Embed Size (px)
Citation preview

J. Phytopathology 131, 184—192 (1991)© 1991 Paul Parey Scientific Publishers, Berlin and HamburgISSN 0931-1785
Department of Plant and Forest Protection,Swedish University of Agricultural Sdences
Intra- and Interspecific Variation in Plant Responseto Inoculation with Deleterious Rhizosphere Pseudomonads
BOEL AsTROM
Author's address: Department of Plant and Forest Protection, Swedish University of AgriculturalSciences, Box 7044, S-750 07 Uppsala, Sweden
Received May 2, 1990; accepted June 27, 1990
Abstract
Accessions of wheat, spinach, lettuce and different Brassica species were tested in greenhouseexperiments for reaction to inoculation with two isolates of growth-inhibitory rhizosphere bacteria.Seedlings grown in non-sterile soil were inoculated with bacterial suspension and shoot dry weightwas measured after five weeks. Large differences were found between the plant species tested in theiraverage sensitivity to each bacterial isolate, and in the majority of plant species, significant differenceswere also found between accessions in the response to one or both isolates. These findings suggestthat, in addition to the variation between plant species, intraspecific variation in the reaction todeleterious bacteria is a common feature in plants. This supports the hypothesis that plant reaction torhizosphere bacteria is under genetic control. The results funher indicate specificity in the inter-actions between plants and bacterial isolates, both at the plant species level and at the accession level.
Zusammenfassung
Intra- und interspezifische Variation der Pflanzenreaktiongegenuber einer Inokulation mit schadlichen Rhizospharen Pseudomonaden
In Gewachshausversuchen wurde die Reaktion verschiedener Herkiinfte von Weizen, Spinat,Salat und unterschiedlichen Brassica spp. nach einer Inokulation mit zwei wachstumshemmendenRhizospharebakterien-Isolaten untersucht. Samlinge, die in nicht steriler Erde wuchsen, wurden miteiner Bakteriensuspension inokuliert und das Sprofitrockengewicht wurde nach weiteren fiinf Wochenermittelt. Gro6e Unterschiede wurden in der durchschnittlichen Empfindlichkeit der getestetenPflanzenspezies gegen beide bakterielle Isolate festgestellt, und bei den meisten Pflanzenspezieswurden signifikante Unterschiede auch zwischen den Linien in ihrer Reaktion auf einen oder beidenIsolaten festgestellt. Diese Ergebnisse deuten daraufhin, dafi zusatzlich zu Variationen zwischen denPflanzenspezies, eine intraspezifische Variation in der Reaktion auf schadliche Bakterien eine haufigvorkommende Eigenschaft bei Pflanzen ist. Diese Beobachtung unterstiitzt die Hypothese, dafi diePflanzenreaklion auf Rhizosphare Bakterien unter genetischer Kontrolle ist. Die Ergebnisse zeigenweiterhin, dafi es eine SpezHizitat der Interaktionen zwischen PHanzen und Bakterienisolaten, sowohlauf Pflatizenspezi(»ebene als auch auf Ebene der Herkiinfte gibt.
U.S. Copyri t Clearance Cenwr Code Sutement: 0931-1785/91/3103-0184$02.50/0

Intra- and Interspecific Variation in Plant Response 185
The surface and cortex of the plant root and the zone of soil influenced byroot excretions are colonized by a variety of micro-organisms. This rhizospheremicroflora includes groups of organisms that are widely recognized as having amore or less major impact on nutrient uptake and/or plant growth, e.g. mycorrhi-zal fungi, nitrogen-fixing bacteria and root-infecting pathogenic fungi (KATZNEL-SON 1965, CURL and TRUELOVE 1986), but also less well-known, saprophyticorganisms that are potentially beneficial or detrimental to plants (SuSLOW 1982,BURR and CAESAR 1984). The expression "deleterious rhizobacteria" has beencoined for such root-associated bacteria that retard plant growth, with or withoutcausing symptoms on roots and/or shoots (SusLOW and SCHROTH 1982, FREDRICK-SON and ELLIOTT 1985 a, GERHARDSON et al. 1985, SCHIPPERS et al. 1987). Deleteri-ous bacteria may be classified as 'exopathogens', i.e. non-parasitic pathogens thataffect the plant through the release of harmful substances (WoLTZ 1978).Although their role in plant ecosystems and crop productivity is at present notfully understood, recent research indicates that deleterious bacteria, at least incertain situations, may be of economic importance (GARDNER et al. 1984,FREDRICKSON and ELLIOTT 1985 a, CAMPBELL et al. 1986, SCHIPPERS et al. 1987,TuRCO et al. 1990). NEAL et al. (1970, 1973) and others have demonstrated thatthe plant's genetic constitution may have a considerable influence on specificmicrobial groups in the rhizosphere. However, the extent of influence of theplant genotype on interactions between plants and deleterious bacteria has notbeen elucidated.
