Embed Size (px)
Citation preview

Iron regulated outer membrane proteins (IROMPs) as potential targets against carbapenem-resistant Acinetobacter spp. isolated from a Medical Centre in Malaysia
Wong Eng Hwa, Geetha Subramaniam, Marzida Bt. Mansor*, Ong Siok Yan, Gracie*, Deepa Anbazhagan & Sekaran Shamala Devi
Departments of Medical Microbiology & *Anaesthesiology, Faculty of Medicine, University Malaya Kuala Lumpur, Malaysia
Received December 8, 2008
Background & objectives: Carbapenem-resistant Acinetobacter spp. have gained increasing significance as opportunistic pathogens in hospitalized patients. Carbapenem resistance is often associated with the loss and/or decrease in outer membrane proteins (OMP) and overexpression of multidrug efflux systems. However, carbapenem-hydrolysing β-lactamases of Ambler Class B (metallo-enzymes) and Ambler Class D (oxacillinases) have also been detected in Acinetobacter spp. In this study we have investigated the role of the iron regulated outer membrane protein (IROMPs) and the loss of a 29-kDa OMP in carbapenem resistance of Acinetobacter calcoaceticus. Methods: Carbapenem resistant clinical isolates (n=39) of Acinetobacter baumannii / calcoaceticus were used. Identification of Acinetobacter spp. at species level was done by amplified ribosomal DNA restriction analysis (ARDRA). MIC was evaluated using agar dilution method according to CLSI standards. Presence of outer membrane proteins were determined by SDS-PAGE. A representative strain of A. calcoaceticus, S26 with the loss of 29-kDa OMP was selected for further analysis as strain S26 had unique resistance mechanism, that is, the presence of IMP-4 metallo-β-lactamases. IROMPs were expressed under iron deficit conditions. Bands corresponding to IROMPs were excised from SDS-PAGE and used to immunize rabbits for the production of polyclonal antibodies. The antibodies raised against IROMPs were detected by an in-house ELISA and then used for bactericidal activity against carbapenem resistant A. baumannii / calcoaceticus. Results: All isolates were resistant to all antibiotics including imipenem and meropenem and had loss of a 29-kDa OMP. The polyclonal antibodies showed bactericidal effect against the organism tested and it specifically killed the bacteria grown in iron deficit medium. Interpretation & conclusions: In this study, a 29-kDa OMP has been identified to be the major outer membrane protein in A. baumannii / calcoaceticus and loss of this porin and overexpression of IROMPs have contributed to carbapenem resistance. Polyclonal antibodies raised against IROMPs may have a role in antimicrobial therapy in these isolates.
Key words Acinetobacter spp. - imipenem - iron regulated outer membrane protein (IROMPs) - meropenem - multiresistant - outer membrane protein (OMP) - SDS-PAGE
Indian J Med Res 131, April 2010, pp 578-583
578

Acinetobacter baumannii / calcoaceticus are oxidase-negative, non-motile bacteria which appear as Gram-negative coccobacilli. These bacteria are widely found in nature, mostly in water and soil. This opportunistic pathogen has also been isolated from the normal flora of human skin, respiratory tract, gastrointestinal tract and various other sites in patients, especially in immunocompromised patients1. A. baumannii / calcoaceticus is often associated with hospital-acquired infections such as blood stream infections, meningitis, ventilator-associated pneumonia, wound, and urinary tract infections. It has been reported worldwide and is now becoming one of the most difficult healthcare-associated infections to control and treat2. The treatment of A. baumannii/ calcoaceticus infections is becoming a serious problem due to multiple acquired resistances following indiscriminate use of antibiotics and often carbapenems remain as the only alternative drug for the treatment of Acinetobacter spp. infection.
The common carbapenem resistance mechanisms in A. baumannii / calcoaceticus include acquisition of carbapenemases3-7, β-lactamases8 which are capable of hydrolyzing carbapenems9, reduced affinity of penicillin binding proteins10, and low permeability of outer membrane protein11-14. However, a few studies have shown the importance of the outer membrane protein in carbapenem resistant A. baumannii / calcoaceticus isolates 11-13,16,17. Almost all the previously reported studies have shown the role of OMP towards carbapenem resistant A. baumannii / calcoaceticus involving one mechanism.
