Embed Size (px)
Citation preview

doi:10.1182/blood-2012-01-402370Prepublished online July 12, 2012;
Lee, Jenifer Widger, Tibor Keler, Lélia Delamarre and Ira MellmanBithi Chatterjee, Anna Smed-Sörensen, Lillian Cohn, Cécile Chalouni, Richard Vandlen, Byoung-Chul regulate the efficiency of cross presentation by human dendritic cellsInternalization and endosomal degradation of receptor-bound antigens
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

1
Internalization and endosomal degradation of receptor-bound antigens
regulate the efficiency of cross presentation by human dendritic cells
Bithi Chatterjee1,2, Anna Smed-Sörensen2,3, Lillian Cohn2, Cécile Chalouni2, Richard
Vandlen2, Byoung-Chul Lee2, Jenifer Widger4, Tibor Keler4, Lélia Delamarre2, and Ira
Mellman2*
1Department of Cell Biology, Yale University School of Medicine, New Haven, CT 06510;
2Genentech, South San Francisco, CA 94080; 3Department of Microbiology, Tumor and
Cell Biology, Karolinska Institutet, Stockholm, Sweden; 4Celldex Therapeutics,
Phillipsburg, NJ 08865
*Correspondence should be addressed to:
Ira Mellman, Genentech, 1 DNA Way, Mail Stop 212, South San Francisco, CA 94080
email: [email protected]
Short title: Degradation controls cross presentation efficiency
Keywords: cross presentation, endocytosis, degradation, MHC class I, dendritic cells
Blood First Edition Paper, prepublished online July 12, 2012; DOI 10.1182/blood-2012-01-402370
Copyright © 2012 American Society of Hematology
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

2
Abstract
Dendritic cells (DCs) can capture extracellular antigens and load resultant peptides
on to MHC class I molecules, a process termed cross presentation. The mechanisms of
cross presentation remain incompletely understood, particularly in primary human DCs.
One unknown is the extent to which antigen delivery to distinct endocytic compartments
determines cross presentation efficiency, possibly by influencing antigen egress to the
cytosol. We addressed the problem directly and quantitatively by comparing the cross
presentation of identical antigens conjugated to antibodies against different DC receptors
that are targeted to early or late endosomes at distinct efficiencies. In human BDCA1+
and monocyte-derived DCs, CD40 and mannose receptor (MR) targeted antibody
conjugates to early endosomes while DEC205 targeted antigen primarily to late
compartments. Surprisingly, the receptor least efficient at internalization, CD40, was the
most efficient at cross presentation. This did not reflect DC activation by CD40, but rather
its relatively poor uptake or intra-endosomal degradation as compared to MR or DEC205.
Thus, while both early and late endosomes appear to support cross presentation in
human DCs, internalization efficiency, especially to late compartments, may be a negative
predictor of activity when selecting receptors for vaccine development.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

3
Introduction
Dendritic cells (DCs) are professional antigen presenting cells that process proteins
and load resultant peptides on to major histocompatability complex (MHC) molecules.
Upon activation, DCs migrate to secondary lymphoid organs and present antigen to their
cognate T cells, for the induction of adaptive immune responses 1. Since DCs are
essential for the initiation of adaptive immune responses, they are attractive targets for
enhancing prophylactic and therapeutic vaccination strategies 2-4 and have been shown to
activate anti-tumor immune responses in murine cancer models 5,6.
In human cancer, clinical evidence is now accumulating to suggest that the
induction or activation of CD8+ T cell immunity can contribute to the arrest of tumor
growth and patient survival 7. In principle, targeting tumor antigens to DCs may enhance
protective CD8+ T cell responses due to the ability of DCs to cross present exogenous
antigens 8. In cross presentation, exogenous proteins are endocytosed, processed, and
loaded on to MHC class I molecules for presentation to CD8+ T cells. The efficiency of
cross presentation can be improved >100-fold when receptors found on the surface of
DCs are targeted specifically 6.
Multiple DC populations exhibit some capacity for cross presentation in vitro, but
certain subpopulations (CD8+ DCs in the mouse, BDCA3+ DCs in human) are thought to
be particularly adept in vivo 8. DCs exhibit a variety of surface receptors that can
internalize antigen for cross presentation, with some being subpopulation-specific.
Pioneering studies from Steinman and co-workers have shown cross presentation using
the C-type lectin receptor DEC205, which is expressed by CD8+ mouse DCs as well as
by multiple human DC subsets 9,10. Another lectin receptor, CLEC9A/DNGR-1, is
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

4
expressed by mouse CD8+ DCs and human BDCA3+ DCs, and also mediates cross
presentation 5. Indeed, a variety of receptors expressed by human or mouse DCs (DCIR,
Langerin, mannose receptor (MR), DC-SIGN, CLEC12A) enable cross presentation 10-15.
Which of these receptors is most efficient and why remains poorly understood, particularly
in human DCs.
A few recent studies have begun to address the relative roles of early endosomes
and late endosomes/lysosomes in cross presentation. Burgdorf et al. suggested for
mouse DCs that cross presentation of soluble ovalbumin internalized by MR, presumably
targeted to early endosomes, is more efficient than cross presentation following fluid
phase uptake, which also delivers an unspecified portion of soluble ovalbumin to late
endosomes 16-18. Early endosomes also may host the cross presentation of liposome-
encapsulated hen egg lysozyme, but in a fashion independent of the proteasomal
pathway 19,20. Antigen coupled to DC-SIGN antibodies also suggested that early
endosomal targeting may facilitate cross presentation by human DC-SIGN transgenic
mouse DCs 13. In all of these studies, however, there was limited quantitation or
characterization of antigen accumulation and fate, and direct and effective comparisons
were not made between the various uptake routes. Where the effects of targeting different
receptors was compared, endosomal targeting was not well studied and the conjugates
were poorly characterized for the possibility of aggregates 21,22, an important consideration
if any of these platforms are to be transferred to the clinic for use in humans. Aggregates
also tend to be transferred to lysosomes and may have altered degradation properties
23,24. Finally, suggestions that early endosomal targeting is optimal appear inconsistent
with more extensive investigations of DEC205, which is targeted primarily to late
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

5
endocytic compartments 25,26, yet appears to mediate the efficient cross presentation of
proteins coupled to or fused to anti-DEC205 antibodies 6,9,12.
To establish directly and quantitatively the endosomal requirements for efficient
cross presentation using a clinically relevant and scalable platform, we used elongated
peptide antigens conjugated to monoclonal antibodies against three receptors that traffic
to distinct cellular compartments: CD40, MR, and DEC205. Interestingly, we found an
inverse relationship between internalization or antigen degradation and cross
presentation: antigens destined for more degradative late endosomes were poorly cross
presented relative to the same antigens targeted to early endosomes, an effect that was
independent of the amount of antigen internalized.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

6
Materials and methods
Cell isolation and culture
This study was approved by the Genentech Institutional Review Board. Our procedures
for the isolation of DCs and T cells from blood have been described previously 27. Healthy
donors were leukapheresed and enriched populations of lymphocytes and monocytes
were obtained by counterflow elutriation. BDCA1+ DCs were isolated from elutriated
monocytes using the CD1c DC isolation kit and AutoMACS technology (Miltenyi). Cells
were cultured overnight in R10 (RPMI 1640 + Glutamax with 10% FCS, 100 units/mL
penicillin + 100 μg/mL streptomycin, 10 mM Hepes) (Gibco/Invitrogen), and 2 ng/mL GM-
CSF (Peprotech). Monocyte-derived DCs (MoDCs) were derived in 5-7 d from CD14+
monocytes cultured in R10 with 100 ng/mL GM-CSF (Peprotech) and 6.5 ng/mL IL-4
(R&D Systems). To mature DCs, DCs were exposed to 0.2 μg/mL lipopolysaccharide
(LPS, Sigma) overnight. For autologous CD8+ T cell assays with MoDCs, elutriated HLA-
A*0201+ monocytes were frozen in anticipation of donor recall. CD8+ T cells were
isolated from HLA-A*0201+ elutriated lymphocytes using the CD8+ T cell isolation kit
(Miltenyi). For CD4 T cell assays using an NY-ESO peptide specific human CD4 T cell
clone (CCNE-1 415; obtained from Cassian Yee), HLA-DPB1*0401 DCs were obtained as
above.
Antibodies
Anti-DEC205 (3G9; Kd, 0.3 nM) and anti-MR (B11; Kd, 0.7 nM) used in our studies were
obtained in collaboration with Celldex Therapeutics, and the anti-CD40 (S2C6; Kd, 0.12
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

