Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 248, No. 7, Issue of April 10, pp. 2542-2548, 1973
Printed in U.S.A.
Intestinal Epithelial Cell Surface Membrane
Glycoprotein Synthesis
II. GLYCOSYLTRANSFERASES AND ENDOGENOUS ACCEPTORS OE‘ THE UNDIFFER.ENTIATED CELI, SURFACE MEMBRANE*
(Received for publication, October 2, 1972)
MILTON M. WEISER$
From the Department of &!edicine, Harvard Medical School and Medical Services ((:astr~o~r~testi??al CTllit),
Massachusetts General Hospital, Boston, Massachusetts 02114
SUMMARY
The cell surface membranes of mitotically active crypt cells from rat intestinal epithelium were found to have glycosyltransferase activities and endogenous acceptors not found on the differentiating villus cells. Rat intestinal epi- thelial cells were isolated as intact cells from different levels of the villus and crypt areas representing sequential stages of cellular differentiation. Incubation of these cells with UDP-N-acetyl[l-14C]glucosamine, UDP-[UJ4C]glucose, UDP- [l-3H]galactose, GDP-[UJ4C]mannose, and GDP-[U-Wlfucose demonstrated up to a lo-fold greater incorporation into crypt cells as compared to villus cells. In marked contrast, incubation with CMP-[4,5,6,7,8, 9-14C]sialic acid showed that radioactive sialic acid was preferably transferred to villus cell rather than to crypt cell surface. Effective trans- fer of sugars to exogenous acceptors gave further indication for the presence of galactosyltransferase enzyme activity on crypt cells rather than villus cells, and of sialyltransferase enzyme activity as being predominantly on villus cells. Neither glycosyltransferase activities nor endogenous ac- ceptors were released into the incubation medium. Analy- sis of labeled endogenous product by dialysis, acid precipita- tion, differential solvent extraction, graded acid hydrolysis, high voltage electrophoresis, and descending chromatog- raphy suggested that most of the label was present as the original monosaccharide derived from the nucleotide sugar, and that it had been incorporated into a membrane-asso- ciated glycoprotein. Incubation of human fetal intestinal epithelial cells with UDP-[l-3H]galactose also showed ga- lactosyltransferase enzyme activity to an exogenous as well as an endogenous acceptor. These results suggest that the plasma membrane of the mitotically active undifferentiated crypt cell and the fetal cell both contain (a) active glycosyl- transferase enzymes and (b) acceptor sites which are glyco- proteins with incomplete polysaccharide chains.
* This research was supported by grants from the National In- stitutes of Health (AM-03014) and the American Cancer Society (BC-93).
1 Recipient of the Massachusetts Cancer Society Scholar Award.
Evidence from tissue culture systems has suggested that cell surface membrane glycoproteins play a significant role in cell adhesion and contact inhibition and that alterations in these glycoproteins are related to the ability of a cell to enter mitosis
or to exhibit changes in cell behavior as seen after viral trans- formation (l-4). In addition, Roth ef al. (5) have demonstrated galactosyltransferase enzyme activity on cell surface membranes of embryonic chicken neural retina cells in tissue culture and they have suggested that these enzymes are involved in deter- mining cell behavior. The feto-embryonic cell membrane of neural retina cells from chicken embryo (6) and human fetal intestinal epithelial cells (7) could be further distinguished from the mature adult cell surface membrane by their differential agglutination with concanavalin A.
In the first paper of this series a gradient of cell surface mem- brane glycoprotein synthesis was demonstrated in rat intestinal epithelium. The gradient proceeded from very low rates in the undifferentiated crypt cell to constantly increasing rates for the differentiating villus cell. These data suggest that glycosylation of membrane-associated protein ma)- be an integral part of the process of cell differentiation (8).
The present study extends these observations by examining the ability of isolated rat and human fetal intestinal epithelial cells to transfer labeled monosaccharides from nucleotide sugars to endogenous acceptors present on the cell surface and to exog- enous acceptors. The surface membrane of the undifferentiated mitotically active crypt cell and of human fetal intestinal cells was characterized by active glgcosyltransferase activity ac- companied by the presence of appropriate endogenous acceptors. A significant exception was the sialyltransferase :endogenous acceptor system which appeared to be more a feature of the differentiating villus cell.
METHODS
Xaterials and General ~~~ethods-R:Ldioactive materials were purchased from Kew England Nuclrar and Amershan-Searle. Nonradioactive nucleotide sugars wcrc bought from Sigma. Protein was determined by the method of Lowry et al. (9). The nonfasting Sprague-Hawley female rats (175 to 225 g) obtained from Holtzman Company were killed, the small intestine was
2542
2543
removed, and isolated illtcstinal cell preparations were made as previously described (8). Purified brush borders were prepared by the method of Forstner et al. (10). Human fetuses were ob- tained by hysterotomy with their length varying from 7 to 12 cm (crown to rump). Fetal intestinal isolated cell preparations were prepared as previously described (7). For estimation of glycosyltransferasc activities the cells were washed and resus- pended in 0.1 M cacodylate-HCI buffer, pH 7.2, with 0.154 M NaCl to a concentration of approximately 2 X lo6 cells per ml.
