Embed Size (px)
Citation preview

Investigating the Influence of D4 Receptor Modulation on Alcohol Addiction-Relevant Behaviours
by
Aaron Kim
A thesis submitted in conformity with the requirements for the degree of Master of Science
Department of Pharmacology & Toxicology
University of Toronto
© Copyright by Aaron Kim 2018

ii
Examining the Influence of D4 Receptor Modulation on Alcohol Addiction-Relevant Behaviours
Aaron Kim
Master of Science
Department of Pharmacology & Toxicology
University of Toronto
2018
Abstract
The recent development of selective dopamine D4 receptor ligands has made it possible to
further examine their effects on addiction-relevant behaviours. This study investigated the effects of D4
receptor modulation, using the selective antagonist L-745,870 and agonist PD 168,077, on alcohol self-
administration and reinstatement induced either by cue or stress.
Although activation of the D4 receptor was ineffective, inactivation attenuated alcohol self-
administration without affecting food self-administration. Initial analyses of reinstatement experiments
found no significant effects, but by distilling for robust responders, alcohol-seeking was reduced by D4
receptor inactivation under stress-induced reinstatement.
The culmination of this work deepens existing lines of evidence that the D4 receptor is involved
in addictive disorders, and suggests that D4 receptor blockade diminishes motivation for alcohol-taking
without influencing natural food rewards. Furthermore, there appears to be a plausible effect of D4
receptor blockade interfering with stress- but not cue-induced alcohol-seeking.

iii
Acknowledgements Every step of the way, I struggled with finding the perfect way to concisely articulate facts but I did not anticipate struggling even more with sentiments. During this extensive journey, I have had to encounter my limitations regularly, but I overcame them each time with the help of everyone around me. And so, I would like to express my sincere gratitude to all those that made this thesis possible (a list proportionate to the support I received): First and foremost, I would like to thank my supervisor, Dr. Le Foll, for his support and more importantly, for allowing me the freedom to work at my own pace. Pamela Sabioni for her endless encouragement and humour. Abhiram Pushparaj, Andrew Loughlin, Annie Peng, and Jeremy Watts, for their valuable friendship and technical assistance. Casey Suchit and Jack Bilodeau for always being welcoming and kind. William Ngo for his probe-like insight and encouragement, Kenneth Yip for pumping out work sessions with me when I was losing steam, Thomas Truong for maintaining my physical health, and Anh Chu for helping me cross the finish line. And finally, Alani Van, the love of my life, for your unconditional support and understanding; you have been there for me every step of the way.

iv
Table of Contents Acknowledgements ...................................................................................................................................... iii
Table of Contents ......................................................................................................................................... iv
List of Abbreviations ................................................................................................................................... vii
List of Figures ................................................................................................................................................ x
CHAPTER 1. Introduction .............................................................................................................................. 1
1.1 Addiction ............................................................................................................................................. 1
1.1.1 Defining Addiction........................................................................................................................ 1
1.1.2 Addiction: A Health Burden for the World .................................................................................. 1
1.1.3 Alcohol and Disease ..................................................................................................................... 2
1.1.4 Current Therapeutic Drugs for Alcohol Use Disorders................................................................. 3
1.2 Animal Models .................................................................................................................................... 4
1.2.1 Identifying Novel Therapies with Animal Models ........................................................................ 4
1.2.2 Voluntary Consumption Models .................................................................................................. 6
1.2.3 Operant Self-Administration Models ........................................................................................... 6
1.2.4 Reinstatement Models ................................................................................................................. 8
1.3 Dopamine ............................................................................................................................................ 9
1.3.1 Brief History of Dopamine ........................................................................................................... 9
1.3.2 Dopamine Biochemistry ............................................................................................................. 11
1.3.3 Dopaminergic Pathways ............................................................................................................ 12
1.4 The Dopamine Hypothesis of Addiction ........................................................................................... 14
1.4.1 Hypotheses of Dopamine and Reward ...................................................................................... 14
1.4.2 Theories of Executive Function .................................................................................................. 16
1.4.3 Theories of Relapse .................................................................................................................... 17
1.4.4 Synthesis and Challenges of the Dopamine Hypothesis ............................................................ 22
1.5 The Dopamine D4 Receptor .............................................................................................................. 24
1.5.1 The Dopamine D4 Receptor and Antipsychotics ....................................................................... 24
1.5.2 D4 Receptor Distribution ........................................................................................................... 25
1.5.3 D4 Receptor Genetics and Polymorphisms ................................................................................ 25
1.5.4 D4 Receptor Signalling ............................................................................................................... 26
1.5.5 D4 Receptor VNTR Polymorphism ............................................................................................. 27
1.6 Role of the D4 Receptor in Addiction ............................................................................................... 29

v
1.6.1 Early D4 Behavioural Studies ..................................................................................................... 29
1.6.2 Addiction-Relevant Neuroadaptations of the D4 Receptor ....................................................... 31
1.6.3 The Role of D4 Receptors in Gambling Disorders ...................................................................... 32
1.6.4 The D4 Receptor and Alcohol .................................................................................................... 33
1.6.5 The D4 Receptor and Other Substances of Abuse ..................................................................... 36
1.7 Experimental Rationale ..................................................................................................................... 39
1.7.1 Summary Overview .................................................................................................................... 39
1.7.2 Experimental Rationale .............................................................................................................. 39
CHAPTER 2. Methods .................................................................................................................................. 40
2.1 Animals .............................................................................................................................................. 40
2.2 Drug Preparation ............................................................................................................................... 41
2.3 Alcohol Training ................................................................................................................................ 41
2.3.1 Two-Bottle Choice Alcohol Acquisition ...................................................................................... 41
2.3.2 Operant Alcohol Self-Administration ......................................................................................... 42
2.4 Sorting ............................................................................................................................................... 44
2.5 Alcohol Self-Administration Testing .................................................................................................. 44
2.6 Extinction .......................................................................................................................................... 45
2.7 Cue-Induced Reinstatement Testing ................................................................................................. 45
2.8 Stress-Induced Reinstatement Testing ............................................................................................. 45
2.9 Cohort Sequence ............................................................................................................................... 46
2.10 Food Testing .................................................................................................................................... 46
2.11 Data Analysis ................................................................................................................................... 47
CHAPTER 3. Results ..................................................................................................................................... 47
3.1 Alcohol Consumption Training .......................................................................................................... 47
3.1.1 Two-Bottle Choice ...................................................................................................................... 47
3.1.2 Operant Oral Self-Administration .............................................................................................. 51
3.2 Alcohol Self-Administration Testing .................................................................................................. 52
3.2.1 The Effects of the Selective DRD4 Antagonist, L-745,870, on Alcohol Self-Administration ...... 52
3.2.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Alcohol Self-Administration ........... 53
3.3 Cue-Induced Reinstatement ............................................................................................................. 55
3.3.1 The Effects of the Selective DRD4 Antagonist L-745,870 on Cue-Induced Reinstatement ....... 55
3.3.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Cue-Induced Reinstatement .......... 57
3.4 Stress-Induced Reinstatement .......................................................................................................... 59

vi
3.4.1 The Effects of the Selective DRD4 Antagonist L-745,870 on Stress-Induced Reinstatement .... 59
3.4.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Stress-Induced Reinstatement ...... 62
3.5 Food Control ..................................................................................................................................... 65
3.5.1 Effects of L-745,870 on Food Self-Administration ..................................................................... 65
CHAPTER 4. Discussion ................................................................................................................................ 66
4.1 Overall Outcomes ............................................................................................................................. 66
4.2 Overview of Relevant Literature ....................................................................................................... 67
4.2.1 Summary of D4 Receptor Literature ........................................................................................... 67
4.3 Experimental Paradigms ................................................................................................................... 68
4.3.1 Operant Self-Administration Paradigm ...................................................................................... 68
4.3.2 Cue-Induced Reinstatement ...................................................................................................... 69
4.3.3 Stress-Induced Reinstatement ................................................................................................... 71
4.4 Theoretical Implications of Findings ................................................................................................. 73
4.4.1 Dopamine Reward Hypothesis ................................................................................................... 73
4.4.2 Theories of Executive Function .................................................................................................. 73
4.5 Challenges and Limitations ............................................................................................................... 75
4.6 Conclusions ....................................................................................................................................... 78
4.7 Future Directions .............................................................................................................................. 78
References .................................................................................................................................................. 80

vii
List of Abbreviations
2BC two-bottle choice 6-OHDA 6-hydroxydopamine AADC aromatic L-amino acid decarboxylase AAF alcohol-attributable fraction AC adenylyl cyclase ACTH adrenocorticotropic hormone ADE alcohol deprivation effect ADH alcohol dehydrogenase ADHD attention-deficit hyperactivity disorder ALDH acetaldehyde dehydrogenase AMPA α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid ANOVA analysis of variance ASDR age-standardized death rate ATP adenosine triphosphate AUD alcohol use disorder BAC blood alcohol content BLA basolateral amygdala BNST bed nucleus of stria terminalis bp base pairs CaMKII Ca2+/calmodulin-dependent protein kinase cAMP cyclic adenosine monophosphate CBT cognitive behavioural therapy CCAC Canadian Council on Animal Care CeA central nucleus of the amygdala CRF corticotropin-releasing factor CS conditioned stimulus DALY disability-adjusted life year DARPP-32 dopamine and cAMP-regulated phosphoprotein DMSO dimethyl sulfoxide dPFC dorsal prefrontal cortex
DRD1 dopamine receptor D1
DRD2 dopamine receptor D2
DRD3 dopamine receptor D3
DRD4 dopamine receptor D4
DRD5 dopamine receptor D5 DSM-5 Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition EPM elevated plus-maze fMRI-BOLD functional magnetic resonance imaging blood-oxygen-level-dependent FR fixed-ratio

viii
GABA γ-aminobutyric acid GBD Global Burden of Disease GPCR G-protein-coupled receptors HPA hypothalamic-pituitary-adrenal i.p. intraperitoneal IR immunoreactivity Kir3 potassium inwardly rectifying channel L-allele long allele L-DOPA L-3,4-dihydroxyphenylalanine LSD lysergic acid diethylamide LTN lateral tegmental nucleus MFB medial forebrain bundle MOR µ-opioid receptor mPFC medial prefrontal cortex NAc nucleus accumbens NAM negative allosteric modulator NCD non-communicable diseases NE norepinephrine NHP non-human primates NMDA N-methyl-D-aspartate OFC orbitofrontal cortex PAM positive allosteric modulator PCP phencyclidine PD pharmacodynamics PET positron emission tomography PFC prefrontal cortex PK pharmacokinetics PKA protein kinase A PLCγ phosphoinositide phospholipase C-γ PPP1CA protein phosphatase 1 PVN paraventricular nucleus rGT rat gambling task rSMT rodent slot machine task S-Allele short allele SNc substantia nigra pars compacta SNP single nucleotide polymorphism SPECT single-photon emission computer tomography SUD substance use disorder TCI Temperament and Character Inventory TH tyrosine hydroxylase VNTR variable number tandem repeat VP ventral pallidum VTA ventral tegmental area

ix
WHO World Health Organization YLD years lost due to disability YLL years of life lost

x
List of Figures
Figure 1 Aggregate two-bottle choice training over 25 days Figure 2 Separate two-bottle choice training over 25 days Figure 3 Alcohol dose over the last 10 days Figure 4 Aggregate operant self-administration training Figure 5 Effects of the DRD4 antagonist L-745,870 on alcohol self-administration Figure 6 Effects of the DRD4 agonist PD-168,077 on alcohol self-administration Figure 7 Effects of the DRD4 antagonist L-745,870 on cue-induced reinstatement Figure 8 Effects of the DRD4 antagonist L-745,870 on cue-induced reinstatement Figure 9 Effects of the DRD4 agonist PD 168,077 on cue-induced reinstatement Figure 10 Effects of the DRD4 antagonist L-745,870 on stress-induced
reinstatement Figure 11 Effects of the DRD4 antagonist L-745,870 on stress-induced
reinstatement with a threshold of ≥40 active lever presses for both vehicle tests
Figure 12 Effects of DRD4 agonist PD-168,077 on stress-induced reinstatement Figure 13 Effects of dopamine agonist PD-168,077 on stress-induced reinstatement
with a threshold of ≥20 active lever presses for both vehicle tests Figure 14 Food self-administration under a FR3 schedule of reinforcement

1
CHAPTER 1. Introduction 1.1 Addiction 1.1.1 Defining Addiction
The term ‘addiction’ originates from the Latin verb addico, referring to the state of being ‘given
over’. In one context, it was used as a label for bond slaves, and in a favourable sense, to express
devotion (Alexander & Schweighofer, 1988; Lewis & Short, 1879). The essence of the latter definition
endured until the alcohol temperance and anti-opium movements of the 19th century, where its usage
became gradually restricted to an association with drugs of abuse and their harmful effects (Levine,
1978). The advent of these movements prompted the establishment of addiction as a legitimate illness
falling into the domain of public health (Parssinen & Kerner, 1980). And through extensive research and
neurological understanding, the definition has progressed to encompass behavioural addictions,
including gambling disorders. Today, addiction is understood as a chronic relapsing disorder that
involves impaired response inhibition and enhanced salience attribution at the cognitive level,
underlying compulsive seeking and taking of rewarding stimuli at the behavioural level, despite harmful
consequences (Goldstein & Volkow, 2002; National Institute on Drug Abuse, 2014).
Standards for the classification and diagnosis of addiction are perpetually changing in efforts to
better serve patients. For instance, the widely accepted nomenclature system presented by the
Diagnostic and Statistical Manual of Mental Disorders (5th ed.; DSM-5; American Psychiatric Association,
2013) recently replaced their dichotomy of substance abuse and dependence with a unified category of
‘substance use disorders’ (SUDs). As a result, alcohol abuse and alcohol dependence have been
integrated under alcohol use disorders (AUDs).
1.1.2 Addiction: A Health Burden for the World Historically, the allocation of resources and awareness towards mental and substance use
disorders were minimal and considered a low priority concern for public health (Ustün, 1999). Once the
World Development Report (World Bank, 1993) shed light on the importance of investing in health and

2
its potential translatable economic growth, public attention shifted away from mortality, and towards
morbidity, paving the way towards a closer examination of mental and substance use disorders.
Subsequent epidemiological studies have determined that substance-related and addictive disorders
play a substantial role in the global burden of disease (Degenhardt et al., 2013; Lim et al., 2012).
The burden of disease attributable to mental and substance use disorders is generally measured
in terms of disability-adjusted life years (DALYs), alternatively understood as the difference between the
current and ideal health status of a population. The calculation of DALYs takes the sum of years of life
lost to premature mortality (YLL) and the years lost due to disability (YLD), an estimate of quality life lost
due to health conditions (World Health Organization, 2008).
Through the Global Burden of Disease Study (GBD) 2010, Whiteford (2013) estimated that the
burden of mental and substance use disorders accounted for 7.4% (183.9 million) of all DALYs worldwide
and were also the leading cause of YLDs. And from this category, AUDs accounted for 9.6% of DALYs and
had the highest YLLs (44.4%) with the largest burden occurring at 25-50 years of age (Whiteford et al.,
2013). Another analysis of the GBD 2010 by Lim et al. (2012) found that the contribution of risk factors
to disease burden changed significantly since 1990, moving away from communicable diseases in
children, and towards non-communicable diseases (NCD) in adults. And as a consequence of this
change, alcohol use was identified as the 3rd leading risk factor for global disease burden (Lim et al.,
2012).
1.1.3 Alcohol and Disease As with most substance use disorders, AUDs are highly comorbid with mood, anxiety, conduct,
and antisocial behaviour disorders (Merikangas et al., 1998). And beyond mental disorders, alcohol use
is linked to many other negative health consequences. A review by Boffetta and Hashibe (2006)
summarized that alcohol consumption is causally associated with cancers of the oral cavity, liver, colon,
rectum, and breast. Although not fully understood, possible mechanisms for the carcinogenic effects of

3
alcohol include: facilitating solubility of tobacco’s carcinogens, genotoxic effects of the metabolite
acetaldehyde, production of reactive oxygen species, and changes in folate metabolism (Wright &
Morgan, 2013).
Alcohol use is also known to cause numerous diseases such as alcoholic polyneuropathy,
cardiomyopathy, gastritis, hepatitis, cirrhosis, and hemorrhagic stroke (Rehm et al., 2004; Reynolds et
al., 2003). A report by the World Bank (1993) estimated that from the annual 2 million alcohol-related
deaths worldwide, liver cirrhosis was responsible for 50%, liver cancer for 35%, and 10% from AUDs.
Additionally, prenatal alcohol exposure is associated with many detrimental effects such as deficiencies
in cognitive functioning, fetal alcohol syndrome, premature birth, and intrauterine growth retardation
(Connor & Streissguth, 1996).
While modest alcohol consumption at 2.5 to 14.9 g (≤1 drink) per day has been found to have
protective effects such as lower stroke incidence and risk reduction to coronary heart disease, even low
levels of alcohol intake have been associated with an increased risk of liver cirrhosis (Corrao et al., 1998;
Klatsky, 1999; Ronksley et al., 2011). Higher levels of alcohol consumption were found to be associated
with acute and chronic pancreatitis, high blood pressure, and cardiovascular disease, ischemic stroke in
particular (Durbec & Sarles, 1978; Mukamal, 2005; Thakker, 1998).
A global status report from the World Health Organization (WHO; 2014) estimated that the 2010
prevalence of AUDs in Canada in those aged 15 years and older was 6.8% for both sexes. The report
found that the age-standardized death rate (ASDR) for liver cirrhosis in males was 10.6 (per 100,000)
and that the alcohol-attributable fraction (AAF) was 62.5%. Whereas the ASDR for road traffic accidents
in males was 11.0 (per 100,000) and that the AAF was 13.8% (WHO, 2014).
1.1.4 Current Therapeutic Drugs for Alcohol Use Disorders In recent history, several therapeutic approaches have been utilized to treat AUDs. A previously
common aversion strategy used disulfiram, an irreversible inhibitor of acetaldehyde dehydrogenase

4
(ALDH). Normally, alcohol is oxidized to acetaldehyde by alcohol dehydrogenase (ADH) and
subsequently converted into acetic acid by ALDH; disulfiram blocks the latter step, increasing the
concentration of acetaldehyde in the human body. As acetaldehyde is responsible for aversive
‘hangover’ symptoms, disulfiram treatment causes them to be experienced immediately upon alcohol
consumption. As an aversion strategy, along with the inability to influence alcohol craving, disulfiram is
naturally prone to poor compliance, and has since been replaced by newer treatments such as
naltrexone and acamprosate.
Naltrexone acts as a competitive antagonist of the µ-opioid receptor (MOR), and although the
pharmacodynamics are not fully elucidated, it is believed to generate its therapeutic effect via the
dopaminergic mesolimbic pathway. Although naltrexone is effective in reducing relapse to heavy
drinking, its overall benefit has been regarded as ‘modest’ (Garbutt, 2010). The pharmacodynamics of
acamprosate are even more poorly understood but physiological studies have indicated weak N-methyl-
D-aspartate (NMDA) receptor antagonism and potential positive allosteric modulation of γ-aminobutyric
acid (GABAA) receptors (Littleton & Zieglgänsberger, 2003). Conversely, acamprosate is effective in
maintaining abstinence but only when detoxified before treatment (Maisel et al., 2013). Thus,
investigating alternative interventions for AUDs could usher in a new era of treatment strategies better
suited to addressing neurophysiological processes relevant to addiction.
1.2 Animal Models 1.2.1 Identifying Novel Therapies with Animal Models
The vast negative health ramifications brought on by substances of abuse demand further
exploration of addictive processes and therapeutic strategies. As conducting human studies are often
challenging due to safety concerns, as well as ethical and experimental constraints, researchers rely on
animal models. Similar to cell cultures for biological studies, animals are crucial for behavioural
investigations. With the use of animal models, important information on efficacy, toxicity, and
pharmacokinetics-pharmacodynamics (PK; PD) can be collected on novel drugs to ensure safety prior to

5
human testing (Tabakoff & Hoffman, 2000). One of the most significant advantages of animal models is
that they allow for highly invasive procedures that would otherwise be infeasible in humans (i.e. brain
lesioning).
Although unable to fully emulate humans, the relative simplicity of animal models allow for
efficiency, and when carefully designed, complex human disorders can be reduced into component parts
for further study. And continuous improvements on these models can yield a better understanding of
disorders under investigation to facilitate the discovery of new therapeutic strategies.
Despite improving efficiency, there is no guarantee that a model will be adequate for what it is
intended to study, which is why validity must be appropriately accounted for. Validity refers to the
extent that an assessment accurately measures what it intends to measure. Our understanding of
addiction in the clinical environment relies heavily on the validity of animal models. As described by
Willner (1991), the most relevant forms of validity in preclinical models are: 1) face, 2) predictive, and 3)
construct validity.
Face validity is a subjective evaluation of whether a test measures what it claims to measure. In
animal models, this refers to whether the animal’s behaviour corresponds to what is seen in humans.
Indeed, animal models tend to have high face validity as they are often able to exhibit aspects of the
human condition being studied. This is apparent in the similarity between animals that present with
voluntary drug-seeking and the behaviour of humans with addiction. Naturally, non-human primates
have a high degree of face validity as they are most closely related to humans. However, due to their
high maintenance costs and the substantial amount of expertise required, rodent models are preferable.
Predictive validity refers to the degree in which an operationalization can predict outcomes in
the same construct that is measured elsewhere. Animal models are valuable for their high predictive
validity as they are informative in how effective treatments will be on humans, particularly when it
comes to assessing the potential abuse liability of novel drugs in humans.

6
Construct validity refers to the extent that an operationalization, and what it attempts to
measure, are grounded in theory and the existing literature. The theoretical rationale must be sound for
there to be construct validity connecting the human condition and the animal model.
As addictive disorders are inherently complex, animal models provide an avenue to reduce them
into manageable parts for deeper study. And by generating the component characteristics of addiction
in rodents, it is possible to measure changes in behaviour, biochemistry, and gene expression. The
animal models relevant to this thesis will be briefly summarized along with their advantages and
disadvantages.
1.2.2 Voluntary Consumption Models Many different variants of assessing voluntary alcohol intake exist, but the most prevalent is the
two-bottle choice (2BC) paradigm. In 2BC, animals are presented with two bottles: one containing an
alcohol solution, and the other, a non-alcoholic fluid such as water or a sucrose solution. Depending on
the experimental design, access to the bottles is either available throughout or limited during the day.
This paradigm is used to assess general avidity for alcohol and demonstrates that
heterogeneous animal populations display varying levels of alcohol consumption that are comparable to
humans. While this model appears to have high face validity, it is difficult to identify the underlying
impetus behind their behaviour and so, it is unclear as to how accurately it represents the actual human
condition of addiction (Tabakoff & Hoffman, 2000). A further disadvantage of this model is the relative
coarseness of its measurement and its potential to be affected by unrelated factors such as taste and
calories (Heilig & Koob, 2007).
1.2.3 Operant Self-Administration Models Measuring motivation to obtain a drug reward in animals may be difficult in voluntary
consumption models, but this limitation can be overcome through operant self-administration
approaches. These models employ the behavioural concept of reinforcement: with positive
reinforcement, the presentation of a rewarding stimulus is used to increase the probability of a

7
response, whereas with negative reinforcement, the removal of an aversive stimulus or negative
internal state (i.e. withdrawal) is used to decrease the probability of a response. Operant self-
administration models use positive reinforcers such as food or addictive substances to reliably increase
the probability of a specific behaviour (i.e. lever-pressing). This allows for the means to examine
motivation for voluntary responding as well as the direct reinforcing capacity of a stimulus. The major
advantages of operant self-administration models are their high face and construct validity. Drugs that
have high abuse liability in humans are able to reinforce self-administration in animals and adjustments
in dose are capable of affecting responding. Additionally, the theoretical rationale for the
operationalization of motivation is strongly supported by the literature, satisfying construct validity
(Kalivas et al., 2006).
Operant self-administration models commonly utilize a FR schedule of reinforcement in which
the delivery of the reinforcement is contingent on a fixed number of responses. The number of
responses is interpreted as the amount of effort an animal is willing to commit to receiving a rewarding
stimulus and can be further assessed by manipulating the FR requirement. This operationalization is able
to show that alcohol consumption is due to pharmacological motivation instead of other factors such as
appetite or thirst (Tabakoff & Hoffman, 2000). This approach is beneficial because it produces a
quantifiable stable behaviour and allows a large number of animals to learn the operant behaviour with
relative ease (Roberts & Zito, 1987; Schindler et al., 2002).
Under FR schedules of reinforcement, animals will self-administer addictive substances in
attempts to maintain constant levels of dopamine in the nucleus accumbens (NAc; Robbins & Everitt,
1992). Over a range of doses that are able to sustain stable responding, changes in dose are followed by
compensatory behaviour; when dose is decreased, animals will increase their responding, and an
increase in dose will decrease their response rate (Heidbreder, 2011).