In a previous study, significant genetic variation was recorded in wheat{Triticum aestivum L.) and in pea {Pisum spp.) in the reaction to root inoculationwith different deleterious bacteria (ASTROM and GERHARDSON 1988). The aim ofthe present study was to confirm these results using a wheat material of greaterdiversity than the one previously tested, and to investigate whether intraspecificvariation in reaction to deleterious bacteria is of more common occurrence inplants. For this purpose, accessions of wheat, spinach {Spinada oleracea L.),lettuce {Lactuca sativa L.) and Brassica spp. were inoculated with two plantgrowth-inhibiting Pseudomonas isolates, and shoot weight, and in one experi-ment also root weight, was recorded.
Materials and Methods
Plant material
Seeds from 20 accessions of spring wheat originating from different parts of the world wereobtained from the Zentralinstitut fiir Genetik und Kulturpflanzenforschung, Gatersleben, DDR;Vavilov Institute of Plant Husbandry, Leningrad, USSR; Germplasm Institute, Bari, Italy, andUSDA National Small Grain Collection, Beltsviile, USA. Accessions of Brassica spp. were suppliedby Svalof AB, Svalov; Hammenhogs Fro AB, Hammenhog; Weibullsholm Plant Breeding Institute,Landskrona; and by The Department of Plant Breeding, Uppsala; all in Sweden. Cultivars andbreeding lines of spinach were obtained from Nordreco AB, Bjuv, Sweden, and from WeibullsholmPlant Breeding Institute, and lettuce cultivars were supplied by Hammenkogs Fro AB, Hammenfaog,Sweden.
186 ASTROM
Bacterial isolates and inoculum preparation
Two deleterious bacterial isolates, A 112 {Pseudomonas fluorescens [Trevisan] Migula) and A313 {Pseudomonas sp., non-fluorescent), originally isolated from plant roots (ASTRGM and GERHARD-SON 1988) were used. Inoculum was prepared by harvesting cells from two-day-old agar plate cultures(King's Medium B; KING et al 1954) and suspending them in Standard nutrient broth (SNB;ALSTROM 1987). For each plant, inoculum concentration was adjusted to a suitable level as determinedin pre-tests. Inoculum concentration was checked by plating serial dilutions on King's Medium B andcounting the number of cfu (colony forming units).