@osyltransferase Enzyme Assays--To a 0.1 ml suspension of intact cells in 0.1 M cacodylate-HCl buffer, pH 7.2, containing 0.154 M NaCl was added 0.01 ml of 0.1 M MnCl2 and 0.01 ml of radioactive nuclcotidc sugars (see figures and tables for specific activities and concentrations). The cell suspension was then incubated at 37” for 45 or 90 min. Endogenous product was assayed in two ways. (a) The samples were precipitated by t,he addition of 2 ml of 57, cold trichloroacetic acid, filtered through glass fiber filters, 2.4 cm in diameter (Reeve Angel grade 934AH), and washed with 10 ml of cold 5% trichloroacetic acid followed by 10 ml of absolute ethanol. The filters were then placed in counting vials, dried in air, covered with 10 ml of a toluene-based scintillator (4 g of 2, %diphenylosazole, 50 mg of 1,4-bis[2-(5.phenylosazoyl)]benzene, per liter of toluene made wit’h LiquiHnor from New England Nuclear), and counted in a liquid scintillation counter. (b) The reaction was stopped by placing the reaction mixture in ice and adding 0.08 ml of 2% sodium tetraborate. Aliquots of the total mixture were then placed on Whatman No. 3MM paper and subjected to high voltage paper electrophoresis, 2000 volts, 30 to 60 min, through 1’s sodium tetraboratc. Radioactive areas were located by a Packard radiochromatogram scanner. Under these conditions the endogenous product remained at the origin and the nucleo- tide sugars moved anodally a significant distance, as did most of the non-nucleotidc sugars. Appropriate areas were then cut out and counted in a liquid scintillation counter as above. When esogenous acceptors for galactosylt~ransferase activity (S-acetyl- glucosamine) and for the sialyltransferase activity (lactose) were used their labclcd products, lactosamine and neuromine lactose, migrated naar the origin but still separated from the endogenous product at the origin (see Fig. 2). In the assay mixture either 0.05 ml of 0.5 M 12’.acetylglucosamine (0.14 M
final concentration) or 0.01 ml of 0.25 M lactose (0.02 M final concentration) was added when transferase activities to exog- enous acceptors were evaluated. All values were calculated by comparison with appropriate zero time controls. On occasion, cells were separated from incubation medium by centrifugation and cells and medium evaluated separately for product.
Products Idenlificafion-For product identification larger assay volumes were used which consisted of 0.8 ml of cells suspended in the cacodylate-HCI-NaCl buffer, 0.1 ml of 0.1 M MnClz and 0.1 ml of the nucleotide sugar. The reaction mixture was treated as above. The endogenous product always accompanied the cells after centrifugation and these were homogenized and di- alyzed overnight against 0.1 M KC1 and then against Hz0 for 2 days. After dialysis the endogenous product was recovered completely as a precipitate, resuspended in minimum volume of Hz0 and aliquots subjected to acid hydrolysis. The products of the sialyltransferase activity were hydrolyzed with 0.1 s acetic acid, IOO”, 60 min under vacuum. It was then evaporated to dryness with repeated water washes using a Buchler rotary evaporator to remove the acetic acid. The hydrolytic products were then separated by high voltage paper electrophoresis (1 o/o sodium tetraboratc, 2000 volts, 30 min) and by descending paper
(Whatman No. 1) chromatography with (a) butanol-acetic acid-Hz0 (3:2:1,28 hours), and (b) butanol-pyridine-Hz0 (6:4:3, 40 hours). The products of the galactosyltransferase and N-ac- tylglucosaminyltransferase activities were hydrolyzed with 1.0 N
HCl, 100” for 18 hours under vacuum and the mixture deionized with 0.2 M dioctylmethylamine in chloroform. These hydrolytic products were also separated by sodium tetraborate high voltage electrophoresis as well as by descending paper chromatography with (c) pyridine-ethylacetate-HzO-acetic acid (5: 5 :3 : I,24 hours) and cd) butanol-ethanol-Hz0 (10 : 1: 2, 5 days).
RESULTS
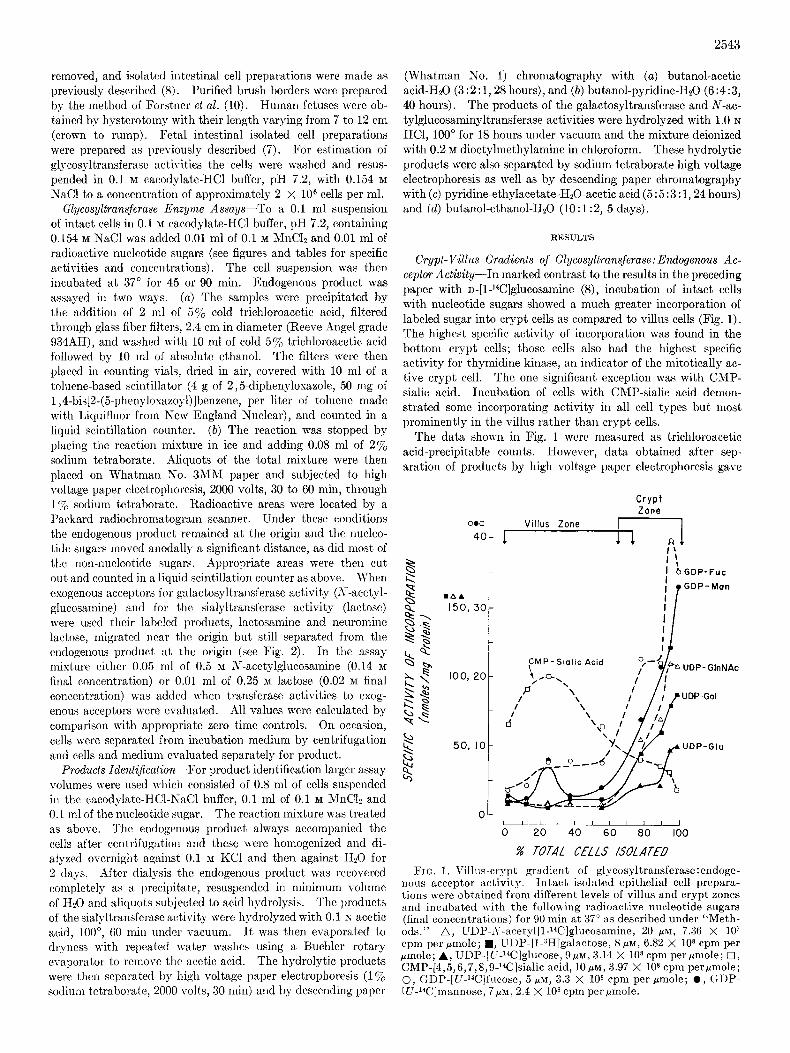
Crypt-Villus Gradients of Glycosyltransjerase:Endogenous Ac- ceptor Activity-In marked contrast to the results in the preceding paper with D-[1-14C]glucosamine (8), incubation of intact cells with nucleotide sugars showed a much greater incorporation of labeled sugar into crypt cells as compared to villus cells (Fig. 1). The highest specific activity of incorporation was found in the bottom crypt cells; those cells also had the highest specific activity for thymidine kinase, an indicator of the mitotically ac- tive crypt cell. The one significant exception was with CMP- sialic acid. Incubation of cells with CMP-sialic acid demon- strated some incorporating activity in all cell types but most prominently in the villus rather than crypt cells.
