Embed Size (px)
Citation preview

Isana Rodrigues Silva
AVALIAÇÃO DO PERFIL DE METILAÇÃO DO DNA EM GENES SUPRESSORES TUMORAIS E
SEQUENCIAS REPETITIVAS EM TRABALHADORES DA CONSTRUÇÃO CIVIL EXPOSTOS A AGENTES
CARCINOGÊNICOS AMBIENTAIS E OCUPACIONAIS
Dissertação apresentada ao Programa de Pós-
graduação do Hospital de Câncer de Barretos Fundação
PIO XII para obtenção do Título de Mestre em Ciências da
Saúde.
Área de concentração: Oncologia
Orientador: Prof. Dr. Henrique César Santejo Silveira
Barretos, SP 2016

Isana Rodrigues Silva
AVALIAÇÃO DO PERFIL DE METILAÇÃO DO DNA EM GENES SUPRESSORES TUMORAIS E
SEQUENCIAS REPETITIVAS EM TRABALHADORES DA CONSTRUÇÃO CIVIL EXPOSTOS A AGENTES
CARCINOGÊNICOS AMBIENTAIS E OCUPACIONAIS
Dissertação apresentada ao Programa de Pós-
graduação do Hospital de Câncer de Barretos
Fundação PIO XII para obtenção do Título de Mestre
em Ciências da Saúde.
Área de concentração: Oncologia
Orientador: Prof. Dr. Henrique César Santejo Silveira
Barretos, SP 2016

FICHA CATALOGRÁFICA Preparada por Martins Fideles dos Santos Neto CRB 8/9570
Biblioteca da Fundação Pio XII – Hospital de Câncer de Barretos
S586a Silva, Isana Rodrigues.
Avaliação do perfil de metilação do DNA em genes supressores tumorais e sequências repetitivas em trabalhadores da construção civil expostos a agentes carcinogênicos ambientais e ocupacionais / Isana Rodrigues Silva. - Barretos, SP 2016.
105 f. : il. Orientador: Dr. Henrique César Santejo Silveira. Dissertação (Mestrado em Ciências da Saúde) – Fundação Pio XII – Hospital
de Câncer de Barretos, 2016. 1. Metilação. 2. Biomarcadores. 3. Epigenética. 4. Pirosequenciamento. 5.
Câncer. 6. Construção Civil I. Autor. II. Silveira, Henrique César Santejo. III. Título

FOLHA DE APROVAÇÃO
Isana Rodrigues Silva
Avaliação do perfil de metilação do DNA em genes supressores tumorais e sequências repetitivas em
trabalhadores da construção civil expostos a agentes carcinogênicos ambientais e ocupacionais.
Dissertação apresentada ao Programa de Pós-Graduação da Fundação Pio XII – Hospital de Câncer de
Barretos para obtenção do Título de Mestre em Ciências da Saúde - Área de Concentração: Oncologia
Data da aprovação: 01/04/2016
Banca Examinadora:
Prof.ª Dra. Elza Tiemi Sakamoto Hojo
Instituição: Faculdade de Filosofia Ciências e Letras de Ribeirão Preto, Departamento de Biologia - USP
Prof.ª Dra. Mariana Brait Rodrigues de Oliveira
Instituição: Johns Hopkins University
Prof. Dr. Henrique Cesar Santejo Silveira
Orientador
Prof. Dr. Adhemar Longatto Filho
Presidente da Banca

SUPORTE À PESQUISA POR AGÊNCIA DE FOMENTO
Este trabalho recebeu apoio da Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)
através de Auxílio à Pesquisa – Regular (processo número 2012/24279-1).
As opiniões, hipóteses e conclusões ou recomendações expressas neste material são de
responsabilidade dos autores e não necessariamente refletem a visão da FAPESP.

“Esta dissertação foi elaborada e está apresentada de acordo com as normas da Pós-
Graduação do Hospital de Câncer de Barretos – Fundação Pio XII, baseando-se no Regimento do
Programa de Pós-Graduação em Oncologia e no Manual de Apresentação de Dissertações e Teses
do Hospital de Câncer de Barretos. Os pesquisadores declaram ainda que este trabalho foi
realizado em concordância com o Código de Boas Práticas Científicas (FAPESP), não havendo nada
em seu conteúdo que possa ser considerado como plágio, fabricação ou falsificação de dados. As
opiniões, hipóteses e conclusões ou recomendações expressas neste material são de
responsabilidade dos autores e não necessariamente refletem a visão da Fundação Pio XII –
Hospital de Câncer de Barretos.”
“Embora o Núcleo de Apoio ao Pesquisador do Hospital de Câncer de Barretos tenha
realizado as análises estatísticas e orientado sua interpretação, a descrição da metodologia
estatística, a apresentação dos resultados e suas conclusões são de inteira responsabilidade dos
pesquisadores envolvidos. Os pesquisadores declaram não ter qualquer conflito de interesse
relacionado a este estudo.”

Dedico este trabalho a Deus, por dar sabedoria todos os dias
para que eu pudesse suportar momentos e situações difíceis.
Aos meus amados pais Elisabete Rodrigues Silva e Ari Souza
da Silva que sempre acreditaram na minha capacidade, sem
eles nenhum dos meus objetivos teriam se tornado realidade.
Ao meu irmão Igor Rodrigues Silva por acreditar na
profissional que sou. E ao meu avô José Rodrigues (em
memória) que esteve presente comigo quando precisei, com
vibrações positivas.

AGRADECIMENTOS
“Você pode sonhar, criar, desenhar e construir o lugar mais bonito do mundo. Mas é
necessário ter pessoas para transformar seu sonho em realidade.” (Walt Disney). Neste momento,
venho expressar todo meu agradecimento e gratidão, do fundo do meu coração à todas as
pessoas que contribuíram com a realização desse trabalho. Com toda certeza, sem vocês esse
sonho não teria se concretizado. A gratidão é o único tesouro dos humildes. (William
Shakespeare). Muito obrigada.
Agradeço a Deus, por me conceder sabedoria e me ajudar a suportar momentos difíceis. E
no final você vai olhar para trás e agradecer cada tropeço. Acredite, Deus não falha.
A minha mãe, Elisabete, que de tudo sabe um pouco e qualquer situação ou problema que
eu estava passando, sempre tinha a palavra certa nos momentos certos que me faziam pensar.
Sempre acreditou em mim, na minha capacidade, quantas vezes, só de estar ao seu lado eu
recuperava todas as minhas energias para continuar.
Ao meu pai, Ari, meu grande herói, que sempre acredita no meu potencial, que sou capaz de
realizar minhas metas, reconhecendo meu esforço e auxiliando em tudo que precisava. Meu total
suporte. Obrigada por torcer por mim.
Ao meu irmão, Igor, que uma vez disse que sou um exemplo para ele. Espero estar sendo o
melhor exemplo possível, mesmo com defeitos, mas tentando acertar sempre. Obrigada pela
torcida.
Quantas vezes voltei para casa apenas para poder sentir a vibração positiva de vocês e
recarregar minhas energias para poder voltar e conseguir prosseguir.
Ao meu grande amigo, inesquecível, Luiz Manoel, que acompanhou todos os meus
momentos bons e ruins de muito perto, aguentando muitas vezes lágrimas e também sorrisos.
Também me escutando, o que me aliviava muitas vezes. Fazia de tudo para me distrair e me deixar
feliz. Obrigada por entender os momentos em que precisava estudar. Obrigada por estar comigo
em todos os momentos. Fez a diferença. Obrigada por tudo.
A toda minha família que de alguma forma me proporcionaram chegar até aqui, minha tia
Silvia, onde morei por um ano, minha avó Mileide (em memória), que também me cedeu moradia
por um ano no começo dos estudos. Obrigada.

Se você passar por uma guerra no trabalho, mas tiver paz quando chegar em casa, será um
ser humano feliz. Mas, se você tiver alegria fora de casa e viver uma guerra na sua família, a
infelicidade será sua amiga. (Augusto Cury).
Ao meu orientador, Dr. Henrique César Santejo Silveira, que me cedeu a oportunidade de
realizar o mestrado em uma instituição respeitada e contribuiu para mais um avanço na minha
formação profissional.
Ao grande amigo e profissional, Msc. André Van Helvoort Lengert, desde o início me
incentivando, acreditando em mim sempre, na minha capacidade. Foi um dos responsáveis pelo
grande aprendizado que obtive durante esse tempo. Sem sua ajuda eu não teria concluído essa
dissertação. Meus sinceros agradecimentos, muito obrigada.
À Dra. Adriane Feijó Evangelista, pela grande ajuda e disponibilidade. Pronta sempre a me
socorrer em momentos tensos, mesmo estando de férias. Obrigada pelas dicas e por passar seus
conhecimentos que não são poucos todas as vezes que precisava.
À Dra. Lídia Maria Rebolho Batista Arantes, por toda ajuda com seus conhecimentos e
sempre disposta a auxiliar, seja qual fosse a questão. Com certeza umas das pessoas essenciais
para a concretização desse trabalho. Meu muito obrigada.
Costumo dizer que devo ter acumulado alguns carmas positivos ao longo da vida, por ter
tantas pessoas boas que me rodeiam e estão dispostos a ajudar. Sempre que estava precisando de
algo, Deus colocava alguém especial no meu caminho pronto a me ajudar e se tornando mais um
amigo(a), foi assim com a Dra. Hellen Kuasne, que me deu muito auxílio na parte de metodologia,
sua ajuda foi essencial para o decorrer desse trabalho. Obrigada por cada dúvida sanada, sempre
com muito carinho e prontidão. Muito obrigada.
Aos amigos que vou levar no meu coração para toda a vida, que estiveram presentes
durante minha caminhada, minha eterna amiga, amiga de alma, de outras vidas, Manoela
Catarina Aguena Souza Ramos, o maior presente que ganhei durante minha caminhada foi ter
conhecido você e hoje poder te chamar de “amiga”. Obrigada por cada momento, foram os
melhores, por toda ajuda, por me socorrer várias vezes no início nos meus desesperos, por querer
ajudar mesmo estando morta de cansaço. Eu me considero uma pessoa de muita sorte por ter te
conhecido e ter tido a oportunidade de conviver com você. Mesmo não estando presente, não
teve um dia sequer que não pensava “Ah, se a Manú estivesse aqui”. Finalizo esse trabalho com
orgulho e também dedico à você. Obrigada amiga.

À minha amiga, Aline Oliveira da Rocha, obrigada por cada momento de desabafo que
tivemos, paciência e carinho, por acreditar que eu chegaria até o fim, o reconhecimento é
essencial, obrigada. Às amigas, Msc.Taciane, Fernanda Franco Munari, Isabella Alves Brunetti,
Karen Borba, Msc. Tatiane Siqueira, Rhafaella Causin, Caroline Rogeri e Msc. Danielle, obrigada a
todas por todo o auxílio e ajuda, contribuindo com seus conhecimentos e parceria. Vocês fizeram
o meu dia-a-dia que não eram fáceis, se tornarem mais leves com os sorrisos e descontrações.
Ficou mais fácil com vocês.
Agradeço à família CPOM pela convivência nesses últimos anos, Adriana Carlone (Dry),
Adriana Lorenzi, Mathias, Maraisa, Ana Luiza, Rafael Tiveron, Renato, Carolzinha, Carol Laus,
Vânia, Estela, Paula, Paulinha, Karina, Natália Campacci, Nathália Campanella, Tati Honda,
Marcela, Viviane, Ana Laura, Ana Gabriela, Aline Coelho, Úrsula, Weder, Cintia, Vitor, à
Fernanda Cury, pelas conversas e toda paciência no início.
A todos os pesquisadores do CPOM e IEP, pelas sugestões em reuniões e journal.
Aos membros da banca de acompanhamento, Dra. Mariana Brait, sempre muito gentil,
contribuindo para o trabalho se concretizar de maneira excelente. Dr. Rui Reis também se
disponibilizando a ajudar através de críticas, sugestões e contribuições a este trabalho. Foi uma
honra tê-los em minha banca.
Ao programa de pós-graduação, em especial à Silvana Rodrigues e Brenda Honda.
Aos amigos e colaboradores do Núcleo de Apoio ao Pesquisador (NAP), por todo
comprometimento, desde o início, meu muito obrigada a toda a equipe, Tamira, Thaís, Cleyton,
Marcos, Jamile e meu amigo Marco por toda colaboração na parte estatística, me ajudando a ter
vários raciocínios, deixando a estatística mais fácil de ser entendida (se é que isso é possível), por
todos os e-mails, principalmente os fora de horário de expediente e também finais de semana,
pronto sempre a me ajudar e sanar minhas dúvidas. Obrigada por tudo. Aos amigos do
condomínio, Ronaldo, Monise, Kleyton, obrigada por fazer meus dias mais divertidos.
Por fim, chego ao final de mais um desafio que tracei em minha vida e com muito esforço e
dedicação, concluo com êxito. Durante toda a jornada, aprendi muito, aprendi como ser e como
não ser, como lidar com várias situações. Chorei muito, mas também sorri muito. Conheci e
convivi com pessoas maravilhosas que me fizeram ser alguém melhor. E agora o sentimento é
apenas de gratidão à todos que contribuíram e muito orgulho por ter realizado um trabalho com
muita ética. “Aquele que é possuidor de verdadeira sabedoria não se envaidece por seus êxitos;

ele mantém sempre a humildade, atribuindo a Deus o mérito da sua vitória.” (Masaharu
Taniguchi).
O aprendizado é constante.
O meu sincero Muito Obrigada a todos.

O AMOR
Quando propus a teoria da relatividade, muito poucos me entenderam e o que vou agora revelar a você,
para que transmita à humanidade, também chocará o mundo, com sua incompreensão e preconceitos.
Peço ainda que aguarde todo o tempo necessário, anos, décadas, até que a sociedade tenha avançado o
suficiente para aceitar o que explicarei em seguida para você.
Há uma força extremamente poderosa para a qual a ciência até agora não
encontrou uma explicação formal. É uma força que inclui e governa todas as outras, existindo por trás de
qualquer fenômeno que opere no universo e que ainda não foi identificada por nós.
Esta força universal é o AMOR.
Quando os cientistas estavam procurando uma teoria unificada do Universo esqueceram a mais invisível e
poderosa de todas as forças.
O Amor é Luz, dado que ilumina aquele que dá e o que recebe.
O Amor é gravidade, porque faz com que as pessoas se sintam atraídas umas pelas outras.
O Amor é potência, pois multiplica (potencia) o melhor que temos, permitindo assim que a humanidade
não se extinga em seu egoísmo cego.
O Amor revela e desvela. Por amor, vivemos e morremos. O Amor é Deus e Deus é Amor.
Esta força tudo explica e dá SENTIDO à vida. Esta é a variável que temos
ignorado por muito tempo, talvez porque o amor provoca medo, sendo o único poder no universo que o
homem ainda não aprendeu a dirigir a seu favor.
Para dar visibilidade ao amor, eu fiz uma substituição simples na minha
equação mais famosa. Se em vez de E = mc², aceitarmos que a energia para curar o mundo pode ser obtido
através do amor multiplicado pela velocidade da luz ao quadrado (energia de cura = amor x velocidade da
luz ²), chegaremos à conclusão de que o amor é a força mais poderosa que existe, porque não tem limites.
Após o fracasso da humanidade no uso e controle das outras forças do
universo, que se voltaram contra nós, é urgente que nos alimentemos de outro tipo de energia. Se
queremos que a nossa espécie sobreviva, se quisermos encontrar sentido na vida, se queremos salvar o
mundo e todos os seres sensíveis que nele habitam, o amor é a única e a resposta última.
Talvez ainda não estejamos preparados para fabricar uma bomba de amor, uma criação suficientemente
poderosa para destruir todo o ódio, egoísmo e ganância que assolam o planeta. No entanto, cada indivíduo
carrega dentro de si um pequeno, mas poderoso gerador de amor, cuja energia aguarda para ser libertada.
Quando aprendemos a dar e receber esta energia universal, provaremos que o amor tudo vence, tudo
transcende e tudo pode, porque o amor é a quintessência da vida.
Albert Einstein

ÍNDICE
1 INTRODUÇÃO 1
1.1 Câncer Ocupacional 1
1.2 Exposição ocupacional em trabalhadores da construção civil 3
1.3 Epigenética 7
1.4 Metilação do DNA para determinação dos sujeitos com alto risco ao desenvolvimento de câncer 12
2 JUSTIFICATIVA 15
3 OBJETIVOS 16
3.1 Objetivo geral 16
3.2 Objetivos específicos 16
4 MATERIAL E MÉTODOS 17
4.1 Local de estudo 17
4.2 Desenho do estudo 17
4.3 Sujeitos da pesquisa 17
4.4 Critérios de inclusão 17
4.5 Critérios de exclusão 18
4.6 Coleta de dados 18
4.7 Coleta de sangue 18
4.8 Determinação da metilação do DNA na região promotora de genes 18
4.9 Análise estatística dos dados 24
4.10 Aspectos Éticos 25
5 RESULTADOS 26
5.1 Coleta e caracterização sócio-demográficas dos grupos 26
5.2 Avaliação do perfil de metilação do DNA entre trabalhadores da construção civil (expostos) e
controles não-expostos (caso x controle) 30
5.3 Avaliação do nível de metilação dos genes com as covariáveis hábito de bebida e fumante passivo
para os grupos caso e controle. 39
5.4 Avaliação do nível de metilação dos genes considerando as covariáveis hábito de bebida, fumante
passivo e ex-fumantes para os grupos caso/controle. 42

