Embed Size (px)
Citation preview

Cytokines T Cell Sensitivity to IL-2 Family+IRF4 Modulates CD8
Jie SunSu Huang, Yingjia Shen, Duy Pham, Li Jiang, Zheng Wang, Mark H. Kaplan, Guangjun Zhang and
http://www.immunohorizons.org/content/1/6/92https://doi.org/10.4049/immunohorizons.1700020doi:
2017, 1 (6) 92-100ImmunoHorizons
This information is current as of July 16, 2018.
Referenceshttp://www.immunohorizons.org/content/1/6/92.full#ref-list-1
, 8 of which you can access for free at: cites 42 articlesThis article
Email Alertshttp://www.immunohorizons.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
ISSN 2573-7732.All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is an open access journal published byImmunoHorizons
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
by guest on July 16, 2018
http://ww
w.im
munohorizons.org/
Dow
nloaded from

IRF4 Modulates CD8+ T Cell Sensitivity to IL-2 Family Cytokines
Su Huang,*,† Yingjia Shen,‡ Duy Pham,† Li Jiang,* Zheng Wang,† Mark H. Kaplan,† Guangjun Zhang,§ and Jie Sun*,†
*Thoracic Diseases Research Unit, Division of Pulmonary and Critical Care, Department of Medicine, Mayo Clinic College of Medicine, Rochester,
MN 55905; †Herman B Wells Pediatric Research Center, Department of Pediatrics, Indiana University School of Medicine, Indianapolis, IN 46202;‡Key Laboratory of the Ministry of Education for Coastal and Wetland Ecosystems, College of Environment and Ecology, Xiamen University, Xiamen
361005, China; and §Department of Comparative Pathobiology, Center for Cancer Research, Purdue University, West Lafayette, IN 47907
ABSTRACT
IFN regulatory factor 4 (IRF4) is a key transcription factor that promotes effector CD8+ T cell differentiation and expansion. The roles
of IRF4 in regulating the CD8+ T cell response to cytokines have not been explored. In this article, we show that IL-2 and IL-15
signaling and STAT5 activation regulate IRF4 expression in CD8+ T cells. Gene-expression profile analysis has also revealed that IRF4
is required for expression of the receptors of IL-2 family cytokines CD122 and CD127. We found that IRF4 binds directly to CD122 and
CD127 gene loci, indicating that it may directly promote CD122 and CD127 gene transcription. As a consequence, IRF4-deficient CD8+
T cells show diminished sensitivity to IL-2, IL-15, and IL-7 treatment in vitro. Furthermore, we found that IRF4-deficient CD8+ T cells had
lower expression of CD122 and CD127 in vivo during influenza virus infection. These data suggest that IRF4 regulates the sensitivity of
CD8+ T cells to IL-2 family cytokines, which correlates with the diminished effector and memory CD8+ T cell responses in IRF4-deficient
CD8+ T cells. ImmunoHorizons, 2017, 1: 92–100.
INTRODUCTION
As an indispensable part of adaptive immunity, CD8+ T cellsprotect the host from intracellular pathogens and cancerouscells. Three major signals, including TCR signaling, cell surfacecoreceptor signaling, and cytokine receptor signaling, cooperateto direct CD8+ T cell activation, effector differentiation, effectorcell expansion, contraction, and the formation of memory CTLsduring a robust immune response (1). Initially, TCR binding to theAg/MHC complex, with the help of signals from cell surfacecoreceptors, leads to the assembly and activation of a proximalcytoplasm complex of molecules, which subsequently activatesdownstream signaling pathways, such as PI3K and MAPK (2).These signaling pathways further relay activation signals to
various intracellular transcription programs that direct CD8+
T cell activation and proliferation, as well as effector and memoryT cell formation (3). Activation of naive CD8+ T cells also leads tothe expression of various cytokines and cytokine receptors, and thesubsequent downstream cytokine signaling further modifies CD8+
T cell transcriptional programs to assist, guide, and sustain CD8+
T cell responses (4, 5).Among various cytokines that are important in CD8+ T cell–
mediated immunity, IL-2 family cytokines, including IL-2, IL-7,and IL-15, play critical roles in regulating effector and memoryCD8Tcell responses (5–12). IL-2, producedbyactivatedCD4+ andCD8+ T cells, acts through paracrine and autocrinemechanisms topromote CD8 T cell proliferation and effector differentiation(i.e., gaining the ability to express various cytokines and cytotoxic
Received for publication May 31, 2017. Accepted for publication July 25, 2017.
Address correspondence and reprint requests to: Dr. Jie Sun, Mayo Clinic College of Medicine, 200 1st Street SW, Stabile 8-26, Rochester, MN 55905. E-mailaddress: [email protected]
ORCID: 0000-0002-2923-8245 (M.H.K.).
The RNA sequencing data presented in this article have been submitted to the Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/) under accession numberGSE101510.
This work was supported by National Institutes of Health Grants AI112844, AG047156, and AI076458 (to J.S.) and R01 AI095282 and AI057459 (to M.H.K.).
Abbreviations used in this article: BMDC, bone marrow–derived dendritic cell; ChIP, chromatin immunoprecipitation; DC, dendritic cell; ICS, intracellular staining; IRF4,IFN regulatory factor 4; KEGG, Kyoto Encyclopedia of Genes and Genomes; MLN, mediastinal lymph node; RNA-seq, RNA sequencing; STAT5-CA, constitutive activeSTAT5; WT, wild-type.
This article is distributed under the terms of the CC BY 4.0 Unported license.
Copyright © 2017 The Authors
92 https://doi.org/10.4049/immunohorizons.1700020
RESEARCH ARTICLE
Adaptive Immunity
ImmunoHorizons is published by The American Association of Immunologists, Inc.
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
molecules) (4, 13–16). IL-2 signaling also promotes the recallresponses of memory CD8 T cells (15, 17). IL-15 and IL-7 signalingcan promote effector CD8+ T cell proliferation and survival in thetissue, and they appear to be essential for memory CD8+ T cellmaintenance and function (3, 9, 18). IL-2 induces intracellularsignals through the IL-2R complex, consisting of CD25 (IL-2Ra),CD122 (IL-2/IL-15Rb), and CD132 (common g-chain). TheIL-15R complex consists of a unique IL-15Ra subunit alongwith CD122 and CD132 subunits, whereas the IL-7R complexincludes CD127 (IL-7Ra) and CD132. Stimulation of the recep-tor complexes by IL-2, IL-15, and IL-7 induces the activationof STAT5, which mediates the majority of the downstreameffects of these cytokines (5).
A number of transcription factors have been identified to playimportant roles in regulating effector and memory CD8+ T cellresponses. For instance, IFN regulatory factor 4 (IRF4), BATF, T-bet, Blimp-1, and Id2 promote effectorCD8+T cell expansion and/or differentiation, whereas Eomes, Tcf1, BCL6, and Id3 areconsidered to support the generation of memory CD8+ T cells (3).