The data shown in Fig. 1 wcrc measured as trichloroacetic acid-precipitable counts. However, data obtained after sep- aration of products by high voltage paper electrophoresis gave
Crypt Zone
0.0 Villus Zone
4or -T
50, IO-
i ’ I bGDP-Fuc ir GDP- Man
FIG. 1. Villas-crypt gradient of glycosyltransferase:endoge- nous acceptor activity. Intact isolated epithelial cell prepara- tions were obtained from different levels of villus and crypt zones and incubated with the follovc ing radioactive nucleotide sugars (final concentrations) for 90 min ai 37” as described under “Meth- ads.” A, UDP-,Y-acetyl[l-Wlglucosamine, 20 PM, 7.36 X 10’ cpm per pmole; w, UDP-[l-3H]galactose, 8 pM, 6.82 X lo* cpm per pmole; A, UDP-[C-14C]glncose, (3p~, 3.14 X lOa cpm per @mole; 0, CMP-[4,5,6,7,8,9-W]sialic acid, lops, 3.97 X lo* cpm perpmole; 0, GDP-[U-Wlfncose, 5 PM, 3.3 X IO* cpm per rmole; 0, (+DP- [LI-Wlmannose, ‘7~~1,2.4 X lo* cpm per pmole.
2544
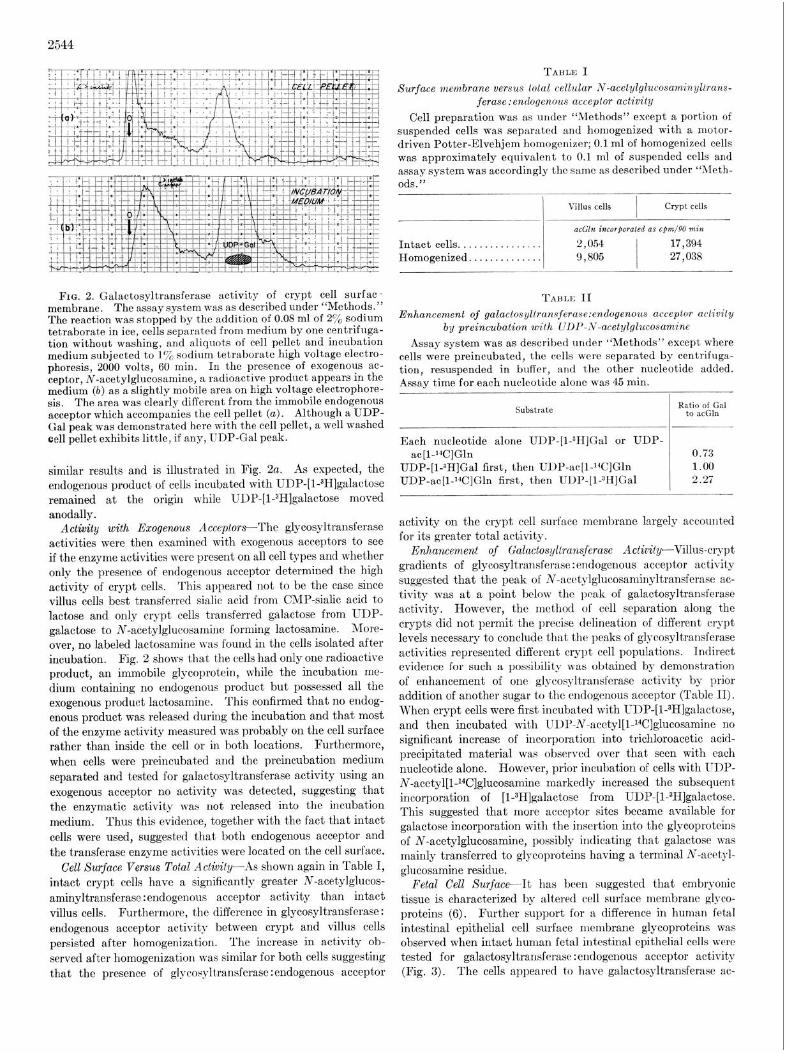
FIG. 2. Galactosyltransferase activity of crypt cell surfac- membrane. The assay system was as described under “Methods.” The reaction was stopped by the addition of 0.08 ml of 2% sodium tetraborate in ice, cells separated from medium by one centrifuga- tion without washing, and aliquots of cell pellet and incubation medium subjected to 1% sodium tetraborate high voltage electro- phoresis, 2000 volts, 60 min. In the presence of exogenous ac- ceptor, N-acetylglucosamine, a radioactive product appears in the medium (b) as a slightly mobile area on high voltage electrophore- sis. The area was clearly diffcrcnt from the immobile endogenous acceptor which accompanies the cell pellet (a). Although a UDP- Gal peak was demonstrated here with the cell pellet, a well washed cell pellet exhibits little, if any, UDP-Gal peak.
similar results and is illustrated in Fig. 2~. As expected, the endogenous product of cells incubated with UDP-[l-3H]galactose remained at the origin while UDP-[l-3H]galactose moved anodally.
