited to one or two
Electronic supplementary material The online version of this article(doi:10.1007/s12080-015-0251-2) contains supplementary material,which is available to authorized users.
B. Bachelot (*) :M. UriarteDepartment of Ecology, Evolution and Environmental Biology,Columbia University, 1200 Amsterdam Avenue, NewYork, NY 10027, USAe-mail: [email protected]
K. McGuireDepartment of Biology, Barnard College, Columbia University, NewYork, NY 10027, USA
Theor EcolDOI 10.1007/s12080-015-0251-2

aspects of interspecific interactions. In natural systems, organ-isms are simultaneously involved in mutualism, competition,and predation. Each process interacts with each other, limitingour ability to understand their joint effects from studies thatinvestigate them in isolation (Fontaine et al. 2011; Georgelinand Loeuille 2014). Therefore, it is important to combine mu-tualism and food web dynamics into our understanding ofcommunity dynamics and species coexistence (Rai et al.1983; Addicott and Freedman 1984; Freedman et al. 1987;Ringel et al. 1996; Jang 2002; Bronstein et al. 2003; Meliánet al. 2009; Loeuille 2010; Mougi and Kondoh 2012; Hollandet al. 2013; Georgelin and Loeuille 2014; Mougi and Kondoh2014). Recently, there has been a regained interest in combin-ing interaction types such as mutualism and antagonism mo-tivated by the recognition that indirect interactions can alterthe effects of mutualism and antagonism (Holland et al. 2013;Tang et al. 2014), affect resilience of dynamical systems toperturbations (Georgelin and Loeuille 2014), and promote sta-bility in complex systems (Mougi and Kondoh 2014), whichtend to be unstable when investigating interaction types inisolation (Allesina and Tang 2012).
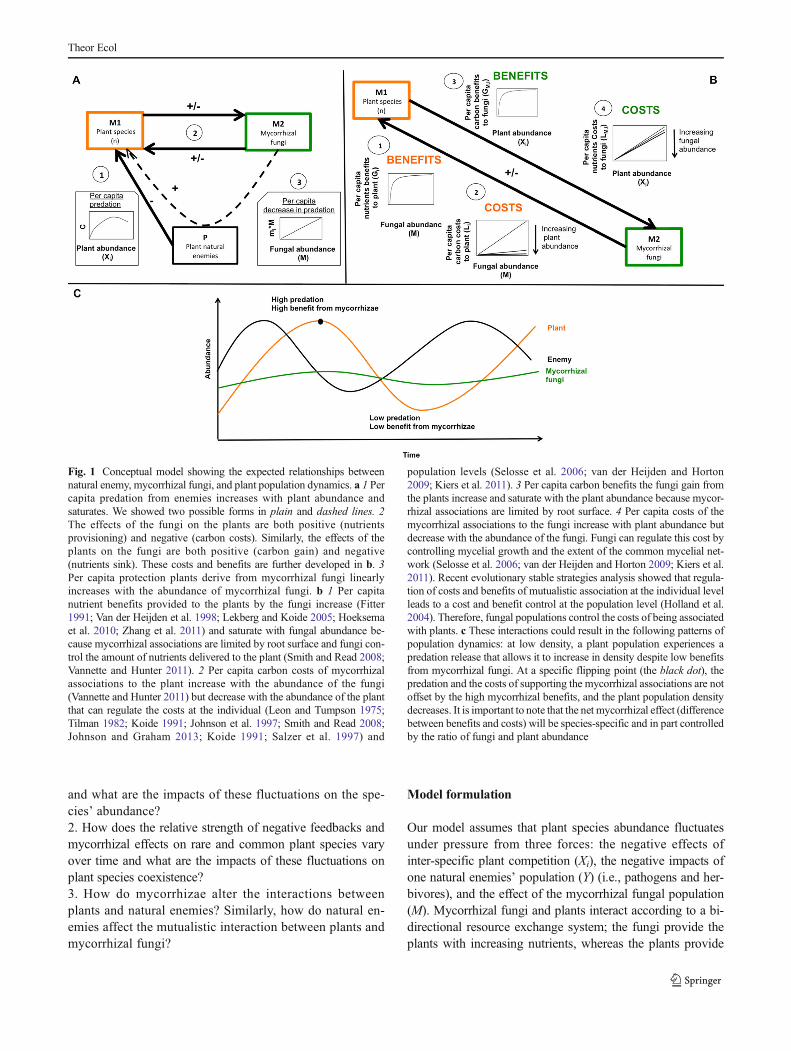
In this study, we investigate the joint role of predation,competition, and mutualistic associations with arbuscular my-corrhizal fungi for the dynamics of multiple plant species andfor species coexistence. In highly diverse plant communities,such as tropical forests, negative density-dependent factors(also known as Janzen-Connell (J-C) effects; Janzen 1970;Connell 1971; Connell et al. 1984) are the most frequentlystudied mechanisms that could explain the tree species coex-istence (reviewed inWright 2002). Janzen-Connell effects op-erate through the attraction of species-specific enemies such asseed predators, herbivores, or pathogens to the seedlings ofcanopy trees, which reduce conspecific seedling survivorshipnear the adult trees at high conspecific seedling density, leav-ing ecological space for heterospecific seedlings to recruit.The prediction from the J-C hypothesis is that the per capitapredation rate of a plant species increases with plant speciesabundance at local and community scales (Fig. 1a(1)).
In contrast to the large body of research on the importanceof negative feedbacks for species coexistence in plant com-munities and of positive feedbacks for plant invasion(Eppstein et al. 2006; Levine et al. 2006; Eppstein andMolofsky 2007), the role of fluctuating positive and negativemycorrhizal feedbacks on plant community dynamics remainsunderexplored both theoretically (Bruno et al. 2003) and em-pirically (Booth and Hoeksema 2010). Yet, theoretical studieshave shown the importance of fluctuating mutualisms in sta-bilizing interspecific interactions (Holland et al. 2013). Unlikesoil pathogens, which only have negative effects on their hosttrees, mycorrhizal fungi can have variable impacts on theirhost plant along a continuum from mutualism to parasitism(Fig. 1a(2), Johnson et al. 1997; Johnson and Graham 2013).If recent theoretical studies have incorporated costs and
benefits in analysis of mutualism (Neuhauser and Fargione2004; Holland and DeAngelis 2010; Holland et al. 2013),costs are usually lacking and mutualism is generally unidirec-tional in theoretical studies that combine mutualism and an-tagonism (Holland et al. 2013; Georgelin and Loeuille 2014;Mougi and Kondoh 2014).
The relative effect of mycorrhization on a plant is typi-cally gauged in terms of the quantity of limiting resourcesextracted from soil versus the amount of carbon needed tomaintain the symbiosis. Evidence for positive mycorrhizaleffects on host plants is extensive and has been attributed toa variety of mechanisms including greater plant nutrientuptake (Smith and Read 2008), defense against enemies(Gange and West 1994), and drought resistance (Augeet al. 1987). The nutrient benefits a plant species derivesfrom mycorrhizae increase with mycorrhizal fungal abun-dance as root colonization increases (Fitter 1991; Van derHeijden et al. 1998; Lekberg and Koide 2005; Hoeksemaet al. 2010; Zhang et al. 2011) and saturates when all avail-able roots have been colonized (Smith and Read 2008;Vannette and Hunter 2011; Fig. 1b(1)). Studies have alsouncovered negative feedbacks to plants from mycorrhizalfungal associations, which result from greater carbon costswith higher fungal abundance (Vannette and Hunter 2011).However, the cost of mycorrhizal associations to the plantsaturates with plant abundance (Leon and Tumpson 1975;Kiers et al. 2011; Fig. 1b(2)).
Additionally, mycorrhization has been repeatedly dem-onstrated to minimize the risk of infection by root patho-gens (Newsham et al. 1995) and to maximize response ef-ficiency of plant to natural enemies (Bi et al. 2007). Con-sequently, increased mycorrhizal colonization should becorrelated with decreased vulnerability to enemies, poten-tially leading to weaker negative enemy-mediated densitydependence and lower number of natural enemies(Fig. 1a(3)). Bi et al. (2007) provide an extensive reviewof the mechanisms underlying this effect, which includeincreased production of allelochemicals such as phenolsby the plants, induction of plant defense genes, activationof the jasmonate pathway that is crucial to respond to at-tacks by plant enemies, and attraction of predators of plantnatural enemies (Hoffman et al. 2011a, b).
Here, we present a mathematical model to explore thejoint effects of negative enemy and mycorrhizal fungalfeedbacks on plant community dynamics and species coex-istence. We then interpret the results of our model in light ofexisting empirical work on several aspects of the feedbackswe examine. Specifically, we address the followingquestions:
1. How does the relative strength of negative feedbacksfrom natural enemies and the magnitude and direction ofmycorrhizal effects vary over time within a plant species
Theor Ecol
and what are the impacts of these fluctuations on the spe-cies’ abundance?2. How does the relative strength of negative feedbacks andmycorrhizal effects on rare and common plant species varyover time and what are the impacts of these fluctuations onplant species coexistence?3. How do mycorrhizae alter the interactions betweenplants and natural enemies? Similarly, how do natural en-emies affect the mutualistic interaction between plants andmycorrhizal fungi?
