Embed Size (px)
Citation preview

J. Zool.. Lond. (1984) 203, 95-101
Interspecific spawning between Common frogs ( R a m temporaria) and Common toads (Bufo bufo)
C . J . R E A D I N G Institute of’ Terrestrial Ecology, Furzebrook Research Station, Wareham, Dorset, U.K.
(Accepted 20 September 1983)
(With 4 figures in the text)
The interaction between Common frogs (Rana iemporaria) and Common toads (Bufo buJb) during the breeding season was studied at a small pond on Portland, Dorset. Although the frogs started and finished spawning earlier than the toads there was a period during which interspecific spawning between female toads and male frogs took place. This appears to have resulted from reduced male toad activity caused by the cold breeding season given that spatial, temporal and behavioural species separation did not occur.
Page Contents Introduction . . . . . . . . . . . . . . . . . . . . 95 Methods 95 . . . . . . . . . . . . . . . . . . . . . . Results 96 Discussion 99 References . . . . . . . . . . . . . . . . . . . . 101
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Introduction The indescriminate pairing behaviour of both male Common frogs (Rana temporaria L.)
and male Common toads (Buf’o bofo L.) during the breeding season is well known. Smith (1951) states that male toads will readily grab any object similar in girth to a female and that males of both species have been found in amplexus with fish. He also states that in those exceptional years when both frogs and toads spawned together, mixed couples were often found. Usually, however, when both species breed in the same pond they either select different sites within it in which to spawn (Smith, 1951; Cooke, 1975; Strijbosch, 1979) or they spawn at different times (Cooke, 1977).
In this paper the occurrence is described and the causes of interspecific spawning between breeding populations of Common frogs and Common toads’ at a small pond on Portland, Dorset in 1983 are investigated.
Methods During the spring of I983 a mixed breeding population ofCommon toad (Bufb bufo L.) and Common
frog (Rana temporaria L.) was studied at a small, shallow, man-made pond on Portland, Dorset. A detailed description of the methods used to catch and mark the animals can be found in Reading &
0022-5460/84/0 I0095 + 07 $03.00/0 95
0 I984 The Zoological Society of London
96 C . J. READING
20
15
10
5
0
( a )
February March
1983
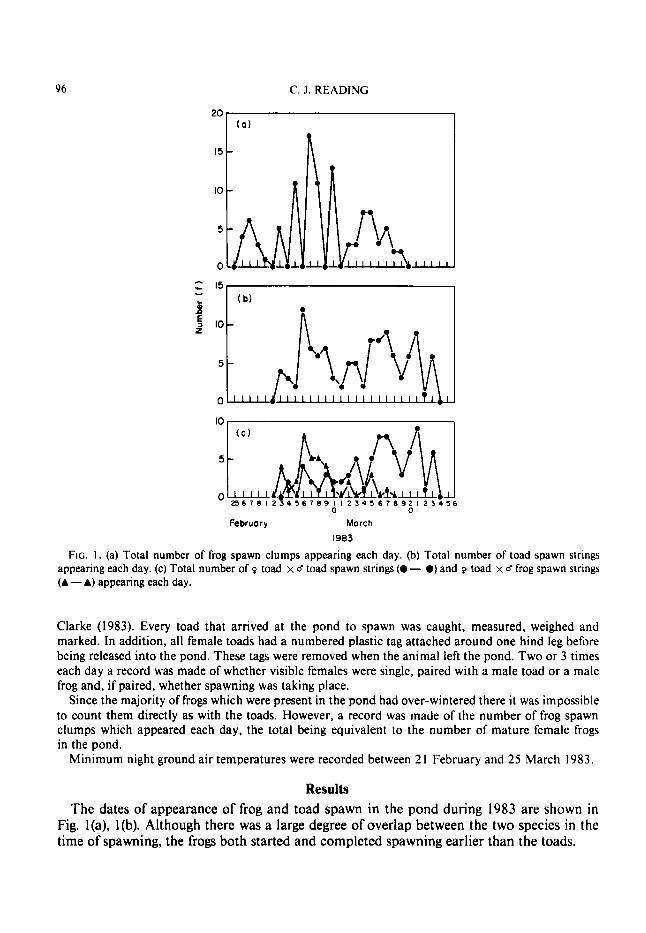
FIG. 1 . (a) Total number of frog spawn clumps appearing each day. (b) Total number of toad spawn strings appearing each day. (c) Total number of 9 toad x d toad spawn strings (0 - 0) and 9 toad x d frog spawn strings (A - A) appearing each day.