Numerous loss-of-function and gain-of-function studies havedemonstrated the independent actions of individual transcrip-tion factors, as well as synergy and cooperation among those
transcriptions factors in governing effector and memory CD8+
T cell responses. IRF4, in particular, serves as an integrator tocoordinate the expression and function ofmultiple transcriptionalfactors for effector CD8+ T cell differentiation and expansion(19–24). IRF4 interacts with BATF and is required for theexpression of Hif1a, T-bet, and Blimp1 for sustaining effectorCD8+ T cell differentiation and expansion (19, 21–24). IRF4 alsorepresses Tcf1, Eomes, and BCL6 expression, whichmay promotethe formation of terminal effector CD8T cells overmemory T cellgeneration (19, 22–24).
Given the importance of IL-2, IL-7, and IL-15 in regulatingeffector and memory CD8+ T cell responses, we have investigatedthe potential functional interactions of these cytokines and IRF4.ThroughRNAsequencing (RNA-seq) analysis,we found that IRF4is required for the expression of IL-2 family cytokine receptorsCD122 and CD127. Furthermore, chromatin immunoprecipitation(ChIP) analysis showed that IRF4 directly binds to CD122 andCD127 gene loci, indicating that it may directly promote thetranscription of CD122 and CD127 genes. We found that IRF4-deficient CD8 T cells were less sensitive to IL-2, IL-15, and IL-7treatment during in vitro culture, and IRF4 deficiency in CD8+
T cells resulted in diminished expression of CD122 and CD127 in
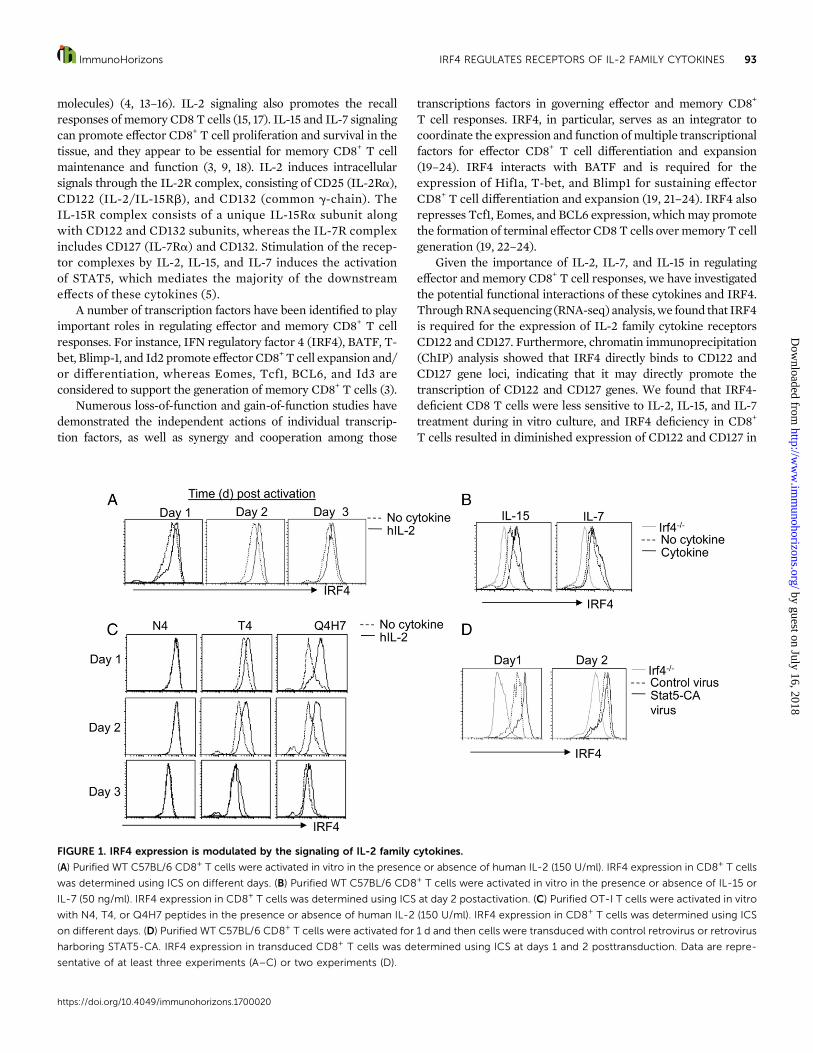
FIGURE 1. IRF4 expression is modulated by the signaling of IL-2 family cytokines.
(A) Purified WT C57BL/6 CD8+ T cells were activated in vitro in the presence or absence of human IL-2 (150 U/ml). IRF4 expression in CD8+ T cells
was determined using ICS on different days. (B) Purified WT C57BL/6 CD8+ T cells were activated in vitro in the presence or absence of IL-15 or
IL-7 (50 ng/ml). IRF4 expression in CD8+ T cells was determined using ICS at day 2 postactivation. (C) Purified OT-I T cells were activated in vitro
with N4, T4, or Q4H7 peptides in the presence or absence of human IL-2 (150 U/ml). IRF4 expression in CD8+ T cells was determined using ICS
on different days. (D) Purified WT C57BL/6 CD8+ T cells were activated for 1 d and then cells were transduced with control retrovirus or retrovirus
harboring STAT5-CA. IRF4 expression in transduced CD8+ T cells was determined using ICS at days 1 and 2 posttransduction. Data are repre-
sentative of at least three experiments (A–C) or two experiments (D).
https://doi.org/10.4049/immunohorizons.1700020
ImmunoHorizons IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES 93
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
influenza-specific CD8+ T cells in vivo. These data suggested thatIRF4 regulates the sensitivity of CD8+ T cells to IL-2 familycytokines.
MATERIALS AND METHODS
Mouse and infectionWild-type (WT)C57BL/6micewerepurchased fromTheJacksonLaboratory. OVA-specific TCR-transgenic (OT-I) mice were bredin-house. Irf4DCD8mice were generated by crossing Irf4 fl/flmicewith mice harboring the E8i-cre transgene, as previously de-scribed (22, 25). Controls for Irf4DCD8micewere littermate Irf4fl/flmice without the E8i-cre transgene. Irf4 fl/fl and Irf4DCD8 micewere on a C57BL/6 background. All mice were housed in a specificpathogen–free environment, and all animal experiments wereperformed in accordance with protocols approved by the IndianaUniversity Institutional Animal Care and Use Committee. In-fluenza A/PR8/34 (;200 PFU permouse) and recombinant PR8-OVA (;400 PFUpermouse) infectionwas performed as describedbefore (26).