Iron is one of the most important nutrients required by all forms of living organisms including microorganisms. However, the availability of iron is poor in the environment as most of it is found complexed with transferrin, lactoferrin, ferritin, haem, and haemoglobin18,19. In order to survive in a human host, bacteria form siderophores called iron-regulated outer membrane proteins (IROMPs). It has been reported that monoclonal antibodies raised against IROMPs of A. baumannii are bactericidal20. Here, in this study, we have incorporated another important factor to further analyze our findings to investigate the survival of carbapenem resistant A. calcoaceticus with the loss of 29-kDa outer membrane protein in iron deficit conditions.
Material & Methods
Bacterial strains and susceptibility testing: Isolates of carbapenem-resistant Acinetobacter spp. (n=39)
were obtained from patients who were hospitalized in University Malaya Medical Centre, Kuala Lumpur, Malaysia, from August 2003 until March 2004. This included one representative strain of A. calcoaceticus S26 which had a unique resistance mechanism, i.e., the presence of IMP-4 metallo-b-lactamases. The strains were obtained from invasive and non invasive sites which include blood, tracheal secretion, sputum, throat swab, peritoneal fluid, wound, bronchial lavage, and urine. Besides that, these strains were also selected based on their resistance to carbapenems, imipenem and meropenem. API20NE System (bioMérieux SA, Marcy-1’Etoile, France)21 and growth at 44◦C was performed to identify and differentiate between A. baumannii and A. calcoaceticus22. Further species level identification was done by amplified ribosomal DNA restriction analysis (ARDRA) as described by Johannes et al23. Briefly, 1500 bp fragment of the 16S rRNA gene was amplified using the universal primers - forward 5’- GGCTCAGATTGAAACGCTGGCGGC-3 and reverse 5’ TACCTTGTTACGACTTCACCCCA-3’. The PCR reaction mixture of the final volume of 50 µl contained 20 pM of each primer, 200 µM deoxynucleotide triphosphate, 1x reaction buffer, 1.5 mM MgCl2, 2.5 U Taq DNA polymerase (Fermentas, Lithuania, USA) and approximately 1 µl of template DNA. The thermocycle protocol used was: an initial denaturation step at 94◦C for 5 min, followed by 25 cycles of denaturation at 94◦C for 45 sec annealing at 58◦C for 45 sec, elongation at 72◦C for 3 min, followed by a final extension step at 72◦C for 7 min. The restriction enzymes AluI, CfoI, MboI, RsaI and MspI (Gibco BRL, Paisley, UK) were used in separate reactions to digest the amplimers, according to the manufactures’ instructions. Restriction fragments were resolved in 2 per cent agarose gel and the profiles were compared with those reported earlier23,24.
Minimum inhibitory concentrations (MICs) for the A. baumannii / calcoaceticus isolates were determined and interpreted according to Clinical Laboratory Standard Institute (CLSI, 2005)25. The antibiotics used in this study include imipenem, meropenem, ceftazidime, cefotaxime, gentamicin, amikacin, ciprofloxacin and ampicillin.
Outer membrane protein extraction: Outer membrane proteins were prepared based on the modified method of Matsuyama et al26. Briefly, A. calcoaceticus was grown on 50 ml of iron rich medium and on iron-deficit medium to early stationary growth phase. Cells were pelleted by centrifugation for 10,000 X g for 20 min at 4◦C, washed twice with 10 mM phosphate buffer, pH
HWA et al: IROMPS & CARBAPENEM RESISTANCE IN ACINETOBACTER SPP. 579

7.2. The cells were then resuspended in 20 ml of the same buffer before being subjected to cell disruption by sonication for 30s at 30 KHz output with 30 sec intervals. Cell debris was removed by centrifugation at 10,000 X g for 20 min at 4◦C. The supernatant was transferred to an ultra tube and the outer membrane fraction was pelleted by ultra centrifugation at 100, 000 X g for 35 min at 4◦C. The pellet was then resuspended in 20 ml of 10 mM phosphate buffer, pH 7.2 containing 2 per cent (v/v) Triton X-100. The suspension was incubated at 37◦C for 20 min. The Triton X-100 insoluble fraction was recovered by ultracentrifugation at 100,000 X g for 35 min at 4◦C. The pellet was then washed once with 10 ml phosphate buffer, pH 7.2 and recovered by centrifugation at 100,000 X g for 35 min at 4◦C. The protein pellet was resuspended in 100 µl of Laemmli buffer (Sigma-Aldrich, St. Louis, USA) and stored at -20◦C till use. The concentration of protein was determined using Bradford method27 and the protein samples were analyzed via SDS-PAGE.