Clarke (1983). Every toad that arrived at the pond to spawn was caught, measured, weighed and marked. In addition, all female toads had a numbered plastic tag attached around one hind leg before being released into the pond. These tags were removed when the animal left the pond. Two or 3 times each day a record was made of whether visible females were single, paired with a male toad or a male frog and, if paired, whether spawning was taking place.
Since the majority of frogs which were present in the pond had over-wintered there it was impossible to count them directly as with the toads. However, a record was made of the number of frog spawn clumps which appeared each day, the total being equivalent to the number of mature female frogs in the pond.
Minimum night ground air temperatures were recorded between 2 1 February and 25 March 1983.
Results The dates of appearance of frog and toad spawn in the pond during 1983 are shown in
Fig. l(a), l(b). Although there was a large degree of overlap between the two species in the time of spawning, the frogs both started and completed spawning earlier than the toads.

INTERSPECIFIC SPAWNING BETWEEN FROGS A N D TOADS 91
TABLE I S p i w t n u n runk corrdation coelficienrs
Toad x Toad Toad x Frog Total Toad spawn spawn spawn
Frog spawn r , = -0.05 r,,=O.43 r,=0,1 I P>0.05 P<O.OS P>0.05
Toad x Toad spawn r , = -0.16 r,r=0.85 P>0.05 P<O.OI
Toad x Frog spawn r, = 0.46 P<O.OI
The frogs produced a peak of spawning activity between 2 and 1 1 March in which 55% (57 clumps) of the total frog spawn was laid. This was preceded (25 February-2 March) and followed ( 1 1-20 March) by periods of reduced spawning activity in which 14% (14 clumps) and 3 1% (32 clumps) respectively, of the total spawn was produced.
The toads showed two peaks of spawning activity, the first between 2 and 1 1 March pro- duced 40% (46 strings) and the second larger peak between 14 and 24 March produced 51% (58 strings) of the total spawn laid. These figures for the toad refer only to those females in which spawning was observed.
The appearance of toad spawn can, however, be divided into spawn produced by Q toad x d toad pairings and spawn produced by Q toad x d frog pairings, (Fig. I(c)). This shows that the first peak of toad spawning activity was mainly the result of toadxfrog pairings (65%), whilst the second was the result of toadx toad pairings (91%). These figures do not include two females observed spawning without a mate and eight females who spawned par- tially with both a male frog and a male toad. During the whole spawning period a minimum of 37 (30%) out of 124 observed female toads, spawned with male frogs.
The times of appearance of frog spawn, toad x toad spawn and toad x frog spawn were compared using the Spearman Rank Correlation (Table I). Although no significant corre- lation was found between the appearances of frog spawn and toad x toad spawn ( r , = -0.05, f 5 0 . 0 5 ) or toadxtoad spawn and toadxfrog spawn ( r ,= -0.16, P>0-05) one was found between the appearance of frog spawn and toad x frog spawn (r,=0.43, P<0.05).
Most of the female toads (87%) which were observed spawning with male frogs arrived at the pond paired with male toads. This suggested that male frogs were achieving takeovers during fights with paired male toads. The body length of male toads losing a female to a male frog was therefore compared, using the Student's t-test, with the body length of those spawning with a female. No significant difference was found (t=0.96, d.f. =91, P>0.05). Similarly, female toads found spawning with male frogs were not significantly different in body length (d=0.82, P>O.IO) from those spawning with male toads.