Quantitative RT-PCRTotal RNA from cultured cells, MACS beads–enriched in vivoCD8+ T cells, or lung homogenates was extracted with an RNeasyKit (QIAGEN) and treated with DNase I (Invitrogen). Randomprimers and Superscript II (both from Invitrogen) were used tosynthesize first-strand cDNAs from equivalent amounts of RNAof each sample. RT-PCR was performed with SYBR Green PCRMaster Mix (Applied Biosystems). Data were generated with thecomparative threshold cycle method (27) by normalizing tohypoxanthine guanine phosphoribosyl transferase.
T cell cocultureBonemarrow–deriveddendritic cells (BMDCs)were generated aspreviously described (28). CD8+ T cells were isolated from thespleen and lymph nodes of the indicated mice using MACS,according to the manufacturer’s protocol (Miltenyi Biotec). DCswere mixed with CD8+ T cells (5 3 104) at a 1:10 ratio in round-bottom 96-well plates in the presence of 0.1 mg/ml anti-CD3, aspreviously described (26). For activation of OT-I TCR-transgenicT cells, CD8+ T cells from OT-I mice were isolated and culturedwith SIINFEKL (N4), SIITFEKL (T4), or SIIQFEHL (Q4H7)
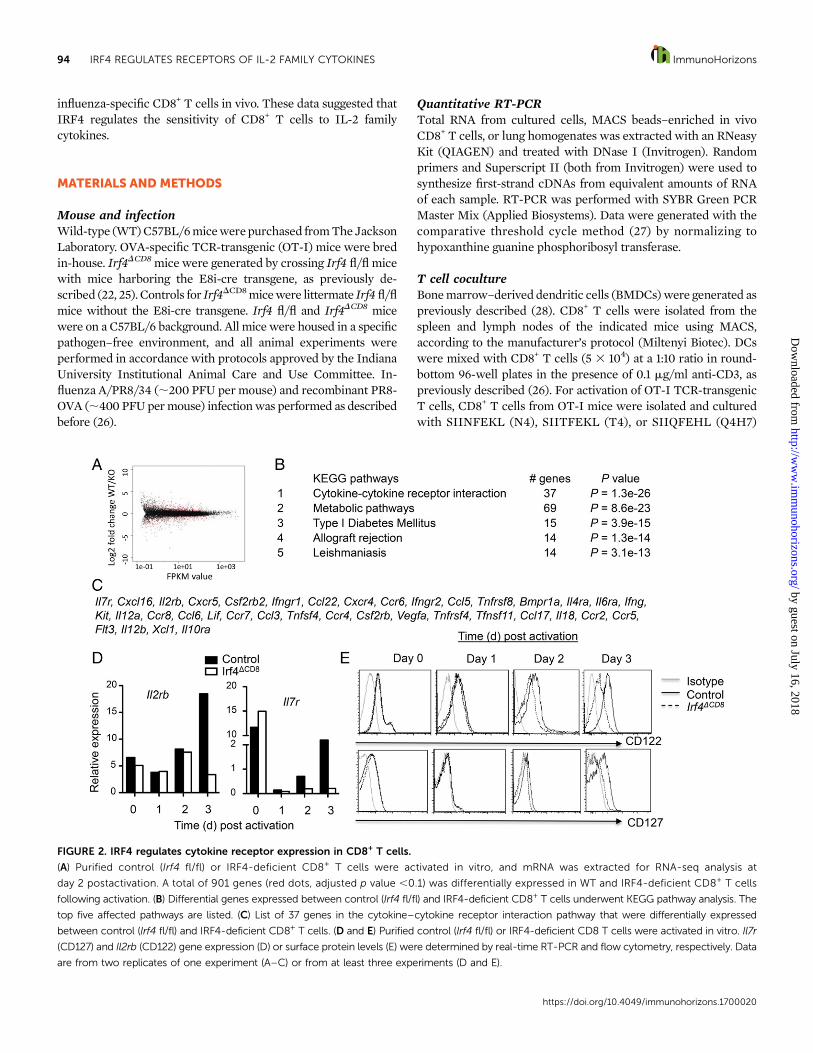
FIGURE 2. IRF4 regulates cytokine receptor expression in CD8+ T cells.
(A) Purified control (Irf4 fl/fl) or IRF4-deficient CD8+ T cells were activated in vitro, and mRNA was extracted for RNA-seq analysis at
day 2 postactivation. A total of 901 genes (red dots, adjusted p value ,0.1) was differentially expressed in WT and IRF4-deficient CD8+ T cells
following activation. (B) Differential genes expressed between control (Irf4 fl/fl) and IRF4-deficient CD8+ T cells underwent KEGG pathway analysis. The
top five affected pathways are listed. (C) List of 37 genes in the cytokine–cytokine receptor interaction pathway that were differentially expressed
between control (Irf4 fl/fl) and IRF4-deficient CD8+ T cells. (D and E) Purified control (Irf4 fl/fl) or IRF4-deficient CD8 T cells were activated in vitro. Il7r
(CD127) and Il2rb (CD122) gene expression (D) or surface protein levels (E) were determined by real-time RT-PCR and flow cytometry, respectively. Data
are from two replicates of one experiment (A–C) or from at least three experiments (D and E).
https://doi.org/10.4049/immunohorizons.1700020
94 IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES ImmunoHorizons
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
peptides (4 ng/ml). In some experiments, the indicated concen-trations of human IL-2, IL-7, and IL-15 were included in theculture.
RNA-seqTotal RNA from culturedWT or IRF4-deficient CD8+ T cells wasextracted with an RNeasy Kit (QIAGEN). Sequencing librarieswere constructed from the total RNA using the Illumina TruSeqStranded mRNA Library Prep Kit, largely as directed by the kit’sdirections butwith somemodifications. Fragmentation time in theelute–fragment–prime stepwas reduced from8 to 4min, resultingin cDNA ranging in length from 100 to 1000 bp. SubsequentAMPure purification was performed at a sample/AMPure ratio of1.8:1 (v/v). The resulting libraries were composed of ampliconslargely ranging in length from 200 to 1000 bp. Subsequently, 8cycles of PCR amplification were undertaken, rather than the 15cycles specified by the instructions. Individual libraries werepooled based on titers using KAPA Library Quantification on anApplied Biosystems Step-One qPCR machine. A single pool of allsix librarieswas clustered onone lane of aHiSEquation 2500HighOutput 23 100–bases read chemistry run.
For bioinformatics analysis, we first processed reads using acustom Perl script to remove unreliable reads and to trim se-quences with low-quality sections of each read based on the fol-lowing criteria: any reads with uncalled bases and Phred qualityscore of 2 were rejected, and reads were trimmed when the qual-ity score was ,10 or the average score of three continuous bases
was ,20. Then processed reads were mapped against the refer-ence mouse genome (GRCm38.p5) using Tophat2 (29). To quan-tify the read count per transcript, HTSeq (30) was used, withgenome annotation downloaded from Ensembl (release 88). Differ-ential expression analyses were performed using the DESeq2package (31, 32). Genes with false discovery rate, 10% (adjustedp value,0.1) and.2-fold change were considered significant. Allsignificant genes (.2-fold change and false discovery rate, 10%)were analyzed by WebGestalt (32) for Gene Ontology and KyotoEncyclopedia of Genes and Genomes (KEGG) pathway enrich-ment using a hypergeometric test, with a significance level of0.01. RNA-seq data have been submitted to the Gene Expres-sion Omnibus under accession number GSE101510 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE101510).