5.5 Análise do perfil de metilação do DNA nos trabalhadores da construção civil expostos no 1º dia de
trabalho da semana (segunda- feira) e 5º dia de trabalho da semana (sexta- feira). 44
6 DISCUSSÃO 47
7 CONCLUSÕES 52
REFERÊNCIAS 53
ANEXOS
Anexo A – Questionário sócio-demográfico aplicado aos grupos 62
Anexo B – Condições estabelecidas de ciclagem da PCR dos genes supressores tumorais e sequências
repetitivas (°C). 67
Anexo C - Sequência dos primers e região analisada dos genes analisados 68
Anexo D – Sequência genômica dos genes de acordo com o banco de dados da UCSC 69
Anexo E – Matriz de exposição aplicado aos trabalhadores da construção. 71
Anexo F – Parecer consubstanciado do CEP 73
Anexo G – Análise dos motifs das regiões promotoras dos genes CDKN2A, MLH1 e APC utilizando o banco
de dados JASPAR. 80

LISTA DE FIGURAS
Figura 1 - Esquerda: Representação esquemática de modificações epigenéticas. no DNA e nas histonas.
Direita: Esquema representando os microRNAs, que são pequenos RNAs não-codificantes, que reconhece
um RNAm alvo por homologia de sequência, levando a clivagem e o bloqueio da tradução. (Adaptado de
Mansuy et al., 201162). 8
Figura 2 – Representação da ligação do grupo CH3 no carbono 5 da citosina (C) gerando a 5-metilcitosina,
sendo este processo mediado pelas enzimas DNA metiltransferases (DNMT’s). (Retirado e modificado de
Lorincz73). 10
Figura 3 – Representação esquemética do protocolo de conversão por bissulfito de sódio. Adaptado de
Epitech Bisulfite kit (Qiagen)111. 20
Figura 4 - Representação esquemática do sistema enzimático de pirosequenciamento. Cada dNTP é
incorporada na cadeia de DNA pela enzima polimerase, a qual adiciona as bases de acordo com a fita
molde, liberando assim a molécula de pirofosfato (PPi). A enzima ATP sulfurilase converte o PPi em ATP que
serve como substrato para a produção de luz pela enzima luciferase. A luz produzida é detectada como
evidência de que a incorporação de nucleotídeos foi eficiente (Retirado e adaptado de Ahmadian et al.113).
22
Figura 5 – Representação dos pirogramas de cada região promotora analisada para cada gene. (A)
Pirograma representando a região da sequência ALU. (B) Pirograma representando a região promotora do
gene RASSF1A. (C) Pirograma representando controle metilado. (D) Pirograma representando controle não-
metilado. (E) Pirograma representando a região promotora do gene APC. (F) Pirograma representando a
região da sequência LINE-1. (G) Pirograma representando a região promotora do gene MGMT. (H)
Pirograma representando a região promotora do gene MLH1. (I) Pirograma representando a região
promotora do gene CDKN2A. (J) Pirograma representando a região promotora do gene TP53. 23
Figura 6 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho atual
(1º grupo). 28
Figura 7 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho em
empregos anteriores (1º grupo). 28
Figura 8 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho atual
(2º grupo). 30
Figura 9 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho em
empregos anteriores (2º grupo). 30

LISTA DE TABELAS
Tabela 1 - Características sócio-demográficas dos participantes do primeiro grupo coletado (caso x
controle). 27
Tabela 2 - Características sócio-demográficos dos participantes do segundo grupo coletado (curto período
de exposição no ambiente da construção civil). 29
Tabela 3 - Avaliação do perfil de metilação do DNA na região promotora dos genes TP53, MGMT, RASSF1A
e sequência repetitiva ALU em trabalhadores da construção civil e no grupo controle. 31
Tabela 4 – Avaliação dos níveis de metilação do DNA da região promotora dos genes CDKN2A, MLH1, APC e
sequência LINE-1 em trabalhadores da construção civil e do grupo controle. 34
Tabela 5 - Nível de metilação do gene CDKN2A analisado de acordo com agentes expostos no ambiente de
trabalho atual. 35
Tabela 6 - Nível de metilação do gene CDKN2A analisado de acordo com agentes ocupacionais no ambiente
de trabalho em empregos anteriores. 35
Tabela 7 - Nível de metilação do gene MLH1 analisados de acordo com agentes ocupacionais no ambiente
de trabalho em empregos anteriores. 36
Tabela 8 - Nível de metilação do gene MLH1 analisados de acordo com agentes expostos no ambiente de
trabalho atual. 36
Tabela 9 - Nível de metilação do gene APC (Hot CpG sites) analisados de acordo com agentes ocupacionais
no ambiente de trabalho em empregos anteriores. 37
Tabela 10 - Nível de metilação do gene APC (Hot CpG sites) analisados de acordo com agentes ocupacionais
no ambiente de trabalho atual. 38
Tabela 11 - Nível de metilação da sequência repetitiva LINE-1 analisado de acordo com agentes
ocupacionais no ambiente de trabalho em empregos anteriores. 38
Tabela 12 - Nível de metilação da sequência repetitiva LINE-1 analisado de acordo com agentes
ocupacionais no ambiente de trabalho atual. 39
Tabela 13 - Níveis de metilação das regiões promotoras dos genes CDKN2A, MLH1, APC e da sequência
repetitiva LINE-1 em relação à exposição passiva ao fumo (grupo caso). 41
Tabela 14 - Níveis de metilação das regiões promotoras dos genes CDKN2A, MLH1, APC e da sequência
repetitiva LINE-1 em relação ao hábito de bebida (grupo caso). 41
Tabela 15 - Modelo de Regressão Linear Múltipla para porcentagem (%) de metilação dos genes CDKN2A,
MLH1, APC e sequência repetitiva LINE-1 em relação a hábito de fumante passivo, bebida alcoólica e ex-
fumantes (grupo caso x controle). 43

Tabela 16 - Exemplo de predição de acordo com o modelo de regressão linear para a média de
porcentagem (%) de metilação. 43
Tabela 17 - Análise da metilação do DNA da região promotora dos genes MGMT, RASSF1A, APC, MLH1 e
sequências ALU e LINE-1 em trabalhadores expostos no 1º e 5º dia da semana. 45
Tabela 18 - Análise da metilação do DNA da região promotora do gene CDKN2A em trabalhadores expostos
no 1º e 5º dia da semana. 46

LISTA DE ABREVIATURAS
ADP Adenosina difosfato
ALU Elementos intercalados curtos
APS Adenosina 5-fosfosulfato
ATP Trifosfato de Adenosina
CCD Charge Coupled Device
DNA Ácido Desoxirribonucleico
DNMTs DNA metiltransferases
dNTP Desoxirribonucleotídeos Fosfatados
EDTA Ácido etilenodiamino tetra-acético
EPA Agência de Proteção Ambiental
HPAs Hidrocarboneto Policíclicos Aromáticos
IARC Agência Internacional de Investigação do Câncer
KB Quilobase (1000 pares de bases)
LINE1 Elementos Intercalados longos
miRNA Micro ácido ribonucleico
mL Mililitro
MMR Mecanismo de reparo
MNs Micronúcleos
MP Material Particulado
NaOH Cloreto de Sódio
NAP Núcleo de Apoio ao Pesquisador
OMS Organização Mundial da Saúde
OSHA Segurança Ocupacional e administração da Saúde
PB Pares de bases
PBS Solução Salina fosfatada tamponada
PCR Reação em Cadeia da Polimerase
pH Potencial hidrogeniônico
PIB Produto Interno Bruto
PPi Pirofosfato
RNA Ácido Ribonucléico
RNAm RNA mensageiro
RPM Rotação por minuto

siRNA Small interfering RNA
TCLE Termo de Consentimento Livre e Esclarecido
UV Ultra-violeta

LISTA DE SÍMBOLO
% Porcentagem
R$ Real
µm Micrometro
°C Graus Celsius
mL Mililitro
µL Microlitro
Nm Nanometro
µg Micrograma
µM Micromolar
M Molar
> Maior que
≥ Maior ou igual a
< Menor que
≤ Menor ou igual a
ᵝ Beta

RESUMO
Rodrigues IS. Avaliação do perfil de metilação do DNA em genes supressores tumorais e
sequências repetitivas em trabalhadores da construção civil expostos a agentes carcinogênicos
ambientais e ocupacionais. Dissertação (Mestrado). Barretos: Hospital de Câncer de Barretos;
2016.
JUSTIFICATIVA: A exposição ambiental tem sido associada a um risco aumentado de
desenvolvimento de câncer, onde a metilação do DNA em regiões promotoras de genes
supressores de tumor tem surgido como possível biomarcador promissor para câncer, incluindo o
câncer de pulmão. No setor da construção civil a exposição dos trabalhadores a materiais
contendo amianto, sílica e metais, além da exposição à radiação solar, são fatores capazes de
interferir na estabilidade do genoma humano, provocando quebras no DNA e mudanças nos
padrões epigenéticos. Até o momento, nenhum estudo avaliou os níveis de metilação do DNA em
trabalhadores da construção, os quais podem estar associados ao desenvolvimento de câncer de
pulmão devido a exposição ocupacional. OBJETIVO: Este trabalho buscou avaliar possíveis
biomarcadores epigenéticos para associação do potencial risco de desenvolvimento de câncer em
trabalhadores da construção civil expostos a agentes carcinogênicos ambientais e ocupacionais.
Além disso, também foi avaliado o perfil de metilação do DNA no grupo de trabalhadores no
primeiro dia de trabalho da semana e no quinto dia de exposição, a fim de detectar diferenças na
metilação em um curto período de tempo. MATERIAIS E MÉTODOS: Foram selecionados seis
genes supressores de tumor (APC, RASSF1A, CDKN2A, TP53, MGMT, MLH1) e duas sequências
repetitivas (ALU e LINE-1), que já foram demonstrados hipermetilados e hipometilados,
respectivamente, em diversos tipos de exposição. O DNA foi extraído a partir de leucócitos de 58
amostras de trabalhadores do setor da construção (caso) e 50 trabalhadores do setor
administrativo (controle). Além disso, foram extraídas 39 amostras dos trabalhadores do setor da
construção, no primeiro e quinto dia da semana de trabalho. Posteriormente, as amostras foram
submetidas a conversão de bissulfito de sódio e os níveis de metilação das amostras foram
avaliadas pela metodologia de pirosequenciamento. Foram utilizados modelos de regressão linear
múltipla para verificação da relação conjunta de alguns fatores de confusão como: fumante
passivo, ex-fumante, etilismo. Para as análises estatísticas foi considerado um nível de significância
de 5%, p valor <0,05. RESULTADOS: Verificamos nas análises de metilação, médias maiores para os
genes CDKN2A (p=0,05), MLH1 (p=0,007), APC (p<0,001) e média menor dos níveis de metilação

da sequência repetitiva LINE-1 (p=0,005), para o grupo caso em relação ao controle. Sugere-se que
tais diferenças nos níveis de metilação são decorrentes da exposição a concreto (p=0,03), pó de
madeira (p=0,02) e sílica (p=0,05). Fatores de confusão, tais como, hábito de bebida, exposição
passiva ao cigarro e o critério de ser ex-fumante não influenciaram nos níveis de metilação dos
genes. Nos trabalhadores da construção coletados no 1º e 5º dia da semana, observaram-se níveis
maiores de metilação para o gene CDNK2A (p=0,04) no 5º dia de exposição, sugerindo que um
curto tempo de exposição pode influenciar nos níveis de metilação desse gene. CONCLUSÃO:
Estes achados sugerem que o ambiente da construção civil pode influenciar no perfil de metilação
de genes envolvidos no processo de carcinogênese. Além disso, pode levar a alterações na
metilação global, proporcionando instabilidade genômica. Nossos resultados sugerem que a
hipometilação de LINE-1 e hipermetilação de APC, CDKN2A e MLH1 podem ser utilizados como
biomarcadores para exposição ambiental e ocupacional, porém, investigações adicionais são
necessárias a fim de confirmar nossos achados.
Palavras Chave: Metilação do DNA; Biomarcadores; Epigenética; Pirosequenciamento; Câncer;
Trabalhadores da Construção.

ABSTRACT
Rodrigues. IS. Evaluation of DNA methylation profile in tumor supressor genes and repetitive
sequences on construction workers exposed to environmental and occupational carcinogens.
Dissertation (Master’s degree). Barretos: Hospital de Câncer de Barretos; 2016.
BACKGROUND: The environmental exposure has been linked to an increased risk of cancer, where
the DNA methylation in promoter regions of tumor suppressor genes has emerged as promising
potential biomarker for cancer, including lung cancer. In the construction sector the exposure of
workers to materials containing asbestos, silica and metals, and exposure to solar radiation, are
factors that can affect the stability of the human genome, causing breaks in DNA and epigenetic
alterations. To date, no study has evaluated the DNA methylation levels in construction workers,
which may be associated to the risk of lung cancer development due to the work environment.
OBJECTIVE: This study aimed to evaluate possible epigenetic biomarkers for association of the
potential risk of developing cancer in construction workers exposed to environmental and
occupational carcinogens. Furthermore, it was also evaluated the DNA methylation profile in
workers in the first and fifth day of exposure in a working week in order to detect the methylation
differences within a short period of time. MATERIALS AND METHODS: We selected six tumor
suppressor genes (APC, RASSF1A, CDKN2A, TP53, MGMT, MLH1) and two repetitive sequences
(ALU and LINE-1), which have been demonstrated hypermethylated and hypomethylated,
respectively, in various types of exposition. DNA was extracted from white blood cells of 58
samples of construction sector workers (case) and 50 workers in the administrative sector
(control). In addition, samples were collected from 39 workers from the construction sector in the
first and fifth day of the working week. Subsequently, the samples were subjected to sodium
bisulfite conversion and sample methylation levels were assessed by pyrosequencing
methodology. Multiple linear regression models were used to check the joint relationship of some
confounding factors such as passive smoker, ex-smoker, alcohol consumption. For statistical
analysis it was considered a significance level of 5%, p value <0.05. RESULTS: We verified higher
methylation levels for CDKN2A (p = 0.05), MLH1 (p = 0.007), APC (p <0.001) and lower methylation
levels of LINE-1 repetitive sequence (p = 0.005) for the case group compared to control. It is
suggested that such differences in methylation levels are from exposure to concrete (p = 0.03),
wood powder (p = 0.02) and silica (p = 0.05). Confounding factors, such as alcohol consumption,
passive smoker and ex-smoker did not influence the methylation levels of genes. In the

construction workers collected on 1st and 5th day, there were higher levels of CDKN2A
methylation (p = 0.04) on the 5th day of exposure, suggesting that a short exposure time can
influence the methylation levels of this gene. CONCLUSION: These findings suggest that the
construction environment can influence the methylation profile of genes involved in
carcinogenesis. Moreover, it can lead to changes in global methylation, providing genomic
instability. Our results suggest that hypomethylation of LINE-1 and hypermethylation of APC,
MLH1 and CDKN2A can be used as biomarkers for environmental and occupational exposure,
however, further investigations are needed to confirm our findings.
Keyword: DNA methylation; Biomarkers; Epigenetics; Pyrosequencing; Cancer; Construction
workers.

