Embed Size (px)
Citation preview

Journal of Neuroimmunology 105 (2000) 103–108www.elsevier.com/ locate / jneuroin
Interleukin-4-dependent induction of preproenkephalin in antigen-specificT helper-type 2 (Th2) cells
a,b a a,c a,c aTakashi Yahata , Chie Yahata , Akio Ohta , Masashi Sekimoto , Hidemitsu Kitamura ,a,b b d a a cKenji Iwakabe , Sonoko Habu , Shinzi Azuma , Minoru Nakui , Marimo Sato , Toshiaki Koda ,
a,c ,*Takashi NishimuraaSection of Genetic Engineering, Research Center for Genetic Engineering and Cell Transplantation, Tokai University School of Medicine, Bohseidai,
Isehara 259-1193, JapanbDepartment of Immunology, Tokai University School of Medicine, Bohseidai, Isehara 259-1193, Japan
cSection of Bacterial Infection, Institute of Immunological Science, Hokkaido University, Sapporo 060-0815, JapandResearch & Development, Eicom Corporation, Kyoto 612-8474, Japan
Received 13 May 1999; received in revised form 15 December 1999; accepted 23 December 1999
Abstract
Naive Th cells obtained from OVA -specific DO11.10 TCR-Tg mice did not express preproenkephalin (PPE) mRNA. However,323–339
culture of naive Th cells with OVA peptide (OVA-pep) plus IL-2 under Th2-inducing conditions for 7 days resulted in an induction323–339
of PPE mRNA. The PPE mRNA was also induced by culturing with OVA-pep plus IL-2 (neutral condition). However, PPE mRNAinduction under neutral conditions was totally abrogated by addition of anti-IL-4 mAb. The existence of methionine-enkephalin was alsodemonstrated in peptidase-digested peptides derived from Th2 cell lysate. These results demonstrate that IL-4 is a critical factor for theinduction of PPE mRNA in freshly expanded antigen-specific Th2 cells. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Th1/Th2; Preproenkephalin; IL-4; RDA
1. Introduction DNA (cDNA) specifically expressed in Th1 or Th2 cellsby a representational difference analysis (RDA) method
It has been demonstrated that cellular and humoral (Diatchenko et al., 1996). As a result, preproenkephalinimmunity are critically regulated by the balance between (PPE) cDNA was cloned from a cDNA library preparedtwo distinct Th subsets, Th1 and Th2 cells, (Th1/Th2 from the messenger RNA (mRNA) of Th2 cells. PPE is abalance) through the production of cytokines (Mosmann precursor protein of enkephalins, which act upon nerveand Sad, 1996; Ohta et al., 1997). Generally, Th1 and Th2 cells expressing the receptor and regulates many physio-cells cross-regulate each other to maintain a balance. For logical functions, including pain perception and analgesiaexample, Th1-derived IFN-g blocks the production of IL-4 (Wybran et al., 1987; Konig et al., 1996). Therefore, ifby Th2 cells (Hsieh et al., 1993). Conversely, Th2-derived PPE and its processed opioid peptides were specificallyIL-4 inhibits the production of IFN-g by Th1 cells (Hsieh expressed in Th2 cells, one could speculate that Th1/Th2et al., 1992). It has been proposed that an imbalance balance may play an important role in neuroimmunology.between Th1 and Th2 cells may underlie the development The expression of PPE mRNA in Th cells was firstlyof many immune diseases (Librau et al., 1995; O’Garra reported by Zurawski et al. (1986). Moreover, Cherwinskiand Murphy, 1995). Therefore, it is of importance to define et al. (1987) suggested that PPE mRNA is stronglyunique functional molecules selectively expressed in Th1 expressed in long-term cultured Th2 cell clones thoughor Th2 cells. some Th1 clones also expressed it. However, it remains
For this purpose, we tried to isolate complementary unclear whether primary-induced antigen-specific Th2 cellsexpress PPE mRNA and how PPE mRNA is induced inTh2 cells from naive Th cells. Thus, we examined theexpression and the regulation of PPE mRNA in antigen-*Corresponding author. Tel. / fax: 181-11-707-6835.
