Embed Size (px)
Citation preview

Interactions of cyclic adenosine monophosphate, brain-derivedneurotrophic factor, and glial cell line-derived neurotrophic factortreatment on the survival and growth of postnatal mesencephalic
dopamine neurons in vitro
Jesus Lara,1 Kiyoshi Kusano, Shirley House, and Harold Gainer*Laboratory of Neurochemistry, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, MD 20892-4120, USA
Received 16 April 2002; revised 30 September 2002; accepted 1 October 2002
Abstract
The survival of rat postnatal mesencephalic dopamine (DA) neurons in dissociated cell cultures was studied by examining thecombinatorial effects of dibutyryl cyclic adenosine monophosphate (db-cAMP), glial cell line-derived neurotrophic factor (GDNF), andbrain-derived neurotrophic factor (BDNF), as well as selective inhibitors of protein kinase A (PKA), and mitogen-activated protein kinase(MAPK). Postnatal DA neurons were maintained for 14 days in vitro, and were identified by immunohistochemistry using tyrosinehydroxylase antibody. The survival and growth of DA neurons was significantly increased by the inclusion of either �100 �M db-cAMPor 10 �M Forskolin plus 100 �M IBMX in the culture medium. Neither 10–50 ng/ml GDNF nor 50 ng/ml BDNF alone significantlyincreased DA neuron survival in vitro. However, the combined use of GDNF and BDNF did increase DA neuron survival, and the additionof either db-cAMP or IBMX/Forskolin to media containing these neurotrophins markedly increased DA neuron survival and growth. ThecAMP inhibitor Rp-cAMP, the cAMP-dependent protein kinase A inhibitor H89, and the MAP kinase (MAPK) pathway inhibitor PD98059significantly reduced the survival of DA neurons when applied alone in the absence of added growth factors. Application of GDNF plusBDNF, or db-cAMP significantly protected the DA neurons from the deleterious effects on survival of either 20 �M H89 or 20 �M PD98059. The results suggest that BDNF, GDNF, and cAMP produce convergent signals to activate PKA and MAPK pathways which areinvolved in the survival of postnatal mesencephalic DA neurons in vitro.Published by Elsevier Science (USA).
Keywords: cAMP; GDNF; BDNF; Mesencephalon; Dopaminergic neurons; Parkinson’s disease; Tyrosine hydroxylase; Substantia nigra; MAP kinase
Introduction
Neurotrophins have been actively studied as possibleneuroprotective agents for degenerating substantia nigra do-pamine (DA) neurons in Parkinson’s disease (Alexi et al.,2000; Collier and Sortwell, 1999; Dunnett and Bjorklund,1999; Unsicker, 1994). Among these, brain-derived neuro-
trophic factor (BDNF) and glial-derived neurotrophic factor(GDNF) have been of particular interest, and these factorshave been shown to protect DA neurons from degenerationboth in vivo (GDNF: (Beck et al., 1995; Choi-Lundberg etal., 1997; Kordower et al., 2000; Sauer et al., 1995; Sinclairet al., 1996; Tomac et al., 1995); BDNF: (Shults et al., 1995;Tsukahara et al., 1995)) and in vitro (GDNF: (Burke et al.,1998; Clarkson et al., 1995; Lin et al., 1993; Schatz et al.,1999); BDNF: (Hyman et al., 1991; Knusel et al., 1991;Studer et al., 1995)). These neurotrophins are also synthe-sized and released from the mesencephalic DA neuronsthemselves (Pochon et al., 1997; Seroogy and Gall, 1993),and the receptors for both BDNF (Feng et al., 1999; Merlioet al., 1992) and GDNF (Feng et al., 1999; Jing et al., 1996,
* Corresponding author. National Institutes of Health, NINDS-LNC,Building 36, Room 4D04, Bethesda, MD 20892, USA. Fax: �1-301-496-1339.
E-mail address: [email protected] (H. Gainer).1 Present address: Centro Universitario de Investigaciones Biomedicas,
Universidad de Colima, Ave 25 de Julio #965, C.P. 28045, Colima VillaSan Sebastian, Colima, Col., Mexico.
R
Available online at www.sciencedirect.com
Experimental Neurology 180 (2003) 32–45 www.elsevier.com/locate/yexnr
0014-4886/03/$ – see front matter Published by Elsevier Science (USA).doi:10.1016/S0014-4886(02)00028-6

1997; Matsuo et al., 2000; Nostrat et al., 1997; Takahashi etal., 1988; Trupp et al., 1994, 1997) have been found local-ized within these neurons, indicating that these neurotro-phins and their receptors appear to be in place for autocrineor paracrine actions. In addition, when these growth factorsare used together in vitro, they are often more effective inimproving the survival, growth, and differentiation of var-ious types of neurons (Du and Iacovetti, 1995; Erickson etal., 2001; Franke et al., 2000; Goldberg and Barres, 2000;Ogilvie et al., 2000; Yan et al., 1999). However, theirinteractions with the cellular signaling mechanisms in themesencephalic DA neurons that are responsible for en-hanced survival are not well understood.
Signaling through cAMP-dependent pathways has beenrecognized as playing an important role in the maturationand long-term survival of mesencephalic DA neurons(Branton et al., 1998; Michel and Agid, 1996). The eleva-tion of intracellular cAMP can promote DA neuron survival,and its survival promoting effects in mesencephalic DAneurons appear similar to those found in other CNS neurons(Bonni et al., 1999; Goldberg and Barres, 2000). In addition,the increase of cAMP levels in neurons has been reported toenhance their responsiveness to either GDNF or BDNF(Engele and Franke, 1996; Goldberg and Barres, 2000;Meyer-Franke et al., 1995, 1998).
In this article, we studied the individual and combinedeffects of raising intracellular cAMP, and extracellular ap-plications of GDNF and/or BDNF on the survival andgrowth of postnatal rat mesencephalic DA neurons in vitro.We chose to study postnatal neurons, in part, because it hasbeen reported that the natural cell death period for DAneurons in the substantia nigra occurs predominantly in theearly postnatal period (Oo and Burke, 1997) and also be-cause postnatal neurons are more differentiated and thedamage to their axons and dendrites during the preparationof dissociated cells for culture would presumably be moresevere, thereby providing an additional challenge for theirsurvival in vitro. We found that the most effective condi-tions to maintain the DA neurons in vitro is the simulta-neous presence of db-cAMP, GDNF, and BDNF in theculture medium. These data and the results of other exper-iments using selective protein kinase inhibitors suggest thatthere are convergent signaling pathways that are involved inthe maintenance of DA neuron survival in vitro.
Materials and methods
Cell culture and drug treatments
Coronal mesencephalic slices from postnatal (PN) 2- to5-day-old Sprague–Dawley rats were made with the use ofMcIlwain tissue slicer at a thickness of about 350 �m. Allprocedures were carried out in accordance with the NIHGuidelines on the Care and Use of Animals and an animal
study protocol approved by the NINDS Animal Care and UseCommittee.
The substantia nigra and ventral tegmental areas were man-ually dissected out from the individual slices (see Fig. 1 for theboundaries of the slices taken for culture), and treated with 20U/ml papain and 2 � 103 U/ml DNase in Earle’s balanced saltsolution (BSS) for 90 min at 37°C (Worthington BiochemicalCorp., Lakewood, NJ, USA). Enzyme-treated tissues werethen mechanically dispersed by gentle trituration through a10-ml pipette and further enzyme action was stopped by add-ing 10 mg/ml ovomucoid protease inhibitor with 10 mg/mlbovine serum albumin (both from Worthington) in Earle’sBSS. The suspended cells were plated at a density of about 1.2� 106 cells (including neurons and other cells) on to each22-mm-diameter glass coverslip, which had been pretreatedwith 10 �g/ml poly-D-lysine (Sigma, St. Louis, MO, USA) andcoated with matrigel (Collaborative Biochemical Products,Bedford, MA, USA). Approximately 30 coverslips were pro-duced from 10 pups. Each coverslip was kept in an individual35-mm Petri dish, and the cells were covered by 2 ml of culturemedium and maintained in a humidified incubator at 37°C with5% CO2. In most of the experiments, the control culture me-dium consisted of Neurobasal A plus B27 (Life Technologies,Gaithersburg, MD, USA), 0.5 mM glutamine, 5 mg/ml glu-cose, 1 mM sodium pyruvate, 0.075% (w/v) sodium bicarbon-ate, 10 mM HEPES, and Pen-Strep (100 U/ml penicillin and100 �g/ml streptomycin) supplemented with 10% heat-inacti-vated horse serum (SCM). Neurobasal A and B27 are knownto contain various neuroprotective agents such as antioxidants,steroid hormones, vitamins, bovine albumin (Brewer et al.,1993). Most of the studies described here were carried out inSCM but some experiments were also carried out in serum-freemedium (SFM) for comparison. Similar to the observations ofothers (Cardozo, 1993; Masuko et al., 1992; Michel and Agid,1996; Rayport et al., 1992); we found that the mesencephalicDA neurons did not thrive in culture medium in the absence ofthe horse serum (see Fig. 2). This was primarily due to the poorsurvival of the astrocytes in SFM which aid in the neuronalattachment to the surface of the glass coverslip. The cultures inSCM routinely received a single dose of 8 � 10�5 M 5-fluoro-2deoxyuridine (FdUR; Sigma) on Day 3 for 4 days, to suppressthe overproliferation of nonneuronal cells, whereas in SFM theFdUR treatment was not used. The cultures were fed twice aweek with fresh medium and were usually maintained for 14days in vitro (div), but some cultures were maintained forlonger periods (up to 60 div).