SDS-PAGE: SDS-PAGE analysis was performed with the mini-protein II system (8.2 by 7.2 by 0.75 cm; BioRad Laboratories, USA). The stacking gel and separating gel contained 4 and 12 per cent acrylamide respectively. A total of 25 µg of protein of each sample was denatured by boiling for 10 min and cooled immediately prior to loading. Electrophoresis was carried out at constant voltage at 100 V for 90 min. The protein gels were visualized by staining with Coomassie blue R (Sigma, St. Louis, USA). The apparent molecular weights of the proteins detected were estimated by comparison with a broad-range protein standard obtained from New England Biolabs, United Kingdom. One well served as a negative control which contained only sample dilution buffer and loading dye.
Extraction of IROMPs: IROMPs were characterized according to a modified method of Goel and Kapil20. Briefly, Acinetobacter calcoaceticus were grown in iron depleted chemically defined medium (CDMFe-) and in iron rich (CDMFe+) medium. Medium without iron (CDMFe-) contained less than 0.04 mM Fe3+, and for medium with iron (CDMFe+), FeSO4 was added to give a concentration of 60 mM. Overnight cultures in CDMFe- and CDMFe+ were allowed to grow at 37◦C with aeration (150 rpm) until late log phase at optical density of (OD650) of 0.4. The cultures were then subjected to OMPs extraction according to the modified method of Matsuyama et al26. IROMPs extracted from the SDS-PAGE gel were used as antigen to immunize rabbits.
Preparation of polyclonal sera: To raise polyclonal antibodies in rabbit, a first dose of IROMP (300 µg/ml) was given intraperitoneally (ip) mixed with 250 µl of complete Freund’s adjuvant (CFA) followed by three doses (300 µg/ml each) in Freund’s incomplete adjuvant at intervals of one week. A final booster was given in normal saline, intraperitoneally, just four days before sacrifice. The experiments reported herein were conducted according to the principles in the Guidelines for the Care and Use of Laboratory Animals28. An ELISA was carried out to test the reactivity of the polyclonal antibodies from rabbit sera with the IROMP (10 µg/ml). Briefly, 96-well microtitter plates (Nunc, Roskilde, Denmark) were coated overnight at 4◦C with antigen diluted in phosphate buffer saline (PBS). The plates were washed four times by dispensing 250 µl PBS-T (PBS + 0.05% Tween) to each well. Then, the plates were blocked with 1 per cent BSA in PBS for 1 h at 37oC, and washed four times with PBS-T. One hundred microlitres of rabbit sera diluted 2-fold serially was added to the wells. The plate was incubated for 1 h at room temperature and washed four times with PBS-T. Goat anti-rabbit IgG horseradish peroxidase (Incstar Corporation, Stilwater Minnesota, USA) diluted at 1:2,000 was added to the wells. After 1 h of incubation at room temperature, the plates were washed four times with PBS-T and the last wash was with PBS only before the addition of the 2, 2’-azinobis (3-ethylbenzthiazoline sulphonic acid) (ABTS) substrate (Kirkegaard & Perry Laboratories Inc., Gaithersburg, Meryland, USA). Optical densities at 410 nm (OD410) were measured after 15 min of incubation at room temperature.
Bactericidal activity: Total viable count was performed to demonstrate bactericidal activity of polyclonal antibodies raised against IROMPs according to the modified method described by Poolman et al29. First, a standard growth curve was plotted and used as control. Briefly, 1 ml of known amount of A. calcoaceticus culture approximately 108 cfu/ml was inoculated into CDMFe- medium and CDMFe+ medium. The cultures were vortexed well and incubated at 37◦C with shaking at 250 rpm. Every half an hour, 10 µl of the culture suspensions were diluted in 90 µl of Luria broth in a test tube and followed by a 10-fold serial dilution until to 101 cfu/ml. Each serially diluted culture suspensions about 100 µl each was spread onto Mueller-Hinton agar plates. The plates were incubated at 37◦C overnight. Total viable count was done between 18 and 24 h. This procedure was repeated every half hour until the bacterial growth reached late log phase. Following
580 INDIAN J MED RES, APRIL 2010

this, total viable count method was performed on polyclonal antibodies to obtain bactericidal activity. In this experiment, polyclonal antibodies with 2 different dilution factors were used to obtain different concentration of antisera; 1:5 and 1:10. The appropriate concentrations of polyclonal antibodies were added to the A. calcoaceticus culture medium at early stationary phase. Finally, the growth curve graphs were plotted and the data obtained were analyzed.