ChIPChIP assays were performed as described (33). In brief, activatedWT or IRF4-deficient CD8+ T cells were cross-linked for 10 minwith 1% formaldehyde and fragmentized by sonication. Cross-linked cells were precleared with salmon sperm DNA, BSA, andprotein G agarose (IRF4 ChIP) bead slurry (50%). Cell extractswere incubatedwithAbs to goat polyclonal IRF4M-17 (SantaCruzBiotechnology), H3K27ac (Millipore), and normal goat IgG (SantaCruz Biotechnology) overnight at 4°C. The immunocomplexeswere precipitated with Protein A agarose (H3K27ac ChIP) orProtein G agarose (IRF4 ChIP) beads at 4°C for 2 h, washed, andeluted, and cross-links were reversed at 65°C overnight. DNAwas
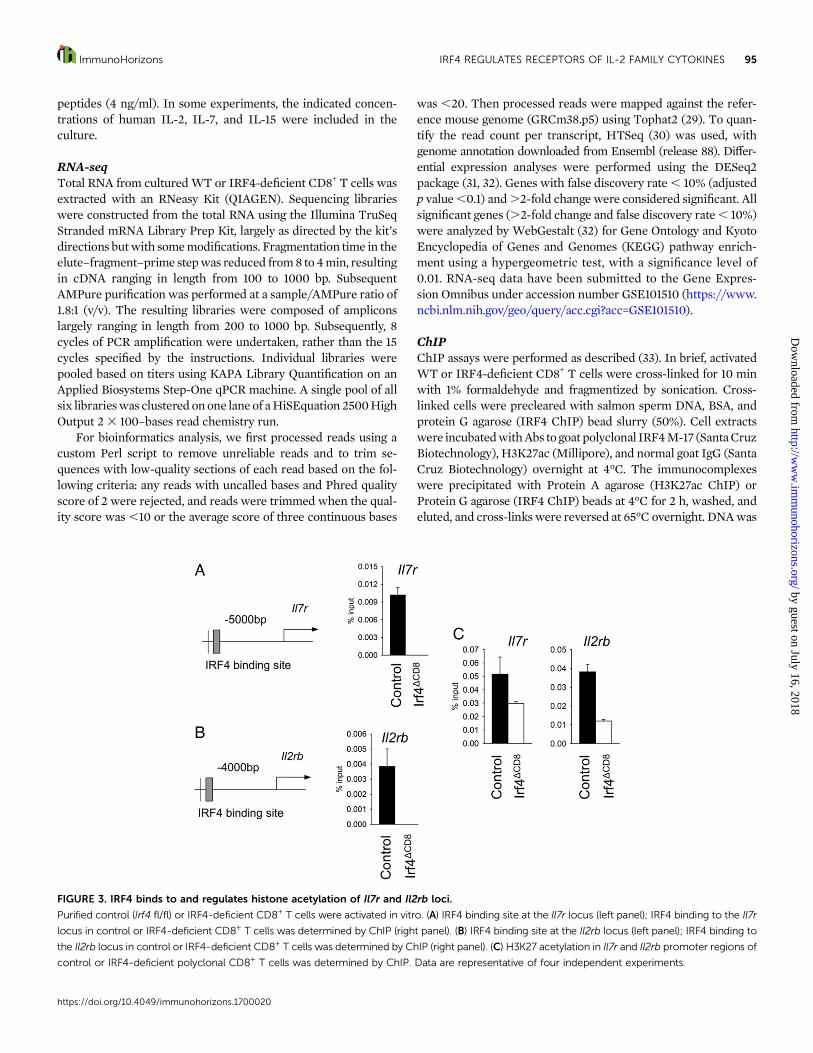
FIGURE 3. IRF4 binds to and regulates histone acetylation of Il7r and Il2rb loci.
Purified control (Irf4 fl/fl) or IRF4-deficient CD8+ T cells were activated in vitro. (A) IRF4 binding site at the Il7r locus (left panel); IRF4 binding to the Il7r
locus in control or IRF4-deficient CD8+ T cells was determined by ChIP (right panel). (B) IRF4 binding site at the Il2rb locus (left panel); IRF4 binding to
the Il2rb locus in control or IRF4-deficient CD8+ T cells was determined by ChIP (right panel). (C) H3K27 acetylation in Il7r and Il2rb promoter regions of
control or IRF4-deficient polyclonal CD8+ T cells was determined by ChIP. Data are representative of four independent experiments.
https://doi.org/10.4049/immunohorizons.1700020
ImmunoHorizons IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES 95
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
purified, suspended in H2O, and analyzed by quantitative PCR, aspreviously described (34). Primers for IRF4 binding to IL-2rb andIL-7ra lociwere designedagainst theirpromoter region, accordingto current Ensembl annotation. The IRF4 binding sites for Il2rband Il7r were determined with the TRANSFAC Transcriptionfactor binding site database. The following primers were used forquantitative PCR: Il7r forward 59-GCCCCAGCATTTTGAAGTT-39, Il7r reverse 59-GTGTCTTCTTCTTTCTGC-39, Il2rb forward59-CCTGCTTCCCTTCTGTTCTG-39, and Il2rb reverse 59-TCT-TCGAGGCCAACATGTCT-39.
Retroviral transductionConstitutive active STAT5 (STAT5-CA) and control retroviralplasmids were kindly provided by Dr. J. Zhu (National In-stitutes of Health) (35). Retroviral stocks were prepared bycalcium phosphate transfection of Phoenix GP cells. Themedium was refreshed 12 h after transfection, and viralsupernatants were collected 24 and 48 h later. For retroviraltransduction, CD8+ T cells were stimulated with BMDCs plusanti-CD3. At day 1 of the culture, cells were transduced withretroviruses through spin infection (2500 rpm, 90 min), asprevious described (36). After transduction, cells were ana-lyzed by flow cytometry.
Flow cytometry analysisFluorescence-conjugated FACS Abs were purchased from BioL-egend, BD Biosciences, or eBioscience. Intracellular staining(ICS) of IRF4 was performed using a Foxp3 Staining Buffer Set(eBioscience). For IRF4staining, IRF4-deficientCD8+Tcellswereused as the negative staining control. Cells were acquired usinga FACSCalibur (BD Biosciences) or Attune Acoustic FocusingCytometer (Invitrogen), and data were analyzed with FlowJosoftware (TreeStar).