Model formulation
Our model assumes that plant species abundance fluctuatesunder pressure from three forces: the negative effects ofinter-specific plant competition (Xi), the negative impacts ofone natural enemies’ population (Y) (i.e., pathogens and her-bivores), and the effect of the mycorrhizal fungal population(M). Mycorrhizal fungi and plants interact according to a bi-directional resource exchange system; the fungi provide theplants with increasing nutrients, whereas the plants provide
Fig. 1 Conceptual model showing the expected relationships betweennatural enemy, mycorrhizal fungi, and plant population dynamics. a 1 Percapita predation from enemies increases with plant abundance andsaturates. We showed two possible forms in plain and dashed lines. 2The effects of the fungi on the plants are both positive (nutrientsprovisioning) and negative (carbon costs). Similarly, the effects of theplants on the fungi are both positive (carbon gain) and negative(nutrients sink). These costs and benefits are further developed in b. 3Per capita protection plants derive from mycorrhizal fungi linearlyincreases with the abundance of mycorrhizal fungi. b 1 Per capitanutrient benefits provided to the plants by the fungi increase (Fitter1991; Van der Heijden et al. 1998; Lekberg and Koide 2005; Hoeksemaet al. 2010; Zhang et al. 2011) and saturate with fungal abundance be-cause mycorrhizal associations are limited by root surface and fungi con-trol the amount of nutrients delivered to the plant (Smith and Read 2008;Vannette and Hunter 2011). 2 Per capita carbon costs of mycorrhizalassociations to the plant increase with the abundance of the fungi(Vannette and Hunter 2011) but decrease with the abundance of the plantthat can regulate the costs at the individual (Leon and Tumpson 1975;Tilman 1982; Koide 1991; Johnson et al. 1997; Smith and Read 2008;Johnson and Graham 2013; Koide 1991; Salzer et al. 1997) and
population levels (Selosse et al. 2006; van der Heijden and Horton2009; Kiers et al. 2011). 3 Per capita carbon benefits the fungi gain fromthe plants increase and saturate with the plant abundance because mycor-rhizal associations are limited by root surface. 4 Per capita costs of themycorrhizal associations to the fungi increase with plant abundance butdecrease with the abundance of the fungi. Fungi can regulate this cost bycontrolling mycelial growth and the extent of the common mycelial net-work (Selosse et al. 2006; van der Heijden and Horton 2009; Kiers et al.2011). Recent evolutionary stable strategies analysis showed that regula-tion of costs and benefits of mutualistic association at the individual levelleads to a cost and benefit control at the population level (Holland et al.2004). Therefore, fungal populations control the costs of being associatedwith plants. c These interactions could result in the following patterns ofpopulation dynamics: at low density, a plant population experiences apredation release that allows it to increase in density despite low benefitsfrom mycorrhizal fungi. At a specific flipping point (the black dot), thepredation and the costs of supporting the mycorrhizal associations are notoffset by the high mycorrhizal benefits, and the plant population densitydecreases. It is important to note that the net mycorrhizal effect (differencebetween benefits and costs) will be species-specific and in part controlledby the ratio of fungi and plant abundance
Theor Ecol

the fungi with carbon (Smith and Read 2008). Furthermore,mycorrhizal colonization lowers predation from plant enemiesby increasing plant defenses (Pozo and Azcón-Aguilar 2007;Bi et al. 2007). To understand the dynamics that would resultfrom these complex enemy–plant–fungal interactions, we
couple a multispecies, continuous, nonlinear prey–predatormodel (enemies-plant) with a bidirectional resource exchangemodel to represent the interactions between plants and fungi.The dynamics of plant species i (Xi) at time t are described bythe following equation:
dX i tð Þdt
¼ X i tð Þ bi þ αiGi RM→X i ; tð Þ−βiLi RX i→M ; tð Þ− Ci X ; tð Þ−miM tð Þ½ �Y tð Þ−Xnj¼1
ai; jX j tð Þ" #
ð1Þ
where Xi(t) and Y(t) represent population abundance of plantspecies i (prey) and enemies, respectively, bi represents the percapita exponential rate of increase of plant species i, ai,j des-ignates the per capita interspecific competition among plantspecies, ai,i represents intraspecific competition (self-limita-tion), M(t) is the abundance of the mycorrhizal fungal popu-lation, XiCi(X,t) is the functional response of natural enemiesto plant species i, and therefore Ci(X,t) is the per capita rate ofmortality from one enemy (Fig. 1a(1)). We used a classicalHolling type III functional response (Holling 1959; Smoutet al. 2010):
X iCi X ; tð Þ ¼ X i tð Þ ciX i tð Þ1þ
Xjw jc jX j tð Þ2
; withallci andwi > 0
ð2Þ
where wi is the handling time of plant species i, and ci is theencounter rate of natural enemy with plant species i. Thisenables us to evaluate natural enemy behavior that causesdensity-dependent mortality (Janzen 1970; Connell 1971;Murdoch 1975; Pacala and Crawley 1992; Mordecai 2011),while acknowledging that natural enemies might satiate athigh plant density (Silvertown 1980). Furthermore, it intro-duces the notion that natural enemy behavior might not beinfluenced by very low plant density (Real 1977; Krebs1974; Jeschke et al. 2002). In a diverse community, very rareplant species might escape predation because natural enemiesmight not find them (Jaenike 1990; Pacala and Crawley 1992;Hierro et al. 2005) or not target them due to change in diet/hostbreadth with plant availability (Fox and Morrow 1981; Ste-phens and Krebs 1986). Therefore, a Holling type III func-tional response is the best choice to investigate the joint effectof natural enemies and mycorrhizal fungi on plant dynamics.Although we will present results for a type III response here,more general assumptions about functional responses yieldsimilar qualitative results (see Appendix 1). The term mi×M(t) in Eq. 1 modifies the functional response of the enemybecause mycorrhizal fungi provide plants with increased per
capita protection (mi) against enemies (Fig. 1a(3); Gange andWest 1994; Pozo and Azcón-Aguilar 2007; Bi et al. 2007). Inorder to prevent that a net positive effect arises from the inter-actions between natural enemies and fungi, we bounded themycorrhizal fungal abundance so that Ci(X,t)−miM(t)≥0. Fi-nally, the plant nutritionally benefits (Fig. 1a(2), b(1),αiGi RM→X i ; tð Þ) and pays a carbon cost (Fig. 1a(2), b(2),βiLi RX i→M ; tð Þ) from the mycorrhizal associations (Smithand Read 2008; Johnson and Graham 2013). The functionsGi(RM−>Xi, t) represent the per capita increase in resourceplants’ gain from the mycorrhizal associations (Fig. 1b(1);Smith and Read 2008), whereas Li(RXi−>M, t) represents theper capita carbon losses to the associations (Fig. 1b(2); Olssonet al. 2010). The parameter αi represents the conversion of thegain in a unit of resource per capita to a gain in abundance ofthe population of the plant species i. In contrast, βi stands forthe conversion of per capita loss in carbon to a loss in abun-dance of the population of the plant species i. We use the termnet mycorrhizal effect for the plant species i to denote thedifference between the benefits (nutrition and protection fromenemies) and the costs (carbon): αi Gi(RM−>Xi, t)+miM(t)Y(t)and βi Li(RXi−>M, t).
The dynamics of the mycorrhizal fungal population(M) directly depend on the balance between the carbongains and nutrient losses they derive from plant species1 to n (Koide and Elliott 1989; Fitter 1991). In an effort toincrease model tractability, we did not account for differ-ent mycorrhizal fungal species. Growing evidence hasshown that the specific community of mycorrhizal fungiassociated with a plant rather than the identity of individ-ual mycorrhizal fungi controls the outcomes of the asso-ciations (Johnson and Graham 2013; Toju et al. 2013).Therefore, considering the mycorrhizal fungal communityrather than individual mycorrhizal fungal species does notdramatically hinder the realism of our model. The dynam-ics of the mycorrhizal fungal population are described bya consumer–resource (C-R) model (Rosenzweig andMacArthur 1963) similar to the bidirectional C-R modelproposed by Holland and DeAngelis (2010) as follows(Fig. 1a(2)):
Theor Ecol

dM tð Þdt
¼ M tð ÞXni¼1
αM ;iGM ;i RX i→M ; tð Þ−βM ;iLM ;i RM→X i ; tð Þ� �−rM
!ð3Þ
where GM,i (RXi−>M, t) and LM,i (RM−>Xi, t), respectively, rep-resent the per capita resource gains and losses of the fungalpopulation to plant species i, αM,i represents the conversion ofthe gain in resource from plant species i to fungal populationabundance (Smith and Read 2008), and βM,imodulates the percapita loss in resource to plant species i in loss in fungalabundance (respectively Fig. 1b(3, 4), and rM is the per capitaexponential rate of mortality of the mycorrhizal fungi in theabsence of plants.
Studies have shown that the effects of mycorrhizal associ-ations are context-specific, and the same mycorrhizal fungalspecies could have positive, negative, and neutral effects(Smith and Read 2008; Johnson and Graham 2013; Tojuet al. 2013). To account for this level of variation in the effectof mycorrhizal fungi, we use plant species-specific resourcegain and loss functions (G and L in Eqs. 1 and 3) and plantspecies-specific numerical responses (α and β in Eqs. 1 and3). The mycorrhizal fungi are strictly obligate and cannot sur-vive without the plants (Smith and Read 2008). Therefore, thedynamics of the fungi are solely controlled by the exchange ofresources with the plants.