Results & Discussion
All the 39 isolates were identified as A. baumannii-calcoaceticus complex using the API20NE system. Growing at 44◦C showed that 36 were identified as A. baumannii and 3 were A. calcoaceticus. The species were further confirmed by amplified ribosomal DNA restriction analysis (Fig. 1). All isolates were multi-resistant to all the antimicrobials tested including imipenem and meropenem (Table). In a previous study we have shown that Class 1 integrons were detected
in 31 of 39 isolates8. Two of the isolates harboured an integron-borne blaIMP-4 metallo-β-lactamase, 1 of which was located on a 36-kb plasmid.
The OMP profile via SDS-PAGE showed that all the carbapenem resistant A. baumannii / calcoaceticus isolates had a missing OMP band at about 29-kDa (marked by the red arrow) as shown in Fig. 2 suggesting that loss of a 29-kDa OMP is one of the leading factors to carbapenem resistance among these A. baumannii / calcoaceticus isolates.
The S26 isolate was used as representative of carbapenem resistant isolate with the loss of 29-kDa OMP and its survival when grown in iron deficit medium, was investigated. S26 was seen to overexpress bands corresponding to IROMPs in comparison to the isolate which was grown in the presence of iron in the medium (Fig. 3).
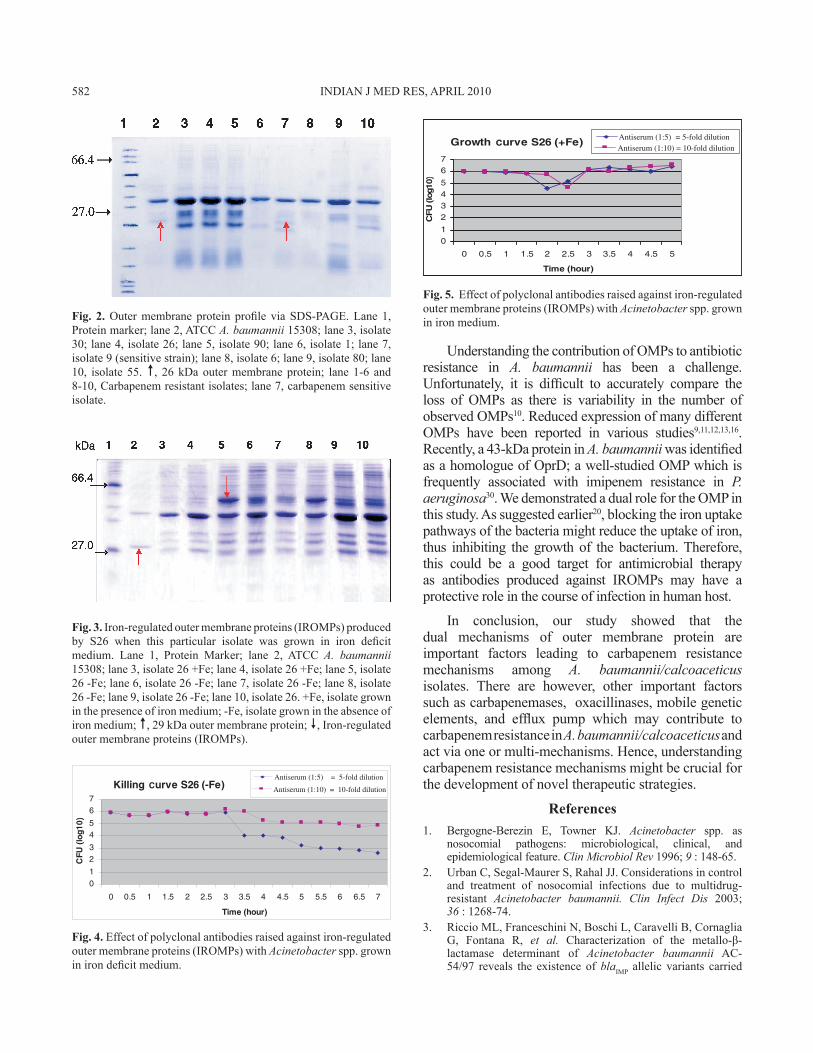
The polyclonal antibodies raised against IROMPs in rabbits showed bactericidal activity against the organism tested and specifically killed the bacteria grown in iron deficit medium in a dose dependant manner (Fig. 4). Five-fold dilution of the polyclonal antibody reduced the growth of A. calcoaceticus from 106 to 104 cfu/ml from the time point of 3 to 3.5 h up to 7 h suggesting that the polyclonal antibodies raised against IROMPs are bactericidal. However, in the presence of iron, the bactericidal effect of the polyclonal antibodies was only observed up to the time point of 1.5 to 2 h (Fig. 5) after which the organism recovered from the bactericidal effect and started to grow well.