8
Similar to the effects of a decrease in drug dose, Yokel & Wise (1975) observed that stimulant
intake increased following neuroleptic treatment, which partially blocks dopamine receptors. These
increases in responding were interpreted as a reduction in the reinforcing value of the stimulant, where
intake had to increase to achieve previous levels of reinforcement. However, Roberts & Zito (1987)
argued against this interpretation as they found that 6-hydroxydopamine (6-OHDA)-induced destruction
of dopamine terminals in the NAc resulted in a decrease of stimulant self-administration rather than an
increase.
To add to this complexity, preclinical self-administration studies have established a
characteristic inverted U-shaped function between drug dose and response for many major drugs of
abuse, including alcohol (Corrigall & Coen, 1989; Grahame & Grose, 2003). For both very low and high
relevant doses, response rates were significantly lower when compared to acquisition doses. In fact,
substantial suppression of the reinforcer displays extinction-like behaviour where animals initially
increase responding, followed by a cessation of responding (Yokel & Wise, 1975). As a result of these
issues, self-administration using an FR schedule can be difficult to discriminate whether a treatment has
potentiating or diminishing effects on a reinforcer.
On another note, operant alcohol self-administration can also be used to model the withdrawal
symptoms of alcohol craving observed in alcoholics, contributing to face and predictive validity. In a
phenomenon known as the alcohol deprivation effect (ADE), after a period of forced abstinence, animals
that regularly consumed alcohol will exhibit a relapse-like drinking state when alcohol is made available
again.
1.2.4 Reinstatement Models Even after successful detoxification and extended abstinence, high rates of relapse in drug-
seeking and drug-taking pose significant challenges to the long-term treatment of addiction. Given the

9
complexity of addiction, the ability to examine the aspect of relapse susceptibility in preclinical models is
instrumental in developing novel therapies.
The reinstatement paradigm extends on the operant self-administration model and has three
general steps: acquisition, extinction, and reinstatement-testing. Animals undergo self-administration
training where drug delivery is typically paired with the presentation of a conditioned stimulus (CS) such
as a discrete cue (i.e. light or tone). Subsequently, the animal enters a period of forced abstinence,
during which the drug reinforcer and paired CS are withheld, leading to the gradual extinction of drug-
seeking behaviour.
Once the animal is sufficiently extinguished, drug-seeking behaviour is reinstated and measured
in response to different conditions: re-exposure to the CS (cue-induced reinstatement), a non-
contingent drug prime (drug-induced reinstatement), or a stressor (stress-induced reinstatement).
These methods of inducing reinstatement have been demonstrated in the animal literature for a variety
of drugs of abuse, including cocaine, opiates (Stewart, de Wit, & Eikelboom, 1984), nicotine (Feltenstein
et al., 2012), and alcohol (Chiamulera et al. 1995). Similar to operant self-administration models, the
reinstatement paradigm exhibits strong face and construct validity as each of the relapse-inducing
methods are able to induce craving and relapse in humans as well (Sinha & Li, 2007). The theoretical
frameworks and neurocircuitry underlying relapse and different reinstatement methods will be
discussed at length in a later chapter.
1.3 Dopamine 1.3.1 Brief History of Dopamine
Evidently, an abundance of research has established dopaminergic transmission as one of the
major mechanisms in which addiction develops (Koob & Le Moal, 2001). And so an understanding of the
history behind dopamine and its pathways can shed deeper insights on the neurobiology of addiction.
Initially synthesized in 1910, dopamine (3,4-dihydroxyphenethylamine) was classified as a
sympathomimetic drug and went largely unnoticed for several decades (Barger & Dale, 1910; Barger &

10
Ewins, 1910). During efforts to reach consilience for monoamine oxidase as the enzyme responsible for
catalyzing the degradation of adrenaline and other amines, dopamine was inadvertently found to be a
suitable substrate as well (Blaschko, Richter, & Schlossmann, 1937). Shortly afterwards, Holtz et al.
(1938) showed that the naturally occurring aromatic L-amino acid decarboxylase (AADC) catalyzed the
reaction of L-3,4-dihydroxyphenylalanine (levodopa or L-DOPA) to dopamine, which then led Blaschko
(1939) to deduce that dopamine was a precursor to norepinephrine (NE) and epinephrine. By the late
1950s, dopamine was detected in the human brain as well as the corpus striatum of the dog brain,
leading to its recognition as a neurotransmitter (Bertler & Rosengren, 1959; Carlsson, 1959;
Hornykiewicz, 1986; Montagu, 1957).
The following decades were marked by significant clinical developments in dopamine research.
At the time, antipsychotics were known to be effective in treating schizophrenia, but its principal mode
of action was unclear. Carlsson's (1963) speculation that dopamine blockade was responsible for this
effect was later corroborated by numerous studies (Iversen, 1975; Snyder et al., 1974). Among them was
the discovery that dopamine and haloperidol both bound to the same site, which came to be known as
the D2 receptor (Seeman et al., 1976). A significant landmark was reached when a successful high-dose
oral regimen of L-DOPA achieved sustained improvement in patients with Parkinson’s disease by
eliminating rigidity and tremors (Cotzias, Van Woert, & Schiffer, 1967). Afterwards, the field of
dopamine research saw tremendous growth on all fronts (Hornykiewicz, 1986).
Building on previous findings, early preclinical studies found that dopamine blockade by
antipsychotics suppressed the initiation of voluntary movement and impaired acquisition of food-
reinforced lever-pressing, while excessive activation resulted in sustained hyperactivity as well as
impaired discriminative ability and response inhibition (Ahlenius, 1979; Wise & Schwartz, 1981). It was
believed that dopamine blockade interfered with the hedonic impact of rewarding stimuli (Wise, 1982).

11
Dopamine is now known to play a neuromodulatory role in several key functions of the brain
such as motor control, motivation, reward, learning, working memory, and endocrine regulation. Most
importantly, dopamine mediates the primary reinforcing aspects of addictive drugs, which will later be
further elaborated upon (Girault & Greengard, 2004; Greengard, 2001; Schultz et al., 1997).
1.3.2 Dopamine Biochemistry 1.3.2.1 Dopamine Synthesis
Dopamine belongs to a group of catecholamines, organic compounds containing a catechol (a
benzene ring with two hydroxyl groups) and side-chain amine. Dopamine is generally synthesized in two
steps. First, the amino acid tyrosine is converted into L-DOPA by tyrosine hydroxylase (TH) with the
cofactors: molecular oxygen, iron, and tetrahydrobiopterin (Nagatsu et al., 1964). In the second step, L-
DOPA is converted into dopamine via AADC with pyridoxal phosphate as a cofactor (Holtz, Credner, &
Koepp, 1942; Molinoff & Axelrod, 1971). In neuronal groups that do not possess dopamine β-
hydroxylase, an enzyme responsible for converting dopamine to NE, dopamine is the end product.
1.3.2.2 Dopamine Signalling Fast excitatory or inhibitory synapses directly open ligand-gated ion channels and are activated
by neurotransmitters such as glutamate and GABA, respectively. Unlike these methods of transmission,
slow synaptic transmission is much more complex, and depends on altering intracellular levels of second
messenger molecules to produce cascading effects.
Dopamine receptors belong to a class of G-protein-coupled receptors (GPCRs) that trigger slow-
acting but long-lasting effects by changing the levels of cyclic adenosine monophosphate (cAMP) from
adenosine triphosphate (ATP) via adenylyl-cyclase (Kebabian, Petzold, & Greengard, 1972). The cAMP
signaling molecules activate protein kinase A (PKA) which goes on to phosphorylate several other
downstream targets (Walsh, Perkins, & Krebs, 1968). These phosphorylation cascades result in the
modulation of glutamate receptors, voltage-gated ion channels, and transcription factors. Of note, the
phosphorylation of the dopamine- and cAMP-regulated phosphoprotein (DARPP-32) leads to the

12
inhibition of the potent protein phosphatase 1 (PPP1CA), which is involved in the transfer of information
between the striatum and prefrontal cortex (Ivar Walaas, Aswad, & Greengard, 1983; Meyer-Lindenberg
et al., 2007).
1.3.2.3 Dopamine Receptor Subtypes The presence of dopamine receptors in the brain was first anticipated from the finding of
dopamine-sensitive adenylyl cyclase (AC; Kebabian et al., 1972), and 2 major dopamine receptors were
identified: D1 and D2 (Kebabian & Calne, 1979). Shortly after the successful cloning of the dopamine D1
receptor gene (Dearry et al., 1990) and the D2 receptor gene (Bunzow et al., 1988), three new dopamine
receptor subtypes were identified: the D3 receptor (Sokoloff et al., 1990), the D4 receptor (Van Tol et al.,
1991), and the D5 receptor (Sunahara et al., 1991), referred to as the DRD1, DRD2, DRD3, DRD4, and
DRD5, respectively.
The dopamine receptors have since been classified into two groups based on their amino-acid
sequence homology and their ability to stimulate or inhibit AC. The D1 and D5 receptors are grouped into
the D1-like receptor family and are coupled to the G protein Gs which activates AC, increasing
intracellular concentrations of cAMP. Conversely, the D2, D3, and D4 receptors are grouped in the D2-like
receptor family, and are coupled to the G protein Gi which inhibits AC, inhibiting the formation of cAMP.
As the dopamine subtypes exhibit diverse pharmacological profiles and expression levels in the brain,
they have been implicated in different functional roles, especially in the development of addiction.
1.3.3 Dopaminergic Pathways Four major dopaminergic pathways have been identified wherein dopamine is synthesized and
released: the nigrostriatal, mesolimbic, mesocortical, and tuberoinfundibular (Iversen & Iversen, 2007;
Marsden, 2006).
The nigrostriatal pathway originates in the dopamine-synthesizing neurons of the substantia
nigra pars compacta (midbrain nucleus) and projects to the dorsal striatum (caudate nucleus and

13
putamen), and is primarily associated with Parkinson’s disease. In this pathway, the D1 and D2 receptors
of the striatum are important in modulating simple and complex motor behaviour (Barnes et al., 2005).
In the mesolimbic pathway, dopaminergic cell bodies originate from the midbrain ventral
tegmental area (VTA), and connect to the basal forebrain, in particular, the ventral striatum (consisting
of the NAc and olfactory tubercle), amygdala, and frontal and limbic cortices (Heimer et al., 1997). The
NAc is predominantly composed of inhibitory medium spiny neurons, and once activated by dopamine,
they release GABA onto the ventral pallidum (VP). This pathway is generally tied to motivation between
affect and action (Mogenson, Jones, & Yim, 1980).
The mesocortical pathway also begins in the VTA but projects to different areas of the cortex
(medial, prefrontal, cingulate, and entorhinal cortex) and is largely populated by D1 receptors
(Sawaguchi & Goldman-Rakic, 1991). Activation of D1 receptors in the prefrontal cortex were found to
modulate spatial working memory (Williams & Goldman-Rakic, 1995). The mesolimbic and mesocortical
pathways are both associated with reward, learning, and schizophrenia, often jointly referred to as the
mesocorticolimbic pathway.
Finally, the tuberoinfundibular pathway originates from the arcuate and periventricular nuclei of
the hypothalamus, and via the hypophyseal portal system, dopamine released to the median eminence
reaches the intermediate lobe of the pituitary gland (Makara, Harris, & Spyer, 1972). This pathway is
important in the D2 receptor-mediated inhibition of prolactin. Accordingly, D2 receptor antagonism was
found to be the mechanism of action for antipsychotics due to the detection of altered plasma protein
levels (Johnstone et al., 1978).
Of the aforementioned pathways, the mesolimbic system is the most critically involved in the
positive reinforcing effects of drugs of abuse, developing, and maintaining addiction. In this circuit, the
dopaminergic cell bodies originate from the VTA, and connect to the basal forebrain consisting of the
nucleus accumbens, olfactory tubercle, amygdala, and frontal and limbic cortices.

14
Drugs of abuse primarily stimulate dopamine transmission from the VTA to the nucleus
accumbens and caudate (Imperato & Di Chiara, 1986; Imperato, Mulas, & Di Chiara, 1986). Once
addiction is established, dopamine levels increase in the nucleus accumbens even during initiation and
execution of drug-seeking behaviour (Phillips et al., 2003). It is proposed that this transmission is
responsible for encoding information about salient events, including sensitivity to rewarding and
aversive stimuli (Kapur, 2004; Reynolds & Berridge, 2002). Dopaminergic projections from the VTA to
the amygdala are imperative for reward-based, associative learning (Rosenkranz & Grace, 2002). And
together, both the accumbens and amygdala have been found to be associated with the reinforcing
effects of psychostimulants and the motivational aspects of drug withdrawal (Koob & Le Moal, 2001).
1.4 The Dopamine Hypothesis of Addiction Theories of addiction have established the importance of the mesocorticolimbic dopamine
system and extended amygdala; drugs of abuse stimulate dopamine release from the VTA neurons of
the midbrain to the NAc of the striatal complex. Although variations exist based on substance, a
common neuropathway involves dopaminergic projections from the VTA to the PFC and basolateral
amygdala (BLA), with both regions sending excitatory projections to the NAc, and finally the NAc sending
inhibitory projections to the VP. The following section will present a summary on the progression of
dopamine theories of addiction and relapse, as well as an expansion on the underlying neurobiology of
different reinstatement paradigms.
1.4.1 Hypotheses of Dopamine and Reward The early mesolimbic dopamine reward hypothesis of addiction postulated that drugs of abuse
activated reward sites that affected a common dopaminergic synapse, a theory that continues to shape
addiction research to this day. Although vaguely defined at the time, the fibers of the medial forebrain
bundle (MFB) and the VTA dopamine system of the midbrain were identified to be involved in the
reward system. And from the MFB, a group of dopaminergic neurons projecting from the VTA to the
NAc, now known as the mesolimbic pathway, emerged as a key component of the reward system. Early

15
drug studies supported this theory: amphetamine and cocaine directly increased dopamine levels in the
NAc, and alcohol was speculated to disinhibit noradrenergic inhibitory control over dopaminergic
neurons (Wise, 1980).
The subsequent psychomotor theory of addiction sought to synthesize addiction with operant
reinforcement, and asserted that a drug’s ability to induce psychomotor activation was indicative of its
abuse potential. Proponents of this theory argued that all drugs of abuse increased forward locomotion
via activation of the mesolimbic dopaminergic system, wherein a common mechanism was responsible
for both psychomotor stimulation and positive reinforcement (Wise & Bozarth, 1987). Although some
aspects of this theory have been disputed, it nevertheless generated an emphasis on the importance of
the mesolimbic dopamine system in the positive reinforcing effects of drugs.
Afterwards, the mesolimbic dopamine reward hypothesis of addiction was revisited and revised.
Instead of an emphasis on primary reward function, the theory shifted to place more importance on the
role of the mesolimbic dopamine system in habit formation and ‘craving’. This version argues that the
mesolimbic dopamine system is activated by both natural rewards and drugs of abuse (directly), and by
extension, their associated habits are consolidated by the same reinforcement processes (Wise, 2002).
This development was sparked by observations of non-human primates (NHP) that were trained
to respond to fruit juice. The naturally rewarding stimulus initially activated midbrain dopamine neurons
but over repeated testing, became less responsive to the actual reward, and more responsive to stimuli
that predicted reward (Schultz et al., 1993). This led to several conclusions: 1) midbrain dopamine
neurons were involved in arousal and motivational processes which controlled behavioural responses, 2)
activation of the midbrain dopamine system established reward-response habits, and 3) reward-
predictive stimuli produced the most arousal, thereby acting as conditioned rewards themselves.

16
1.4.2 Theories of Executive Function While mesolimbic dopamine hypotheses were being refined, the role of executive function in
addiction was becoming increasingly recognized. Theories related to executive function have centred on
the frontal cortex and its role in inhibitory response control, asserting that continued drug use leads to
frontal cortical dysfunction, progressively diminishing the capacity to inhibit reward-seeking behaviour.
A prevalent theory focusing on motivational circuits proposes that compulsive drug-seeking behaviour is
a product of a functional synergism between: 1) enhanced conditioned reward due to amygdalar
dysfunction, and 2) impaired inhibitory control driven by frontal cortical dysfunction (Jentsch & Taylor,
1999).
As the amygdala directs incentive learning and mediates the incentive value of conditioned
stimuli, amygdalar dysfunction is hypothesized to unduly facilitate the acquisition of stimulus-reward
associations and enhance the incentive properties of the drug and its associated stimuli. This process is
presumably caused by hyperactive dopaminergic projections from the VTA to the amygdala. With
regards to drug-seeking behaviour, two important amygdalar regions have been identified: the BLA and
the central nucleus of the amygdala (CeA). Excitotoxic lesion studies have demonstrated that the CeA is
involved in the potentiation of conditioned rewards while the BLA is involved in cue-induced
reinstatement (Meil & See, 1997; Robledo et al., 1996).
On the other hand, the impairment in modulating reward-related behaviour is presumably
driven by frontal cortical dopamine hypofunction. The frontal cortex is argued to regulate internal
motivational states to seek natural rewards via inhibitory control mechanisms as frontal cortical lesions
have led to cognitive impairments and deficits in inhibition (Milner, 1982). Although drug administration
studies found diverging levels of dopamine transmission in the frontal cortex depending on dose and
duration, it was concluded that dopaminergic dysregulation was responsible for the loss of inhibition in
reward-seeking behaviour (Jentsch & Taylor, 1999). Furthermore, dopaminergic hypofunction in the

17
frontal cortex was found to activate subcortical dopamine systems, as evidenced by locomotor activity
caused by the resulting NAc activation (Louilot et al., 1989; Tassin et al., 1978).
Brain imaging studies have further complemented the executive function theory. Abnormalities
found in the PFC and anterior cingulate gyrus of subjects with addiction led Volkow and colleagues to
hypothesize that a control circuit existed in drug addiction. They proposed that 4 interconnected circuits
were disrupted in addiction: 1) reward, 2) motivation, 3) memory and learning, and 4) control (Volkow
et al., 2003). Under normal circumstances, these circuits work concurrently to produce a balanced
output response but during addiction, the enhancement of reward, motivation, and learning are
presumed to overwhelm the inhibitory control exerted by the PFC.
Imaging studies localized the reward circuit to the NAc and VP as drugs of abuse acutely
increased extracellular dopamine in the striatum, and subjects that developed dependence exhibited
long-lasting decreases of striatal D2 receptors. Thus, it was deduced that the enduring decreases in
dopaminergic activity reduced the ability of natural reinforcers to stimulate reward circuits, increasing
susceptibility to drug-seeking in efforts to maintain previous levels of stimulation (Volkow et al., 2002).
The motivation circuit has been localized to the orbitofrontal cortex (OFC) of the PFC, and
subcallosal cortex as imaging studies have shown hyperactivity in those areas during intoxication and
presentation of drug-associated cues. The memory and learning circuit has been localized to the
amygdala and hippocampus, while the control circuit has been localized to the PFC and anterior
cingulate. In particular, it was demonstrated that disruption of the PFC impaired inhibitory control and
decision-making by increasing the tendency to choose immediate rewards over delayed rewards.
1.4.3 Theories of Relapse The aforementioned theories aim to characterize the mechanisms involved in the development
and maintenance of addiction, but the phenomenon of relapse appears to engage a different set of
processes. As addiction is established, drug-seeking behaviour becomes more dependent on

18
glutamatergic projections from the PFC to the NAc, and relapse susceptibility shifts from regulated to
compulsive as prefrontal regulation is diminished (Cardinal & Everitt, 2004). During regulated relapse, a
conscious decision is made involving active decision-making and declarative memory, whereas in
compulsive relapse, a conscious decision is absent, relying on procedural memories to guide behaviour
(Kalivas & O’Brien, 2007).
‘Craving’ is considered to be one of the primary motivating factors behind the precipitation of
relapse and is a crucial measurement for clinical relapse studies (Tiffany & Carter, 1998). Whereas
reinforcement is a process in which desirable or aversive stimuli are used to increase behavioural
responses, craving is the enduring physiological state and affect evoked by the stimuli that drives the
behaviour. ‘Positive craving’ refers to the motivation for the positive affect that accompanies positive
reinforcement, such as excitement and anticipation of a reward, and ‘negative craving’ refers to the
motivation for the removal of a negative affective state, such as anxiety, dysphoria, and depression
(Littleton, 2000).
Early preclinical studies of reinstatement often used cocaine for its highly positively reinforcing
properties and ability to evoke positive craving. Alcohol, on the other hand, is less directly reinforcing
but is still able to induce both positive and negative reinforcing pharmacological effects, producing both
types of craving. Thus, it should be taken into consideration that findings from early drug-conditioning
models using cocaine may not necessarily transfer over to the study of alcohol.
Contributing to the complexity of relapse, reinstatement can be induced by the drug itself (drug-
prime), drug-associated cues, or stressors, each with varying neurocircuitry. But despite these variances,
all priming stimuli are hypothesized to share a final common neuropathway of the medial prefrontal
cortex (mPFC) as an input, and the NAc core as an output (Kalivas & McFarland, 2003). Ultimately,
dopamine release in the NAc is considered to be the final event that triggers relapse to drug-seeking
behaviour by all reinstating stimuli (Self & Nestler, 1998).

19
1.4.3.1 Drug-Induced Reinstatement Reinstatement of drug-seeking can be induced by administering pharmacological agents that
elicit equivalent experiences to the initial drug of abuse. Although drug-induced reinstatement has been
demonstrated using a variety of substances, studies have primarily focused on cocaine due to its robust
effects (Lê et al., 1998; Self & Nestler, 1998). Cocaine-induced reinstatement is purportedly mediated by
a neurocircuit consisting of the dorsal prefrontal cortex (dPFC), NAc core, and VP, but not the BLA, and is
initiated by dopamine release in the dPFC (Grimm & See, 2000).
Cocaine-induced reinstatement has been elicited by systemic injections of D2-like receptor
agonists, direct infusions of cocaine and dopamine into the NAc and mPFC, and by dopamine agonists in
the NAc shell (McFarland & Kalivas, 2001; Tran-Nguyen et al., 1998). Reinstatement has also been
induced by glutamatergic activation of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)
receptors in the NAc, as well as NMDA receptors in both the NAc and VTA dopamine neurons, further
demonstrating a shared neurocircuit with the PFC (Cornish & Kalivas, 2000).
Conversely, cocaine-induced reinstatement is seemingly inhibited through the opposite
processes: mPFC and NAc shell injections of dopamine receptor antagonists, inactivation of dopamine
neurons in the VTA by GABAergic agonists, and reversible inactivation of the VP and NAc. However,
there have been contradictory findings regarding whether inactivation of the NAc shell or core blocks
cocaine-induced reinstatement (Shaham et al., 2003).
Interestingly, drugs from different classes have been able to produce a ‘cross-priming’ effect as
they innervate the same mesolimbic dopamine circuit: opiates are able to reinstate responding in
animals that self-administer psychostimulants, and the infusion of amphetamine into the NAc is able to
reinstate heroin-seeking behaviour (de Wit & Stewart, 1981; Stewart & Vezina, 1988).
1.4.3.2 Cue-Induced Reinstatement An arguably more complex means of inducing reinstatement is through re-exposure to
environmental cues. Initially, drug use induces transient neuroplastic changes corresponding with the

20
acquisition phase, an associative learning process that pairs a neutral stimulus with a drug, and
establishes a behavioural response to the stimulus. Through repeated drug use, the neuroplastic
changes gradually become stable and the previously neutral stimulus develops its own reinforcing
properties and becomes a CS (Stewart et al., 1984). Following abstinence, the CS can evoke drug-craving
and physiological arousal in the absence of a primary reinforcer, perpetuating drug-seeking behaviour
and relapse (See et al., 2003).
Cue-induced reinstatement is proposed to be dependent on dopaminergic innervation of the
BLA and the mPFC, innervating glutamatergic projections to the NAc core. The BLA complex plays a
crucial role in cue-induced reinstatement and is necessary for the acquisition of associative learning with
drug-paired stimuli. Cue-induced reinstatement appears to be mediated by dopaminergic inputs to the
BLA as intra-BLA infusions of D1-like receptor antagonists are able to block reinstatement (See et al.,
2001). Furthermore, excitotoxic lesions and reversible inactivation of the BLA significantly attenuates
cue-induced reinstatement without affecting drug self-administration or drug-primed reinstatement
with cocaine (Meil & See, 1997). Cue-induced reinstatement is also inhibited through the inactivation of:
the anterior cingulate, VTA, NAc, and the lateral OFC (Di Ciano & Everitt, 2004; Weiss et al., 2000).
On the other hand, stimulating dopamine release in the BLA using amphetamines was able to
potentiate responding in the presence of a CS. This supplements the idea that dopamine release in the
amygdala plays a critical role in cue-induced drug-seeking behaviour; dopamine may be able to enhance
the salience of drug-paired stimuli by driving the retrieval and utilization of those associations (See et al.,
2003). Interestingly, inactivation of the NAc attenuated cocaine self-administration without affecting
conditioned lever responding lending support that the BLA but not the NAc is essential for cue-induced
reinstatement (Grimm & See, 2000).
Although it is reasonable to expect that the ability of drug-related cues to reinstate drug-seeking
would gradually diminish over time, an opposing pattern is seen. A delayed-onset craving syndrome,

21
termed ‘incubation’, progressively strengthens the ability of drug-related cues to induce reinstatement
for a period as abstinence length increases, and remains high for extended periods (Grimm et al., 2001).
Signaling pathways in the VTA and neuronal activation of the CeA have been implicated in this
incubation process (Pickens et al., 2011).
1.4.3.3 Stress-Induced Reinstatement Stress is an adaptive mechanism that prepares organisms for new environmental challenges,
however, it becomes maladaptive when it increases vulnerability to drug abuse and relapse after
periods of abstinence (Sinha, 2001). In animal studies, exposure to stressful stimuli (often electric foot-
shock pulses) are used to induce reinstatement (Weiss et al., 2001).
Stressful events typically activate the hypothalamic-pituitary-adrenal (HPA) axis by initiating the
secretion of corticotropin-releasing factor (CRF) from the paraventricular nucleus (PVN) of the
hypothalamus. The neuropeptide hormone CRF stimulates the synthesis of adrenocorticotropic
hormone (ACTH) from the anterior pituitary gland, which then increases the production and release of
cortisol from the adrenal gland. This signaling cascade produces a stress response that includes the
suppression of the immune system, and increases in metabolism, blood sugar, and blood pressure.
However, it has been identified that CRF neurons in the CeA, independent of the HPA axis, are
responsible for mediating stress-related drug-seeking behaviour as well as the anxiogenic and stress-like
symptoms experienced during withdrawal from drugs (Dunn & Berridge, 1990; Koob et al., 1994). During
stress exposure, CRF is released into the CeA, containing a high concentration of CRF receptors (Shekhar
et al., 2005).
Stressful stimuli activate glutamatergic projections from the PFC to VTA while stress-induced
corticosterone secretions also activate VTA neurons, triggering VTA dopamine release in the NAc (Self &
Nestler, 1998; Shaham et al., 1996). Stress-induced reinstatement also involves adrenergic projections
from the lateral tegmental nucleus (LTN) to the extended amygdala, in particular the CeA and lateral bed
nucleus of stria terminalis (BNST).

22
During acute alcohol withdrawal, extracellular CRF levels are substantially elevated in the CeA of
rats, and so administration of CRF antagonists have been shown to decrease the reinstating effects of
stress (Pich et al., 1995). In addition, stress-induced reinstatement has also been prevented by the
inactivation of the CeA, BNST, PFC, VTA, and NAc (Kalivas & McFarland, 2003; McFarland & Kalivas,
2001). Stress-induced reinstatement has also been attenuated by injections of noradrenergic
antagonists into both the CeA and lateral BNST, as well as CRF antagonists into the lateral BNST, but not
the BLA (Leri et al., 2002; Shaham et al., 2000).
1.4.4 Synthesis and Challenges of the Dopamine Hypothesis As a synthesis of the literature, Koob and Le Moal (2006) have discerned three general
neurocircuits: a drug-reinforcement circuit, a drug- and cue-induced reinstatement circuit, and a drug-
seeking circuit. The drug-reinforcement circuit is primarily involved with reward and stress. The acute
reinforcing effects of drugs are modulated by dopaminergic projections from the VTA to the NAc, but
involve many other brain areas including the extended amygdala (CeA and BNST), arcuate nucleus of the
hypothalamus, and ventral striatal-ventral pallidal-thalamic-cortical loops. Furthermore, stress is
mediated by CRF and NE projections between the extended amygdala and the pontine nuclei of the
brainstem, and is responsible for the integration of rewarding or aversive stimuli. A combination of a
reduction of function in the extended amygdala reward system and engagement of stress neurocircuitry
are proposed to give rise to the effects of withdrawal and stress-induced relapse.
The drug- and cue-induced reinstatement circuit is presumed to evoke craving and consists of
afferent projections from the PFC (anterior cingulate, prelimbic cortex, and orbitofrontal cortex) and BLA
to the extended amygdala and NAc. More specifically, a critical role exists for the PFC in drug-induced
reinstatement, and the BLA in cue-induced reinstatement. Lastly, the drug-seeking circuit controls
behavioural output, and dysfunction leads to compulsivity. In this circuit, motivation is processed

23
through the extended amygdala reward pathway, and translated into action via the NAc, VP, thalamus,
and motor cortex (Koob & Le Moal, 2006).
Thus far, dopamine transmission has been associated with a variety of rewarding activities
ranging from gambling, eating, gaming, and placebo responses, further supporting the dopamine theory
of addiction. Additionally, dopamine release is alleged to be responsible for the euphoric effect caused
by substances of abuse. With the onset of radiotracer imaging techniques such as 11C-raclopride
positron emission tomography (PET) and single-photon emission computer tomography (SPECT), studies
were able to indirectly measure in vivo dopamine release in the human striatum, and found that drug-
induced dopamine increases were correlated with increased ratings of euphoria (Laruelle et al., 1995).
This led to the interpretation that euphoria was mediated by the release of dopamine, and that any drug
inducing dopamine release had the potential risk of being abused (Nutt et al., 2015).
Studies have since established that striatal dopamine receptor availability and dopamine release
are reduced in individuals with stimulant or alcohol use disorders (Volkow et al., 2007), attributed to a
downregulation of postsynaptic dopamine receptors. However, these striatal changes have been
inconsistent with other substances such as nicotine and opiates (Martinez et al., 2012; Scott et al.,
2006).
These findings raise an issue with the dopamine hypothesis of addiction: if striatal dopamine is
responsible for pleasure, then lower receptor availability should diminish this experience. However, the
opposite effect occurs in which those with lower levels of striatal dopamine receptors report more
pleasurable effects. Further complicating the matter, increasing dopamine receptor levels in the ventral
striatum of dependent rats reduces drug intake (Thanos et al., 2005). Thus, it should be taken into
consideration that a unifying dopamine theory of addiction may be much more complex, and that other
neurotransmitters may play a greater role than previously thought.