1
1 INTRODUÇÃO
1.1 Câncer Ocupacional
O câncer está entre as doenças que mais acometem a população mundial, sendo que no ano
de 2016/2017 as estimativas apontam para aproximadamente 600 mil novos casos, onde traquéia,
brônquios e pulmão em homens estima-se 17.330 novos casos e 10.890 em mulheres1.
Hoje no país, o câncer é um problema de saúde pública, sendo assim, controle e prevenção
deverão ser priorizados em todas as regiões, desde as mais desenvolvidas socioeconomicamente
até às mais desiguais2.
Podemos salientar que aproximadamente 80% dos casos de câncer estão relacionados a
fatores ambientais, em maior ou menor grau, evitáveis3. Esses fatores envolvem água, terra, ar,
ambiente de consumo (alimentos, medicamentos, fumo, álcool e produtos domésticos), ambiente
cultural (estilo, costumes e hábitos de vida) e ambiente ocupacional. Muitas substâncias ou a
mistura destas presentes no ambiente ocupacional e que levam ao aumento da incidência de
neoplasias ou uma redução no período de latência entre a exposição e o aparecimento da doença,
são consideradas substâncias com um potencial cancerígeno ocupacional4. Estima-se que
aproximadamente 10,8% dos casos de câncer (excluindo pele não melanoma) em homens e 2,2%
dos casos de câncer em mulheres são causados por exposição ocupacional5.
De acordo com a Occupational Safety and Health Administration (OSHA), é considerado um
potencial carcinógeno ocupacional qualquer substância, combinação ou mistura de substâncias
que causem aumento da incidência de neoplasias ou redução substancial no período de latência
entre a exposição e o aparecimento da doença em humanos ou mamíferos6.
No Brasil, estima-se que cerca de 23.040 dos casos de câncer para o ano de 2015 serão
desenvolvidos devido ao seu ambiente de trabalho1. A primeira observação entre a ocupação e
câncer foi descrita por Pott em 1775, que observou uma grande frequência de câncer da bolsa
escrotal em limpadores de chaminé em Londres, na Inglaterra, relacionando a incidência de câncer
nesta classe de trabalhadores decorrente a exposição a fuligem7. Atualmente, principalmente no
ambiente de trabalho industrial são introduzidas um número grande de substâncias e pouco se
sabe sobre a relação dos efeitos tóxicos ou não que essas substancias podem ocasionar9.
Entre os cânceres ocupacionais, pelo menos 19 tipos de tumores estão relacionados à
ocupação e ao ambiente de trabalho, entre eles os tipos, de pulmão, pele, fígado, laringe, bexiga e

2
leucemias3. Em relação ao câncer de pulmão, 90% dos casos em homens são atribuídos pelo
hábito de fumar8,9. Além disso, hoje é sabido que a influência ambiental e ocupacional são fatores
que contribuem como fator de risco para o câncer de pulmão. Entre os casos de câncer de pulmão
causadas decorrentes da exposição ocupacional podemos salientar que tem sido relatado em
cerca de 7,9% a 16,5% nos homens e 1,4% para 4,5% em mulheres10. Os principais fatores que
contribuem para esta porcentagem, são as exposições a amianto, as emissões dos motores diesel,
e outras misturas contendo hidrocarbonetos aromáticos policíclicos, sílica cristalina, arsênico e
alguns metais pesados11,12.
A grande maioria das associações da incidência de câncer e causas ocupacionais tem sido
demonstrada por estudos epidemiológicos. Em 1965 a IARC (International Agency for Research on
Cancer) foi criada pela Organização Mundial da Saúde (OMS), assim, o objetivo da IARC é
promover a colaboração internacional na pesquisa do câncer, reunindo competências em
epidemiologia, ciências laboratoriais e bioestatística para identificar as causas do câncer de modo
que podem ser adoptadas medidas de prevenção. Dessa forma, a agência ficou a cargo de
reconhecer o potencial ou provável caráter cancerígeno das substâncias, agentes ou outras formas
de exposição. Em relação a exposição ocupacional a IARC tem o papel de reconhecer os ambientes
complexos e as múltiplas exposições que ocorrem no ambiente de trabalho e que permitem a
caracterização de agentes isolados6. A IARC reúne anualmente grupos de especialistas
internacionais para elaborar suas publicações de monografias temáticas, possibilitando a
elaboração de um consenso das pesquisas6. Os resultados servem de referência para
recomendação da OMS. Os agentes ou substâncias são classificados em quatro grupos: o Grupo 1
o agente (mistura) é cancerígeno para humanos (evidência epidemiológica suficiente para
carcinogenicidade em seres humanos). O Grupo 2A, o agente é provavelmente cancerígeno para
humanos (provavelmente cancerígeno em seres humanos, segundo evidências limitadas em seres
humanos e evidências suficientes em animais). O grupo 2B possivelmente cancerígeno em seres
humanos segundo evidência suficiente em animais, porém, evidências inadequadas em seres
humanos ou limitada nesses com evidência suficiente em animais. O grupo 3 o agente não é
classificado como cancerígeno para humanos. E o grupo 4 o agente ou mistura é provavelmente
não cancerígeno para humanos6.
A questão saúde-trabalho deixou de ser analisada apenas como um simples indicador. Hoje
as condições do ambiente e segurança do trabalho e suas influências na saúde do trabalhador

3
passam a ser compreendidas como garantias essenciais para a qualidade de vida e os direitos dos
trabalhadores13.
Ao longo das últimas décadas, ocorreu um declínio nos cânceres atribuíveis a exposição
ambiental e ocupacional a carcinógenos, tais como, amianto, arsênico e poluição do ar em países
desenvolvidos. Por outro lado, em países em desenvolvimento ou subdesenvolvidos, à medida
que industrialização se expande ocorrerá o aumento da exposição a substancias e agentes
carcinogênicos o que tornará as populações mais susceptíveis ao câncer14. Um estudo realizado
por Ziech et al.15 sugere que a exposição ambiental, a agentes químicos e biológicos
desempenham um importante papel na ocorrência de câncer, como os fatores de estilo e hábito
de vida contribuindo como uma parte do todo. Por outro lado, os agentes carcinogênicos
presentes no meio ambiente e na ocupação estejam envolvidos significativamente e contribuindo
para a carcinogênese humana, interpretações quanto à extensão de tal contribuição permanecem
em grande parte obscura.
1.2 Exposição ocupacional em trabalhadores da construção civil
Uma população que apresenta risco ao câncer ocupacional são os trabalhadores da
construção civil, devido à exposição diária e por longos períodos a diversos agentes reconhecidos
como nocivos à saúde. Esses trabalhadores utilizam materiais que são constituídos por diversas
substâncias classificadas pela IARC como carcinogênico ou potencialmente carcinogênico aos seres
humanos. Uma destas substâncias é a sílica cristalina na forma de pó de quartzo, no ambiente de
trabalho pode ocorrer à exposição através de diversas tarefas como a mistura de concreto, corte,
furação, jateamento, demolição e limpeza e quando inalada pode levar a silicose e potencialmente
aumentar o risco ao desenvolvimento de câncer de pulmão e de outras doenças autoimunes18.
A sílica é um mineral duro e dos mais abundantes na crosta terrestre, sendo encontrado em
rochas e areias, muito utilizado como produto final, subproduto ou matéria prima em vários
processos industriais, como na indústria do cimento (argila, areia, pedras e terra diatomácea), de
construção civil, fundição e indústria de mineral não metálico (cerâmicos vidros e fundições).
Segundo a OMS (Organização Mundial da Saúde) a sílica cristalina está classificada como Grupo I,
reconhecidamente cancerígena para seres humanos. A exposição ocupacional a esta substância
ocorre por meio de inalação de poeira, podendo ocasionar a silicose, que é uma fibrose pulmonar
difusa, nodular, intersticial, causada por uma reação dos tecidos à inalação do pó, aumentando o
risco ao desenvolvimento de câncer pulmonar e de outras doenças autoimunes. A sílica é capaz de

4
ativar processos inflamatórios que podem resultar em hiperplasia epitelial associada a
neoplasias19.
Sellappa et al.20 sugerem que trabalhadores expostos aos constituintes do cimento e
concreto podem ter um risco aumentado para o desenvolvimento de câncer de pulmão, tumores
gastrointestinais e dermatites, além de ocasionar alterações genéticas nas células somáticas e
germinativas.
Outro efeito nocivo à saúde dos trabalhadores da construção é o contato com a poeira
composta por metais, como o cobalto e tungstênio. Estes são responsáveis por doenças
pulmonares, com significativas taxas de morbidade e mortalidade. Os principais sintomas
relatados pelos trabalhadores com doença pulmonar por metais são inespecíficos: tosse seca,
dispneia progressiva aos esforços, cianose, dor no peito, fadiga e perda de peso21.
Outra substância presente no ambiente ocupacional destes trabalhadores e potencialmente
carcinogênica é o amianto, também chamado de asbesto, e está relacionado ao desenvolvimento
de câncer de pulmão e de pleura, além da asbestose, mesotelioma pleural e peritoneal, câncer de
laringe, ovário, colo retal, faringe, estômago e lesões pleurais benignas, além de câncer de
estômago, intestino, esôfago, pâncreas, rins e mesotelioma7-22,24,25,25-26.
A exposição ao amianto no ambiente da construção civil pode ocorrer através de tarefas
específicas como o isolamento, a demolição e renovação da construção, e ainda, está presente na
confecção de caixas d’água, telhas onduladas e planas, tubulações, divisórias, tintas e
revestimentos18,19. A Organização Mundial da Saúde (OMS) e IARC classificam este agente como
potencialmente carcinogênico para os humanos em qualquer estágio de produção, transformação
e utilização. Dentre os casos de câncer ocupacional, 70% representam o câncer de pulmão, onde
de 1% a 2% das mortes são atribuídas ao asbesto27,28. O Brasil é o quinto maior produtor de
asbesto, é autossuficiente e exporta 30% do excedente da produção. Estima-se que a população
brasileira exposta diretamente seja de 500 mil pessoas, sendo 20 mil ligadas à exposição
ocupacional em mineração e produção de cimento-amianto. Este produto já foi proibido em todos
os países membros da União Européia, desde 2005 e em alguns países da América latina, com
exceção do Brasil, que apenas instituiu normas regulamentadoras19.
O amianto se divide em dois grupos: as serpentinas e os anfibólios. As serpentinas se
apresentam em fibras curvas e maleáveis chamadas de crisotila ou “amianto branco”, os anfibólios
representam menos de 5% do amianto consumido no mundo, ambos são carcinogênicos para os
humanos29. A asbestose é uma doença causada pelas fibras de asbestos que se depositam nos

5
alvéolos pulmonares e provocam uma reação inflamatória e em seguida uma fibrose, acarretando
em uma rigidez e reduzindo a capacidade de se realizar trocas gasosas, ocorrendo a perda da
elasticidade pulmonar e respiratória. Menegozzo et al.25 realizaram um estudo a fim de se verificar
as causas de mortalidade entre os trabalhadores expostos ao asbesto. Foram analisados 1247
trabalhadores entre os anos de 1950 e 2005 e observaram que as principais causas de morte
foram: câncer do trato respiratório e peritoneal, pleural, de laringe e estômago, além das doenças
respiratórias, como pneumoconiose e asbestose. Foi observado um maior número de asbestose
entre os trabalhadores com até nove anos de exposição e a incidência de câncer de pulmão, maior
entre aqueles com 20 a 29 anos em contato com a substância. Barbieri et al.30 observaram que
pessoas nunca expostas ao amianto, porém que residem próximas a uma fábrica de cimento,
estavam desenvolvendo mesotelioma maligno. Além disso foi encontrado altos níveis da
substância nos pulmões destas pessoas. Consonni et al.201531 avaliaram o risco de
desenvolvimento de câncer de pulmão em pedreiros considerando os efeitos em conjunto dos
agentes cancerígenos ocupacionais, dentre eles sílica e amianto, e evidenciaram o aumento do
risco de câncer de pulmão em pedreiros, sendo a maior associação pela exposição a sílica cristalina
e em menor grau para o amianto.
Além disso, análises moleculares em trabalhadores expostos ao asbesto revelaram que, os
polimorfismos de deleção no gene GSTT1 e GSTM1 estão relacionados com alterações fibróticas e
maior espessura das placas pleurais respectivamente32. Já os polimorfismos nos genes XRCC1,
XRCC3, XPD e OGG1 estão relacionados com a incidência de mesotelioma maligno em sujeitos
expostos ao asbesto33.
Outro fator de exposição que atinge esses trabalhadores é a exposição à luz solar diária, que
quando tomada as devidas precauções é extremamente útil ao nosso organismo, pois estimula a
produção de vitamina D34,35-36. Além disso, ajuda a prevenir doenças autoimunes, infecções,
doenças cardiovasculares, câncer de mama, próstata e cólon36,37-38. No entanto, quando a
exposição solar ocorre frequentemente e em excesso, acarreta graves danos à saúde, como o
câncer, cujos tipos mais frequentes são os de pele e de lábio37. Outra consequência da exposição
solar são os danos aos olhos, como cataratas, neoplasias da conjuntiva e melanoma ocular35,36-38.
Além disso, em relação ao carcinoma basocelular também é uma consequência causada pela
exposição a luz solar, e representa 75% dos tumores de pele não melanoma. Mais de 80% dos
casos de carcinoma de células basais são localizadas em áreas expostas ao sol, tais como a cabeça

6
e pescoço. Este tipo de câncer é raro em áreas sem exposição aos raios ultravioleta, especialmente
nas regiões genitais e perianais40.
Outro agente nocivo a saúde dos trabalhadores da construção civil é o comprometimento da
qualidade do ar, causado pela emissão de contaminantes ou poluentes atmosféricos, também
pode ser um fator para o desenvolvimento de diversas doenças, entre elas o câncer de pulmão.
Segundo Lim et al.41 em 2010 foram registrados mais de 3 milhões de mortes em consequência
dessa exposição. Os poluentes atmosféricos podem ser categorizados em gases, poluentes
orgânicos resistentes, metais pesados e material particulado (MP)42. Nos Estados Unidos, a
Environmental Protection Agency (EPA) realizou uma caracterização genérica das fontes de
emissão do MP e concluíram que o setor da construção civil representou 13% da origem deste
poluente. Neste setor o MP pode ser constituído por pó de cimento, gesso, cal, argamassa
industrializada, poeira devido às escavações ou circulação de veículos ou vento, amianto e outras
fontes43.
Em um estudo realizado por Cardoso & Araújo44, foram relacionados os principais aspectos
ambientais do canteiro de obras de edificações aos incômodos e poluição gerados em diferentes
fases e atividades da obra, a emissão de MP foi relevante nas fases de serviços preliminares,
infraestrutura, estrutura, revestimentos verticais, pinturas, assentamento de pisos e
pavimentação. Também são fontes de MP os setores de produções dos materiais utilizados no
ramo da construção civil, tais como na extração de agregados ou na moagem de matérias primas e
na produção de cal e cimento.
O material particulado (MP) é uma mistura complexa de componentes como ácidos,
produtos químicos orgânicos, metais e poeiras do solo, que podem ser partículas sólidas e
líquidas, em suspensão na atmosfera45. Suas composições químicas e físicas variam de acordo com
origem, localização, clima e época do ano46,47-48. O MP pode ser emitido tanto de fonte
antropogênica ou biogênica, resultando em uma mistura complexa de partículas extremamente
pequenas e de gotículas líquidas. Quando emitidas diretamente na atmosfera são denominadas de
partículas primárias e quando formadas a partir de reações químicas entre componentes
preexistentes são chamadas de secundárias. As emissões atmosféricas destes compostos têm sido
associadas a efeitos adversos agudos e crônicos na saúde humana49,50.
Uma das categorias do MP são as partículas inaláveis, apresentando diâmetro aerodinâmico
menor ou igual a 10µm e são classificadas em partículas grossas (10-2.5 µm), finas (≤ 2.5µm) e
ultrafinas (≤ 0.1µm). O tamanho é um fator importante, o qual influência a forma como as

7
partículas vão se depositar no trato respiratório: MP10 depositam-se principalmente no trato
respiratório superior, enquanto partículas finas e ultrafinas são capazes de atingir os alvéolos
pulmonares. Estudos epidemiológicos demonstram forte associação entre exposição ao MP fino e
ultrafino com a morbidade e mortalidade de doenças cardiorrespiratórias50,51.
Recentemente, a IARC classificou o MP presente na poluição do ar como carcinogênico para
humanos (Grupo 1). Esta determinação foi baseada em evidências sobre a relação do MP2.5 e MP10
ao risco de câncer de pulmão52.
Diversos mecanismos são propostos para explicar a ligação entre MP e o risco de câncer de
pulmão, incluindo a alteração na expressão de genes, respostas inflamatórias e o estresse
oxidativo, no entanto esta relação permanece mal compreendida44-53,54,51,55-56. O MP pode levar a
inflamação e induzir a liberação de espécies reativas de oxigênio, tanto local como
sistemicamente, o que por sua vez, pode contribuir para alterações epigenéticas e instabilidade
genômica51-52,53,57,58-59.
1.3 Epigenética
O genoma humano possui cerca de 25.000 genes e a expressão de cada gene é fortemente
regulada para controlar a função de cada célula. No núcleo, o modelo para informação genética é
codificado em segmentos de DNA, os quais são transcritos em moléculas de RNA. Estas moléculas
são transportadas a partir do núcleo para o citoplasma, onde eles são traduzidos em proteínas. A
atividade dos genes é controlada ao nível do DNA, RNA, e proteína. No nível do DNA, a atividade
do gene pode ser estruturalmente regulada, com histonas para controlar tanto a acessibilidade
física dos fatores de transcrição a um segmento de DNA, quanto para a disponibilidade de fatores
de transcrição. (ARTIGO SYLVIA K. SHENOUDA & SURESH K. ALAHARI)
Aparentemente, há muitas maneiras pelas quais a função do gene pode ser alterada, o que
reflete a relativa facilidade com que a função do gene pode ser perdida. Em contraste, a ativação
de um proto-oncogene pode exigir alterações mais específicas que conferem novas propriedades
ao produto do gene. Além disso, a conversão de 5-metilcitosina à timina em sequências CpG,
podem ser utilizado para estimar a taxa espontânea de alterações de bases induzidas. A metilação
de resíduos citosina em regiões CpG, é um processo endógeno, uma vez que nenhum fator
exógeno foi encontrado ainda que altere a frequência de desaminação de citosinas metiladas em
CpG ou a eficácia de reparação de mutações. Assim, é razoável supor que uma diminuição da

8
proporção de CpG seria indicativo de um efeito direto sobre o DNA exercido por um agente
químico ou físico.
A epigenética refere-se ao estudo de mecanismos que alteram a expressão dos genes, sem
qualquer alteração na sequência primária do DNA60, esses mecanismos epigenéticos são herdáveis
e reversíveis.
Os mecanismos epigenéticos são essenciais para o desenvolvimento normal e manutenção
dos padrões de expressão gênica específicos para os tecidos em mamíferos. A alteração de
processos epigenéticos podem desencadear a função alterada dos genes levando a transformação
celular. As mudanças globais no panorama epigenético são uma marca registrada do câncer,
envolvidas na iniciação e progressão da doença, tradicionalmente, visto como uma doença
genética, tornou-se uma realidade o envolvimento de anormalidades epigenéticas juntamente
com alterações genéticas61. Os principais fenômenos classificados como alterações epigenéticas
são: a metilação do DNA (metilação sobre as bases de citosina do DNA, em dinucleotídeos CG,
comumente referido como CpG), modificações de histonas (modificações pós-traducionais que
alteram a sua interação com proteínas nucleares, tais como acetilação e metilação), RNA não-
codificadores e (miRNA, siRNA)60(Figura1).
Figura 1 - Esquerda: Representação esquemática de modificações epigenéticas. no DNA e nas histonas. Direita: Esquema representando os microRNAs, que são pequenos RNAs não-codificantes, que reconhece um RNAm alvo por homologia de sequência, levando a clivagem e o bloqueio da tradução. (Adaptado de Mansuy et al., 2011
62).