Inoculation procedure and experimental conditions
Seeds of test plants were sown in plastic sowing trays (with pots 4 cm wide and 6 cm deep forwheat, and 3.2 cm wide and 4.5 cm deep for other plants), containing a 1 : 1 mixture of field soil(ASTROM and GERHARDSON 1988) and greenhouse peat soil (GERHARDSON et al 1985). Afteremergence, the stands were thinned to three plants per pot, with the exception of lettuce, which wasgrown as single plants. Six to eight days after sowing, the plants were watered with 5 ml of bacterialsuspension per pot, except for wheat, which received 10 ml due to the larger pot volume. Controlplants were treated with corresponding volumes of SNB only. Controls without SNB were not
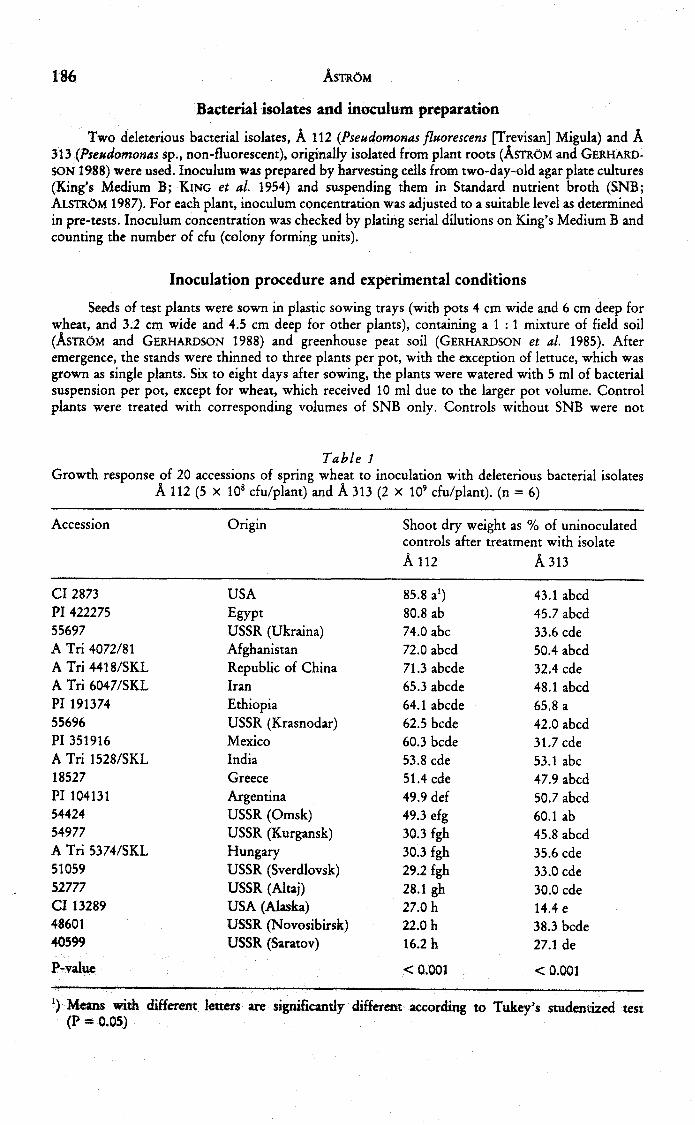
Table 1Growth response of 20 accessions of spring wheat to inoculation with deleterious bacterial isolates
A 112 (5 X 10 cfu/plant) and A 313 (2 X 10' cfu/plant). (n = 6)
Accession
CI 2873PI 42227555697A Tri 4072/81ATri4418/SKLA Tri 6047/SKLPI 19137455696PI 351916A Tri 1528/SKL18527PI 1041315442454977A Tri 5374/SKL5105952777CI 132894860140599
P-valuc
Origin
USAEgyptUSSR (Ukraina)AfghanistanRepublic of ChinaIranEthiopiaUSSR (Krasnodar)MexicoIndiaGreeceArgentinaUSSR (Omsk)USSR (Kurgansk)HungaryUSSR (Sverdlovsk)USSR (Altai)USA (Alaska)USSR (Novosibirsk)USSR (Saratov)
Shoot dry weight as % of uninoculatedcontrols after treatment with isolate
A 112
85.8 a')80.8 ab74.0 abc72.0 abed71.3 abcde65.3 abcde64.1 abcde62.5 bcde60.3 bcde53.8 cde51.4 cde49.9 def49.3 efg30.3 fgh30.3 fgh29.2 fgh28.1 gh27.0 h22.0 h16.2 h
< 0.001
A 313
43.1 abed45.7 abed33.6 cde50.4 abed32.4 cde48.1 abed65.8 a42.0 abed31.7 cde53.1 abc47.9 abed50.7 abed60.1 ab45.8 abed35.6 cde33.0 cde30.0 cde14.4 e38.3 bcde27.1 de
< 0.001
') Mesuis with different letters are significantly different according to Tukey's studentized test(P » 0.05)
Intra- and Interspecific Variation in Plant Response 187
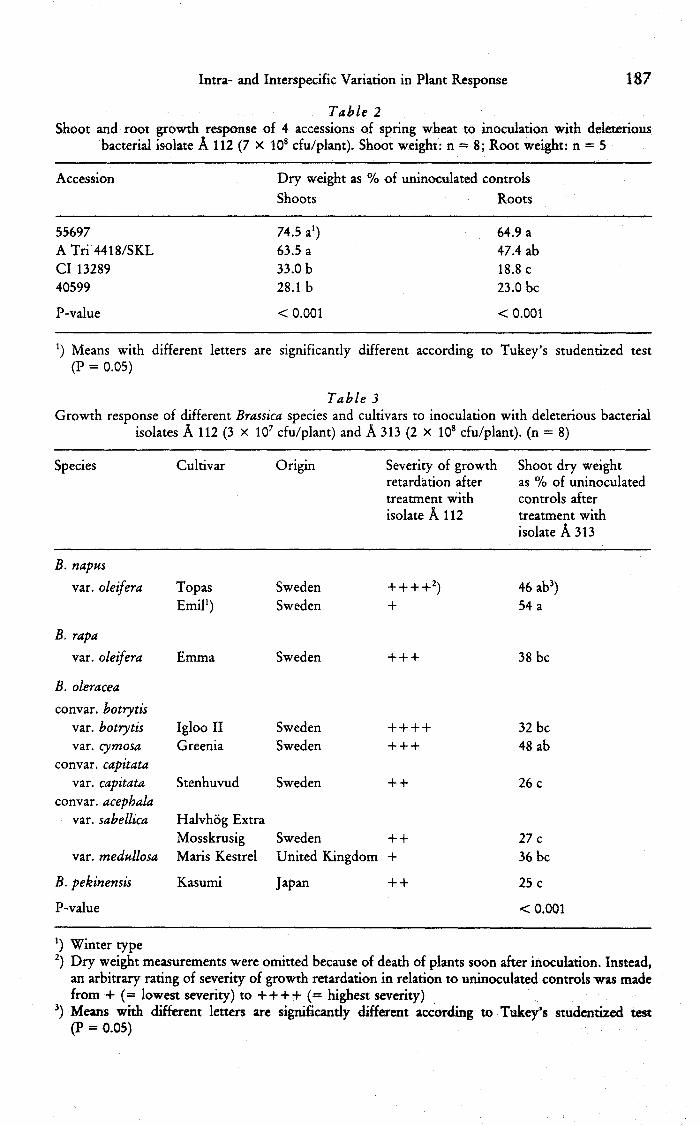
Table 2Shoot and root growth response of 4 accessions of spring wheat to inoculation with deleterious
bacterial isolate A 112 (7 X 10 cfu/piant). Shoot weight: n = 8; Root weight: n = 5
Accession Dry weight as % of uninoculated controlsShoots Roots
55697ATri4418/SKLCl 1328940599
P-value
74.5 aO63.5 a33.0 b28.1b
< 0.001
64.9 a47.4 ab18.8 c23.0 be
< 0.001