7
nM) was obtained in collaboration with Seattle Genetics. Binding affinities were
determined by flow cytometry.
Antibodies used for flow cytometry/immunofluorescence were as follows: anti-CD3 (SK7),
CD4 (Leu-3a/SK3), CD8 (SK1), CD8 (RPA-T8), CD11c (B-ly6), CD14 (MΦP9), CD19
(Leu-12), CD40 (5C3), CD71 (M-A712), CD83 (HB15e), CD86 (FUN-1), CD206 (19.2),
DEC205 (MG38), HLA-DR (TU36), IFNγ (B27), IL-2 (MQ1-17H12), Lamp1 (H4A3) and
TNFα (6401.1111) (BD Biosciences); anti-HLA-DR (L243) (Biolegend); anti-HLA-ABC
(W6/32) (eBioscience); anti-CD1a (NA1/34) (DAKO); anti-EEA1 (Cell Signaling);
secondary reagents (steptavidin Alexa 555/647, anti-rabbit Alexa 546, anti-FITC Alexa
488) (Invitrogen/Molecular Probes).
Synthesis of antibody-peptide conjugates
Antibodies in PBS buffer with 50mM potassium phosphate (pH 7.5) were reacted with a
10-15 fold molar excess of N-succinimidyl S-acetylthioacetate (Pierce) for 4-6 h at 20°C.
Excess reagents were removed by dialysis in PBS resulting in antibodies with 3-6 blocked
thiol groups. Blocking groups were then removed with 20 mM hydroxylamine (Sigma) and
peptides containing an N-terminal maleimide (Elim Biopharmaceuticals) were added to
approximately 5-6 fold molar excess over antibody and allowed to react with the new thiol
groups for 2-4 h at 20°C. Excess peptide and aggregated antibodies were removed by gel
filtration in PBS. Electrospray mass spectrometry was used to estimate the average
number of peptides per antibody. The peptides conjugated to antibodies were
(immunodominant epitopes underlined and sequence position indicated):
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

8
LTKGILGFVFTLTVPSER (influenza M1 58-66), QAGILARNLVPMVATVQGQNL
(cytomegalovirus (CMV) pp65 495-503), or QQLSLLMWITQAFLPVFLAQPPSGQRR (NY-
ESO 157-170).
Immunofluorescence
Following culture or accumulation with 1 μg/mL anti-CD40 (S2C6), anti-MR (B11), or anti-
DEC205 (3G9) antibodies covalently conjugated to Alexa 488 (Alexa 488 monoclonal
antibody kit, Invitrogen), BDCA1+ DCs or MoDCs were washed and spotted on Alcian
blue coated coverslips for 10-15 min at room temperature. Cells were fixed in 4%
paraformaldehyde (PFA, Electron Microscopy Source) for 15 min, permeabilized in 0.05%
saponin (Sigma), and counterstained. When biotinylated antibodies were used,
endogenous biotin was blocked first by using an excess of unlabeled streptavidin and
biotin (Endogenous biotin blocking kit, Invitrogen). Following labeling, coverslips were
mounted onto glass slides using Prolong Gold with DAPI (Invitrogen/Molecular Probes).
Images were acquired on a Leica SP5 confocal microscope, using a 100x oil objective
(NA: 1.47) with zoom 7, and Leica’s LAS imaging software (Leica Microsystems).
Antibody accumulation experiments
Cells were continuously incubated with 1 μg/mL anti-CD40 (S2C6), anti-MR (B11), or anti-
DEC205 (3G9) Alexa 488 antibodies for indicated times, in the presence or absence of
the serine/cysteine protease inhibitor leupeptin (5 mM) (Roche) and the acidophilic weak
base ammonium chloride (NH4Cl) (10 mM). Following incubations, cells were stained for
CD14 and maturation markers, fixed, acquired using a BD FACSCanto II, and analyzed
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

9
for Alexa 488 signal using FlowJo. For direct receptor comparisons, MFI was normalized
for surface antibody signal (4°C) and the number of fluors per antibody.
Antigen presentation assays
MHC class I:
HLA-A*0201 DCs were incubated for 4-6 h in the presence of antibody-peptide conjugates
(0.1-10 μg/mL) or other reagents of interest (peptide control 2.5-250 ng/mL, Proimmune;
epoxomicin, 0.1 μM, Calbiochem; DMSO, 0.1 μM, Sigma). Following antigen uptake, DCs
were washed extensively to remove free antibody, peptide, or inhibitor, and cocultured
with carboxyfluorescein succinmidyl ester (CFSE) (Invitrogen) labeled CD8+ T cells for 8-
10 d at a DC:T cell ratio of 1:30, in the presence of 20 units/mL IL-2 (Roche) and 200
ng/mL LPS (Sigma). After coculture, influenza M1 or CMV pp65 peptide specific CD8+ T
cells were labeled with a corresponding PE-pentamer (ProImmune), acquired using a BD
FACSCanto II, and gated on CD3+CD8+ (CD14/CD4/CD19-) cells. CFSElo/pentamerhi
cells were analyzed using FlowJo.
MHC class II:
HLA-DPB1*0401 DCs were incubated for 1.5 h in the presence of antibody conjugates
(0.1-10 μg/mL) or other reagents of interest (peptide control 2.5-25 μg/mL, Elim
Biopharmaceuticals; SEB 1μg/mL, Sigma). Following antigen uptake, DCs were washed
and cocultured with an NY-ESO peptide specific human CD4 T cell clone for 7-9 h at
DC:T ratios of 1:1, 1:3, and 1:9. Two hours into the coculture, Brefeldin A (BFA;
eBioscience) was added to prevent cytokine secretion. Following coculture, cells were
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

10
stained for surface markers, fixed, permeabilized, and labeled for intracellular IFNγ, IL-2,
and TNFα. Cells were acquired with a BD FACSCanto II, and gated on CD3+CD4+CD1c-
cells, followed by Flowjo analysis.
Pulse-chase experiments
A488 conjugated antibodies of interest were pulsed with cells at 37°C or 4°C for 1 h at 4
μg/mL, in the presence or absence of 5 mM leupeptin and 10 mM NH4Cl. Cells were
washed and chased for the indicated times, in the presence or absence of leupeptin and
NH4Cl. Cells were counterstained with CD14 and maturation markers, washed, and fixed.
Cells were acquired using a BD FACSCanto II, and analyzed for Alexa 488 signal using
FlowJo.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

11
Results Antibodies are targeted to distinct cellular compartments
To compare quantitatively the ability of different DC endosomal compartments to
facilitate cross presentation, we selected three receptors suspected to have distinct
intracellular destinations or rates of internalization: mannose receptor (MR), CD40, and
DEC205. We first confirmed their surface expression on primary human BDCA1+ DCs
and monocyte-derived DCs (MoDCs) (Fig. 1A-C). With the exception of MR on BDCA1+
DCs, both CD40 and DEC205 were expressed at variably high levels by both immature
and mature BDCA1+ DCs and MoDCs. MR expression was low but detectable on
immature BDCA1+ DCs, but this amount further decreased upon maturation. As expected,
maturation enhanced CD40 and DEC205 expression in both DC types. When comparing
relative receptor expression levels, both DC types expressed significant amounts of each
receptor before and after maturation by LPS, except for MR whose expression was low in
immature BDCA1+ DCs (Fig. 1D). All antibodies exhibited sub-nanomolar affinity (anti-
CD40: 0.12 nM, anti-DEC205: 0.3 nM, anti-MR: 0.7 nM), and were used at saturation.
We next characterized the ability of each receptor to be internalized and its
subsequent subcellular localization in both BDCA1+ DCs and MoDCs. As expected from
previous studies, anti-DEC205 antibodies accumulated largely in late endosomes. Very
little was found on the surface of BDCA1+ DCs and MoDCs (Fig. 2A and 2D, first panel),
and a small amount appeared to be associated with early endosomes as marked by EEA1
(Fig. 2A and 2D, second panel). The bulk of anti-DEC205 antibody overlapped with the
late endosomal marker Lamp1 (Fig. 2A and 2D, third panel), and the signal was enhanced
in cells treated with the protease inhibitor leupeptin together with the acidophilic weak
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