Quantifying the costs and benefits plants derive frommycorrhizal associations and vice versa remains challeng-ing (Johnson et al. 1997; Johnson and Graham 2013). In ourmodel, we used classical resource exchange functions suchas the one described by Holland and DeAngelis (2010)(Fig. 1b(1, 2)):
Gi RM→X i ; tð Þ ¼ giM tð Þhi þM tð Þ ð4Þ
Li RX i→M ; tð Þ ¼ liM tð Þki þ X i tð Þ ð5Þ
In parallel,GM,i and LM,i for the fungi in Eq. 3 take the form(Fig. 1b(3, 4)):
GM ;i RX i→M ; tð Þ ¼ gM ;iX i tð ÞhM ;i þ X i tð Þ ð6Þ
LM ;i RM→X i ; tð Þ ¼ lM ;iX i tð ÞkM ;i þM tð Þ ð7Þ
where gi and gM,i are the saturation levels of resource gains,and hi and hM,i are the half saturation constants for plant
species i and for fungal populationM, respectively. Similarly,li and lM,i are the saturation levels of the loss of resources, andki and kM,i are the half saturation constants. Parameters g and lrepresent the per capita interaction strength of one species onthe other. We assumed per capita resource gain to plant speciesi (the Gi function in Eq. 1) a saturating (Michaelis-Menten)function of the abundance of fungi associated with each plantspecies i=1 to n (Fig. 1b(1)). Similarly, benefits to the mycor-rhizal fungi (the GM,i function in Eq. 3) are described byMichaelis-Menten functions of the plant abundance associat-ed with the fungal population (Fig. 1b(3)). This functionalform captures the assumption that the benefits extracted byfungi and plants are limited by the extent of fine root surfacearea and that the two symbiotic partners (i.e., plants and fungi)can control these benefits. Per capita resources lost by plantspecies i (the Li function in Eq. 1, Fig. 1b(2)) increase linearlywith increasing fungal populations (Eq. 2), while resourceslost by the fungi (the LM,i functions in Eq. 3) increase linearlywith greater abundance of plant species i population (Eq. 7,Fig. 1b(4)). The per capita loss of resources functions saturateswith the supplier species abundance (denominators in Eqs. 5and 7, Fig. 1b(2, 4)). In Eq. 5 and Fig. 1b(2), the rationale forthe denominator is that plants have the potential to controlfungal colonization to maximize the benefits of the mycorrhi-zal associations while minimizing the costs (Koide 1991;Johnson et al. 1997; Smith and Read 2008; Johnson and Gra-ham 2013). Similarly, the rational for the denominator in Eq. 7and Fig. 1b(4) is that fungi are able to overcome plant controlsof the colonization (Johnson et al. 1997; Smith and Read2008; Johnson and Graham. 2013). In other words, plantsand fungi can control the costs of the mycorrhizal associationsat the individual level and thus at the population level due toevolutionary stable strategies of mutualism (Holland et al.2004). Another motivation behind the denominator of the costfunction arises from the fact that plants and mycorrhizal fungiare connected via common mycelial network, allowing themto share the costs of the mycorrhizal association with eachother (Simard and Durall 2004). We acknowledge that theform we are using to represent the mycorrhizal effect is asimplification because we do not know the exact forms ofper capita resource gains (theGi andGM,i functions) and losses(the Li and LM,i functions). These simple forms of consumer–resource interactions have been shown to be accurate in somebiological systems (Holland 2002; Holland et al. 2002; Hol-land and DeAngelis 2006; Holland and DeAngelis 2010; Hol-land et al. 2013).
Finally, enemy dynamics (Y) are described as one popula-tion of enemies exhibiting host preferences rather than spe-cialization. We assume that predation increases with plantabundance (Fig. 1a(1)). The assumption of host preferencescan lead to the simulation of inappropriate population dynam-ics for the few plants that have species-specific interactionswith specialist pathogens and herbivores. However, in these
Theor Ecol

associations between plants and specialist enemies, plantabundance dynamics are likely to be tightly dependent onthe dynamics of their own specialist enemies and less depen-dent on other plant species present in the community. There-fore, ignoring the distinction between host-specific and gen-eralist interactions in the system should not affect the validityof our conclusions especially given the low rates of host spe-cialization of natural enemies in the tropics (Novotny et al.2010). The dynamics of plant natural enemies are a function ofplant species and mycorrhizal fungal abundances as follows:
dY tð Þdt
¼ Y tð ÞXni¼1
di Ci X ; tð Þ−miM tð Þð ÞX i tð Þ½ �−eY tð Þ−r" #
ð8Þ
where di Ci (X,t)Xi(t) represents the per capita numerical re-sponse of enemies to plant i, mi the per capita protectionagainst enemies provided by the mycorrhizal associationswith plant species i (Fig. 1a(3)), M the abundance of the my-corrhizal fungal population, e is the per capita competitioncoefficient for the enemy species (self-limitation), and r theper capita exponential rate of mortality of the enemy species inthe absence of plant (i.e., starvation rate). Therefore, the my-corrhizal fungi have two effects on the plant natural enemypopulation (Wootton 1994; Strauss and Irwin 2004;Koricheva et al. 2009; Bardgett and Wardle 2010; Barberet al. 2012): An indirect effect that arises frommodifying plantpopulation growth (depicted in the resource exchange func-tions,G and L), and a direct effect that emerges from change innatural enemy functional response due to increased plant pro-tection (−miM).
We chose to co-model community dynamics in continuoustime, which is more suitable to model populations of organ-isms with short life spans (natural enemies and mycorrhizalfungi) and very long-lived ones (trees, Wangersky 1978). Thefinal system is a nonlinear prey–predator model that combinescompetition among plant species, predation, and potential fa-cilitation by mycorrhizal fungi if the overall net mycorrhizaleffect is positive. This last effect is a fluctuation-dependentmechanism (Chesson 2000). Over time, as plant abundanceincreases, the facilitation effect increases until a flipping pointafter which this facilitation effect levels off, eventually becom-ing a negative density-dependent effect driven by a growingenemy population and the carbon and other costs associatedwith the mycorrhizal symbiosis (Fig. 1c). The formulation ofthe net mycorrhizal effect via bidirectional resource/consumersystem (Johnson et al. 1997; Holland and DeAngelis 2010)enables this effect to become negative and introduces an ad-ditional negative covariance between fitness and density sim-ilar to the one produced by the effect of predation (Chesson2000). Our model is one of the first attempts to combine
mutualistic interactions and food web dynamics. Hollandet al. (2013) used resource exchange functions to investigatethe stability of a system that included one plant species, onemutualistic pollinator, and one nectar-parasite. In their system,the mutualistic pollinator gains benefits from the plant withoutpaying a cost, and the nectar-parasite does not alter the behav-ior of the mutualistic pollinator (unidirectional mutualism).Our study expands on Holland et al. system (2013) and aimsto explore community dynamics and coexistence of n plantspecies that are involved in bidirectional exchange with my-corrhizal fungi, where each plant and fungus experiences ben-efits and costs. Furthermore, natural enemies whose functionalresponse is affected by mycorrhizae target the n plant species.
To evaluate the system, we assume that the mycorrhizalfungal population does not tend towards infinity but ratherexhibits an upper bound (v). We defined this upper bound toensure that the protection conveyed bymycorrhizal fungi doesnot result in a positive effect of natural enemies on plant pop-
ulation (for all plant species i, mi≤Cli
max M tð Þð Þ defCliv ). In other
words, the mycorrhizal fungi might enhance plant protectionagainst natural enemies, but this enhancement will not resultin a total protection. If the mycorrhizal fungal population goesto extinction, then our model becomes a classical multispeciesprey–predator type system. Therefore, in our work, we inves-tigate the conditions to maintain plant species coexistencewhen the mycorrhizal fungal population remains between astrictly positive lower bound ( ) and an upper bound (v). Themodel was evaluated for n plant species (Appendix 1). Spe-cifically, we analyzed the model to find conditions allowingthe populations to remain between a positive lower abundanceboundary and a finite upper abundance boundary. In otherwords, we evaluate the conditions enabling the coexistenceof n plant species in the system. We also analyzed a simpleversion of the model involving one plant species and a fixedmycorrhizal fungal population in order to understand howmycorrhizal fungi alter the prey–predator dynamics(Appendix 2). Similarly, we investigated the case of a fixednatural enemy population to understand its effect on the mu-tualistic interactions (Appendix 2). We illustrated the impor-tant results of the model using simulation to represent fluctu-ations in abundance over time within one plant species and fortwo coexisting plant species: one rare and one common. Inthis paper, the distinction between a rare and a common plantspecies is a relative notion based upon the relative density ofeach plant species. Finally, we investigated the dynamics ofone equilibrium by using simulations and changing the valueof each parameter one by one (Tables 2 and 3 in Appendix 3).Specifically, we wanted to assess how small changes in eachparameter of the model with two plant populations wouldchange the dynamics of the equilibrium (stable, periodic, notperiodic, or unstable). All simulations were accomplishedusing Mathematica v. 7.0. (Wolfram Research, Inc. 2008).
Theor Ecol
Results and discussion
Variability of the relative strength of negative feedbacksand net mycorrhizal effect on plant species
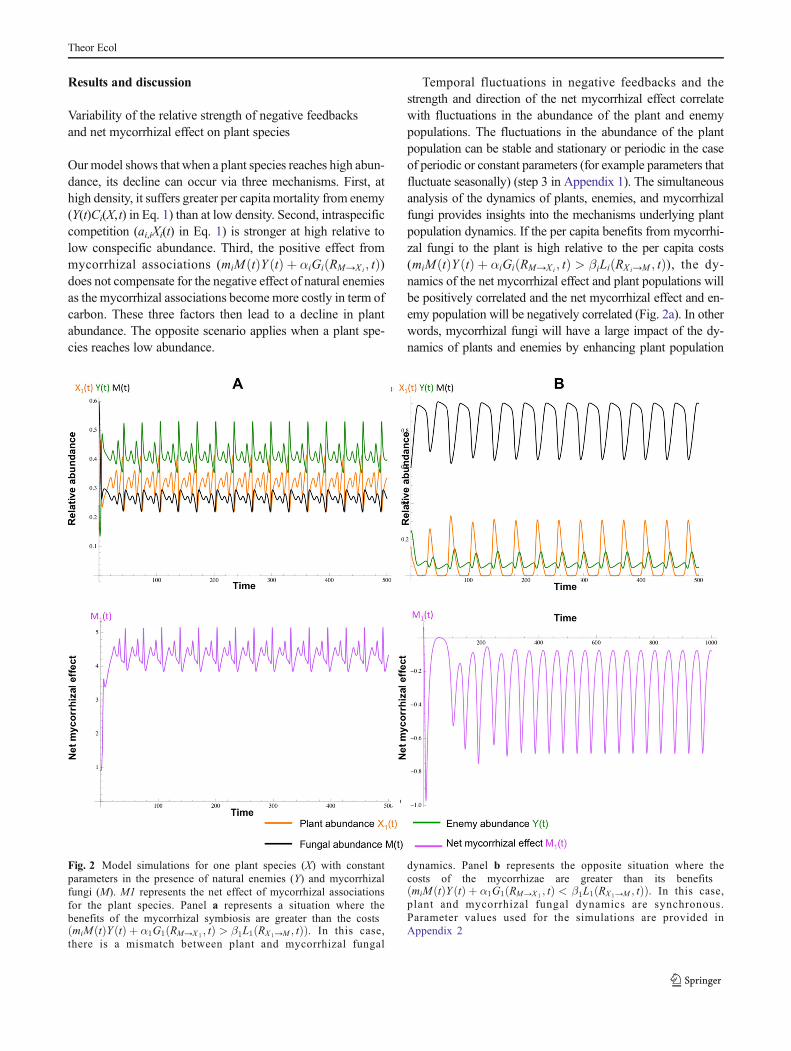
Our model shows that when a plant species reaches high abun-dance, its decline can occur via three mechanisms. First, athigh density, it suffers greater per capita mortality from enemy(Y(t)Ci(X,t) in Eq. 1) than at low density. Second, intraspecificcompetition (ai,iXi(t) in Eq. 1) is stronger at high relative tolow conspecific abundance. Third, the positive effect frommycorrhizal associations (miM tð ÞY tð Þ þ αiGi RM→X i ; tð Þ)does not compensate for the negative effect of natural enemiesas the mycorrhizal associations become more costly in term ofcarbon. These three factors then lead to a decline in plantabundance. The opposite scenario applies when a plant spe-cies reaches low abundance.