Activity with Exogenous Acceplors-The glycosyltransferase activities were then examined with exogenous acceptors to see if the enzyme activities were present on all cell types and whether only the presence of endogenous acceptor determined the high activity of crypt cells. This appeared not t’o be the case since villus cells best transferred sialic acid from CMP-sialic acid to lactose and only crypt cells transferred galactose from UDP- galactose to N-acetylglucosamine forming lactosamine. More- over, no labeled lactosamine was found in the cells isolated after incubation. Fig. 2 shows that the cells had only one radioactive product, an immobile glycoprotein, while the incubation me- dium containing no endogenous product but possessed all the exogenous product lactosamine. This confirmed that no endog- enous product was released during the incubation and that most of the enzyme activity measured was probably on the cell surface rather than inside the cell or in both locations. Furthermore, when cells were preincubated and the preincubation medium separated and tested for galactosyltransferase activity using an exogenous acceptor no activity was detected, suggesting that the enzymatic activity was not released into the incubation medium. Thus this evidence, together with the fact that intact cells were used, suggested that both endogenous acceptor and the transferase enzyme activities were located on the cell surface.
Cell Surface Versus Total Activity--As shown again in Table I, intact crypt cells have a significantly greater N-acetylglucos- aminyltransferase :endogenous acceptor activity than intact villus cells. Furthermore, the difference in glycosyltransferase: endogenous acceptor activity between crypt and villus cells persisted after homogenization. The increase in activity ob- served after homogenization was similar for both cells suggesting that the presence of glycosyltransferase:endogenous acceptor
TABLE I Surface membrane versus Lola1 cellular iv-acetylglucosaminl/ltrans-
ferase:endogenous acceptor activity Cell preparation was as under “Methods” except a portion of
suspended cells was separated and homogenized with a motor- driven Potter-Elvehjem homogenizer; 0.1 ml of homogenized cells was approximately equivalent to 0.1 ml of suspended cells and assay system was accordingly the same as described under “Meth- ods.”
Villus cells I
crypt cells
acGln incorfmrated as cfm/OO min
Intact cells.. . . . . . . . . . . . . . 2,054 17,394 Homogenized. . . . . . . . . . . 9,805
I 27,038
Tnnrz II Enhancement of galactosyliransjerase:endogenous acceptor activity
by preincubation with UDP-N-acetylglucosamine Assay system was as described under “Methods” except where
cells were preincubated, the cells were separated by centrifuga- tion, resuspended in buffer, and the other nucleotide added. Assay time for each nucleotide alone was 45 min.
Substrate R~tpd&%l
Each nucleotide alone UDP-[1-311]Gal or UDP- ac[l-%]Gln 0.73
UDP-[I-3HJGal first, then UDP-ac[l-‘4C]Gln 1.00 UDP-ac[l-“C]Gln first, then UDP-[1-3H]Gal 2.27
activity on the crypt cell surface membrane largely accounted for its greater total activity.
Enhancement of Galuctosyllransjerase Activity-Villus-crypt gradients of glycosyltransferase : endogenous acceptor activity suggested that the peak of N-acetylglucosaminyltransferasc ac- tivity was at a point below the peak of galactosyltransferase activity. However, the m&hod of cell separation along the crypts did not permit the precise delineation of different crypt levels necessary to conclude that the peaks of glycosyltransferase activities represented different crypt cell populations. Indirect evidence for such a possibility was obtained by demonstration of enhancement of one glycosyltransferase activity by prior addition of another sugar to the cndogenous acceptor (Table II). When crypt cells were first incubated with UDP-[l-3H]galactose, and then incubated with IJDP-N-acetyl[l-14C]glucosamine no significant increase of incorporation into trichloroacetic acid- precipitated material was observed over that seen with each nucleotide alone. However, prior incubation of cells with UDP- N-acetyl[l-l%]glucosamine markedly increased the subsequent incorporation of [l-3H]galactose from UDP-[l-3H]galactose. This suggested that more acceptor sites became available for galactose incorporation with the insertion into the glycoproteins of N-acetylglucosamine, possibly indicating that galactose was mainly transferred to glycoproteins having a terminal N-acetyl- glucosamine residue.