9
As modificações das histonas é uma alteração epigenética frequentemente encontrados nas
células tumorais. As histonas são proteínas conservadas evolutivamente que têm domínios
globulares carboxi-terminal cruciais para a formação do nucleossomo, e as caudas amino-
terminais flexíveis que se projetam a partir da parte central do nucleossomo e conectando os
nucleossomos adjacentes para formarem e ordenarem as estruturas da cromatina63. Cerca de oito
modificações pós-traducionais ocorrem nas histonas incluindo a metilação, acetilação,
fosforilação, ubiquitinação, sumoilação, biotinilação, e ribosilação de ADP, e foram identificados
no núcleo das histonas H2A, H2B, H3, H4, e na família das histonas ligantes H164,65. Estas marcas
nas histonas são essenciais para a organização da cromatina, manutenção da estabilidade do
genoma, silenciamento de sequencias repetitivas do DNA, que regulam a progressão do ciclo
celular, o reconhecimento de sítios de danos e reparação do DNA e manutenção de expressão
adequada de informação genética63.
Outra classe de moléculas que participam dos eventos epigenéticos são os microRNAs. De
acordo com os estudos da última década, os microRNAs emergiram como moléculas-chave na
regulação das funções celulares em mamíferos. Os microRNAs são reguladores fundamentais
relacionados a várias doenças, estas moléculas são sequencias curtas de DNA (aproximadamente
22 nucleotídeos) que regulam a transcrição de genes no nível pós-transcricional66. Desde a
primeira descoberta dos microRNAs em Caenorhabditis elegans Lee et al.38, centenas de
microRNAs foram identificados em eucariotos por influenciarem processos fisiológicos, tais como,
o desenvolvimento, crescimento, diferenciação, reações imunológicas, e a adaptação ao
stress39,66.
Os estudos demonstram que a expressão alterada dos microRNAs pode promover condições
patológicas. O papel dos microRNAs na biologia do câncer tem sido verificada nos últimos anos e
os estudos demonstram sua importância no controle da expressão de RNAm alvo que promovem
ou facilitam o crescimento do tumor, invasão, angiogênese67. Além disso, o perfil de expressão dos
microRNAs nos tumores tem sido relacionados por definirem subtipos, sobrevivência do paciente,
metástases e resposta ao tratamento67,68. Por outro lado, sabe-se que a exposição ambiental,
química, biológica ou agentes físicos podem ser responsáveis por doenças humanas, incluindo o
câncer, o descobrimento das relações entre a exposição aos carcinógenos ambientais e a
expressão de microRNAs pode contribuir para melhor elucidar os mecanismos iniciais da doença e
pode potencialmente conduzir ao desenvolvimento de indicadores úteis de exposição a
substâncias tóxicas ou novos biomarcadores para testes de carcinogenicidade69,70.

10
1.3.1 Metilação do DNA
A metilação do DNA é o mecanismo de regulação epigenética mais estudado e é crucial para
o desenvolvimento normal e manutenção da homeostase celular71,72. Além disso, desempenha um
importante papel em vários processos biológicos, que incluem a inativação do cromossomo X,
regulação da expressão gênica, imprinting e manutenção da estabilidade cromossômica.
Biologicamente consiste na ligação covalente de um radical metil (CH3) em resíduos de citosina
em dinucleotídeos CpG60 (Figura 2).
Figura 2 – Representação da ligação do grupo CH3 no carbono 5 da citosina (C) gerando a 5-metilcitosina, sendo este processo mediado pelas enzimas DNA metiltransferases (DNMT’s). (Retirado e modificado de Lorincz
73).
Os dinucleotídeos CpG não estão uniformemente distribuídas em todo o genoma humano,
mas estão concentradas em trechos curtos do DNA ricos em CpG chamados de ilhas CpG e nas
regiões de sequencias repetitivas, como por exemplo, repetições centroméricas e elementos
retronsposons61. Existem cerca de 28 milhões de sítios CpG no genoma e a maioria dos
dinucleotídeos CpG (70 a 80%) são metilados nas células normais74,75-76. As ilhas CpG são grupos de
sítios CpG que variam entre 200 a 3000 pb e que apresentam uma porcentagem maior que 50 %
de CG61,67. Além disso as ilhas de CpG estão preferencialmente localizadas na região 5’ dos genes
ocupando as regiões regulatórias ou promotoras, cerca de 60% das regiões promotoras dos genes
humanos contem em sua estrutura ilhas CpG55,68. Usualmente nas células normais, as ilhas CpG
não metiladas na região promotora está associada com a expressão dos genes durante a
diferenciação. Por outro lado, ilhas CpG metiladas nas regiões promotoras estão diretamente
ligadas a repressão da expressão dos genes61,77. Além disso, no genoma margeando as ilhas CpG
existem as shores, regiões de baixa densidade em CpG que estão localizadas aproximadamente a 2
Kb das ilhas CpG78. Ainda podemos salientar que além das shores e ilhas CpG são encontradas as
shelves, regiões de 2 Kb que flanqueiam as shores. Perfis de metilação diferencial encontrados nas
shores em vários tipos de câncer tem sido associado com alterações na expressão gênica76,79.

11
As DNA metiltransferases têm um motivo catalítico altamente conservado que estabelece as
padrões de metilação do DNA em sequências alvo do genoma. A distribuição de dos grupamentos
metil demarca regiões de silenciamento da transcrição genica ou de potencial transcrição, criando
uma assinatura epigenética estável76. Alterações nos padrões de metilação do DNA são bem
estudadas em câncer e uma redução global de metilação e hipermetilação em genes específicos,
tais como, em genes supressores tumorais, tem sido descrito como uma característica marcante
de neoplasia80. Sabe-se que o mecanismo de metilação do DNA também pode alterar padrões de
expressão gênica81,82. A metilação do DNA é mitóticamente estável e pode ser propagada pela
divisão celular. Entretanto, estudos têm demonstrado que a metilação do DNA pode alterar
rapidamente em resposta a fatores ambientais. A hipermetilação em genes supressores tumorais
é amplamente caracterizada por representar um dos primeiros passos envolvidos na
carcinogênese humana83. Cada alteração transitória na metilação do DNA pode refletir em uma
condição de estresse na célula, associado com alterações no processo de apoptose, no controle do
ciclo e na proliferação celular. Isto pode levar ao acúmulo de danos genéticos e epigenéticos
persistentes, depois de repetidas exposições a possíveis agentes carcinogênicos82.
Além da análise de genes pontuais, a avaliação dos padrões de metilação do genoma como
um todo, são realizadas em sequências repetitivas de DNA, como os elementos retrotransposons
Alu repeats e LINE-1 (Long Interspersed Nuclear Element-1), são de grande interesse82,84.
Anteriormente, considerado como sequencias sem função no genoma, elementos repetitivos têm
se tornado cada vez mais reconhecido como uma importante classe de reguladores da expressão
gênica72,85-86. Além disso, a hipometilação global em elementos repetitivos pode levar a rearranjos
cromossômicos e ativação de oncogênese87,76. Por serem abundantes no genoma humano, em
torno de 55% consiste destes elementos, o padrão de metilação nestas sequências é considerado
um possível preditor da metilação global84,88. Adicionalmente, sabe-se que a hipometilação destes
elementos aumenta sua atividade retrotranspositiva, o que pode acarretar alterações genômicas,
tais como, mutagênese insercional, interferência na transcrição, e a instabilidade genômica com
consequente variação da regulação transcricional72-75,90,78,84-80. Além disso, a ativação de
sequencias repetitivas de DNA por meio da hipometilação também pode contribuir para
translocações destas sequencias hipometiladas, levando afrouxamento da cromatina91. Elementos
repetitivos compreendem quase a metade de todo o genoma e são um dos principais
contribuintes na hipometilação relacionada ao câncer. A associação entre a hipometilação de

12
elementos repetitivos e exposição à vários fatores ambientais tem sido claramente demonstrada
tanto na população geral quanto em indivíduos expostos ocupacionalmente51,92-93.
1.4 Metilação do DNA para determinação dos sujeitos com alto risco ao desenvolvimento de
câncer
A exposição de trabalhadores a determinados ambientes ocupacionais pode estar associado
ao aparecimento de vários tipos de doenças, como o câncer. Alguns agentes presentes nos
ambientes ocupacionais são classificados como carcinogênicos com potencial bem reconhecido,
no entanto, os mecanismos moleculares subjacentes às suas associações com os riscos de câncer
permanecem pouco compreendidos. Dessa forma a utilização de biomarcadores genéticos e
epigenéticos podem elucidar mecanismos envolvidos com a iniciação da carcinogênese, em
populações expostas.
Um estudo realizado em trabalhadores com elevada exposição aos hidrocarbonetos
policiclicos aromáticos (HPAs), revelou uma hipometilação na região promotora do gene p53, que
pode estar envolvido com mecanismos adicionais associados a instabilidade genética induzidos
por HPAs, e pode mediar o risco de câncer de pulmão93. Em trabalhadores de fundição, com alta
exposição ao material particulado (MP), foram comparados os padrões de metilação nos
leucócitos do sangue dos genes supressores tumorais APC, p16, p53 e RASSF1A, no primeiro dia e
no último dia da semana de trabalho, período de curta exposição ao MP e foi observado
hipermetilação dos genes p16 e APC e hipometilação de RASSF1A e p5382. O padrão alterado de
metilação do DNA em genes supressores tumorais, como o APC, p16, p53 e RASSF1A é geralmente
observada em DNA de leucócitos do sangue de pacientes com câncer de pulmão73-94,95-96.
Umemura et al.97 demonstraram hipermetilação em genes supressores de tumor no DNA do soro
de pacientes com silicose e câncer de pulmão. Este estudo sugere que a aberrante metilação em
genes supressores de tumor pode facilitar a detecção precoce do câncer de pulmão em pacientes
com silicose. O p16 foi o primeiro gene identificado como hipermetilado, nos cânceres de
pulmão96. Estudos tem demonstrado que a expressão de p16 é inativado pela metilação em uma
prevalência de 60% a 70% em câncer de pulmão primário e com baixa frequência de mutações28,98.
Estudos em animais demonstraram a hipermetilação global do DNA em esperma de camundongos
após a exposição à poluição do ar, bem como a hipermetilação no p16 em pulmões de ratos, após
a exposição ambiental ao MP2.5 concentrado56,99.

13
Um estudo de meta-análise avaliou o papel da metilação dos genes em pacientes com
câncer de pulmão de não pequenas células e a relação com o hábito de fumar. Foi verificada uma
associação significativa no perfil de metilação entre os genes CDKN2A, RASSF1, MGMT, RARB,
DAPK, WIF1 e FHIT e o tabagismo em pacientes com câncer de pulmão. A hipermetilação de p16
foi um fator de risco comum ao tabagismo nos pacientes100. Vários investigadores têm
demonstrado que ilhas CpG nas regiões promotoras dos genes MGMT e MLH1 são hipermetiladas
em vários tipos de câncer e a aberrante hipermetilação reprime a expressão de ambos os
genes101,102-103. O gene MLH1 é um dos participantes da via de reparo MMR, essa via é um sistema
essencial pelo qual as células corrigem erros na replicação do DNA para manutenção da fidelidade
do genoma104,105. O MGMT codifica a O6-metilguanina-DNA-metiltransferase enzima de reparo
que desempenha um importante papel na defesa do DNA celular contra efeitos citotóxicos e
carcinogênicos de agentes alquilantes103. Em um estudo realizado na China em trabalhadores da
indústria do plástico, ambiente rico em cloreto de vinila, reconhecido como agente carcinogênico
para humanos, foi detectada a hipermetilação de MGMT em grupo destes trabalhadores que
obtiveram uma aumentada instabilidade cromossômica avaliada por MNs103. Por outro lado Duan
et al.106 verificaram a hipometilação do gene MGMT em um grupo de trabalhadores expostos ao
MP associado a HPAs comparados aos indivíduos não expostos. Outro estudo realizado em uma
população com prolongada exposição ao arsênio foi verificado o aumento da metilação do DNA
em genes supressores tumorais, como, p16 e no gene de reparo MLH1107.
A avaliação dos padrões de metilação entre as sequências repetitivas de DNA, como os
elementos Alu e LINE-1, são utilizadas como biomarcadores epigenéticos73,75. Estudos in vitro e in
vivo tem demonstrado uma hipometilação nestes elementos em resposta à exposição a partículas
de poluição ambiental, sendo a variação na metilação um evento sistêmico consequente da
cascata de reações inflamatórias desencadeada pela inalação de partículas poluentes, esses
resultados demonstram a possibilidade de que essas mudanças sejam responsáveis por respostas
adaptativas e determinem efeitos ambientais na saúde humana92. No trabalho de Duan et al.106 foi
determinado em trabalhadores com alta exposição a hidrocarbonetos policíclicos aromáticos
(HPAs), a hipometilação da sequência repetitiva LINE-1, sendo assim os autores sugeriram a
utilização deste marcador para avaliação da exposição aos HPAs. Demetriou & Vineis91
evidenciaram que a poluição do ar influência nos níveis de metilação globais, e essas mudanças
epigenéticas podem contribuir para a carcinogênese tanto quanto as alterações genéticas.

14
A grande maioria dos estudos envolvendo marcadores de metilação do DNA em populações
expostas ocupacionalmente são realizados em leucócitos do sangue periférico, devido a estas
células serem as primeiros a responderem aos mediadores inflamatórios liberados nos pulmões
após a exposição ambiental108. A metilação do DNA regula as funções dos leucócitos em nível
transcricional e, portanto, afeta a migração e o recrutamento dos leucócitos da corrente
sanguínea para o pulmão109. A metilação do DNA no sangue periférico tem sido apontada como
um marcador sensível para mudanças agudas e de curto prazo no epigenoma108.
Sendo assim, a utilização da análise de metilação do DNA como um possível biomarcador,
com intuito de predizer a incidência de risco de câncer em populações expostas como os
trabalhadores da construção civil, pode melhorar a avaliação da exposição mas também aumentar
a compreensão dos mecanismos subjacentes a esta tipo de exposição que esta classe de
trabalhadores estão submetidos, podendo elucidar o risco de câncer.

15
2 JUSTIFICATIVA
O universo de empresas na indústria da construção civil, abrange em torno de 11,9 mil
empresas ativas que empregaram cerca de 3 milhões de pessoas. Somente na região Sudeste o
número total de empresas da construção civil é de 50.008, com 1.611.861 pessoas trabalhando no
setor 110.
Diante da representatividade do setor da construção civil no cenário da economia brasileira,
responsável por considerável parte no PIB em nosso país e sua relação social, tendo em vista o
grande número de empregos gerados, é essencial pensar na qualidade de vida e no impacto a
saúde dos trabalhadores deste setor. Haja vista, que milhões de trabalhadores estão expostos
ocupacionalmente a diversos fatores e agentes nocivos à saúde. A busca de possíveis
biomarcadores para avaliação de danos citotóxicos é uma importante ferramenta para a
determinação de populações de risco para o possível desenvolvimento de câncer. Além disso, o
aparecimento do câncer é provocado por mutações no DNA e também por mudanças
epigenéticas. Sendo assim, os estudos de metilação do DNA em regiões promotoras de genes
alvos podem contribuir na elucidação dos mecanismos moleculares importantes para o processo
de carcinogênese e apontar potenciais biomarcadores.

16
3 OBJETIVOS
3.1 Objetivo geral
Avaliação de possíveis biomarcadores epigenéticos para associação do potencial risco de
desenvolvimento de câncer em trabalhadores da construção civil expostos a agentes
carcinogênicos ambientais e ocupacionais.
3.2 Objetivos específicos
Avaliar o perfil de metilação do DNA nos leucócitos do sangue dos trabalhadores da
construção civil em comparação ao grupo controle (não expostos), nas regiões
promotoras dos genes APC, RASSF1A, p16, p53, MGMT, MLH1 e das sequencias
repetitivas ALU e LINE-1, utilizando a metodologia de pirosequenciamento.
Comparar o perfil de metilação do DNA dos genes alvos escolhidos nos leucócitos do
sangue no grupo de trabalhadores da construção civil no primeiro dia de trabalho da
semana e no quinto dia de exposição.
Correlacionar os dados do questionário ocupacional, com os níveis de metilação dos
genes supressores tumorais analisados e das sequências repetitivas ALU e LINE-1.