In experiments, cultures were treated individually by theaddition to the medium of 50 ng/ml BDNF (Peprotech, Inc.,Rocky Hill, NJ, USA), 10 ng/ml GDNF (Peprotech), and100–1000 �M db-cAMP (Sigma), or by various combina-tions of these agents. These concentrations for BDNF (Hy-man et al., 1994; Studer et al., 1995) and GDNF (Burke etal., 1998; Clarkson et al., 1995; Sautter et al., 1998) hadpreviously been shown to produce optimal survival anddifferentiation effects on prenatal mesencephalic DA neu-rons. We also compared the GDNF and BDNF obtained
33J. Lara et al. / Experimental Neurology 180 (2003) 32–45

from RD Systems (Minneapolis, MN, USA) with theseneurotrophins obtained from Peprotech, and found no dif-ferences in their effect on DA neuron survival (P � 0.05).The 100 �M db-cAMP concentration was lower than thatused in most studies, but this had been shown to potentiateGDNF’s effects when they were applied together (Brantonet al., 1998; Engele and Franke, 1996; Hartikka et al., 1992;Michele and Agid, 1996). Effects of various modulators ofcAMP activity and protein kinase inhibitors were also em-ployed. All drugs were purchased from Sigma. These in-cluded 200 �M Sp-cAMP, 200 �M 8 Br-cAMP, 400 �MRp-cAMP, 10 �M Forskolin, 100 �M 3-isobutyl-1-meth-ylxanthine (IBMX), 20 �M H89 ((N-2-[p-bromocin-namylamino]ethyl)-5-isoquinolinesulfonamide hydrochlo-ride) and 20 �M PD98059 (2-(2-amino-3-methoxyphenyl)-4H-1-benzopyran-4-one). Stock solutions of some of thesedrugs were made in dimethyl sulfoxide (DMSO). Controlexperiments showed that the resulting DMSO concentration(�0.4%) in the media from the vehicle had no effect oneither cell survival or neurite outgrowth. Higher concentra-
tions (�20 �M) of H89 tended to damage the underlyingastrocyte layer, which could confound the evaluation of theinhibitor effect on the DA neuron survival. Hence, we didnot exceed the concentration 20 �M of H89 in these studies.In contrast, in the PD98059-treated cultures the astrocyteswere not at all effected by the 20 �M concentration used.When 20 �M H89 and 20 �M PD98059 were appliedsimultaneously for 14 days, there was some astrocyte dam-age. DMSO was used as the vehicle for both inhibitors, witha final concentration of 0.4% DMSO in the medium. Inthese experiments, the controls also contained the sameconcentrations of DMSO in the media, and preliminaryexperiments showed no significant effect of the 0.4%DMSO in the media on the survival or neurite outgrowth ofthe DA neurons.
Immunohistochemical techniques
After tissue culture the neurons grown on the coverslipwere fixed by 4% paraformaldehyde in phosphate-buffered
Fig. 1. Representative anterior to posterior coronal sections (A–D) of the PN2 rat mesencephalon, illustrating the areas containing tyrosine hydroxylase (TH)immunostained neurons in the substantia nigra (SN), and ventral tegmental area (VTA), that were selected for tissue culture in this study. The cells in thespecific regions shown between the dotted line (dorsal) and the base of the mesencephalon were specifically isolated and dissociated for these cultures (seeMaterials and methods).
34 J. Lara et al. / Experimental Neurology 180 (2003) 32–45

saline (PBS) for 1 h at room temperature (RT). After severalwashes in PBS, the coverslips were incubated in PBS con-taining 10% normal goat serum, 0.3% Triton X-100, and0.1% Na azide, for 1 h to permeabilize the cell membrane,and to prevent nonspecific antibody binding. They werethen incubated overnight in the primary antibody, poly-clonal rabbit tyrosine hydroxylase antibody (Catalog No.TE101, Protos Biotech Corp., New York, NY, USA), di-luted 1:2000 in PBS at 4°C. On Day 2 the coverslips wererinsed three times in PBS and then incubated in goat poly-clonal anti-rabbit biotinylated secondary antibody (VectorLaboratories, Burlingame, CA, USA) at a 1:500 dilution for1.5 h at RT. This was followed by rinsing three times inPBS and incubation in avidin–biotinylated horseradish per-oxidase at 1:600 dilution (Vectastin Elite ABC kit, VectorLabs., Burlingame, CA, USA) for 1.5 h at RT. Followingthis incubation, they were rinsed three times in PBS andreacted with 3, 3�diaminobenzidine tetrahydrochloride(DAB, Sigma) containing solution, which consisted of 10mg DAB, 12 mg NH4Cl, 40 mg imidazole, 120 mg glucose,24 mg NiSO4, and 50 units of glucose oxidase (Sigma) in 30ml PBS. Following the immunostaining, the preparationswere dehydrated by ethanol, cleared in xylene, and mountedusing Permount (Fisher, Pittsburgh, PA, USA).
Double label immunofluorescence
To examine the distribution of two antigens within aculture, the cultures were processed sequentially using two
different antibodies and chromogens which produced redand green colors. The localization of the first antigen wassimilar to that described above. First, to identify DA neu-rons, a polyclonal rabbit tyrosine hydroxylase antibody(Protos) at dilution of 1:2000 was used, and this was de-tected by the fluorescent dye Alexa 594 (red), conjugatedwith goat anti-rabbit IgG (H � L) (Molecular Probe, Eu-gene, OR, USA) used at 1:200 dilution. A mouse monoclo-nal antibody against microtuble-associated protein-2(MAP-2 (2a � 2b)) (Sigma) was used at dilution of 1:2000to label neurons in the culture, and was detected by thefluorescent dye Alexa 488 (green) conjugated with goatanti-mouse IgG (Molecular Probe). A similar protocol wasused for double labeling tyrosine hydroxylase-immunoreac-tive (TH-ir) neurons and for calbindin or calretinin calciumbinding proteins. Calbindin and calretinin antibodies wereobtained from Chemicon (Temecula, CA, USA) and wereused at 1:1,500 dilutions. The coverslips were mountedusing Gel/Mount (Biomeda Co., Foster City, CA, USA),and the slides were viewed under an epifluorescence micro-scope (Nikon Eclipse E400, Japan) and photographed.
Analyses of cell numbers and soma sizes
The number of DA neurons was determined by countingall the TH-ir neurons on the coverslip under a microscope at200� magnification. For the analysis of cell size, somaprofile areas of 50 randomly selected multipolar neurons,which came from 10 coverslips derived from five indepen-dent cultures in each of the treated and control groups, weremeasured using the Neurolucida program (Version 3.18,MicroBrightField, Inc., Colchester, VT, USA).
Statistical analysis
Quantitative data were expressed as mean � SEM, and nrepresents the number of coverslips analyzed, from a min-imum of three independent experiments. Multiple compar-isons against a single control group were made by one-wayanalysis of variance (ANOVA) followed by Newman–Keuls multiple comparison test using the GraphPad Prismprogram (Version 2.0, San Diego, CA, USA). A two-tailedt test was used to analyze the inhibition data in Table 1. Inall cases, P � 0.05 was considered to be a statisticallysignificant difference.
Results
Characteristics of postnatal mesencephalic DA neurons indissociated cell culture
Comparison of the survival of mesencephalic DA neu-rons from a single dissociated cell culture group (3–21 div)in SCM versus SFM is shown in Fig 2. About 60% of theTH-ir neurons present after 3 div survived by 14 div and
Fig. 2. Comparison of the survival of dissociated postnatal mesencephalicdopaminergic neurons in serum containing (SCM) versus serum-free(SFM) culture media. Dissociated ventral mesencephalic cells derived from13 PN-5 rats were plated onto 40 glass coverslips (22 mm in diameter) andmaintained in the Neurobasal A and B27-based culture medium with orwithout 10% horse serum (see Materials and methods). The numbers oftyrosine hydroxylase immunoreactive (TH-ir) neurons surviving per cov-erslip after 3–21 days in vitro are shown. The numbers of TH-ir cellspresent in the serum-containing medium after 3 div was 219 � 30.5, n �5. The symbols and the error bars at each data point represent the mean �SEM of results from 5 coverslips.