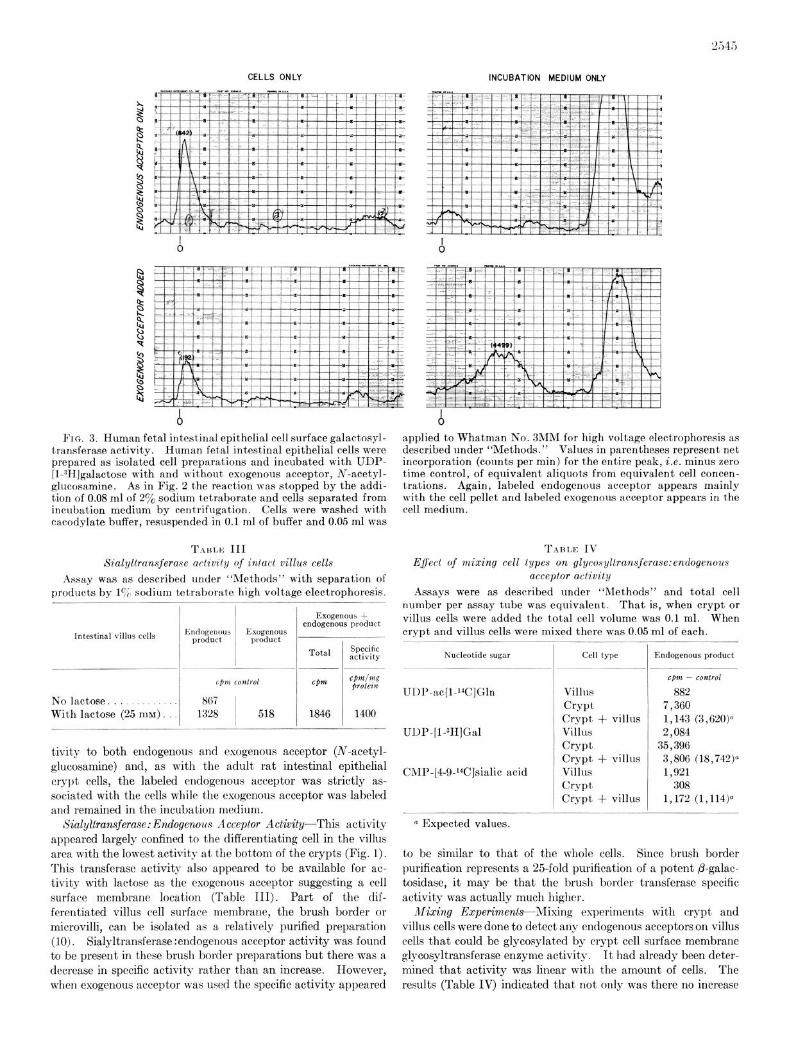
Fetal Cell Surface-It has been suggested that embryonic tissue is characterized by altered cell surface membrane glyco- proteins (6). Further support for a difference in human fetal intestinal epithelial cell surface membrane glycoproteins was observed when intact human fetal intestinal epithelial cells were tested for galactosyltransferasc :endogenous acceptor activity (Fig. 3). The cells appeared to have galactosyltransferase ac-
CELLS ONLY INCUBATION MEDIUM ONLY
Fro. 3. Human fetal intest,inal epithelial cell surface galactosyl- transferase activity. Human fetal intestinal epithelial cells were prepared as isolated cell preparations and incubated with UDP- [I-3HJgalactose with and without exogenous acceptor, Ar-acetyl- glucosamine. As in Fig. 2 the reaction was stopped by the addi- tion of 0.08 ml of 2% sodium tetraborate and cells separated from incubation medium by centrifugation. Cells were washed with cacodylate buffer, resuspended in 0.1 ml of buffer and 0.05 ml was
Tnn11s III Sialyllransferase aciiviiy of inlact villus cells
Assay was as described under “Methods” with separation of products by lo/;> sodium tetraborate high voltage electrophoresis.
Intestinal villus cells
No lactose. With lactose (25 mM) -
Endogenous 1 Exogenous product / product
-
Exogenous + endogenous product
Total Specific activity
1846 1400
tivity to both endogenous and exogenous acceptor (N-acetyl- glucosamine) and, as with the adult rat intestinal epithelial crypt cells, the labeled endogenous acceptor was strictly as- sociated with the cells while the exogenous acceptor was labeled and remained in the incubation medium.
Sialyltransferase: Endogerwus Acceptor Activity-This activity appeared largely confined to the differentiating cell in the villus area with the lowest activity at the bottom of the crypts (Fig. 1). This transferase activity also appeared to be available for ac- tivity with lactose as the exogenous acceptor suggesting a cell surface membrane location (Table III). Part of the dif- ferentiated villus cell surface membrane, the brush border or microvilli, can be isolated as a relatively purified preparation (10). Sialyltransferase :endogenous acceptor activity was found to be present in these brush border preparations but there was a decrease in specific activity rather than an increase. However, when exogenous acceptor was used the specific activity appeared
applied to Whatman No. 3MM for high voltage electrophoresis as described under “Methods.” Values in parentheses represent net incorporation (counts per min) for the entire peak, i.e. minus zero time control, of equivalent aliquots from equivalent cell concen- trations. Again, labeled endogenous acceptor appears mainly with the cell pellet and labeled exogenous acceptor appears in the cell medium.
Effecl of mixing cell types on glyconyllransferase:endogenous TABI,E IV
acceptor activity
Assays were as described under ‘(Methods” and total cell number per assay tube was equivalent. That is, when crypt or villus cells were added the total cell volume was 0.1 ml. When crypt and villus cells were mixed there was 0.05 ml of each.
Nucleotide sugar Cell type Endogenous product
Villlis Crypt Crypt + villus Villus Crypt Crypt + villus Villus Crypt Crypt + villus
cpm - control
882 7,360 1,143 (3,620)0 2,084
35,396 3,806 (18,742)c 1,921
308 1,172 (1,114)0
CMP-[4-9.‘4C]sialic acid
o Expected values.
to be similar to that of the whole cells. Since brush border purification represents a 25-fold purification of a potent @-galac- tosidase, it may be that the brush border transferase specific activity was actually much higher.