17
4 MATERIAL E MÉTODOS
4.1 Local de estudo
O estudo foi realizado no Hospital de Câncer de Barretos, localizado na cidade de Barretos,
estado de São Paulo.
4.2 Desenho do estudo
O estudo é observacional prospectivo.
O estudo foi dividido em dois grupos, sendo o primeiro grupo de 58 trabalhadores da
construção civil (caso) e 50 trabalhadores do setor administrativo (controle), sendo a primeira
etapa do estudo uma comparação caso x controle.
No segundo grupo foram analisados 39 trabalhadores da construção civil, que foram
coletados em dois dias diferentes da semana, a coleta foi realizada do mesmo indivíduo sendo a
primeira no 1° dia de trabalho da semana (segunda-feira) e a segunda coleta no 5° dia de trabalho
(sexta-feira), a fim de detectar as potenciais diferenças no nível de metilação do DNA dos genes
propostos em curto período de tempo de exposição ocupacional no ambiente de trabalho da
construção civil. Além disso, as análises com este grupo proporcionam a verificação do nível de
metilação do DNA dos indivíduos trabalhando no mesmo local de trabalho sem interferentes de
outros participantes que não tem a exposição e com características socioeconômicas diferentes.
4.3 Sujeitos da pesquisa
Os sujeitos da pesquisa foram os trabalhadores da construção civil do Hospital de Câncer de
Barretos (grupo estudo) e trabalhadores do setor administrativo desta mesma instituição (grupo
controle) pareados por idade. O tamanho amostral foi calculado para homens não fumantes com
nível de 5% de significância e 10% de precisão. O número do cálculo amostral foi calculado pelo
NAP (Núcleo de apoio à pesquisa do Hospital de Câncer de Barretos), considerando os critérios de
inclusão e exclusão descritos abaixo.
4.4 Critérios de inclusão
Foram incluídas no estudo homens com mais de 18 anos, trabalhadores da construção civil
do Hospital de Câncer de Barretos e ex-fumantes com pelo menos um ano de término do hábito.

18
4.5 Critérios de exclusão
Foram excluídos todos os indivíduos fumantes, usuários de drogas, trabalhadores com
doenças infecciosas ou doenças crônicas (como as doenças autoimunes), expostos a outros
agentes, como raios X e xilol. E para análise do segundo grupo, também, foram excluídos do
estudo os trabalhadores que faltaram no trabalho durante os cinco dias de exposição.
4.6 Coleta de dados
Para este estudo cada participante respondeu um questionário que continha questões
referentes às características sócio-demográfico, rotina ocupacional e a saúde geral e bucal, o
questionário pode ser visualizado no anexo (A). No preenchimento do questionário foi
apresentado o Termo de Consentimento Livre e Esclarecido (TCLE) para assinatura, e tanto os
trabalhadores da construção civil quanto os trabalhadores do administrativo foram informados
dos potenciais fatores de risco. O questionário utilizado foi adaptado a partir de Boffeta et al. 1999
e Sartor, 2003.
4.7 Coleta de sangue
Foram coletados 2 tubos de 5mL de amostras biológicas (sangue) para cada indivíduo, em
tubos Vacutainer contendo EDTA, que posteriormente foram submetidos a extração do DNA
genômico.
4.8 Determinação da metilação do DNA na região promotora de genes
4.8.1 Extração de DNA de sangue periférico
A partir da amostra coletada, o tubo Vacutainer foi centrifugado a 3500 rpm por 10 minutos
à 4°C. O sobrenadante contendo o anel de leucócitos, com aproximadamente 500uL, foi retirado e
transferido para um tubo falcon de 15 ml. Foi adicionado 12 ml de solução de lise composta por
bicarbonato de amônio (1X) e cloreto de amônio (10X) que posteriormente foram imersos em gelo
por 20 minutos. Após esse período, os tubos foram homogeneizados e centrifugados por 10
minutos a 3500 rpm a 4°C e o pellet obtido foi ressuspendido em 20uL de PBS 1X e transferido
para tubos de 1,5 mL. Os próximos passos foram seguidos de acordo com o DNeasy Blood&Tissue
Kit (Qiagen), conforme instruções do fabricante e a amostra final foi eluída em 60uL em água

19
estéril deionizada (milliQ). O DNA extraído foi quantificado por espectrofotometria em NanoDrop
2000C (ND-1000 Spectrophotometer v.3.0.1, Termo Scientific). Este aparelho quantifica ácidos
nucleicos e proteína através da leitura do comprimento de onda absorvido. A absorbância de
260nm indica o pico de absorbância de UV do DNA, a absorbância de 280nm o pico de absorção de
UV de proteínas e 230nm o pico de absorção de UV para contaminantes orgânicos.
Posteriormente, calculando as razões 260/280 e 260/230 é possível obter a pureza do DNA a ser
analisado. As amostras foram quantificadas utilizando 1uL de DNA e estavam dentro dos valores
de razões aceitáveis para trabalhar com o DNA, entre 1,6 e 2,3. Os DNA’s foram estocados em
freezer (-80°C).
4.8.2 Conversão de Bissulfito
O DNA genômico foi submetido ao tratamento de conversão por bissulfito de sódio de
acordo como descrito no kit Epitech Bisulfite kit (Qiagen) conforme instruções do fabricante. O
tratamento consiste em converter as citosinas não metiladas em uracilas através da perda do
grupo amina na base citosina. Enquanto que as citosinas metiladas permanecem protegidas e
inalteradas.
O método é composto pelas seguintes etapas: desnaturação, sulfonação, deaminação,
purificação, dessulfonação e eluição das amostras. Foram utilizados 1ug de DNA, obtendo um
volume final de 20 uL, estes foram colocados em microtubos de 0,2uL, em seguida adicionado
85uL de bissulfito mix e 35uL de “DNA Protect” Epitech Bisulfite kit (Qiagen) formulado para evitar
a fragmentação do DNA geralmente associada ao tratamento com bissulfito a altas temperaturas e
de baixo valores de pH. Ele também fornece a desnaturação efetiva de DNA, resultando na cadeia
simples do mesmo, necessário para a conversão completa de citosinas. Além disso, o DNA Protect
Buffer contém um corante indicador de pH, tal como uma mistura de controle para permitir
confirmação do pH correto para conversão de citosinas. Após a adição do bissulfito mix e DNA
Protect, as amostras foram homogeneizadas e incubadas em termociclador. O programa de ciclo
térmico fornece uma série de passos de incubação necessários, otimizados para a desnaturação
térmica do DNA e subsequente dessulfonação e desaminação de citosinas, permitindo maiores
taxas de conversão das mesmas. Após a incubação, as amostras foram centrifugadas e transferidas
para tubos limpos de 1,5mL e adicionado buffer BL que promove a ligação do DNA convertido na
membrana da coluna de spin-MinElute DNA de cadeia simples. Subsequente a este passo, o DNA é
ligado à membrana que é lavada utilizando o buffer BW que remove eficientemente o bissulfito de

20
sódio residual. Logo após, a dessulfonação ocorre usando Buffer BD, a membrana é novamente
lavada utilizando tampão BW antes da eluição a partir da membrana da coluna utilizando tampão
EB (Figura 3).
Figura 3 – Representação esquemética do protocolo de conversão por bissulfito de sódio. Adaptado de Epitech Bisulfite kit (Qiagen)
111.
4.8.3 PCR e eletroforese em gel de agarose
Após extração de DNA e tratamento com bissulfito de sódio, cada amostra foi submetida à
amplificação pela metodologia de PCR com um dos primers marcados com biotina. As reações de
PCR (Reação em Cadeia da Polimerase) foram realizadas em um volume de 50 uL contendo: 2 uL
do DNA convertido, 25uL de HotStarTaq Master Mix (Qiagen), 1uL de cada primer na concentração
de 10 uM (foward e reverse). As condições de ciclagem foram estabelecidas para cada conjunto de
Reação Bissulfito
DNA com DNA Protect
Ciclo término (5h)
Transferir DNA para o microtubo de 1,5mL
com tampão BL.
Adicionar etanol
Transferir para coluna do Kit
Adicionar Buffer BW
Vincular
Lavagem
Dessulfonação Adicionar Buffer BW
Lavagem (2X) Adicionar Buffer BW
Adicionar etanol
Adicionar Buffer EB
Eluição

21
primers (Anexo B). Cinco uL do produto de PCR foram submetidos a análise por eletroforese
utilizando-se gel de agarose 2%. O marcador molecular de 1Kb (Invitrogen) foi utilizado para
verificação do tamanho do amplicon e a imagem foi visualizada no Imagem Quant LAS 4000 Mini.
Foram utilizados os desenhos dos primers dos genes APC, RASSF1A e TP53 conforme descrito
por Baccarelli et al.112. Para os conjuntos de primers do gene MGMT foram desenhados utilizando-
se o Software PyroMark Assay Design (Qiagen) seguindo os parâmetros recomendados: tamanho
dos primers entre 18 e 21 pares de bases, com mais de 50% de G+C, temperatura de anelamento
55 a 62°C, evitando a formação de harpain e loop. Os genes MLH1, CDKN2A e as sequências
repetitivas ALU e LINE-1 foram utilizados os primers conforme gentilmente cedidos os desenhos
pelo Prof. Dr. Zdenko Herceg do IARC (Anexo C e D).
4.8.4 Pirosequenciamento
Para a preparação das amostras para o pirosequenciamento, os 45uL de produto da PCR
foram transferidos para uma placa de 96 poços e agitados em um mix contendo beads de sefarose
(Strepatavidin Sepharose GE Healthcare) e tampão de ligação (binding buffer – Qiagen) de acordo
com a quantidade de amostra. A seguir a placa foi submetida a agitação por 10 minutos a 1.400
rpm. Logo após, a placa contendo o produto de PCR foi submetida a bomba vácuo (PrepStation).
Na ferramenta PrepStation o produto de PCR é ligado nas beads, ficando preso a mesma.
Assim, foi iniciado as etapas de lavagem, primeiro em 100mL de solução etanol 70%, segundo em
100mL de solução denaturante de NaOH 0,2M (Qiagen) e em seguida lavado com tampão de
lavagem Wash buffer (Qiagen). Após as lavagens, os produtos são transferidos para placa de
pirosequenciamento (PSQ 96 low plate) (Qiagen) contendo primer de sequenciamento na
concentração de 0,4uM diluídos em tampão de anelamento (anneling buffer – Qiagen).
Após a transferência, a placa é colocada em um termo bloco à 80ºC por 5 minutos a fim de
proporcionar o anelamento dos primers. Em seguida, a placa é colocada em temperatura
ambiente por 10 minutos. Por fim, a placa contendo os produtos ligados aos primers de
sequenciamento foram transferidos para o equipamento pirosequenciador PyroMark Q96 ID
System (Qiagen) acompanhado do cartucho contendo as enzimas (ATP sulfurilase, luciferase e
apirase), substratos (adenosina 5fosfosulfato (APS) e luciferina) e dNTPs.
O primeiro dos quatro dessoxinucleotide (dNTPs) é adicionado a reação. A DNA polimerase
catalisa a incorporação da dNTP à fita de DNA. Cada evento de incorporação é acompanhada da
liberação do pirofosfato (PPi) em uma quantidade equimolar à quantidade de nucleotídeos

22
incorporados. A enzima ATP sulfurilase converte o PPi em ATP na presença de APS. Este ATP dirige
a conversão mediada pela luciferase da luciferina em oxiluciferina que gera uma luz visível numa
quantidade proporcional à quantidade de ATP. A luz produzida é detectada por uma câmera CCD
(charge coupled device) que é vista como um pico no pirograma. A Apirase, uma enzima que
degrada nucleotídeos, degrada constantemente os dNTPs não incorporados e o excesso de ATP,
quando a degradação é completada, outro dNTP é adicionado. A adição de dNTPs é realizada uma
de cada vez. À medida que o processo continua, a fita de DNA complementar é formada e a
sequência é determinada a partir dos picos do pirograma (Figura 4).
A metodologia de pirosequenciamento, é um procedimento qualitativo quanto quantitativo.
É possível identificar a quantidade exata de metilação de dinucleotídeos CpGs específicos de
maneira rápida e eficaz. É possível verificar a qualidade da conversão por bissulfito de sódio, uma
vez que podem ser inseridos na ordem de dispensação de nucleotídeos controles que podem
indicar a ocorrência de conversão parcial do DNA conforme exemplificado na Figura 5.
Figura 4 - Representação esquemática do sistema enzimático de pirosequenciamento. Cada dNTP é incorporada na cadeia de DNA pela enzima polimerase, a qual adiciona as bases de acordo com a fita molde, liberando assim a molécula de pirofosfato (PPi). A enzima ATP sulfurilase converte o PPi em ATP que serve como substrato para a produção de luz pela enzima luciferase. A luz produzida é detectada como evidência de que a incorporação de nucleotídeos foi eficiente (Retirado e adaptado de Ahmadian et al.
113).
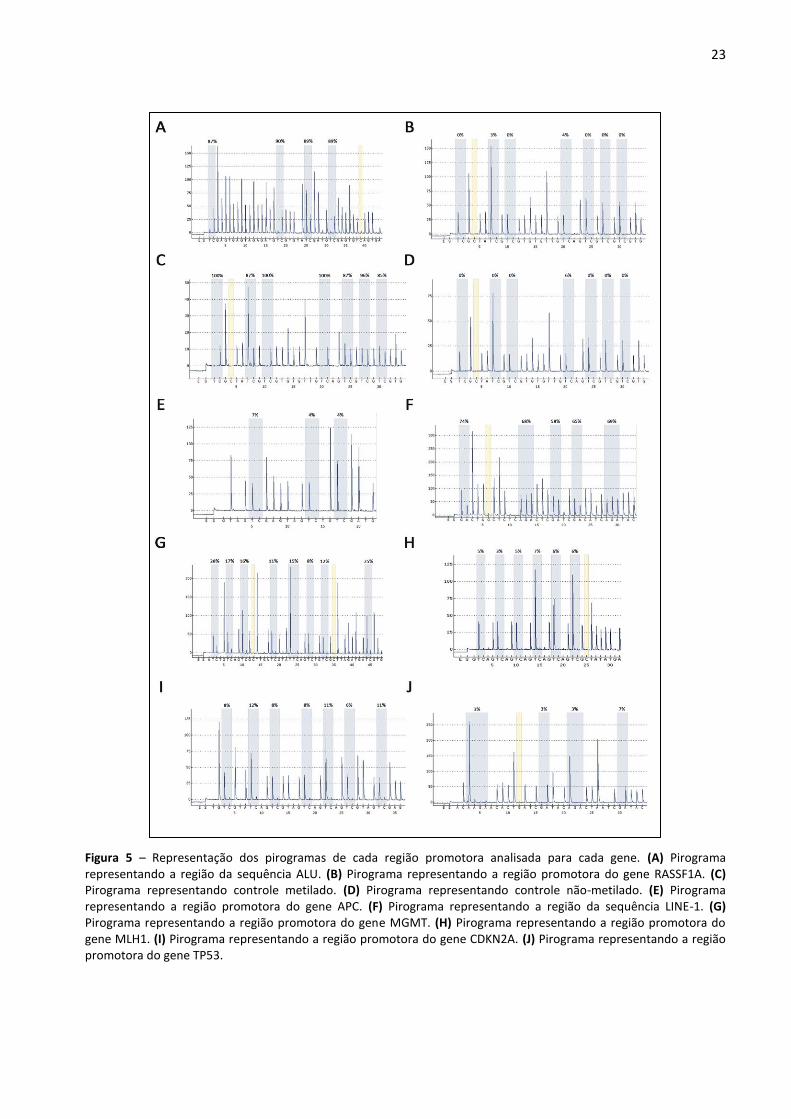
23
Figura 5 – Representação dos pirogramas de cada região promotora analisada para cada gene. (A) Pirograma representando a região da sequência ALU. (B) Pirograma representando a região promotora do gene RASSF1A. (C) Pirograma representando controle metilado. (D) Pirograma representando controle não-metilado. (E) Pirograma representando a região promotora do gene APC. (F) Pirograma representando a região da sequência LINE-1. (G) Pirograma representando a região promotora do gene MGMT. (H) Pirograma representando a região promotora do gene MLH1. (I) Pirograma representando a região promotora do gene CDKN2A. (J) Pirograma representando a região promotora do gene TP53.