12
base ammonium chloride (inh), indicating that antibody degradation was occurring in this
compartment (Lamp1; Fig. 2A and 2D, fourth panel). Therefore, in untreated cells the
amount of anti-DEC205 delivery to late compartments was underestimated.
When MR antibody was incubated continuously with BDCA1+ DCs and MoDCs, it
was mainly found intracellularly with relatively little on the cell surface (Fig. 2B and 2E,
first panel). Anti-MR was found in early endosomes as indicated by the partial overlap with
EEA1 (Fig. 2B and 2E, second panel) and minimal overlap with Lamp1 (Fig. 2B and 2E
third panel). This pattern was maintained even in the presence of protease and
acidification inhibitors (Fig. 2B and 2E fourth panel), suggesting that anti-MR antibodies
did not reach late compartments as significantly as anti-DEC205 and did not escape
detection due to their degradation.
Following continuous incubation in BDCA1+ DCs and MoDCs, anti-CD40 antibody
was largely found on the surface (Fig. 2C and 2F, first panel), in early endosomes (EEA1;
Fig. 2C and 2F, second panel), and minimally in late endosomes (Lamp1; Fig. 2C and 2F,
third panel). This pattern was similar in the presence of protease and acidification
inhibitors (Fig. 2C and 2F, fourth panel). Thus, like MR, anti-CD40 internalized to early
endosomes, but appeared to do so much less efficiently. Similar results were obtained for
all antibodies at earlier and later time points of internalization (data not shown), and the
extent of overlap was quantified (Fig. S1).
The patterns observed by antibody internalization were confirmed by indirect
immunofluorescence detection of the receptors themselves: CD40 was mainly found on
the surface, while MR and DEC205 were mainly found intracellularly, in early and in late
endosomes, respectively (data not shown). In sum, we have characterized the targeting
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

13
and localization of antibodies against three receptors in two different human DC types:
DEC205 in late endosomes, MR in early endosomes, and CD40 on the surface and in
early endosomes.
CD40 accumulates the least efficiently of the three receptors
To quantify the ability of DEC205, MR, and CD40 to accumulate antigen, we
determined the time course of continuous anti-receptor antibody uptake by flow cytometry.
As shown in Figure 3 for BDCA1+ DCs (A) and MoDCs (B), MR antibody was more
effectively accumulated than either CD40 or DEC205 antibodies (top panels). Inhibiting
proteolysis by the addition of the protease and acidification inhibitors (inh) did not
markedly affect the accumulation of either MR or CD40 antibodies, but did significantly
enhance the amount of cell-associated DEC205 antibody, particularly in MoDCs (Fig. 3B,
bottom panel; p <0.05). This finding is consistent with the observations that MoDCs exhibit
higher levels of lysosomal enzymes than BDCA1+ DCs 28,29 and that DEC205 antibodies
are delivered predominantly to protease-containing late endosomes. In all cases, isotype
controls were minimally internalized (muIso corresponds to anti-CD40; huIso corresponds
to anti-MR and anti-DEC205). Taken together, these data indicate that MR and DEC205
antibodies are internalized by both DC types to a greater extent than CD40 antibodies.
Targeting CD40 and MR is more efficient for cross presentation than targeting
DEC205
Having characterized the internalization and intracellular destinations of antibodies
to CD40, DEC205, and MR, we next compared their relative abilities to mediate cross
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

14
presentation. For this purpose, each high affinity antibody was chemically coupled to
either of two immunodominant HLA-A*0201 epitopes, influenza M1 (58-66) and
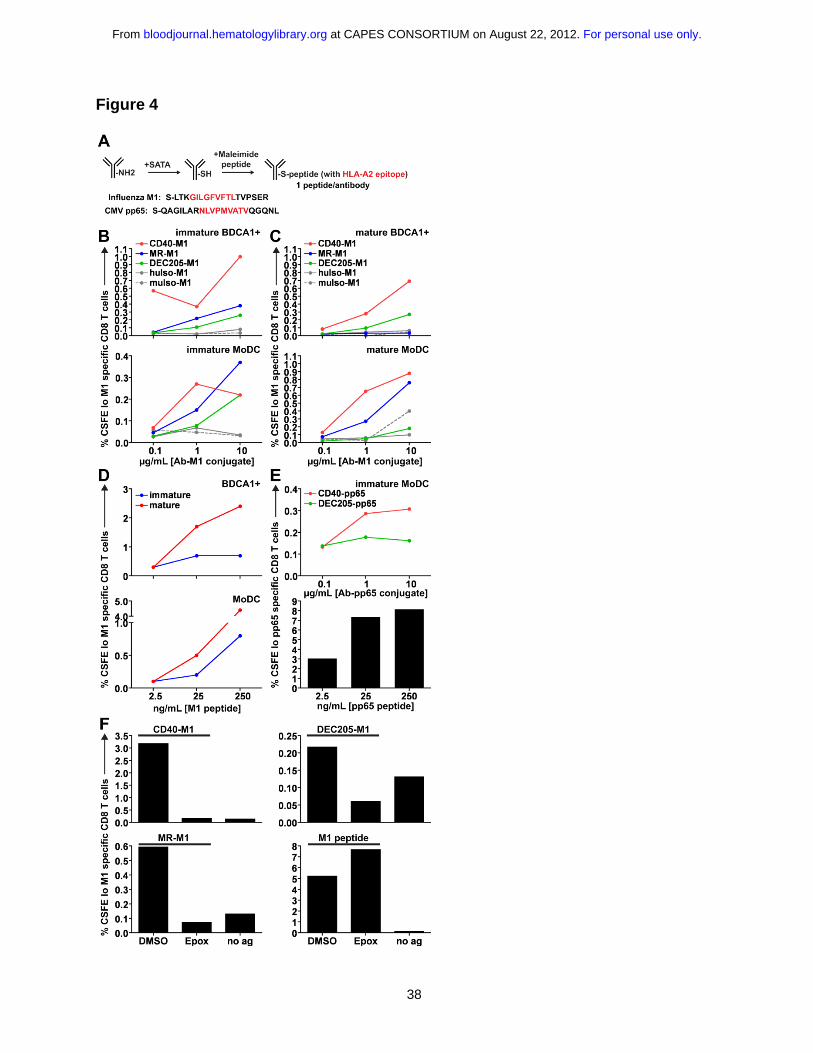
cytomegalovirus (CMV) pp65 (495-503), using the strategy outlined in Figure 4A. Both
epitopes (red residues, Fig. 4A) were extended by several residues of naturally occurring
peptide sequence on the amino- and carboxy-terminal ends (black residues, Fig. 4A). We
determined that the antibodies were conjugated to similar numbers of peptides per
molecule by mass spectrometry and remained >95% monomeric by gel filtration
chromatography (data not shown).
Using these antibody conjugates, we performed recall cross presentation assays
using human HLA-A*0201+ DCs and autologous CD8+ T cells from donors with
detectable memory responses against influenza or CMV (Fig. S2A). The percent of total
CD8+ T cells that were dividing and M1 or pp65 specific was assessed by CFSE dilution
and pentamer staining. In immature BDCA1+ DCs and MoDCs fed with M1 peptide
coupled antibodies, targeting antibodies to the early endosomal receptors CD40 and MR
was generally superior for cross presentation as compared to DEC205 (Fig. 4B and Fig.
S2B), which surprisingly, was the most inefficient for cross presentation. Interestingly, the
receptor that was least efficient in accumulating antigen, CD40, was the most efficient for
cross presentation, indicating that intracellular antigen accumulation was not the rate-
limiting step in determining cross presentation efficiency.
We also assessed the ability of mature DCs to cross present receptor targeted
antigen, as matured mouse DCs are able to present antigens captured via receptor-
mediated endocytosis 26. We found that mature BDCA1+ DCs and MoDCs were able to
induce efficient cross presentation via CD40, and in mature MoDCs, targeting MR also led
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