Temporal fluctuations in negative feedbacks and thestrength and direction of the net mycorrhizal effect correlatewith fluctuations in the abundance of the plant and enemypopulations. The fluctuations in the abundance of the plantpopulation can be stable and stationary or periodic in the caseof periodic or constant parameters (for example parameters thatfluctuate seasonally) (step 3 in Appendix 1). The simultaneousanalysis of the dynamics of plants, enemies, and mycorrhizalfungi provides insights into the mechanisms underlying plantpopulation dynamics. If the per capita benefits from mycorrhi-zal fungi to the plant is high relative to the per capita costs(miM tð ÞY tð Þ þ αiGi RM→X i ; tð Þ > βiLi RX i→M ; tð Þ), the dy-namics of the net mycorrhizal effect and plant populations willbe positively correlated and the net mycorrhizal effect and en-emy population will be negatively correlated (Fig. 2a). In otherwords, mycorrhizal fungi will have a large impact of the dy-namics of plants and enemies by enhancing plant population
Fig. 2 Model simulations for one plant species (X) with constantparameters in the presence of natural enemies (Y) and mycorrhizalfungi (M). M1 represents the net effect of mycorrhizal associationsfor the plant species. Panel a represents a situation where thebenefits of the mycorrhizal symbiosis are greater than the costsmiM tð ÞY tð Þ þ α1G1 RM→X 1
; tð Þ > β1L1 RX 1→M ; tð Þð Þ. In this case,there is a mismatch between plant and mycorrhizal fungal
dynamics. Panel b represents the opposite situation where thecosts of the mycorrhizae are greater than its benefitsmiM tð ÞY tð Þ þ α1G1 RM→X 1
; tð Þ < β1L1 RX 1→M ; tð Þð Þ. In this case,plant and mycorrhizal fungal dynamics are synchronous.Parameter values used for the simulations are provided inAppendix 2
Theor Ecol

growth and impeding predation from enemies. Conversely, ifthe per capita benefits from mycorrhizal associations are lowerthan its per capita costs for both fungi and plant populations( miM tð ÞY tð Þ þ αiGi RM→X i ; tð Þ < βiLi RX i→M ; tð Þ andmiMtð ÞY tð Þ þαM ;iGM ;i RX i→M ; tð Þ < βM ;iLM ;i RM→X i ; tð Þ) , t h e
net mycorrhizal effect to the plants and the plant populationsize will remain positively correlated, but the net mycorrhizaleffect to the plants and the enemy population might becomepositively correlated (Fig. 2b). In other words, the mycorrhizalfungi will not be beneficial to the plants but rather they mightindirectly enhance the enemy population. In other words, thiscould occur if by enhancing plant growth, mycorrhizal fungiindirectly benefit plant enemies. If the net mycorrhizal effectfluctuates between positive and negative (The sign of miM tð ÞY tð Þ þ αiGi RM→X i ; tð Þ−βiLi RX i→M ; tð Þ will change over timedue to fluctuating plant and fungal populations, similar to theparasitism/mutualism continuum proposed by Johnson andGraham 2013), the dynamics will exhibit a variety of patternsranging from positive to negative correlations between the net
mycorrhizal effect and the natural enemy population size(Fig. 3).
Our model shows that the joint impacts of enemies andmycorrhizal fungi can lead to temporal fluctuations in theabundance of a single plant species. The impacts of thesecombined feedbacks on plant dynamics have received scantattention (Van der Putten et al. 2009; Bardgett and Wardle2010). While these interactions are complex and likely to bespecies-specific (Wardle 2002), evidence from herbivory stud-ies suggests that mycorrhizal fungi and plant natural enemiesare interacting (Bi et al. 2007). The various dynamic patternsobtained with our model suggest that the interactions betweenplant natural enemies and mycorrhizal fungi are often but notalways negative. In support of our findings, a number of em-pirical studies have uncovered negative correlations betweenherbivory and the abundance, phenology, and colonization ofmycorrhizal fungi, lending support to the idea that top-downeffects can modulate the strength and direction of mycorrhizaleffects on plants. For example, Gehring and Whitham (1994)
Fig. 3 Model simulations for one plant species (X) in the presence ofnatural enemies (Y) and mycorrhizal fungi (M) with constant parameters.M1 represents the net effect of mycorrhizal associations for the plantspecies. Panels a and b illustrate two situations where over time the net
effect of mycorrhizal associations is fluctuating between positive andnegative. Parameter values used for the simulations are provided inAppendix 2
Theor Ecol

found a decrease in mycorrhizal colonization following defo-liation for 23 of 37 studied plants. Recent reviews, however,highlight how the complex interaction between plant enemiesand mycorrhizal fungi can vary with the frequency, timing,and type of herbivory (Klironomos et al. 2004; Saravesiet al. 2008).
Impact of negative feedbacks relative strength and netmycorrhizal effect fluctuations on species abundanceand coexistence
Our model predicts coexistence of n plant species under thejoint effects of mycorrhizal fungi, plant natural enemies, andplant competition, which together establish lower and upperabundance constraints, offering an additional mechanism toexplain the high diversity of plant species in diverse commu-nities such as tropical forests. This result is the first theoreticalevidence showing that the joint effects of plant natural ene-mies and mycorrhizal fungi can lead to plant species coexis-tence. This result is consistent with theoretical work showingthat mutualism alone cannot lead to species coexistence(McGill 2005) unless mutualistic populations are regulatedby other mechanisms (e.g., plant natural enemies in our mod-el, Simonsen and Stinchcombe 2014).
Our model predicts that each plant species will fluctuatebetween low and high abundance constraints that depend onthe magnitude of negative feedbacks from natural enemies,plant, and mycorrhizal fungi. In order to reach coexistence,the parameters of the model must satisfy three conditions (Ap-pendix 1). First, the per capita exponential rate of growth ofthe plant population i (bi) must be greater than the sum of (1)the negative per capita effects due to predation (Ci
uq, where qis the upper bound of enemy population and Ci
u is themaximum value of the per capita mortality of plant species ifrom enemy, see Appendix 1), (2) the per capita interspecific
competition ( ∑n
j¼1; j≠iai; jp j
h i, where pj is the upper limit of plant
species j population size), and (3) the per capita costs of my-
corrhizal associations (βilivki, where v is the upper bound of
mycorrhizal fungal population) (step 2), that is:
bi > Cui qþ
Xnj¼1; j≠i
ai; jp j
h iþ βi
liv
kið9Þ
This first condition ensures that plant populations avoidextinction. Second, the persistence of plant species requiresthat the enemy population does not become overdominant andthat this upper limit of natural enemy population is biological-ly reachable (in other words, it is not an infinite or a negativenumber). This condition requires that the per capita enemyexponential rate of mortality (r) is strictly lower than the total
of the maximum value of the per capita numerical responses
of enemies to plants (∑n
i¼1diC
ui pi) (step 1 in Appendix 1):
r <Xni¼1
diCui pi ð10Þ
This condition ensures the existence of an upper bound (q)for the enemy population that depends only on the per capita
enemy numerical responses (∑n
i¼1diC
ui pi), the per capita com-
petition among enemies (e), and the per capita enemy expo-nential rate of mortality (r), as follows:
Y tð Þ≤X n
i¼1diC
ui pi−r
edefq ð11Þ
A third condition required to prevent enemy populationextinction is that the total of the minimum value of the per
capita numerical responses of enemies to plants (∑n
i¼1diC
liepi� �
;
where epi is the lower limit of plant species i population size)
protected by mycorrhizal fungi (∑n
i¼1−dimivpið Þ; where v is the
upper bound of mycorrhizal fungal population) must be great-er than the per capita enemy exponential rate of mortality (r)(step 2 in Appendix 1):
Xni¼1
diCliepi−dimivpi
� �> r ð12Þ
If this condition is met (Eq. 12), then the condition to pre-vent enemies from becoming overdominant (Eq. 10) is alsomet. Thus, we found that two necessary conditions must bemet to maintain species coexistence: (1) the per capita expo-nential rate of growth of the plant population must be greaterthan the sum of the negative effects due to per capita preda-tion, per capita interspecific competition, and the per capitacosts of mycorrhizal associations, and (2) the total of the min-imum value of the per capita numerical responses of enemiesto plants protected by mycorrhizal fungi must be greater thanthe per capita enemy exponential rate of mortality. In otherwords, the protection conveyed by the mycorrhizal associa-tion is not strong enough to dramatically decrease the enemypopulation (food web cascade; Bruno et al. 2003; Cantrell andCosner 2001; McGill 2005). Overall, these two conditionsreflect the balance between regulation and facilitation in thesystem. More empirical and experimental work is required toevaluate the realism of these two conditions.