&fizing Ezrperimenla-Mixing experiments with crypt and villus cells were done to detect any endogenous acceptors on villus cells that could be glycosylated by crypt ccl1 surface mcmbranc glycosyltransferase enzyme activity. It had already been deter- mined that activity was linear with the amount of cells. The results (Table IV) indicated that not only was there no increase
2546
(b) (b
(a)
(b)
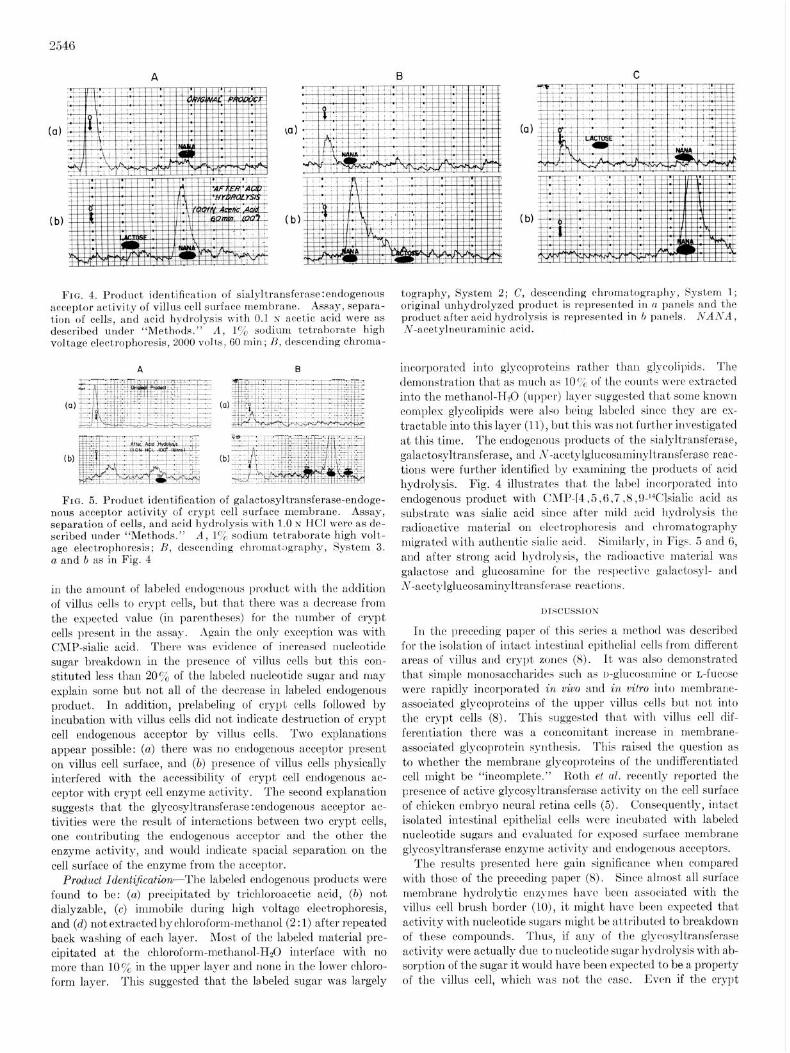
FIG. 4. Product identification of sialyltransfernse:endogenous acceptor achtivity of villus cell surface membrane. Assay, separa- tion of cells, and acid hydrolysis v\ith 0.1 N acetic acid were as described under “Methods.” A, 170 sodium tetraborate high voltage electrophoresis, 2000 volts, 60 min; H, descending chroma-
(b)
FIG. 5. Product identification of galactosyltransferase-endoge- nous acceptor activity of crypt cell surface membrane. Assay, separation of cells, and acid hydrolysis with 1.0 N KC1 were as de- scribed nnder “Methods.” A, 17p sodium tetraborate high volt- age electrophoresis; H, descending chrornat,,graphy, System 3. a and b as in Fig. 4
in the amount of labeled endogenous product nith the addition of villus cells to crypt cells, but that there was a decrease from the expected value (in parentheses) for the number of crypt cells present in the assay. Again the only exception was with CMP-sialic acid. There was evidence of increased nucleotide
sugar breakdown in the presence of villus cells but this con- stituted less than 20% of the labeled uucleotide sugar and may explain some but not all of the decrease in labeled endogenous product. In addition, prelabeling of crypt cells followed by incubation with villus cells did not indicate destruction of crypt cell endogenous acceptor by villus cells. Two explanations appear possible: (a) there was no endogenous acceptor present on villus cell surface, and (b) presence of villus cells physically interfered with the accessibility of crypt cell cndogenous ac- ceptor with crypt cell enzyme activity. The second explanation suggests that the glycosyltransferase :endogrnous acceptor ac- tivities were the result of interactions between two crypt cells, one contributing the endogenous acceptor and the other the enzyme activity, and would indicate spatial separation on the cell surface of the enzyme from the acceptor.
Product Ident&ation--The labeled endogenous products were found to be: (a) precipitated by trichloroacetic acid, (a) not dialyzable, (c) immobile during high voltage elcctrophoresis, and (d) not extracted by chloroform-methauol (2 : 1) after repeated back washing of each layer. Most of the labeled material pre- cipitated at the chloroform-methanol-EL0 interface with no more than 10% in the upper layer and none in the lower chloro- form layer. This suggested that the labeled sugar was largely
tography, System 2; C, descending chromatography, System 1; original unhydrolyzed product is represented in n panels and the product after acid hydrolysis is reproser~tetf in 7, panels. NANA, N-acet.ylnenraminic acid.
incorporated into glycoproteins rather thau glycoliyids. The demonstration that as much as 10c& of the couuts were extracted into the methanol-H20 (upper) layer suggested that some known complex glycolipids were also being labeled since they are cs- tractable into this layer (II), but this was not furt’hcr investigated at this time. The endogenous products of the sialyltransferase, galactospltransferase, and S-acctylglucosamiilyltJransferase reac- tions were further ident,ified by examining the products of acid hydrolysis. Fig. 4 illustrates that t,he label incorporated into endogenous product with CAlI-‘- .5,6,7,8 ,OJ‘Clsialic acid as substrate was sialic acid since aft’er mild acid hydrolysis t’he radioactive material on electrophorc~sis aud chromatography migrated with authentic sialic acid. Similnrly, in Figs. 5 and 6, and after st,rong acid hydrolysis, the radioactive material was galactose and glucosamine for the rrspectivc galactosyl- and N-acct~ylglucosaminyltransferasc reactions.