15
to efficient cross presentation (Figure 4C). Targeting MR in mature BDCA1+ DCs resulted
in little to no cross presentation, likely due to the absence of MR on the surface upon
maturation (Fig. 1A). In both immature and mature DCs, targeting DEC205 resulted in
similarly low levels of cross presentation (Fig. S3), and was unable to induce cross
presentation as efficiently as targeting MR or CD40, except at the highest antibody
concentrations used in immature MoDCs (Fig. 4B). Figure 4D shows donor matched pre-
processed HLA-A*0201+ peptide controls, with mature DCs presenting surface loaded
influenza M1 peptide more efficiently than immature DCs, in line with their higher surface
expression of MHC class I. CD40 antibodies coupled to the CMV pp65 peptide also
resulted in superior cross presentation as compared to DEC205-pp65 antibodies (Fig. 4E).
Thus, receptor-mediated delivery to early endosomes is a relevant determinant of cross
presentation efficiency for multiple epitopes. We determined whether a cytosolic
intermediate was involved for antibody-targeted cross presentation to both early and late
endosomes by inhibiting proteasomal activity. Cross presentation via all three receptors
required proteasomal degradation for the generation of peptides, while surface peptide
loading and presentation remained intact in the presence of epoxomicin (Fig. 4F).
Having shown that MHC class I cross presentation was superior when targeting
CD40 or MR over DEC205, we next asked if the same held true for processing and
presentation on MHC class II. We chemically coupled anti-CD40 and anti-DEC205 to an
extended MHC class II peptide from the NY-ESO protein that had a cysteine to alanine
mutation to facilitate conjugation (Fig. 5A, mutated residue bolded). The mutation did not
appear to reduce the ability of the peptide to be recognized by NY-ESO specific T cells
(Fig. 5B). Therefore, we next incubated BDCA1+ DCs with the antibody conjugates, and
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

16
cocultured DCs together with the NY-ESO peptide specific CD4 T cell clone; activation
was measured by monitoring the production of three cytokines (IFNγ, IL-2, and TNFα). As
found for cross presentation of other antigens on MHC class I, anti-CD40 was far more
efficient at mediating the presentation of a NY-ESO MHC class II epitope than was anti-
DEC205 over a 100-fold range of antibody concentrations (Fig. 5C, 5D, and 5E) and at
different DC: T cell ratios (Fig. 5F). Thus, antibody targeting to CD40 is not only superior
for cross presentation, but also for MHC class II presentation.
Taken together, these data indicate that antigen accumulation is not necessarily a
good predictor of cross presentation efficiency. Instead, receptors that target to early
compartments are quantitatively more adept at delivering antigen into the cross
presentation pathway than late endosomes, a finding that also seems to hold true for
MHC class II presentation. Interestingly, both endosomal destinations required
proteasomal degradation. Additionally, a receptor (CD40) that internalizes slowly but
targets to early endosomes appeared superior for antigen cross presentation.
Anti-CD40 does not enhance cross presentation by triggering DC maturation
We next asked if the anti-CD40 antibody’s superiority for cross presentation might
reflect its ability to generate an activation signal in DCs, especially since CD40 signaling is
known to enhance cross presentation in general 6,30. MoDCs were incubated overnight
with unlabeled CD40, MR, or DEC205 antibodies and assayed for maturation. Anti-CD40
slightly activated MoDCs, as gauged by CD86 and CD83 upregulation on a small cell
population. HLA-ABC (MHC class I) surface levels remained unaffected by the addition of
anti-CD40, indicating that the upregulation of HLA-ABC by anti-CD40 signaling was not
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

17
responsible for enhanced cross presentation (Fig. 6A). Anti-DEC205 and anti-MR
antibodies did not cause upregulation of any surface markers (Fig. 6A). Analysis of
supernatants collected from MoDCs treated overnight with these antibodies showed non-
detectable levels of IL-12p70 and minimal amounts of TNFα in response to anti-CD40 (Fig.
6B). These experiments indicate that the addition of the anti-CD40 antibody used here
(S2C6) induced a very low but detectable level of DC activation, consistent with this
antibody’s previous characterization as a very weak agonist in vitro.
To determine if the low level of CD40-mediated DC activation might itself enhance
cross presentation, we asked if the addition of anti-CD40 antibodies would enhance cross
presentation by the anti-DEC205-M1 and anti-MR-M1 antibody conjugates. DCs were
incubated with either anti-DEC205-M1 or anti-MR-M1 together with 1 μg/mL unconjugated
anti-CD40 or isotype control during the antigen uptake phase. In immature and mature
MoDCs (Fig. 6C-D) and in immature BDCA1+ DCs (Fig. 6E), the addition of CD40
antibodies did not enhance DEC205 cross presentation. In co-uptake experiments, we
observed that both MR and CD40 likely trafficked through the same compartments (data
not shown), and it is thought that antigen presentation is enhanced when both antigen and
agonist originate from the same compartment 31. However, when we co-administered MR-
M1 and anti-CD40, this did not enhance cross presentation via MR (Fig. 6C-E), indicating
that the addition of this anti-CD40 antibody did not enhance cross presentation by
triggering DC activation. Indeed, adding anti-CD40 antibodies also did not improve MHC
class II presentation by DEC205 (Fig. 6F), indicating that any anti-CD40 signal imparted
was negligible for both antigen presentation pathways tested.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