35J. Lara et al. / Experimental Neurology 180 (2003) 32–45

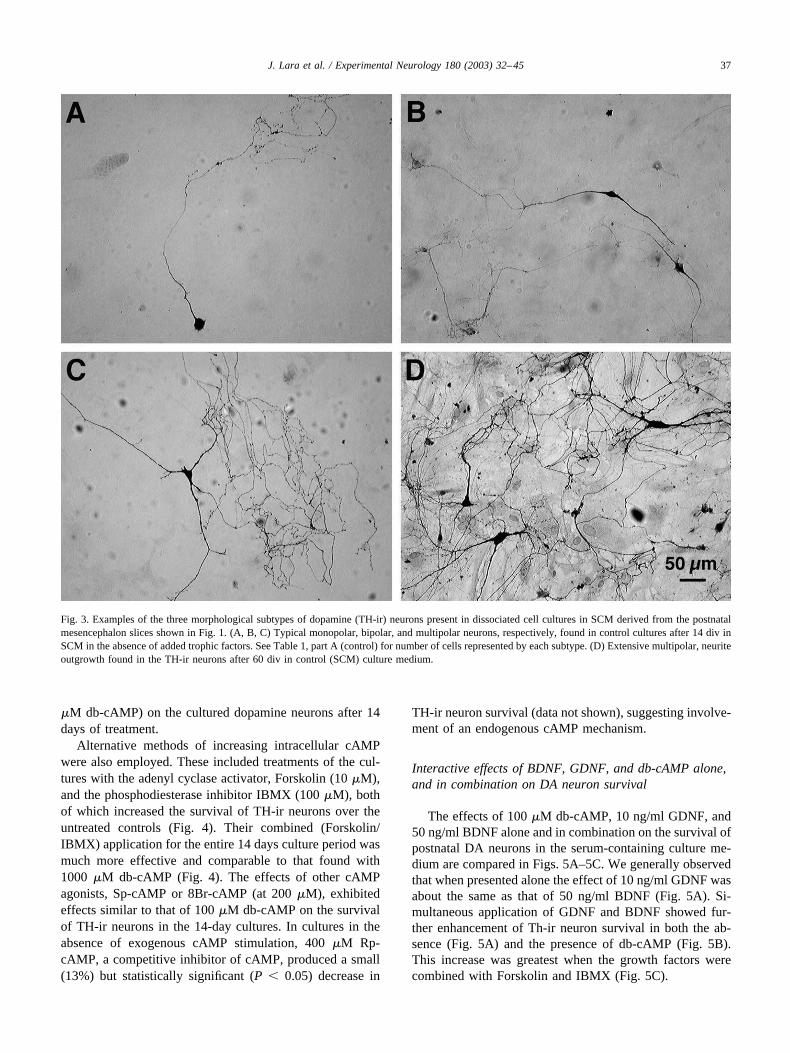
about 50% survived by 21 div in SCM. After 21 div the lossof TH-ir neurons in SCM occurred at a slower rate, and after60 div about 60 (or 27%) TH-ir neurons/coverslip (n � 8)were found in these long-term cultures. These TH-ir neu-rons were morphologically robust and their tendency to befound in clusters (Fig. 3D) suggests that intrinsic factorsexchanged between these neurons might be contributing totheir long-term survival in the absence of exogenous neu-rotrophins. In contrast, in SFM only 30% of the TH-irneurons survived after 14 div, and about 16% after 21 div(Fig. 2). These findings of a lower basal survival rate for DAneurons in SFM as opposed to SCM are similar to thatreported in other studies (Cardozo, 1993; Masuko et al.,1992; Michel and Agid, 1996; Rayport et al., 1992), andhence, we used SCM in all of the experiments describedbelow.
Under these SCM culture conditions, the number of DAneurons detected by TH-ir that were present after 14 div was189 � 18/coverslip (Table 1). These TH-ir neurons repre-sented about 10% of the average, total number of neurons(i.e., the number of MAP2-ir neurons) present on the cov-erslips (n � 11 coverslips fom two independent culturesessions). Three morphological subtypes of TH-ir neuronswere consistently observed in control cultures (Fig. 3).These were monopolar (Fig. 3A), bipolar (Fig. 3B), andmultipolar (Fig. 3C) neurons, representing 14, 31, and 55%,respectively, of the total population of TH-ir neurons on thecoverslip (see Table 1, control). The average soma diameterof the monopolar neurons was small (�15 �m), but therewere also some monopolar neurons that possessed some-what larger somas and longer neurites with varicosities(e.g., Fig. 3A). Since monopolar DA neurons are not usuallypresent in the mesencephalic DA neuron population in vivo,it is possible that this morphological subtype is a product ofthe tissue culture conditions. Comparisons of the numbersof cells represented by each of these subtypes of TH-ir
neurons in the control and various experimental conditionsafter 14 div are summarized in Table 1. As is found in vivo,the cultured TH-ir neurons also contained calcium bindingproteins. Approximately half of the TH-ir neurons con-tained calbindin immunoreactivity, and most of the restwere calretinin immunoreactive (not illustrated).
c-AMP enhances the survival of postnatal DA neurons invitro
The effects of various concentrations of db-cAMP on thesurvival and growth of TH-ir neurons were examined bytreating the cultures continuously with this agent for 14days. The number of TH-ir neurons surviving in these cul-tures increased with increasing db-cAMP concentration(Fig. 4), and at 1000 �M db-cAMP their soma sizes weremuch larger and with thicker and longer neurites. One reportin the literature (Hartikka et al., 1992) suggested that treat-ment of mesencephalon cultures with db-cAMP (1000 �M)for only the initial 3 days was more beneficial for survivaland growth of embryonic dopamine neurons than moreprolonged treatments. We also examined this possibility onthese postnatal dopamine neurons by employing differentdurations of treatment. These variations included comparingan initial 3-day treatment with db-cAMP (at 100 or 1000�M) with groups that were continuously treated at these twoconcentrations for the total 14 div. In another experiment,cultures were treated with 100 �M db-cAMP for variouslengths of time during the 21-day culture period: the firstgroup with the initial 3-day treatment only, a second groupby repeating 3 days of treatment twice (on Days 0–3 andDays 11–14), and a third group by treating continuously for21 days. In all of these studies, the increases in postnataldopamine neuron survival after db-cAMP treatment washighest in the continuously treated group, and there were noobvious deleterious effects of the high concentrations (1000
Table 1Morphological subtypes of TH-ir neurons after 14 div under the influence of various growth factor combinations
Condition n TH-ir neuron number Total
Monopolar Bipolar Multipolar
A. No db-cAMPControl 60 27 � 4 59 � 5 104 � 10 189 � 18GDNF 24 28 � 3 57 � 4 139 � 14 224 � 20BDNF 35 21 � 2 55 � 7 143 � 12 219 � 19GDNF � BDNF 23 47 � 8 123 � 10 241 � 24 411 � 41*
B. db-cAMP 14 daysControl (db-cAMP) 40 51 � 11 101 � 19 173 � 25 329 � 24*GDNF 26 63 � 6 120 � 8 221 � 12 405 � 24*BDNF 23 75 � 9 150 � 14 281 � 16 505 � 39*GDNF � BDNF 28 93 � 13 186 � 20 378 � 31 657 � 48*
C. IBMX/Forskolin 14 daysGDNF � BDNF 11 265 � 40 269 � 17 419 � 35 953 � 82*
Note. Cultures were incubated in the absence or presence of db-cAMP (100 �M) for 14 days, and various combinations of GDNF (10 ng/ml), and BDNF(50 ng/ml), IBMX (100 �M) plus Forskolin (10 �M). TH-ir cells counts shown are shown as means � SEM (n equals number of coverslips).
* Significantly increased compared to control (no db-cAMP), P � 0.05.
36 J. Lara et al. / Experimental Neurology 180 (2003) 32–45
�M db-cAMP) on the cultured dopamine neurons after 14days of treatment.
Alternative methods of increasing intracellular cAMPwere also employed. These included treatments of the cul-tures with the adenyl cyclase activator, Forskolin (10 �M),and the phosphodiesterase inhibitor IBMX (100 �M), bothof which increased the survival of TH-ir neurons over theuntreated controls (Fig. 4). Their combined (Forskolin/IBMX) application for the entire 14 days culture period wasmuch more effective and comparable to that found with1000 �M db-cAMP (Fig. 4). The effects of other cAMPagonists, Sp-cAMP or 8Br-cAMP (at 200 �M), exhibitedeffects similar to that of 100 �M db-cAMP on the survivalof TH-ir neurons in the 14-day cultures. In cultures in theabsence of exogenous cAMP stimulation, 400 �M Rp-cAMP, a competitive inhibitor of cAMP, produced a small(13%) but statistically significant (P � 0.05) decrease in
TH-ir neuron survival (data not shown), suggesting involve-ment of an endogenous cAMP mechanism.