24
1.5 The Dopamine D4 Receptor 1.5.1 The Dopamine D4 Receptor and Antipsychotics
Although the D3 and D4 receptors belong to the D2-like receptor class, studies have often
misattributed their findings to the D2 receptor itself due to its abundance and distribution in reward
structures (Marsden, 2006). And relative to other D2-like receptors, fewer studies have examined the D4
receptor. A contributing factor for this has been the absence of truly selective compounds for the D4
receptor, and so most early studies relied on the usage of D4 knockout mice. But the motivation to
increase the development of selective D4 receptor ligands was partly due to the initial success of the
antipsychotic clozapine.
Typical antipsychotics act through D2 receptor blockade, but are prone to treatment-resistance
and adverse effects. In 1971, clozapine, the first atypical antipsychotic, was shown to be effective for
treatment-resistant schizophrenia, and more importantly, had reduced extrapyramidal symptoms.
Clozapine treatment also reduced alcohol use in schizophrenia patients with comorbid substance abuse
(Marcus & Snyder, 1995). The efficacy of clozapine was hypothesized to be attributed to its ability to
block the D4 receptor (Seeman & Van Tol, 1994), and so the development of the selective D4 receptor
antagonist L-745,870 was highly anticipated to also produce antipsychotic effects.
Unfortunately, clinical experiments using L-745,870 on patients with acute schizophrenia were
ineffective (Kramer et al., 1997). In order to determine if the efficacy of clozapine was due to anxiolytic
properties modulated by D4 receptors, the elevated plus-maze (EPM) test, which measures anxiety
based on an animal’s aversion to open spaces, was utilized. Once again, L-745,870 produced no
significant behavioural changes in mice, suggesting that D4 receptor antagonism does not modulate
anxiety-related behaviours (Cao & Rodgers, 1997). Nevertheless, the initial attention incited by
clozapine shifted research focus towards the D4 receptor, giving rise to numerous investigations on its
specific functions and polymorphisms. Further behavioural studies of the D4 receptor have also
demonstrated potential therapeutic effects in the realm of addiction, which will be discussed later.

25
1.5.2 D4 Receptor Distribution While the D2 and D4 receptors share similar distributions, D4 receptor expression is significantly
lower (Rondou et al., 2010). Due to the historical absence of selective ligands, D4 receptors were difficult
to identify and localize. But through technical advancements, D4 receptor mRNA have been detected in
the retina, cerebral cortex, amygdala, hypothalamus, pituitary, and striatum via Northern blot and RT-
PCR analyses (Cohen et al., 1992; Matsumoto et al., 1995; Van Tol et al., 1991), in situ hybridization
(Meador-Woodruff et al., 1997), and ligand binding (Defagot et al., 2000; Primus et al., 1997). D4
expression levels were highest in the retina, followed by the PFC, hippocampus, amygdala, and
hypothalamus, with low expression levels in striatal areas. Further confirmatory immunohistochemistry
studies localized the D4 receptor to GABAergic neurons in the PFC, hippocampus, substantia nigra pars
reticula, and globus pallidus (Khan et al., 1998; Mrzljak et al., 1996; Rivera et al., 2002). In the striatum,
D4 receptors are mainly expressed in the dendritic shafts and spines of post-synaptic neurons (Rivera et
al., 2002). D4 receptors have also been found outside of the CNS in the cardiac atrium, lymphocytes, and
kidney (Bondy et al., 1996; O’Malley et al., 1992; Seamans & Yang, 2004).
While D4 receptors have been detected in numerous brain regions, the most notable are the
PFC, amygdala, and hippocampus, for their implication in cognition and the development and
maintenance of addiction. The amygdala and hippocampus are responsible for incentive learning and
memory, whereas the PFC is involved in inhibitory response control. Thus, the presence of D4 receptors
in these areas is indicative of possible involvement in these processes.
1.5.3 D4 Receptor Genetics and Polymorphisms The human DRD4 is located on chromosome 11, contains four exons, and is flanked by two CpG
islands, one on each of the 5’ and 3’ regions; the DRD4 promoter is located in the CpG islands of the 5’-
region (Kamakura et al., 1997). The transcription initiation site and promoter sequences are located
within 400-500 base pairs (bp) upstream from the translation initiation codon, with the promoter being
highly rich in guanine and cytosine nucleotides (Van Tol et al., 1992).

26
Numerous polymorphisms have been associated with the DRD4: a single nucleotide
polymorphism (SNP) in the promoter region (C-521T), a 12-bp tandem duplication in exon 1, a 13-bp
deletion of bases 235 to 247 near the 3´ end of exon 1, and a substitution of valine by glycine at the
beginning of exon 3 (Catalano et al., 1993; Nöthen et al., 1994).
The most notable DRD4 polymorphism is a variable number tandem repeat (VNTR) found in the
third exon in the region that encodes the third intracellular loop of the receptor (Van Tol et al., 1992).
The VNTR consists of a 48-bp sequence, coding for 16 amino acids, in which alleles have been identified
with anywhere from 2 to 11 (32 to 176 amino acids) repeats (denoted as D4.2 to D4.11). As a result of
these repeats, the VNTR is responsible for encoding up to 20 different protein variants of the receptor
(Asghari et al., 1994). Allele variants with less than 7 repeats are referred to as ‘short alleles’ (S-allele),
and those with greater than 7 repeats are referred to as ‘long alleles’ (L-allele). The most predominant
DRD4 VNTR allele variants are the D4.4, D4.7, and D2.2, occurring at global mean frequencies of 64.3%,
20.6%, and 8.2%, respectively (Chang et al. 1996).
1.5.4 D4 Receptor Signalling As a member of the D2-like receptor subfamily, the D4 receptor exhibits a pharmacological
profile similar to that of the D2 and D3 receptors. The D4 receptor is most potently activated by the
endogenous ligand dopamine, but can also be activated by epinephrine and norepinephrine, albeit with
5-fold lower potency (Lanau et al., 1997).
The D4 receptor is coupled to the G protein αi/o subunits, and when activated, inhibits AC which
reduces the formation of the second messenger cAMP. Compared to the D4.2 and D4.4 variants, the D4.7
has a two-fold reduced potency for dopamine to inhibit cAMP formation (Asghari et al., 1995), whereas
the D4.10 variant is two-fold more potent (Jovanovic et al., 1999).
Depending on the cell type, the D4 receptor is able to influence intracellular calcium levels.
Activation of the D4 receptor has been reported to lead to phosphoinositide phospholipase C-γ (PLCγ)

27
activation, releasing calcium in CA1 hippocampal neurons (Pangalos & Davies, 2002), as well as
activation of Ca2+/calmodulin-dependent protein kinase (CaMKII) which in turn influences AMPA
receptors (Lane et al., 2008).
The D4 receptor is also reported to interact with G protein-coupled potassium inwardly
rectifying channels (Kir3) that hyperpolarize cells and inhibit their firing rates (Lavine et al., 2002). In
comparison to the D4.4 variant, dopamine is five-fold more potent on D4.2 and D4.7 (Wedemeyer et al.,
2007). Taking into account the biochemical differences found in the D4 receptor allelic variants suggests
that therapeutic drugs targeting the D4 receptor may have varying efficacy in individuals.
Furthermore, D4 receptors expressed in GABAergic neurons of the PFC have been found to
regulate GABAA receptor current and trafficking. D4 agonists inhibited postsynaptic GABAA receptor
currents in PFC pyramidal neurons, an effect that was blocked by D4 antagonists (Wang et al., 2002). In
another study, D4 receptor activation led to actin depolymerization, attenuating the transport of GABAA
receptors to the plasma membrane, resulting in reduced functional GABAA receptor density and current
(Graziane et al., 2009). As GABAA receptor-mediated inhibitory transmission in the PFC is implicated in
working memory, D4 receptors appear to mediate this process.
1.5.5 D4 Receptor VNTR Polymorphism At the molecular level, the VNTR polymorphism has been associated with variations in D4
receptor expression, ligand binding, and cAMP formation (Asghari et al., 1995; Cohen et al., 1992). These
variations are purported to have much larger implications as the VNTR has been linked to addictive and
mental disorders, as well as differences in personality traits.
As it is unlikely for the development of complex disorders to be caused by one specific gene, and
more likely caused by a combination of genetic and environmental factors, using the approach of
identifying a behavioural phenotype related to the overarching disorder has allowed for increased
statistical power to detect associations with particular candidate genes.

28
With this approach in mind, cue-elicited craving for tobacco was used as a phenotype to
examine the effects of the VNTR on nicotine dependence. Smokers that were heterozygous or
homozygous for L-alleles demonstrated greater craving, arousal, and attention to smoking cues in
comparison to those with S-alleles (Hutchison et al., 2002a). These findings were later corroborated by
functional magnetic resonance imaging blood-oxygen-level-dependent (fMRI-BOLD) responses to
smoking cues in nicotine dependent adults (McClernon et al., 2007).
Similar findings were discovered with alcohol as participants that were homozygous or
heterozygous for L-alleles displayed significantly higher craving after alcohol consumption in comparison
to the control beverage (Hutchison et al., 2002b). Conversely, genotyping via Southern blot and PCR
analysis was used to determine that the S-alleles, D4.3 and D4.6, were 3- and 4-fold more prevalent,
respectively, for those with alcohol addiction when compared to control populations (George et al.,
1993).
Furthermore, the D4.7 allele was found to be highly associated with the personality trait of
novelty-seeking and attention-deficit hyperactivity disorder (ADHD), accounting for a minimum of 25-
50% of observed cases (Ebstein et al., 1996; Grady et al., 2003). The association between the D4 receptor
and novelty-seeking is particularly interesting due to its implications in drug abuse and cognition.
Indeed, the personality trait of novelty-seeking has been found to mediate the association
between the D4 receptor and heavy drinking behaviour in D4.7 allele carriers (Laucht et al., 2007).
Carriers of the D4.7 allele reported drinking higher amounts of alcohol in one occasion and greater heavy
drinking activity than those without, and scored higher on novelty-seeking as assessed by the
Temperament and Character Inventory (TCI). Although novelty-seeking was also associated with
drinking, once this trait was taken into account, the effect of the D4.7 allele on drinking behaviour
became nonsignificant, meeting the conditions for mediation. In concordance with the novelty-seeking

29
trait, the D4.7 allele was also associated with higher measures of sexual desire, arousal, a wider variety of
sexual behaviors, and promiscuity (Halley et al., 2016).
However, some of these findings have been challenged as another study found no evidence of
the VNTR polymorphism being linked to novelty-seeking, alcoholism, or smoking (Luciano et al., 2004). A
subsequent meta-analysis also did not find any associations between the VNTR and novelty-seeking,
although the C-521T SNP of the D4 receptor appeared to account for phenotypic variance on novelty-
seeking and impulsivity traits (Munafò et al., 2008).
1.6 Role of the D4 Receptor in Addiction 1.6.1 Early D4 Behavioural Studies
Early efforts to clarify the role of the D4 receptor in vivo utilized genetically modified mice, in
which the D4 receptor gene was ‘knocked-out’. Although complete null allele mutants have yet to be
produced, D4 knockout mice were generated by using recombinant clones lacking exon II of the D4 gene
(Rubinstein et al., 1997). The removal of exon II resulted in the splicing of exon 1 and 3, causing a
reading frame shift and introducing a premature stop codon during translation. By disrupting the
expression of the D4 receptor protein, it may be possible to disentangle its functional role from any
remarkable behavioural effects.
Rubinstein and colleagues (1997) observed that D4 knockout mice exhibited reduced overall
locomotor activity and sensitivity to alcohol, cocaine, and methamphetamine. Although the D4 knockout
mice displayed reduced spontaneous locomotion and rearing activity in both novel and familiar open
field environments, they significantly outperformed wildtype mice on the rotarod, a performance test
measuring the ability to sustain complex coordinated movements over time.
The authors hypothesized that the reduced locomotion was either caused by the absence of D4
receptors during development, resulting in irregular dopamine signaling or the lack of D4 receptors in
limbic regions, reducing motivation to explore the environment (Rubinstein et al., 1997). Interestingly,
the biosynthesis and metabolism of dopamine in the dorsal striatum (caudate and putamen) was

30
significantly increased in D4 knockout mice. The authors surmised that the elevated striatal dopamine
levels may have contributed to the enhanced performance of the mutant mice on the rotarod.
Since drugs of abuse alter dopamine levels in the brain, Rubinstein et al. (1997) investigated the
effects of various drugs on D4 knockout mice in a spontaneous horizontal locomotion test. They found
that locomotor stimulation by alcohol, cocaine, and methamphetamine, was much greater in D4
knockout mice relative to wild-type littermates. As D4 receptors regulate GABA receptor current and
trafficking, the authors suggested that the locomotor sensitivity seen in D4 knockout mice was due to a
disruption in balance between the D4 receptor-mediated inhibitory tone and the D1-like receptor-
mediated excitation. Taken together, these findings from D4 knockout mice suggest that the D4 receptor
is involved in modulating various locomotor behaviours, as well as neurotransmission between the
cortex, basal ganglia, and thalamus (Rubinstein et al., 1997).
A study examining the influence of D4 receptors on novelty-seeking found that D4 knockout mice
were less responsive to novelty in open field, emergence, and novel object tests (Dulawa et al., 1999).
The greatest difference was found in the novel object test, a free exploration paradigm that measures
an animal’s tendency to approach novel stimuli in a familiar environment. D4 knockout mice spent
significantly less time exploring the novel object, indicating that the D4 receptor does indeed play a role
in modulating approach behaviour during exploration.
Expanding upon these findings, the C57BL/6J mouse strain exhibited increased exploration of a
novel object when administered with the partial D4 receptor agonist RO-10-5824 without any effects on
locomotor activity in either novel or familiar environments (Powell et al., 2003). The significance of
these results is the indication that D4 receptor stimulation can enhance novelty-seeking, and by
extension, an increased susceptibility to drug-taking behaviour. Although difficult to generalize to
humans, these findings do however complement genetic association studies linking the D4 VNTR to the
personality trait of novelty-seeking.

31
1.6.2 Addiction-Relevant Neuroadaptations of the D4 Receptor Of relevance to this thesis are the neuroadaptations of the D4 receptor induced by alcohol.
Repeated administration of alcohol has been reported to potentiate drug-induced psychomotor
activation, or behavioural sensitization, in mice (Masur et al., 1986). This is hypothesized to reflect
sensitization to the motivational properties of alcohol, contributing to increased susceptibility to
addiction and relapse. However, species and strain seem to play a role in the activating effects of alcohol
and behavioural sensitization. Although chronic alcohol exposure in Swiss mice showed an increase in
locomotor activity, no such effects were seen in male Wistar rats (Masur et al., 1986). In fact, Wistar rats
exhibited decreased locomotor activity to acute alcohol treatment, but developed tolerance soon
afterwards, displaying similar activity levels to the control group over chronic treatment.
In order to further examine the effects of alcohol-induced behavioural sensitization on the D4
receptor, D4 receptor binding levels were assessed via quantitative autoradiography using the
radioligand [3H]nemonapride, a selective antagonist for D2-like receptors, and raclopride, a selective
antagonist for D2 and D3 receptors to prevent binding to sites other than D4 receptors. Dopamine D4
receptor binding levels were found to be increased in striatal and limbic areas of mice exposed to
chronic alcohol treatment (Quadros et al., 2005). Although locomotor activity tests were used to
categorize mice as ‘sensitized’ or ‘non-sensitized’, both groups showed higher D4 binding densities than
saline controls in the posterior caudate-putamen and olfactory tubercle. However, only sensitized mice
had higher D4 binding at the lateral septal nucleus. Thus, increased D4 binding levels in alcohol-treated
groups suggest that repeated alcohol administration induces the upregulation of D4 receptors in specific
brain regions, generating greater sensitization to the effects of alcohol. As the upregulated brain regions
are components of the mesolimbic dopaminergic pathway, the role of the D4 receptor is further
implicated in the drug addiction process.

32
1.6.3 The Role of D4 Receptors in Gambling Disorders D4 receptors have also been implicated in the decision-making processes of gambling, an
addictive disorder. In order to determine the neurochemical processes underlying gambling, animal
models emulating human gambling-related decision-making have been developed.
A recently developed model is the rat gambling task (rGT), an analogue of the Iowa gambling
task, in which the optimal strategy to yield the greatest gains is to avoid high-risk high-reward options,
and pursue low-risk low-reward options. Using the rGT, the D2-like antagonist eticlopride was found to
significantly enhance the choice of the optimal option (Zeeb et al., 2009).
Another model named the rodent slot machine task (rSMT) focuses on the ‘near-miss’ effect in
which gamblers perceive certain losing outcomes as being closer to a win than others (i.e. two lucky 7’s
instead of three). Due to this perception, near-misses disproportionately heighten expectations of
reward, and induce similar physiological responses as winning outcomes, further propagating gambling
behaviours (Griffiths, 1991). In the rSMT, a collect lever press results in either a large reward in response
to the illumination of 3 lights (winning outcome), or a time penalty in response to losing outcomes
indicated by 2, 1, or 0 lights, referred to as near-miss, near-loss, or clear-loss trials, respectively. During
baseline behavioural testing in the rSMT, rats consistently demonstrate a preference for the collect lever
on 2-light loss trials, analogous to the near-miss effect (Cocker et al., 2014).
Contrary to previous rGT findings, eticlopride had no effects on rSMT performance whereas the
D2-like receptor agonist quinpirole dose-dependently increased erroneous collect responses, with a
pronounced effect on the 2-light loss trials (Winstanley et al., 2011). However, it should be noted that all
doses of quinpirole and high doses of eticlopride increased response latency on the collect lever,
regardless of trial type. While motor activity may be reduced, the results of this study suggest that D2-
like receptors mediate reward expectancies, and that enhanced activation may impair the ability to
discriminate between salient and irrelevant information.

33
Further examination with selective D2 and D3 ligands yielded no effects on the rSMT, but the
selective D4 agonist PD 168,077 impaired performance, similar to the effects of quinpirole. Importantly,
the selective D4 antagonist L-745,870 improved rSMT performance, particularly on non-win trials where
the last light was off, and when co-administered with quinpirole, attenuated its impairing effects,
returning performance to near baseline levels (Cocker et al., 2014). Thus, this study demonstrates that
D4 receptors, rather than D2 or D3 receptors, are specifically responsible for mediating erroneous
expectations of reward in the rSMT. The impaired choice behaviour induced by D4 activation is
hypothesized to be caused by an enhancement in salience of reward signals, while the effects of D4
receptor blockade may be due to either a reduction in the salience of putative win signals, or an
increase in the salience of non-reward signals counteracting heightened reward expectations.
A return to the rGT sought to identify the dopamine receptor subtypes responsible for the
previously mentioned enhanced performance induced by eticlopride. Although none of the selective D3
or D4 ligands affected decision-making under the rGT, the D4 ligands did however affect latency
measures (Di Ciano et al., 2015). The D4 antagonist L-745,870 dose-dependently increased collect
latency while a decrease was seen with high doses of the D4 agonist PD 168,077. Despite the lack of
support for selective D2, D3, or D4 ligands on decision-making, the change in collect latency suggests a
possible effect on motivation to accept rewards.
1.6.4 The D4 Receptor and Alcohol 1.6.4.1 Mechanisms of Action of Alcohol and Dopamine
As the most relevant substance of abuse to this thesis is alcohol, this section will delve further
into its pharmacology and involvement of dopamine. Alcohol interacts with both excitatory and
inhibitory systems in the CNS to produce the characteristic effects of elevated mood, euphoria,
anxiolysis, and sedation. Alcohol mainly exerts its effects by binding to GABAA receptors as a positive
allosteric modulator (PAM), increasing the effects of the inhibitory neurotransmitter GABA. The binding

34
of GABA to GABAA receptors triggers the opening of chloride channels, allowing the entry of chloride
anions to hyperpolarize the postsynaptic neuron, decreasing its firing rate. As a PAM, alcohol
potentiates the effects of GABA by either increasing the frequency or duration of the opening of
chloride channels.
Secondarily, alcohol acts as a negative allosteric modulator (NAM) of NMDA and AMPA
receptors, inhibiting the excitatory effects of glutamate (Davies, 2003). Glutamatergic activation of
NMDA receptors results in the opening of non-selective cation channels; the influx of sodium and
calcium ions depolarizes the neuron.
As discussed in previous chapters, alcohol has been found to stimulate dopamine release in the
NAc. However, direct exposure of moderate concentrations of alcohol to dopamine terminals in the rat
NAc had no effects on dopamine release or uptake, and high concentrations of alcohol decreased
dopamine levels (Budygin et al., 2001). As with many substances of abuse, dopamine release in the NAc
occurs through the activation of dopaminergic neurons in the VTA. Indeed, alcohol was found to have
direct excitatory effects on rat dopamine VTA neurons in vitro (Brodie et al., 1990).
Subsequent investigations determined that alcohol reduces the current of delayed rectifier K+
channels (Appel et al., 2003; Brodie et al., 1999). Normally, the efflux of K+ ions are responsible for the
repolarization and hyperpolarization phase of the action potential, and when hyperpolarized, the
neuron is unable to generate subsequent action potentials, termed as the refractory period. However,
by decreasing the efflux of K+ ions, the amplitude of the hyperpolarization is reduced, which circumvents
the refractory period, further propagating the spontaneous firing of dopamine VTA neurons.
In addition, alcohol indirectly influences VTA dopaminergic activity through GABAA receptors.
Low doses of alcohol have been reported to stimulate GABAA receptors on GABAergic interneurons in
the VTA, disinhibiting dopaminergic activity (Mereu & Gessa, 1985). Conversely, high doses of GABAA
agonists inhibited dopaminergic VTA cells (Westerink et al., 1996). Based on these findings, it has been

35
argued that GABAA receptors are partially responsible for the stimulant effects induced by low doses of
alcohol, and depressant effects elicited by high doses (Pierce & Kumaresan, 2006).
1.6.4.2 The D4 Receptor and Alcohol In order to determine the behavioural effects of dopamine on alcohol, this section will
summarize relevant findings. Once again, the initial lack of selective D4 receptor ligands have hindered
the examination of its specific behavioural effects. Nonetheless, early studies using D2-like receptor
ligands may still be relevant due to their potential effects through the D4 receptor.
For instance, the anxiolytic buspirone, an agonist of the serotonin subtype 5-HT1A receptor was
reported to reduce alcohol consumption in Macaque monkeys (Collins & Myers, 1987). But radioligand
binding studies have confirmed that buspirone also has high affinity for the human D4 receptor, acting as
an antagonist (Bergman et al., 2013), thus alluding to a possible effect on addiction via the D4 receptor.
In a study using oral self-administration of alcohol in rats, both the D2-like agonist and
antagonist (apomorphine and pimozide, respectively) decreased total responding, but produced
different patterns of responding; apomorphine dose-dependently decreased responding at the onset of
the session while pimozide had no effect on early responding but exhibited an earlier cessation of
responding (Samson et al., 1990).
In mice, alcohol produces dose-dependent locomotor activity (Risinger et al., 1994) and
conditioned taste aversion (Sherman et al., 1988), and when paired with environmental cues, results in a
conditioned place preference (Risinger & Oakes, 1996). The blockade of D4 receptors by clozapine
eliminated alcohol-stimulated locomotor activity, but did not influence place- or taste-conditioning
(Thrasher et al., 1999). And as mentioned earlier, the study by Rubinstein and colleagues (1997)
demonstrated that D4 knockout mice exhibited greater locomotor stimulation by alcohol. Since D4
receptor activation has been implicated in the downregulation of GABA receptors, it has been suggested
that the locomotor sensitivity may be caused by a lack of D4 receptor-mediated inhibitory tone. From

36
these findings, it appeared as if D4 receptors were only involved in the stimulant properties of alcohol,
but not the motivational or aversive effects of alcohol.
In another study, male Wistar rats were found to self-administer alcohol directly into the
posterior VTA via stereotactic implantation of guide cannulas. Administration of the D2-like agonist
quinpirole significantly reduced alcohol self-administration (measured by lever presses), and the co-
administration of the D2-like antagonist sulpiride induced reinstatement of self-administration
behaviours (Rodd et al., 2004). These results indicated that alcohol did in fact have reinforcing effects
within the posterior VTA.
1.6.5 The D4 Receptor and Other Substances of Abuse 1.6.5.1 Stimulants Amphetamine
As with individuals with ADHD, amphetamine administration reduces locomotor activity in
hyperactive mouse mutants. L-745,870 induced a significant overall reduction in amphetamine-
mediated locomotor activity in both mutant and control mice without affecting baseline activity in
control mice but did not appear to modify amphetamine-mediated locomotor activity (Fan et al., 2010).
In rats trained to discriminate amphetamine from saline, the administration of D4 agonist WAY-100,635
potentiated drug appropriate responding, whereas L-745,870 partially blocked the discriminative
stimulus effect without disruption (Marona-Lewicka & Nichols, 2011). Thus, it seems that the D4
receptor plays a role in modulating the interoceptive effects induced by substances of abuse.
Cocaine
Drug discrimination studies found L-745,870 to be ineffective for cocaine responding (Costanza
& Terry, 1998). Although selective D2 antagonists dose-dependently increased cocaine self-
administration, no significant influence was seen with L-745,870 (Caine et al., 2002). Thus, D4 receptors
do not appear to mediate the reinforcing effects of cocaine. Since cocaine inhibits the reuptake of

37
dopamine by blocking dopamine transporter proteins, it seems that D4 receptors do not directly
influence these mechanisms.
Nicotine
While D4 receptor blockade by L-745,870 had no effect on nicotine self-administration, it
significantly reduced both cue- and nicotine-induced reinstatement of nicotine-seeking behaviour,
without influencing reinstatement of food-seeking behaviour (Yan et al., 2012). However, the activation
of D4 receptors by PD 168,077 did not reinstate extinguished nicotine- or food-seeking behaviour (Yan et
al., 2012). Altogether, D4 receptors may not be involved in the reinforcing effects of nicotine but they do
appear to play an important role in the relapse processes of nicotine-seeking behaviour.
1.6.5.2 Opioids Morphine
A study by Mamiya and colleagues (2004) found that L-745,870 significantly attenuated
withdrawal behaviour precipitated by naloxone in morphine-dependent mice. Naloxone administration
in morphine-dependent mice normally elevates cAMP levels in the thalamus, and the co-administration
of D2-like antagonists prevents the regulation of cAMP, inhibiting the expression of withdrawal
behaviour.
The D4 agonist PD 168,077 was found to prevent morphine-induced activation of the
nigrostriatal dopamine pathway and alterations of substantia nigra pars compacta (SNc) morphology. An
examination of behavioural effects revealed that PD 168,077 counteracted morphine-induced
locomotor activity, suppressed the rewarding properties of morphine measured by conditioned place
preference, and prevented the development of withdrawal syndromes in rats. Additionally, L-745,870
counteracted the preventative effects of PD 168,077 on morphine-induced alterations of SNc
morphology (Rivera et al., 2016).