18
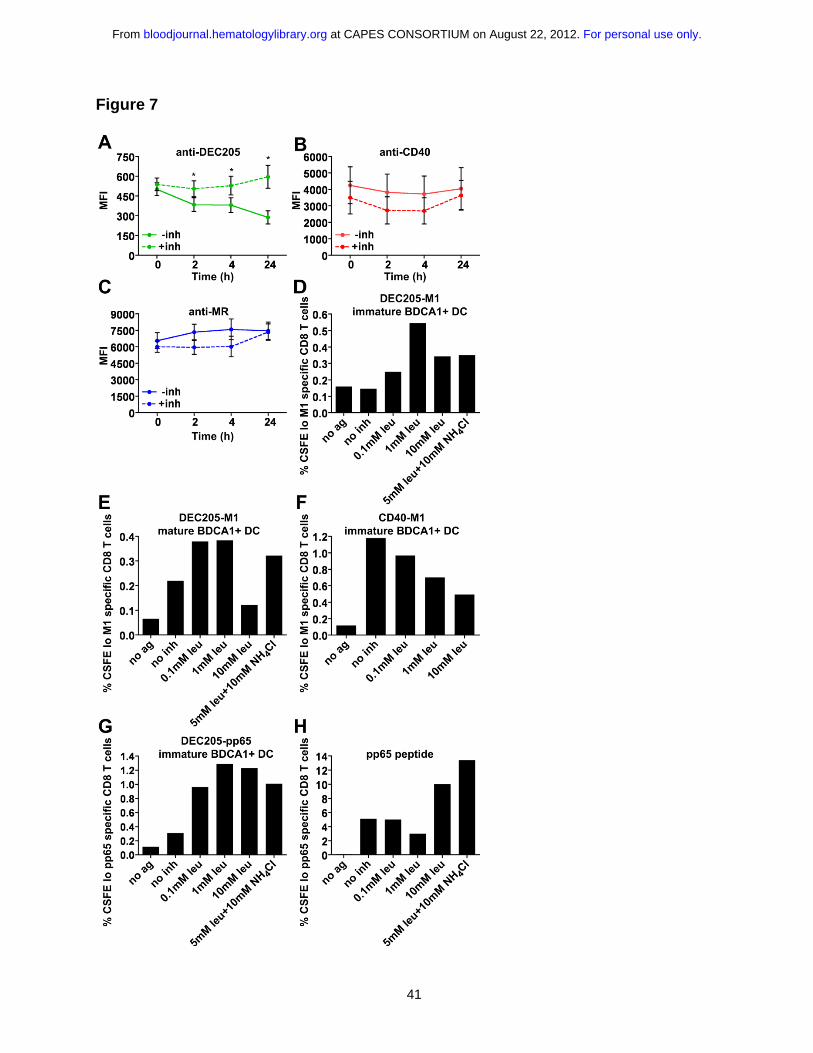
DEC205 antibodies are degraded over time, whereas CD40 and MR antibodies
remain stable
Having ruled out a role for CD40 signaling in enhancing cross presentation, and
having established that the extent of antigen uptake is not a key factor in cross
presentation efficiency, we wished to understand if differences in antibody degradation
were responsible, as suggested by our earlier accumulation experiments. To more directly
monitor antibody degradation, we pulsed MoDCs with Alexa 488 conjugated antibodies for
1 h at 4°C or 37°C, followed by washing and chasing for the indicated times. After 24 h,
there was a significant loss of anti-DEC205 signal (Fig. 7A) while both anti-CD40 and anti-
MR signal remained relatively constant (Fig. 7B, C), indicating that anti-DEC205 was
exposed to greater degradative forces. Parallel experiments were performed in cells
treated with leupeptin and ammonium chloride (inh) to inhibit endosomal-lysosomal
proteolysis. While we did not see an appreciable change in anti-MR and anti-CD40
fluorescence levels upon inhibitor addition (dashed lines), the inhibitors did rescue the
anti-DEC205 antibody signal, consistent with this antibody having been delivered to late
endosomes and subjected to degradation.
Inhibiting degradation rescues DEC205 targeted cross presentation
Conceivably, delivery to degradative compartments reduces cross presentation by
causing intra-lysosomal degradation of the internalized antibody-peptide conjugates prior
to release into the cytosol for proteasomal processing. Indeed, this difference might be
accentuated in our system, which relied on the use of relatively small peptide conjugates
that could be more easily digested than intact protein antigens. Resistance to degradation
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

19
is well known to be important for MHC class II presentation 1,32-34, with antigens more
resistant to degradation being more efficiently processed and presented. Our MHC class
II data may also reflect the differences in degradative capacity between the endosomal
compartments, as targeting CD40 elicited superior CD4+ T cell responses as compared to
DEC205 (Fig. 5). The same may be true in the case of cross presentation, given studies
showing enhancement when antigen presenting cells were treated with chloroquine 19,35.
To examine this possibility, we added protease and acidification inhibitors during the
antigen capture phase of our CD8+ T cell assay. As shown in Figure 7, the addition of
protease and acidification inhibitors enhanced cross presentation via DEC205 in both
immature (panel D) and mature DCs (panel E). This was true for influenza M1 and CMV
pp65 peptide conjugates (Fig. 7D, G). Interestingly, addition of increasing concentrations
of protease inhibitors reduced cross presentation via CD40 (Fig. 7F), indicating that some
level of proteolysis was required for escape to the cytosol and subsequent cross
presentation. Presentation of unconjugated peptide was largely unaffected by the inhibitor
treatments, except at the very highest concentrations (Fig. 7H). Thus, cross presentation
of antigens delivered to highly degradative compartments could be enhanced by inhibiting
the degradative capacity of these compartments. In sum, by targeting three receptors with
distinct endosomal localization patterns, we show that targeting early endosomal
receptors with antibody-peptide conjugates results in more efficient cross presentation,
due to the lowered degradative capacity of this compartment. Additionally, we find that
under conditions where proteases are inhibited, both late and early endosomal
compartments are able to support cross presentation, arguing against the existence of
specializations unique to a single endosome population.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

20
Discussion DCs harbor a variety of specializations that together enable their capacity for
antigen presentation and the control of T cell stimulation 1. Although a number of these
specializations affect the endosomal and phagosomal apparatus 36,37, the formation of
MHC class II-peptide complexes does not appear to be linked to any one intracellular site.
The situation is less clear in the case of cross presentation on MHC class I molecules.
Although antigen can be cross presented regardless of internalization mode 38-40, whether
it must access a specific endosomal compartment was uncertain. Previous reports have
implicated early endosomes as essential for cross presentation, based on a qualitative
comparison of receptor-mediated versus fluid phase ovalbumin uptake and the possible
presence of the TAP2 translocator by immunofluorescence microscopy 16,18, while other
studies with little emphasis on intracellular trafficking have determined that many
receptors are able to cross present equivalently with great efficiency 10,12,21,22.
We avoided making any assumptions concerning which endosomal compartment
was essential or what specializations it might possess for cross presentation by using high
affinity antibodies that targeted antigen to distinct compartments. This approach allowed a
direct, quantitative assessment of the relative importance of antigen internalization,
localization, and degradation in determining the efficiency of cross presentation in two
different human DC populations. Our results suggest that both early and late endosomal
compartments are capable of serving as antigen portals for cytosolic entry and cross
presentation. Late compartments appear to be less efficient for some antigens, however,
given their higher concentration of lysosomal enzymes, which degrade antigens before
they can be released into the cytosol. Accordingly, we find that the rapidity of degradation
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

21
can explain at least some of the compartment-specific differences in cross presentation.
The fact that inhibiting proteolysis enhances the ability of late endosomes and lysosomes
to allow cross presentation of accumulated antigens also means that it is unlikely that
early endosomes alone contain the specializations required for antigen egress or for re-
importation for loading onto endosomal MHC class I molecules, as proposed previously 18.
Our ability to detect a relationship between degradation and presentation from late
compartments may reflect the nature of the antigen delivery platform we have developed,
i.e., the use of extended peptides chemically coupled to anti-receptor antibodies. Previous
work has relied mostly on the use of heavy chain fusions of full-length antigens 9,10, which
may be inherently more resistant to degradation. In our hands, such fusions also have a
significant tendency to aggregate and to be proteolysed during production, which would
complicate potential scale up for application in human patients. Our peptide platform
offers the advantage of scalability and combinatorial flexibility (allowing for delivery of
different peptides), although the potential decrease in peptide antigen stability may require
the targeting of such constructs to early endosomes. Indeed, protein stability may be a
reason why targeting DEC205 is inferior for cross presentation; peptide antigens are likely
more susceptible to degradation, which would decrease their ability to survive long
enough to escape into the cytosol. The use of more susceptible peptide antigens may
also explain why anti-CD40 was superior to anti-DEC205 for targeting MHC class II
presentation. Given that an inverse relationship between antigen stability and formation of
peptide-MHC class II complexes is known to exist 33, it is perhaps not too surprising that a
similar situation might exist for cross presented antigens.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