In t,he preceding paper of t’his series a mrthotl was described for the isolation of intact intestinal epithelial cells from different areas of villus and crypt zones (8). It was also demonstrated that simple monosaccharides such as n-glucosamine or L-fucose were rapidly incorporated in viva and in vi/ro into membrane- associated glycoproteins of the upper villus cells but not into the crypt cells (8). This suggeskd that with villus cell dif-
ferentiation there was a concomitant increase in membrane- associated glycoprotein synthesis. This raised t’he question as to whether the membrane glycoproteins of the undifferent’iated cell might be “incomplete.” Roth et al. recently reported the
presence of active glycosyltra,nsferase activity on the cell surface of chicken embryo neural ret’ina cells (5). Consequently, intact isolated intestinal epithelial cells were incubat,ed with labeled nucleotide sugars and evaluated for exposed surface membrane glycosyltransferase enzyme activity and endogrnous acceptors.
The results presented here gain significance when compared with those of the preceding paper (8). Since almost all surface membrane hydrolytic enzymes have been associated lrith the villus cell brush border (lo), it might havr been expected that activity wit’h nucleotide sugars might be attributed to breakdown of t’hese compounds. Thus, if any of t,he 81y(:osyltransf~rase
activity were actually due to nucleotide sugar hydrolysis with ab- sorption of t’he sugar it would have been expected to be a property of the villus cell, which was not thr case. Ii:vcn if the crypt

‘Fro. 6. Product, identification of A‘-acetylglucosnminyltransferase-endogenoLls acceptor activity of crypt cell surface membrane. Assay, separation of cells, and acid hydrolysis with 1.0 N HCl were as described under “Methods”. A, I:/, sodium tetraborate high voltage electrophoresis; f1, descending chromatography, System 3; C, descending chromatography, System 4. a and 0 as in Fig. 4.
cell was capable of hyclrolyzing the nucleotide sugars it would not hare been capable of incorporating the free sugar into glyco- proteins (8). Furthermore, as shown in Table I the intracellular glycosyltransferase : endogenous acceptor activities for both villus and crypt cells appeared to be similar and the marked increased activity of intact crypt cells over intact villus cells probably represented surface membrane activity. The per- &tent although low activity seen with intact villus cells (Fig. 1 and Table T) could represent a small degree of leakiness of the cells for the nucleotidr sugars. It could also represent activity present on the villus lateral or basal membrane, or the presence of crypt cells in villus fractions. Villus cell fractions were not totally free of crypt ~11s as was evident by low levels of thymi- dine kinase activity and low [Me-311]thymidine incorporation during early labeling periods (8). Quantitatively these crypt cell markers indicate that 5 to 10% of the crypt cell activities wwc prcscnt in villus cell fractions. The glycosyltransferase: cndogcnous ac*cclptor activity found in villus cell fractions was also 5 to 10% of the cryl)t cell fraction activity. In addition, it has been demonstrated that concanavalin A derivatized nylon fibers could select from Tillus cell fractions cells which appeared t,o bc crypt cells by thymidine incorporation studies as well as by possessing high glycosyltransfcrasc:cndogenous acceptor ac- tivity.’ This selection of crypt cells by concanavalin A dcriva- tized nylon fibers caannot give a quantitative estimate of the number of crypt cells in villus cell fractions.
Nevertheless, the data has indicated that it was mainly the intact crypt cells that wcrc capable of transferring sugars from a nucleotide precursor to both endogenous and exogenous accep- tors. It, would appear then that the most likely explanation is that the crypt cell surface membrane has (a) exposed and active glycosyltransferase enzymes, and (b) that this membrane also has an cndogenous acceptor which must be incomplete if capable of accepting a sugar.
Tn addition to evidence that surface membrane glycoproteins are incompletely glycosylatcd in normal cells during mitosis and in virally transformed cells (3), cvidencr has been presented by Moscona showing that embryonic tissue is also characterized by altered surface membrane glycoprotcins (6) as detected by a marked agglutination of these cells by concanavalin A. This has also been found to be the case with human fetal intestinal cells (‘7). The prrsent results further suggest the presence of incompletely glycosylated membrane glycoprotcin and exposed glycosyltransferases on the fetal intestinal cell surface membrane.
* U. Podolsky and AI. ;\I. Weiser, unpublished experiments.
It could be argued that what was demonstrated in these cx- periments was the isolation of cells a,ctively synthesizing new cell membrane iu an early phase of differentiation and not, neces- sarily, a prerequisite cell membrane alteration for mitosis. Gnder these circumstances one might expect that there would be some order by which specific sugars were added. The data in Table II showing enhancement of galactosyltransferase ac- tivity by preincubation with TJDP-N-acctylglucosamine suggest that the order was N-acetylglucosamine followed by galactosc. Thus, either during or after mitosis, the cell surface membrane glycoproteins of the mitotically active crypt cells were charac- teristically incomplete and cont’aincd active glycosyltransferase enzymes. In contrast, the mixing experiments (Table IV) sug- gested that the villus cell surface membrane glycoproteins were either complct’cd before insertion into the membrane or inac- cessible since these villus cells had not demonstrated available endogcnous acceptor activit,y when enzyme activity was present (as the crypt cell surface mcmbranct).