38
Acute activation of D4 receptors by PD 168,077 decreased MOR immunoreactivity (IR) in the
striatum, whereas L-745,870 blocked the downregulation of MOR IR. These results suggest that D4
receptor activation reduces MOR expression (Gago et al., 2007).
1.6.5.3 Hallucinogens Phencyclidine
A prominent hallucinogen is phencyclidine (PCP), a dissociative drug used in animal models of
schizophrenia to induce locomotor hyperactivity, stereotypy, and social isolation. Primarily, PCP acts as a
noncompetitive NMDA receptor antagonist, and when used in humans, can induce a psychotic state
resembling schizophrenia. As an antipsychotic, clozapine has been reported to alleviate cognitive deficits
in monkeys treated with PCP (Jentsch et al., 1997) and acute PCP-induced hyperlocomotion in rats
(Maurel-Remy et al., 1995). D2-like agonists are able to potentiate PCP-induced deficits in social
behaviour, an effect that is attenuated by D2/D3 antagonists. However, the D4 antagonist L-745,870 had
no effect on the behaviour of rats treated with PCP (Sams-Dodd, 1998).
Lysergic acid diethylamide
Through drug discrimination studies, a biphasic pharmacological profile has been established for
lysergic acid diethylamide (LSD): 1) an initial 5-HT2A receptor-mediated phase lasting approximately 1
hour, and 2) a delayed phase mediated by D2-like receptors occurring 60-90 minutes after LSD
administration. A study by Marona-Lewicka (2009) found that the D4 agonists shifted the dose-response
curve of LSD leftward, with the D4 agonist WAY 100635 mimicking the LSD cue during the delayed phase,
while D4 antagonists attenuated the LSD cue during both phases with a more pronounced effect in the
second phase. Thus, these findings suggest that the D4 receptor modulates discriminative stimulus
properties of LSD (Marona-Lewicka et al., 2009).

39
1.7 Experimental Rationale 1.7.1 Summary Overview
Initial administration of addictive substances induces neuroplastic changes in the reward
circuitry of the brain, involving memory and associative learning processes. Over repeated drug
administrations, these processes gradually become dependent on other neurocircuits, increasing
susceptibility to drug-seeking behaviours. Aversive internal states induced by periods of abstinence are
alleviated by drug-taking, promoting negatively reinforcing behaviour. This results in a vicious and
deeply entrenched positive and negative reinforcement cycle of drug-seeking.
Since the revelation of the involvement of the dopaminergic mesolimbic pathway in the reward
system, dopamine has been the focus of intense research in addiction. Dopamine transmission has been
the source of many theoretical accounts of addiction from its involvement in many different aspects of
pathways that reinforce addictive behaviours.
Due to variations in pharmacology and expression, the dopamine receptor subtypes exhibit
different functional roles. And though other dopamine D2-like receptors have undergone significant
examination, the D4 receptor has only recently begun to be investigated due to the development of
novel selective ligands. The role of the D4 receptor in the maintenance and relapse of alcohol addiction
remains unclear, and thus is the aim of this investigative endeavour.
Alcohol, a common substance of abuse, innervates the aforementioned drug-reward circuits and
establishes addiction-related behaviours. This is robustly demonstrated in rats that self-administer
alcohol. Thus, operant oral self-administration of alcohol was used to examine the effects of the D4
receptor on drug-taking. For relapse, cue- and stress-induced reinstatement paradigms were used in
order to examine the effects of D4 receptor modulations on drug-seeking.
1.7.2 Experimental Rationale Hypothesis

40
Through operant oral self-administration of alcohol, rats will heterogeneously acquire
behavioural responses associated with alcohol. It is hypothesized that selective ligands for the D4
receptor will modulate the effects of alcohol self-administration as well as cue-induced and stress-
induced reinstatement.
Purpose of Investigation
The purpose of the experiments is as follows:
1. Are modulations of the D4 receptor able to influence alcohol self-administration?
2. Are modulations of the D4 receptor able to influence alcohol reinstatement through the use of
cues or stress?
These experiments will address these inquiries by determining if any behavioural effects can be
gathered from direct drug-taking behaviour, or drug-seeking behaviours as induced by cues or stress.
CHAPTER 2. Methods 2.1 Animals
A total of 110 naïve male Wistar rats, initially weighing 200-225g, were obtained from Charles
River, Lachine, QC. All animals were pair-housed in a temperature controlled (21-22 °C) vivarium on a
12-h reverse light-dark cycle, with lights on from 7 p.m. to 7 a.m. Animals were allowed to acclimatize to
the colony room with access to ad libitum water and food for 1 week prior to being single-housed and
food restricted to 4 pellets of standard lab chow per day (approximately 20-25g) for training and
experiments. All experimental procedures described were carried out in compliance with the guidelines
of the Canadian Council on Animal Care (CCAC) and the guidelines of the Animal Care and Use
Committee of the National Institute on Drug Abuse.

41
2.2 Drug Preparation Alcohol solutions were prepared daily from a 95% stock solution which was diluted with filtered
water to create the appropriate concentrations of 3, 6, or 12%.
The DRD4 antagonist L-745,870 (3-[4-(4-chlorophenyl) piperazin-1-yl] methyl-1H-pyrrolo [2, 3-b]
pyridine) hydrochloride (Tocris) was used for its high DRD4 selectivity (> 2000-fold) compared to other
dopamine receptor subtypes and its low-moderate affinity (IC50 <300 nM) for non-dopaminergic
receptors (Patel et al., 1997). L-745,870 is reported to have Ki values of 0.43, 960, and 2300 nM for the
D4, D2, and D3 receptor subtypes respectively, with no appreciable binding (Ki > 10,000 nM) for the D1
and D5 subtypes (Kulagowski et al., 1996). And although described as a D4 receptor antagonist, L-745,870
can also act as a partial agonist (Gazi et al., 1999).
L-745,870 was dissolved with saline to create a 10 mg/mL solution, and then diluted into 5, 1,
and 0.1 mg/mL concentrations. Once prepared, any unused solutions were stored aliquoted in vials at -
20°C or below and used within 1 week. A saline solution was used for vehicle injections.
The drug PD 168,077 (N-([4-(2-cyano-phenyl) piperazine-1-yl] methyl)-3-methylbenzamide)
maleate (Tocris) was used for its potency as a D4 receptor agonist with Ki values of 8.7, 2810, and 3740
nM for D4, D3, and D2 receptor subtypes, respectively (Glase et al., 1997). PD 168,077 was dissolved in
saline and dimethyl sulfoxide (DMSO) to a concentration of 10 mg/ml, and then diluted accordingly to 5,
1, and 0.1 mg/ml. A solution of saline and 10% DMSO was used for vehicle injections.
All drugs were administered via intraperitoneal (i.p.) injection. The L-745,870 and PD 168,077
solutions were injected at 30 and 10 minutes, respectively, before all testing sessions.
2.3 Alcohol Training 2.3.1 Two-Bottle Choice Alcohol Acquisition
Procedures for alcohol training were similar to those previously reported (Kostowski & Dyr,
1992; Rezvani et al., 1991; Yan et al., 2012). For initial training, all animals received limited access (30
minutes per day) to alcohol via a 2BC paradigm, a prominent variant of voluntary consumption models

42
(Tabakoff & Hoffman, 2000). In this method, two Richter tubes were presented: one containing filtered
water and the other, unsweetened alcohol. Alcohol acclimatization occurred by gradually escalating
concentrations from 3, 6, to 12%, over 5, 5, and 15 days, respectively, for a total of 25 days. After each
training session, animals were returned to their home cages and fed. Based on available cages outfitted
for 2BC training, animals were separated into four groups: two groups of 30, and two groups of 25.
Measurements for body weight (g) were taken before each session, whereas volume of alcohol
and water consumed (mL) were taken after each session. These measurements were used to calculate
total dose of alcohol (using an alcohol density of 0.789g/mL) and alcohol preference, expressed as a
percentage of alcohol against total fluid consumption.
Calculations are as follows:
Dose of EtOH (g/kg) =∆EtOH (mL) × EtOH % × EtOH density (g/mL)
Body Weight (kg)
EtOH Preference (%) =∆EtOH
∆H20 + ∆𝐸𝐸𝐸𝐸𝐸𝐸𝐸𝐸× 100%
2.3.2 Operant Alcohol Self-Administration After 2BC training was complete, animals were reallocated into 9 groups of 12 and a final group
of 2 (order was unchanged), and continued training in operant chambers to self-administer alcohol
under a FR schedule.
The operant chambers were ventilated, sound-attenuating cubicles, and dispensed alcohol using
a variable infusion rate syringe pump (Med Associates, St. Albans, VT). The chambers were outfitted
with a recessed sipper tray receptacle located between 2 response levers, a 28-V white cue light
positioned above each lever, and a ceiling-mounted house light on the same side which signalled the
start of each session when illuminated.

43
One of the levers was assigned as the inactive lever and had no programmed consequence. The
other was assigned as the active lever, and once a fixed number of presses were reached, the following
events would occur simultaneously for 5 seconds: the house light would turn off, the corresponding cue
light would turn on, and the syringe pump would dispense alcohol to the sipper tray as reinforcement.
All active and inactive lever presses were recorded throughout the session, but during those 5 seconds,
active lever presses did not count towards the required FR schedule. Active and inactive levers were
evenly counterbalanced in each group with lever assignment remaining the same for each animal
throughout all experiments.
Animals were trained on a fixed-ratio 1 (FR1) schedule for 15 days, where 1 active lever press
resulted in alcohol reinforcement, then on an FR2 schedule for 5 days, and FR3 for 15 days. Session
duration was 1 hour with no reinforcement limit, and upon meeting lever press criteria, the syringe
pump (pushbutton setting of 42) would dispense alcohol at a flow rate of 38 µL/s for 5 seconds,
delivering a total volume of 0.19 mL.
Upon completion of each session, any amount of unconsumed alcohol in the sipper tray was
aspirated with a 1 mL syringe, measured and recorded. This value was subtracted from the volume of
alcohol dispensed to calculate the number of reinforcements and volume of alcohol consumed. In order
to habituate animals to injection procedures, i.p. saline injections were administered 8 days before
testing sessions.
The mean ± SEM of active lever presses, inactive lever presses, reinforcements, and total dose of
alcohol over the last 2 days of operant FR training were 42.95±2.09, 3.34±0.57, 12.8±0.61, and
0.51±0.02, respectively.

44
2.4 Sorting Active lever responding and alcohol dose were carefully monitored during the last 15 days of
FR3 alcohol self-administration training in order to apply the appropriate inclusion and exclusion criteria
before continuing on to further testing sessions.
Animals with an average dose of alcohol intake of ≥ 0.4 g/kg over the last 3 consecutive sessions
were to be included, of which 86 out of 110 met this criterion. Animals with a variation of < 20% in
active lever pressing over the last 3 consecutive sessions were also to be included, of which, 50 met this
criteria. As only 47 of the 110 animals met both criteria, inclusion was broadened to be based mainly on
dose of alcohol intake instead.
Initially, animals with an average of < 10 active lever presses for the last 3 consecutive days were
excluded due to low responding, a total of 8. This criterion was, however, expanded to < 23 active lever
presses to exclude 6 more animals, for a total of 14.
The subsequent 12 animals with the lowest average alcohol dose were placed into a
maintenance group to continue FR3 alcohol self-administration. Although rats consuming < 0.4 g/kg
have normally been excluded in previous studies, these animals were kept for potential future use in the
event that they reached the inclusion criteria.
Those that met both criteria, and had the highest alcohol dose averages, were allocated into 2
groups of 12 to be tested on alcohol self-administration. The rest of the 60 rats were allocated into 5
groups of 12 for extinction, to later be tested on reinstatement induced by either cue or stress.
2.5 Alcohol Self-Administration Testing In alcohol self-administration testing, animals were exposed to identical conditions as in FR3
training. Animals were exposed to 5 different doses of their respective drug at either 0 (vehicle), 0.1, 1,
5, or 10 mg/kg, using a variation of the Latin rectangle design where each dose occurred exactly once for
each subject. Testing sessions were separated by 2 baseline days of alcohol self-administration to ensure
stabilization and the washout of previous treatments before the next test session.

45
2.6 Extinction During extinction, the house light remained for the duration of the 1 hour session. Active lever
presses were not reinforced by alcohol, and the associated cue light was withheld. Responses on both
levers were recorded but had no programmed consequences. Extinction was conducted until the last 2
consecutive sessions met the criteria of <20 active lever presses per session or <20% of the average of
active lever presses that occurred over the last 5 days of alcohol self-administration.
Five groups of rats (n=12 for all groups) were initially allocated to the extinction phase.
2.7 Cue-Induced Reinstatement Testing Cue-induced reinstatement testing was similar to FR3 SA sessions, with several exceptions. At
the beginning of the session, the house light would turn on for 4 seconds, followed by a single
presentation of the visual cue (house light off and cue light on) for 5 seconds. The house light
subsequently turned on again until the active lever press criteria for FR3 was met. Although the
contingent presentation of the cue light and activation of the syringe pump still occurred for 5 seconds,
the syringe containing alcohol was removed before the session, and would therefore no longer dispense
an alcohol reinforcement. As with FR3 SA sessions, during this period, active lever presses did not count
towards another cue presentation but were recorded. All recorded responses on the active lever served
as a measure of reinstatement. Responses on the inactive lever were also recorded but had no
consequence. Three testing days occurred: vehicle, drug, and vehicle respectively, each separated by at
least two extinction sessions.
2.8 Stress-Induced Reinstatement Testing Stress-induced reinstatement sessions were similar to FR3 SA sessions as well, with several
differences. Prior to each session, animals were exposed to a period of inescapable, intermittent foot
shock in the operant chamber for 10 minutes while the house light was on. During this time, lever
presses had no programmed consequences.

46
Foot shock (current intensity 0.8 mA, train length 0.5 s) was administered via a modular shock
floor (Model E10-10SF, CoulBourn Instruments) and a micro controlled shock source (Med Associates,
St. Albans, VT) under a variable-interval 40-s schedule (interval range: 10-70s). After termination of foot
shock, responses on levers were recorded for 30 minutes.
2.9 Cohort Sequence Out of the 5 groups of rats (n=12 for each) that underwent extinction, 3 groups were used to
investigate the effect of drugs on cue-induced reinstatement, extinguished for 2 sessions, and then
tested again under stress-induced reinstatement. The other 2 groups underwent testing in the opposite
sequence.
The two groups of rats tested for alcohol SA were exposed to extinction training and then tested
for the effects of cue-induced reinstatement, extinguished again, and tested for stress-induced
reinstatement.
The final group of rats that was run under the maintenance condition was extinguished and
tested on stress-induced reinstatement.
2.10 Food Testing A new group of animals were ordered to conduct food self-administration (n=10). They were
trained to self-administer food under FR1, FR2, FR3, for 5, 3, and 12 days respectively. During the last 8
days of training, animals were injected with saline i.p. to habituate them to injection procedures.
Animals then underwent testing to examine the effects of the D4 antagonist L-745,870 at different
doses, each separated by 2 baseline sessions.
For food SA sessions, the apparatus, stimuli associated with food delivery, and the schedule of
acquisition were the same as those for the alcohol experiments, with the exception that animals
received a food pellet (45 mg dustless precision pellets; Bioserv, Laurel, Maryland) instead of alcohol
reinforcement.

47
2.11 Data Analysis For analyses, one-way repeated-measures analysis of variance (ANOVA) was used followed by
post hoc Tukey tests for the difference in the number of active vs inactive lever presses during
acquisition of alcohol self-administration, alcohol extinction training, acquisition of food self-
administration, food extinction training, and acquisition of alcohol discrimination.
Graph Pad Prism 6.00 was used to perform all analyses, and a result of p ≤ 0.05 was considered
to be statistically significant.
CHAPTER 3. Results 3.1 Alcohol Consumption Training 3.1.1 Two-Bottle Choice
As an aggregate (n=110), animals appeared to follow anticipated trends during the 2BC training
sessions. Animals consumed a stable volume of alcohol for the first 5 days at the 3% concentration
(Figure 1A). For each subsequent increase in alcohol concentration, there was an initial reduction in the
volume of alcohol consumed, most noticeably at 12% on day 11, followed by a gradual increase as they
became accustomed to the new concentrations. Although volume of alcohol consumption fluctuated
during these transitions, the volume of water consumed remained at relatively low levels (Figure 1B),
reflected by a consistently high percentage in alcohol preference (Figure 1C). An exception was the
anomalous upward spike in volume of water, and corresponding downward spike in alcohol preference
on day 14, potentially due to partial feeding before the session. A repeated measures ANOVA showed a
significant effect of “Day” on volume of water consumed [F(7.7,800.4)=13.6, p<.0001], and Tukey’s
pairwise comparisons indicated the volume of water consumed on day 14 was significantly higher than
all others (p<.0001) except for day 8.
Lastly, the total dose of alcohol intake saw a stable and gradual increase (Figure 1D), even
throughout escalations in alcohol concentration. A repeated measures ANOVA showed an effect of

48
“Alcohol Concentration” on the dose of alcohol intake [F(1.7,188.3)=326.2, p<.001]. Post hoc analyses
with Bonferroni correction revealed that each increase in alcohol concentration significantly increased
dose of alcohol intake (p<.01).
FIGURE 1
Figure 1. Aggregate two-bottle choice training over 25 days (n=110). Alcohol was presented at escalating
concentrations of 3, 6, and 12% over 5, 5, and 15 days, respectively. (A) Volume of alcohol consumed
gradually increased over training whereas (B) water consumption gradually decreased. (C) Alcohol
preference is expressed as a percentage of total volume of liquid consumed, and remains relatively
stable. (D) Total dose of alcohol intake gradually increased after each successive increase in alcohol
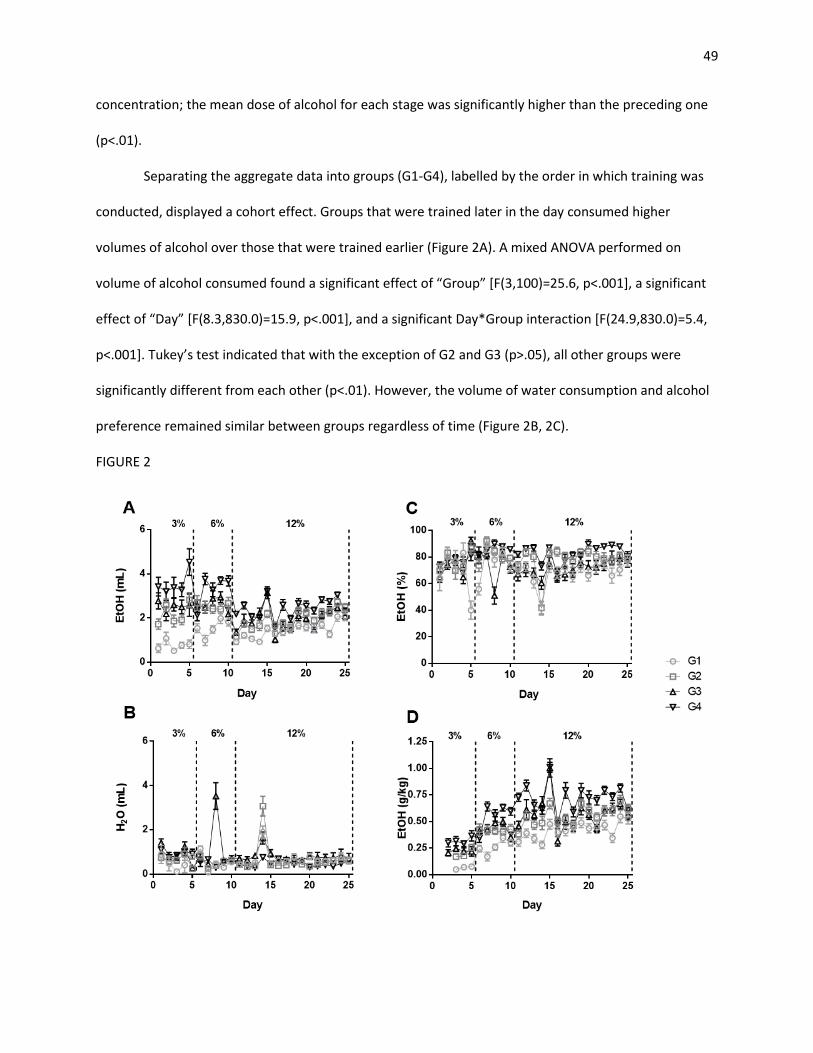
49
concentration; the mean dose of alcohol for each stage was significantly higher than the preceding one
(p<.01).
Separating the aggregate data into groups (G1-G4), labelled by the order in which training was
conducted, displayed a cohort effect. Groups that were trained later in the day consumed higher
volumes of alcohol over those that were trained earlier (Figure 2A). A mixed ANOVA performed on
volume of alcohol consumed found a significant effect of “Group” [F(3,100)=25.6, p<.001], a significant
effect of “Day” [F(8.3,830.0)=15.9, p<.001], and a significant Day*Group interaction [F(24.9,830.0)=5.4,
p<.001]. Tukey’s test indicated that with the exception of G2 and G3 (p>.05), all other groups were
significantly different from each other (p<.01). However, the volume of water consumption and alcohol
preference remained similar between groups regardless of time (Figure 2B, 2C).
FIGURE 2
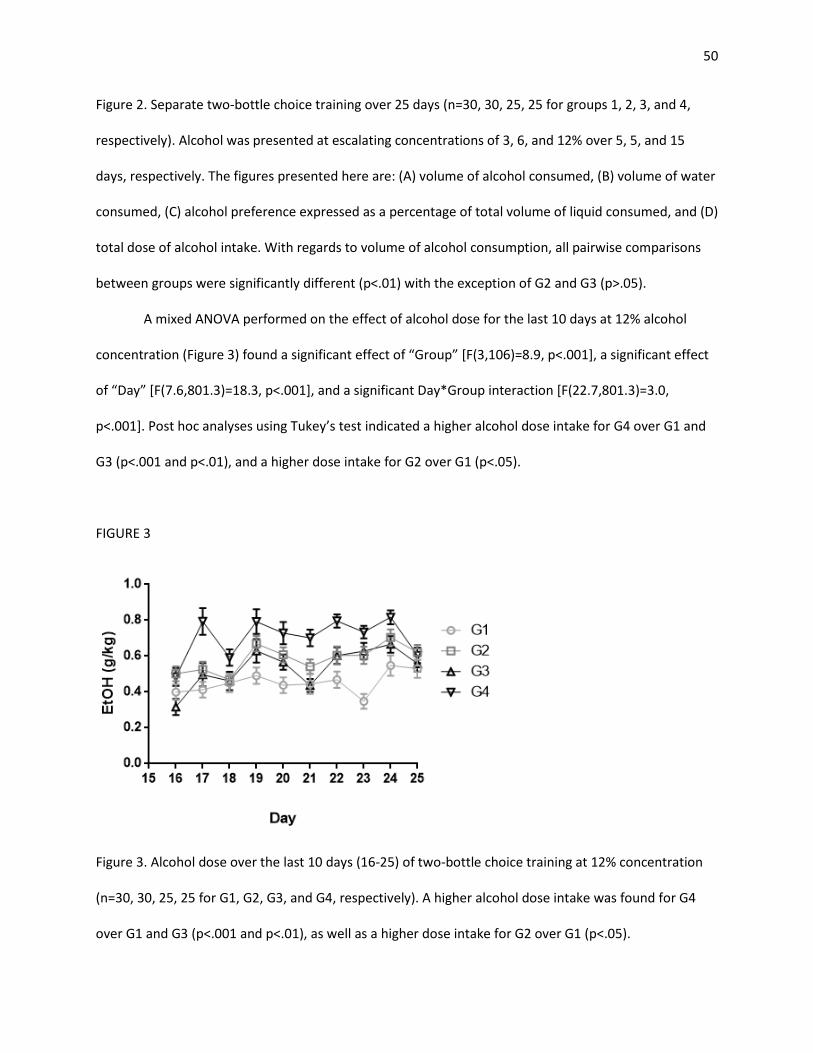
50
Figure 2. Separate two-bottle choice training over 25 days (n=30, 30, 25, 25 for groups 1, 2, 3, and 4,
respectively). Alcohol was presented at escalating concentrations of 3, 6, and 12% over 5, 5, and 15
days, respectively. The figures presented here are: (A) volume of alcohol consumed, (B) volume of water
consumed, (C) alcohol preference expressed as a percentage of total volume of liquid consumed, and (D)
total dose of alcohol intake. With regards to volume of alcohol consumption, all pairwise comparisons
between groups were significantly different (p<.01) with the exception of G2 and G3 (p>.05).
A mixed ANOVA performed on the effect of alcohol dose for the last 10 days at 12% alcohol
concentration (Figure 3) found a significant effect of “Group” [F(3,106)=8.9, p<.001], a significant effect
of “Day” [F(7.6,801.3)=18.3, p<.001], and a significant Day*Group interaction [F(22.7,801.3)=3.0,
p<.001]. Post hoc analyses using Tukey’s test indicated a higher alcohol dose intake for G4 over G1 and
G3 (p<.001 and p<.01), and a higher dose intake for G2 over G1 (p<.05).
FIGURE 3
Figure 3. Alcohol dose over the last 10 days (16-25) of two-bottle choice training at 12% concentration
(n=30, 30, 25, 25 for G1, G2, G3, and G4, respectively). A higher alcohol dose intake was found for G4
over G1 and G3 (p<.001 and p<.01), as well as a higher dose intake for G2 over G1 (p<.05).