E-mail address: [email protected] (T. Nishimura) specific naive or activated Th cell subsets obtained from
0165-5728/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0165-5728( 00 )00188-0

104 T. Yahata et al. / Journal of Neuroimmunology 105 (2000) 103 –108
OVA-specific DO11.10 T cell receptor transgenic (TCR- CD4 mAb (Pharmingen), fixed by 4% paraformaldehyde,Tg) mice. and permeabilized by 0.5% Triton X-100, 5 mM EDTA,
In this paper, we demonstrate that: (i) PPE mRNA is 50 mM NaCl (pH 7.5). Then, intracellular cytokines werehighly expressed in freshly induced OVA-specific Th2 stained with FITC-conjugated anti-IFN-g mAb and PE-cells, but not in Th1 cells; (ii) the induction of PPE mRNA conjugated anti-IL-4 mAb (Pharmingen).in Th2 cells is totally dependent on IL-4; and (iii)physiologically active methionine-enkephalin (Met-Enk) 2.3. mAb blockingwas detected in Th2 cell lysate after digestion with
1peptidases. Thus, this paper extends previous findings CD4 T cells were cultured under neutral conditions(Zurawski et al., 1986; Cherwinski et al., 1987) and further (APC, OVA peptide and IL-2) with or without anti-323–339proposes that Th1/Th2 balance may play an important role IL-4 mAb (50 mg/ml), anti-IL-10 mAb (5 mg/ml; JES5-in interconnections between the nervous and immune 2A5, Pharmingen) or anti-IFN-g mAb (50 mg/ml). After 7systems. days of culture, cells were harvested and analyzed by
Northern hybridization as described below.
2. Materials and methods2.4. mRNA preparation
2.1. MiceTh1 and Th2 cells were treated with 20 ng/ml of
phorbol 12-myristate 13-acetate (PMA; purchased fromOVA -specific DO11.10 TCR-Tg mice maintained 1323–339 Sigma) for 2 h and poly(A) mRNA was purified with the
on a BALB/c background were kindly donated by DrFastTrack 2.0 kit (Invitrogen, San Diego, CA,USA) ac-
K.M. Murphy (Washington University School of Medicine,cording to the manufacturer’s recommendation.
St. Louis, MO, USA) (Murphy et al., 1990) and main-tained in our animal center. All animals were used at 5–8
2.5. cDNA subtraction and library screeningweeks of age.
Complementary DNAs were synthesized from 2 mg of2.2. Induction of Th1 and Th2 cells from DO11.101poly(A) RNA, and subtractive hybridization was per-TCR-Tg mice
formed using a PCR-Select cDNA Subtraction kit (Clon-tech, Palo Alto, CA, USA) as previously described (Diat-Th1 cells were induced from FACSvantage (Becton-
1 1 chenko et al., 1996). Representative cDNAs were preparedDickinson, San Jose, CA, USA)-sorted CD4 CD45RBfrom RsaI restriction enzyme digestion of cDNAs. Th2naive T cells obtained from DO11.10 TCR-Tg mousecDNA was ligated to adapter 1 or 2R (Clontech) and bothspleen cells by culturing with 10 mg/ml of OVA323–339of the adapter ligated Th2 cDNAs were hybridized withpeptide (kind gift of Fujiya, Hadano, Japan), 100 U/ml ofTh1 cDNA at 688C. Eight hours later, both reactionIL-2 (kind gift of Shionogi Pharmaceutical, Osaka, Japan),mixtures were combined and incubated at 688C for aIL-12 (20 U/ml; kindly donated by Genetics Institute,further 20 h. Subtracted Th2-specific cDNAs were am-MA), IFN-g (1 ng/ml; Pharmingen, San Diego, CA, USA)plified by PCR using adapter-specific primer and thenand anti IL-4 mAb (50 mg/ml; 11B11, ATCC, Rockville,