) Means with different letters are significantly different according to Tukey's studentized test(P = 0.05)
Table 3Growth response of different Brassica species and cultivars to inoculation with deleterious bacterial
isolates A 112 (3 x 10 cfu/plant) and A 313 (2 X 10' cfu/plant). (n = 8)
Species Cultivar Origin Severity of growthretardation aftertreatment withisolate A 112
Shoot dry weightas % of uninoculatedcontrols aftertreatment withisolate A 313
B. napus
var. oleifera
B. rapa
var. oleifera
B. oleracea
eonvar. botrytisvar. botrytisvar. cymosa
eonvar. capitatavar. capitata
eonvar. acephalavar. sabellica
var. medullosa
B. pekinensis
P-value
TopasEmiP)
Emma
Igloo IIGreenia
Stenhuvud
SwedenSweden 4-
Sweden
SwedenSweden
Sweden
4-4-4-
4-4-4-
4-4-
Halvhog ExtraMosskrusig Sweden 4- 4-Maris Kestrel United Kingdom 4-
Kasumi Japan 4-4-
46 ab')54 a
38 be
32 be48 ab
26 e
27 c36 be
25 e
< 0.001
Winter typeDry weight measurements were omitted because of death of plants soon after inoculation. Instead,an arbitrary rating of severity of growth retardation in relation to uninoculated controls was madefrom 4- (= lowest severity) to 4-4-4-+ (= highest severity)Means with different letters are significantly di^erent according to Tukey's studentized test(P = 0.05)

188 ASTROM
included since SNB in itself has been found to have no, or very small, effects on plant growth(ASTROM, unpublished). One day after inoculation, the plants were transferred to larger pots (10 cmdiameter and 8 cm deep) filled with soil mixture, and then placed in the greenhouse (14 h light/10 hdark, 18 to 22 °C/15 to 18 °C) with natural and artificial (Philips Mercury lamps, 400 W) light givingapproximately 10 kLux at plant height. Water was supplied daily, and nutrient solution (Superba S;Supra Ceres, Landskrona, Sweden) was added three to four times a week. Five weeks afterinoculation, shoots were harvested and dry weight per pot was determined after drying samples for 24h at 105 °C. To test for reproducibility of results, wheat accessions showing extreme reactions wereselected for a repeated test using the same experimental procedure. In this experiment, root weightwas determined after washing root systems free from soil and drying them as above.
Experimental design and statistical evaluation