51
3.1.2 Operant Oral Self-Administration Animals responded as anticipated during the FR alcohol oral self-administration training.
Animals gradually increased in active lever pressing throughout training procedures with some
fluctuations occurring when transitioning to higher FR schedules. Conversely, inactive lever pressing
started at similar numbers as active lever but immediately decreased and continued to gradually
decrease (Figure 4A). A repeated measures ANOVA revealed a significant effect of “Day” on active lever
press [F(12.3,1336.8)=68.8, p<.001] and inactive lever press [F(14.5,1574.9)=12.6, p<.001].
Volume of alcohol consumed (Figure 4B), number of reinforcements (Figure 4C), and total dose
of alcohol (Figure 4D), all presented similar patterns of initially decreasing after each change in response
requirement, followed by a gradual increase as they adapted to the new schedule.
FIGURE 4

52
Figure 4. Aggregate operant self-administration training over 35 days (n=110) on a FR1, FR2, and FR3
schedule, for 15, 5, and 15 days, respectively. The figures presented here are (A) active and inactive
lever press, (B) volume of alcohol consumed, (C) number of alcohol reinforcements, (D) total dose of
alcohol intake. A significant effect of “Day” was found for both active lever press and inactive lever press
(p<.001).
3.2 Alcohol Self-Administration Testing 3.2.1 The Effects of the Selective DRD4 Antagonist, L-745,870, on Alcohol Self-Administration
A one-way ANOVA was performed on the number of active lever presses that rats (n=12) made
during alcohol self-administration testing sessions (Figure 5A) and showed a significant effect of “Dose”
[F(4,44)=14.5, p<.001]. Tukey’s pairwise comparisons indicated that the 10 mg/kg (i.p.) dose of L-
745,870 significantly reduced the number of active lever presses when compared with all other doses
(p<.0001 for vehicle, doses 0.5, 1, and 10 mg/kg, and p<.01 for 5 mg/kg). All other pairwise comparisons
were not significant. The ANOVA performed on the number of inactive lever presses that rats made
during alcohol self-administration testing sessions (Figure 5B) showed no significant effects of Dose
[F(1.6,17.7)=2.5, p>.05].
For the number of reinforcements earned during the alcohol self-administration testing sessions
(Figure 5C), the ANOVA showed a significant effect of “Dose” [F(4,44)=12.7, p<.001], reflecting the
results of active lever presses. Further pairwise comparisons revealed that the 10 mg/kg dose of L-
745,870 significantly reduced the number of reinforcements when compared to all other doses (p<.0001
for vehicle, doses 0.5, 1, and 10 mg/kg, and p<.01 for 5 mg/kg). All other pairwise comparisons were not
significant.
The ANOVA performed on the total alcohol intake (g/kg) of rats during self-administration
testing sessions (Figure 5D) showed a main effect of “Dose” [F(2.3,24.8)=12.4, p<.001]. Once again, the
10 mg/kg dose of L-745,870 significantly reduced the amount of alcohol intake when compared to all

53
doses (p<.0001 for vehicle, doses 0.5, 1, and 10 mg/kg, and p<.01 for 5 mg/kg). There were no other
significant pairwise comparisons.
FIGURE 5
Figure 5. Effects of the DRD4 antagonist L-745,870 on alcohol self-administration under a fixed-ratio 3
(FR3) schedule of reinforcement (n = 12). Data are expressed as mean (±SEM) number of active lever
presses (A), inactive lever presses (B), alcohol reinforcements earned (C), or amount of alcohol intake in
g/kg (D). The symbols **, ****, and ns, indicate p<.01, p<.0001, and non-significance, respectively.
3.2.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Alcohol Self-Administration The one-way ANOVA performed on all measures of interest during alcohol self-administration
testing sessions when treated with the dopamine agonist PD-168,077 yielded no significant effects.
Testing sessions showed no significant effects of “Dose” on active [Figure 6A; F(4,44)=2.2, p=.083] or

54
inactive [Figure 6B; F(2.1,23.4)=.41, p=.68] lever presses of rats (n=12). Similarly, no significant effects of
“Dose” were seen on reinforcements consumed [Figure 6C; F(1.8,19.6)=1.7, p=.21] or total dose of
alcohol intake [Figure 6D; F(1.9,20.4)=1.7, p=.21].
FIGURE 6
Figure 6. Effects of the DRD4 agonist PD-168,077 on alcohol self-administration under a fixed-ratio 3
(FR3) schedule of reinforcement (n = 12). Data are expressed as mean (±SEM) number of active lever
presses (A), inactive lever presses (B), alcohol reinforcements earned (C), or amount of alcohol intake in
g/kg (D). No significant effects were indicated for any of the graphs.
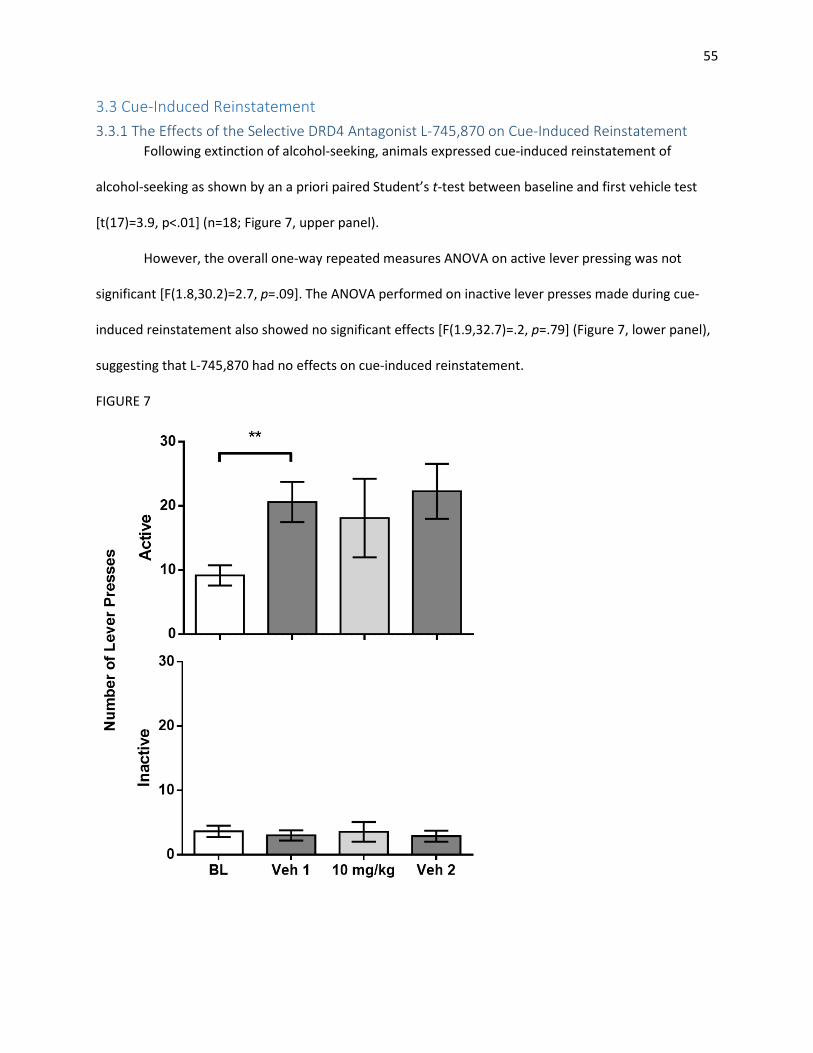
55
3.3 Cue-Induced Reinstatement 3.3.1 The Effects of the Selective DRD4 Antagonist L-745,870 on Cue-Induced Reinstatement
Following extinction of alcohol-seeking, animals expressed cue-induced reinstatement of
alcohol-seeking as shown by an a priori paired Student’s t-test between baseline and first vehicle test
[t(17)=3.9, p<.01] (n=18; Figure 7, upper panel).
However, the overall one-way repeated measures ANOVA on active lever pressing was not
significant [F(1.8,30.2)=2.7, p=.09]. The ANOVA performed on inactive lever presses made during cue-
induced reinstatement also showed no significant effects [F(1.9,32.7)=.2, p=.79] (Figure 7, lower panel),
suggesting that L-745,870 had no effects on cue-induced reinstatement.
FIGURE 7

56
Figure 7. Effects of the DRD4 antagonist L-745,870 on cue-induced reinstatement of alcohol-seeking
behaviour under a fixed-ratio 3 (FR3) schedule of reinforcement (n = 18). Data are expressed as mean
(±SEM) of the number of active lever presses (upper panel), inactive lever presses (lower panel).
Presented here are the baseline session (BL), first vehicle test, 10 mg/kg (i.p.) L-745,870 test, and second
vehicle test. Significance of p<.01 is indicated by (**).
In order to distill the data to only contain subjects that exhibited cue-induced reinstatement
behaviour, the inclusion threshold for active lever pressing was increased to ≥10 for both vehicle tests.
Further increases of active lever pressing threshold would result in too low of a sample size.
The results of the distillation were similar to preceding findings. The baseline extinction session
was significantly different to the first vehicle test [t(12)=3.8, p<.01] (n=13; Figure 8, upper panel).
However, the one-way ANOVA yielded no significant effects on active lever pressing [F(1.6,19.1)=2.7,
p=.10] or inactive lever pressing [F(1.8,22.1)=.1, p=.86] (Figure 8, lower panel). Thus, it appears that
while cue-induced reinstatement was achieved for the first vehicle test, L-745,870 had no significant
effects on active lever pressing.
FIGURE 8

57
Figure 8. Effects of the DRD4 antagonist L-745,870 on cue-induced reinstatement of alcohol-seeking (n =
13) with vehicle test thresholds of ≥10 active lever presses. Data are expressed as mean (±SEM) number
of active lever presses (upper panel), inactive lever presses (lower panel). Presented here are the
baseline (BL), first vehicle test, 10 mg/kg (i.p.) L-745,870 test, and second vehicle test. Significance of
p<.01 is indicated by (**).
3.3.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Cue-Induced Reinstatement An a priori paired Student’s t-test performed between baseline and the first vehicle test
indicated that animals expressed cue-induced reinstatement of alcohol-seeking [t(16)=2.26, p<.05]
(n=17; Figure 9, upper panel).

58
Overall one-way ANOVAs were not significant for active lever pressing [F(1.9,30.4)=2.2, p=.13] or
inactive lever pressing [F(2.15,34.4)=2.7, p=.08] (Figure 9, lower panel), suggesting that PD 168,077 has
no effects on cue-induced reinstatement.
Additionally, applying a similar inclusion threshold as done previously with L-745,870 cue-
induced reinstatement would yield too low of a sample size to perform any meaningful analyses, and so
was not conducted.
FIGURE 9
Figure 9. Effects of the DRD4 agonist PD 168,077 on cue-induced reinstatement of alcohol-seeking
behaviour under a fixed-ratio 3 (FR3) schedule of reinforcement (n = 17). Data are expressed as mean
(±SEM) of the number of active lever presses (upper panel), inactive lever presses (lower panel).

59
Presented here are the baseline session (BL), first vehicle test, 10 mg/kg (i.p.) PD 168,077 test, and
second vehicle test. Significance of p<.05 is indicated by (*).
3.4 Stress-Induced Reinstatement 3.4.1 The Effects of the Selective DRD4 Antagonist L-745,870 on Stress-Induced Reinstatement
After extinction of alcohol-seeking, animals exhibited stress-induced reinstatement indicated by
an a priori Student’s t-test between baseline and the first vehicle test [t(33)=6.5, p<.0001] (n=34; Figure
10, upper panel). The overall one-way repeated measures ANOVA on active lever pressing was
significant [F(1.9,61.6)=23.7, p<.0001].
Post hoc Tukey’s test revealed that active lever presses were significantly higher in the first
vehicle test when compared to the baseline and 10 mg/kg (i.p.) L-745,870 dose (p<.0001). Active lever
presses in the second vehicle test were significantly higher than the baseline and lower than the first
vehicle test (p<.01), and the baseline was significantly lower than the 10 mg/kg dose (p<.05).
The one-way ANOVA on inactive lever pressing was significant as well [F(2.0,65.6)=8.6, p=<.001]
(Figure 10, lower panel). Pairwise comparisons via Tukey’s test revealed that the baseline had
significantly fewer inactive lever presses than all other conditions (p<.01, p<.05 for second vehicle test),
and the first vehicle test was significantly higher than the 10 mg/kg (i.p.) dose (p<.05).
FIGURE 10

60
Figure 10. Effects of the DRD4 antagonist L-745,870 on stress-induced reinstatement of alcohol-seeking
behaviour under a FR3 schedule of reinforcement (n=34). Data are expressed as mean (±SEM) number
of active lever presses (upper panel), inactive lever presses (lower panel). Presented here are the
baseline (BL), first vehicle test, 10 mg/kg (i.p.) L-745,870 test, and second vehicle test. Asterisks are with
regards to BL where significance values of p<.0001, p<.01, and p<.05, are indicated by (****), (**), and
(*), respectively.
As shock data is prone to producing noise and erratic data, the inclusion threshold for active
lever pressing was increased to ≥40 for both vehicle tests in an attempt to only capture subjects that
were responsive to stress-induced reinstatement.
A one-way repeated-measures ANOVA on active lever pressing (n=7) yielded a significant overall
effect [F(3,18)=16.6, p<.0001] (Figure 11, upper panel). The following post hoc Tukey’s test revealed that

61
active lever presses for the first and second vehicle test were significantly higher than baseline (p<.0001
and p<.001 respectively). Additionally, active lever presses for both vehicle tests were significantly
higher than the 10 mg/kg (i.p.) dose (p<.01). No significant effects were found for inactive lever presses
[F(1.1,6.5)=2.4, p=.17] (Figure 11, lower panel).
FIGURE 11
Figure 11. Effects of the DRD4 antagonist L-745,870 on stress-induced reinstatement of alcohol-seeking
(n=7) with a threshold of ≥40 active lever presses for both vehicle tests. Data are expressed as mean
(±SEM) number of active lever presses (upper panel), inactive lever presses (lower panel). Presented
here are the baseline (BL), first vehicle test, 10 mg/kg (i.p.) L-745,870 test, and second vehicle test. The
asterisks (****) and (***) represent comparisons to BL indicating significance values of p<.0001 and

62
p<.001, respectively, whereas (**) represents comparisons to both vehicle tests with a significance of
p<.01.
3.4.2 The Effects of the Selective DRD4 Agonist PD-168,077 on Stress-Induced Reinstatement Animals appeared to display stress-induced reinstatement of alcohol-seeking behaviour
indicated by a Student’s t-test between baseline and the first vehicle test [t(25)=3.8, p=<.001] (n=26;
Figure 12, upper panel). The one-way repeated measures ANOVA on active lever pressing was significant
[F(3,75)=6.2, p<.001]. Pairwise comparisons via Tukey’s test revealed that active lever presses for the
baseline session was significantly lower than the first vehicle test, the 10 mg/kg (i.p.) PD 168,077 dose,
and the second vehicle test (p<.001, p<.01, and p<.05, respectively).
The ANOVA on inactive lever pressing was significant as well [F(2.5,62.7)=5.8, p<.01] (Figure 12,
lower panel). Pairwise comparisons via Tukey’s test revealed that inactive lever presses for baseline was
significantly lower compared to all other conditions (p<.01, p<.05 for second vehicle test).
FIGURE 12

63
Figure 12. Effects of DRD4 agonist PD-168,077 on stress-induced reinstatement of alcohol-seeking
behaviour (n=26). Data are expressed as means (±SEM) of the number of active lever presses (upper
panel), inactive lever presses (lower panel). Presented here are the baseline (BL), first vehicle test, 10
mg/kg (i.p.) PD 168,077 test, and second vehicle test. The asterisks (***), (**), and (*) represent
comparisons to BL indicating significance values of p<.001, p<.01, and p<.05, respectively.
Once again, the data was distilled to include subjects that reached active lever pressing of ≥20
for both vehicle tests; further threshold increases would result in too low of a sample size. An a priori t-
test between baseline and the first vehicle test was significant [t(7)=2.7, p<.05] (n=8; Figure 13, upper
panel), indicating that the effect of stress-induced reinstatement remained.
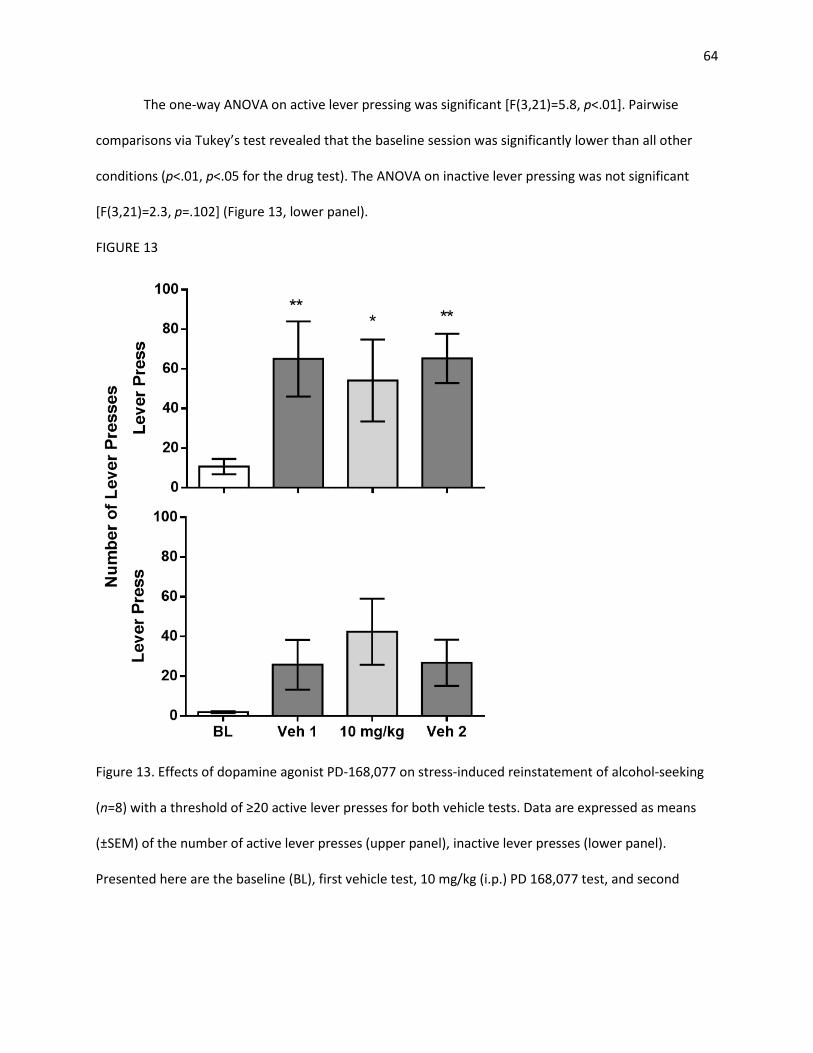
64
The one-way ANOVA on active lever pressing was significant [F(3,21)=5.8, p<.01]. Pairwise
comparisons via Tukey’s test revealed that the baseline session was significantly lower than all other
conditions (p<.01, p<.05 for the drug test). The ANOVA on inactive lever pressing was not significant
[F(3,21)=2.3, p=.102] (Figure 13, lower panel).
FIGURE 13
Figure 13. Effects of dopamine agonist PD-168,077 on stress-induced reinstatement of alcohol-seeking
(n=8) with a threshold of ≥20 active lever presses for both vehicle tests. Data are expressed as means
(±SEM) of the number of active lever presses (upper panel), inactive lever presses (lower panel).
Presented here are the baseline (BL), first vehicle test, 10 mg/kg (i.p.) PD 168,077 test, and second

65
vehicle test. The asterisks (**) and (*) represent comparisons to BL indicating significance values of
p<.01 and p<.05, respectively.
3.5 Food Control 3.5.1 Effects of L-745,870 on Food Self-Administration
A repeated measures one-way ANOVA (n=10) was used for all measures pertaining to food self-
administration. No overall significant effects were found for active [F(2.7,24.2)=3.0, p>.05] or inactive
lever pressing [F(1.4,12.3)=0.8, p>.05] (Figure 14A), as well as number of reinforcements
[F(2.8,25.4)=1.1, p>.05] (Figure 14B). Further post hoc multiple comparisons found no significant
differences.
FIGURE 14

66
Figure 14. Food self-administration under a fixed-ratio 3 (FR3) schedule of reinforcement (n=10). Data
are expressed as means (±SEM) of the number of active and inactive lever presses (A), and number of
food reinforcements earned (B).
CHAPTER 4. Discussion 4.1 Overall Outcomes
The experiments conducted here aimed to examine the effects of D4 receptor modulation on
alcohol addiction. And through the use of selective ligands, it was possible to attain a deeper
understanding of the role of the D4 receptor on specific addiction-relevant behaviours, contributing to
the groundwork necessary for the development of future therapeutic strategies. The overall outcomes
of this investigation are summarised briefly as follows:
The selective D4 receptor antagonist L-745,870 was found to attenuate alcohol self-
administration, without affecting food self-administration, suggesting that D4 receptor blockade
influences motivational processes for alcohol-taking without affecting natural rewards. As for cue- and
stress-induced reinstatement, initial analyses found no effects of D4 receptor modulation. However, by
distilling for robust responders, alcohol-seeking appeared to be reduced by D4 receptor blockade under
the stress-induced reinstatement paradigm.
The results obtained here further supports the assertion that the D4 receptor does indeed play a
role in addictive disorders, where D4 receptor blockade attenuates alcohol self-administration, and
although there seems to be a plausible effect of D4 receptor blockade on stress-induced reinstatement,
D4 receptor modulation does not appear to influence reinstatement mediated by cues. Given these
findings, the data suggests that D4 receptors are a potential therapeutic target for curbing AUDs, and
possibly relapse.

67
4.2 Overview of Relevant Literature 4.2.1 Summary of D4 Receptor Literature
To date, relatively few studies have explored the D4 receptor with regards to addictive disorders
and even less so for alcohol disorders specifically. In fact, previous pharmacological studies may have
misattributed their effects to other D2-like receptors due to the late discovery of the D4 receptor and
absence of selective D4 ligands. But in accordance with the literature to date, the experiments in this
study deepen existing lines of evidence that point to the D4 receptor being responsible for reductions in
alcohol consumption.
In an early preclinical study, the anxiolytic buspirone was found to reduce alcohol consumption
in macaque monkeys, an effect earlier attributed to other dopamine receptors (Collins & Myers, 1987).
It was later revealed that buspirone actually binds to D4 receptors with a much higher affinity than other
D2-like receptors (Bergman et al., 2013), suggesting that D4 receptor blockade may have contributed
more significantly to the reduction of alcohol consumption.
Later on, D4 receptor blockade by L-745,870 was reported to attenuate nicotine- and cue-
induced reinstatement of nicotine-seeking behavior in rats (Yan et al., 2012). Additionally, L-745,870
significantly attenuated the withdrawal syndrome precipitated by naloxone in morphine-dependent
mice (Mamiya et al., 2004). Thus, several lines of research in the preclinical setting seem to point
towards the therapeutic potential for D4 receptor antagonism, not only in relapse, but in reducing
aversive withdrawal symptoms as well. And in a more recent study, DRD4 deficient mice were less
anxious (as demonstrated by greater novelty-seeking behaviour and exploratory behaviour), and
consumed less alcohol than controls (Thanos et al., 2015). These findings suggest that the DRD4 gene
does indeed play a role in exploratory and anxiolytic behaviour in males, behaviours that were positively
correlated with increased alcohol consumption.
In the clinical setting, it was earlier reported that clozapine treatment reduced comorbid
substance abuse, including alcohol, in schizophrenia patients (Marcus & Snyder, 1995). And due to

68
clozapine’s high affinity for D4 receptors, interest in its therapeutic potential for addiction has been
galvanized. Since then, most clinical studies on this topic have diverted attention towards understanding
the effects of variations in the DRD4 gene and its associations with personality traits and novelty-seeking
behaviours. However, there is still progress being made on its effects on substances on abuse; for
instance, one study reported a correlation between alcohol intake and the DRD4 genotype (Luciano et
al., 2004).
Having taken into consideration the involvement of the D4 receptor in addictive disorders, we
conducted this study to better understand how pharmacologically targeting this receptor would
influence behaviours relevant to alcohol disorders. Taken together, our findings are generally aligned
with many previous preclinical and clinical studies, the implications of which, will be examined further in
the following sections.
4.3 Experimental Paradigms 4.3.1 Operant Self-Administration Paradigm
The operant self-administration paradigm was used to assess motivation to obtain alcohol,
where the number of active lever responses were interpreted as a reflection of the amount of effort an
animal was willing to commit to receive a rewarding stimulus. And by confining alcohol reinforcements
to a requisite FR response, the motivation for drug-taking behaviour could be constrained to the
pharmacological effect of alcohol instead of other factors. Thus, the attenuation of alcohol self-
administration elicited by L-745,870 suggests that the motivation to commit effort towards obtaining a
drug reward was decreased, likely through reducing the direct reinforcing capacity of alcohol.
Interestingly, in the previously mentioned study by Yan and colleagues (2012), L-745,870 did not
influence nicotine self-administration, suggesting that the effects of D4 receptor blockade are dependent
on the drug of abuse.
Another important point to consider however is that previous studies have shown that
moderate adjustments of drug dose have been followed by compensatory behaviours in order to sustain

69
constant levels of dopamine (Heidbreder, 2011). This phenomenon was demonstrated further by
alterations of not just the drug dose itself, but the downstream effects as well; the neuroleptic
pimozide, a D2-like antagonist, was reported to increase responding for amphetamines in rats (Yokel &
Wise, 1975). It was hypothesized that dopamine blockade reduced the amphetamine’s rewarding
properties, causing animals to consume higher doses to achieve previous levels of drug satiety. Thus, it
is imperative that potential therapeutic drugs do not have the adverse effect of increasing drug-taking.
In the current study, this compensatory effect was not observed with the administration of L-
745,870 and appeared to only decrease drug responding and alcohol intake, suggesting an attenuation
of motivational processes for drug-taking. As a paradigm with high construct validity, the results of the
experiment suggest that D4 receptor blockade will have low abuse liability in humans, lending further
support to pursue D4 receptor modulation as a prospective novel therapy for AUDs.
In previous studies, the D4 receptor has been shown to play a role in locomotor activity, and so it
is plausible that the reductions in lever pressing and alcohol intake by D4 blockade were due to
decreases locomotor activity rather than motivational processes. However, no significant effects on food
self-administration were found with D4 receptor blockade for all doses. Thus, it is unlikely that
reductions in alcohol self-administration by L-745,870 were due to decreases in locomotor activity.
In addition, these results suggest that the motivation to obtain natural rewards is unaffected by
D4 receptor blockade, and more specifically, influences circuits involved with drugs of abuse. Taken
together, these findings support selective D4 receptor blockade as a therapeutic strategy to attenuate
drug-taking while sparing motivational processes involving natural rewards.
4.3.2 Cue-Induced Reinstatement Dopaminergic innervation of the BLA and the mPFC are purported to be key components in the
cue-induced reinstatement neurocircuit, which in turn send glutamatergic projections to the NAc core.
Although cue-induced reinstatement has been found to be inhibited by the inactivation of other brain

70
regions such as the anterior cingulate, VTA, NAc, and lateral OFC, the BLA appears to be an essential
component distinguishing this circuit from other modes of reinstatement.
This is a sensible claim as the BLA is especially necessary for the acquisition of associative
learning in which dopamine transmission is believed to enhance the salience of drug-paired stimuli. In
support of this, pharmacological lesioning and inactivation of the BLA have been found to attenuate cue-
induced reinstatement of cocaine, whereas amphetamine infusions stimulating dopamine release in the
BLA potentiated conditioned-cue reinstatement (Meil & See, 1997; See et al., 2003). This circuit was
believed to be primarily mediated by D1-like receptors as intra-BLA infusions of D1-like receptor
antagonists reduced cue-induced reinstatement of cocaine (See et al., 2001). However, more recently,
systemic D4 receptor blockade by L-745,870 was found to attenuate cue-induced reinstatement of
nicotine-seeking behaviour without affecting the reinstatement of food-seeking (Yan et al., 2012). And
although PD 168,077 did not reinstate extinguished nicotine-seeking behaviour, it is unknown if D4
receptor agonism would have potentiated cue-induced nicotine-seeking as this experiment was not
conducted (Yan et al., 2012). Since D4 receptors are highly expressed in the basal and central nuclei of
the amygdala (Xiang et al., 2008), it would be apt to presume that the ligands may have been acting on
these regions.
Given these findings, it would be reasonable to anticipate that cue-induced reinstatement of
alcohol would similarly be enhanced or reduced with dopaminergic activation or blockade, respectively.
But while reinstatement behaviour was elicited under vehicle treatment, the administration of either D4
receptor ligands did not yield any significant effects on cue-induced alcohol-seeking in our experiments.
These contradictory findings seem to suggest that reinstatement processes induced by cues are
even more distinctly dependent on the specific drug of abuse and dopamine receptor subtype than
previously thought. This is especially perplexing as D1-like and D2-like family receptors have opposing
effects on cAMP levels, and subsequent downstream processes, and thus, these findings would be

71
better reconciled if the effects of D4 receptor blockade on drug-seeking occurred elsewhere from the
BLA. But even so, as there were no effects of D4 receptor modulation on alcohol-seeking, this suggests
that the reinstatement of alcohol by cues is not dependent on D4 receptors, and that this process may
reside in a different neurocircuit than that of nicotine. This may, in part, be due to alcohol’s unique
biochemical properties, variety in binding of ion channels, and lower primary reinforcing effects relative
to nicotine.
4.3.3 Stress-Induced Reinstatement Although cue- and stress-induced reinstatement appear outwardly similar, their underlying
mechanisms are dependent on different neurocircuits. Stressful events are known to primarily activate
the HPA axis, but its downstream signaling cascades intertwine with reward circuits, leading to relapse.
In terms of neurotransmission, stressful stimuli have been found to activate the release of glutamate
from the PFC to the VTA as well as induce the production of corticosterone, stimulating dopaminergic
projections from the VTA to the NAc (Self & Nestler, 1998).
For this circuit, the CeA has been revealed as a crucial region for receiving and processing pain
information, functioning as the major output of the amygdala (Hasanein et al., 2008). Indeed, the
release of CRF in the CeA is responsible for mediating stress-related drug-seeking behaviours and
alcohol withdrawal symptoms (Koob et al., 1994). And accordingly, injections of noradrenergic and CRF
antagonists into the CeA and lateral BNST have been found to attenuate stress-induced reinstatement
(Pich et al., 1995).
As D4 mRNA are highly expressed in the CeA, it is likely that they also contribute to the stress-
mediated reinstatement of drug-seeking. Furthermore, noradrenaline and adrenaline are high affinity D4
receptor agonists, albeit at higher concentrations (50 to 100-fold) than dopamine (Newman-Tancredi et
al., 1997). Thus, it is possible that the effects of increased noradrenaline levels are involved in inducing
stress-induced reinstatement in the CeA which may then be mitigated by D4 receptor blockade.