1 directly cloned into T/A vector (Invitrogen). A clonedMD, USA). Th2 cells were induced from naive CD4 Tsubtracted Th2-specific cDNA library was screened bycells by culturing with OVA peptide, IL-2, IL-4 (1323–339hybridization with cDNA probes from Th1 and Th2. Theng/ml; Wako Pure Chemical Industries, Osaka, Japan),differentially expressed clones were sequenced using prim-anti-IL-12 mAb (50 mg/ml; C15.1 and C15.6, kind gifters to the M13 region of pCRII and BigDyeE Terminatorfrom Dr G. Trinchieri, Wistar Institute of Anatomy andDNA sequencing kit (Perkin-Elmer, Foster City, CA,Biology, Philadelphia, PA, USA), and anti-IFN-g mAb (50USA) by ABI PRISME 310 Genetic Analyzer (Perkin-mg/ml; R4-6A2, Pharmingen). Th1 and Th2 cells culturedElmer). Sequence comparisons were made with sequencesfor 7–10 days were used for the experiments. The skewingin GenBank using NCBI BLAST server system.profile of Th1 and Th2 cells was examined by measuring
their IFN-g and IL-4 producing ability with ELISA kits(Amersham International, Buckingamshire, UK). Further- 2.6. Northern hybridizationmore, it was also confirmed by intracellular cytokinestaining as described previously (Nishimura et al., 1999). Th1 or Th2 mRNA (1 mg per lane) were electrophoresedBriefly, cells were stimulated with PMA (20 ng/ml) and using 1.0% agarose gel containing formaldehyde and thenA23187 (200 ng/ml). After 4 h of activation, brefeldin A transferred to nylon membrane. Probe was amplified by(10 mg/ml; Sigma, St. Louis, MO, USA) was added to the PCR using nested primer pair (Clontech) and labeled with
32cultures and the cells were cultured for an additional 2 h. [a- P]dCTP using Random Primer DNA labeling Kit ver.Cells were harvested, stained with PerCP-conjugated anti- 2 (Takara Shuzo, Shiga, Japan). Hybridization was exe-

T. Yahata et al. / Journal of Neuroimmunology 105 (2000) 103 –108 105
cuted using ExpressHybE Hybridization Solution (Clon-tech) according to manufacturer’s recommendation.
2.7. Detection of preproenkephalin (PPE)
PPE was purified and digested according to a method ofLewis et al. (1980) with some modifications. Th1 and Th2
7cells (5310 cells) were washed twice with PBS andsuspended in 5 ml of 1 M acetic acid, 20 mM HCl, 0.1%2-mercaptoethanol, 1 mM phenylmethylsulfonylfluoride, 1mg/ml pepstatin. Cells were sonicated and the extractcentrifuged at 140 0003g for 60 min. The supernatant waschromatographed on Sephadex G-75 with 1 M acetic acid,20 mM HCl, 0.1% 2-mercaptoethanol. Each fraction waslyophilized, dissolved in 0.5 ml of 50 mM Tris–HCl (pH8.5), and digested at 378C with 2 mg/ml of TPCK-treatedtrypsin for 16 h followed by 0.2 mg/ml of carboxy-peptidase B for 2 h. Proteases were heat-inactivated at908C for 20 min and resulted portions were evaporated todryness. Met-enkephalin in protease-digested fractions was Fig. 1. Northern blot analysis demonstrated Th2 cell-specific expression
of PPE. Th1 and Th2 cells were induced from DO11.10 TCR-Tg mice asseparated on Eicompak MA-5ODS (Eicom, Kyoto, Japan)described in Section 2, and analyzed for their expression of PPE mRNAwith 0.1 M acetate-citrate (pH 3.9), 20.5% methanol andat 7 days. In addition to the cells with no further restimulation (2),detected by electron capture detector (ECD-300; Eicom)mRNA was also isolated from the cells restimulated for 4 h with PMA
(Arita and Kimura, 1988). (20 ng/ml). Northern hybridization analysis was carried out using controlprobe specific for GAPDH (lower) or PPE cDNA probe derived from Th2cells (upper). The data shown are representative of three consistentexperiments.