One important rsception was t.hc Galyltransfcrase activity which n-as highest with villus cells and lowest with crypt cells (Fig. I). Since sialic acid terminates chain growth when joined to a N-acctylgalactosarnine residue (4) but is also the sugar with the most labile linkage (4), it might be argued t.hat the presence of sialylt,ransfsrase activity on the surface membrane is for con- stant repair. The presence of sialic acid on membrane glyco- proteins has been given greater functional significance than that of other sugars (12-17). Sialic acid is one of the factors which appears to give to t,he mcmbrant: its net negative electrical charge (18). The presence of increased or decreased sialic acid residues on t,umor aud transformed cell membranes has been debated for a number of years but the most recent studies (19, 20) have shown a decreased membrane sialic acid content after viral transformation. No direct evidence has been presented for any sialic acid-mediat,ed control of cell turnover.
Cell surface membrane glycoproteins are believed to be in- complete during mitosis, in ea,rly sparse growth of tissue culture cells, and after viral transformation (l-3). The properties of decreased cell adhesion and contact inhibition under these con- ditions of cell growth or transformation may be due to these alterations in membrane glycoprotein. In fact, Roseman (4) has proposed that surface membrane glycosyltransferases inter- act with incomplete glycoproteins of another cell membrane and thereby produce specific cell adhesive forces. The results de- scribed in the present report arc compatible with espcriments in tissue culture systems (l-3) and with chicken embryo cells (5). However, there is no evidence that intestinal epithelial crypt

cells or fetal intestinal cells are less adhesive to adjacent cells or to the basement membrane. In fact, it was easier to obtain isolated epithelial cell preparat’ions of single cells with villus cells than with crypt cells which remained in clumps. This was not true for fetal cells which were easily prepared as isolated cells. Thus, at least for the intestinal epithelial system, there was no consistent relationship between cell adhesive forces and the incompleteness of surface membrane glycoproteins.
It has been suggested that the cell surface membrane is ac- tually an outgrowth from the Golgi membrane and, therefore, would have glycosyltransferases imbedded in its unit membrane structure (21, 22). As long as the cell is relatively primitive and rapidly dividing, requiring rapid synthesis of new surface membranes, it will have a surface membrane which retains the characteristics of its source, the Golgi membrane. It would then follow that one explanation for finding glycosyltransferase ac- tivity on crypt and fetal (sell surface membranes may be that the “early” cell surface membrane is, in fact, externalized Golgi membrane. Regardless of the origin, the presence of these en- zymes and of incompletely glycosylated glycoproteins on the cell surface membrane appears to be a consistent and valuable identifying feature of the undifferentiated cell state.
2. BURGER. M. M. (1969) Proc. Nat. Acad. Sci. U. S. A. 62, 994- 1001 ’ ~ ’
3. Fox, T. O., SHEPPARD, J. R., AND BURGKR, M. M. (1971) Proc. Nat. Acad. Sci. U. S. A. 68. 244-247
4. ROSEM~N, S. (1970) Chewa. Ph&. I,ipids 6, 270-297 5. ROTH, S., MCGUIRF:, E. J., I\ND I<osc:M.\N, S. (1971) J. Cell
Biol. 61, 536-547 6. MOSCON~, A. A. (1971) Science 171, 905 7. WEISER, M. M. (1972) Science 177, 525-526 8. WEISER, M. M. (1973) J. Hiol. ChewL. 248, 2536-2541 9. LOWRY, 0. H., ROSEBROUGII, N. J., Fauu, A. L., AND RAND.~LL,
R. J. (1951) J. Biol. Chem. 193, 265-275 10. FORSTNER, G. G., SABI~SIN, S. M., .\ND ISSI?I,BACHER, K. J.
(1968) Biochem. J. 106, 381-390 11. SIDDIQUI, B., AND H.\Ic,\MoI~I, S. (1970) Cancer Res. 30, 2930-
2936 12. WARREN, L., FUHRER, J. P., AND BUCK, C. A. (1972) Proc. Xat.
Acad. Sci. U. S. A. 69, 18381842 13. COOK, G. M. W., HKARD, D. I~., AND SI’:.\M.IN, G. V. F. (1960)
Nature 188, 1011-1012 14. LIINGLEY, 0. K., .\ND AMHROSIC, E. J. (1964) Xature 204, 53-54 15. GLICK, J. L., GOLD~XRG, A. I~., I\~~ P.\RDXE, A. B. (1966)
Cancer Res. 26, 1774-1777 16. MESTER, L., SZADADOS, L., BORN, G. V. R., AND MICH.\I+ F.
(1972) Kature New Biol. 236, 213-214 17. PRICER, W. IX., JR., .\ND ASHWISLL, G. (1971) J. Biol. Chews.
246, 4825-4833
Acknowledgments-I thauk Miss Ireue Kozdrowski for valuable 18. COOK, G. M. W. (1968) Biol. Rev. 43,3633391
technical assistance. I especially thank Dr. Kurt J. Isselbacher 19. Wu, H. C., MEESAN, E., BLACK, P. H., AND ROBBINS, P. W.
(1969) Biochemistry 8, 250992517 for his support in this work, as well as his critical review of this 20. GRIMES, W. J. (1970) Biochemistry 9, 5083-5092 manuscript. 21. WAGNER, R. R., AND CYNKIN, M. A. (1969) Arch. Biochewl.
REFERENCES Biophys. 129, 242-247
22. SCHACHTER, H., JANBAL, I., HUDGIN, R.. L., PINTERIC, L., 1. OPPGNHKIMGR, S. B., EDIDEN, M., ORR, C. W., AND ROSEMAN, MCGUIRE, E. J., AND RORICM~ZN, S. (1970) J. Biol. Chem. 246,
S. (1969) Proc. Nat. Acad. Sci. U. S. A. 63, 1395-1402 1090-1100





