72
Though not initially apparent in our experiments, D4 receptor antagonism appeared to attenuate
stress-induced reinstatement in robust responders. Stress-induced reinstatement was evoked as
indicated by higher active lever pressing on vehicle tests, however the results from the administration of
L-745,870 were less clear, warranting further examination. Raising the active lever threshold in both
vehicle tests for inclusion allowed for the distillation of highly responsive subjects to stress-induced
reinstatement. As expected, vehicle tests had significantly higher active lever presses than baseline
(p<.001), but more importantly, both vehicle tests were significantly higher than the 10 mg/kg dose
(p<.01), suggesting an attenuation effect on reinstatement.
In light of these results, it does seem plausible that D4 receptor antagonism occurred at the CeA,
reducing the effects of noradrenaline, and overall output of pain signalling from the amygdala to
decrease drug-seeking. And while unclear what effects D4 receptor antagonism may have had on the
neurotransmission of other systems involving CRF and adrenergic projections, it is possible that
blockade also occurred at the VTA, preventing the stimulation of the NAc by dopamine.
Conversely, in another study, L-745,870 had no significant effects on mice in an EPM test, and
was surmised not to modulate anxiety-related behaviours (Cao & Rodgers, 1997). Thus, it may be the
case that D4 blockade produces no observable effects with low-intensity anxiety, or that there were no
perceived means to alleviate the aversive environment. Thus, it may be the case that the anxiety elicited
by the EPM test was not intensive enough to produce observable effects by D4 blockade, or that there
were no perceived means to suitably alleviate the aversive experience.
Adding consistency to the preceding experiments, no discernable behavioural effects were
observed for PD 168,077 on stress-induced reinstatement, thus pointing towards D4 agonism having no
overall effects on drug-taking or -seeking.

73
4.4 Theoretical Implications of Findings 4.4.1 Dopamine Reward Hypothesis
In one of the most established theoretical frameworks, the mesolimbic dopamine reward
hypothesis asserts that dopamine transmission from midbrain VTA neurons to the NAc is the primary
mechanism involved in the development and maintenance of addiction. This has been corroborated by
numerous imaging studies in which dopamine levels and receptors in the NAc were acutely elevated
during drug use and chronically reduced in drug-dependent subjects. This hypothesis has since
progressed towards emphasizing a role for dopamine transmission on habit formation and ‘craving’ over
prolonged periods, rather than solely regulating incentive salience for rewarding stimuli in the short-
term.
As this framework is well suited for explaining the results obtained from self-administration
models, it was anticipated that alcohol-induced elevations of dopamine in the NAc would be reduced by
D4 receptor blockade. Further speculation using this framework would suggest that the reductions in
alcohol-taking were caused by the disruption of either automatic goal-directed habit processes or the
craving sensation of alcohol. However, it should then follow that D4 receptor activation would increase
alcohol-taking behaviour, but this effect was not observed. Thus, within the confines of this framework,
these findings would suggest that the disruption was perhaps due to interference in habitual processes
rather than craving or rewarding aspects of alcohol, otherwise we would have observed increased
alcohol-taking.
4.4.2 Theories of Executive Function Theories of executive function of addiction focus on motivational neurocircuits and propose that
the inhibitory control exerted by the PFC becomes progressively overwhelmed by impaired reward and
learning circuits over sustained drug abuse (Nora D. Volkow et al., 2003). These theories assert that the
development of addiction is caused by dysfunction primarily originating from the frontal cortex and
amygdala (Jentsch & Taylor, 1999), which is especially pertinent for our study as the highest levels of D4

74
receptor expression occurs in these regions. As animals were habituated to self-administer alcohol
during training, it is likely that enduring neuroplastic changes occurred in multiple brain regions
including the aforementioned frontal cortex and amygdala, impairing inhibitory control and enhancing
conditioned rewards, respectively.
Theoretical frameworks of executive function primarily revolve around the frontal cortex for its
role in regulating internal motivational states via inhibitory control mechanisms. And as dopamine in the
frontal cortex is believed to oppose neurocircuits such as reward and motivation to produce balanced
responses, dysregulation of dopaminergic transmission is purported to be responsible for the lack of
inhibitory control (Nora D. Volkow et al., 2003).
With basal levels of dopamine activating D4 receptors, downregulating GABA receptors and
currents in postsynaptic PFC neurons (Wang et al., 2002), the equilibrium inhibitory tone is even further
reduced from drug acquisition. And so it is possible that D4 receptor blockade in this region counteracts
the basal downregulation of GABA receptors, allowing for increased inhibitory control, and assisting in
the overcoming of the aforementioned reward and memory circuits. Along this line of interpretation
however, further activation of D4 receptors via agonists should further impair inhibitory control, but this
effect was not observed in any of the paradigms.
On the other hand, hyperactive dopaminergic projections from the VTA to the amygdala are
purported to augment the acquisition of stimulus-reward associations, where the CeA potentiates
conditioned rewards, and the BLA is crucial for cue-induced reinstatement, but not primary
reinforcement (Meil & See, 1997; Robledo et al., 1996). And so, blockade of D4 receptors in the
amygdala should theoretically attenuate the established hyperactive dopaminergic transmission
between the VTA and amygdala, and effectively reduce the incentive properties of alcohol.
Indeed, this is aligned with the results found during alcohol operant self-administration testing
with L-745,870, where active lever-pressing was reduced. But once again, it would be expected that

75
further dopaminergic activation via PD 168,077 would cause the opposite effect, increasing alcohol self-
administration, but this effect was not seen. A possible explanation for this is that once addiction is
established, a hyperactive dopaminergic state may result in the downregulation of dopamine receptors
themselves. Thus, the maximum response may have already been elicited once all receptors were
occupied, rendering additional dopamine agonists ineffective. Alternatively, the dose of the agonist may
have been too low for an effect to occur, seeing as how the antagonist was only efficacious at its highest
dose.
As for cue-induced reinstatement, similar results would be predicted where blockade would
have decreased alcohol-seeking, and activation would have increased alcohol-seeking, but no significant
effects were observed. However, the reduction of stress-induced reinstatement by D4 receptor blockade
is interesting because it suggests a different set of neurocircuits were affected, calling for further
examination. Thus, these findings do seem to be congruous with the theory of executive function
involving the amygdala of learning and memory, albeit with new inquiries.
4.5 Challenges and Limitations There were several challenges and limitations to this study that should be taken into
consideration with the interpretation of the aforementioned results. Although necessary for
acclimatization and habituation before experimental procedures, repeated handling and i.p. injections
may have acted as a prolonged stressor, increasing cortisol levels. More specifically, chronically elevated
cortisol levels could have influenced behavioural output during reinstatement paradigms, blunting the
intensity of stress induced by foot-shock, and interfering with the effect of cues and their associated
pathways. This would have explained, in part, why drug-seeking was not highly evoked during
reinstatement procedures. Additionally, as it was challenging to elicit drug-seeking behaviour, especially
when induced by cue, perhaps a compound cue (i.e. tone and light) could have been more effective in
inducing reinstatement than simply just a cue-light (See et al., 1999).

76
Due to variations in metabolism among subjects, orally self-administering alcohol can result in
different levels of dose and effect, potentially impacting behaviour and motivational processes.
Although it is presumed that subjects naturally reach an equilibrium reinforcing dose, other confounds
related to oral intake may still be present. Thus, to ensure consistent blood alcohol levels and
pharmacological effects, intravenous administration could have been utilized (Tabakoff & Hoffman,
2000). Although intravenous alcohol self-administration may be difficult to sustain in rodents, it would
however allow for more precise control over these variables, bypassing possible confounds.
Furthermore, blood alcohol content (BAC) obtained through traditional calculations may not accurately
correlate with absolute alcohol intake due several variables such as: pattern of responding, circadian
time points during testing, and body weight to fat ratio differences (June & Gilpin, 2010). Thus, operant
alcohol self-administration procedures could have been validated by assessing the post-session BAC via
tail blood samples.
Furthermore, the pharmacodynamics of alcohol are notoriously elusive. At this point, it is known
that alcohol induces behavioural effects similar to drugs that target GABAA receptors. And indeed,
recent research has pointed towards a subclass of GABA receptors that are bound at low millimolar
concentrations of alcohol to enhance GABAergic neurotransmission, thereby increasing tonic inhibition
(Santhakumar et al., 2007). But there are also contradictory reports showing no effect at higher
concentration of alcohol in vivo or in vitro (Lobo & Harris, 2008). Thus, despite being studied extensively,
the precise mechanisms of action of alcohol continue to remain unclear, and so it is uncertain how other
systems and brain regions are concurrently affected by alcohol, as well as how they interact with the
effects of D4 receptor modulation.
In terms of biochemistry, a potential limitation in this study is the difference in dissociation
constants of the D4 receptor ligands used, and the possible ramifications of this discrepancy. As the Ki
values of L-745,870 and PD 168,077 for D4 receptors are 0.43 and 8.7, respectively, it is unclear on

77
whether the magnitude of the responses elicited can be compared equivalently. Biological profiling has
suggested that 0.035 mg/kg of L-745,870 can occupy 50% of D4 receptors in the mouse brain, whereas
1.0 mg/kg occupies up to 90% (Patel et al., 1997). However, seeing as PD 168,077 appears to have low
binding affinities for other dopamine or serotonin receptors (Glase et al., 1997), D4 receptor occupancy
does not seem to be an issue.
One of the more evident limitations of this investigation is the absence of intracranial
procedures as we opted for the less invasive approach of systemic injection. The use of direct
intracranial administration of therapeutic ligands as well as pharmacological inactivation or lesioning
could have offered powerful insights into how key regions are affected by D4 receptor agents. Thus, we
were unable to localize which areas of interest were affected, and more importantly, which regions
were responsible for the results found in this study. As such, this would be a subject of great interest in
future studies.
Additionally, the D4 receptor agents used in this study were administered only once during
testing sessions to detect immediate and acute effects, but did not examine their chronic effects. This
study could have investigated the effects of sustained pharmacological modulation of D4 receptors on
alcohol self-administration and reinstatement procedures, providing further insights on long-term
effects on alterations of neuroplasticity as well as predictive validity for potential future clinical
investigations. And so, conducting an additional set of experiments would have strengthened the
current conclusions.
The inconsistent effects of reinstatement in this study was a significant limitation in and of itself,
in particular, the results obtained by cue and the fact that the data had to be distilled. For instance, the
results of the second vehicle test under L-745,870 appeared to reinstate to similar levels as the first
vehicle test, but did not do so under PD 168,077 testing. Although this could have been due to a
sustained effect of PD 168,077 attenuating reinstatement during the second vehicle test, because of the

78
relatively low proportion of animals reinstating, it is difficult to draw any further conclusions. A possible
explanation for low response rates is that cue acquisition was not strong enough, however, other
studies have been found to robustly reproduce cue-induced reinstatement. Because of these
inconsistencies, it is challenging to affirm the certainty of the effects obtained from the reinstatement
procedures as a whole.
4.6 Conclusions The main findings from this study demonstrate that the D4 receptor is associated with
motivational processes of alcohol intake, as well as stress-mediated relapse behaviours, while sparing
the incentive for natural rewards. The therapeutic effects are generally elicited by D4 receptor blockade
whereas activation seems to have no effects on alcohol-related behaviours. Thus, pharmacological
strategies involving D4 receptor modulation may be plausible for substance dependence without the
potential for abuse or adverse side effects.
These findings contribute to the current literature as it adds another means of understanding
the crucial role of D4 receptors on addictive disorders on animals, and by extension, the potential for
humans. And with sufficiently compiled evidence, the field can move forward from gene association
studies and towards the progression of clinical trials of D4 ligands on addictive disorders. However, it
should also be taken into consideration that the overall role of the D4 receptor is incredibly intricate and
thus, will be difficult to completely understand its full nature.
4.7 Future Directions The natural prospective direction for this line of research is to pursue a deeper comprehension
of how the D4 receptor is involved in cognitive processes and behaviours relevant to addictive disorders.
And in doing so, the therapeutic landscape in the near future may see patients with AUDs utilizing non-
invasive pharmacological interventions targeting the D4 receptor in conjunction with current
psychosocial interventions, such as cognitive behavioural therapy (CBT), for optimal efficacy.

79
Although the work presented here, along with the recent literature, has established a critical
role for the D4 receptor in the neurocircuitry of addiction, several research goals have now become
more apparent. Further work remains to be conducted to identify the distinct roles of the D4 receptor
with respect to the different subregions of the brain as well as their directly interconnected
glutamatergic and GABAergic projections. And while this pursuit would seemingly require an endless
amount of neurophysiological and behavioural research in the preclinical sector, the immediate goals
are to further characterize the existing D4 neurocircuits involved in addiction.
Ultimately, the long-term goal is to make sufficient advancements in the preclinical sector to
warrant the initiation of early phase clinical trials of D4 receptor modulation as a potential therapeutic
strategy for addictive disorders including AUDs.

80
References
Ahlenius, S. (1979). An analysis of behavioural effects produced by drug-induced changes of
dopaminergic neurotransmission in the brain. Scandinavian Journal of Psychology, 20(1), 59–64.
https://doi.org/10.1111/j.1467-9450.1979.tb00683.x
Alexander, B. K., & Schweighofer, A. R. (1988). Defining" addiction.". Canadian Psychology/Psychologie
Canadienne, 29(2), 151.
American Psychiatric Association (Ed.). (2013). Diagnostic and statistical manual of mental disorders:
DSM-5 (5th ed). Washington, D.C: American Psychiatric Association.
Appel, S. B., Liu, Z., McElvain, M. A., & Brodie, M. S. (2003). Ethanol Excitation of Dopaminergic Ventral
Tegmental Area Neurons Is Blocked by Quinidine. Journal of Pharmacology and Experimental
Therapeutics, 306(2), 437–446. https://doi.org/10.1124/jpet.103.050963
Asghari, V., Sanyal, S., Buchwaldt, S., Paterson, A., Jovanovic, V., & Van Tol, H. H. M. (1995). Modulation
of Intracellular Cyclic AMP Levels by Different Human Dopamine D4 Receptor Variants. Journal
of Neurochemistry, 65(3), 1157–1165. https://doi.org/10.1046/j.1471-4159.1995.65031157.x
Asghari, V., Schoots, O., van Kats, S., Ohara, K., Jovanovic, V., Guan, H.-C., … Van Tol, H. H. (1994).
Dopamine D4 receptor repeat: analysis of different native and mutant forms of the human and
rat genes. Molecular Pharmacology, 46(2), 364–373.
Barger, G., & Dale, H. H. (1910). Chemical structure and sympathomimetic action of amines. The Journal
of Physiology, 41(1–2), 19–59.
Barger, G., & Ewins, A. J. (1910). CCXXXVII.—Some phenolic derivatives of β-phenylethylamine. Journal
of the Chemical Society, Transactions, 97(0), 2253–2261.
https://doi.org/10.1039/CT9109702253

81
Barnes, T. D., Kubota, Y., Hu, D., Jin, D. Z., & Graybiel, A. M. (2005). Activity of striatal neurons reflects
dynamic encoding and recoding of procedural memories. Nature, 437(7062), 1158–1161.
https://doi.org/10.1038/nature04053
Bergman, J., Roof, R. A., Furman, C. A., Conroy, J. L., Mello, N. K., Sibley, D. R., & Skolnick, P. (2013).
Modification of cocaine self-administration by buspirone (buspar®): potential involvement of D3
and D4 dopamine receptors. The International Journal of Neuropsychopharmacology, 16(2),
445–458. https://doi.org/10.1017/S1461145712000661
Bertler, Å., & Rosengren, E. (1959). Occurrence and distribution of dopamine in brain and other tissues.
Experientia, 15(1), 10–11. https://doi.org/10.1007/BF02157069
Blaschko, H. (1939). The specific action of L-dopa decarboxylase. The Journal of Physiology, 96, 50–51.
https://doi.org/10.1113/jphysiol.1939.sp003786
Blaschko, H., Richter, D., & Schlossmann, H. (1937). The oxidation of adrenaline and other amines.
Biochemical Journal, 31(12), 2187.
Boffetta, P., & Hashibe, M. (2006). Alcohol and cancer. The Lancet Oncology, 7(2), 149–156.
https://doi.org/10.1016/S1470-2045(06)70577-0
Bondy, B., de Jonge, S., Pander, S., Primbs, J., & Ackenheil, M. (1996). Identification of dopamine D4
receptor mRNA in circulating human lymphocytes using nested polymerase chain reaction.
Journal of Neuroimmunology, 71(1), 139–144. https://doi.org/10.1016/S0165-5728(96)00148-8
Brodie, M. S., McElvain, M. A., Bunney, E. B., & Appel, S. B. (1999). Pharmacological Reduction of Small
Conductance Calcium-Activated Potassium Current (SK) Potentiates the Excitatory Effect of
Ethanol on Ventral Tegmental Area Dopamine Neurons. Journal of Pharmacology and
Experimental Therapeutics, 290(1), 325–333.

82
Brodie, M. S., Shefner, S. A., & Dunwiddie, T. V. (1990). Ethanol increases the firing rate of dopamine
neurons of the rat ventral tegmental area in vitro. Brain Research, 508(1), 65–69.
https://doi.org/10.1016/0006-8993(90)91118-Z
Budygin, E. A., Phillips, P. E., Wightman, R. M., & Jones, S. R. (2001). Terminal effects of ethanol on
dopamine dynamics in rat nucleus accumbens: an in vitro voltammetric study. Synapse (New
York, N.Y.), 42(2), 77–79. https://doi.org/10.1002/syn.1101
Bunzow, J. R., Tol, H. H. M. V., Grandy, D. K., Albert, P., Salon, J., Christie, M., … Civelli, O. (1988). Cloning
and expression of a rat D2 dopamine receptor cDNA. Nature, 336(6201), 783–787.
https://doi.org/10.1038/336783a0
Caine, S. B., Negus, S. S., Mello, N. K., Patel, S., Bristow, L., Kulagowski, J., … Borrelli, E. (2002). Role of
Dopamine D2-like Receptors in Cocaine Self-Administration: Studies with D2 Receptor Mutant
Mice and Novel D2 Receptor Antagonists. Journal of Neuroscience, 22(7), 2977–2988.
Cardinal, R. N., & Everitt, B. J. (2004). Neural and psychological mechanisms underlying appetitive
learning: links to drug addiction. Current Opinion in Neurobiology, 14(2), 156–162.
https://doi.org/10.1016/j.conb.2004.03.004
Carlsson, A. (1959). The Occurrence, Distribution and Physiological Role of Catecholamines in the
Nervous System. Pharmacological Reviews, 11(2), 490–493.
Carlsson, A., & Lindqvist, M. (1963). Effect of Chlorpromazine or Haloperidol on Formation of 3-
Methoxytyramine and Normetanephrine in Mouse Brain. Acta Pharmacologica et Toxicologica,
20(2), 140–144. https://doi.org/10.1111/j.1600-0773.1963.tb01730.x
Catalano, M., Nobile, M., Novelli, E., Nöthen, M. M., & Smeraldi, E. (1993). Distribution of a novel
mutation in the first exon of the human dopamine D4 receptor gene in psychotic patients.
Biological Psychiatry, 34(7), 459–464. https://doi.org/10.1016/0006-3223(93)90236-7

83
Chang, F.-M., Kidd, J. R., Livak, K. J., Pakstis, A. J., & Kidd, K. K. (1996). The world-wide distribution of
allele frequencies at the human dopamine D4 receptor locus. Human Genetics, 98(1), 91–101.
https://doi.org/10.1007/s004390050166
Chiamulera, C., Valerio, E., & Tessari, M. (1995). Resumption of ethanol-seeking behaviour in rats.
Behavioural Pharmacology, 6(1), 32–39.
Cocker, P. J., Le Foll, B., Rogers, R. D., & Winstanley, C. A. (2014). A Selective Role for Dopamine D4
Receptors in Modulating Reward Expectancy in a Rodent Slot Machine Task. Biological
Psychiatry, 75(10), 817–824. https://doi.org/10.1016/j.biopsych.2013.08.026
Cohen, A. I., Todd, R. D., Harmon, S., & O’Malley, K. L. (1992). Photoreceptors of mouse retinas possess
D4 receptors coupled to adenylate cyclase. Proceedings of the National Academy of Sciences of
the United States of America, 89(24), 12093.
Collins, D. M., & Myers, R. D. (1987). Buspirone attenuates volitional alcohol intake in the chronically
drinking monkey. Alcohol, 4(1), 49–56. https://doi.org/10.1016/0741-8329(87)90060-7
Connor, P. D., & Streissguth, A. P. (1996). Effects of prenatal exposure to alcohol across the life span.
Alcohol Research and Health, 20(3), 170.
Cornish, J. L., & Kalivas, P. W. (2000). Glutamate Transmission in the Nucleus Accumbens Mediates
Relapse in Cocaine Addiction. Journal of Neuroscience, 20(15), RC89-RC89.
Corrao, G., Bagnardi, V., Zambon, A., & Torchio, P. (1998). Meta-Analysis of Alcohol Intake in Relation to
Risk of Liver Cirrhosis. Alcohol and Alcoholism, 33(4), 381–392.
Corrigall, W. A., & Coen, K. M. (1989). Nicotine maintains robust self-administration in rats on a limited-
access schedule. Psychopharmacology, 99(4), 473–478. https://doi.org/10.1007/BF00589894
Costanza, R. M., & Terry, P. (1998). The dopamine D4 receptor antagonist L-745,870: effects in rats
discriminating cocaine from saline. European Journal of Pharmacology, 345(2), 129–132.
https://doi.org/10.1016/S0014-2999(97)01603-8

84
Cotzias, G. C., Van Woert, M. H., & Schiffer, L. M. (1967). Aromatic Amino Acids and Modification of
Parkinsonism. New England Journal of Medicine, 276(7), 374–379.
https://doi.org/10.1056/NEJM196702162760703
Davies, M. (2003). The role of GABAA receptors in mediating the effects of alcohol in the central nervous
system. Journal of Psychiatry and Neuroscience, 28(4), 263–274.
de Wit, H., & Stewart, J. (1981). Reinstatement of cocaine-reinforced responding in the rat.
Psychopharmacology, 75(2), 134–143. https://doi.org/10.1007/BF00432175
Dearry, A., Gingrich, J. A., Falardeau, P., Fremeau, R. T., Bates, M. D., & Caron, M. G. (1990). Molecular
cloning and expression of the gene for a human D1 dopamine receptor. Nature, 347(6288), 72–
76. https://doi.org/10.1038/347072a0
Defagot, M. C., Falzone, T. L., Low, M. J., Grandy, D. K., Rubinstein, M., & Antonelli, M. C. (2000).
Quantitative analysis of the dopamine D4 receptor in the mouse brain. Journal of Neuroscience
Research, 59(2), 202–208. https://doi.org/10.1002/(SICI)1097-4547(20000115)59:2<202::AID-
JNR6>3.0.CO;2-5
Degenhardt, L., Whiteford, H. A., Ferrari, A. J., Baxter, A. J., Charlson, F. J., Hall, W. D., … Vos, T. (2013).
Global burden of disease attributable to illicit drug use and dependence: findings from the
Global Burden of Disease Study 2010. The Lancet, 382(9904), 1564–1574.
https://doi.org/10.1016/S0140-6736(13)61530-5
Di Ciano, P., & Everitt, B. J. (2004). Contribution of the ventral tegmental area to cocaine-seeking
maintained by a drug-paired conditioned stimulus in rats. European Journal of Neuroscience,
19(6), 1661–1667. https://doi.org/10.1111/j.1460-9568.2004.03232.x
Di Ciano, P., Pushparaj, A., Kim, A., Hatch, J., Masood, T., Ramzi, A., … Le Foll, B. (2015). The Impact of
Selective Dopamine D2, D3 and D4 Ligands on the Rat Gambling Task. PLOS ONE, 10(9),
e0136267. https://doi.org/10.1371/journal.pone.0136267