Because the eggs produced from a toadxfrog spawning are infertile, ways in which a female toad might increase her chance of losing an attached male frog were investigated. First, spawning might be delayed, thus increasing the probability of a male toad achieving a takeover. The time spent in the pond before spawning was therefore calculated for all
98 C. J . READING
0
0
0 b
0
0 0
0
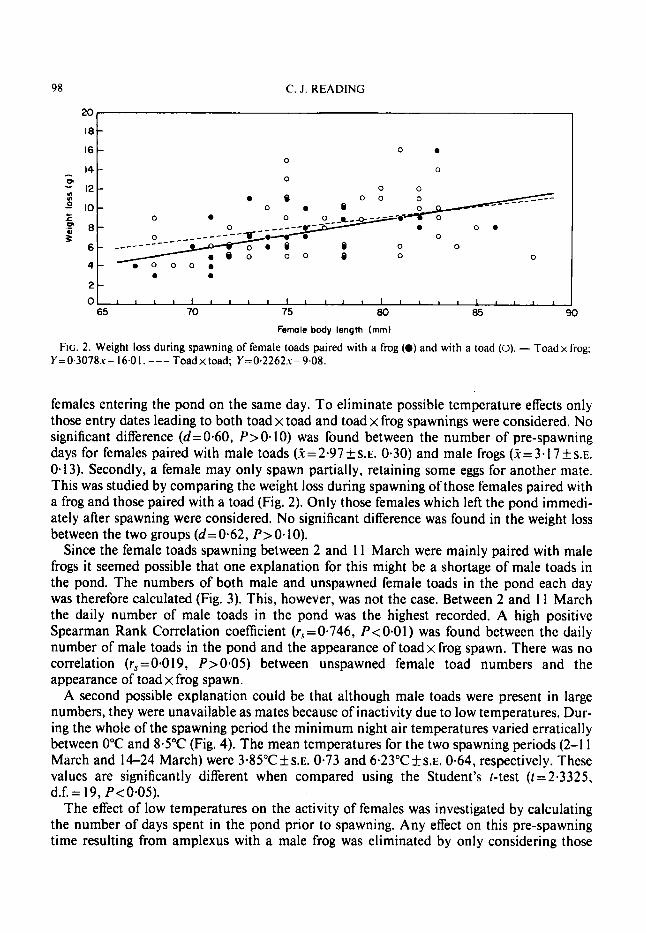
FIG. 2. Weight loss during spawning of female toads paired with a frog (0) and with a toad (0). - Toad x frog; Y=0.3078.u- 16.01. --- Toadxtoad; Y=0.2262s-9.08.
females entering the pond on the same day. To eliminate possible temperature effects only those entry dates leading to both toad x toad and toad x frog spawnings were considered. No significant difference (d=0.60, P>O. 10) was found between the number of pre-spawning days for females paired with male toads ( 3 = 2 + 9 7 f s . ~ . 0.30) and male frogs (5=3.17+s.~. 0.13). Secondly, a female may only spawn partially, retaining some eggs for another mate. This was studied by comparing the weight loss during spawning of those females paired with a frog and those paired with a toad (Fig. 2). Only those females which left the pond immedi- ately after spawning were considered. No significant difference was found in the weight loss between the two groups (d=0-62, P>O-lO).
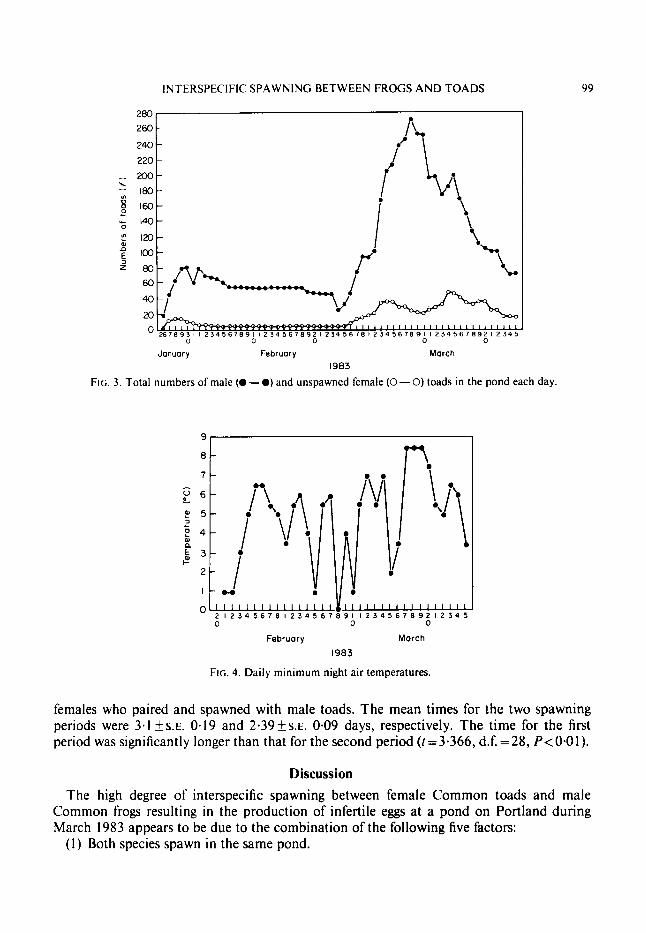
Since the female toads spawning between 2 and 1 1 March were mainly paired with male frogs it seemed possible that one explanation for this might be a shortage of male toads in the pond. The numbers of both male and unspawned female toads in the pond each day was therefore calculated (Fig. 3). This, however, was not the case. Between 2 and 1 1 March the daily number of male toads in the pond was the highest recorded. A high positive Spearman Rank Correlation coefficient (r,y=0.746, P < O - O I ) was found between the daily number of male toads in the pond and the appearance of toad x frog spawn. There was no correlation (rs =0-019, P>0.05) between unspawned female toad numbers and the appearance of toad x frog spawn.