Interactive effects of BDNF, GDNF, and db-cAMP alone,and in combination on DA neuron survival
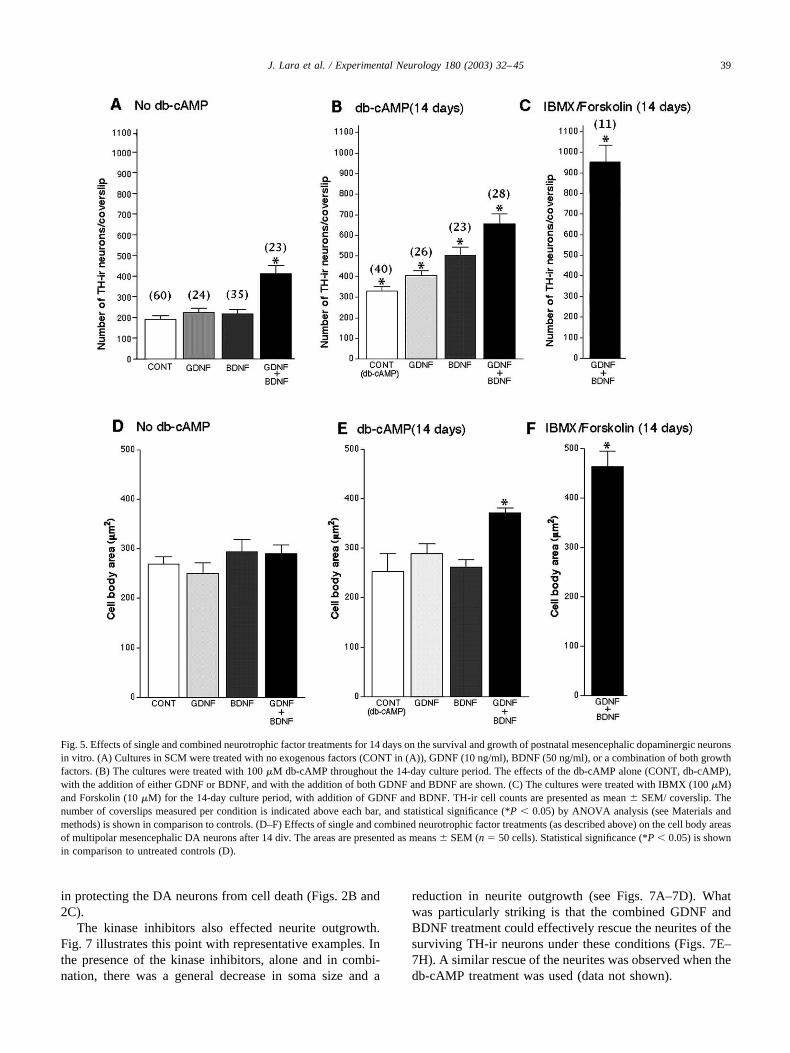
The effects of 100 �M db-cAMP, 10 ng/ml GDNF, and50 ng/ml BDNF alone and in combination on the survival ofpostnatal DA neurons in the serum-containing culture me-dium are compared in Figs. 5A–5C. We generally observedthat when presented alone the effect of 10 ng/ml GDNF wasabout the same as that of 50 ng/ml BDNF (Fig. 5A). Si-multaneous application of GDNF and BDNF showed fur-ther enhancement of Th-ir neuron survival in both the ab-sence (Fig. 5A) and the presence of db-cAMP (Fig. 5B).This increase was greatest when the growth factors werecombined with Forskolin and IBMX (Fig. 5C).
Fig. 3. Examples of the three morphological subtypes of dopamine (TH-ir) neurons present in dissociated cell cultures in SCM derived from the postnatalmesencephalon slices shown in Fig. 1. (A, B, C) Typical monopolar, bipolar, and multipolar neurons, respectively, found in control cultures after 14 div inSCM in the absence of added trophic factors. See Table 1, part A (control) for number of cells represented by each subtype. (D) Extensive multipolar, neuriteoutgrowth found in the TH-ir neurons after 60 div in control (SCM) culture medium.
37J. Lara et al. / Experimental Neurology 180 (2003) 32–45

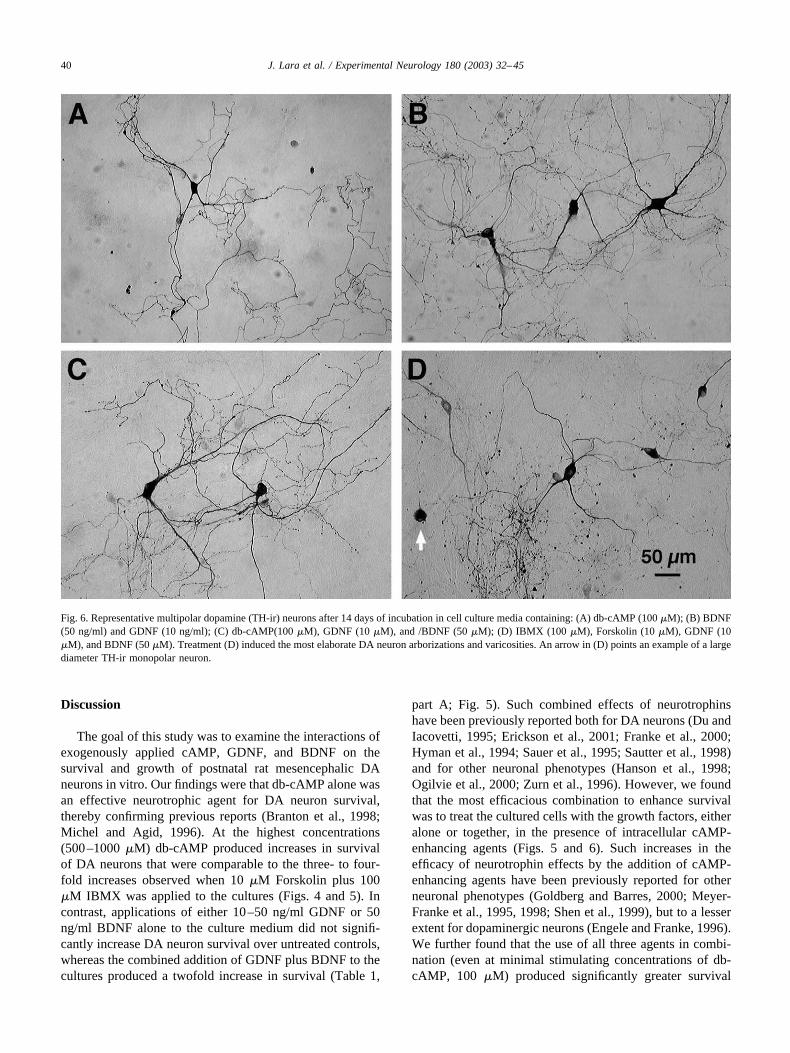
The effects of individual and combined treatments ofGDNF, BDNF, and db-cAMP on the soma areas of themultipolar TH-ir neurons are shown in Figs. 5D–5F.Individual and combined growth factor treatments did notsignificantly affect soma size (Figs. 5D and 5E). How-ever, significant increases in soma area were observedwhen the two growth factors were used together withdb-cAMP (Fig. 5E) and in the combined Forskolin/IBMX/GDNF/BDNF-treated cultures (Fig. 5F). Effectsof the above neurotrophins on the neurite lengths of themultipolar TH-ir neurons were found to be similar tothose effects on the soma areas (data not shown). Ingeneral, larger cell bodies and more elaborate neuriteswere found in the db-cAMP and IBMX/Forskolin-treatedcultures (Fig. 6).
Table 1 summarizes the effects of the various treatmentson the distribution of TH-ir neuronal subtypes. The distri-bution of morphological subtypes did not appear to changeafter treatments with growth factors and 100 �M db-cAMPfor 14 div. For example, in the presence of db-cAMP for 14days these percentages were 16, 30, and 53%, respectively,very similar to the 14, 31, and 55% found in the controlmedia (Table 1). These ratios were slightly changed to 28,28, and 44%, respectively, in the Forskolin/IBMX/BDNF/GDNF 14-day-treated cultures, largely due to the increaseof monopolar neurons that occur under these conditions (seeTable 1, part C).
Effects of protein kinase inhibitors on the survival ofpostnatal mesencephalic DA neurons in vitro
In view of the beneficial effect that increases in intracel-lular cAMP had on the survival of postnatal TH-ir neuronsin the cultures (Figs. 4–6), we next studied whether theinhibition of various downstream protein kinases could in-fluence the survival of the TH-ir neurons cultured in SCM inthe absence or presence of added neurotrophic factors (Ta-ble 2, Fig. 7). We specifically examined effects of theprotein kinase A inhibitor H89, and the inhibitor of MEK,PD98059, a critical activating kinase in the MAP kinasepathway. These inhibitor treatments were carried out duringentire cell culture period (14 d), from the time of cellplating, and renewed whenever the culture media werechanged. We found that the EC50 for the reduction in sur-vival of the DA neurons was between 10 and 20 �M forH89 and about 20 �M for PD98059.