24
4.8.5 DNA controle metilado e não-metilado
Para as análises realizadas no pirosequenciador foram utilizados DNA controles obtidos da (Qiagen).
O controle utilizado de DNA metilado passa pelo processo de metilação in vitro e após este passo por
conversão de bissulfito. Também é utilizado um DNA não-metilado. A fonte do DNA são de sangue humano
constituído por um pool de vários doadores anônimos, onde todo o genoma é representado. O DNA foi
preparado a partir de células do sangue periférico (baixo grau de metilação). O sangue foi coletado de
doadores diferentes, tanto homens como mulheres, para evitar possíveis mutações de apenas um doador.
O DNA não metilado é obtida por amplificação de todo o genoma. Os controles utilizados possuem uma
faixa de porcentagem de metilação do DNA aceitável, o qual no controle metilado é acima de 85% e no
não-metilado igual ou abaixo de 5%, quando submetidos no pirosequenciador.
4.9 Análise estatística dos dados
Para as análises dos dados foi utilizado o software SPSS para Windows versão 21.0.
Inicialmente, os dados foram tabulados considerando as estatísticas descritivas (média, desvio
padrão, mediana, mínimo, máximo) para os dados quantitativos e tabelas de frequência para os
qualitativos.
Basicamente existem dois tipos básicos de distribuição dos dados: Normal ou Anormal,
também conhecida como distribuição livre. A distribuição normal ou Gaussiana (relativo à Carl
Gauss) apresenta uma forma semelhante a uma curva em sino quando os dados contínuos estão
dispostos em uma curva de distribuição. Desta forma, podemos visualizar os dados que se
concentram em torno de uma média e se dispersam simetricamente a partir desse ponto central.
Muitos testes estatísticos, como o teste t de Student, requerem uma distribuição normal. Quando
a curva de distribuição dos dados não apresenta uma forma de sino é chamada de assimétrica,
anormal ou de livre distribuição. Os testes usados para dados com distribuição anormal são
conhecidos como estatísticas não-paramétricas (exemplo: teste de Mann-Whitney). Assim, torna-
se imprescindível, antes da escolha do teste estatístico para dados contínuos, examinar a
distribuição dos dados que neste trabalho foi realizada através do teste Shapiro-Will e
Kolmogorov-Smirnov. Se dados contínuos apresentam uma distribuição anormal, é necessário
escolher um teste não-paramétrico ou transformar os dados em uma distribuição normal,
aplicando, como exemplo, uma transformação logarítmica114.
Contudo, antes de realizar o teste estatístico para comparar a associação entre a metilação e
o desfecho (Caso ou Controle) realizamos um teste para verificação da normalidade dos dados e

25
baseado nesses resultados optamos pelo Teste T (paramétrico) ou Teste Man-Whitney (não
paramétrico). Com isso, para as análises das diferenças entre os grupos considerando as
características demográficas foram utilizados os testes não paramétrico Qui-quadrado ou Exato de
Fisher, optando pelo Exato de Fisher apenas quando os pressupostos do teste de Qui-quadrado
não foram contemplados.
Para a análise do perfil de metilação dos trabalhadores da construção civil e do grupo
controle, foram realizadas as análises da distribuição dos grupos se paramétrico ou não
paramétrico, assim foram utilizados para as análises os testes estatísticos Teste-T ou Mann-
Whitney, respectivamente. Para as análises de metilação do DNA foram calculados as médias e/ou
mediana geral de cada gene para cada grupo e também foi calculado as diferenças para cada sítio
de cada gene. Para as análises de correlação entre o perfil de metilação do DNA dos genes que
apresentaram significância, o grupo de trabalhadores da construção civil foi dividido pela média,
de acordo com o hábito de bebida e a informação se fumantes passivo. A partir desta divisão foi
aplicado o Teste-T e/ou Mann-Whitney para avaliação estatística.
Para avaliar a metilação do DNA nos genes dos trabalhadores da construção civil (caso) no 1º
e 5º dia de exposição foi utilizado o Teste T pareado (paramétrico) ou Teste de Wilcoxon (não
pareado), após a verificação da normalidade através do teste Shapiro-Wilk. Em todas as análises
foi considerado um nível de significância de 5%, p valor <0,05.
Nas análises realizadas para verificação da presença ou ausência de motifs ligantes para
fatores de transcrição foi utilizado o banco de dados JASPAR e as análises foram realizadas em
ambiente R utilizando-se o pacote R/Bioconductor conforme descrito por Mathelier et al.115
4.10 Aspectos Éticos
Este estudo foi submetido ao Comitê de Ética em Pesquisa do Hospital de Câncer de
Barretos e encontra-se aprovado (586/2012). Os pesquisadores garantem o sigilo de todos os
participantes da pesquisa, não divulgando o nome ou qualquer outra informação que possa
identificar os envolvidos no estudo.

26
5 RESULTADOS
5.1 Coleta e caracterização sócio-demográficas dos grupos
A população do estudo foi composta por indivíduos do sexo masculino e não tabagista. No
primeiro grupo analisado foram incluídos 58 indivíduos no grupo dos trabalhadores da construção
civil (caso) e 50 trabalhadores do setor administrativo (controle), totalizando 108 indivíduos. No
segundo grupo foram analisados 39 indivíduos da construção civil no primeiro dia de trabalho da
semana e no quinto dia para avaliação do perfil de metilação dos genes propostos em curto
período de tempo de exposição no ambiente da construção civil.
Os participantes da pesquisa de ambos os grupos responderam a um questionário referente
as características sócio-demográficas, dados ocupacionais, tais como, histórico ocupacional, tempo
de duração no emprego atual e anteriores e aos hábitos de tabagismo e consumo de bebida
alcoólica. O grupo da construção civil respondeu a um questionário contendo perguntas
referentes a algumas substâncias possivelmente presentes no ambiente da construção e que de
acordo com a literatura podem contribuir com efeitos genotóxicos e na carcinogênese. Além disso,
foram entrevistados também em relação ao uso de EPI (equipamento de proteção individual),
tempo de exposição (em anos trabalhados) e quantidade (em horas) de exposição aos materiais e
substâncias conforme apresentados no (anexo D). O grupo de funcionários do setor administrativo
(grupo controle) também responderam ao mesmo questionário. No entanto, dados referentes à
exposição não foram aplicados, pois na função atual eles não tem nenhum contato com poeira,
fumaça ou substâncias tóxicas que façam parte do ambiente ocupacional, como estabelecidos nos
critérios de inclusão e exclusão do estudo.
Os trabalhadores da construção civil e do setor administrativo, pertencentes ao primeiro
grupo analisado, apresentaram médias de idade semelhantes (caso: 39 ± 13 anos; controle: 32 ±
10 anos). A média de duração de exposição ocupacional no grupo caso foi de 10 anos, enquanto
que no grupo controle não foi reportado histórico de exposição para os carcinógenos relacionados
com a construção civil ou qualquer agente físico ou químico no local de trabalho. A maioria dos
participantes, sendo 61% dos casos e 83% dos controles, consumiam bebidas alcoólicas. Além
disso, 62,7% dos casos e 31,1 % dos controles foram expostos à fumaça do cigarro como fumantes
passivos (Tabela 1).
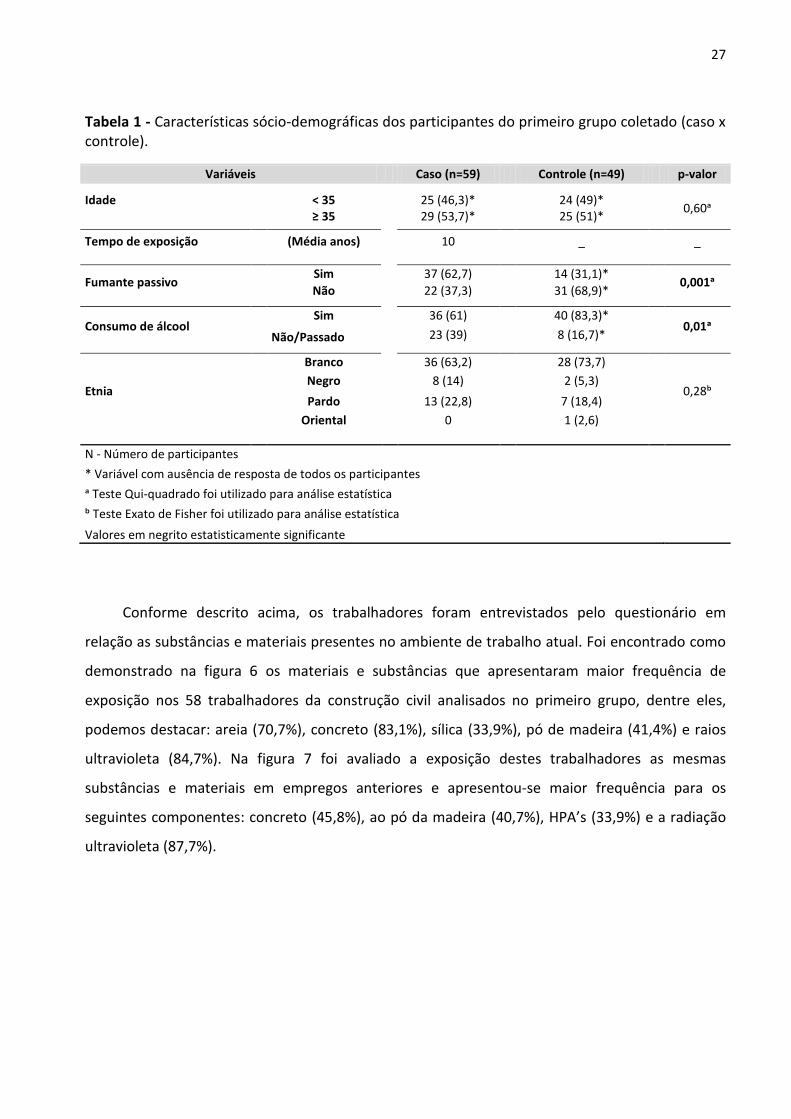
27
Tabela 1 - Características sócio-demográficas dos participantes do primeiro grupo coletado (caso x controle).
Variáveis Caso (n=59) Controle (n=49) p-valor
Idade
< 35
25 (46,3)*
24 (49)*
0,60ᵃ
≥ 35
29 (53,7)*
25 (51)*
Tempo de exposição (Média anos) 10 _ _
Fumante passivo
Sim
37 (62,7) 14 (31,1)* 0,001ᵃ
Não
22 (37,3)
31 (68,9)*
Consumo de álcool Sim
36 (61) 40 (83,3)*
0,01ᵃ
Não/Passado
23 (39)
8 (16,7)*
Etnia
Branco
36 (63,2) 28 (73,7)
0,28ᵇ
Negro
8 (14)
2 (5,3)
Pardo
13 (22,8)
7 (18,4)
Oriental
0
1 (2,6)
N - Número de participantes * Variável com ausência de resposta de todos os participantes ᵃ Teste Qui-quadrado foi utilizado para análise estatística ᵇ Teste Exato de Fisher foi utilizado para análise estatística Valores em negrito estatisticamente significante
Conforme descrito acima, os trabalhadores foram entrevistados pelo questionário em
relação as substâncias e materiais presentes no ambiente de trabalho atual. Foi encontrado como
demonstrado na figura 6 os materiais e substâncias que apresentaram maior frequência de
exposição nos 58 trabalhadores da construção civil analisados no primeiro grupo, dentre eles,
podemos destacar: areia (70,7%), concreto (83,1%), sílica (33,9%), pó de madeira (41,4%) e raios
ultravioleta (84,7%). Na figura 7 foi avaliado a exposição destes trabalhadores as mesmas
substâncias e materiais em empregos anteriores e apresentou-se maior frequência para os
seguintes componentes: concreto (45,8%), ao pó da madeira (40,7%), HPA’s (33,9%) e a radiação
ultravioleta (87,7%).
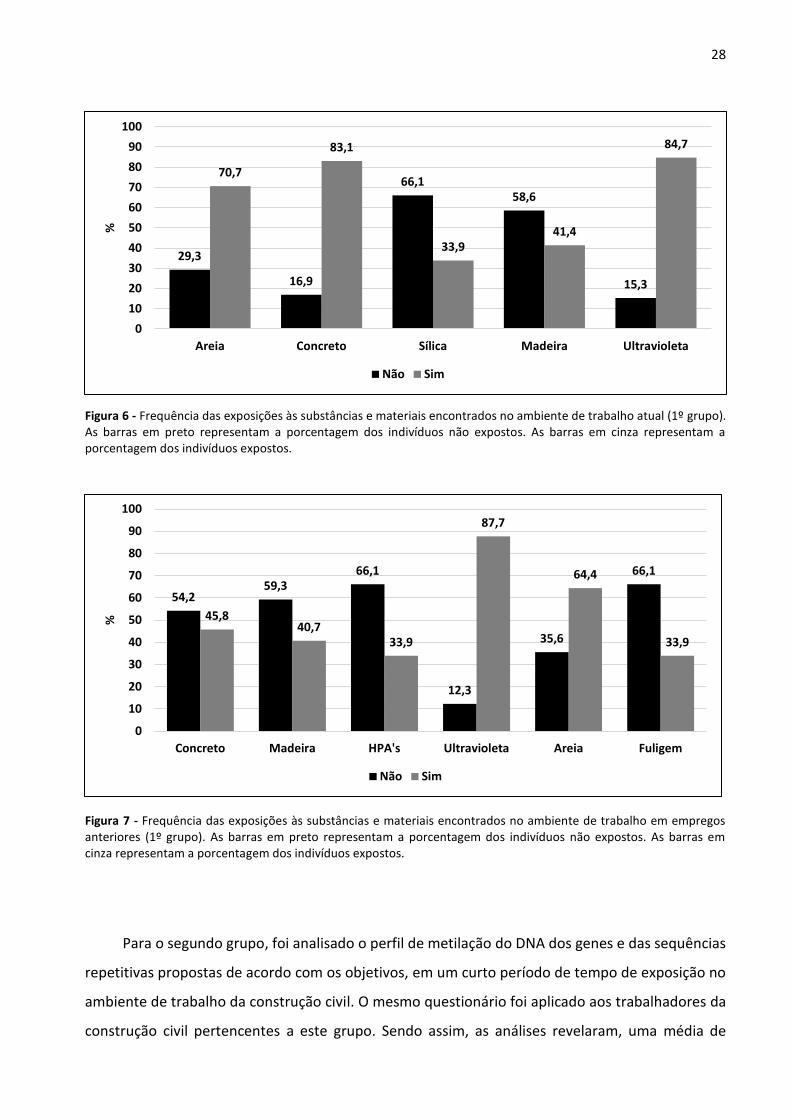
28
Figura 6 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho atual (1º grupo). As barras em preto representam a porcentagem dos indivíduos não expostos. As barras em cinza representam a porcentagem dos indivíduos expostos.
Figura 7 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho em empregos anteriores (1º grupo). As barras em preto representam a porcentagem dos indivíduos não expostos. As barras em cinza representam a porcentagem dos indivíduos expostos.
Para o segundo grupo, foi analisado o perfil de metilação do DNA dos genes e das sequências
repetitivas propostas de acordo com os objetivos, em um curto período de tempo de exposição no
ambiente de trabalho da construção civil. O mesmo questionário foi aplicado aos trabalhadores da
construção civil pertencentes a este grupo. Sendo assim, as análises revelaram, uma média de
54,2 59,3
66,1
12,3
35,6
66,1
45,8 40,7
33,9
87,7
64,4
33,9
0
10
20
30
40
50
60
70
80
90
100
Concreto Madeira HPA's Ultravioleta Areia Fuligem
%
Não Sim
29,3
16,9
66,1 58,6
15,3
70,7
83,1
33,9 41,4
84,7
0
10
20
30
40
50
60
70
80
90
100
Areia Concreto Sílica Madeira Ultravioleta
%
Não Sim
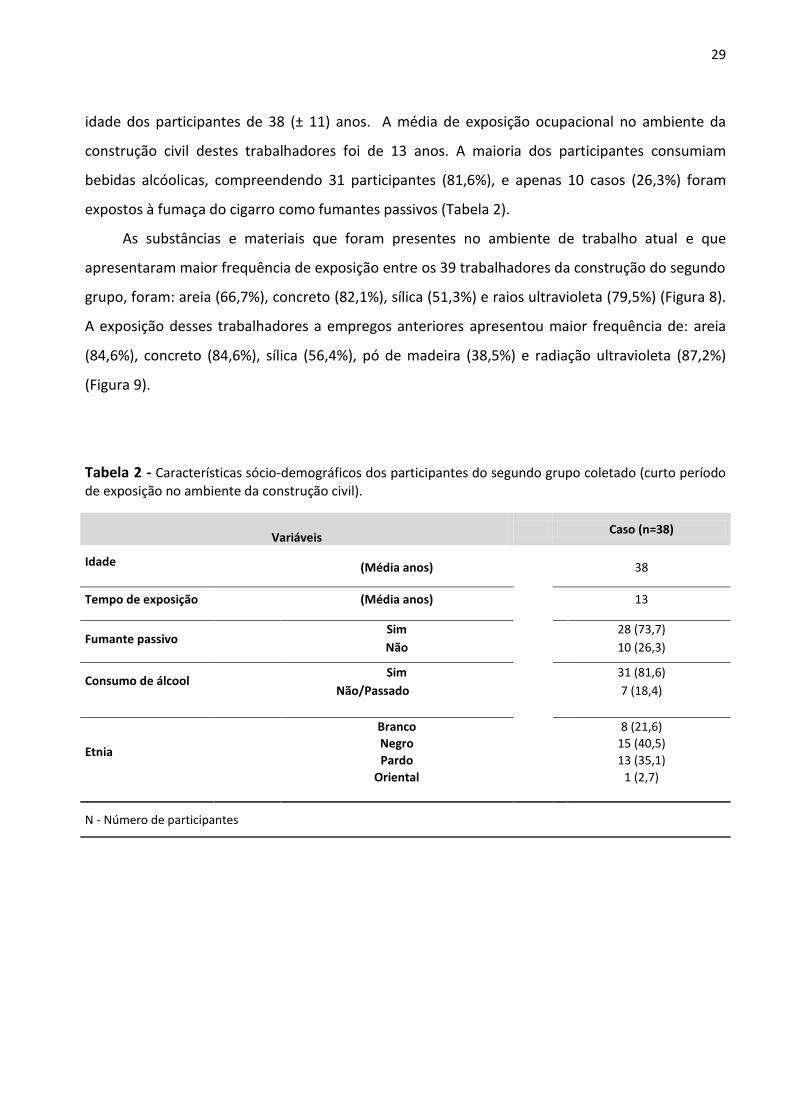
29
idade dos participantes de 38 (± 11) anos. A média de exposição ocupacional no ambiente da
construção civil destes trabalhadores foi de 13 anos. A maioria dos participantes consumiam
bebidas alcóolicas, compreendendo 31 participantes (81,6%), e apenas 10 casos (26,3%) foram
expostos à fumaça do cigarro como fumantes passivos (Tabela 2).
As substâncias e materiais que foram presentes no ambiente de trabalho atual e que
apresentaram maior frequência de exposição entre os 39 trabalhadores da construção do segundo
grupo, foram: areia (66,7%), concreto (82,1%), sílica (51,3%) e raios ultravioleta (79,5%) (Figura 8).
A exposição desses trabalhadores a empregos anteriores apresentou maior frequência de: areia
(84,6%), concreto (84,6%), sílica (56,4%), pó de madeira (38,5%) e radiação ultravioleta (87,2%)
(Figura 9).
Tabela 2 - Características sócio-demográficos dos participantes do segundo grupo coletado (curto período de exposição no ambiente da construção civil).
Variáveis
Caso (n=38)
Idade (Média anos) 38
Tempo de exposição (Média anos) 13
Fumante passivo Sim
28 (73,7)
Não
10 (26,3)
Consumo de álcool
Sim
31 (81,6)
Não/Passado
7 (18,4)
Etnia
Branco
8 (21,6)
Negro
15 (40,5)
Pardo
13 (35,1)
Oriental
1 (2,7)
N - Número de participantes
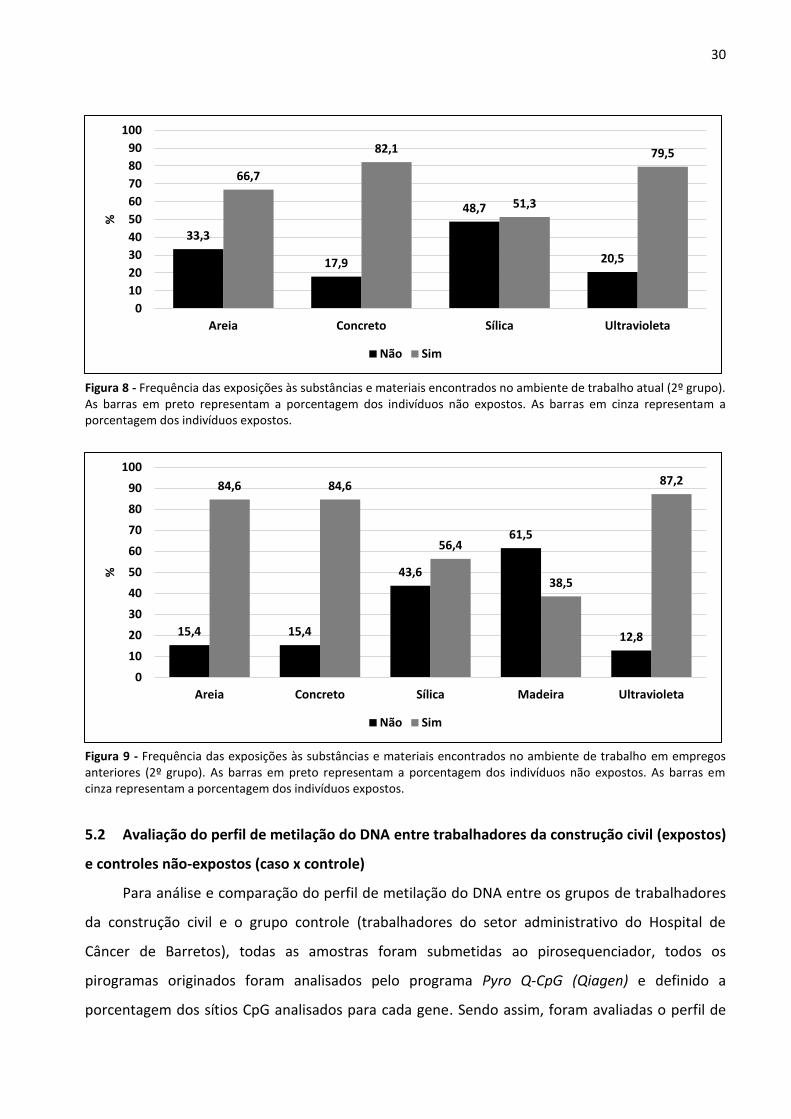
30
Figura 8 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho atual (2º grupo). As barras em preto representam a porcentagem dos indivíduos não expostos. As barras em cinza representam a porcentagem dos indivíduos expostos.
Figura 9 - Frequência das exposições às substâncias e materiais encontrados no ambiente de trabalho em empregos anteriores (2º grupo). As barras em preto representam a porcentagem dos indivíduos não expostos. As barras em cinza representam a porcentagem dos indivíduos expostos.
5.2 Avaliação do perfil de metilação do DNA entre trabalhadores da construção civil (expostos)
e controles não-expostos (caso x controle)
Para análise e comparação do perfil de metilação do DNA entre os grupos de trabalhadores
da construção civil e o grupo controle (trabalhadores do setor administrativo do Hospital de
Câncer de Barretos), todas as amostras foram submetidas ao pirosequenciador, todos os
pirogramas originados foram analisados pelo programa Pyro Q-CpG (Qiagen) e definido a
porcentagem dos sítios CpG analisados para cada gene. Sendo assim, foram avaliadas o perfil de
33,3
17,9
48,7
20,5
66,7
82,1
51,3
79,5
0
10
20
30
40
50
60
70
80
90
100
Areia Concreto Sílica Ultravioleta
%
Não Sim
15,4 15,4
43,6
61,5
12,8
84,6 84,6
56,4
38,5
87,2
0
10
20
30
40
50
60
70
80
90
100
Areia Concreto Sílica Madeira Ultravioleta
%
Não Sim
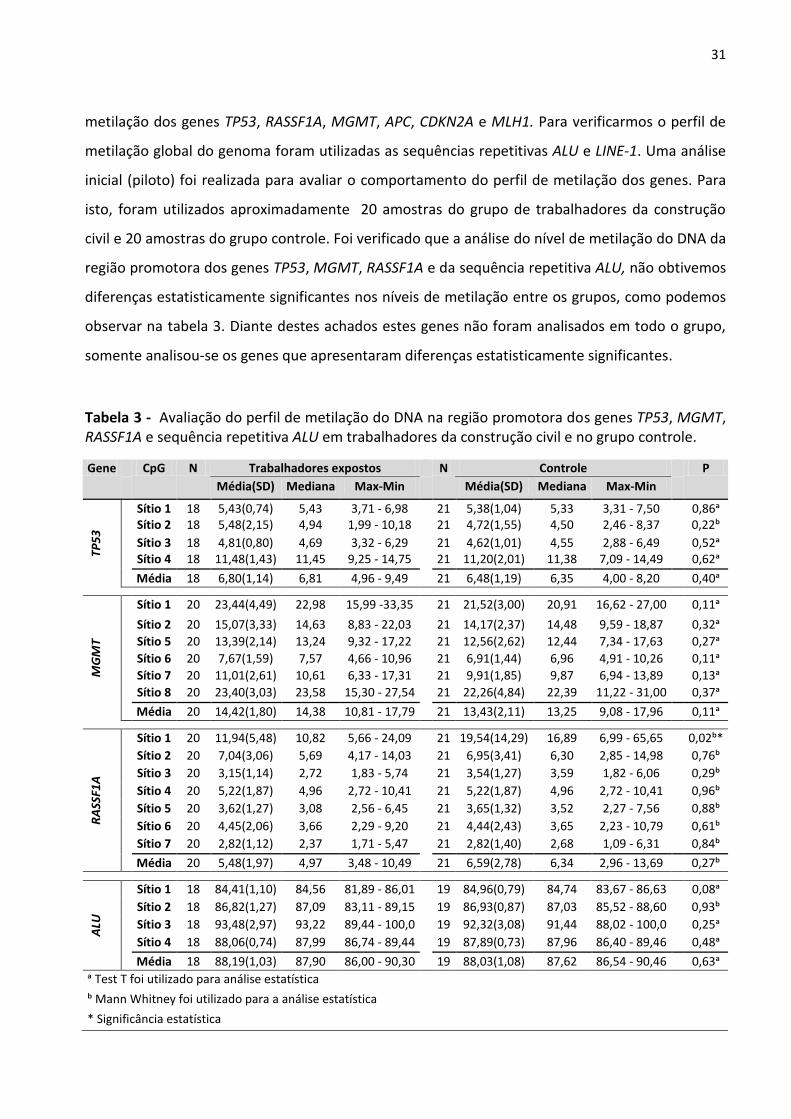
31
metilação dos genes TP53, RASSF1A, MGMT, APC, CDKN2A e MLH1. Para verificarmos o perfil de
metilação global do genoma foram utilizadas as sequências repetitivas ALU e LINE-1. Uma análise
inicial (piloto) foi realizada para avaliar o comportamento do perfil de metilação dos genes. Para
isto, foram utilizados aproximadamente 20 amostras do grupo de trabalhadores da construção
civil e 20 amostras do grupo controle. Foi verificado que a análise do nível de metilação do DNA da
região promotora dos genes TP53, MGMT, RASSF1A e da sequência repetitiva ALU, não obtivemos
diferenças estatisticamente significantes nos níveis de metilação entre os grupos, como podemos
observar na tabela 3. Diante destes achados estes genes não foram analisados em todo o grupo,
somente analisou-se os genes que apresentaram diferenças estatisticamente significantes.
Tabela 3 - Avaliação do perfil de metilação do DNA na região promotora dos genes TP53, MGMT, RASSF1A e sequência repetitiva ALU em trabalhadores da construção civil e no grupo controle.
Gene CpG N Trabalhadores expostos N Controle P
Média(SD) Mediana Max-Min Média(SD) Mediana Max-Min
TP5
3
Sítio 1 18 5,43(0,74) 5,43 3,71 - 6,98 21 5,38(1,04) 5,33 3,31 - 7,50 0,86ᵃ
Sítio 2 18 5,48(2,15) 4,94 1,99 - 10,18 21 4,72(1,55) 4,50 2,46 - 8,37 0,22ᵇ
Sítio 3 18 4,81(0,80) 4,69 3,32 - 6,29 21 4,62(1,01) 4,55 2,88 - 6,49 0,52ᵃ
Sítio 4 18 11,48(1,43) 11,45 9,25 - 14,75 21 11,20(2,01) 11,38 7,09 - 14,49 0,62ᵃ
Média 18 6,80(1,14) 6,81 4,96 - 9,49 21 6,48(1,19) 6,35 4,00 - 8,20 0,40ᵃ
MG
MT
Sítio 1 20 23,44(4,49) 22,98 15,99 -33,35 21 21,52(3,00) 20,91 16,62 - 27,00 0,11ᵃ
Sítio 2 20 15,07(3,33) 14,63 8,83 - 22,03 21 14,17(2,37) 14,48 9,59 - 18,87 0,32ᵃ
Sítio 5 20 13,39(2,14) 13,24 9,32 - 17,22 21 12,56(2,62) 12,44 7,34 - 17,63 0,27ᵃ
Sítio 6 20 7,67(1,59) 7,57 4,66 - 10,96 21 6,91(1,44) 6,96 4,91 - 10,26 0,11ᵃ
Sítio 7 20 11,01(2,61) 10,61 6,33 - 17,31 21 9,91(1,85) 9,87 6,94 - 13,89 0,13ᵃ
Sítio 8 20 23,40(3,03) 23,58 15,30 - 27,54 21 22,26(4,84) 22,39 11,22 - 31,00 0,37ᵃ
Média 20 14,42(1,80) 14,38 10,81 - 17,79 21 13,43(2,11) 13,25 9,08 - 17,96 0,11ᵃ
RA
SSF1
A
Sítio 1 20 11,94(5,48) 10,82 5,66 - 24,09 21 19,54(14,29) 16,89 6,99 - 65,65 0,02ᵇ*
Sítio 2 20 7,04(3,06) 5,69 4,17 - 14,03 21 6,95(3,41) 6,30 2,85 - 14,98 0,76ᵇ
Sítio 3 20 3,15(1,14) 2,72 1,83 - 5,74 21 3,54(1,27) 3,59 1,82 - 6,06 0,29ᵇ
Sítio 4 20 5,22(1,87) 4,96 2,72 - 10,41 21 5,22(1,87) 4,96 2,72 - 10,41 0,96ᵇ
Sítio 5 20 3,62(1,27) 3,08 2,56 - 6,45 21 3,65(1,32) 3,52 2,27 - 7,56 0,88ᵇ
Sítio 6 20 4,45(2,06) 3,66 2,29 - 9,20 21 4,44(2,43) 3,65 2,23 - 10,79 0,61ᵇ
Sítio 7 20 2,82(1,12) 2,37 1,71 - 5,47 21 2,82(1,40) 2,68 1,09 - 6,31 0,84ᵇ
Média 20 5,48(1,97) 4,97 3,48 - 10,49 21 6,59(2,78) 6,34 2,96 - 13,69 0,27ᵇ
ALU
Sítio 1 18 84,41(1,10) 84,56 81,89 - 86,01 19 84,96(0,79) 84,74 83,67 - 86,63 0,08ᵃ
Sítio 2 18 86,82(1,27) 87,09 83,11 - 89,15 19 86,93(0,87) 87,03 85,52 - 88,60 0,93ᵇ
Sítio 3 18 93,48(2,97) 93,22 89,44 - 100,0 19 92,32(3,08) 91,44 88,02 - 100,0 0,25ᵃ
Sítio 4 18 88,06(0,74) 87,99 86,74 - 89,44 19 87,89(0,73) 87,96 86,40 - 89,46 0,48ᵃ
Média 18 88,19(1,03) 87,90 86,00 - 90,30 19 88,03(1,08) 87,62 86,54 - 90,46 0,63ᵃ
ᵃ Test T foi utilizado para análise estatística
ᵇ Mann Whitney foi utilizado para a análise estatística
* Significância estatística