Eight or ten replicates were used in all cases except in the first wheat experiment, where sixreplicates were used. Determinations of root weight in wheat were made on five replicates. All weightdata were transformed to values relative to control plants, and subjected to analysis of variance (SASPackage, SAS Institute Inc., N. Carolina, USA). Mean relative weights of different accessions werecompared using Tukey's studentized range test at P = 0.05.
Results
Significant differences between wheat accessions were recorded in thegrowth response to inoculation with both bacterial isolates tested (Table 1).Relative shoot growth of selected wheat accessions after inoculation with A 112(Table 2) and A 313 (results not shown here) was similar in the second test, withthe exception of one accession (CI 13289), which was less affected by A 313 thancould be expected from the first test. Table 2 also shows that root growth wasseverely inhibited by bacterial inoculation, and that the differences recorded inshoot growth response were paralleled by differences in root growth response.
Table 4Growth response of different accessions of B. nigra and B. rapa var. oleifera to inoculation with
deleterious bacterial isolates A 112 (6 x 10' cfu/plant) and A 313 (2 X 10* cfu/plant). (n = 8)
Species
B. nigra
B. rapa
var. oleifera
P-value
Accession
cv. Giebracv. Saratovska
cv. Per')cv. Tykocv. TobinWW 438')WW1681
Origin
DDRUSSR
SwedenSwedenCanadaSwedenSweden
Shoot dry weight as % of uninoculatedcontrols after treatment with isolateA 112
76.9 ab )70.9 ab
90.1 a72.5 ab70.5 ab68.6 ab61.3 b
0.036
A 313
61.7 b59.3 b
71.0 b65.7 b74.3 ab88.1a61.6 b
< 0.001
') Winter type) Means with different letters are significantly different according to Tukey's studentized test
(P = 0.05)

Intra- and Interspecific Variation in Plant Response 189
Tables 3 and 4 show that significant variation was found in the reaction toisolate A 313, both among Brassica species, and among accessions withinB. oleracea and B. rapa. Accessions of B. rapa also differed significantly inreaction to isolate A 112 (Table 4). Due to extensive early plant death in one of thetwo experiments with Brassica, probably caused by too high inoculum level, dryweight was not recorded for plants inoculated with A 112. However, subjectiveratings of cultivar reactions indicate that considerable variation in the response tothis isolate is to be found also in B. napus and B. oleracea (Table 3).