Statistical analysisData are mean 6 SEM of values from individual mice (in vivoexperiments) or mean 6 SD of values from triplicate analysis ofthe same sample (in vitro cell numbers and ChIP analysis). Apaired or unpaired two-tailed Student t test was used. We con-sider p values,0.05 significant.
RESULTS
IL-2 and IL-15 modulate IRF4 expression in CD8+ T cellsPrevious reports have established that IRF4 is vital for thegeneration of effector and memory CD8+ T cells (19–24). Theexpression of IRF4 is primarily regulated by TCR signaling,although type I IFNs were recently shown to promote IRF4expression in CD8+ T cells (37), indicating that IRF4 expressioncould be potentially modulated by cytokines. Because the IL-2family cytokines IL-2, IL-15, and IL-7 are potent modulators ofeffector andmemory cell responses (1, 5),we investigatedwhetherIL-2, IL-15, and/or IL-7 regulate IRF4 expression in CD8+ T cells.To this end, we first stimulated WT CD8+ T cells in vitro with
BMDCs and anti-CD3 in the absence or presence of human IL-2.We found that human IL-2 inclusion enhanced IRF4 expressionin CD8+ T cells (Fig. 1A). Like IL-2, IL-15 could boost IRF4expression, whereas IL-7 only modestly increased IRF4 expres-sion inCD8T cells (Fig. 1B). These data suggested that IL-2 familycytokines differentially regulate IRF4 expression in CD8+ T cells.
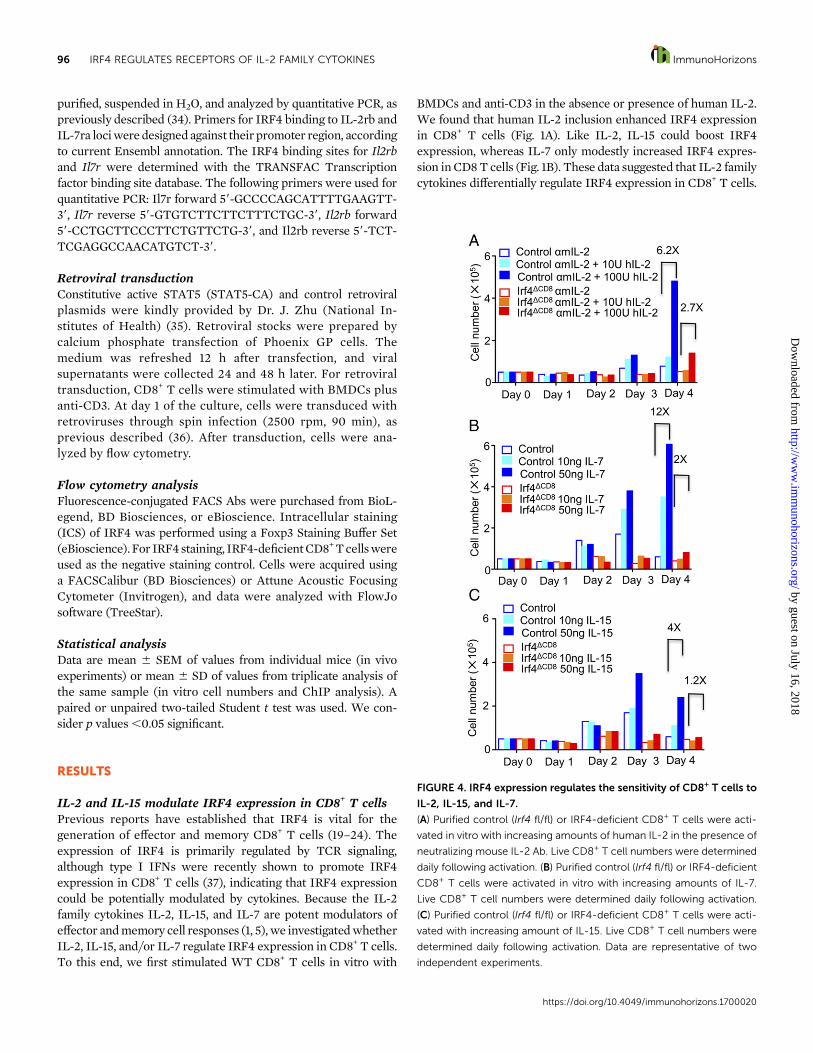
FIGURE 4. IRF4 expression regulates the sensitivity of CD8+ T cells to
IL-2, IL-15, and IL-7.
(A) Purified control (Irf4 fl/fl) or IRF4-deficient CD8+ T cells were acti-
vated in vitro with increasing amounts of human IL-2 in the presence of
neutralizing mouse IL-2 Ab. Live CD8+ T cell numbers were determined
daily following activation. (B) Purified control (Irf4 fl/fl) or IRF4-deficient
CD8+ T cells were activated in vitro with increasing amounts of IL-7.
Live CD8+ T cell numbers were determined daily following activation.
(C) Purified control (Irf4 fl/fl) or IRF4-deficient CD8+ T cells were acti-
vated with increasing amount of IL-15. Live CD8+ T cell numbers were
determined daily following activation. Data are representative of two
independent experiments.