Table. MICs of β-lactams for S26 (representative strain used for animal study) and other 38 isolates
MIC (µg/ml) for isolates (Σn=39) β-lactams Animal study
(n=1) S26Other isolates
(n=38)Imipenem 32 16 - 64Meropenem 128 32 - 128Ceftazidime 256 8 - 512Cefotaxime 512 32 - >512n = number of isolatesΣn = total number of isolates
Fig. 1. RE Digestion of 16S rDNA using AluI, CfoI and MboI enzymes for species level identification. Lane 1, 100 bp; lanes 2-5: A. baumannii: lane 2, isolate 6; lane 3, isolate 12; lane 4, isolate 13; lane 5, isolate 14 ; lanes 6-8: A. calcoaceticus: lane 6, isolate 9; lane 7, isolate 26; lane 8, isolate 90; lane 9, 100 bp; lane 10-13: A. baumannii: lane 10, isolate 6; lane 11, isolate 12; lane 12, isolate 13; lane 13, isolate 14; lane 14-16: A. calcoaceticus: lane 14, isolate 9; lane 15, isolate 26; lane 16, isolate 90; lane 17, 100 bp; lane 18-22: A. baumannii: lane 18, isolate 6; lane 19, isolate 12; lane 20, isolate 13; lane 21, isolate 14; lane 22, isolate 15; lane 23-25: A. calcoaceticus: lane 23, isolate 9; lane 24, isolate 26; lane 25, isolate 90; lane 26, 100 bp; lane 27, A. baumannii ATCC 15308 (AluI digest); lane 28, A. baumannii ATCC 15308 (CfoI digest); lane 29, A. baumannii ATCC 15308 (MboI digest); lane 30, 100 bp.
HWA et al: IROMPS & CARBAPENEM RESISTANCE IN ACINETOBACTER SPP. 581
Understanding the contribution of OMPs to antibiotic resistance in A. baumannii has been a challenge. Unfortunately, it is difficult to accurately compare the loss of OMPs as there is variability in the number of observed OMPs10. Reduced expression of many different OMPs have been reported in various studies9,11,12,13,16. Recently, a 43-kDa protein in A. baumannii was identified as a homologue of OprD; a well-studied OMP which is frequently associated with imipenem resistance in P. aeruginosa30. We demonstrated a dual role for the OMP in this study. As suggested earlier20, blocking the iron uptake pathways of the bacteria might reduce the uptake of iron, thus inhibiting the growth of the bacterium. Therefore, this could be a good target for antimicrobial therapy as antibodies produced against IROMPs may have a protective role in the course of infection in human host.
In conclusion, our study showed that the dual mechanisms of outer membrane protein are important factors leading to carbapenem resistance mechanisms among A. baumannii/calcoaceticus isolates. There are however, other important factors such as carbapenemases, oxacillinases, mobile genetic elements, and efflux pump which may contribute to carbapenem resistance in A. baumannii/calcoaceticus and act via one or multi-mechanisms. Hence, understanding carbapenem resistance mechanisms might be crucial for the development of novel therapeutic strategies.
ReferencesBergogne-Berezin E, Towner KJ. 1. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological feature. Clin Microbiol Rev 1996; 9 : 148-65.Urban C, Segal-Maurer S, Rahal JJ. Considerations in control 2. and treatment of nosocomial infections due to multidrug-resistant Acinetobacter baumannii. Clin Infect Dis 2003; 36 : 1268-74.Riccio ML, Franceschini N, Boschi L, Caravelli B, Cornaglia 3. G, Fontana R, et al. Characterization of the metallo-β-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of blaIMP allelic variants carried
Fig. 2. Outer membrane protein profile via SDS-PAGE. Lane 1, Protein marker; lane 2, ATCC A. baumannii 15308; lane 3, isolate 30; lane 4, isolate 26; lane 5, isolate 90; lane 6, isolate 1; lane 7, isolate 9 (sensitive strain); lane 8, isolate 6; lane 9, isolate 80; lane 10, isolate 55. �, 26 kDa outer membrane protein; lane 1-6 and 8-10, Carbapenem resistant isolates; lane 7, carbapenem sensitive isolate.