Effects on the survival of DA neurons by 20 �M H89, 20�M PD98059, and media containing both inhibitors werecompared in control cultures, and in media containing 10ng/ml GDNF and 50 ng/ml BDNF, or 500 �M db-cAMP.These results are summarized in Table 2. The large multi-polar DA neurons were particularly vulnerable to the dele-terious effects of H89. They became swollen and monopolardue to the poor outgrowth and retraction of their neurites(Fig. 7B). Often there were many fragmented, swollen neu-rites present, which were sometimes difficult to distinguishfrom TH-ir cell bodies without neurites. Therefore, wecounted only the numbers of TH-ir bipolar and multipolarneurons for the analysis shown in Table 2. In the absence ofinhibitors the increase in survival of the DA neurons, abovethe untreated control cultures, was 215% in 10 ng/ml GDNFand 50 ng/ml BDNF-containing medium and was 188% inthe 500 �M db-cAMP-containing medium. The H89 at 20�M decreased TH-ir neuron survival to 28%, and 20 �MPD 98059 decreased the survival of the TH-ir neurons toabout 54% of control levels. Use of both inhibitors togetherproduced an additional reduction in cell survival to 10%(Table 2, part A). The effects of the above inhibitors werealso studied in the presence of the neurotrophic factorsGDNF and BDNF (Table 2, part B) and db-cAMP (Table 2,part C). Inhibitors were applied together with the combinedneurotrophic factors or 500 �M db-cAMP for the entire 14days of culture. While the 20 �M H89 and or 20 �M PD98059 significantly decreased the growth factor or db-cAMP-enhanced survival of the TH-ir neurons, there wasalso a significant protection of the DA neurons from thedeleterious effects of the inhibitors (Table 2, parts B and C).In fact, application of the PD98059 inhibitor to 500 �Mdb-cAMP-treated cultures did not significantly diminishtheir enhanced neuronal survival (Table 2, part C). Whenthe H89 and PD 98059 inhibitors were used together, thecombined GDNF- and BDNF-containing medium wasmuch more effective than the db-cAMP-containing medium
Fig. 4. Effects of various concentrations of db-cAMP and other cellularcAMP-enhancing agents (IBMX, Forskolin) on the survival of postnatalDA neurons in the dissociated cell culture. Each treatment was carriedout for the entire 14 days of culture in SCM. The data are expressed asmeans and SEMs; asterisks represent significant differences (P � 0.05)from control (Cont) values using an ANOVA test (see Materials andmethods).
38 J. Lara et al. / Experimental Neurology 180 (2003) 32–45
in protecting the DA neurons from cell death (Figs. 2B and2C).
The kinase inhibitors also effected neurite outgrowth.Fig. 7 illustrates this point with representative examples. Inthe presence of the kinase inhibitors, alone and in combi-nation, there was a general decrease in soma size and a
reduction in neurite outgrowth (see Figs. 7A–7D). Whatwas particularly striking is that the combined GDNF andBDNF treatment could effectively rescue the neurites of thesurviving TH-ir neurons under these conditions (Figs. 7E–7H). A similar rescue of the neurites was observed when thedb-cAMP treatment was used (data not shown).
Fig. 5. Effects of single and combined neurotrophic factor treatments for 14 days on the survival and growth of postnatal mesencephalic dopaminergic neuronsin vitro. (A) Cultures in SCM were treated with no exogenous factors (CONT in (A)), GDNF (10 ng/ml), BDNF (50 ng/ml), or a combination of both growthfactors. (B) The cultures were treated with 100 �M db-cAMP throughout the 14-day culture period. The effects of the db-cAMP alone (CONT, db-cAMP),with the addition of either GDNF or BDNF, and with the addition of both GDNF and BDNF are shown. (C) The cultures were treated with IBMX (100 �M)and Forskolin (10 �M) for the 14-day culture period, with addition of GDNF and BDNF. TH-ir cell counts are presented as mean � SEM/ coverslip. Thenumber of coverslips measured per condition is indicated above each bar, and statistical significance (*P � 0.05) by ANOVA analysis (see Materials andmethods) is shown in comparison to controls. (D–F) Effects of single and combined neurotrophic factor treatments (as described above) on the cell body areasof multipolar mesencephalic DA neurons after 14 div. The areas are presented as means � SEM (n � 50 cells). Statistical significance (*P � 0.05) is shownin comparison to untreated controls (D).
39J. Lara et al. / Experimental Neurology 180 (2003) 32–45
Discussion
The goal of this study was to examine the interactions ofexogenously applied cAMP, GDNF, and BDNF on thesurvival and growth of postnatal rat mesencephalic DAneurons in vitro. Our findings were that db-cAMP alone wasan effective neurotrophic agent for DA neuron survival,thereby confirming previous reports (Branton et al., 1998;Michel and Agid, 1996). At the highest concentrations(500–1000 �M) db-cAMP produced increases in survivalof DA neurons that were comparable to the three- to four-fold increases observed when 10 �M Forskolin plus 100�M IBMX was applied to the cultures (Figs. 4 and 5). Incontrast, applications of either 10–50 ng/ml GDNF or 50ng/ml BDNF alone to the culture medium did not signifi-cantly increase DA neuron survival over untreated controls,whereas the combined addition of GDNF plus BDNF to thecultures produced a twofold increase in survival (Table 1,
part A; Fig. 5). Such combined effects of neurotrophinshave been previously reported both for DA neurons (Du andIacovetti, 1995; Erickson et al., 2001; Franke et al., 2000;Hyman et al., 1994; Sauer et al., 1995; Sautter et al., 1998)and for other neuronal phenotypes (Hanson et al., 1998;Ogilvie et al., 2000; Zurn et al., 1996). However, we foundthat the most efficacious combination to enhance survivalwas to treat the cultured cells with the growth factors, eitheralone or together, in the presence of intracellular cAMP-enhancing agents (Figs. 5 and 6). Such increases in theefficacy of neurotrophin effects by the addition of cAMP-enhancing agents have been previously reported for otherneuronal phenotypes (Goldberg and Barres, 2000; Meyer-Franke et al., 1995, 1998; Shen et al., 1999), but to a lesserextent for dopaminergic neurons (Engele and Franke, 1996).We further found that the use of all three agents in combi-nation (even at minimal stimulating concentrations of db-cAMP, 100 �M) produced significantly greater survival
Fig. 6. Representative multipolar dopamine (TH-ir) neurons after 14 days of incubation in cell culture media containing: (A) db-cAMP (100 �M); (B) BDNF(50 ng/ml) and GDNF (10 ng/ml); (C) db-cAMP(100 �M), GDNF (10 �M), and /BDNF (50 �M); (D) IBMX (100 �M), Forskolin (10 �M), GDNF (10�M), and BDNF (50 �M). Treatment (D) induced the most elaborate DA neuron arborizations and varicosities. An arrow in (D) points an example of a largediameter TH-ir monopolar neuron.
40 J. Lara et al. / Experimental Neurology 180 (2003) 32–45

41J. Lara et al. / Experimental Neurology 180 (2003) 32–45

(Figs. 5A and 5B), and maximal cAMP stimulation (inIBMX plus Forskolin) produced the greatest survival of theDA neurons (Fig. 5C, summarized in Table 1). Similarpatterns of action of the neurotrophic agents were seen intheir effects on perikaryal soma size (Figs. 5D–5F, Fig. 6).
We performed the above experiments in SCM, since inour experience (Fig. 2) and that of others (Cardozo, 1993;Masuko et al., 1992; Michel and Agid, 1996; Rayport et al.,1992), the presence of horse serum in the medium greatlyimproves the survival of the DA neurons. However, itshould be noted here that we found similar enhancing ef-fects of cAMP and GDNF and BDNF and their combina-tions on the survival of the DA neurons in SFM (not illus-trated). Although the responses are smaller in SFM than
those in SCM, the general response patterns are similar inthe two conditions. Hence, the interaction of unknown se-rum factors with these agents appears to be supportive ofbut not essential for their effects. The inclusions of thesegrowth factors and db-cAMP in the serum-free culture me-dium also improved the growth and survival of nonneuronalcells in the culture, and this could also be indirectly con-tributing to the improved growth and survival of the TH-irneurons.