https://doi.org/10.4049/immunohorizons.1700020
96 IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES ImmunoHorizons
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from
Because IRF4 expression in CD8+ T cells is influenced by thestrength of TCR stimulation (22, 23, 38), we next examinedwhether IL-2 supplementation could differentially regulate IRF4expression under strong and weak TCR stimulation. To this end,we activated OT-I T cells with BMDCs plus OVA (OVA257–264)SINNFEKLpeptide (N4) or its low-affinity alteredpeptide ligandsSINTFEKL (T4) and SINQFEHL (Q4H7) in the presence orabsence of human IL-2. As previously reported, high-affinity TCR
stimulation induced stronger and sustained IRF4 expression (N4stimulation), whereas weak TCR stimulation induced lower andtransient IRF4 expression (T4 or Q4H7 stimulation) (22, 23). IL-2enhanced IRF4 expression in CD8+ T cells stimulated with T4 orQ4H7, but not N4, suggesting that IL-2 preferentially boostsIRF4 expression in CD8+ T cells stimulated with weak TCRsignals (Fig. 1C). STAT5 is an important downstreamtranscription factor that mediates the effects of IL-2 and
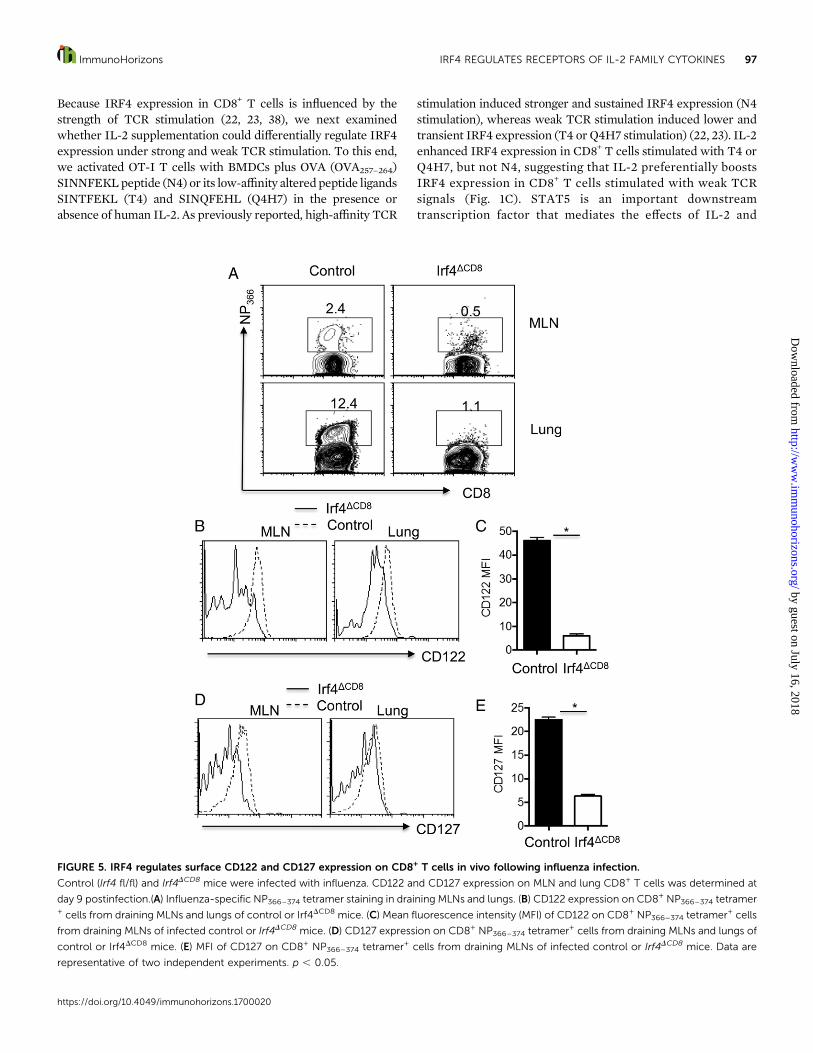
FIGURE 5. IRF4 regulates surface CD122 and CD127 expression on CD8+ T cells in vivo following influenza infection.
Control (Irf4 fl/fl) and Irf4DCD8 mice were infected with influenza. CD122 and CD127 expression on MLN and lung CD8+ T cells was determined at
day 9 postinfection.(A) Influenza-specific NP366–374 tetramer staining in draining MLNs and lungs. (B) CD122 expression on CD8+ NP366–374 tetramer+ cells from draining MLNs and lungs of control or Irf4DCD8 mice. (C) Mean fluorescence intensity (MFI) of CD122 on CD8+ NP366–374 tetramer+ cells
from draining MLNs of infected control or Irf4DCD8 mice. (D) CD127 expression on CD8+ NP366–374 tetramer+ cells from draining MLNs and lungs of
control or Irf4DCD8 mice. (E) MFI of CD127 on CD8+ NP366–374 tetramer+ cells from draining MLNs of infected control or Irf4DCD8 mice. Data are
representative of two independent experiments. p , 0.05.
https://doi.org/10.4049/immunohorizons.1700020
ImmunoHorizons IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES 97
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from

IL-15 in T cells (5, 39). Therefore, we examined whetherSTAT5 activation promotes IRF4 expression. To this end, wetransduced activated CD8+ T cells with control retrovirus orretrovirus containing STAT5-CA (35). We found that consti-tutive STAT5 activation promoted IRF4 expression in CD8+
T cells (Fig. 1D). Althoughwe could not rule out the possibilitythat STAT5-independent pathways, such as the PI3K/AKTsignaling pathway, may regulate IRF4 expression in CD8+
T cells (40), these data suggested that STAT5 activation down-stream of IL-2 and IL-15 signaling contributes to the upregulationof IRF4 expression in CD8+ T cells.
Impaired cytokine receptor expression in IRF4-deficientCD8+ T cellsTo better understand the underlying molecular mechanismsby which IRF4 regulates CD8+ T cell responses, we performednext-generation RNA-seq analysis on activated control andIRF4-deficient CD8+ T cells. We found that .900 genes weredifferentially regulated in control and IRF4-knockout T cells (Fig.2A). We next performed KEGG pathway analysis of the genesthat were differentially expressed between control and IRF4-deficient CD8+ T cells. As shown in Fig. 2B, the cytokine–cytokinereceptor interaction pathway is the pathway that is most signif-icantly differentially regulated in control and IRF4-deficientCD8+
Tcells: 37 genes of thispathwayweredifferentially expressed (Fig.2C). As reported previously (23), we found that IRF4-deficientCD8+ T cells had diminished expression of many genes relatedto cellular metabolism (Fig. 2B). Among the cytokine receptorsthat are differentially expressed between control and IRF4-deficient CD8+ T cells, we found that Il7r (CD127) and Il2rb(CD122, shared by IL-2 and IL-15 signaling) were downregulatedin IRF4-deficient CD8+ T cells (Fig. 2C). We confirmed the RNA-seq data using real-time RT-PCR (Fig. 2D). Flow cytometryanalysis of the surface expression of CD127 and CD122 furtherconfirmed the diminished expression of CD127 and CD122 onIRF4-deficient CD8+ T cells (Fig. 2E). Given that IL-2 and IL-15could potently regulate IRF4 expression, these data suggestedthat a cytokine feed-forward loop exists on IRF4 regulation topotentially promote CD8+ T cell activation, expansion, and ef-fector differentiation.
IRF4 binds and regulates epigenetic modifications ofcytokine receptor gene lociIRF4 can work as a transcriptional activator to induce manydownstream genes, including cell-cycle related genes, metabolicgenes, andeffectormolecules (20–24, 38).Therefore,weexaminedwhether IRF4bindsdirectly toCD127 (Il7r) andCD122 (Il2rb) geneloci to potentially regulate their expression.We found that Il7r andIl2rb loci contain IRF4 binding sites using TRANSFAC (Fig. 3A,3B). ChIP assay confirmed IRF4 binding to Il7r and Il2rb loci inWT CD8+ T cells but not in IRF4-deficient CD8+ T cells (Fig. 3A,3B). Epigenetic chromatinmodifications, such as the acetylation ofH3K27, usually mark open chromatin that is accessible for genetranscription. Therefore, we examined H3K27 acetylation in theIl7r and Il2rb promoter regions in activated control and IRF4-
deficient CD8+ T cells. We found that IRF4 deficiency resulted inreduced H3K27 acetylation in the Il7r and Il2rb gene loci.Together, these data demonstrated that IRF4 directly binds andpromotes epigenetic modifications at CD122 and CD127 gene loci.