22
We were, however, surprised by the observation that targeting CD40 mediated
more efficient cross presentation than MR. Both receptors targeted internalized peptide-
antibody conjugates to early endosomes, but CD40 was substantially less efficient at
antibody uptake. The enhanced cross presentation was not due to maturation effects of
anti-CD40, despite the fact that CD40 stimulation can be important for activating cross
presentation by mouse DCs 6,30. The one obvious difference was in the localization of
antibodies even after extended time points. Whereas the bulk of anti-MR was internalized,
most anti-CD40 remained at the plasma membrane from which it was presumably
continuously but more slowly internalized, or to which it was more rapidly recycled.
Conceivably, maximum antigen stability may be achieved when antibodies are
internalized slowly to early and recycling endosomes. This limits the time that the
antibody-peptide conjugate spends intracellularly, where early endosomal proteases
(albeit fewer) gain access to antigens 41,42, and provides a continuous “time-release” pool
of antigen that might be used over extended periods for the continuous formation of
peptide-MHC class I complexes. Indeed, preliminary cross presentation data from another
slowly internalizing receptor, CD11c, appears to support this hypothesis (B.C. and I.M.,
unpublished data, February 1, 2012). Whatever the underlying mechanism, this
observation was unexpected and indicates that rapid endocytosis and antigen
accumulation may not always be the best criteria for choosing receptors to target vaccines
to DCs.
Our data may additionally point to some cell type specific differences in cross
presentation efficiency. Both DC types examined exhibited similar patterns of receptor
expression, localization, antibody accumulation, and cross presentation. However, while
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

23
MoDCs were more endocytic overall, they also appeared to be more degradative,
especially when we assayed for DEC205 targeting. We generally see that immature
MoDCs exhibit lower cross presentation efficiency than BDCA1+ DCs, and one reason for
this may be due to higher protease levels in MoDCs as compared to BDCA1+ DCs 28,29.
The existence of distinct DC subsets that have variable capacities for cross presentation
may mean that any such specializations could also be cell type specific. Indeed, recent in
vivo work in the mouse has demonstrated that as long as a given antibody (DEC205,
Langerin, CLEC9A/DNGR-1) can target CD8α+ cross presenting DCs, cross presentation
will occur 10. Recent studies on DC subtypes have implicated BDCA3+ DCs as the human
equivalent of the mouse CD8α+ DCs 43-46, a DC type that is key for efficient cross
presentation in vivo in mice 47-49. Initial data from these groups indicates that BDCA3+
DCs are superior for cross presentation, compared to BDCA1+ DCs. Our data indicate
that degradation of antigen affects the efficiency of cross presentation by human DCs,
including BDCA1+ DCs, and that slower antigen internalization to early endosomes may
be superior. Therefore, it may be possible that differential antigen handling by BDCA3+
DCs may be the reason for these differences, potentially through altered antigen
accumulation or degradation, or perhaps through increased access to the cytosol.
Further studies to elucidate the requirements for efficient cross presentation will be
key for efficient DC targeting. Since we see that multiple DC types are able to cross
present, it will be advantageous to target more than one type of DC simultaneously to
elicit the most efficient CD8+ T cell responses. Based on our results, this will best be
accomplished not only by taking into account surface receptor expression, but also by
determining the intracellular localization and degradation of the trafficking antibodies of
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

24
interest.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

25
Authorship
Contributions: B.C. and I.M. designed the experiments with help from A.S.-S.; B.C.
performed the experiments; B.C. and I.M. analyzed the data; A.S.-S., L.D., and I.M.
supervised the project; L.C. assisted with primary DC isolation; C.C. assisted with imaging
analysis; R.V. and B-C. L. generated the antibody-peptide conjugates; J.W. and T.K.
provided the B11 and 3G9 antibodies; B.C. and I.M. wrote the paper; and all authors
approved this manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests. B.C.,
L.C., C.C., R.V., B-C. L, L.D., and I.M. are employees of Genentech. J.W. and T.K. are
employees of Celldex Therapeutics.
Acknowledgements We would like to thank the Genentech Research Blood Program and Blood Centers of the
Pacific for donor coordination and leukapheresis, as well as Laurie Gilmour and Yelena
Dayter for technical assistance with elutriations. We would especially like to thank
members of the Mellman lab for advice and fruitful discussions, and Peter Ebert for the
critical reading of this manuscript. The authors dedicate this manuscript to the memory of
our mentor and dear friend, Ralph Steinman.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

26
References
1. Trombetta ES, Mellman I. Cell biology of antigen processing in vitro and in vivo. Annu Rev Immunol. 2005;23:975-1028.
2. Delamarre L, Mellman I. Harnessing dendritic cells for immunotherapy. Seminars in immunology. 2011;23(1):2-11.
3. Caminschi I, Lahoud MH, Shortman K. Enhancing immune responses by targeting antigen to DC. Eur J Immunol. 2009;39(4):931-938.
4. Palucka K, Banchereau J, Mellman I. Designing vaccines based on biology of human dendritic cell subsets. Immunity. 2010;33(4):464-478.
5. Sancho D, Mourao-Sa D, Joffre OP, et al. Tumor therapy in mice via antigen targeting to a novel, DC-restricted C-type lectin. J Clin Invest. 2008;118(6):2098-2110.
6. Bonifaz LC, Bonnyay DP, Charalambous A, et al. In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptor improves T cell vaccination. J Exp Med. 2004;199(6):815-824.
7. Mellman I, Coukos G, Dranoff G. Cancer immunotherapy comes of age. Nature. 2011:In press.
8. Segura E, Villadangos JA. Antigen presentation by dendritic cells in vivo. Curr Opin Immunol. 2009;21(1):105-110.
9. Bozzacco L, Trumpfheller C, Siegal FP, et al. DEC-205 receptor on dendritic cells mediates presentation of HIV gag protein to CD8+ T cells in a spectrum of human MHC I haplotypes. Proc Natl Acad Sci U S A. 2007;104(4):1289-1294.
10. Idoyaga J, Lubkin A, Fiorese C, et al. Comparable T helper 1 (Th1) and CD8 T-cell immunity by targeting HIV gag p24 to CD8 dendritic cells within antibodies to Langerin, DEC205, and Clec9A. Proc Natl Acad Sci U S A. 2011;108(6):2384-2389.
11. Klechevsky E, Flamar AL, Cao Y, et al. Cross-priming CD8+ T cells by targeting antigens to human dendritic cells through DCIR. Blood. 2010;116(10):1685-1697.
12. Tsuji T, Matsuzaki J, Kelly MP, et al. Antibody-targeted NY-ESO-1 to mannose receptor or DEC-205 in vitro elicits dual human CD8+ and CD4+ T cell responses with broad antigen specificity. J Immunol. 2011;186(2):1218-1227.
13. Tacken PJ, Ginter W, Berod L, et al. Targeting DC-SIGN via its neck region leads to prolonged antigen residence in early endosomes, delayed lysosomal degradation and cross-presentation. Blood. 2011;118(15):4111-4119.
14. Tacken PJ, Joosten B, Reddy A, et al. No advantage of cell-penetrating peptides over receptor-specific antibodies in targeting antigen to human dendritic cells for cross-presentation. J Immunol. 2008;180(11):7687-7696.
15. Lahoud MH, Proietto AI, Ahmet F, et al. The C-type lectin Clec12A present on mouse and human dendritic cells can serve as a target for antigen delivery and enhancement of antibody responses. J Immunol. 2009;182(12):7587-7594.
16. Burgdorf S, Kautz A, Bohnert V, Knolle PA, Kurts C. Distinct pathways of antigen uptake and intracellular routing in CD4 and CD8 T cell activation. Science. 2007;316(5824):612-616.
17. Zehner M, Chasan AI, Schuette V, et al. Mannose receptor polyubiquitination regulates endosomal recruitment of p97 and cytosolic antigen translocation for cross-presentation. Proc Natl Acad Sci U S A. 2011;108(24):9933-9938.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