3. Results and discussion
On the quest for unknown functional molecules ex- point. Cherwinski et al. (1987) suggested expression ofpressed only in Th2 cells, we isolated some cDNA clones PPE mRNA in some Th1 clones; however, we could notby the RDA method from cDNA library prepared from detect PPE mRNA in Th1 cells even after 14 days ofmRNA of Th2 cells. Fig. 1 shows Th2 cell-specific culture (data not shown). Therefore, in these freshlyexpression of one of such clones. DNA sequence analysis induced Th1 or Th2 cells, we concluded that PPE mRNArevealed that this cDNA clone was identical with cDNA was selectively induced in Th2 cells, but not in Th1 cells.encoding PPE (Zurawski et al., 1986) (data not shown). Recently, it has been accepted that IL-12, IFN-g andBecause Th1 cells did not express PPE mRNA even after IL-18 are critical factors for the differentiation of Th1 cellsstimulation with polyclonal T cell activator, PMA (Fig. 1), (Trinchieri, 1994; Takeda et al., 1998), whereas IL-4 is anit was strongly suggested that PPE mRNA is exclusively essential factor for the development of Th2 cells (Hsieh etexpressed in Th2 cells. This result is consistent with a al., 1992; Mosmann and Sad, 1996; Nishimura et al.,previous report by Cherwinski et al. (1987) showing that 1997). Moreover, IL-10 is also involved in the Th2Th2 clones expressed higher levels of PPE mRNA than differentiation through the inhibition of IL-12 productionTh1 clones. To examine the correlation between PPE from APC (Moore et al., 1993). Therefore, our result thatmRNA expression and Th2 differentiation, the kinetics of PPE mRNA was induced in Th2 cells after culture of naivePPE mRNA expression during differentiation into Th1 or Th cells under Th2 conditions (Fig. 2B) strongly suggestedTh2 cells from naive Th cells were examined. Based on that IL-4 and/or IL-10 were key factors for the inductionIFN-g production from Th1 cells and IL-4 production from of PPE mRNA in Th2 cells. To evaluate this, naive ThTh2 cells, the polarization of naive Th cells to Th1 or Th2 cells were cultured under neutral conditions (OVA-pepcells commenced at 2 days and was complete around 7 plus IL-2 without any Th1- or Th2-inducing cytokines) indays of culture (Fig. 2A). The FACS profile of intracellular the presence or absence of either mAb against IL-4, IL-10cytokine staining also demonstrated biased expression of or IFN-g. As shown in Fig. 3, Th cells expanded for 7 daysIFN-g and IL-4 in Th1 and Th2 cells, respectively (Fig. under neutral conditions expressed PPE mRNA. However,2B). In parallel with the differentiation of Th2 cells, addition of anti-IL-4 mAb into the culture completelyupregulation of PPE mRNA was demonstrated in Th2 cells abrogated the upregulation of PPE mRNA. In contrast, noonly after 7 days (Fig. 2C). In contrast, no significant PPE significant inhibition was detected by the addition of anti-mRNA expression was induced in Th1 cells at any time IL-10 mAb or anti-IFN-g mAb. Therefore, endogenously
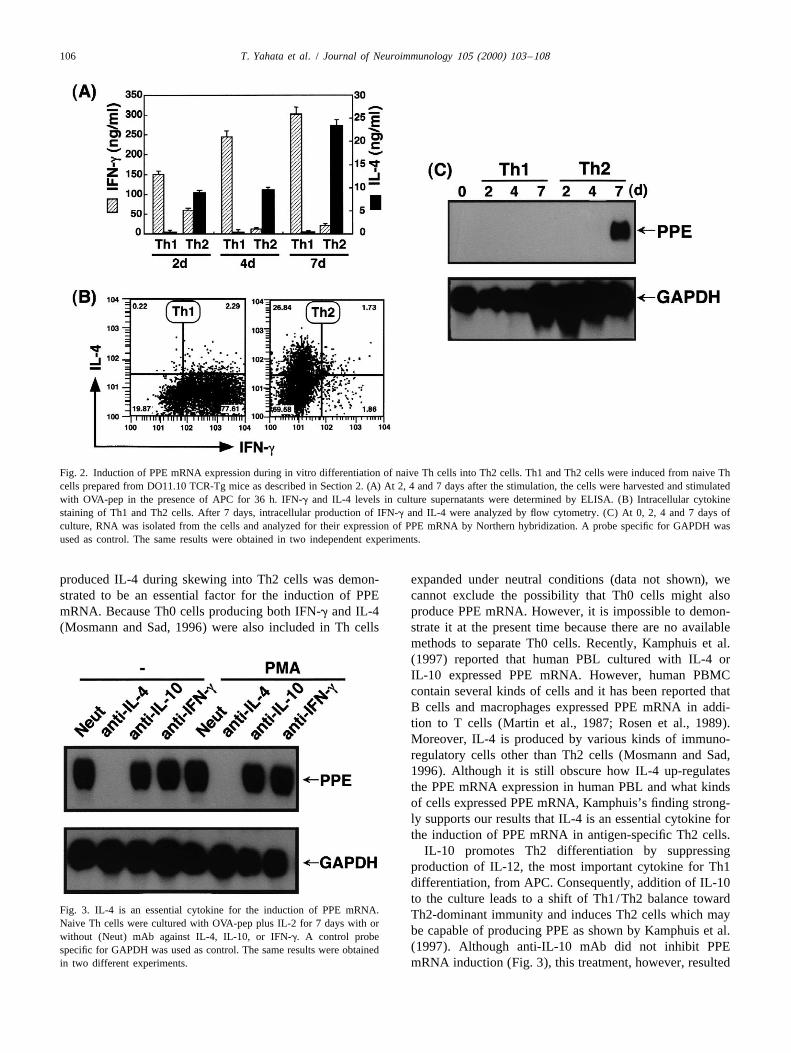
106 T. Yahata et al. / Journal of Neuroimmunology 105 (2000) 103 –108
Fig. 2. Induction of PPE mRNA expression during in vitro differentiation of naive Th cells into Th2 cells. Th1 and Th2 cells were induced from naive Thcells prepared from DO11.10 TCR-Tg mice as described in Section 2. (A) At 2, 4 and 7 days after the stimulation, the cells were harvested and stimulatedwith OVA-pep in the presence of APC for 36 h. IFN-g and IL-4 levels in culture supernatants were determined by ELISA. (B) Intracellular cytokinestaining of Th1 and Th2 cells. After 7 days, intracellular production of IFN-g and IL-4 were analyzed by flow cytometry. (C) At 0, 2, 4 and 7 days ofculture, RNA was isolated from the cells and analyzed for their expression of PPE mRNA by Northern hybridization. A probe specific for GAPDH wasused as control. The same results were obtained in two independent experiments.