IRF4 regulates CD8 T cell expansion in responseto cytokinesBecause IRF4 regulates CD127 and CD122 expression on CD8+
T cells, we next examined whether IRF4 potentiates CD8+ T cellexpansion in response to cytokine signaling. To this end, weexamined whether IRF4 is required for cytokine-induced CD8+
T cell proliferation in vitro. We activated CD8+ T cells in theabsence or presence of increasing concentrations of IL-2, IL-7, orIL-15. As expected, addition of IL-2, IL-7, or IL-15 increasedcontrol CD8+ T cell expansion (Fig. 4). However, IRF4 deficiencyimpaired CD8+ T cell expansion in response to IL-2, IL-7, or IL-15(Fig. 4), suggesting that IRF4 regulates the response of CD8+
T cells to IL-2 family cytokines.
IRF4 is required for the surface expression of CD127 andCD122 on CD8+ T cells in vivoTo examine whether IRF4 is required for cytokine receptorexpression on CD8+ T cells in vivo, we infected WT or Irf4DCD8
micewith influenzaA/PR/8 and then examinedCD127 andCD122expression on Ag-specific NP366–374 tetramer+ CD8+ cells at day 9postinfection. As previously reported (22), Irf4DCD8micemountedgreatly diminished influenza-specific T cell responses in vivo (Fig.5A). In accordance with the diminished CD122 observed in vitro,CD122 expression on influenza-specific CD8+ T cells fromdraining mediastinal lymph nodes (MLNs) and the lungs werediminished in the absence of IRF4 (Fig. 5B, 5C). Similarly, weobserveddiminishedCD127expressionon influenza-specificCD8+
T cells in vivo (Figs. 5D, 5E). These data suggest that IRF4 isrequired for CD122 and CD127 expression on Ag-specific CD8+
T cells in vivo (Fig. 6).
FIGURE 6. Integrated model of IRF4 and IL-2 family cytokine
signaling in regulating effector and memory CD8+ T cell responses.
https://doi.org/10.4049/immunohorizons.1700020
98 IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES ImmunoHorizons
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from

DISCUSSION
Taken together, we have identified that IL-2 family cytokines IL-2and IL-15 could modulate IRF4 expression via STAT5 signalingand, conversely, that IRF4 is vital for expression of the receptors ofIL-2 family cytokines, including CD122 and CD127. Thus, thesedata suggest that IRF4 may be a key factor regulating a cytokinefeed-forward loop to promote CD8+ T cell responses. Further-more, because IRF4 expression is also controlled by the strengthof TCR stimulation, IRF4 may link graded TCR signals to CD8+
T cell sensitivity to cytokines (Fig. 6). In this model, productiveTCR stimulation promotes IRF4 expression, which leads to thesynthesis of key cytokine receptors and sensitizes CD8+ T cell re-sponses to cytokines. Subsequently, cytokines further potentiateand sustain IRF4 expression in activated CD8+ T cells to promoteeffector CTL responses. Understanding the exact mechanisms bywhich IRF4 modulates the interplay of TCR and cytokine signal-ing to promote effective CTL responses could provide new in-sights into the molecular regulation of CD8+ T cell responsesagainstpathogens andcancerous cells; inparticular, IL-2 andIL-15have been used as promising antitumor agents (41, 42).
DISCLOSURES
The authors have no financial conflicts of interest.
ACKNOWLEDGMENTS
We thank Dr. Jinfang Zhu for the STAT5-CA plasmids and the NationalInstitutes of Health Tetramer Core Facility for tetramers to detectinfluenza-specific CD8+ T cells.
REFERENCES
1. Bevan, M. J. 2011. Memory T cells as an occupying force. Eur. J.Immunol. 41: 1192–1195.
2. Chakraborty, A. K., and A. Weiss. 2014. Insights into the initiation ofTCR signaling. Nat. Immunol. 15: 798–807.
3. Kaech, S. M., and W. Cui. 2012. Transcriptional control of effector andmemory CD8+ T cell differentiation. Nat. Rev. Immunol. 12: 749–761.
4. Zhang, N., and M. J. Bevan. 2011. CD8(+) T cells: foot soldiers of theimmune system. Immunity 35: 161–168.
5. Liao, W., J. X. Lin, and W. J. Leonard. 2013. Interleukin-2 at thecrossroads of effector responses, tolerance, and immunotherapy. Im-munity 38: 13–25.
6. Boyman, O., and J. Sprent. 2012. The role of interleukin-2 duringhomeostasis and activation of the immune system. Nat. Rev. Immunol.12: 180–190.
7. Cox, M. A., L. E. Harrington, and A. J. Zajac. 2011. Cytokines and theinception of CD8 T cell responses. Trends Immunol. 32: 180–186.
8. Kim, M. T., and J. T. Harty. 2014. Impact of inflammatory cytokineson effector and memory CD8+ T cells. Front. Immunol. 5: 295.
9. Verbist, K. C., and K. D. Klonowski. 2012. Functions of IL-15 in anti-viral immunity: multiplicity and variety. Cytokine 59: 467–478.
10. Steel, J. C., T. A. Waldmann, and J. C. Morris. 2012. Interleukin-15biology and its therapeutic implications in cancer. Trends Pharmacol.Sci. 33: 35–41.
11. Osborne, L. C., and N. Abraham. 2010. Regulation of memory T cellsby gc cytokines. Cytokine 50: 105–113.
12. Boyman, O., J. F. Purton, C. D. Surh, and J. Sprent. 2007. Cytokinesand T-cell homeostasis. Curr. Opin. Immunol. 19: 320–326.
13. Pipkin, M. E., J. A. Sacks, F. Cruz-Guilloty, M. G. Lichtenheld, M. J.Bevan, and A. Rao. 2010. Interleukin-2 and inflammation inducedistinct transcriptional programs that promote the differentiation ofeffector cytolytic T cells. Immunity 32: 79–90.
14. Kamimura, D., and M. J. Bevan. 2007. Naive CD8+ T cells differen-tiate into protective memory-like cells after IL-2 anti IL-2 complextreatment in vivo. J. Exp. Med. 204: 1803–1812.
15. Feau, S., R. Arens, S. Togher, and S. P. Schoenberger. 2011. AutocrineIL-2 is required for secondary population expansion of CD8(+)memory T cells. Nat. Immunol. 12: 908–913.
16. Kalia, V., S. Sarkar, S. Subramaniam, W. N. Haining, K. A. Smith, andR. Ahmed. 2010. Prolonged interleukin-2Ralpha expression on virus-specific CD8+ T cells favors terminal-effector differentiation in vivo.Immunity 32: 91–103.
17. Williams, M. A., A. J. Tyznik, and M. J. Bevan. 2006. Interleukin-2signals during priming are required for secondary expansion of CD8+memory T cells. Nature 441: 890–893.