Theor Ecol

If the two conditions described in inequalities Eqs. 9 and 12are met, the abundance of plant species i (Xi) will fluctuatebetween the following upper (pi) and lower (epi) limits (seeAppendix 1 for more details):
epidef bi−βiliv
ki−X n
j¼1; j≠iai; jp j
h i−Cu
i q
ai;i≤X i≤
bi þ αigiai;i
defpi
ð13Þ
The parameters q and v represent the upper boundary ofenemy and mycorrhizal fungal abundance, respectively (Ap-pendix 1). The upper limit of plant abundance depends on theper capita benefits to the plant species i (αigi) associated withthe mycorrhizal associations and the per capita exponentialrate of increase of plant species i (bi), whereas the lowerboundary is a function of the maximum per capita costs of
mycorrhizal associations to the plant species i (βilivki), the per
capita competition among plants (aij), and the maximum percapita predation effect (Ci
u). Different parameters will lead todifferent model outputs: If the per capita benefits gained viamycorrhizal fungi are lower in rare relative to common plant
species (αrgr<αcgc) as expected in light of empirical work(Moora et al. 2004), given similar per capita intraspecific com-petition (ai,i) and per capita growth rate (bi), the maximumabundance of a rare plant species (pr) will remain lower thanthat of a common plant species (pc):
pr ¼br þ αrgr
ar;r≤bc þ αcgc
ac;c¼ pc ð14Þ
Therefore, inequality Eq. 14 predicts that lower frequencyof association with beneficial fungi could explain the relativerarity of a plant species. Alternatively, lower growth rate orhigher intraspecific competition in rare relative to commonplant species could explain rarity.
The lower limit of plant abundance can take a lower valuefor common species than for rare species when the effects ofper capita predation (Cc
uq) plus the per capita costs of mycor-
rhizal associations (βilivki) are greater in a common relative to a
rare plant species (βclcvkcþ Cu
cq > βrlrvkrþ Cu
rq), given similar
per capita intraspecific and interspecific competition (ai,j) andper capita growth rate (bi):
epc ¼ bc−βclcv
kc−X n
j¼1; j≠cac; jp j
h i−Cu
cq
ac;c≤br−βr
lrv
kr−X n
j¼1; j≠rar; jp j
h i−Cu
r q
ar;r¼ epr ð15Þ
Greater effect of natural enemies on common relative torare plant species is predicted by the J-C hypothesis (Janzen1970; Connell 1971; Connell et al. 1984). Furthermore, sev-eral empirical studies supported this prediction (Bachelot andKobe 2013; Bagchi et al. 2014). However, if plant naturalenemies equally target common and rare plant species (Cc
uq=Cruq), then the costs that arise from mycorrhizal associations
could explain rarity (if rare species pay greater per capita costsfor the association than common species) or large fluctuationsin common plant species abundance (if common plant speciesexperiences greater per capita costs than rare plant species).
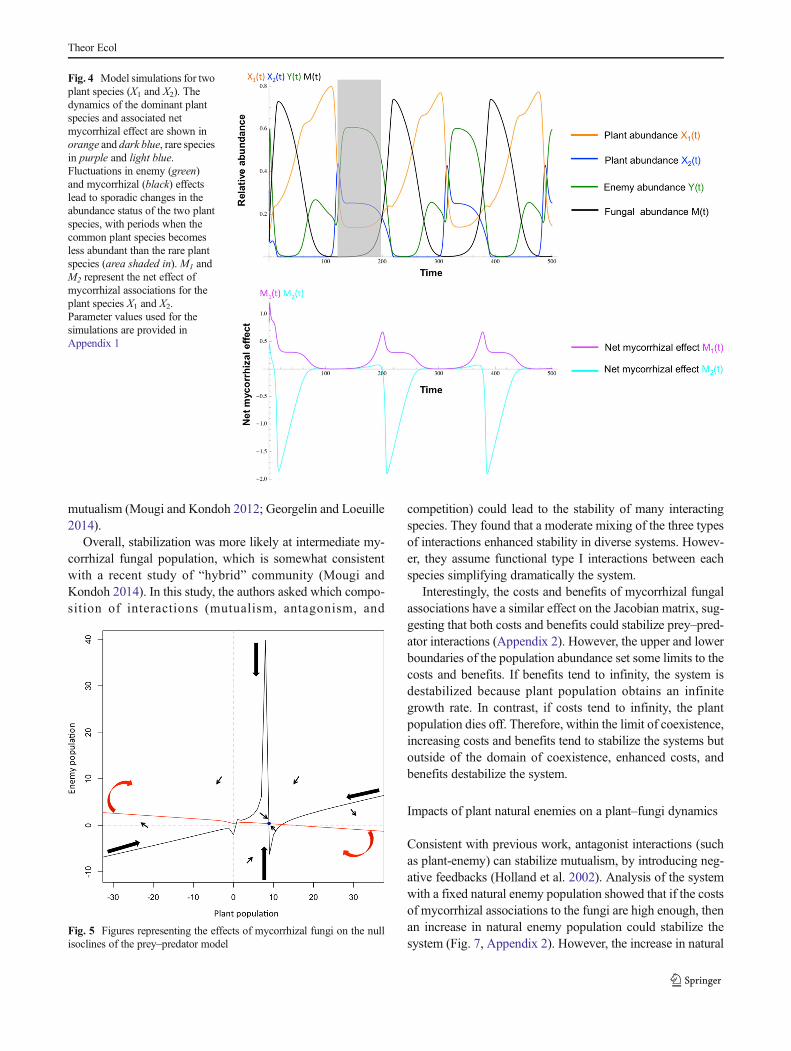
Overall, variation in the strength and direction of enemyand mycorrhizal fungal feedbacks confines the population ofeach plant species between an upper limit pi and a lower limitepi, ensuring plant species coexistence because no species willgo extinct or become overdominant. Furthermore, commonplant species are likely to reach higher abundance and canexperience greater fluctuations in abundance over time thanrare plant species (Fig. 4).
Beside change in the strength of various interactionsamong species, the hierarchical structure of the interactionscan greatly impact species coexistence (Kondoh et al. 2010;Mougi and Kondoh 2012; Suweis et al. 2014; Mougi andKondoh 2014). In our model, however, we did not incorporatehierarchical structure since each species interact with every
other species. Future work should investigate how networkstructure would affect natural enemies–plants–mycorrhizalfungi interactions.
Impacts of mycorrhizal associations on a plant-enemydynamics
Consistent with recent work (Holland et al. 2013; Georgelinand Loeuille 2014), the third analysis of the system showedthat adding mutualism to a prey–predator system could stabi-lize or destabilize the interactions. Specifically, our analysisrevealed complex changes in the dynamics of the system withincreasing mycorrhizal fungal abundance (Appendix 2). If theprotection conveyed by the fungi to the plants is small enough,mutualism stabilizes the interactions between plants and nat-ural enemies, suggesting that mutualism can damper the inter-actions between plants and natural enemies (Figs. 5 and 6).However, as mycorrhizal fungal abundance continues to in-crease, the natural enemy population can be pushed to extinc-tion, resulting in the collapse of the system (Fig. 6). Whenprotection conveyed by mycorrhizal fungi to plants is large,mutualism destabilizes the system, which can introduce cycledynamics and collapse (Fig. 6). Similar potential stabilizationand destabilization effects of mutualism were also detected inpollinator–herbivore–plant systems involving unidirectional
Theor Ecol
mutualism (Mougi and Kondoh 2012; Georgelin and Loeuille2014).
Overall, stabilization was more likely at intermediate my-corrhizal fungal population, which is somewhat consistentwith a recent study of “hybrid” community (Mougi andKondoh 2014). In this study, the authors asked which compo-sition of interactions (mutualism, antagonism, and
competition) could lead to the stability of many interactingspecies. They found that a moderate mixing of the three typesof interactions enhanced stability in diverse systems. Howev-er, they assume functional type I interactions between eachspecies simplifying dramatically the system.
Interestingly, the costs and benefits of mycorrhizal fungalassociations have a similar effect on the Jacobian matrix, sug-gesting that both costs and benefits could stabilize prey–pred-ator interactions (Appendix 2). However, the upper and lowerboundaries of the population abundance set some limits to thecosts and benefits. If benefits tend to infinity, the system isdestabilized because plant population obtains an infinitegrowth rate. In contrast, if costs tend to infinity, the plantpopulation dies off. Therefore, within the limit of coexistence,increasing costs and benefits tend to stabilize the systems butoutside of the domain of coexistence, enhanced costs, andbenefits destabilize the system.