While there is clear evidence for the value of cAMP,GDNF, and BDNF as survival factors for mesencepalic DAneurons, there is less understanding about the subsequentsignaling and downstream mechanisms that are involved.Brain-derived neurotrophic factor and GDNF activate dif-ferent receptor systems (Baloh et al., 2000; Lewin andBarde, 1996; Unsicker and Krieglstein, 2000; Van Weeringand Bos, 1998), TrkB is the specific tyrosine kinase receptorfor BDNF, whereas GDNF interacts with another specificcell-surface receptor, GFR�-1, whose biological effects aremediated through the interaction of a multicomponent re-ceptor complex of GFR�-1 and another tyrosine kinasereceptor, Ret (Airaksinen et al., 1999; Jing et al., 1996;Treanor et al., 1996; Trupp et al., 1999). Both TrkB and Rettyrosine kinase receptors are able to activate a mitogen-activated protein kinase (MAPK) pathway (Marsh et al.,1993; Worby et al., 1996) which can converge with cAMP-activated PKA pathways to phosphorylate and activate thecyclic-AMP responsive element-binding protein (CREB),which in turn is well known to promote neuronal growthand survival (Bonni et al., 1999; Finkbeiner, 2000; Riccio etal., 1999; Walton and Dragunow, 2000). Thus, the enhancedeffects of the combined neurotrophins and cAMP on DAneuron survival could be a consequence of the synergisticcrosstalk known to occur between these signaling pathways(Bornfeldt and Krebs, 1999; Chang and Karin, 2001; Rich-ards, 2001; Shaywitz and Greenberg, 1999; Walton andDragunow, 2000). In addition, the increase of cAMP levelsin neurons activates protein kinase A-dependent signalingpathways, which could then lead to increased GDNF orBDNF receptor densities on the plasma membrane by re-cruitment from intracellular sites (Engele and Franke, 1996;Goldberg and Barres, 2000; Meyer-Franke et al., 1998).
While the details of the cellular signaling pathways
Table 2Effects of protein kinase inhibitors on the survival of postnatalmesencephalic dopaminergic neurons
Conditiona %Survivalb
mean � SEMn
AUntreated control cultures 100 � 13 42�20 �M H89 28 � 5c 16�20 �M PD98059 54 � 7c 23�20 �M H89 � 20�M PD98059 10 � 4c 14
BGDNF � BDNF-treated cultures 215 � 14 23�20 �M H89 86 � 10c 10�20 �M PD98059 146 � 15c 9�20 �M H89 � 20 �M PD98059 76 � 7c 15
Cdb-cAMP-treated cultures 181 � 31c 28�20 �M H89 73 � 9c 16�20 �M PD98059 181 � 16 21�20 �M H89 � 20 �M PD98059 25 � 5c 16
a Each treatment was carried out for the entire 14 days of the cultureperiod. Concentrations of growth factors used were 10 ng/ml GDNF, 50ng/ml BDNF, and 500 �M db-cAMP.
b The means of bipolar and multipolar TH� neurons in the untreatedcontrol cultures in these experiments was taken as 100% survival. Eachcondition had two to five independent culture sessions with matchedcontrols, and with n equaling the total number of experimental coverslipsin each experiment.
c Statistically significant differences of inhibitor-treated from respectiveinhibitor-untreated (control) groups in the same culture session as shown inparts A, B, and C, P � 0.05.
Fig. 7. Representative illustrations of the effects of H89 and PD98059 on the growth and survival of TH-ir neurons in control (A–D) and GDNF plus BDNF(E–H)-containing culture media. Each treatment was carried out for the entire 14 days of culture. (A) TH-ir neurons after 14 days in control culture medium.(B) Effects of 20 �M H-89, showing loss of bipolar and multipolar TH-ir neurons leaving only monopolar TH-ir neurons with greatly reduced neurites. (C)Effects of 20 �M PD98059, also showing loss of bipolar and multipolar TH-ir neurons and reduction of neurites but to a lesser extent than in theH89-contained medium (see (B)) (D) A TH-ir neuron with a reduced soma size and a single, thin neurite observed among the few surviving TH-ir neuronsin the H89- and PD98059-containing medium (see Table 2, part A). (E) TH-ir neurons grown in BDNF (50 ng/ml) � GDNF (10 ng/ml)-containing medium.Somas of TH-ir neurons are larger and their neurites are more elaborate than in the controls (see (A)). (F) TH-ir neurons grown in 20 �M H89- plus GDNF-and BDNF-containing medium. The deleterious effects of the 20 �M H89 on neurite outgrowth (see (B)) were mitigated by the simultaneous presence ofthe growth factors. (G) TH-ir neurons grown in 20 �M PD98059- plus GDNF- and BDNF-containing medium. The deleterious effects of the 20 �M PD98059on neurite outgrowth in the control medium (see (C)) were significantly reduced by the simultaneous presence of the growth factors. (H) TH-ir neurons grownin 20 �M H89- and 20 �M PD98059- plus GDNF- and BDNF-containing medium. The combined GDNF and BDNF treatment significantly effected thecombined effects of H89 � PD98059 in that the surviving TH-ir neurons had large somas and more robust neurites.
42 J. Lara et al. / Experimental Neurology 180 (2003) 32–45

remain to be determined, there appears to be a generalconsensus not only that more DA neurons are activatedby the combined application of different neurotrophins,but also that the distinct signaling pathways activated bydifferent neurotrophins act cooperatively to support thesurvival of DA neurons by ultimately phosphorylatingMAPK. The activated MAPK could contribute to in-creased survival by induction of the antiapoptic proteinsBcl-2 and Bcl-XL, or by inhibiting the proapoptic pro-teins BAD or BAX (Berridge et al., 1993; Bonni et al.,1999; Franke et al., 2000; Harada et al., 1999; Riccio etal., 1999; Sautter et al., 1998). Activation of cAMP-dependent protein kinase A by db-cAMP and other cel-lular cAMP-elevating agents (e.g., Forskolin, IBMX)could also inactivate the proapoptic molecule BAD(Harada et al., 1999), and promote DA neuron survival.Consistent with this view we found that the inhibitor H89decreased TH-ir neuron survival (Table 2). It has alsobeen shown that activation of protein kinase A activatesthe serine–threonine kinase B-raf, which in turn activatesMAP kinase that in turn promotes the cell survival (Ric-cio et al., 1999; Vossler et al., 1997). Our finding of areduced TH-ir neuron survival by the MEK inhibitorPD98059 (Table 2) is also consistent with the aboveview. Overall, our data suggest that the combined appli-cation of GDNF, BDNF, and db-cAMP might activateboth the MAP kinase and the PKA signaling pathways,which together maximally promote the survival andgrowth of DA neurons in vitro. In the present study, themost dramatic increase in number and size of TH-irneurons was seen in the combined Forskolin; IBMX-,GDNF-, and BDNF-treated culture.
It has previously been demonstrated that GDNF is aneffective neurotrophic factor for midbrain DA neurons invivo (Beck et al., 1995; Tomac et al., 1995), and thuswould seem to be an efficacious candidate for growthfactor-based therapy of Parkinson’s disease (Bjorklund etal., 1997; Bowenkamp et al., 1995; Gash et al., 1998). Wefind that the ability of GDNF alone effect to promote thesurvival of cultured postnatal dopaminergic neurons ap-pears to be minimal (Fig. 5A). However, the combinedtreatment using GDNF, BDNF, and db-cAMP greatlyinfluences not only survival but also cell body size andneurite outgrowth of DA neurons. This suggests multiplesignaling pathways may be involved in maintaining thewell-being of DA neurons and the possibility that undersome conditions degenerating DA neurons in vivo mightbe rescued even more effectively by the combined treat-ment, as opposed to GDNF alone.
Acknowledgments
We thank Drs. Heather Cameron and Bing-jun Zhang forhelp in using the Neurolucida program.
References
Airaksinen, M.S., Titievsky, A., Saarma, M., 1999. GDNF family neuro-trophic factor signaling: four masters, one servant? Mol. Cell Neurosci.13, 313–325.
Alexi, T., Borlongan, C.V., Faull, R.L.M., Williams, C.E., Clark, R.G.,Gluckman, P.D., Hughes, P.E., 2000. Neuroprotective strategies forbasal ganglia degeneration: Parkinson’s and Huntington’s diseases.Prog. Neurobiol. 60, 409–470.
Baloh, R.H., Enomoto, H., Johnson Jr., E.M., Milbrandt, J., 2000. TheGDNF family ligands and receptors—Implications for neural develop-ment. Curr. Opin. Neurobiol. 10, 103–110.
Beck, K.D., Valverde, J., Alexi, T., Poulsen, K., Moffat, B., Vandlen, R.A.,Rosenthal, A., Hefti, A., 1995. GDNF protects mesencephalic dopa-minergic neurons from axotomy-induced degeneration in the adultbrain. Nature 373, 339–341.
Berridge, M.V., Tan, A.S., Hilton, C.J., 1993. Cyclic adenosine monophos-phate promotes cell survival and retards apoptosis in a factor-dependentbone marrow-derived cell line. Exp. Hematol. 21, 269–276.