27
18. Burgdorf S, Scholz C, Kautz A, Tampe R, Kurts C. Spatial and mechanistic separation of cross-presentation and endogenous antigen presentation. Nat Immunol. 2008;9(5):558-566.
19. Belizaire R, Unanue ER. Targeting proteins to distinct subcellular compartments reveals unique requirements for MHC class I and II presentation. Proc Natl Acad Sci U S A. 2009;106(41):17463-17468.
20. Harding CV, Collins DS, Slot JW, Geuze HJ, Unanue ER. Liposome-encapsulated antigens are processed in lysosomes, recycled, and presented to T cells. Cell. 1991;64(2):393-401.
21. Castro FV, Tutt AL, White AL, et al. CD11c provides an effective immunotarget for the generation of both CD4 and CD8 T cell responses. Eur J Immunol. 2008;38(8):2263-2273.
22. Kratzer R, Mauvais FX, Burgevin A, Barilleau E, van Endert P. Fusion proteins for versatile antigen targeting to cell surface receptors reveal differential capacity to prime immune responses. J Immunol. 2010;184(12):6855-6864.
23. Mellman I, Plutner H. Internalization and degradation of macrophage Fc receptors bound to polyvalent immune complexes. J Cell Biol. 1984;98(4):1170-1177.
24. Mellman I, Plutner H, Ukkonen P. Internalization and rapid recycling of macrophage Fc receptors tagged with monovalent antireceptor antibody: possible role of a prelysosomal compartment. J Cell Biol. 1984;98(4):1163-1169.
25. Mahnke K, Guo M, Lee S, et al. The dendritic cell receptor for endocytosis, DEC-205, can recycle and enhance antigen presentation via major histocompatibility complex class II-positive lysosomal compartments. J Cell Biol. 2000;151(3):673-684.
26. Platt CD, Ma JK, Chalouni C, et al. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc Natl Acad Sci U S A. 2010;107(9):4287-4292.
27. Smed-Sorensen A, Lore K, Vasudevan J, et al. Differential susceptibility to human immunodeficiency virus type 1 infection of myeloid and plasmacytoid dendritic cells. Journal of virology. 2005;79(14):8861-8869.
28. Burster T, Beck A, Tolosa E, et al. Differential processing of autoantigens in lysosomes from human monocyte-derived and peripheral blood dendritic cells. J Immunol. 2005;175(9):5940-5949.
29. McCurley N, Mellman I. Monocyte-derived dendritic cells exhibit increased levels of lysosomal proteolysis as compared to other human dendritic cell populations. PLoS One. 2010;5(8):e11949.
30. Delamarre L, Holcombe H, Mellman I. Presentation of exogenous antigens on major histocompatibility complex (MHC) class I and MHC class II molecules is differentially regulated during dendritic cell maturation. J Exp Med. 2003;198(1):111-122.
31. Blander JM, Medzhitov R. Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature. 2006;440(7085):808-812.
32. Delamarre L, Pack M, Chang H, Mellman I, Trombetta ES. Differential lysosomal proteolysis in antigen-presenting cells determines antigen fate. Science. 2005;307(5715):1630-1634.
33. Delamarre L, Couture R, Mellman I, Trombetta ES. Enhancing immunogenicity by limiting susceptibility to lysosomal proteolysis. J Exp Med. 2006;203(9):2049-2055.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

28
34. Ziegler HK, Unanue ER. Decrease in macrophage antigen catabolism caused by ammonia and chloroquine is associated with inhibition of antigen presentation to T cells. Proc Natl Acad Sci U S A. 1982;79(1):175-178.
35. Accapezzato D, Visco V, Francavilla V, et al. Chloroquine enhances human CD8+ T cell responses against soluble antigens in vivo. J Exp Med. 2005;202(6):817-828.
36. Savina A, Jancic C, Hugues S, et al. NOX2 controls phagosomal pH to regulate antigen processing during crosspresentation by dendritic cells. Cell. 2006;126(1):205-218.
37. Trombetta ES, Ebersold M, Garrett W, Pypaert M, Mellman I. Activation of lysosomal function during dendritic cell maturation. Science. 2003;299(5611):1400-1403.
38. Albert ML, Sauter B, Bhardwaj N. Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature. 1998;392(6671):86-89.
39. Iyoda T, Shimoyama S, Liu K, et al. The CD8+ dendritic cell subset selectively endocytoses dying cells in culture and in vivo. J Exp Med. 2002;195(10):1289-1302.
40. Norbury CC, Chambers BJ, Prescott AR, Ljunggren HG, Watts C. Constitutive macropinocytosis allows TAP-dependent major histocompatibility complex class I presentation of exogenous soluble antigen by bone marrow-derived dendritic cells. Eur J Immunol. 1997;27(1):280-288.
41. Chapman HA. Endosomal proteases in antigen presentation. Curr Opin Immunol. 2006;18(1):78-84.
42. Shen L, Sigal LJ, Boes M, Rock KL. Important role of cathepsin S in generating peptides for TAP-independent MHC class I crosspresentation in vivo. Immunity. 2004;21(2):155-165.
43. Crozat K, Guiton R, Contreras V, et al. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8alpha+ dendritic cells. J Exp Med. 2010;207(6):1283-1292.
44. Jongbloed SL, Kassianos AJ, McDonald KJ, et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J Exp Med. 2010;207(6):1247-1260.
45. Poulin LF, Salio M, Griessinger E, et al. Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8alpha+ dendritic cells. J Exp Med. 2010;207(6):1261-1271.
46. Bachem A, Guttler S, Hartung E, et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J Exp Med. 2010;207(6):1273-1281.
47. den Haan JM, Lehar SM, Bevan MJ. CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J Exp Med. 2000;192(12):1685-1696.
48. Pooley JL, Heath WR, Shortman K. Cutting edge: intravenous soluble antigen is presented to CD4 T cells by CD8- dendritic cells, but cross-presented to CD8 T cells by CD8+ dendritic cells. J Immunol. 2001;166(9):5327-5330.
49. Smith CM, Belz GT, Wilson NS, et al. Cutting edge: conventional CD8 alpha+ dendritic cells are preferentially involved in CTL priming after footpad infection with herpes simplex virus-1. J Immunol. 2003;170(9):4437-4440.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

29
Figure legends Figure 1: Human BDCA1+ DCs and MoDCs express mannose receptor, CD40, and
DEC205.
Donor-matched BDCA1+ DCs, or MoDCs were washed and incubated with (A) anti-
mannose receptor (MR), (B) anti-CD40 or (C) anti-DEC205 antibodies for 25 min at 4°C.
DCs were washed, fixed, and analyzed by flow cytometry. Histograms show one
representative of ten independent donors. (D) BDCA1+ DCs (closed symbols) and
MoDCs (open symbols) were incubated with Alexa 488 conjugated anti-MR, CD40, or
DEC205 for 25 min at 4°C, washed, and analyzed by flow cytometry. MFI was normalized
for the number of fluorophores per antibody, in order to directly compare surface levels
between receptors. The graph shows data from at least three independent donors with the
mean MFI depicted.
Figure 2: Targeting to DEC205 results in late endosomal localization while CD40
and mannose receptor antibodies are largely excluded from late endosomes.
(A-C) BDCA1+ DCs or (D-E) MoDCs were continuously incubated with 1 μg/mL of
fluorescently labeled (A and D) anti-DEC205, (B and E) anti-MR or (C and F) anti-CD40
antibodies for 6 h at 37°C. In parallel, DCs were incubated with antibodies in the presence
of protease and acidification inhibitors leupeptin and ammonium chloride (inh). DCs were
seeded on coverslips and fixed, followed by staining for the cell surface (HLA-DR) and
permeabilization and labeling of early endosomes (EEA1) or late endosomes (Lamp1).
Arrows indicate areas of overlap. Images were acquired on a Leica SP5 confocal
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