produced IL-4 during skewing into Th2 cells was demon- expanded under neutral conditions (data not shown), westrated to be an essential factor for the induction of PPE cannot exclude the possibility that Th0 cells might alsomRNA. Because Th0 cells producing both IFN-g and IL-4 produce PPE mRNA. However, it is impossible to demon-(Mosmann and Sad, 1996) were also included in Th cells strate it at the present time because there are no available
methods to separate Th0 cells. Recently, Kamphuis et al.(1997) reported that human PBL cultured with IL-4 orIL-10 expressed PPE mRNA. However, human PBMCcontain several kinds of cells and it has been reported thatB cells and macrophages expressed PPE mRNA in addi-tion to T cells (Martin et al., 1987; Rosen et al., 1989).Moreover, IL-4 is produced by various kinds of immuno-regulatory cells other than Th2 cells (Mosmann and Sad,1996). Although it is still obscure how IL-4 up-regulatesthe PPE mRNA expression in human PBL and what kindsof cells expressed PPE mRNA, Kamphuis’s finding strong-ly supports our results that IL-4 is an essential cytokine forthe induction of PPE mRNA in antigen-specific Th2 cells.
IL-10 promotes Th2 differentiation by suppressingproduction of IL-12, the most important cytokine for Th1differentiation, from APC. Consequently, addition of IL-10to the culture leads to a shift of Th1/Th2 balance toward
Fig. 3. IL-4 is an essential cytokine for the induction of PPE mRNA. Th2-dominant immunity and induces Th2 cells which mayNaive Th cells were cultured with OVA-pep plus IL-2 for 7 days with or
be capable of producing PPE as shown by Kamphuis et al.without (Neut) mAb against IL-4, IL-10, or IFN-g. A control probe(1997). Although anti-IL-10 mAb did not inhibit PPEspecific for GAPDH was used as control. The same results were obtained
in two different experiments. mRNA induction (Fig. 3), this treatment, however, resulted
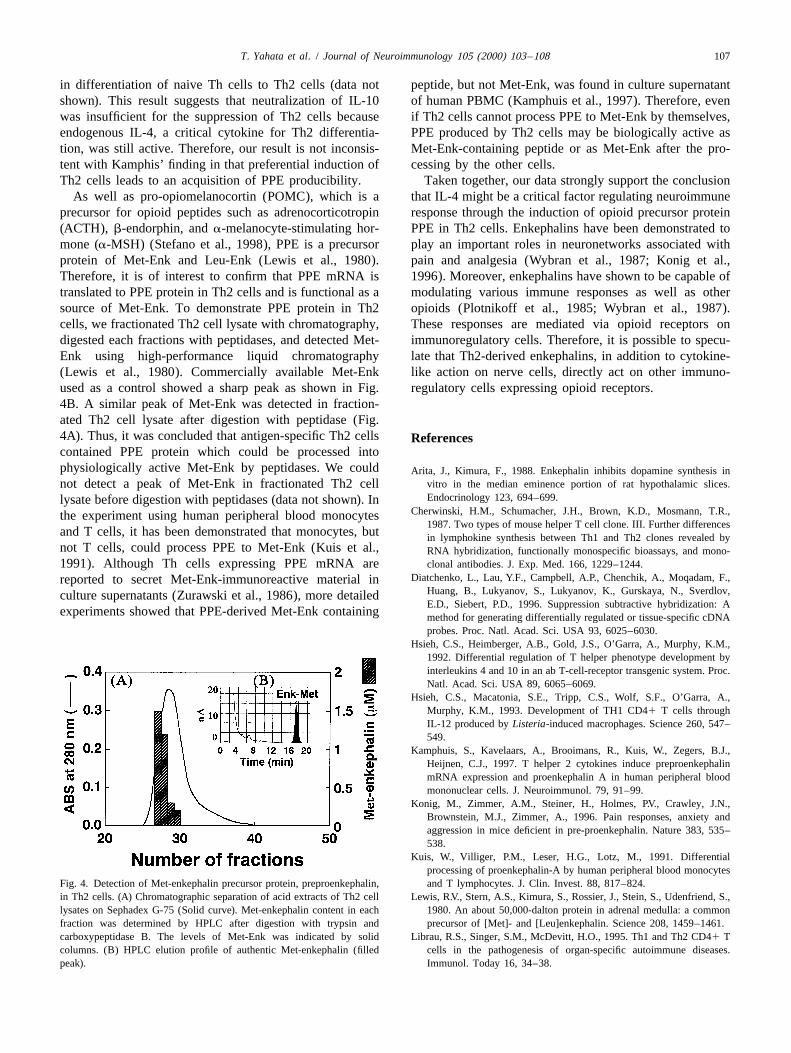
T. Yahata et al. / Journal of Neuroimmunology 105 (2000) 103 –108 107
in differentiation of naive Th cells to Th2 cells (data not peptide, but not Met-Enk, was found in culture supernatantshown). This result suggests that neutralization of IL-10 of human PBMC (Kamphuis et al., 1997). Therefore, evenwas insufficient for the suppression of Th2 cells because if Th2 cells cannot process PPE to Met-Enk by themselves,endogenous IL-4, a critical cytokine for Th2 differentia- PPE produced by Th2 cells may be biologically active astion, was still active. Therefore, our result is not inconsis- Met-Enk-containing peptide or as Met-Enk after the pro-tent with Kamphis’ finding in that preferential induction of cessing by the other cells.Th2 cells leads to an acquisition of PPE producibility. Taken together, our data strongly support the conclusion