Bjorklund, A., Rosenblad, C., Winkler, C., Kirik, D., 1997. Studies onneuroprotective and regenerative effects of GDNF in a partial lesionmodel of Parkinson’s disease. Neurobiol. Dis. 4, 186–200.
Bonni, A, Brunet, A., West, A.E., Datta, S.R., Takasu, M.A., Greenberg,M.E., 1999. Cell survival promoted by the Ras-MAPK signaling path-way by transcription-dependent and -independent mechanisms. Science286, 1358–1362.
Bornfeldt, K.E., Krebs, E.G., 1999. Crosstalk between protein kinase A andgrowth factor receptor signaling pathways in arterial smooth muscle.Cell Signal. 11, 465–477.
Bowenkamp, K.E., Hoffman, A.F., Gerhardt, G.A., Henry, M.A., Biddle,P.T., Hoffer, B.J., Granholm, A.E., 1995. Glial cell line-derived neu-rotrophic factor supports survival of injured midbrain dopaminergicneurons. J. Comp. Neurol. 355, 479–489.
Branton, R.L., Love, R.M., Clarke, D.J., 1998. cAMP included during cellsuspension preparation improves survival of dopaminergic neurons invitro. NeuroReport 9, 3223–3227.
Brewer, J., Torricelli, J.R., Evege, E.K., Price, P.J., 1993. Optimizedsurvival of hippocampal neurons in B27-supplemented Neurobasal, anew serum-free medium combination. J. Neurosci. Res. 35, 567–576.
Burke, R.E., Antonelli, M., Sulzer, D., 1998. Glial cell line-derived neu-rotrophic growth factor inhibits apoptotic death of postnatal substantianigra dopamine neurons in primary culture. J. Neurochem. 71, 517–525.
Cardozo, D.L., 1993. Midbrain dopaminergic neurons from postnatal rat inlong-term primary culture. Neuroscience 56, 409–421.
Chang, L., Karin, M., 2001. Mammalian MAP kinase signaling cascades.Nature 410, 37–40.
Choi-Lundberg, D.L., Lin, Q., Chang, Y.-N., Chiang, Y.L., Hay, C.M.,Mohajeri, H., Davidson, B.L., Bohn, M.C., 1997. Dopaminergic neu-rons protected from degeneration by GDNF gene therapy. Science 275,838–841.
Clarkson, E.D., Zawada, W.M., Freed, C.R., 1995. GDNF reduces apopto-sis in dopaminergic neurons in vitro. NeuroReport 7, 145–149.
Collier, T.J., Sortwell, C.E., 1999. Therapeutic potential of nerve growthfactors in Parkinson’s disease. Drugs Aging 14, 261–287.
Du, X., Iacovetti, L., 1995. Synergy between growth factors and transmit-ters required for catecholamine differentiation in brain neurons. J. Neu-rosci. 15, 5420–5427.
Dunnett, S.B., Bjorklund, A., 1999. Prospects for new restorative andneuroprotective treatments in Parkinson’s disease. Nature 399, A32–A39.
Engele, J., Franke, B., 1996. Effects of glial cell line-derived neurotrophicfactor (GDNF) on dopaminergic neurons require concurrent activationof cAMP-dependent signaling pathways. Cell Tissue Res. 286, 235–240.
43J. Lara et al. / Experimental Neurology 180 (2003) 32–45

Erickson, J.T., Brosenstsch, T.A., Katz, D.M., 2001. Brain-derived neuro-trophic factor and glial line-derived neurotrophic factor are requiredsimultaneously for survival of dopaminergic primary sensory neuronesin vivo. J. Neurosci. 21, 581–589.
Feng, L., Wang, C.Y., Jiang, H., Oho, C., Dugich-Djordjevic, M., Mei, L.,Lu, B., 1999. Differential signaling of glial cell line-derived neurotro-phic factor and brain-derived neurotrophic factor in cultured ventralmesencephalic neurons. Neuroscience 93, 265–273.
Finkbeiner, S., 2000. CREB couples neurotrophin signals to survival mes-sages. Neuron 25, 11–14.
Franke, B., Bayatti, N., Engele, J., 2000. Neurotrophins require distinctextracellular signals to promote the survival of CNS neurons in vitro.Exp. Neurol. 165, 125–135.
Gash, D.M., Zhang, Z., Gerhardt, G., 1998. Neuroprotective and neuro-restorative properties of GDNF. Ann. Neurol. 44, S121–S125.
Goldberg, J.L., Barres, B.A., 2000. The relationship between neuronalsurvival and regeneration. Annu. Rev. Neurosci. 23, 579–612.
Hanson Jr., M.G., Shen, S., Wiemelt, A.P., McMorris, F.A., Barres, B.A.,1998. Cyclic AMP elevation is sufficient to promote the survival ofspinal motor neurons in vitro. J. Neurosci. 18, 7361–7371.
Harada, H., Becknell, B., Wilm, M., Mann, M., Huang, L.J.-S., Taylor,S.S., Scott, J.D., Korsmeyer, S.J., 1999. Phosphorylation and inactiva-tion of BAD by mitochondria-anchored protein kinase A. Mol. Cell. 3,413–422.
Hartikka, J., Staufenbiel, M., Lubbert, H., 1992. Cyclic AMP, but not basicFGF, increases the in vitro survival of mesencephalic dopaminergicneurons and protects them from MPP�-induced degeneration. J. Neu-rosci. Res. 32, 190–201.
Hyman, C., Hofer, M., Barde, Y.A., Juhasz, M., Yancopoulos, G.D.,Squinto, S.P., Lindsay, R.M., 1991. BDNF is a neurotrophic factor fordopaminergic neurons of the substantia nigra. Nature 350, 230–232.
Hyman, C., Juhasz, M., Jackson, C., Wright, P., Ip, N.Y., Lindsay, R.M.,1994. Overlapping and distinct actions of the neurotrophins BDNF,NT-3, NT-4/5, on cultured dopaminergic and GABAergic neurons ofthe ventral mesencephalon. J. Neurosci. 14, 335–347.
Jing, S., Wen, D., Yu, Y., Holst, P.L., Luo, Y., Fan, M., Tamir, R.,Antonio, L., Hu, Z., Cupples, R., Louis, J.C., Hu, S., Altrock, B.W.,Fox, G.M., 1996. GDNF-induced activation of the Ret protein tyrosinekinase is mediated by GDNFR-�, a novel receptor for GDNF. Cell 85,1113–1124.
Jing, S., Yu, Y., Fanf, M., Hu, Z., Holst, P.L., Boone, T., Delaney, J.,Schultz, H., Zhou, R., Fox, G.M., 1997. GFR �-2 and GFR �-3 are twonew receptors for ligands of the GDNF family. J. Biol. Chem. 272,33111–33117.
Knusel, B., Winslow, J.W., Rosenthal, A., Burton, L.E., Seid, D.P., Ni-kolics, K., Hefti, F., 1991. Promotion of central cholinergic and dopa-minergic neuron differentiation by brain-derived neurotrophic factorbut not neurotrophin 3. Proc. Natl. Acad. Sci. USA 88, 961–965.
Kordower, J.H., Emborg, M.E., Bloch, J., Ma, S.Y., Chu, Y., Leventhal, L.,McBride, J., Chen, E.Y., Palfi, S., Roitberg, B.Z., Brown, W.D.,Holden, J.E., Pyzalski, R., Taylor, M.D., Carvey, P., Ling, Z., Trono,D., Hatraye, P., Deglon, N., Aebisher, P., 2000. Neurodegenerationprevented by lentiviral vector delivery of GDNF in primate model ofParkinson’s disease. Science 290, 767–772.
Lewin, G.R., Barde, Y., 1996. Physiology of the neurotrophins. Annu. Rev.Neurosci. 19, 289–317.
Lin, L.-F.H., Doherty, D.H., Lile, J.D., Bektesh, S., Collins, F., 1993.GDNF: a glial cell line-derived neurotrophic factor for midbrain do-paminergic neurons. Science 260, 1130–1132.
Marsh, H.N., Scholz, W.K., Lamballe, F., Klein, R., Nanduri, V., Barbacid,M., Palfrey, H.C., 1993. Signal transduction events mediated by theBDNF receptor gp145trkB in primary hippocampal pyramidal cellculture. J. Neurosci. 13, 4281–4292.
Masuko, S., Nakajima, S., Nakajima, Y., 1992. Dissociated high-puritydopaminergic neuron cultures from the substantia nigra and the ventraltegmental area of the postnatal rat. Neuroscience 49, 347–364.
Matsuo, A., Nakamura, S., Akiguchi, I., 2000. Immunohistochemical lo-calization of glial cell line-derived neurotrophic factor family receptor�-1 in the rat brain: confirmation of expression in various neuronalsystems. Brain Res. 859, 57–71.
Merlio, J.-P., Ernfors, P., Jaber, M., Persson, H., 1992. Molecular cloningof rat trkC and distribution of cells expressing messenger RNAs formembers of the trk family in the rat central nervous system. Neuro-science 51, 513–532.