Table 5Growth response of seven accessions of spinach to inoculation with deleterious bacterial isolates
A 112 (3 X 10* cfu/plant) and A 313 (2 X 10* cfu/plant). (n = 8)
Accession
cv. DominantXFl')cv. Findus no. 3cv. Findus no. 1XH 185')cv. Findus no. 9X 181')
P-value
Origin
DenmarkSwedenSwedenSwedenSwedenSwedenSweden
Shoot dry weight as % of uninoculatedcontrols after treatment with isolate
A 112
58.1 a')50.6 ab50.0 ab45.5 ab40.2 ab40.1 b33.8 b
0.003
A 313
19.7 be32.7 ab29.5 abc17.1c28.7 abc35.2 a18.9 c
0.027
') Breeding line from Nordreco AB) Means with different letters are significantly different according to Tukey's studentized test
(P = 0.05)
Table 6Growth response of lettuce accessions after inoculation with deleterious bacterial isolates A 112
(7 X 10* cfu/plant) and A 313 (1 X lO' cfu/plant). (n = 10)
Accession Origin Shoot dry weight as % of uninoculatedcontrols after treatment with isolate
A 112 A 313
cv. Red Salad BowlAustralian typecv. Salad Bowlcv. Novitacv. Great Lakes 118cv. Montanacv. Bona
P-value
USAUnknownUSAThe NetherlandsUSAThe NetherlandsSweden
89.5 a')69.7 ab67.7 b59.7 b59.1b58.5 b50.7 b
< 0.001
94.6 a100.1 a96.6 an.t.^)98.5 a96.3 an.t.
0.802
') Means with different letters are significantly different according to Tukey's studentized test(P = 0.05)
) n.t. = not tested

190 ASTROM
With both isolate A 112 and A 313, spinach accessions were found to differsignificantly in relative shoot growth after inoculation (Table 5). In lettuce,significant variation was recorded for isolate A 112, while isolate A 313 did notaffect shoot growth in any of the tested cultivars (Table 6).
Discussion
In the five plant species, Triticum aestivum, Spinacia oleracea, Lactucasativa, Brassica rapa and B. oleracea, significant variation was found betweenaccessions in the reaction to inoculation with one or both bacterial isolates tested(Tables 1 to 6). Also, clear differences between B. napus cultivars were observedin the reaction to isolate A 112, although not quantified by shoot weightmeasurements (Table 3). These results, together with earlier reports (ELLIOTT andLYNCH 1984, ASTROM and GERHARDSON 1988), indicate that the degree of growthinhibition recorded after inoculation with deleterious bacteria is largely deter-mined by the plant genotype, and further, that intraspecific variation in thesensitivity to growth-inhibitory bacteria is a general phenomenon in plants. Theresults thereby also imply the possibility of altering plant reaction to deleteriousbacteria by standard plant breeding techniques of crossing and selection.
The greatest range of variation was found in wheat, which could be expectedsince the wheat material represents a worldwide distribution, whereas in the otherplants tested, the number of entries were fewer and of less diverse origin. The lackof variation in lettuce reaction to A 313, on the other hand, is probably due to thegeneral lack of effect of this bacterial isolate on lettuce (Table 6). This finding is inaccordance with GERHARDSON et al. (1985), who found that lettuce growth wasnot affected by some rhizosphere bacteria that were inhibitory to e.g. wheat andoilseed rape, and also with other reported cases of plant species variation in thesensitivity to deleterious bacteria (GARDNER et al. 1984, CHERRINGTON andELLIOTT 1987, FREDRICKSON et al. 1987). However, it cannot be excluded thatlettuce growth may be affected also by isolate A 313 if the inoculum concentra-tion is increased. Several authors have found that the degree of growth inhibitioninduced by deleterious bacteria is dependent on bacterial number/inoculum dose(FREDRICKSON and ELLIOTT 1985 a, 1985 b, ALSTROM 1987, ASTROM and GERHARD-SON 1989) and, from the results presented here, it is evident that the critical levelfor induction of deleterious effects varies considerably with the plant speciestested. Whereas inoculation with isolate A 112 at 3 to 7 X 10 cfu per plantresulted in mean relative shoot weights of 45 and 51 % in spinach and wheat,respectively, a much lower dose, 3X10'' cells per plant, killed all Brassica plants(Table 3). Further, the mean relative shoot weight in spinach after inoculationwith A 313 at 2 X 10 cfu per plant was 26 %; yet lettuce plants were notinhibited by a tenfold increase in inoculum concentration. Thus, for eachbacterial isolate, the plant species and genera tested can be tentatively rankedaccording to their average sensitivities: for A 112 (from most sensitive to leastsensitive) Brassica » spinach, wheat > lettuce; for A 313 spinach > Brassica >wheat > lettuce.