32
5.2.1 Avaliação do perfil de metilação do DNA nos grupos caso e controle dos genes que
apresentaram significância estatística nas análises iniciais.
Para os genes verificados com níveis de metilação do DNA estatisticamente significantes no
piloto foram analisados a amostragem total de ambos os grupos. Sendo assim, foram verificados
os níveis de metilação dos genes CDKN2A, MLH1, APC e avaliação da metilação global pela
sequência repetitiva LINE-1.
Nas análises da região promotora do gene CDKN2A foram avaliados 57 indivíduos da
construção civil (caso) e 44 trabalhadores do setor administrativo (controle) e foi observado uma
média = 4,79%, mediana = 4,44 (0 – 10,73), para o grupo de trabalhadores da construção civil,
quando comparado ao grupo controle com média = 3,63%, mediana = 3,68 (0 – 7,47). Foi
verificada uma média estatisticamente significante no grupo de trabalhadores da construção civil
comparado aos controles. Apesar da significância estatística na média geral considerando todos os
sítios, nas análises sítio por sítio , o padrão dos níveis de metilação do DNA foram inconsistentes.
Dos 7 sítios analisados, somente os sítios 2 e 7 apresentaram diferenças estatísticas significantes
entre os grupos, apresentando p-valor = 0,02 e p-valor = 0,005, respectivamente . Os demais sítios
não demonstraram alterações significantes nos trabalhadores expostos comparados com o
controle, indicando que os outros 2 sítios CpG podem ser os Hot CpG sites e que provavelmente
podem estar participando da regulação da expressão gênica, seguindo conforme descrito por
Duan et al. 106 (Tabela 4) .
Para as análises da região promotora do gene MLH1 foram avaliados 56 indivíduos da
construção civil (caso) e 41 trabalhadores do setor administrativo (controle) e foi observado uma
média = 5,04%, mediana = 5,03% (1,61 – 9,76) para o grupo de trabalhadores da construção civil
quando comparado ao grupo controle com média = 4,22%, mediana = 4,00% (1,74 – 7,49).
Verificou-se uma diferença estatisticamente significante entre os grupos, sendo os níveis de
metilação maiores encontrados no grupo caso comparados com o controle. Para as análises
realizadas para cada sitio CpG, 4 sítios apresentaram diferenças estatísticas significantes entre os
grupos caso e controle, sendo eles os sítios 3 (p= 0,02), sítio 4 (p=0,01), sítio 5 (p=0,001) e sítio 6
(p=0,001). Dois sítios (1 e 2) não demonstraram alterações significantes nos trabalhadores
expostos comparados com o controle. Dessa forma, os 4 sítios que apresentaram diferença
estatística entre os grupos provavelmente são os Hot CpG sites e que possivelmente podem estar
participando da regulação gênica (Tabela 4).