As well as pro-opiomelanocortin (POMC), which is a that IL-4 might be a critical factor regulating neuroimmuneprecursor for opioid peptides such as adrenocorticotropin response through the induction of opioid precursor protein(ACTH), b-endorphin, and a-melanocyte-stimulating hor- PPE in Th2 cells. Enkephalins have been demonstrated tomone (a-MSH) (Stefano et al., 1998), PPE is a precursor play an important roles in neuronetworks associated withprotein of Met-Enk and Leu-Enk (Lewis et al., 1980). pain and analgesia (Wybran et al., 1987; Konig et al.,Therefore, it is of interest to confirm that PPE mRNA is 1996). Moreover, enkephalins have shown to be capable oftranslated to PPE protein in Th2 cells and is functional as a modulating various immune responses as well as othersource of Met-Enk. To demonstrate PPE protein in Th2 opioids (Plotnikoff et al., 1985; Wybran et al., 1987).cells, we fractionated Th2 cell lysate with chromatography, These responses are mediated via opioid receptors ondigested each fractions with peptidases, and detected Met- immunoregulatory cells. Therefore, it is possible to specu-Enk using high-performance liquid chromatography late that Th2-derived enkephalins, in addition to cytokine-(Lewis et al., 1980). Commercially available Met-Enk like action on nerve cells, directly act on other immuno-used as a control showed a sharp peak as shown in Fig. regulatory cells expressing opioid receptors.4B. A similar peak of Met-Enk was detected in fraction-ated Th2 cell lysate after digestion with peptidase (Fig.4A). Thus, it was concluded that antigen-specific Th2 cells Referencescontained PPE protein which could be processed intophysiologically active Met-Enk by peptidases. We could Arita, J., Kimura, F., 1988. Enkephalin inhibits dopamine synthesis in
vitro in the median eminence portion of rat hypothalamic slices.not detect a peak of Met-Enk in fractionated Th2 cellEndocrinology 123, 694–699.lysate before digestion with peptidases (data not shown). In
Cherwinski, H.M., Schumacher, J.H., Brown, K.D., Mosmann, T.R.,the experiment using human peripheral blood monocytes1987. Two types of mouse helper T cell clone. III. Further differences
and T cells, it has been demonstrated that monocytes, but in lymphokine synthesis between Th1 and Th2 clones revealed bynot T cells, could process PPE to Met-Enk (Kuis et al., RNA hybridization, functionally monospecific bioassays, and mono-1991). Although Th cells expressing PPE mRNA are clonal antibodies. J. Exp. Med. 166, 1229–1244.