A second possible explanation could be that although male toads were present in large numbers, they were unavailable as mates because of inactivity due to low temperatures. Dur- ing the whole of the spawning period the minimum night air temperatures varied erratically between 0°C and 8.5"C (Fig. 4). The mean temperatures for the two spawning periods (2-1 1 March and 14-24 March) were 3.85"Cks.E. 0.73 and 6.23"Cks.~. 0.64, respectively. These values are significantly different when compared using the Student's t-test (f=2.3325, d.f. = 19, P<0*05).
The effect of low temperatures on the activity of females was investigated by calculating the number of days spent in the pond prior to spawning. Any effect on this pre-spawning time resulting from amplexus with a male frog was eliminated by only considering those
INTERSPECIFIC SPAWNING BETWEEN FROGS AND TOADS
260
240
99
- iL - ?' I
' 2 6 7 8 9 3 1 I 2 3 4 5 6 7 8 9 1 1 2 3 4 5 6 7 8 9 2 1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8 9 1 1 2 3 4 5 6 7 8 9 2 1 2 3 4 5 ' 2 6 7 8 9 3 1 I 2 3 4 5 6 7 8 9 1 1 2 3 4 5 6 7 8 9 2 1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8 9 1 1 2 3 4 5 6 7 8 9 2 1 2 3 4 5
January February March 1983
FIG. 3. Total numbers of male (0 - 0 ) and unspawned female (0 - 0) toads in the pond each day.
February Morch
1983
FIG. 4. Daily minimum night air temperatures.
females who paired and spawned with male toads. The mean times for the two spawning periods were 3.1 ~ s . E . 0.19 and 2 . 3 9 k s . ~ . 0.09 days, respectively. The time for the first period was significantly longer than that for the second period ( t=3.366, d.f. =28, P<O.Ol).
Discussion The high degree of interspecific spawning between female Common toads and male
Common frogs resulting in the production of infertile eggs at a pond on Portland during March 1983 appears to be due to the combination of the following five factors:
( I ) Both species spawn in the same pond.

I00 C. J. READING
(2) Both species spawn at the same sites within the pond. (3) Both species spawn at the same time. (4) Mate choice by males of both species is relatively unselective. ( 5 ) A cold breeding season. The occurrence of both species in the study area is the result of a deliberate single
introduction of toads in 1968 and of frogs some years later. The absence of any other suitable spawning sites has led to the use of the same pond by both species.
The pond is small, isolated, man-made and, because it was designed to attract birds, i t has an almost uniformly shallow depth of 18-20 cm. This makes it ideal as a spawning site for frogs, which prefer water up to 15 cm deep, but less suitable for toads which prefer water between 18 and 46 cm deep (Smith, 1951; Frazer, 1966; Cooke, 1975; Strijbosch, 1979). As a result, the toads tend to select the same areas of the pond as the frogs in which to spawn.
Temporal separation of the species during the breeding season normally occurs because frogs are active at lower temperatures than toads and therefore usually spawn earlier (Cooke. 1977). Savage (1939, however, found that frogs which occurred at or near sea-level spawned later than those at altitude. This may explain the large amount of overlap found on Portland between the spawning periods of the two species which occur at approximately 20 m above sea- level.