Meyer-Franke, A., Kaplan, M.R., Pfrieger, F.W., Barres, B.A., 1995.Characterization of the signaling interactions that promote the survivaland growth of developing retinal ganglion cells in culture. Neuron 15,805–819.
Meyer-Franke, A., Wilkinson, G.A., Kruttgen, A., Hu, M., Munro, E.,Hanson Jr., M.G., Reichardt, L.F., Barres, B.A., 1998. Depolarizationand cAMP elevation rapidly recruit TrkB to the plasma membrane ofCNS neurons. Neuron 21, 681–693.
Michel, P.P., Agid, Y., 1996. Chronic activation of the cyclic AMP sig-naling pathway promotes development and long-term survival of mes-encephalic dopaminergic neurons. J. Neurochem. 67, 1633–1642.
Nosrat, C.A., Tomac, A., Hoffer, B.J., Olson, L., 1997. Cellular anddevelopmental patterns of expression of Ret and glial line-derivedneurotrophic factor receptor alpha mRNAs. Exp. Brain Res. 115, 410–422.
Ogilvie, J.M., Speck, J.D., Lett, J.M., 2000. Growth factors in combination,but not individually, rescue rd mouse photoreceptors in organ culture.Exp. Neurol. 161, 676–685.
Oo, T.F., Burke, R.E., 1997. The time course of developmental cell deathin phenotypically defined dopaminergic neurons of the substantia nigra.Dev. Brain Res. 98, 191–196.
Pochon, N.A.-M., Menoud, A., Tseng, J.L., Zurn, A.D., Aebischer, P.,1997. Neuronal GDNF expression in the adult rat nervous systemidentified by in situ hybridization. Eur. J. Neurosci. 9, 463–471.
Rayport, S., Sulzer, D., Shi, W.-X., Sawasdikosol, S., Monaco, J., Batson,D., Rajendran, G., 1992. Identified postnatal mesolimbic dopamineneurons in culture: morphology and electrophysiology. J. Neurosci. 12,4264–4280.
Riccio, A., Ahn, S., Davenport, C.M., Blendy, J.A., Ginty, D.D., 1999.Mediation by a CREB family transcription factor of NGF-dependentsurvival of sympathetic neurons. Science 286, 2358–2361.
Richards, J.S., 2001. New signaling pathways for hormones and cyclicadenosine 3�,5�-monophosphate action in endocrine cells. Mol. Endo-crinol. 15, 209–218.
Sauer, H., Rosenblad, C., Bjorklund, A., 1995. Glial cell line-derivedneurotrophic factor but not transforming growth factor 3 preventsdelayed degeneration of nigral dopaminergic neurons following striatal6-hydroxydopamine lesion. Proc. Natl. Acad. USA 92, 8935–8939.
Sautter, J., Meyer, M., Spenger, C., Seiler, R.W., Widmer, H.R., 1998.Effects of combined BDNF and GDNF treatment on cultured dopami-nergic midbrain neurons. NeuroReport 9, 1093–1096.
Schatz, D.S., Kaufmann, W.A., Saria, A., Humpel, C., 1999. Dopamineneurons in a simple GDNF-treated meso-striatal organotypic co-culturemodel. Exp. Brain Res. 127, 270–278.
Seroogy, K.B., Gall, C.M., 1993. Expression of neurotrophins by midbraindopaminergic neurons. Exp. Neurol. 124, 119–128.
Shaywitz, A.J., Greenberg, M.E., 1999. CREB: a stimulus-induced tran-scription factor activated by a diverse array of extracellular signals.Annu. Rev. Biochem. 68, 821–861.
Shen, S., Wiemelt, A.P., McMorris, F.A., Barres, B.A., 1999. Retinalganglion cells lose trophic responsiveness after axotomy. Neuron 23,285–295.
Shults, C.W., Kimber, T., Altar, C.A., 1995. BDNF attenuates the effectsof intrastriatal injection of 6-hydroxydopamine. NeuroReport 6, 1109–1112.
Sinclair, S.R., Svendsen, C.N., Torres, E.M., Martin, D., Fawcett, J.W.,Dunett, S.B., 1996. GDNF enhances dopaminergic cell survival andfibre outgrowth in embryonic nigral grafts. NeuroReport 7, 2547–2552.
44 J. Lara et al. / Experimental Neurology 180 (2003) 32–45

Studer, L., Spenger, C., Seiler, J.R.W., Altar, C.A., Lindsay, R.M., Hyman,C., 1995. Comparison of the effects of the neurotrophins on the mor-phological structure of dopaminergic neurons in cultures of rat sub-stantia nigra. Eur. J. Neurosci. 7, 223–233.
Takahashi, M., Buma, Y., Imoto, T., Inaguma, Y., Ikeda, H., Hiai, H.,1988. Cloning and expression of the ret proto-oncogene encoding atyrosine kinase with two potential transmembrane domains. Oncogene3, 571–578.
Tomac, A., Lindquist, E., Lin, L.F., Ogren, D., Young, D., Hoffer, B.J.,Olson, L., 1995. Protection and repair of the nigrostriatal dopaminergicsystem by GDNF in vivo. Nature 373, 335–339.
Treanor, J.J.S., Goodman, L., de Sauvage, F., Stone, D.M., Poulsen, K.T.,Beck, C.D., Gray, C., Pollock, R.A., Hefti, F., Philips, H., Goddard, S.,Moore, M.W., Buj-Bello, A., Davies, A.M., Asai, N., Takahashi, M.,Vandlen, R., Henderson, C.E., Rosenthal, A., 1996. Characterization ofa multicomponent receptor for GDNF. Nature 382, 80–83.
Trupp, M., Arenas, E., Fainzilber, M., Nilsson, A.-S., Sieber, B.-A., Grigo-rior, M., Kilkenny, C., Salazar-Grueso, E., Pachnis, V., Arumae, U.,Sariola, H., Saarma, M., Ibanez, C.F., 1996. Functional receptor forGDNF encoded by the c-ret proto-oncogene. Nature 381, 785–789.
Trupp, M., Belluardo, N., Funakoshi, H., Ibanez, C.F., 1997. Complemen-tary and overlapping expression of glial cell line-derived neurotrophicfactor (GDNF), c-ret proto-oncogene, and GDNF receptor-� indicatesmultiple mechanisms of trophic actions in the adult rat CNS. J. Neu-rosci. 17, 3554–3567.
Trupp, M., Scott, R., Whittemore, S.R., Ibanez, C.F., 1999. Ret-dependentand independent mechanisms of glial cell line-derived neurotrophicfactor signaling in neuronal cells. J. Biol. Chem. 274, 20885–20894.
Tsukahara, T., Takeda, M., Shimohama, S., Ohara, O., Hashimoto, N.,1995. Effects of brain-derived neurotrophic factor on 1-methyl-4-phe-nyl-1,2,3,6-tetrahydropyridine-induced parkinsonism in monkeys.Neurosurgery 37, 733–739.
Unsicker, K., 1994. Growth factors in Parkinson’s disease. ProgressGrowth Factor Res. 5, 73–87.
Unsicker, K., Krieglstein, K., 2000. Co-activation of TGF-ss and cytokinesignaling pathways are required for neurotrophic functions. CytokineGrowth Factor Rev. 11, 97–102.
van Weering, D.H., Bos, J.L., 1998. Signal transduction by the receptortyrosine kinase. Ret. Recent Results Cancer Res. 154, 271–281.
Vossler, M.R., Yao, H., York, R.D., Pan, M.-G., Rim, C.S., Stork, P.J.S.,1997. cAMP activates MAP kinase and Elk-1 through a B-Raf- andRap1-dependent pathway. Cell 89, 73–82.
Walton, M.R., Dragunow, I., 2000. Is CREB a key to neuronal survival?Trends Neurosci. 23, 48–53.
Worby, C.A., Vega, Q.C., Zhao, Y., Chao, H.H.-J., Seasholtz, A.F., Dixon,J.E., 1996. Glial cell line derived neurotrophic factor signals throughthe RET receptor and activates mitogen-activated protein kinase.J. Biol. Chem. 271, 23619–23622.
Yan, Q., Wang, J., Matheson, C.R., Urich, J.L., 1999. Glial cell line-derivedneurotrophic factor (GDNF) promotes the survival of axotomized retinalganglion cells in adult rats: comparison to and combination with brain-derived neurotrophic factor (BDNF). J. Neurobiol. 38, 382–390.
Zurn, A.D., Winkel, L., Menoud, A., Djabali, K., Aebischer, P., 1996.Combined effects of GDNF, BDNF, and CNTF on motoneuron differ-entiation in vitro. J. Neurosci. Res. 44, 133–141.
45J. Lara et al. / Experimental Neurology 180 (2003) 32–45





