Diatchenko, L., Lau, Y.F., Campbell, A.P., Chenchik, A., Moqadam, F.,reported to secret Met-Enk-immunoreactive material inHuang, B., Lukyanov, S., Lukyanov, K., Gurskaya, N., Sverdlov,culture supernatants (Zurawski et al., 1986), more detailedE.D., Siebert, P.D., 1996. Suppression subtractive hybridization: A
experiments showed that PPE-derived Met-Enk containing method for generating differentially regulated or tissue-specific cDNAprobes. Proc. Natl. Acad. Sci. USA 93, 6025–6030.
Hsieh, C.S., Heimberger, A.B., Gold, J.S., O’Garra, A., Murphy, K.M.,1992. Differential regulation of T helper phenotype development byinterleukins 4 and 10 in an ab T-cell-receptor transgenic system. Proc.Natl. Acad. Sci. USA 89, 6065–6069.
Hsieh, C.S., Macatonia, S.E., Tripp, C.S., Wolf, S.F., O’Garra, A.,Murphy, K.M., 1993. Development of TH1 CD41 T cells throughIL-12 produced by Listeria-induced macrophages. Science 260, 547–549.
Kamphuis, S., Kavelaars, A., Brooimans, R., Kuis, W., Zegers, B.J.,Heijnen, C.J., 1997. T helper 2 cytokines induce preproenkephalinmRNA expression and proenkephalin A in human peripheral bloodmononuclear cells. J. Neuroimmunol. 79, 91–99.
Konig, M., Zimmer, A.M., Steiner, H., Holmes, P.V., Crawley, J.N.,Brownstein, M.J., Zimmer, A., 1996. Pain responses, anxiety andaggression in mice deficient in pre-proenkephalin. Nature 383, 535–538.
Kuis, W., Villiger, P.M., Leser, H.G., Lotz, M., 1991. Differentialprocessing of proenkephalin-A by human peripheral blood monocytes
Fig. 4. Detection of Met-enkephalin precursor protein, preproenkephalin, and T lymphocytes. J. Clin. Invest. 88, 817–824.in Th2 cells. (A) Chromatographic separation of acid extracts of Th2 cell Lewis, R.V., Stern, A.S., Kimura, S., Rossier, J., Stein, S., Udenfriend, S.,lysates on Sephadex G-75 (Solid curve). Met-enkephalin content in each 1980. An about 50,000-dalton protein in adrenal medulla: a commonfraction was determined by HPLC after digestion with trypsin and precursor of [Met]- and [Leu]enkephalin. Science 208, 1459–1461.carboxypeptidase B. The levels of Met-Enk was indicated by solid Librau, R.S., Singer, S.M., McDevitt, H.O., 1995. Th1 and Th2 CD41 Tcolumns. (B) HPLC elution profile of authentic Met-enkephalin (filled cells in the pathogenesis of organ-specific autoimmune diseases.peak). Immunol. Today 16, 34–38.

108 T. Yahata et al. / Journal of Neuroimmunology 105 (2000) 103 –108
Martin, J., Prystowsky, M.B., Angeletti, R.H., 1987. Preproenkephalin Plotnikoff, N.P., Murgo, A.J., Miller, G.C., Corder, C.N., Faith, R.E.,mRNA in T-cells, macrophages, and mast cells. J. Neurosci. Res. 18, 1985. Enkephalins: immunomodulators. Fed. Proc. 44, 118–122.82–87. Rosen, H., Behar, O., Abramsky, O., Ovadia, H., 1989. Regulated
Moore, K.W., O’Garra, A., de Waal Malefyt, R., Vieira, P., Mosmann, expression of proenkephalin A in normal lymphocytes. J. Immunol.T.R., 1993. Interleukin-10. Annu. Rev. Immunol. 11, 165–190. 143, 3703–3707.