Impacts of plant natural enemies on a plant–fungi dynamics
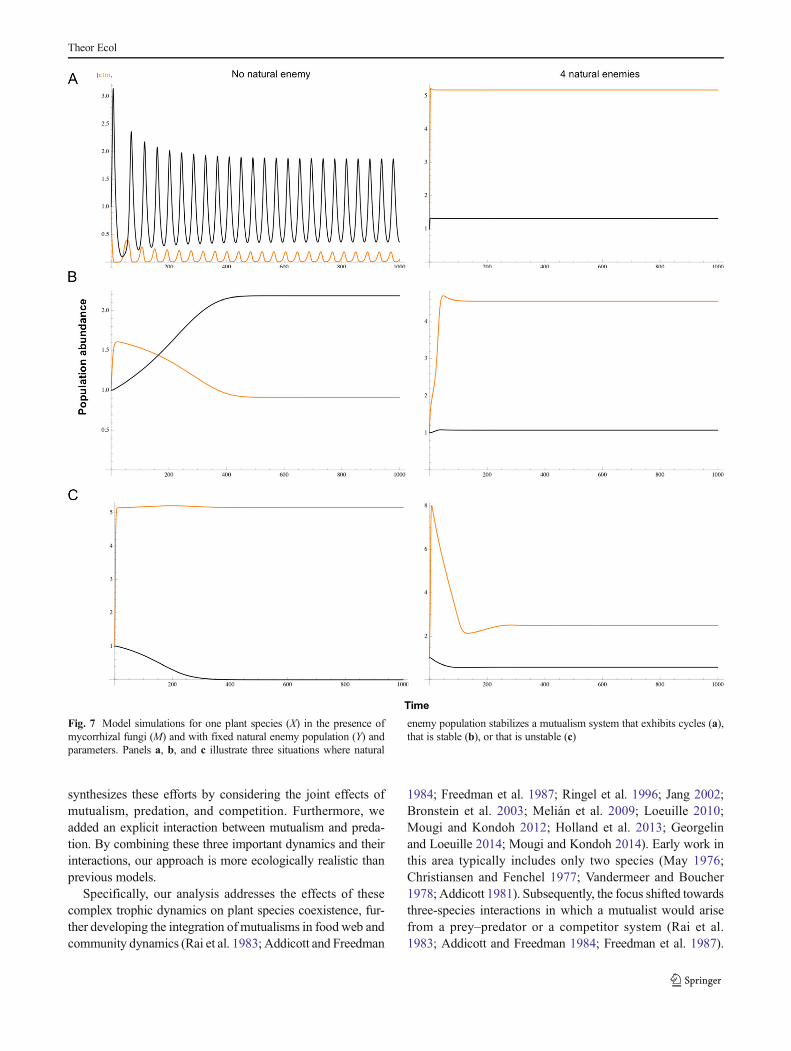
Consistent with previous work, antagonist interactions (suchas plant-enemy) can stabilize mutualism, by introducing neg-ative feedbacks (Holland et al. 2002). Analysis of the systemwith a fixed natural enemy population showed that if the costsof mycorrhizal associations to the fungi are high enough, thenan increase in natural enemy population could stabilize thesystem (Fig. 7, Appendix 2). However, the increase in natural
Fig. 5 Figures representing the effects of mycorrhizal fungi on the nullisoclines of the prey–predator model
Fig. 4 Model simulations for twoplant species (X1 and X2). Thedynamics of the dominant plantspecies and associated netmycorrhizal effect are shown inorange and dark blue, rare speciesin purple and light blue.Fluctuations in enemy (green)and mycorrhizal (black) effectslead to sporadic changes in theabundance status of the two plantspecies, with periods when thecommon plant species becomesless abundant than the rare plantspecies (area shaded in).M1 andM2 represent the net effect ofmycorrhizal associations for theplant species X1 and X2.Parameter values used for thesimulations are provided inAppendix 1
Theor Ecol
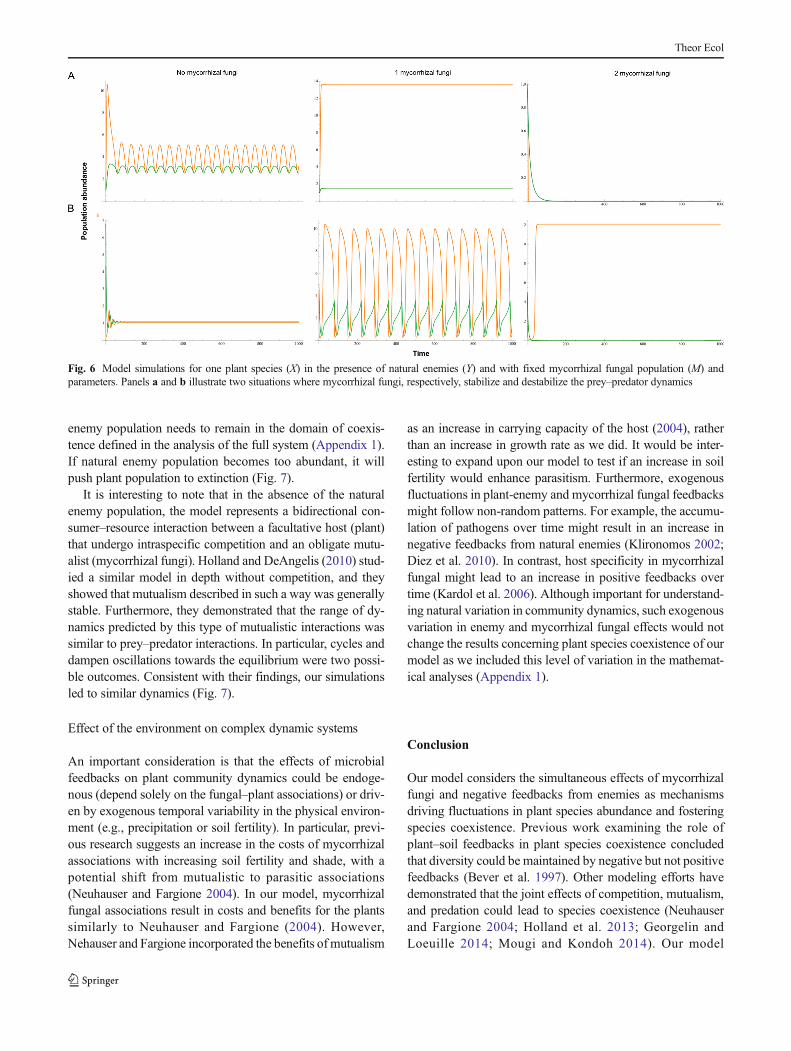
enemy population needs to remain in the domain of coexis-tence defined in the analysis of the full system (Appendix 1).If natural enemy population becomes too abundant, it willpush plant population to extinction (Fig. 7).
It is interesting to note that in the absence of the naturalenemy population, the model represents a bidirectional con-sumer–resource interaction between a facultative host (plant)that undergo intraspecific competition and an obligate mutu-alist (mycorrhizal fungi). Holland and DeAngelis (2010) stud-ied a similar model in depth without competition, and theyshowed that mutualism described in such a way was generallystable. Furthermore, they demonstrated that the range of dy-namics predicted by this type of mutualistic interactions wassimilar to prey–predator interactions. In particular, cycles anddampen oscillations towards the equilibrium were two possi-ble outcomes. Consistent with their findings, our simulationsled to similar dynamics (Fig. 7).
Effect of the environment on complex dynamic systems
An important consideration is that the effects of microbialfeedbacks on plant community dynamics could be endoge-nous (depend solely on the fungal–plant associations) or driv-en by exogenous temporal variability in the physical environ-ment (e.g., precipitation or soil fertility). In particular, previ-ous research suggests an increase in the costs of mycorrhizalassociations with increasing soil fertility and shade, with apotential shift from mutualistic to parasitic associations(Neuhauser and Fargione 2004). In our model, mycorrhizalfungal associations result in costs and benefits for the plantssimilarly to Neuhauser and Fargione (2004). However,Nehauser and Fargione incorporated the benefits of mutualism
as an increase in carrying capacity of the host (2004), ratherthan an increase in growth rate as we did. It would be inter-esting to expand upon our model to test if an increase in soilfertility would enhance parasitism. Furthermore, exogenousfluctuations in plant-enemy and mycorrhizal fungal feedbacksmight follow non-random patterns. For example, the accumu-lation of pathogens over time might result in an increase innegative feedbacks from natural enemies (Klironomos 2002;Diez et al. 2010). In contrast, host specificity in mycorrhizalfungal might lead to an increase in positive feedbacks overtime (Kardol et al. 2006). Although important for understand-ing natural variation in community dynamics, such exogenousvariation in enemy and mycorrhizal fungal effects would notchange the results concerning plant species coexistence of ourmodel as we included this level of variation in the mathemat-ical analyses (Appendix 1).
Conclusion
Our model considers the simultaneous effects of mycorrhizalfungi and negative feedbacks from enemies as mechanismsdriving fluctuations in plant species abundance and fosteringspecies coexistence. Previous work examining the role ofplant–soil feedbacks in plant species coexistence concludedthat diversity could be maintained by negative but not positivefeedbacks (Bever et al. 1997). Other modeling efforts havedemonstrated that the joint effects of competition, mutualism,and predation could lead to species coexistence (Neuhauserand Fargione 2004; Holland et al. 2013; Georgelin andLoeuille 2014; Mougi and Kondoh 2014). Our model
Fig. 6 Model simulations for one plant species (X) in the presence of natural enemies (Y) and with fixed mycorrhizal fungal population (M) andparameters. Panels a and b illustrate two situations where mycorrhizal fungi, respectively, stabilize and destabilize the prey–predator dynamics
Theor Ecol
synthesizes these efforts by considering the joint effects ofmutualism, predation, and competition. Furthermore, weadded an explicit interaction between mutualism and preda-tion. By combining these three important dynamics and theirinteractions, our approach is more ecologically realistic thanprevious models.
Specifically, our analysis addresses the effects of thesecomplex trophic dynamics on plant species coexistence, fur-ther developing the integration of mutualisms in food web andcommunity dynamics (Rai et al. 1983; Addicott and Freedman
1984; Freedman et al. 1987; Ringel et al. 1996; Jang 2002;Bronstein et al. 2003; Melián et al. 2009; Loeuille 2010;Mougi and Kondoh 2012; Holland et al. 2013; Georgelinand Loeuille 2014; Mougi and Kondoh 2014). Early work inthis area typically includes only two species (May 1976;Christiansen and Fenchel 1977; Vandermeer and Boucher1978; Addicott 1981). Subsequently, the focus shifted towardsthree-species interactions in which a mutualist would arisefrom a prey–predator or a competitor system (Rai et al.1983; Addicott and Freedman 1984; Freedman et al. 1987).