Fig. 3. Iron-regulated outer membrane proteins (IROMPs) produced by S26 when this particular isolate was grown in iron deficit medium. Lane 1, Protein Marker; lane 2, ATCC A. baumannii 15308; lane 3, isolate 26 +Fe; lane 4, isolate 26 +Fe; lane 5, isolate 26 -Fe; lane 6, isolate 26 -Fe; lane 7, isolate 26 -Fe; lane 8, isolate 26 -Fe; lane 9, isolate 26 -Fe; lane 10, isolate 26. +Fe, isolate grown in the presence of iron medium; -Fe, isolate grown in the absence of iron medium; �, 29 kDa outer membrane protein; �, Iron-regulated outer membrane proteins (IROMPs).
Fig. 4. Effect of polyclonal antibodies raised against iron-regulated outer membrane proteins (IROMPs) with Acinetobacter spp. grown in iron deficit medium.
Fig. 5. Effect of polyclonal antibodies raised against iron-regulated outer membrane proteins (IROMPs) with Acinetobacter spp. grown in iron medium.
582 INDIAN J MED RES, APRIL 2010

by gene cassettes of different phylogeny. Antimicrob Agents Chemother 2000; 44 : 1229-35.Takashi A, Yomoda S, Kobayashi I, Okubo T, Tsunoda M, 4. Iyobe S. Detection of carbapenemase-producing Acinetobacter baumannii in a hospital. J Clin Microbiol 2000; 38 : 526-9. Poirel L, Naas T, Nicolas D, Collet L, Bellais S, Cavallo JD, 5. et al. Characterization of VIM-2, a carbapenem-hydrolyzing metallo-β-lactamase and its plasmid and integron borne gene from a Pseudomonas aeruginosa clinical isolate in France. Antimicrob Agents Chemother 2000; 44 : 891-7.Poirel L. Nordmann P. Carbapenem resistance in 6. Acinetobacter baumannii: mechanisms and epidemiology. Clin Microbiol Infect 2006; 12 : 826-36.Bonomo A, Szabo D. Mechanisms of multidrug resistance 7. in Acinetobacter species and Pseudomonas aeruginosa. Clin Infect Dis 2006; 43 : S49-56.Eng Hwa W, Subramaniam G, Navaratnam P, Sekaran SD. 8. Detection and characterization of class 1 integrons among carbapenem-resistant isolates of Acinetobacter spp. in Malaysia. J Microbiol Immunol Infect 2009; 42 : 54-62.Bou G, Cervero G, Dominguez MA, Quereda C, Martinez-9. Beltran J. Characterization of nosocomial outbreak caused by a multiresistant Acinetobacter baumannii strain with a carbapenem-hydrolyzing enzyme: High-level carbapenem resistance in A. baumannii is not due solely to the presence of beta-lactamases. J Clin Microbiol 2000; 38 : 3299-305.Fernandez-Cuenca F, Martinez-Martinez L, Conejo MC, Ayala 10. JA, Perea EJ, Pascual A. Relationship between beta-lactamase production, outer membrane protein and penicillin-binding protein profiles on the activity of carbapenem against clinical isolates of Acinetobacter baumannii. J Antimicrob Chemother 2003; 51 : 565-74. Clark RB. Imipenem resistance among 11. Acinetobacter baumannii: association with reduced expression of a 33-36 kDa outer membrane protein. J Antimicrob Chemother 1996; 38 : 245-51.Costa SF, Woodcock J, Gill M, Wise R, Barone AA, Caiafa 12. H, et al. Outer-membrane proteins pattern and detection of β -lactamases in clinical isolates of imipenem-resistant Acinetobacter baumannii from Brazil. Int J Antimicrob Agents 2000; 13 : 175-82.Limansky AS, Mussi MA, Viale AM. Loss of a 29-kilodalton 13. outer membrane protein in Acinetobacter baumannii is associated with imipenem resistance. J Clin Microbiol 2002; 40 : 4776-8.Mussi MA, Relling VM, Limansky A, Viale A. CarO, an 14. Acinetobacter baumannii outer membrane protein involved in carbapenem resistance, is essential for l-ornithine uptake. FEBS Lett 2007; 581 : 5573-8.Yun S-H, Choi CW, Park S-H, Lee J-C, Leem S-H, Choi 15. JS, et al. Proteomic analysis of outer membrane protein from Acinetobacter baumannii DU202 in tetracycline stress condition. J Microbiol 2008; 46 : 720-7.Sato K, Nakae T. Outer membrane permeability of 16. Acinetobacter calcoaceticus and its implication in antibiotic resistance. J Antimicrob Chemother 1991; 28 : 35-45.Siroy A, Molle V, Lemaitre-Guiller C, Vallenet D, Pestel-17. Caron M, Cozzone AJ, et al. Channel formation by CarO, the