18. McGill, J., N. Van Rooijen, and K. L. Legge. 2010. IL-15 trans-presentation by pulmonary dendritic cells promotes effector CD8T cell survival during influenza virus infection. J. Exp. Med. 207:521–534.
19. Nayar, R., E. Schutten, B. Bautista, K. Daniels, A. L. Prince, M. Enos,M. A. Brehm, S. L. Swain, R. M. Welsh, and L. J. Berg. 2014. Gradedlevels of IRF4 regulate CD8+ T cell differentiation and expansion, butnot attrition, in response to acute virus infection. J. Immunol. 192:5881–5893.
20. Huber, M., and M. Lohoff. 2014. IRF4 at the crossroads of effectorT-cell fate decision. Eur. J. Immunol. 44: 1886–1895.
21. Grusdat, M., D. R. McIlwain, H. C. Xu, V. I. Pozdeev, J. Knievel, S. Q.Crome, C. Robert-Tissot, R. J. Dress, A. A. Pandyra, D. E. Speiser, et al.2014. IRF4 and BATF are critical for CD8+ T-cell function followinginfection with LCMV. Cell Death Differ. 21: 1050–1060.
22. Yao, S., B. F. Buzo, D. Pham, L. Jiang, E. J. Taparowsky, M. H. Kaplan,and J. Sun. 2013. Interferon regulatory factor 4 sustains CD8(+) T cellexpansion and effector differentiation. Immunity 39: 833–845.
23. Man, K., M. Miasari, W. Shi, A. Xin, D. C. Henstridge, S. Preston,M. Pellegrini, G. T. Belz, G. K. Smyth, M. A. Febbraio, et al. 2013. Thetranscription factor IRF4 is essential for TCR affinity-mediated met-abolic programming and clonal expansion of T cells. Nat. Immunol.14: 1155–1165.
24. Raczkowski, F., J. Ritter, K. Heesch, V. Schumacher, A. Guralnik,L. Hocker, H. Raifer, M. Klein, T. Bopp, H. Harb, et al. 2013. Thetranscription factor interferon regulatory factor 4 is required for thegeneration of protective effector CD8+ T cells. Proc. Natl. Acad. Sci.USA 110: 15019–15024.
25. Maekawa, Y., Y. Minato, C. Ishifune, T. Kurihara, A. Kitamura,H. Kojima, H. Yagita, M. Sakata-Yanagimoto, T. Saito, I. Taniuchi, et al.2008. Notch2 integrates signaling by the transcription factors RBP-Jand CREB1 to promote T cell cytotoxicity. Nat. Immunol. 9: 1140–1147.
26. Sun, J., R. Madan, C. L. Karp, and T. J. Braciale. 2009. Effector T cellscontrol lung inflammation during acute influenza virus infection byproducing IL-10. Nat. Med. 15: 277–284.
27. Schmittgen, T. D., and K. J. Livak. 2008. Analyzing real-time PCRdata by the comparative C(T) method. Nat. Protoc. 3: 1101–1108.
28. Sun, J., and E. J. Pearce. 2007. Suppression of early IL-4 productionunderlies the failure of CD4 T cells activated by TLR-stimulateddendritic cells to differentiate into Th2 cells. J. Immunol. 178:1635–1644.
29. Kim, D., G. Pertea, C. Trapnell, H. Pimentel, R. Kelley, and S. L. Salzberg.2013. TopHat2: accurate alignment of transcriptomes in the presence ofinsertions, deletions and gene fusions. Genome Biol. 14: R36.
https://doi.org/10.4049/immunohorizons.1700020
ImmunoHorizons IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES 99
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from

30. Anders, S., P. T. Pyl, and W. Huber. 2015. HTSeq–a Python frameworkto work with high-throughput sequencing data. Bioinformatics 31:166–169.
31. Love, M. I., W. Huber, and S. Anders. 2014. Moderated estimation offold change and dispersion for RNA-seq data with DESeq2. GenomeBiol. 15: 550.
32. Wang, J., D. Duncan, Z. Shi, and B. Zhang. 2013. WEB-based GEneSeT anaLysis toolkit (WebGestalt): update 2013. Nucleic Acids Res. 41:W77–W83.
33. Stritesky, G. L., R. Muthukrishnan, S. Sehra, R. Goswami, D. Pham,J. Travers, E. T. Nguyen, D. E. Levy, and M. H. Kaplan. 2011. Thetranscription factor STAT3 is required for T helper 2 cell develop-ment. Immunity 34: 39–49.
34. Pham, D., J. W. Vincentz, A. B. Firulli, and M. H. Kaplan. 2012. Twist1regulates Ifng expression in Th1 cells by interfering with Runx3function. J. Immunol. 189: 832–840.
35. Zhu, J., J. Cote-Sierra, L. Guo, and W. E. Paul. 2003. Stat5 activationplays a critical role in Th2 differentiation. Immunity 19: 739–748.
36. Sun, J., C. J. Krawczyk, and E. J. Pearce. 2008. Suppression of Th2cell development by Notch ligands Delta1 and Delta4. J. Immunol.180: 1655–1661.
37. Jiang, L., S. Yao, S. Huang, J. Wright, T. J. Braciale, and J. Sun. 2016.Type I IFN signaling facilitates the development of IL-10-producingeffector CD8(+) T cells during murine influenza virus infection. Eur. J.Immunol. 46: 2778–2788.
38. Nayar, R., M. Enos, A. Prince, H. Shin, S. Hemmers, J. K. Jiang,U. Klein, C. J. Thomas, and L. J. Berg. 2012. TCR signaling via Teckinase ITK and interferon regulatory factor 4 (IRF4) regulates CD8+T-cell differentiation. Proc. Natl. Acad. Sci. USA 109: E2794–E2802.
39. Jiang, Q., W. Q. Li, F. B. Aiello, R. Mazzucchelli, B. Asefa, A. R.Khaled, and S. K. Durum. 2005. Cell biology of IL-7, a key lympho-trophin. Cytokine Growth Factor Rev. 16: 513–533.
40. Waldmann, T. A. 2006. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Nat. Rev.Immunol. 6: 595–601.
41. Floros, T., and A. A. Tarhini. 2015. Anticancer cytokines: biology andclinical effects of interferon-a2, interleukin (IL)-2, IL-15, IL-21, andIL-12. Semin. Oncol. 42: 539–548.
42. Pulliam, S. R., R. V. Uzhachenko, S. E. Adunyah, and A. Shanker. 2016.Common gamma chain cytokines in combinatorial immune strategiesagainst cancer. Immunol. Lett. 169: 61–72.
https://doi.org/10.4049/immunohorizons.1700020
100 IRF4 REGULATES RECEPTORS OF IL-2 FAMILY CYTOKINES ImmunoHorizons
by guest on July 16, 2018http://w
ww
.imm
unohorizons.org/D
ownloaded from





