Fig. 7 Model simulations for one plant species (X) in the presence ofmycorrhizal fungi (M) and with fixed natural enemy population (Y) andparameters. Panels a, b, and c illustrate three situations where natural
enemy population stabilizes a mutualism system that exhibits cycles (a),that is stable (b), or that is unstable (c)
Theor Ecol

These studies found that different types of mutualisms stabi-lize or destabilize a community: mutualisms that deter preda-tors, compete with a predator, or decrease competitive inter-actions can stabilize interspecific interactions, whereas mutu-alisms that increase prey availability or asymmetric competi-tion might destabilize interactions (Addicott and Freedman1984; Thébault and Fontaine 2010). Consistent with thesestudies, Holland et al. (2013) concluded that a mutualisticpollinator could maintain the stability of the system involvinga plant, a mutualistic pollinator (unidirectional mutualism),and a nectar-parasite. Similarly, our work shows that n plantspecies can coexist with enemy and mycorrhizal fungal pop-ulations because the mycorrhizal fungi and natural enemystabilize each other interactions with plants. Our system ex-pands on previous work by combining competition, predation,and bidirectional mutualism with density-dependent costs andbenefits involving n+2 species. However, our model lacksnetwork structure, which can have dramatic effects on com-munity diversity (Mougi and Kondoh 2014; Suweis et al.2014). Future work should aim at combining network struc-ture, trophic interactions, and community dynamics.
Acknowledgments Benedicte Bachelot was partially supported withfunds from National Science Foundation DEB-524989 to MU. We aregrateful to Dr. Agnes Bachelot, Dr. Alain Bachelot, Dr. Duncan Menge,Dr. Jason Hoeksema, Dr. Stefano Allesina, Dr. Samir Suweis, Dr. Char-lotte Lee, Rachael Eaton, Benton Taylor, Bob Muscarella, NaomiSchwartz, and anonymous reviewers for useful comments on the manu-script. In particular, we are grateful to Axios Review that greatly helpedus improve our manuscript.
References
Addicott JF (1979) A multispecies aphid–ant association: density depen-dence and species-specific effects. Can J Zool 57:558–569
Addicott JF (1981) Properties of 2-species models of mutualism: simula-tion studies. Oecologia 49:42–49
Addicott JF, Freedman HI (1984) On the structure and stability of mutu-alistic systems: analysis of predator–prey and competition models asmodified by the action of a slow-growing mutualist. Theor PopulBiol 26:320–339
Allesina S, Tang S (2012) Stability criteria for complex ecosystems.Nature 483:205–208
Auge RM, Schekel KA,Wample RL (1987) Leaf water and carbohydratestatus of VA mycorrhizal rose exposed to drought stress. Plant Soil302:291–302
Bachelot B, Kobe RK (2013) Rare species advantage? Richness of dam-age types due to natural enemies increases with species abundancein a wet tropical forest. J Ecol 101:846–856
Bagchi R, Gallery RE, Gripenberg S, Gurr SJ, Narayan L, Addis CE,Freckleton RP, Lewis OT (2014) Pathogens and insect herbivoresdrive rainforest plant diversity and composition. Nature 506:85–90
Barber NA, Adler LS, Theis N, Hazzard RV, Kiers ET (2012) Herbivoryreduces plant interactions with above- and belowground antagonistsand mutualists. Ecology 93:1560–1570
Bardgett RD, Wardle DA (2010) Aboveground-belowground linkages.Oxford University Press, Oxford
Bever JD (2003) Soil community feedback and the coexistence of com-petitors: conceptual frameworks and empirical tests. New Phytol157:465–473
Bever JD, Westover KM, Antonovics J (1997) Incorporating the soilcommunity into plant population dynamics: the utility of the feed-back approach. J Ecol 85:561–573
Bi HH, Song YY, Zeng RS (2007) Biochemical and molecular responsesof host plants to mycorrhizal infection and their roles in plant de-fence. Allelopathy J 20:15–27
Booth MG, Hoeksema JD (2010) Mycorrhizal networks counteract com-petitive effects of canopy trees on seedling survival. Ecology 91:2294–2302
Bronstein JL (2001a) The exploitation of mutualisms. Ecol Lett 4:277–287
Bronstein JL (2001b) The costs of mutualism. Am Zool 41:825–839Bronstein J, Wilson W, Morris W (2003) Ecological dynamics of
mutualist/antagonist communities. Am Nat 162:S24–S39Bruno J, Stachowicz J, Bertness M (2003) Inclusion of facilitation into
ecological theory. Trends Ecol Evol 18:119–125Cantrell RS, Cosner C (2001) On the dynamics of predator–prey models
with the Beddington–DeAngelis functional response. J Math AnalAppl 257:206–222
Chesson P (2000)Mechanisms of maintenance of species diversity. AnnuRev Ecol Syst 31:343–366
Chesson P, Kuang JJ (2008) The interaction between predation and com-petition. Nature 456:235–238
Christiansen FB, Fenchel T (1977) Theories of populations in biologicalcommunities. Springer
Connell JH (1971) On the role of natural enemies in preventing compet-itive exclusion in somemarine animals and in rain forest trees. Pages298–312. In: Boer P J, Graadwell G R (eds) Dynamics of numbersin populations
Connell JH, Tracey JG, Webb LJ (1984) Compensatory recruitment,growth, and mortality as factors maintaining rain forest. EcolMonogr 54:141–164
Diez JM, Dickie I, Edwards G, Hulme PE, Sullivan JJ, Duncan RP (2010)Negative soil feedbacks accumulate over time for non-native plantspecies. Ecol Lett 13:803–809
Eppstein MJ, Molofsky J (2007) Invasiveness in plant communities withfeedbacks. Ecol Lett 10:253–263
Eppstein MJ, Bever JD, Molofsky J (2006) Spatio-temporal communitydynamics induced by frequency dependent interactions. Ecol Model197:133–147
Fitter A (1991) Costs and benefits of mycorrhizas: implications for func-tioning under natural conditions. Experientia 47:350–355
Fontaine C, Guimarães PR, Kéfi S, Loeuille N, Memmott J, van derPutten WH, van Veen FJF, Thébault E (2011) The ecological andevolutionary implications of merging different types of networks.Ecol Lett 14:1170–1181
Fox L, Morrow P (1981) Specialization: species property or local phe-nomenon? Science 211:887–893
Freedman HI, Addicott JF, Rai B (1987) Obligate mutualism with apredator: stability and persistence of three-species models. TheorPopul Biol 342:157–175
Gange A, West HM (1994) Interactions between arbuscular mycorrhizalfungi and foliar-feeding insects in Plantago lanceolata L. NewPhytol 128:79–87
Gause G, Witt A (1935) Behavior of mixed populations and the problemof natural selection. Am Nat 69:596–609
Gehring CA, Whitham TG (1994) Comparisons of ectomycorrhizae onpinyon pines (Pinus edulis; Pinaceae) across extremes of soil typeand herbivory. Ecology 81:1509–1516
Georgelin E, Loeuille N (2014) Dynamics of coupled mutualistic andantagonistic interactions, and their implications for ecosystem man-agement. J Theor Biol 346:67–74
Theor Ecol

Hierro JL, Maron JL, Callaway RM (2005) A biogeographical approachto plant invasions: the importance of studying exotics in their intro-duced and native range. Ecology 93:5–15
Hoeksema JD, Chaudhary VB, Gehring CA, Johnson NC, Karst J, KoideRT, Umbanhowar J (2010) A meta-analysis of context-dependencyin plant response to inoculation with mycorrhizal fungi. Ecol Lett13:394–407
Hoffmann D, Vierheilig H, Schausberger P (2011) Mycorrhiza-inducedtrophic cascade enhances fitness and population growth of anacarine predator. Oecologia 166:141–9
HoffmannD, Vierheilig H, Peneder S, Schausberger P (2011)Mycorrhizamodulates aboveground tri-trophic interactions to the fitness benefitof its host plant. Ecological Entomology 36:574–581
Holland JN (2002) Benefits and costs of mutualism: demographic conse-quences in a pollinating seed-consumer interaction. Proc Biol Sci RSoc 269:1405–1412
Holland J, DeAngelis DL (2006) Interspecific population regulation andthe stability of mutualism: fruit abortion and density-dependent mor-tality of pollinating seed-eating insects. Oikos 3:563–571
Holland JN, DeAngelis DL (2010) A consumer-resource approach to thedensity-dependent population dynamics of mutualism. Ecology 91:1286–1295
Holland J, DeAngelis D, Bronstein J (2002) Population dynamics andmutualism: functional responses of benefits and costs. Am Nat 159:231–244
Holland JN, DeAngelis DL, Schultz ST (2004) Evolutionary stability ofmutualism: interspecific population regulation as an evolutionarilystable strategy. Proc R Soc Biol Sci 271:1807–1814
Holland JN, Wang Y, Sun S, DeAngelis DL (2013) Consumer–resourcedynamics of indirect interactions in a mutualism–parasitism foodweb module. Theor Ecol 6:475–493
Holling CS (1959) The components of predation as revealed by a study ofsmall-mammal predation of the European pine sawfly. Can Entomol91:293–320
Jaenike J (1990) Host specialization in phytophageous insects. Annu RevEcol Syst 21:243–273
Jang SRJ (2002) Dynamics of herbivore-plant-pollinator models. J MathBiol 44:129–149
Janzen DH (1970) Herbivores and the number of tree species in tropicalforests. Am Nat 104:501–528