Reprint requests: Dr Shamala Devi Sekaran, Department of Medical Microbiology, Faculty of Medicine, University of Malaya 50603 Kuala Lumpur, Malaysia
e-mail : [email protected]
carbapenem resistance-associated outer membrane protein of Acinetobacter baumannii. Antimicrob Agents Chemother 2005; 49 : 4876-83.Otto BR, Verweij-van Vught MJJ, Maclaren DM. Transferrin 18. and heme-compunds as iron sources for pathogenic bacteria. Crit Microbiol Rev 1992; 18 : 217-33.Noya F, Arias A, Fabiano E. Heme compounds as iron sources 19. for nonpathogenic Rhizobium bacteria. J Bacteriol 1997; 179 : 3076-8.Goel VK, Kapil A. Monoclonal antibodies against the 20. iron regulated outer membrane proteins of Acinetobacter baumannii are bactericidal. BMC Microbiol 2001; 1 : 16-23. Schreckenberger PC, Daneshrar MI, Weyant RS, Hollis DG. 21. Acinetobacter, Chryseobacterium, Moraxella, and other nonfermentative Gram-negative rods. In: Murray PR, Baron EJ, Jorgensen JH, Pfaller MA, Yolken RH, editors. Manual of clinical microbiology, 8th ed. Washington DC: American Society for Microbiology; 2003. p. 749-79.Bouvet PJM, Grimont PAD. Identification and biotyping of 22. clinical isolates of Acinetobacter. Ann Inst Pasteur Microbiol 1987; 138 : 569-78.Koeleman JGM, Stoof J, Biesmans DJ, Savelkoul PHM, 23. Vandenbroucke-Grauls CMJE. Comparison of amplified ribosomal DNA restriction analysis, random amplified polymorphic DNA analysis, and amplified fragment length polymorphism fingerprinting for identification of Acinetobacter genomic species and typing of Acinetobacter baumannii. J Clin Microbiol 1998; 36 : 2522-9.Vaneechoutte M, Dijkshoorn L, Tjernberg I, Elaichouni A, De 24. Vos P, Claeys G, et al. Identification of Acinetobacter genomic species by amplified ribosomal DNA restriction analysis. J Clin Microbiol 1995; 33 : 11-5.Clinical and Laboratory Standards Institute. 25. Performance standards form antimicrobial disk susceptibility testing. Fifteenth informational supplement. Wayne, Pa: Clinical and Laboratory Standards Institute; 2005. M100-S15.Matsuyama SI, Inokuchi K, Mizushima S. Promoter exchange 26. between OmpF and OmpC for osmoregulated major outer membrane proteins of Escherichia coli K12. J Bacteriol 1984; 158 : 1041-7.Bradford MM. A rapid and sensitive method for the 27. quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 1976; 72 : 248-54.Animal Care and Use Committee. Kuala Lumpur, Malaysia: 28. Laboratory Animal Centre, Faculty of Medicine, University of Malaya; 2004. Poolman JT, Wientjes FB, Hopman CTP, Zanen HC. Influence 29. of the length of lipopolysaccharide molecules on the surface exposure of class 1 and 2 outer membrane proteins of Neisseria meningitidis strain 2996 (B:2b:P1.2). In: Schoolnik, editor. The pathogenic Neisseriae. Washington DC: American Society of Microbiology; 1985. p. 562-70. Dupont M, Pages JM, Lafitte D, Siroy A, Bollet C. 30. Identification of an OprD homologue in Acinetobacter baumannii. J Proteome Res 2005; 4 : 2386-90.
HWA et al: IROMPS & CARBAPENEM RESISTANCE IN ACINETOBACTER SPP. 583





