Intra- and.Interspecific Variation in Plant Response 191
The results indicate specificity in the plant — bacterium interactions also atthe accession level. Although some of the accessions were either highly sensitivee.g. wheat accessions CI 13289 and 40599) or insensitive (e.g. PI 191374) to both
112 and A 313, a ranking of all accessions within a species for sensitivity toinoculation gave clearly different rank orders for the different isolates. In wheat,spinach and B. rapa, where the number of accessions tested was sufficiently high,this observation was also confirmed by statistical analysis (Spearman's rankcorrelation test, P = 0.05). These findings point to a true isolate — plantgenotype specificity, and is consistent with the finding that isolates A 112 and A313 affect plants by different mechanisms (ASTROM and GERHARDSON 1989), but isat variance with the results published by ELLIOTT and LYNCH (1984). A rankcorrelation test using their data on plant growth shows that, although cultivardifferences in reaction were found, the ranking of 16 wheat genotypes wasessentially the same for the two inhibitory pseudomonads tested (rj = 0.775,P < 0.001). The properties of the bacteria used by ELLIOTT and LYNCH were notdescribed, but a plausible explanation for this deviation in results is that theirisolates were more similar in character than the two tested in this study.
It was not within the scope of this study to investigate the genetic back-ground for the intraspecific variation in plant response. However, some of theaccessions tested here would seem suitable for using in a detailed analysis ofinheritance of plant response to deleterious bacteria, e.g. wheat accessions 55697and 40599 in relation to isolate A 112 (Tables 1 and 2). In spinach, it was notedthat differential reactions were recorded also with accessions that are geneticallyrelated. Thus, the reaction of Tindus no. 1' to isolate A 313 differed significantlyfrom that of 'Findus no. 9', which is a result of back-crossing with 'Findus no 1'as recurrent parent (R. STEGMARK, personal communication) (Table 5).
The intraspecific variation in plant reaction to bacterial inoculation may haveseveral explanations. Differences recorded in growth response may reflect differ-ences in root colonization by the bacterium applied, but this hypothesis could notbe substantiated in a previous investigation of the two bacteria tested here(ASTROM and GERHARDSON 1988). Another possible explanation is that accessionsdiffer in their sensitivity to toxic substances formed by the growth-inhibitoryisolates as proposed by FREDRICKSON et al. (1987) for species differences.
I thank all those who supplied germplasm, Ms SUSANNE ANDR£ and Ms ANNIKA GUSTAFSSON
for valuable technical assistance, and Dr. B. GERHARDSON and Dr. MAGNHILD UMAERUS for fruitfuldiscussions and critical reading of the manuscript. This work was supported by the Swedish Councilfor Forestry and Agricultural Research.
Literature
ALSTROM, S., 1987: Factors associated with detrimental effects of rhizobacteria on plant growth. Plantand Soil 102, 3—9.
ASTROM, B., and B. GERHARDSON, 1988: Differential reactions of wheat and pea genotypes to rootinoculation with growth-affecting rhizosphere bacteria. Plant and Soil 109, 263--269.
, and , 1989: Wheat cultivar reaaions to deleterious rhizosphere bacteria under gnotobio-tic conditions. Plant and Soil 117, 157—165.

192 ASTROM, Intra- and Interspecific Variation in Plant Response
BURR, T . J., and A. CAESAR, 1984: Beneficial plant bacteria. CRC Critical Reviews in Plant Sciences2, 1—20.