Jeschke J, Kopp M, Tollrian R (2002) Predator functional responses:discriminating between handling and digesting prey. Ecol Monogr72:95–112
Johnson ANC, Graham JH (2013) The continuum concept remains auseful framework for studying mycorrhizal functioning. Plant Soil363:411–419
Johnson ANC, Graham JH, Smith FA (1997) Functioning of mycorrhizalassociations along the mutualism-parasitism continuum. NewPhytol 135:575–586
Kardol P, Bezemer TM, Van der PuttenWH (2006) Temporal variation inplant-soil feedback controls succession. Ecol Lett 9:1080–1088
Kiers ET, Duhamel M, Beesetty Y, Mensah JA, Franken O, VerbruggenE, Fellbaum CR, Kowalchuk GA, Hart MM, Bago A, Palmer TM,West SA, Vandenkoornhuyse P, Jansa J, Bücking H (2011)Reciprocal rewards stabilize cooperation in the mycorrhizal symbi-osis. Science 333:880–882
Klironomos JN (2002) Feedback with soil biota contributes to plant rarityand invasiveness in communities. Nature 417:67–70
Klironomos J, McCune J, Moutoglis P (2004) Species of arbuscular my-corrhizal fungi affect mycorrhizal responses to simulate herbivory.Appl Soil Ecol 26:133–141
Koide R (1991) Tansley review no. 29: nutrient supply, and nutrient toplant response mycorrhizal infection. New Phytol 117:365–386
Koide R, Elliott G (1989) Cost, benefit and efficiency of the vesicular-arbuscular mycorrhizal symbiosis. Funct Ecol 3:252–255
Kondoh M, Kato S, Sakato Y (2010) Food webs are built up with nestedsubwebs. Ecology 91:3123–3130
Koricheva J, Gange AM, Jones T (2009) Effects of mycorrhizal fungi oninsect herbivores: a meta-analysis. Ecology 90:2088–2097
Krebs JR (1974) Behavioral aspects of predation. In: Bateson HG,Klopfer P (eds) Perspectives in ethology. Plenum, New York, pp73–111
Lekberg Y, Koide RT (2005) Is plant performance limited by abundanceof arbuscular mycorrhizal fungi? A meta-analysis of studies pub-lished between 1988 and 2003. New Phytol 168:189–204
Leon JA, Tumpson DB (1975) Competition between two species for twocomplementary or substitutable resources. J Theor Biol 50:185–201
Levine JM, Pachepsky E, Kendall BE, Yelenik SG, Lambers JHR (2006)Plant-soil feedbacks and invasive spread. Ecol Lett 9:1005–1014
Loeuille N (2010) Influence of evolution on the stability of ecologicalcommunities. Ecol Lett 13:1536–1545
May RM (1976) Models for two interacting populations. In: May RM(ed) Theoretical ecology. Saunders, Philadelphia, pp 49–70
McGill B (2005) A mechanistic model of a mutualism and its ecologicaland evolutionary dynamics. Ecol Model 187:413–425
Melián CJ, Bascompte J, Jordano P, Krivan V (2009) Diversity in acomplex ecological network with two interaction types. Oikos118:122–130
Moora M, Opik M, Sent R, Zobel M (2004) Native arbuscular fungalcommunities mycorrhizal influence the seedling performance of raredifferentially and common Pulsatilla species. Funct Ecol 18:554–562
Morales M (2000) Mechanisms and density dependence of benefit in anant-membracid mutualism. Ecology 81:482–489
Mordecai E (2011) Pathogen impacts on plant communities: unifyingtheory, concepts, and empirical work. Ecol Monogr 81:429–441
Mougi A, KondohM (2012) Diversity of interaction types and ecologicalcommunity stability. Science 337:349–351
Mougi A, Kondoh M (2014) Stability of competition-antagonism-mutualism hybrid community and the role of community networkstructure. J Theor Biol 360:54–58
Murdoch W (1975) Diversity, complexity, stability and pest control. JAppl Ecol 12:795–807
Neuhauser C, Fargione JE (2004) A mutualism–parasitism continuummodel and its application to plant–mycorrhizae interactions. EcolModel 177:337–352
Newsham KK, Fitter AH, Watkinson AR (1995) Multi-functionality andbiodiversity in arbuscular mycorrhizas. Trends Ecol Evol 10:407–411
Novotny V, Miller SE, Baje L, Balagawi S, Basset Y, Cizek L, Craft KJ,Dem F, Drew RAI, Hulcr J, Leps J, Lewis OT, Pokon R, StewartAJA, Samuelson GA,Weiblen GD (2010) Guild-specific patterns ofspecies richness and host specialization in plant–herbivore foodwebs from a tropical forest. J Anim Ecol 79:1193–1203
Olsson PA, Rahm J, Aliasgharzad N (2010) Carbon dynamics in mycor-rhizal symbioses is linked to carbon costs and phosphorus benefits.FEMS Microbiol Ecol 72:125–131
Pacala S, CrawleyM (1992) Herbivores and plant diversity. AmNat 140:243–260
PozoMJ, Azcón-Aguilar C (2007) Unraveling mycorrhiza-induced resis-tance. Curr Opin Plant Biol 10:393–398
Rai B, Freedman H, Addicott LJ (1983) Analysis of three species modelsof mutualism in predator–prey and competitive systems. MathBiosci 50:13–50
Real L (1977) The kinetics of functional response. Am Nat 111:289–300Ringel MS, Hu HH, Anderson G (1996) The stability and persistence of
mutualisms embedded in community interactions. Theor Popul Biol50:281–297
Rosenzweig M,MacArthur R (1963) Graphical representation and stabil-ity conditions of predator–prey interactions. Am Nat 97:209–223
Theor Ecol

Roughgarden J (1975) Evolution of marine symbiosis—a simple cost-benefit model. Ecology 56:1201–1208
Salzer P, Hübner B, Sirrenberg A, Hager A (1997) Differential effect ofpurified spruce chitinases and beta-1, 3-glucanases on the activity ofelicitors from ectomycorrhizal fungi. Plant Physiol 114:957–968
Saravesi K, Markkola A, Rautio P, Roitto M, Tuomi J (2008) Defoliationcauses parallel temporal responses in a host tree and its fungal sym-bionts. Oecologia 156:117–123
Schmitt RJ, Holbrook SJ (2003) Mutualism can mediate competition andpromote coexistence. Ecol Lett 6:898–902
Selosse MA, Richard FHX, Simard SW (2006) Mycorrhizal networks:des liaisons dangereuses? Trends Ecol Evol 21:621–628
Silvertown JW (1980) The evolutionary ecology of mast seeding in trees.Biol J Linn Soc 14:235–250
Simard S, Durall D (2004) Mycorrhizal networks: a review of their ex-tent, function, and importance. Can J Bot 82:1140–1165
Simonsen AK, Stinchcombe JR (2014) Herbivory eliminates fitness costsof mutualism exploiters. New Phytol
Smith SE, Read DJ (2008) Mycorrhizal symbiosis, 3rd edn. Academic,London
Smout S, Asseburg C, Matthiopoulos J, Fernández C, Redpath S,Thirgood S, Harwood J (2010) The functional response of a gener-alist predator. PLoS One 5:e10761
Stephens DW, Krebs JR (1986) Foraging theory, vol 1. PrincetonUniversity Press, Princeton, pp 1–100
Strauss S, Irwin R (2004) Ecological and evolutionary consequences ofmultispecies plant-animal interactions. AnnuRev Ecol Evol Syst 35:435–466
Suweis S, Grilli J, Maritan A (2014) Disentangling the effect of hybridinteractions and of the constant effort hypothesis on ecological com-munity stability. Oikos 123:525–532
Tang S, Awar SP, Allesina S (2014) Correlation between interactionstrengths drives stability in large ecological networks. Ecol Lett17:1094–1100
Thébault E, Fontaine C (2010) Stability of ecological communities andthe architecture of mutualistic and trophic networks. Science 329:853–856
Tilman D (1982) Resource competition and community structure.Princeton University Press, Princeton
Toju H, Sato H, Yamamoto S, Kadowaki K, Tanabe AS, Yazawa S,Nishimura O, Agata K (2013) How are plant and fungal communi-ties linked to each other in belowground ecosystems? A massivelyparallel pyrosequencing analysis of the association specificity ofroot-associated fungi and their host plants. Ecol Evol 3:1–13
Van der Heijden MGA, Horton TR (2009) Socialism in soil? The impor-tance of mycorrhizal fungal networks for facilitation in natural eco-systems. J Ecol 97:1139–1150
Van der Heijden MGA, Klironomos J, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR (1998) Mycorrhizalfungal diversity determines plant biodiversity, ecosystem variabilityand productivity. Nature 396:69–72
Van der Putten WH, Bardgett RD, de Ruiter PC, Hol WHG, Meyer KM,Bezemer TM, Bradford MA, Christensen S, Eppinga MB, FukamiT, Hemerik L, Molofsky J, Schädler M, Scherber C, Strauss SY, VosM, Wardle DA (2009) Empirical and theoretical challenges inaboveground-belowground ecology. Oecologia 161:1–14
Vandermeer JH, Boucher DH (1978) Varieties of mutualistic interactionin population models. J Theor Biol 74:549–558
Vannette R, Hunter MD (2011) Plant defense theory re-examined: non-linear expectations based on the costs and benefits of resource mu-tualisms. J Ecol 99:66–76
Wangersky PJ (1978) Lotka-Volterra population models. Annu Rev EcolSyst 9:189–218
Wardle DA (2002) Communities and ecosystems: linking the above-ground and belowground components. Princeton University Press,Princeton
Wolfram Research, Inc. (2008) Mathematica, Version 7.0, ChampaignWootton J (1994) The nature and consequences of indirect effects in
ecological communities. Annu Rev Ecol Syst 25:443–466Wright SJ (2002) Plant diversity in tropical forests: a review of mecha-
nisms of species coexistence. Oecologia 130:1–14Zhang Q, Xu L, Tang J, Bai M, Chen X (2011) Arbuscular mycorrhizal
mediation of biomass-density relationship of Medicago sativa L.under two water conditions in a field experiment. Mycorrhiza 21:269–277
Theor Ecol