85
Dulawa, S. C., Grandy, D. K., Low, M. J., Paulus, M. P., & Geyer, M. A. (1999). Dopamine D4 Receptor-
Knock-Out Mice Exhibit Reduced Exploration of Novel Stimuli. Journal of Neuroscience, 19(21),
9550–9556.
Dunn, A. J., & Berridge, C. W. (1990). Physiological and behavioral responses to corticotropin-releasing
factor administration: is CRF a mediator of anxiety or stress responses? Brain Research Reviews,
15(2), 71–100. https://doi.org/10.1016/0165-0173(90)90012-D
Durbec, J. P., & Sarles, H. (1978). Multicenter Survey of the Etiology of Pancreatic Diseases. Digestion,
18(5–6), 337–350. https://doi.org/10.1159/000198221
Ebstein, R. P., Novick, O., Umansky, R., Priel, B., Osher, Y., Blaine, D., … Belmaker, R. H. (1996). Dopamine
D4 receptor (D4DR) exon III polymorphism associated with the human personality trait of
Novelty Seeking. Nature Genetics, 12(1), 78–80. https://doi.org/10.1038/ng0196-78
Fan, X., Xu, M., & Hess, E. J. (2010). D2 dopamine receptor subtype-mediated hyperactivity and
amphetamine responses in a model of ADHD. Neurobiology of Disease, 37(1), 228–236.
https://doi.org/10.1016/j.nbd.2009.10.009
Feltenstein, M. W., Ghee, S. M., & See, R. E. (2012). Nicotine self-administration and reinstatement of
nicotine-seeking in male and female rats. Drug and Alcohol Dependence, 121(3), 240–246.
https://doi.org/10.1016/j.drugalcdep.2011.09.001
Gago, B., Fuxe, K., Agnati, L., Peñafiel, A., De La Calle, A., & Rivera, A. (2007). Dopamine D4 receptor
activation decreases the expression of μ-opioid receptors in the rat striatum. The Journal of
Comparative Neurology, 502(3), 358–366. https://doi.org/10.1002/cne.21327
Garbutt, J. C. (2010). Efficacy and tolerability of naltrexone in the management of alcohol dependence.
Current Pharmaceutical Design, 16(19), 2091–2097.
Gazi, L., Bobirnac, I., Danzeisen, M., Schüpbach, E., Langenegger, D., Sommer, B., … Schoeffter, P. (1999).
Receptor density as a factor governing the efficacy of the dopamine D4 receptor ligands, L-

86
745,870 and U-101958 at human recombinant D4.4 receptors expressed in CHO cells. British
Journal of Pharmacology, 128(3), 613–620. https://doi.org/10.1038/sj.bjp.0702849
George, S. R., Cheng, R., Nguyen, T., Israel, Y., & Odowd, B. F. (1993). Polymorphisms of the D4
Dopamine Receptor Alleles in Chronic Alcoholism. Biochemical and Biophysical Research
Communications, 196(1), 107–114. https://doi.org/10.1006/bbrc.1993.2222
Girault, J.-A., & Greengard, P. (2004). The Neurobiology of Dopamine Signaling. Archives of Neurology,
61(5), 641–644. https://doi.org/10.1001/archneur.61.5.641
Glase, S. A., Akunne, H. C., Georgic, L. M., Heffner, T. G., MacKenzie, R. G., Manley, P. J., … Wise, L. D.
(1997). Substituted [(4-Phenylpiperazinyl)- methyl]benzamides: Selective Dopamine D4
Agonists. Journal of Medicinal Chemistry, 40(12), 1771–1772.
https://doi.org/10.1021/jm970021c
Goldstein, R. Z., & Volkow, N. D. (2002). Drug Addiction and Its Underlying Neurobiological Basis:
Neuroimaging Evidence for the Involvement of the Frontal Cortex. The American Journal of
Psychiatry, 159(10), 1642–1652.
Grady, D. L., Chi, H.-C., Ding, Y.-C., Smith, M., Wang, E., Schuck, S., … Moyzis, R. K. (2003). High
prevalence of rare dopamine receptor D4 alleles in children diagnosed with attention-deficit
hyperactivity disorder. Molecular Psychiatry, 8(5), 536–545.
https://doi.org/10.1038/sj.mp.4001350
Grahame, N. J., & Grose, A. M. (2003). Blood alcohol concentrations after scheduled access in high-
alcohol-preferring mice. Alcohol, 31(1–2), 99–104.
https://doi.org/10.1016/j.alcohol.2003.08.002
Graziane, N. M., Yuen, E. Y., & Yan, Z. (2009). Dopamine D4 Receptors Regulate GABAA Receptor
Trafficking via an Actin/Cofilin/Myosin-dependent Mechanism. The Journal of Biological
Chemistry, 284(13), 8329–8336. https://doi.org/10.1074/jbc.M807387200

87
Greengard, P. (2001). The Neurobiology of Slow Synaptic Transmission. Science, 294(5544), 1024–1030.
https://doi.org/10.1126/science.294.5544.1024
Griffiths, M. (1991). Psychobiology of the Near-Miss in Fruit Machine Gambling. The Journal of
Psychology, 125(3), 347–357. https://doi.org/10.1080/00223980.1991.10543298
Grimm, J. W., Hope, B. T., Wise, R. A., & Shaham, Y. (2001). Incubation of cocaine craving after
withdrawal. Nature, 412(6843), 141–142. https://doi.org/10.1038/35084134
Grimm, J. W., & See, R. E. (2000). Dissociation of Primary and Secondary Reward-Relevant Limbic Nuclei
in an Animal Model of Relapse. Neuropsychopharmacology, 22(5), 473–479.
https://doi.org/10.1016/S0893-133X(99)00157-8
Halley, A. C., Boretsky, M., Puts, D. A., & Shriver, M. (2016). Self-Reported Sexual Behavioral Interests
and Polymorphisms in the Dopamine Receptor D4 (DRD4) Exon III VNTR in Heterosexual Young
Adults. Archives of Sexual Behavior, 45(8), 2091–2100. https://doi.org/10.1007/s10508-015-
0646-6
Hasanein, P., Mirazi, N., & Javanmardi, K. (2008). GABAA receptors in the central nucleus of amygdala
(CeA) affect on pain modulation. Brain Research, 1241(Supplement C), 36–41.
https://doi.org/10.1016/j.brainres.2008.09.041
Heidbreder, C. (2011). Advances in Animal Models of Drug Addiction. In J. J. Hagan (Ed.), Molecular and
Functional Models in Neuropsychiatry (pp. 213–250). Springer Berlin Heidelberg.
https://doi.org/10.1007/7854_2010_107
Heilig, M., & Koob, G. F. (2007). A key role for corticotropin-releasing factor in alcohol dependence.
Trends in Neurosciences, 30(8), 399–406. https://doi.org/10.1016/j.tins.2007.06.006
Heimer, L., Alheid, de O. J., Groenewegen, H., Haber, S., Harlan, R., & Zahm, D. (1997). The accumbens:
beyond the core-shell dichotomy. The Journal of Neuropsychiatry and Clinical Neurosciences,
9(3), 354–381. https://doi.org/10.1176/jnp.9.3.354

88
Holtz, P., Credner, K., & Koepp, W. (1942). Die enzymatische Entstehung von Oxytyramin im Organismus
und die physiologische Bedeutung der Dopadecarboxylase. Naunyn-Schmiedebergs Archiv für
experimentelle Pathologie und Pharmakologie, 200(2–5), 356–388.
https://doi.org/10.1007/BF01860725
Holtz, P., Heise, R., & Lüdtke, K. (1938). Fermentativer Abbau von l-Dioxyphenylalanin (Dopa) durch
Niere. Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie, 191(1),
87–118. https://doi.org/10.1007/BF01994628
Hornykiewicz, O. (1986). A quarter century of brain dopamine research. London: Macmillan. Retrieved
from
http://books.google.com/books?hl=en&lr=&id=vNllCwAAQBAJ&oi=fnd&pg=PA3&dq=%22(1910)
+as+a+weak+sympathomimetic+(vasopressor)+drug.+It+is%22+%22showed+that+dopamine+wa
s+the+reaction+product+of+the%22+%22immediately+accepted,+and+as+late+as+in+1952+so
me%22+&ots=dBhz2fnP21&sig=GzvjP_LcQeFpy7sm0ww-bLI18VQ
Hutchison, K. E., LaChance, H., Niaura, R., Bryan, A., & Smolen, A. (2002). The DRD4 VNTR polymorphism
influences reactivity to smoking cues. Journal of Abnormal Psychology, 111(1), 134–143.
https://doi.org/10.1037/0021-843X.111.1.134
Hutchison, K. E., McGeary, J., Smolen, A., Bryan, A., & Swift, R. M. (2002). The DRD4 VNTR polymorphism
moderates craving after alcohol consumption. Health Psychology, 21(2), 139–146.
https://doi.org/10.1037//0278-6133.21.2.139
Imperato, A., & Di Chiara, G. (1986). Preferential stimulation of dopamine release in the nucleus
accumbens of freely moving rats by ethanol. Journal of Pharmacology and Experimental
Therapeutics, 239(1), 219–228.

89
Imperato, A., Mulas, A., & Di Chiara, G. (1986). Nicotine preferentially stimulates dopamine release in
the limbic system of freely moving rats. European Journal of Pharmacology, 132(2–3), 337–338.
https://doi.org/10.1016/0014-2999(86)90629-1
Ivar Walaas, S., Aswad, D. W., & Greengard, P. (1983). A dopamine- and cyclic AMP-regulated
phosphoprotein enriched in dopamine-innervated brain regions. Nature, 301(5895), 69–71.
https://doi.org/10.1038/301069a0
Iversen, L. L. (1975). Dopamine Receptors in the Brain. Science, 188(4193), 1084–1089.
Iversen, S. D., & Iversen, L. L. (2007). Dopamine: 50 years in perspective. Trends in Neurosciences, 30(5),
188–193. https://doi.org/10.1016/j.tins.2007.03.002
Jentsch, J. D., Redmond, D. E., Elsworth, J. D., Taylor, J. R., Youngren, K. D., & Roth, R. H. (1997). Enduring
Cognitive Deficits and Cortical Dopamine Dysfunction in Monkeys After Long-Term
Administration of Phencyclidine. Science, 277(5328), 953–955.
https://doi.org/10.1126/science.277.5328.953
Jentsch, J. D., & Taylor, J. R. (1999). Impulsivity resulting from frontostriatal dysfunction in drug abuse:
implications for the control of behavior by reward-related stimuli. Psychopharmacology, 146(4),
373–390.
Johnstone, E. C., Crow, T. J., Frith, C. D., Carney, M. W., & Price, J. S. (1978). Mechanism of the
antipsychotic effect in the treatment of acute schizophrenia. Lancet (London, England), 1(8069),
848–851.
Jovanovic, V., Guan, H. C., & Van Tol, H. H. (1999). Comparative pharmacological and functional analysis
of the human dopamine D4.2 and D4.10 receptor variants. Pharmacogenetics, 9(5), 561–568.
June, H. L., & Gilpin, N. W. (2010). Operant Self-Administration Models for Testing the
Neuropharmacological Basis of Ethanol Consumption in Rats. Current Protocols in Neuroscience /

90
Editorial Board, Jacqueline N. Crawley ... [et Al.], CHAPTER, Unit-9.1226.
https://doi.org/10.1002/0471142301.ns0912s51
Kalivas, P. W., & McFarland, K. (2003). Brain circuitry and the reinstatement of cocaine-seeking behavior.
Psychopharmacology, 168(1–2), 44–56. https://doi.org/10.1007/s00213-003-1393-2
Kalivas, P. W., & O’Brien, C. (2007). Drug Addiction as a Pathology of Staged Neuroplasticity.
Neuropsychopharmacology, 33(1), 166–180. https://doi.org/10.1038/sj.npp.1301564
Kalivas, P. W., Peters, J., & Knackstedt, L. (2006). Animal Models and Brain Circuits in Drug Addiction.
Molecular Interventions, 6(6), 339. https://doi.org/10.1124/mi.6.6.7
Kamakura, S., Iwaki, A., Matsumoto, M., & Fukumaki, Y. (1997). Cloning and characterization of the 5′-
flanking region of the human dopamine D4 receptor gene. Biochemical and Biophysical Research
Communications, 235(2), 321–326.
Kapur, S. (2004). How antipsychotics become anti-‘psychotic’ – from dopamine to salience to psychosis.
Trends in Pharmacological Sciences, 25(8), 402–406. https://doi.org/10.1016/j.tips.2004.06.005
Kebabian, J. W., & Calne, D. B. (1979). Multiple receptors for dopamine. Nature, 277(5692), 93–96.
https://doi.org/10.1038/277093a0
Kebabian, J. W., Petzold, G. L., & Greengard, P. (1972). Dopamine-Sensitive Adenylate Cyclase in Caudate
Nucleus of Rat Brain, and Its Similarity to the “Dopamine Receptor.” Proceedings of the National
Academy of Sciences of the United States of America, 69(8), 2145–2149.
Khan, Z. U., Gutiérrez, A., Martín, R., Peñafiel, A., Rivera, A., & De La Calle, A. (1998). Differential regional
and cellular distribution of dopamine D2-like receptors: An immunocytochemical study of
subtype-specific antibodies in rat and human brain. The Journal of Comparative Neurology,
402(3), 353–371. https://doi.org/10.1002/(SICI)1096-9861(19981221)402:3<353::AID-
CNE5>3.0.CO;2-4

91
Klatsky, A. L. (1999). Moderate drinking and reduced risk of heart disease. Alcohol Research and Health,
23(1), 15–24.
Koob, G. F., Heinrichs, S. C., Menzaghi, F., Pich, E. M., & Britton, K. T. (1994). Corticotropin releasing
factor, stress and behavior. Seminars in Neuroscience, 6(4), 221–229.
https://doi.org/10.1006/smns.1994.1029
Koob, G. F., & Le Moal, M. (2001). Drug Addiction, Dysregulation of Reward, and Allostasis.
Neuropsychopharmacology, 24(2), 97–129. https://doi.org/10.1016/S0893-133X(00)00195-0
Koob, G. F., & Le Moal, M. (2006). CHAPTER 9 - Neurobiological Theories of Addiction. In Neurobiology of
Addiction (pp. 377–428). London: Academic Press. https://doi.org/10.1016/B978-012419239-
3/50046-1
Kostowski, W., & Dyr, W. (1992). Effects of 5-HT-1A receptor agonists on ethanol preference in the rat.
Alcohol, 9(4), 283–286. https://doi.org/10.1016/0741-8329(92)90067-K
Kulagowski, J. J., Broughton, H. B., Curtis, N. R., Mawer, I. M., Ridgill, M. P., Baker, R., … Leeson, P. D.
(1996). 3-[[4-(4-Chlorophenyl)piperazin-1-yl]methyl]-1H-pyrrolo[2,3-b]pyridine: An Antagonist
with High Affinity and Selectivity for the Human Dopamine D4 Receptor. Journal of Medicinal
Chemistry, 39(10), 1941–1942. https://doi.org/10.1021/jm9600712
Lanau, F., Zenner, M.-T., Civelli, O., & Hartman, D. S. (1997). Epinephrine and Norepinephrine Act as
Potent Agonists at the Recombinant Human Dopamine D4 Receptor. Journal of Neurochemistry,
68(2), 804–812. https://doi.org/10.1046/j.1471-4159.1997.68020804.x
Lane, J. R., Powney, B., Wise, A., Rees, S., & Milligan, G. (2008). G protein coupling and ligand selectivity
of the D2L and D3 dopamine receptors. The Journal of Pharmacology and Experimental
Therapeutics, 325(1), 319–330. https://doi.org/10.1124/jpet.107.134296

92
Laruelle, M., Abi-Dargham, A., Dyck, C. H. van, Rosenblatt, W., Zea-Ponce, Y., Zoghbi, S. S., … Innis, R. B.
(1995). SPECT Imaging of Striatal Dopamine Release after Amphetamine Challenge. Journal of
Nuclear Medicine, 36(7), 1182–1190.
Laucht, M., Becker, K., Blomeyer, D., & Schmidt, M. H. (2007). Novelty Seeking Involved in Mediating the
Association Between the Dopamine D4 Receptor Gene Exon III Polymorphism and Heavy
Drinking in Male Adolescents: Results from a High-Risk Community Sample. Biological
Psychiatry, 61(1), 87–92. https://doi.org/10.1016/j.biopsych.2006.05.025
Lavine, N., Ethier, N., Oak, J. N., Pei, L., Liu, F., Trieu, P., … Van Tol, H. H. M. (2002). G protein-coupled
receptors form stable complexes with inwardly rectifying potassium channels and adenylyl
cyclase. The Journal of Biological Chemistry, 277(48), 46010–46019.
https://doi.org/10.1074/jbc.M205035200
Lê, A. D., Quan, B., Juzytch, W., Fletcher, P. J., Joharchi, N., & Shaham, Y. (1998). Reinstatement of
alcohol-seeking by priming injections of alcohol and exposure to stress in rats.
Psychopharmacology, 135(2), 169–174.
Leri, F., Flores, J., Rodaros, D., & Stewart, J. (2002). Blockade of Stress-Induced But Not Cocaine-Induced
Reinstatement by Infusion of Noradrenergic Antagonists into the Bed Nucleus of the Stria
Terminalis or the Central Nucleus of the Amygdala. Journal of Neuroscience, 22(13), 5713–5718.
Levine, H. G. (1978). The discovery of addiction. Changing conceptions of habitual drunkenness in
America. Journal of Studies on Alcohol, 39(1), 143–174.
https://doi.org/10.15288/jsa.1978.39.143
Lewis, C. T., & Short, C. (1879). A Latin Dictionary: Founded on Andrews edition of Freund’s Latin
Dictionary. Retrieved December 28, 2016, from
http://www.perseus.tufts.edu/hopper/text?doc=Perseus:text:1999.04.0059

93
Lim, S. S., Vos, T., Flaxman, A. D., Danaei, G., Shibuya, K., Adair-Rohani, H., … Ezzati, M. (2012). A
comparative risk assessment of burden of disease and injury attributable to 67 risk factors and
risk factor clusters in 21 regions, 1990–2010: a systematic analysis for the Global Burden of
Disease Study 2010. Lancet, 380(9859), 2224–2260. https://doi.org/10.1016/S0140-
6736(12)61766-8
Littleton, J. (2000). Can craving be modeled in animals? The relapse prevention perspective. Addiction,
95(8s2), 83–90. https://doi.org/10.1046/j.1360-0443.95.8s2.18.x
Littleton, J., & Zieglgänsberger, W. (2003). Pharmacological Mechanisms of Naltrexone and Acamprosate
in the Prevention of Relapse in Alcohol Dependence. The American Journal on Addictions, 12,
s3–s11. https://doi.org/10.1111/j.1521-0391.2003.tb00492.x
Lobo, I. A., & Harris, R. A. (2008). GABAA receptors and alcohol. Pharmacology, Biochemistry, and
Behavior, 90(1), 90–94. https://doi.org/10.1016/j.pbb.2008.03.006
Louilot, A., Le Moal, M., & Simon, H. (1989). Opposite influences of dopaminergic pathways to the
prefrontal cortex or the septum on the dopaminergic transmission in the nucleus accumbens. An
in vivo voltammetric study. Neuroscience, 29(1), 45–56. https://doi.org/10.1016/0306-
4522(89)90331-X
Luciano, M., Zhu, G., Kirk, K. m., Whitfield, J. b., Butler, R., Heath, A. c., … Martin, N. g. (2004). Effects of
dopamine receptor D4 variation on alcohol and tobacco use and on novelty seeking:
Multivariate linkage and association analysis. American Journal of Medical Genetics Part B:
Neuropsychiatric Genetics, 124B(1), 113–123. https://doi.org/10.1002/ajmg.b.20077
Maisel, N. C., Blodgett, J. C., Wilbourne, P. L., Humphreys, K., & Finney, J. W. (2013). Meta-analysis of
naltrexone and acamprosate for treating alcohol use disorders: When are these medications
most helpful? Addiction (Abingdon, England), 108(2), 275–293. https://doi.org/10.1111/j.1360-
0443.2012.04054.x

94
Makara, G. B., Harris, M. C., & Spyer, K. M. (1972). Identification and distribution of tuberoinfundibular
neurones. Brain Research, 40(2), 283–290. https://doi.org/10.1016/0006-8993(72)90134-5
Mamiya, T., Matsumura, T., & Ukai, M. (2004). Effects of L-745,870, a Dopamine D4 Receptor
Antagonist, on Naloxone-Induced Morphine Dependence in Mice. Annals of the New York
Academy of Sciences, 1025(1), 424–429. https://doi.org/10.1196/annals.1316.052
Marcus, P., & Snyder, R. (1995). Reduction of comorbid substance abuse with clozapine. The American
Journal of Psychiatry, 152(6), 959.
Marona-Lewicka, D., & Nichols, D. E. (2011). Potential Serotonin 5-HT1A and Dopamine D4 Receptor
Modulation of the Discriminative Stimulus Effects of Amphetamine in Rats. Behavioural
Pharmacology, 22(5–6), 508–515. https://doi.org/10.1097/FBP.0b013e328349fc31
Marsden, C. A. (2006). Dopamine: the rewarding years. British Journal of Pharmacology, 147(Suppl 1),
S136–S144. https://doi.org/10.1038/sj.bjp.0706473
Martinez, D., Saccone, P. A., Liu, F., Slifstein, M., Orlowska, D., Grassetti, A., … Comer, S. D. (2012).
Deficits in dopamine D2 receptors and pre-synaptic dopamine in heroin dependence:
commonalities and differences with other types of addiction. Biological Psychiatry, 71(3), 192–
198. https://doi.org/10.1016/j.biopsych.2011.08.024
Masur, J., de Souza, M. L. O., & Zwicker, A. P. (1986). The excitatory effect of ethanol: Absence in rats,
no tolerance and increased sensitivity in mice. Pharmacology Biochemistry and Behavior, 24(5),
1225–1228. https://doi.org/10.1016/0091-3057(86)90175-9
Matsumoto, M., Hidaka, K., Tada, S., Tasaki, Y., & Yamaguchi, T. (1995). Full-length cDNA cloning and
distribution of human dopamine D4 receptor. Molecular Brain Research, 29(1), 157–162.
https://doi.org/10.1016/0169-328X(94)00245-A
Maurel-Remy, S., Bervoets, K., & Millan, M. J. (1995). Blockade of phencyclidine-induced
hyperlocomotion by clozapine and MDL 100,907 in rats reflects antagonism of 5-HT2A

95
receptors. European Journal of Pharmacology, 280(2), R9–R11. https://doi.org/10.1016/0014-
2999(95)00333-G
McClernon, F. J., Hutchison, K. E., Rose, J. E., & Kozink, R. V. (2007). DRD4 VNTR polymorphism is
associated with transient fMRI-BOLD responses to smoking cues. Psychopharmacology, 194(4),
433–441. https://doi.org/10.1007/s00213-007-0860-6
McFarland, K., & Kalivas, P. W. (2001). The Circuitry Mediating Cocaine-Induced Reinstatement of Drug-
Seeking Behavior. Journal of Neuroscience, 21(21), 8655–8663.
Meador-Woodruff, J. H., Haroutunian, V., Powchik, P., Davidson, M., Davis, K. L., & Watson, S. J. (1997).
Dopamine Receptor Transcript Expression in Striatum and Prefrontal and Occipital Cortex: Focal
Abnormalities in Orbitofrontal Cortex in Schizophrenia. Archives of General Psychiatry, 54(12),
1089–1095. https://doi.org/10.1001/archpsyc.1997.01830240045007
Meil, W. M., & See, R. E. (1997). Lesions of the basolateral amygdala abolish the ability of drug
associated cues to reinstate responding during withdrawal from self-administered cocaine.
Behavioural Brain Research, 87(2), 139–148. https://doi.org/10.1016/S0166-4328(96)02270-X
Mereu, G., & Gessa, G. L. (1985). Low doses of ethanol inhibit the firing of neurons in the substantia
nigra, pars reticulata: a GABAergic effect? Brain Research, 360(1), 325–330.
https://doi.org/10.1016/0006-8993(85)91249-1
Merikangas, K. R., Mehta, R. L., Molnar, B. E., Walters, E. E., Swendsen, J. D., Aguilar-Gaziola, S., …
Kessler, R. C. (1998). Comorbidity of substance use disorders with mood and anxiety disorders:
Results of the international consortium in psychiatric epidemiology. Addictive Behaviors, 23(6),
893–907. https://doi.org/10.1016/S0306-4603(98)00076-8
Meyer-Lindenberg, A., Straub, R. E., Lipska, B. K., Verchinski, B. A., Goldberg, T., Callicott, J. H., …
Weinberger, D. R. (2007). Genetic evidence implicating DARPP-32 in human frontostriatal

96
structure, function, and cognition. The Journal of Clinical Investigation, 117(3), 672–682.
https://doi.org/10.1172/JCI30413
Milner, B. (1982). Some Cognitive Effects of Frontal-Lobe Lesions in Man. Philosophical Transactions of
the Royal Society of London B: Biological Sciences, 298(1089), 211–226.
https://doi.org/10.1098/rstb.1982.0083
Mogenson, G. J., Jones, D. L., & Yim, C. Y. (1980). From motivation to action: Functional interface
between the limbic system and the motor system. Progress in Neurobiology, 14(2–3), 69–97.
https://doi.org/10.1016/0301-0082(80)90018-0
Molinoff, P. B., & Axelrod, J. (1971). Biochemistry of catecholamines. Annual Review of Biochemistry, 40,
465–500. https://doi.org/10.1146/annurev.bi.40.070171.002341
Montagu, K. A. (1957). Catechol Compounds in Rat Tissues and in Brains of Different Animals. Nature,
180(4579), 244–245. https://doi.org/10.1038/180244a0
Mrzljak, L., Bergson, C., Pappy, M., Huff, R., Levenson, R., & Goldman-Rakic, P. S. (1996). Localization of
dopamine D4 receptors in GABAergic neurons of the primate brain. Nature, 381(6579), 245–248.
https://doi.org/10.1038/381245a0
Mukamal, K. J. (2005). Alcohol and Risk for Ischemic Stroke in Men: The Role of Drinking Patterns and
Usual Beverage. Annals of Internal Medicine, 142(1), 11. https://doi.org/10.7326/0003-4819-
142-1-200501040-00007
Munafò, M. R., Yalcin, B., Willis-Owen, S. A., & Flint, J. (2008). Association of the Dopamine D4 Receptor
(DRD4) Gene and Approach-Related Personality Traits: Meta-Analysis and New Data. Biological
Psychiatry, 63(2), 197–206. https://doi.org/10.1016/j.biopsych.2007.04.006
Nagatsu, T., Levitt, M., & Udenfriend, S. (1964). Conversion of L-tyrosine to 3,4-dihydroxyphenylalanine
by cell-free preparations of brain and sympathetically innervated tissues. Biochemical and

97
Biophysical Research Communications, 14(6), 543–549. https://doi.org/10.1016/0006-
291X(64)90266-9
National Institute on Drug Abuse. (2014). Drugs, Brains, and Behavior: The Science of Addiction.
Retrieved December 28, 2016, from https://www.drugabuse.gov/publications/drugs-brains-
behavior-science-addiction/drug-abuse-addiction
Newman-Tancredi, A., Audinot-Bouchez, V., Gobert, A., & Millan, M. J. (1997). Noradrenaline and
adrenaline are high affinity agonists at dopamine D4 receptors. European Journal of
Pharmacology, 319(2), 379–383. https://doi.org/10.1016/S0014-2999(96)00985-5
Nöthen, M. M., Cichon, S., Hemmer, S., Hebebrand, J., Remschmidt, H., Lehmkuhl, G., … Propping, P.
(1994). Human dopamine D4 receptor gene: frequent occurrence of a null allele and observation
of homozygosity. Human Molecular Genetics, 3(12), 2207–2212.
https://doi.org/10.1093/hmg/3.12.2207
Nutt, D. J., Lingford-Hughes, A., Erritzoe, D., & Stokes, P. R. A. (2015). The dopamine theory of addiction:
40 years of highs and lows. Nature Reviews Neuroscience, 16(5), 305–312.
https://doi.org/10.1038/nrn3939
O’Malley, K. L., Harmon, S., Tang, L., & Todd, R. D. (1992). The rat dopamine D4 receptor: sequence,
gene structure, and demonstration of expression in the cardiovascular system. The New
Biologist, 4(2), 137–146.
Pangalos, M. N., & Davies, C. H. (2002). Understanding G protein-coupled receptors and their role in the
CNS. Oxford ; New York : Oxford University Press. Retrieved from
http://trove.nla.gov.au/version/19938326
Parssinen, T. M., & Kerner, K. (1980). Development of the disease model of drug addiction in Britain,
1870-1926. Medical History, 24(3), 275–296.