Mosmann, T.R., Sad, S., 1996. The expanding universe of T-cell subsets: Stefano, G.B., Salzet, B., Fricchione, G.L., 1998. Enkelytin and opioidTh1, Th2 and more. Immunol. Today 17, 138–146. peptide association in invertebrates and vertebrates: immune activa-
Murphy, K.M., Heimberger, A.B., Loh, D.Y., 1990. Induction by antigen tion and pain. Immunol. Today 19, 265–268.of intrathymic apoptosis of CD41CD81TCRlo thymocytes in vivo. Takeda, K., Tsutsui, H., Yoshimoto, T., Adachi, O., Yoshida, N.,Science 250, 1720–1723. Kishimoto, T., Okamura, H., Nakanishi, K., Akira, S., 1998. Defective
Nishimura, T., Santa, K., Yahata, T., Sato, N., Ohta, A., Ohmi, Y., Sato, NK cell activity and Th1 response in IL-18-deficient mice. ImmunityT., Hozumi, K., Habu, S., 1997. Involvement of IL-4-producing 8, 383–390.Vb8.21 CD41 CD62L- CD45RB- T cells in non-MHC gene-con- Trinchieri, G., 1994. Interleukin-12: a cytokine produced by antigen-trolled predisposition toward skewing into T helper type-2 immunity presenting cells with immunoregulatory functions in the generation ofin BALB/c mice. J. Immunol. 158, 5698–5706. T-helper cell type 1 and cytotoxic lymphocytes. Blood 84, 4008–
Nishimura, T., Iwakabe, K., Sekimoto, M., Ohmi, Y., Yahata, T., Nakui, 4027.´M., Sato, T., Habu, S., Tashiro, H., Sato, M., Ohta, A., 1999. Distinct Wybran, J., Schandene, L., Vooren, J.-P.V., Vandermoten, G., Latinne, D.,
`role of antigen-specific T helper type 1 (Th1) and Th2 cells in tumor Sonnet, J., De Bruyere, M., Taelman, H., Plotnikoff, N.P., 1987.eradication in vivo. J. Exp. Med. 190, 617–627. Immunologic properties of methionine-enkephalin, and therapeutic
O’Garra, A., Murphy, K.M., 1995. T-cell subsets in autoimmunity. Curr. implications in AIDS, ARC, and Cancer. Ann. NY Acad. Sci. 496,Opin. Immunol. 5, 880–886. 108–114.
Ohta, A., Sato, N., Yahata, T., Ohmi, Y., Santa, K., Sato, T., Tashiro, H., Zurawski, G., Benedik, M., Kamb, B.J., Abrams, J.S., Zurawski, S.M.,Habu, S., Nishimura, T., 1997. Manipulation of Th1/Th2 balance in Lee, F.D., 1986. Activation of mouse T-helper cells induces abundantvivo by adoptive transfer of antigen-specific Th1 or Th2 cells. J. preproenkephalin mRNA synthesis. Science 232, 772–775.Immunol. Methods 209, 85–92.
![The prevention of 2,4-dinitrochlorobenzene-induced ...€¦ · [5, 6] and T-helper 2 (Th2)-dependent cells [7, 8]. Mast * Correspondence: pm.thehoo@gmail.com; epiko@khu.ac.kr 2Department](https://img.pdfslide.net/doc/110x75/60a198517546aa433c0bf138/the-prevention-of-24-dinitrochlorobenzene-induced-5-6-and-t-helper-2-th2-dependent.jpg)








![Circulating and Tumor-Infiltrating Foxp3 Regulatory T Cell ... · traditional Th1, Th2 helper T cell subsets, Foxp3+ reg-ulatory T cell (Tregs) and IL-17-producing Th17 cells[9]](https://img.pdfslide.net/doc/110x75/5e4b79c0f61ac961cb5bf5de/circulating-and-tumor-infiltrating-foxp3-regulatory-t-cell-traditional-th1.jpg)








![A peer-reviewed version of this preprint was published in ...27 polarizing cytokines (i.e., interferon [IFN]-γ/ Interleukin [IL]-4), and animals with imbalanced 28 Th1/Th2 response](https://img.pdfslide.net/doc/110x75/5ff1ba5a56a8075905798c55/a-peer-reviewed-version-of-this-preprint-was-published-in-27-polarizing-cytokines.jpg)