30
microscope, 100x oil objective (N.A: 1.47), zoom 7. All immunofluorescence experiments
were repeated in at least five independent donors. Scale bar is 5 µm.
Figure 3: Antibodies to DEC205 and mannose receptor are accumulated more
efficiently by DCs than anti-CD40.
(A) BDCA1+ DCs or (B) MoDCs were continuously incubated with 1 μg/mL of
fluorescently labeled anti-MR, anti-CD40, or anti-DEC205 for the indicated times, washed,
counterstained, and fixed (top panels). Middle and bottom panels: data from the top panel,
together with matched samples treated with leupeptin and ammonium chloride (inh)
during the continuous incubation. MuIso is the isotype for anti-CD40, while huIso is the
isotype for anti-MR and anti-DEC205. Graphs depict mean normalized MFI from at least
three independent donors ± SEM. Normalized MFI indicates amount of accumulation; MFI
was calculated by removing the contribution of surface fluorescence (4°C control) and
normalizing the MFI for the number of fluorophores per antibody. Differences in antibody
accumulation in the absence or presence of inhibitors were assessed using the paired t
test and statistically significant differences depicted (*), p<0.05.
Figure 4: Targeting CD40 and mannose receptor leads to superior cross
presentation compared to targeting DEC205.
(A) Schematic for the generation of antibody-peptide conjugates. On average, our peptide
conjugates had 1 peptide per antibody. (B) Matched immature or (C) mature HLA-
A*0201+ DCs were incubated with various concentrations of anti-CD40-M1, anti-MR-M1,
and anti-DEC205-M1 for 4-6 h, washed, and cocultured with autologous CFSE labeled
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

31
CD8+ T cells for 8-10 d in the presence of LPS and IL-2. “Immature” or “mature” refers to
the activation state of the DC prior to antigen uptake and coculture. MuIso-M1 is the
isotype for anti-CD40-M1, while huIso-M1 is the isotype for anti-MR-M1 and anti-DEC205-
M1. Graphs depict frequencies of total CFSElo and Influenza M1 (58-66) specific CD8+ T
cells. One representative experiment of six to eight independent donors (immature
samples) or three to four independent donors (mature samples) is shown. Targeting via
anti-CD40 and MR was judged superior to targeting via anti-DEC205 across six to eight
independent donors using a paired t test at 1 μg/mL for immature DCs (CD40 vs.
DEC205: MoDC p=0.0270, BDCA1+ DC p=0.0337; MR vs. DEC205: MoDC p=0.0264;
BDCA1+ DC was not statistically significant possibly due to variable surface MR
expression). (D) Influenza M1 (58-66) non-extended peptide control for antigen
presentation assay. (E) (Top panel) as in (B), DCs were incubated with anti-CD40-pp65 or
anti-DEC205-pp65 conjugates. (Bottom panel) CMV pp65 (495-503) non-extended
peptide control. (F) DCs were incubated with 1 μg/mL antibody-M1 conjugates or 25
ng/mL M1 peptide ± 0.1 μM epoxomicin or DMSO for 4-6 h, followed by washing and
coculture with autologous CFSE labeled CD8+ T cells. “No ag” shows the background
when DCs and CD8+ T cells are cocultured in the absence of antigen. Graphs depict
frequencies of total CFSElo and influenza M1 (58-66) specific CD8+ T cells. One
representative experiment from at least three independent donors is shown.
Figure 5: Targeting CD40 results in superior MHC class II presentation than
targeting DEC205.
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

32
(A) NY-ESO extended peptide sequence. The amino acid change from cysteine �
alanine is indicated in bold. (B) DPB1*0401+ BDCA1+ DCs were incubated with indicated
doses of irrelevant HIV reverse transcriptase peptide (HIV RTp), WT NY-ESO peptide
(WT NYESOp), or C�A mutated NY-ESO peptide (mut NYESOp) for 1.5 h, followed by
washing and coculture with an NY-ESO peptide specific CD4+ T cell clone. Percentage of
CD4+ T cells with intracellular IFNγ results are shown as one representative of three
independent DC donors. (C-E) 0.1-10 μg/mL antibody-NY-ESO peptide conjugates were
fed to BDCA1+ DCs for 1.5 h, followed by washing and coculture with an NY-ESO specific
CD4+ T cell clone at a DC: T cell ratio of 1:1, and subsequent staining for intracellular
cytokines. The percentage of CD4+ T cells that are positive for (C) IFNγ, (D) IL-2, and (E)
TNFα are depicted. One representative of three independent DC donors is shown. (F) As
in (C-E), using 1 μg/mL antibody-NY-ESO conjugates with varying DC:T cell ratios. The
percentage of CD4+ T cells that are positive for IFNγ is depicted. One representative of
three independent DC donors is shown.
Figure 6: Anti-CD40 does not enhance cross presentation of mannose receptor or
DEC205.
(A) 1 μg/mL of anti-CD40 (S2C6), anti-MR (B11), anti-DEC205 (3G9), isotype (iso), or 200
ng/mL LPS were added to MoDCs overnight. Following overnight culture, supernatants
were harvested and DCs were labeled for surface markers that are normally upregulated
following maturation, as indicated. One representative of three independent donors is
shown. (B) Supernatants from (A) were analyzed for cytokine production using Luminex
technology. IL-12p70 levels were insignificant, except for LPS controls. (C-D) MoDCs or
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

33
(E) BDCA1+ DCs were incubated with 1 μg/mL antibody-M1 peptide conjugates alone or
with antibody-M1 conjugates + 1 μg/mL unconjugated CD40 (S2C6) antibody or 1 μg/mL
isotype (muIso) together for 4-6 h, followed by washing and coculture with autologous
CFSE labeled CD8+ T cells. Graphs depict frequencies of total CFSElo and Influenza M1
(58-66) specific CD8 T cells. One representative of three independent donors is shown.
(F) BDCA1+ DCs were incubated with 1 μg/mL DEC205-NY-ESO peptide conjugates
alone or with DEC205-NY-ESO peptide conjugates + 1 μg/mL unconjugated CD40
(S2C6) antibody or 1 μg/mL isotype (muIso) together for 1.5 h, followed by washing and
coculture with an NY-ESO specific CD4+ T cell clone. Graphs depict percentage of IFNγ+
CD4+ T cells. One representative of two independent DC donors is shown.
Figure 7: DEC205 targeted cross presentation is rescued by the addition of
protease and acidification inhibitors.
4 μg/mL Alexa 488 covalently conjugated (A) anti-DEC205 (3G9), (B) anti-CD40 (S2C6),
or (C) anti-MR (B11) antibodies were internalized by MoDCs for 1 h at 37°C. Cells were
washed extensively and chased for indicated times ± leupeptin and ammonium chloride
(inh), followed by fixation and analysis by flow cytometry. Graphs depict the mean MFI of
three independent donors ± SEM. MFI differences in the absence or presence of inhibitors
were assessed using the paired t test and statistically significant differences depicted (*),
p<0.05. (D) Immature or (E) mature BDCA1+ DCs were incubated with 1 μg/mL DEC205-
M1 conjugates ± indicated inhibitors for 4-6 h, followed by washing and coculture with
autologous CFSE labeled CD8+ T cells. Graphs depict frequencies of CFSElo, influenza
M1 (58-66) specific CD8+ T cells. One representative of three independent donors is
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

34
shown for (D) and one representative of two independent donors is shown for (E). (F)
Immature BDCA1+ DCs were incubated with 1 μg/mL CD40-M1 conjugates ± indicated
inhibitors for 4-6 h, followed by washing and coculture with autologous CFSE labeled
CD8+ T cells. Graphs depict frequencies of total CFSElo and Influenza M1 (58-66) specific
CD8+ T cells. One representative of three independent donors is shown. (G) BDCA1+
DCs were incubated with 1 μg/mL DEC205-pp65 conjugates ± indicated inhibitors for 4-6
h, followed by washing and coculture with autologous CFSE labeled CD8+ T cells. Graphs
depict frequencies of CFSElo, CMV pp65 (495-503) specific CD8+ T cells. One
representative of two independent donors is shown. (H) CMV pp65 (495-503) non-
extended peptide control (25 ng/mL).
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

35
Figure 1
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

36
Figure 2
F
or personal use only. at C
AP
ES
CO
NS
OR
TIU
M on A
ugust 22, 2012. bloodjournal.hem
atologylibrary.orgF
rom

37
Figure 3
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom
38
Figure 4
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

39
Figure 5
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom

40
Figure 6
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom
41
Figure 7
For personal use only. at CAPES CONSORTIUM on August 22, 2012. bloodjournal.hematologylibrary.orgFrom





