During the breeding season male frogs and toads are relatively unselective in their choice of partner and in addition to unspawned females will attempt amplexus with other malcs, dead females and inanimate objects such as sticks (Smith, 195 1). Males ofboth species, when gripped by another male, emit a release call (Savage, 1934) which results in their release, whilst an unspawned female who is gripped by a male remains silent and is not released. Once a pair of either frogs or toads has completed spawning the female is released by the male. Although there is a high risk of interspecific pairing in a mixed breeding population of frogs and toads, spawning between male toads and female frogs was not observed on Portland. This appears to be due to the greater strength and better grip of male frogs enabling them to displace male toads from female frogs.
The lack of mate recognition in male frogs and male toads also occurs in female toads, in as much as they do not delay spawning when paired with a male frog. It is suggested that in the Common frog and Common toad, species recognition behaviour has not evolved because under normal conditions both species would be spatially and/or temporally separated and would therefore not expect to encounter the other at the spawning site.
The last and possibly the most important factor in producing a high degree of interspecific spawning was the cold breeding season of 1983. Between 2 and 1 1 March the minimum ground air temperature did not rise above 6"C, the estimated activity threshold temperature for toads on Portland (Reading & Clarke, 1983). It is, therefore, not surprising that, although male toads were present in the pond in large numbers, they were not able to compete for, or effectively defend female toads against male frogs. Although the female toads were very lethargic during this period they were, however, able to spawn, with the result that 65% of them did so whilst paired with male frogs.
During the second period of toad spawning activity (14-24 March) the mean minimum ground temperature did not fall below the 6°C activity threshold temperature and conse- quently all the male toads were active. The result was that only 9% of the female toads spawned with male frogs during this period. The hypothesis that low temperatures during the breeding season cause an increase in the occurrence of interspecific spawning between

INTERSPECIFIC SPAWNING BETWEEN FROGS AND TOADS 101
frogs and toads is based on one year’s data and therefore needs to be tested over a number of years in which the breeding season temperatures vary.
Lastly, the significant positive correlation between the appearance of frog spawn and the occurrence of interspecific spawning suggests that male frogs were reacting to an increase in the number of spawning female frogs by increasing their pairing activity. Although this reaction may be the result of an olfactory stimulus received from either the spawning pairs or their spawn more work is required before this can be confirmed.
In conclusion, it appears that the large amount of interspecific spawning between male frogs and female toads on Portland in March 1983 was primarily caused by the cold breeding season, given that spatial, temporal and behavioural separation of the species did not occur.
1 would like to thank M. Rogers for the use of his pond during the course of this study and J. P. Dempster, M. G. Morris and J. D. Goss-Custard for their critical examination of the manuscript .
REFERENCES Cooke, A. S. (1975). Spawn site selection and colony size of the frog (Rana femporuria) and the toad (Eufo bufo).
Cooke, A. S . (1977). Spawning dates of the frog (Rana remporaria) and the toad (Eujb bufo) in Britain. Er. J . Herpef.
Frazer, J. F. D. (1966). A breeding colony of toads (Bufo bufo (L.)) in Kent. Er. J. Herpet. 3: 236252. Reading, C. J. & Clarke, R. T. (1983). Male breeding behaviour and mate acquisition in the Common toad, Eufo
Savage, R. M. (1934). The breeding behaviour of the Common frog, Rana temporaria temporaria Linn. and of
Savage, R. M. (1935). The influence of external factors on the spawning date and migration of the Common frog
Smith, M. A. (1951). The British amphibians and reptiles. London: The New Naturalist Series, Collins. Strijbosch, H. (1979). Habitat selection of amphibians during their aquatic phase.. Oikos 33: 363-372.
J . Zool.. Lond. 175: 29-38.
5: 585-589.
bufb L. J . Zool., Lond. 201: 237-246.
the Common toad, Bujo bufi bufo Linn. Proc. zool. SOC. Lond. 1934: 55-70.
(Rana temporaria temporaria). Proc. 2001. Soc. Lond. 1935: 49-9 1.





