CAMPBELL, J . N., K. CONN, L. SORLIE, and F. D. COOK, 1986: Inhibition of growth in canolaseedlings caused by an opportunistic Pseudomonas sp. under laboratory and field conditions.Can. J. Microbiol. 32, 201—207.
CHERRINGTON, C . A., and L. F. ELLIOTT, 1987: Incidence of inhibitory pseudomonads in the PacificNorthwest. Plant and Soil 101, 159—165.
CURL, E. A., and B. TRUELOVE, 1986: The Rhizosphere. Springer-Verlag, Berlin.ELLIOTT, L. F., and J. M. LYNCH, 1984: Pseudomonads as a factor in the growth of winter wheat
(Triticum aestivum L.). Soil. Biol. Biochem. 16, 69—71.FREDRICKSON, J. K., and L. F. ELLIOTT, 1985 a: Colonization of winter wheat roots by inhibitory
rhizobacteria. Soil. Sci. Soc. Am. J. 49, 1172—1177., and , 1985 b: Effects on winter wheat seedling growth by toxin-producing rhizobacteria.
Plant and Soil 83, 399—409., , and J. C. ENGIBOUS, 1987: Crop residues as substrate for host-specific pseudomonads.
Soil. Biol. Biochem. 19, 127—IM.GARDNER, J. M., J. L. CHANDLER, and A. W. FELDMAN, 1984: Growth promotion and inhibition by
antibiotic-producing fluorescent pseudomonads on citrus roots. Plant and Soil 77, 103—113.GERHARDSON, B., S. ALSTROM, and B. RAMERT, 1985: Plant reactions to inoculation of roots with
fungi and bacteria. Phytopath. 2. 114, 108—117.KATZNELSON, H . , 1965: Nature and importance of the rhizosphere. In: K. F. BAKER, and W. C.
SNYDER (eds). Ecology of Soil-borne Plant Pathogens, pp. 187—209. Univ. of CaliforniaPress, Berkeley, Los Angeles.
KING, E . O . , M . K. WARD, and D. E. RANEY, 1954: Two simple media for the demonstration ofpyocyanin and fluorescin. J. Lab. & Clin. Med. 44, 301—307.
NEAL, J. L. Jr., T. G. ATKINSON, and R. I. LARSON, 1970: Changes in the rhizosphere microflora ofspring wheat induced by disomic substitution of a chromosome. Can. J. Microbiol. 16,153—158.
, R. I. LARSON, and T. G. ATKINSON, 1973: Changes in rhizosphere populations of selectedphysiological groups of bacteria related to substitution of specific pairs of chromosomes inspring wheat. Plant and Soil 39, 209—212.
SCHIPPERS, B., A. W. BAKKER, and P. A. H. M. BAKKER, 1987: Interactions of deleterious andbeneficial rhizosphere microorganisms and the effect of cropping practices. Ann. Rev.Phytopathol. 25, 339—358.
SuSLOW, T. V., 1982: Role of root-colonizing bacteria in plant growth. In: M. S. MOUNT and G. H.LACY (eds), Phytopathogenic Procaryotes, Vol. 1, pp. 187—223. Academic Press, London.
, and M. N. SCHROTH, 1982: Role of deleterious rhizobacteria as minor pathogens in reducingcrop growth. Phytopathology 72, 111—115.
TuRCO, R. F., M. BiscHOFF, D. P. BREAKWELL, and D. R. GRIFFITH, 1990: Contribution of soil-borne bacteria to the rotation effect in corn. Plant and Soil 122, 115—120.
WOLTZ, S. S., 1978: Nonparasitic plant pathogens. Ann. Rev. Phytopathol. 16, 403—430.























![[] Microbial Ecology of the Rhizosphere(BookFi.org)](https://img.pdfslide.net/doc/110x75/55cf944c550346f57ba106f1/-microbial-ecology-of-the-rhizospherebookfiorg.jpg)