98
Patel, S., Freedman, S., Chapman, K. L., Emms, F., Fletcher, A. E., Knowles, M., … others. (1997).
Biological profile of L-745,870, a selective antagonist with high affinity for the dopamine D4
receptor. Journal of Pharmacology and Experimental Therapeutics, 283(2), 636–647.
Phillips, P. E. M., Stuber, G. D., Heien, M. L. A. V., Wightman, R. M., & Carelli, R. M. (2003). Subsecond
dopamine release promotes cocaine seeking. Nature, 422(6932), 614–618.
https://doi.org/10.1038/nature01476
Pich, E. M., Lorang, M., Yeganeh, M., Fonseca, F. R. de, Raber, J., Koob, G. F., & Weiss, F. (1995). Increase
of extracellular corticotropin-releasing factor-like immunoreactivity levels in the amygdala of
awake rats during restraint stress and ethanol withdrawal as measured by microdialysis. Journal
of Neuroscience, 15(8), 5439–5447.
Pickens, C. L., Airavaara, M., Theberge, F., Fanous, S., Hope, B. T., & Shaham, Y. (2011). Neurobiology of
the incubation of drug craving. Trends in Neurosciences, 34(8), 411–420.
https://doi.org/10.1016/j.tins.2011.06.001
Pierce, R. C., & Kumaresan, V. (2006). The mesolimbic dopamine system: The final common pathway for
the reinforcing effect of drugs of abuse? Neuroscience & Biobehavioral Reviews, 30(2), 215–238.
https://doi.org/10.1016/j.neubiorev.2005.04.016
Powell, S. B., Paulus, M. P., Hartman, D. S., Godel, T., & Geyer, M. A. (2003). RO-10-5824 is a selective
dopamine D4 receptor agonist that increases novel object exploration in C57 mice.
Neuropharmacology, 44(4), 473–481. https://doi.org/10.1016/S0028-3908(02)00412-4
Primus, R. J., Thurkauf, A., Xu, J., Yevich, E., McInerney, S., Shaw, K., … Gallagher, D. W. (1997). II.
Localization and characterization of dopamine D4 binding sites in rat and human brain by use of
the novel, D4 receptor-selective ligand [3H]NGD 94-1. The Journal of Pharmacology and
Experimental Therapeutics, 282(2), 1020–1027.

99
Quadros, I. M. H., Nobrega, J. N., Hipolide, D. C., & Souza-Formigoni, M. L. O. (2005). Increased brain
dopamine D4-like binding after chronic ethanol is not associated with behavioral sensitization in
mice. Alcohol, 37(2), 99–104. https://doi.org/10.1016/j.alcohol.2005.12.001
Rehm, J., Room, R., Monteiro, M., Gmel, G., Graham, K., Rehn, N., … Jernigan, D. (2004). Alcohol Use. In
Comparative quantification of health risks: global and regional burden of disease attributable to
selected major risk factors (pp. 959–1108). Geneva: World Health Organization. Retrieved from
http://www.who.int/publications/cra/chapters/volume1/0959-1108.pdf
Reynolds, K., Lewis, B., Nolen, J. D. L., Kinney, G. L., Sathya, B., & He, J. (2003). Alcohol Consumption and
Risk of Stroke: A Meta-analysis. JAMA, 289(5), 579–588.
https://doi.org/10.1001/jama.289.5.579
Reynolds, S. M., & Berridge, K. C. (2002). Positive and Negative Motivation in Nucleus Accumbens Shell:
Bivalent Rostrocaudal Gradients for GABA-Elicited Eating, Taste “Liking”/“Disliking” Reactions,
Place Preference/Avoidance, and Fear. Journal of Neuroscience, 22(16), 7308–7320.
Rezvani, A. H., Overstreet, D. H., & Janowsky, D. S. (1991). Drug-induced reductions in ethanol intake in
alcohol preferring and fawn-hooded rats, 433–437.
Risinger, F. O., Malott, D. H., Prather, L. K., Niehus, D. R., & Cunningham, C. L. (1994). Motivational
properties of ethanol in mice selectively bred for ethanol-induced locomotor differences.
Psychopharmacology, 116(2), 207–216. https://doi.org/10.1007/BF02245064
Risinger, F. O., & Oakes, R. A. (1996). Dose- and conditioning trial-dependent ethanol-induced
conditioned place preference in Swiss-Webster mice. Pharmacology Biochemistry and Behavior,
55(1), 117–123. https://doi.org/10.1016/0091-3057(96)00069-X
Rivera, A., Cuéllar, B., Girón, F. J., Grandy, D. K., De La Calle, A., & Moratalla, R. (2002). Dopamine D4
receptors are heterogeneously distributed in the striosomes/matrix compartments of the

100
striatum. Journal of Neurochemistry, 80(2), 219–229. https://doi.org/10.1046/j.0022-
3042.2001.00702.x
Rivera, A., Gago, B., Suárez-Boomgaard, D., Yoshitake, T., Roales-Buján, R., Valderrama-Carvajal, A., …
Fuxe, K. (2016). Dopamine D4 receptor stimulation prevents nigrostriatal dopamine pathway
activation by morphine: relevance for drug addiction. Addiction Biology, n/a-n/a.
https://doi.org/10.1111/adb.12407
Robbins, T. W., & Everitt, B. J. (1992). Functions of dopamine in the dorsal and ventral striatum.
Seminars in Neuroscience, 4(2), 119–127. https://doi.org/10.1016/1044-5765(92)90010-Y
Roberts, D. C. S., & Zito, K. A. (1987). Interpretation of Lesion Effects on Stimulant Self-Administration. In
M. A. Bozarth (Ed.), Methods of assessing the reinforcing properties of abused drugs (pp. 87–
103). New York: Springer-Verlag. Retrieved from http://addictionscience.net/MARPADc04.htm
Robledo, P., Robbins, T. W., & Everitt, B. J. (1996). Effects of excitotoxic lesions of the central amygdaloid
nucleus on the potentiation of reward-related stimuli by intra-accumbens amphetamine.
Behavioral Neuroscience, 110(5), 981–990.
Rodd, Z. A., Melendez, R. I., Bell, R. L., Kuc, K. A., Zhang, Y., Murphy, J. M., & McBride, W. J. (2004).
Intracranial Self-Administration of Ethanol within the Ventral Tegmental Area of Male Wistar
Rats: Evidence for Involvement of Dopamine Neurons. Journal of Neuroscience, 24(5), 1050–
1057. https://doi.org/10.1523/JNEUROSCI.1319-03.2004
Rondou, P., Haegeman, G., & Van Craenenbroeck, K. (2010). The dopamine D4 receptor: biochemical
and signalling properties. Cellular and Molecular Life Sciences, 67(12), 1971–1986.
https://doi.org/10.1007/s00018-010-0293-y
Ronksley, P. E., Brien, S. E., Turner, B. J., Mukamal, K. J., & Ghali, W. A. (2011). Association of alcohol
consumption with selected cardiovascular disease outcomes: a systematic review and meta-
analysis. BMJ, 342, d671. https://doi.org/10.1136/bmj.d671

101
Rosenkranz, J. A., & Grace, A. A. (2002). Dopamine-mediated modulation of odour-evoked amygdala
potentials during pavlovian conditioning. Nature, 417(6886), 282–287.
https://doi.org/10.1038/417282a
Rubinstein, M., Phillips, T. J., Bunzow, J. R., Falzone, T. L., Dziewczapolski, G., Zhang, G., … Grandy, D. K.
(1997). Mice Lacking Dopamine D4 Receptors Are Supersensitive to Ethanol, Cocaine, and
Methamphetamine. Cell, 90(6), 991–1001. https://doi.org/10.1016/S0092-8674(00)80365-7
Sams-Dodd, F. (1998). Effects of dopamine agonists and antagonists on PCP-induced stereotyped
behaviour and social isolation in the rat social interaction test. Psychopharmacology, 135(2),
182–193. https://doi.org/10.1007/s002130050500
Samson, H. H., Tolliver, G. A., & Schwarz-Stevens, K. (1990). Oral ethanol self-administration: A
behavioral pharmacological approach to CNS control mechanisms. Alcohol, 7(3), 187–191.
https://doi.org/10.1016/0741-8329(90)90003-U
Santhakumar, V., Wallner, M., & Otis, T. S. (2007). Ethanol acts directly on extrasynaptic subtypes of
GABAA receptors to increase tonic inhibition. Alcohol (Fayetteville, N.Y.), 41(3), 211–221.
https://doi.org/10.1016/j.alcohol.2007.04.011
Sawaguchi, T., & Goldman-Rakic, P. S. (1991). D1 dopamine receptors in prefrontal cortex: involvement
in working memory. Science, 251(4996), 947–950. https://doi.org/10.1126/science.1825731
Schindler, C. W., Panlilio, L. V., & Goldberg, S. R. (2002). Second-order schedules of drug self-
administration in animals. Psychopharmacology, 163(3–4), 327–344.
https://doi.org/10.1007/s00213-002-1157-4
Schultz, W., Apicella, P., & Ljungberg, T. (1993). Responses of monkey dopamine neurons to reward and
conditioned stimuli during successive steps of learning a delayed response task. Journal of
Neuroscience, 13(3), 900–913.

102
Schultz, W., Dayan, P., & Montague, P. R. (1997). A Neural Substrate of Prediction and Reward. Science,
275(5306), 1593–1599. https://doi.org/10.1126/science.275.5306.1593
Scott, D. J., Domino, E. F., Heitzeg, M. M., Koeppe, R. A., Ni, L., Guthrie, S., & Zubieta, J.-K. (2006).
Smoking Modulation of μ-Opioid and Dopamine D2 Receptor-Mediated Neurotransmission in
Humans. Neuropsychopharmacology, 32(2), 450–457. https://doi.org/10.1038/sj.npp.1301238
Seamans, J. K., & Yang, C. R. (2004). The principal features and mechanisms of dopamine modulation in
the prefrontal cortex. Progress in Neurobiology, 74(1), 1–58.
https://doi.org/10.1016/j.pneurobio.2004.05.006
See, R. E., Fuchs, R. A., Ledford, C. C., & McLaughlin, J. (2003). Drug Addiction, Relapse, and the
Amygdala. Annals of the New York Academy of Sciences, 985(1), 294–307.
https://doi.org/10.1111/j.1749-6632.2003.tb07089.x
See, R. E., Grimm, J. W., Kruzich, P. J., & Rustay, N. (1999). The importance of a compound stimulus in
conditioned drug-seeking behavior following one week of extinction from self-administered
cocaine in rats. Drug and Alcohol Dependence, 57(1), 41–49. https://doi.org/10.1016/S0376-
8716(99)00043-5
See, R. E., Kruzich, P. J., & Grimm, J. W. (2001). Dopamine, but not glutamate, receptor blockade in the
basolateral amygdala attenuates conditioned reward in a rat model of relapse to cocaine-
seeking behavior. Psychopharmacology, 154(3), 301–310.
https://doi.org/10.1007/s002130000636
Seeman, P., Lee, T., Chau-Wong, M., & Wong, K. (1976). Antipsychotic drug doses and
neuroleptic/dopamine receptors. Nature, 261(5562), 717–719.
https://doi.org/10.1038/261717a0
Seeman, P., & Van Tol, H. H. M. (1994). Dopamine receptor pharmacology. Trends in Pharmacological
Sciences, 15(7), 264–270. https://doi.org/10.1016/0165-6147(94)90323-9

103
Self, D. W., & Nestler, E. J. (1998). Relapse to drug-seeking: neural and molecular mechanisms. Drug and
Alcohol Dependence, 51(1–2), 49–60. https://doi.org/10.1016/S0376-8716(98)00065-9
Shaham, Y., Erb, S., & Stewart, J. (2000). Stress-induced relapse to heroin and cocaine seeking in rats: a
review. Brain Research Reviews, 33(1), 13–33. https://doi.org/10.1016/S0165-0173(00)00024-2
Shaham, Y., Rajabi, H., & Stewart, J. (1996). Relapse to heroin-seeking in rats under opioid maintenance:
the effects of stress, heroin priming, and withdrawal. Journal of Neuroscience, 16(5), 1957–1963.
Shaham, Y., Shalev, U., Lu, L., Wit, H. de, & Stewart, J. (2003). The reinstatement model of drug relapse:
history, methodology and major findings. Psychopharmacology, 168(1–2), 3–20.
https://doi.org/10.1007/s00213-002-1224-x
Shekhar, A., Truitt, W., Rainnie, D., & Sajdyk, T. (2005). Role of stress, corticotrophin releasing factor
(CRF) and amygdala plasticity in chronic anxiety. Stress, 8(4), 209–219.
https://doi.org/10.1080/10253890500504557
Sherman, E., Jorenby, D., & Baker, T. (1988). Classical conditioning with alcohol: Acquired preferences
and aversions, tolerance, and urges/craving. Retrieved from
http://psycnet.apa.org/record/1989-97198-006
Sinha, R. (2001). How does stress increase risk of drug abuse and relapse? Psychopharmacology, 158(4),
343–359. https://doi.org/10.1007/s002130100917
Sinha, R., & Li, C.-S. R. (2007). Imaging stress- and cue-induced drug and alcohol craving: association with
relapse and clinical implications. Drug and Alcohol Review, 26(1), 25–31.
https://doi.org/10.1080/09595230601036960
Snyder, S. H., Banerjee, S. P., Yamamura, H. I., & Greenberg, D. (1974). Drugs, Neurotransmitters, and
Schizophrenia. Science, 184(4143), 1243–1253.

104
Sokoloff, P., Giros, B., Martres, M.-P., Bouthenet, M.-L., & Schwartz, J.-C. (1990). Molecular cloning and
characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature,
347(6289), 146–151. https://doi.org/10.1038/347146a0
Stewart, J., de Wit, H., & Eikelboom, R. (1984). Role of unconditioned and conditioned drug effects in the
self-administration of opiates and stimulants. Psychological Review, 91(2), 251–268.
https://doi.org/10.1037/0033-295X.91.2.251
Stewart, J., & Vezina, P. (1988). A comparison of the effects of intra-accumbens injections of
amphetamine and morphine on reinstatement of heroin intravenous self-administration
behavior. Brain Research, 457(2), 287–294. https://doi.org/10.1016/0006-8993(88)90698-1
Sunahara, R. K., Guan, H.-C., O’Dowd, B. F., Seeman, P., Laurier, L. G., Ng, G., … Niznik, H. B. (1991).
Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than
D1. Nature, 350(6319), 614–619. https://doi.org/10.1038/350614a0
Tabakoff, B., & Hoffman, P. L. (2000). Animal models in alcohol research. Alcohol Research and Health,
24(2), 77–84.
Tassin, J.-P., Stinus, L., Simon, H., Blanc, G., Thierry, A.-M., Le Moal, M., … Glowinski, J. (1978).
Relationship between the locomotor hyperactivity induced by A10 lesions and the destruction of
the frontocortical dopaminergic innervation in the rat. Brain Research, 141(2), 267–281.
https://doi.org/10.1016/0006-8993(78)90197-X
Thakker, K. D. (1998). An Overview of Health Risks and Benefits of Alcohol Consumption. Alcoholism:
Clinical and Experimental Research, 22, 285s–298s. https://doi.org/10.1111/j.1530-
0277.1998.tb04381.x
Thanos, P. K., Rivera, S. N., Weaver, K., Grandy, D. K., Rubinstein, M., Umegaki, H., … Volkow, N. D.
(2005). Dopamine D2R DNA transfer in dopamine D2 receptor-deficient mice: Effects on ethanol
drinking. Life Sciences, 77(2), 130–139. https://doi.org/10.1016/j.lfs.2004.10.061

105
Thanos, P. K., Roushdy, K., Sarwar, Z., Rice, O., Ashby, C. R., & Grandy, D. K. (2015). The effect of
dopamine D4 receptor density on novelty seeking, activity, social interaction, and alcohol binge
drinking in adult mice. Synapse, 69(7), 356–364. https://doi.org/10.1002/syn.21822
Thrasher, M. J., Freeman, P. A., & Risinger, F. O. (1999). Clozapine’s Effects on Ethanol’s Motivational
Properties. Alcoholism: Clinical and Experimental Research, 23(8), 1377–1385.
https://doi.org/10.1111/j.1530-0277.1999.tb04360.x
Tiffany, S. T., & Carter, B. L. (1998). Is craving the source of compulsive drug use? Journal of
Psychopharmacology, 12(1), 23–30. https://doi.org/10.1177/026988119801200104
Tran-Nguyen, L. T., Fuchs, R. A., Coffey, G. P., Baker, D. A., O’Dell, L. E., & Neisewander, J. L. (1998).
Time-Dependent Changes in Cocaine-Seeking Behavior and Extracellular Dopamine Levels in the
Amygdala during Cocaine Withdrawal. Neuropsychopharmacology, 19(1), 48–59.
https://doi.org/10.1016/S0893-133X(97)00205-4
Ustün, T. B. (1999). The global burden of mental disorders. American Journal of Public Health, 89(9),
1315–1318.
Van Tol, H., Bunzow, J. R., Guan, H.-C., Sunahara, R. K., Seeman, P., Niznik, H. B., & Civelli, O. (1991).
Cloning of the gene for a human dopamine D4 receptor with high affinity for the antipsychotic
clozapine. Nature, 350(6319), 610–614. https://doi.org/10.1038/350610a0
Van Tol, H. H., Wu, C. M., Guan, H. C., Ohara, K., Bunzow, J. R., Civelli, O., … Jovanovic, V. (1992).
Multiple dopamine D4 receptor variants in the human population. Nature, 358(6382), 149–152.
https://doi.org/10.1038/358149a0
Volkow, N. D., Fowler, J. S., & Wang, G.-J. (2002). Role of dopamine in drug reinforcement and addiction
in humans: results from imaging studies. Behavioural Pharmacology, 13(5–6), 355–366.

106
Volkow, N. D., Fowler, J. S., & Wang, G.-J. (2003). The addicted human brain: insights from imaging
studies. Journal of Clinical Investigation, 111(10), 1444–1451.
https://doi.org/10.1172/JCI200318533
Volkow, N. D., Wang, G.-J., Telang, F., Fowler, J. S., Logan, J., Jayne, M., … Wong, C. (2007). Profound
Decreases in Dopamine Release in Striatum in Detoxified Alcoholics: Possible Orbitofrontal
Involvement. Journal of Neuroscience, 27(46), 12700–12706.
https://doi.org/10.1523/JNEUROSCI.3371-07.2007
Walsh, D. A., Perkins, J. P., & Krebs, E. G. (1968). An Adenosine 3’,5’-Monophosphate-dependant Protein
Kinase from Rabbit Skeletal Muscle. Journal of Biological Chemistry, 243(13), 3763–3765.
Wang, X., Zhong, P., & Yan, Z. (2002). Dopamine D4 receptors modulate GABAergic signaling in
pyramidal neurons of prefrontal cortex. The Journal of Neuroscience: The Official Journal of the
Society for Neuroscience, 22(21), 9185–9193.
Wedemeyer, C., Goutman, J. D., Avale, M. E., Franchini, L. F., Rubinstein, M., & Calvo, D. J. (2007).
Functional activation by central monoamines of human dopamine D4 receptor polymorphic
variants coupled to GIRK channels in Xenopus oocytes. European Journal of Pharmacology,
562(3), 165–173. https://doi.org/10.1016/j.ejphar.2007.01.055
Weiss, F., Ciccocioppo, R., Parsons, L. H., Katner, S., Liu, X., Zorrilla, E. P., … Richter, R. R. (2001).
Compulsive Drug-Seeking Behavior and Relapse. Annals of the New York Academy of Sciences,
937(1), 1–26. https://doi.org/10.1111/j.1749-6632.2001.tb03556.x
Weiss, F., Maldonado-Vlaar, C. S., Parsons, L. H., Kerr, T. M., Smith, D. L., & Ben-Shahar, O. (2000).
Control of cocaine-seeking behavior by drug-associated stimuli in rats: Effects on recovery of
extinguished operant-responding and extracellular dopamine levels in amygdala and nucleus
accumbens. Proceedings of the National Academy of Sciences of the United States of America,
97(8), 4321–4326.

107
Westerink, B. H., Kwint, H. F., & deVries, J. B. (1996). The pharmacology of mesolimbic dopamine
neurons: a dual-probe microdialysis study in the ventral tegmental area and nucleus accumbens
of the rat brain. Journal of Neuroscience, 16(8), 2605–2611.
Whiteford, H. A., Degenhardt, L., Rehm, J., Baxter, A. J., Ferrari, A. J., Erskine, H. E., … others. (2013).
Global burden of disease attributable to mental and substance use disorders: findings from the
Global Burden of Disease Study 2010. The Lancet, 382(9904), 1575–1586.
Williams, G. V., & Goldman-Rakic, P. S. (1995). Modulation of memory fields by dopamine Dl receptors in
prefrontal cortex. Nature, 376(6541), 572–575. https://doi.org/10.1038/376572a0
Willner, P. (1991). Methods for assessing the validity of animal models of human psychopathology.
Animal Models in Psychiatry, I, 1–23.
Winstanley, C. A., Cocker, P. J., & Rogers, R. D. (2011). Dopamine Modulates Reward Expectancy During
Performance of a Slot Machine Task in Rats: Evidence for a ‘Near-miss’ Effect.
Neuropsychopharmacology, 36(5), 913–925. https://doi.org/10.1038/npp.2010.230
Wise, R. A. (1980). Action of drugs of abuse on brain reward systems. Pharmacology Biochemistry and
Behavior, 13, 213–223. https://doi.org/10.1016/S0091-3057(80)80033-5
Wise, R. A. (1982, March). Neuroleptics and operant behavior: The anhedonia hypothesis. Retrieved
February 8, 2017, from /core/journals/behavioral-and-brain-sciences/article/div-
classtitleneuroleptics-and-operant-behavior-the-anhedonia-
hypothesisdiv/EAF66A9C61B9BDD03D35C72E2077620D
Wise, R. A. (2002). Brain Reward Circuitry: Insights from Unsensed Incentives. Neuron, 36(2), 229–240.
https://doi.org/10.1016/S0896-6273(02)00965-0
Wise, R. A., & Bozarth, M. A. (1987). A psychomotor stimulant theory of addiction. Psychological Review,
94(4), 469–492. https://doi.org/10.1037/0033-295X.94.4.469

108
Wise, R. A., & Schwartz, H. V. (1981). Pimozide attenuates acquisition of lever-pressing for food in rats.
Pharmacology Biochemistry and Behavior, 15(4), 655–656. https://doi.org/10.1016/0091-
3057(81)90225-2
World Bank. (1993). World Development Report 1993: Investing in health. Oxford: Oxford Univ. Press.
World Health Organization. (2008). Health statistics and information systems - Metrics: Disability-
Adjusted Life Year (DALY). Retrieved December 29, 2016, from
http://www.who.int/healthinfo/global_burden_disease/metrics_daly/en/
World Health Organization. (2014). Global status report on alcohol and health, 2014. World Health
Organization. Retrieved from
http://apps.who.int/iris/bitstream/10665/112736/1/9789240692763_eng.pdf?ua=1
Wright, G., & Morgan, M. Y. (2013). Alcohol and tobacco misuse: Reducing aerodigestive cancer risk.
World Journal of Hepatology, 5(8), 452–457. https://doi.org/10.4254/wjh.v5.i8.452
Xiang, L., Szebeni, K., Szebeni, A., Klimek, V., Stockmeier, C. A., Karolewicz, B., … Ordway, G. A. (2008).
Dopamine receptor gene expression in human amygdaloid nuclei: Elevated D4 receptor mRNA in
major depression. Brain Research, 1207(Supplement C), 214–224.
https://doi.org/10.1016/j.brainres.2008.02.009
Yan, Y., Pushparaj, A., Le Strat, Y., Gamaleddin, I., Barnes, C., Justinova, Z., … Le Foll, B. (2012). Blockade
of dopamine d4 receptors attenuates reinstatement of extinguished nicotine-seeking behavior
in rats. Neuropsychopharmacology, 37(3), 685–696.
Yokel, R. A., & Wise, R. A. (1975). Increased Lever Pressing for Amphetamine after Pimozide in Rats:
Implications for a Dopamine Theory of Reward. Science, 187(4176), 547–549.
Zeeb, F. D., Robbins, T. W., & Winstanley, C. A. (2009). Serotonergic and Dopaminergic Modulation of
Gambling Behavior as Assessed Using a Novel Rat Gambling Task. Neuropsychopharmacology,
34(10), 2329–2343. https://doi.org/10.1038/npp.2009.62
















![Polymorphisms in the Receptor Tyrosine Kinase MERTK Gene ... · family of structurally related receptor tyrosine kinases that have two identified ligands: GAS6 and protein S [10,11,12]](https://img.pdfslide.net/doc/110x75/5f0d74007e708231d43a6e49/polymorphisms-in-the-receptor-tyrosine-kinase-mertk-gene-family-of-structurally.jpg)