33
Na verificação dos níveis de metilação da região promotora do gene APC, foram avaliados 58
indivíduos da construção civil (caso) e 47 trabalhadores do setor administrativo (controle) e foi
observado uma média = 6,43%, mediana = 5,73% (1,17 – 15,48) para o grupo de trabalhadores da
construção civil quando comparado ao grupo controle com média = 5,47%, mediana = 6,13% (1,29
– 11,54). Não observamos uma diferença estatisticamente significante entre os grupos na média
geral, porém, para esse gene foram analisados 3 sítios CpG. Nas análises para cada sítio CpG,
foram encontrados 2 sítios que apresentaram valores significantes entre os grupos, sendo eles,
sítio 2 (p=0,002) e no sítio 3 (p=0,013) significantes entre os grupos caso e controle. O que pode
indicar serem sítios (Hot CpG sites), que provavelmente podem estar participando da regulação da
expressão gênica. Sendo assim, foram recalculados somente os sítios que apresentaram diferença
estatística (Hot CpG sites) e observou-se uma média aumentada = 4,29%, mediana = 4,29%, (0,81 –
9,61) para grupo dos trabalhadores da construção quando comparado ao grupo controle com
média = 3,11%, mediana = 2,61, (0 – 10,82) com p<0,001 (Tabela 4).
Para verificação da hipótese de participação dos sítios analisados na regulação gênica, foram
realizadas análises in sílico afim de confirmarmos a presença de motifs de ligação de fatores de
transcrição nas sequencias que foram analisadas no pirosequenciador, para os genes CDKN2A,
MLH1 e APC. (Anexo F).
Para as análises de avaliação da metilação global da região repetitiva LINE-1 foram avaliados
57 indivíduos da construção civil (caso) e 44 trabalhadores do setor administrativo (controle) e foi
observado uma média = 66,83%, mediana = 65,93 (62,48 – 72,48) para o grupo de trabalhadores
da construção civil, quando comparado ao grupo controle com média = 68,04%, mediana = 68,63
(64,98 – 70,37). Verificamos uma diferença estatisticamente significante entre os grupos. Dessa
maneira foi observado a média menor nos níveis de metilação do DNA global no grupo caso em
comparação ao grupo controle (p=0,005). Nas análises para cada sítio CpG, foram verificados 5
sítios para a sequência LINE-1, sendo que 2 sítios apresentaram diferenças estatísticas
significantes entre os grupos caso e controle, sendo eles os sítios 1 (p= 0,01), sítio 5 (p=0,005)
(Tabela 4).
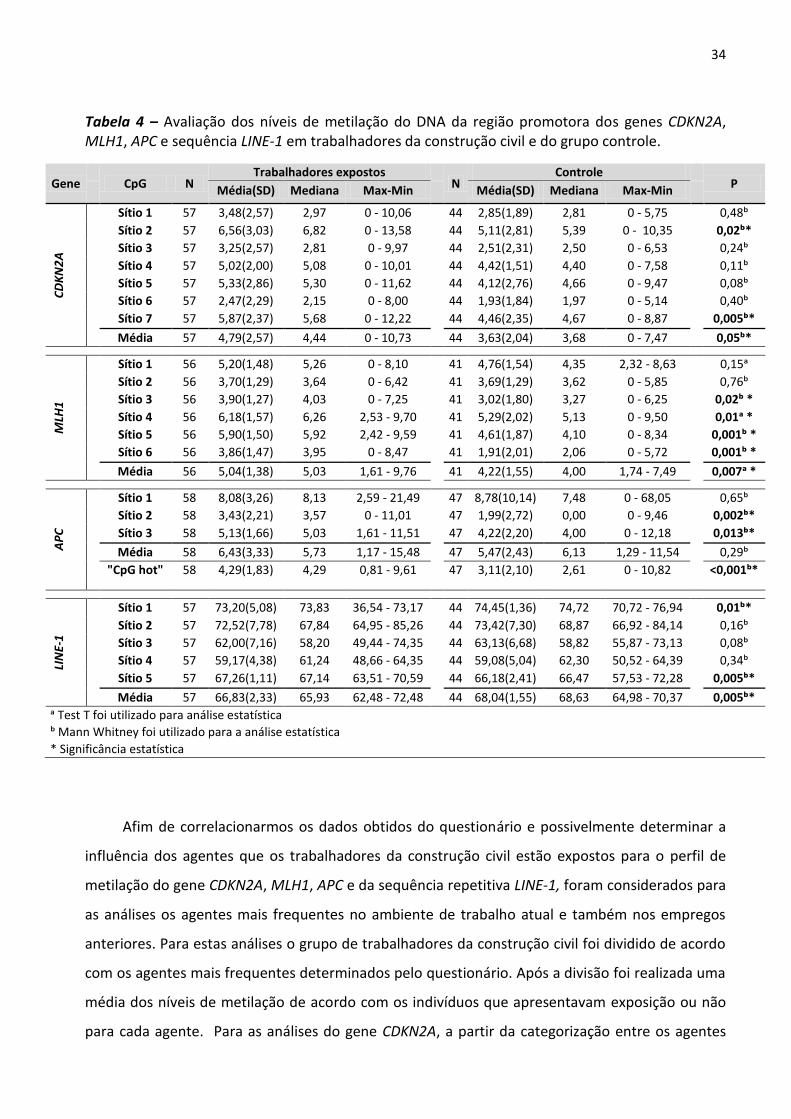
34
Tabela 4 – Avaliação dos níveis de metilação do DNA da região promotora dos genes CDKN2A, MLH1, APC e sequência LINE-1 em trabalhadores da construção civil e do grupo controle.
Gene
CpG N Trabalhadores expostos
N Controle
P Média(SD) Mediana Max-Min Média(SD) Mediana Max-Min
CD
KN
2A
Sítio 1 57 3,48(2,57) 2,97 0 - 10,06 44 2,85(1,89) 2,81 0 - 5,75 0,48ᵇ
Sítio 2 57 6,56(3,03) 6,82 0 - 13,58 44 5,11(2,81) 5,39 0 - 10,35 0,02ᵇ*
Sítio 3 57 3,25(2,57) 2,81 0 - 9,97 44 2,51(2,31) 2,50 0 - 6,53 0,24ᵇ
Sítio 4 57 5,02(2,00) 5,08 0 - 10,01 44 4,42(1,51) 4,40 0 - 7,58 0,11ᵇ
Sítio 5 57 5,33(2,86) 5,30 0 - 11,62 44 4,12(2,76) 4,66 0 - 9,47 0,08ᵇ
Sítio 6 57 2,47(2,29) 2,15 0 - 8,00 44 1,93(1,84) 1,97 0 - 5,14 0,40ᵇ
Sítio 7 57 5,87(2,37) 5,68 0 - 12,22 44 4,46(2,35) 4,67 0 - 8,87 0,005ᵇ*
Média 57 4,79(2,57) 4,44 0 - 10,73 44 3,63(2,04) 3,68 0 - 7,47 0,05ᵇ*
MLH
1
Sítio 1 56 5,20(1,48) 5,26 0 - 8,10 41 4,76(1,54) 4,35 2,32 - 8,63 0,15ᵃ
Sítio 2 56 3,70(1,29) 3,64 0 - 6,42 41 3,69(1,29) 3,62 0 - 5,85 0,76ᵇ
Sítio 3 56 3,90(1,27) 4,03 0 - 7,25 41 3,02(1,80) 3,27 0 - 6,25 0,02ᵇ *
Sítio 4 56 6,18(1,57) 6,26 2,53 - 9,70 41 5,29(2,02) 5,13 0 - 9,50 0,01ᵃ *
Sítio 5 56 5,90(1,50) 5,92 2,42 - 9,59 41 4,61(1,87) 4,10 0 - 8,34 0,001ᵇ *
Sítio 6 56 3,86(1,47) 3,95 0 - 8,47 41 1,91(2,01) 2,06 0 - 5,72 0,001ᵇ *
Média 56 5,04(1,38) 5,03 1,61 - 9,76 41 4,22(1,55) 4,00 1,74 - 7,49 0,007ᵃ *
AP
C
Sítio 1 58 8,08(3,26) 8,13 2,59 - 21,49 47 8,78(10,14) 7,48 0 - 68,05 0,65ᵇ
Sítio 2 58 3,43(2,21) 3,57 0 - 11,01 47 1,99(2,72) 0,00 0 - 9,46 0,002ᵇ*
Sítio 3 58 5,13(1,66) 5,03 1,61 - 11,51 47 4,22(2,20) 4,00 0 - 12,18 0,013ᵇ*
Média 58 6,43(3,33) 5,73 1,17 - 15,48 47 5,47(2,43) 6,13 1,29 - 11,54 0,29ᵇ
"CpG hot" 58 4,29(1,83) 4,29 0,81 - 9,61 47 3,11(2,10) 2,61 0 - 10,82 <0,001ᵇ*
LIN
E-1
Sítio 1 57 73,20(5,08) 73,83 36,54 - 73,17 44 74,45(1,36) 74,72 70,72 - 76,94 0,01ᵇ*
Sítio 2 57 72,52(7,78) 67,84 64,95 - 85,26 44 73,42(7,30) 68,87 66,92 - 84,14 0,16ᵇ
Sítio 3 57 62,00(7,16) 58,20 49,44 - 74,35 44 63,13(6,68) 58,82 55,87 - 73,13 0,08ᵇ
Sítio 4 57 59,17(4,38) 61,24 48,66 - 64,35 44 59,08(5,04) 62,30 50,52 - 64,39 0,34ᵇ
Sítio 5 57 67,26(1,11) 67,14 63,51 - 70,59 44 66,18(2,41) 66,47 57,53 - 72,28 0,005ᵇ*
Média 57 66,83(2,33) 65,93 62,48 - 72,48 44 68,04(1,55) 68,63 64,98 - 70,37 0,005ᵇ*
ᵃ Test T foi utilizado para análise estatística
ᵇ Mann Whitney foi utilizado para a análise estatística
* Significância estatística
Afim de correlacionarmos os dados obtidos do questionário e possivelmente determinar a
influência dos agentes que os trabalhadores da construção civil estão expostos para o perfil de
metilação do gene CDKN2A, MLH1, APC e da sequência repetitiva LINE-1, foram considerados para
as análises os agentes mais frequentes no ambiente de trabalho atual e também nos empregos
anteriores. Para estas análises o grupo de trabalhadores da construção civil foi dividido de acordo
com os agentes mais frequentes determinados pelo questionário. Após a divisão foi realizada uma
média dos níveis de metilação de acordo com os indivíduos que apresentavam exposição ou não
para cada agente. Para as análises do gene CDKN2A, a partir da categorização entre os agentes
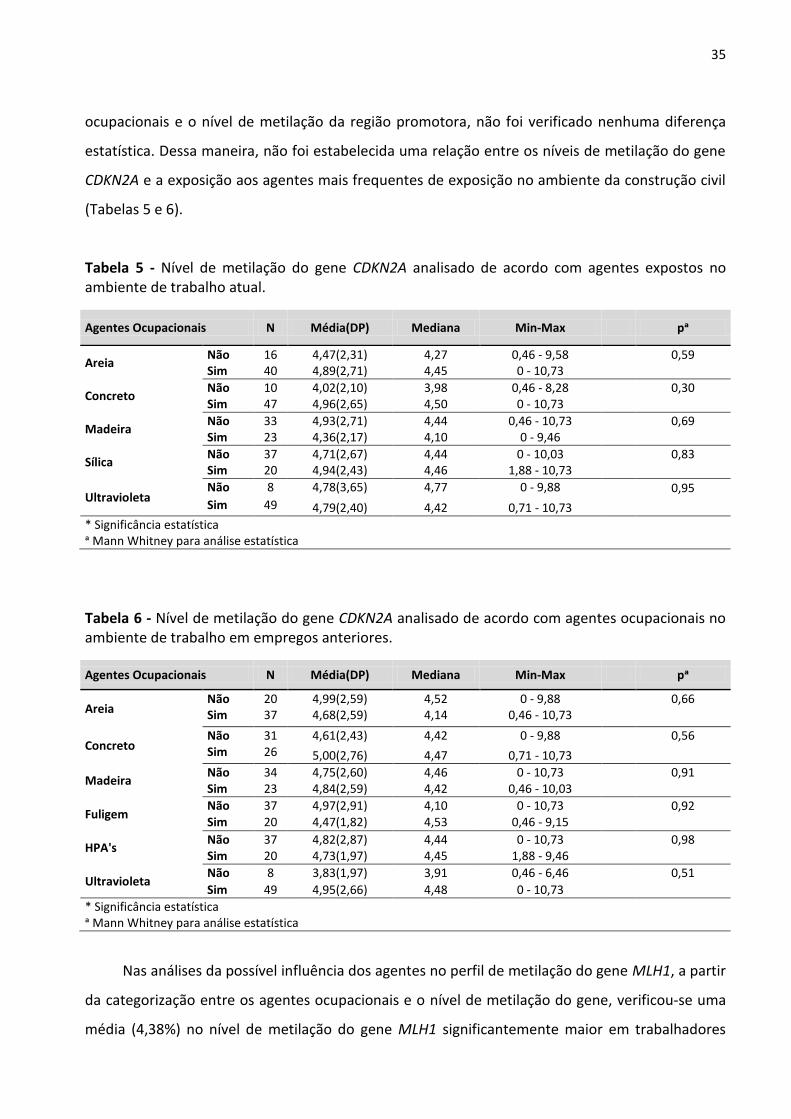
35
ocupacionais e o nível de metilação da região promotora, não foi verificado nenhuma diferença
estatística. Dessa maneira, não foi estabelecida uma relação entre os níveis de metilação do gene
CDKN2A e a exposição aos agentes mais frequentes de exposição no ambiente da construção civil
(Tabelas 5 e 6).
Tabela 5 - Nível de metilação do gene CDKN2A analisado de acordo com agentes expostos no ambiente de trabalho atual.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 16 4,47(2,31) 4,27 0,46 - 9,58 0,59 Sim 40 4,89(2,71) 4,45 0 - 10,73
Concreto Não 10 4,02(2,10) 3,98 0,46 - 8,28 0,30 Sim 47 4,96(2,65) 4,50 0 - 10,73
Madeira Não 33 4,93(2,71) 4,44 0,46 - 10,73 0,69 Sim 23 4,36(2,17) 4,10 0 - 9,46
Sílica Não 37 4,71(2,67) 4,44 0 - 10,03 0,83 Sim 20 4,94(2,43) 4,46 1,88 - 10,73
Ultravioleta Não 8 4,78(3,65) 4,77 0 - 9,88 0,95
Sim 49 4,79(2,40) 4,42 0,71 - 10,73
* Significância estatística ᵃ Mann Whitney para análise estatística
Tabela 6 - Nível de metilação do gene CDKN2A analisado de acordo com agentes ocupacionais no ambiente de trabalho em empregos anteriores.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 20 4,99(2,59) 4,52 0 - 9,88 0,66 Sim 37 4,68(2,59) 4,14 0,46 - 10,73
Concreto Não 31 4,61(2,43) 4,42 0 - 9,88 0,56 Sim 26 5,00(2,76) 4,47 0,71 - 10,73
Madeira Não 34 4,75(2,60) 4,46 0 - 10,73 0,91 Sim 23 4,84(2,59) 4,42 0,46 - 10,03
Fuligem Não 37 4,97(2,91) 4,10 0 - 10,73 0,92 Sim 20 4,47(1,82) 4,53 0,46 - 9,15
HPA's Não 37 4,82(2,87) 4,44 0 - 10,73 0,98 Sim 20 4,73(1,97) 4,45 1,88 - 9,46
Ultravioleta Não 8 3,83(1,97) 3,91 0,46 - 6,46 0,51
Sim 49 4,95(2,66) 4,48 0 - 10,73
* Significância estatística ᵃ Mann Whitney para análise estatística
Nas análises da possível influência dos agentes no perfil de metilação do gene MLH1, a partir
da categorização entre os agentes ocupacionais e o nível de metilação do gene, verificou-se uma
média (4,38%) no nível de metilação do gene MLH1 significantemente maior em trabalhadores
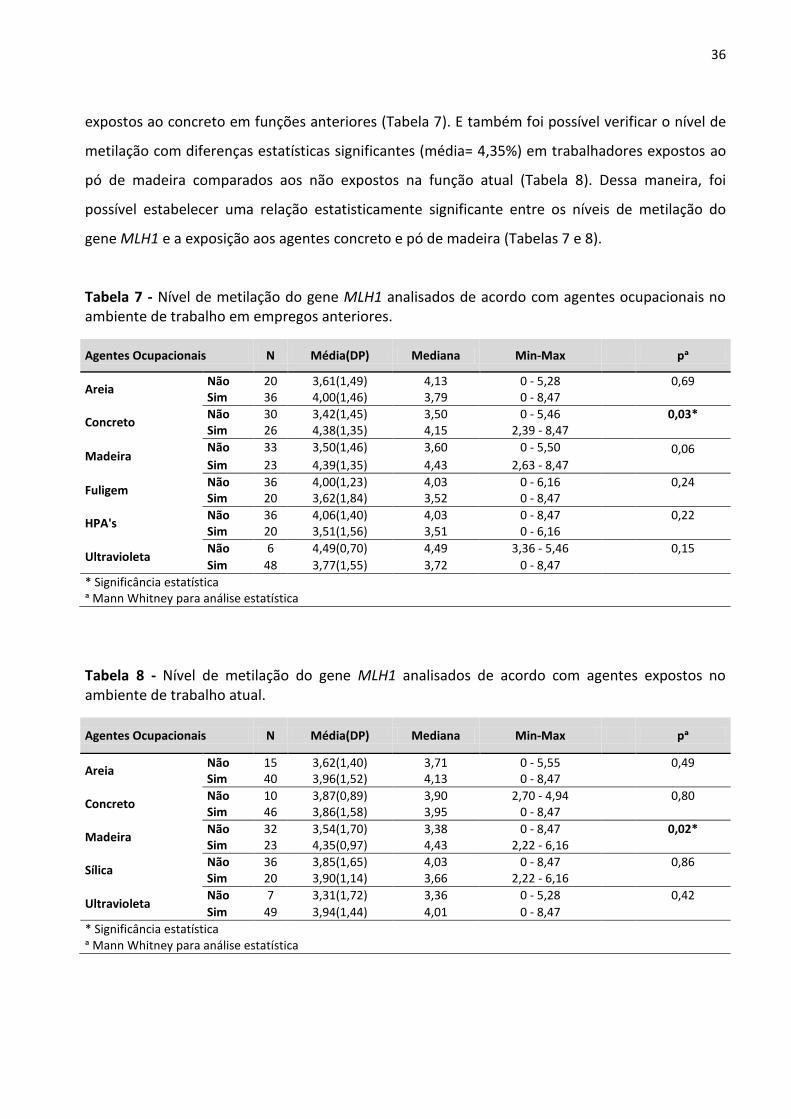
36
expostos ao concreto em funções anteriores (Tabela 7). E também foi possível verificar o nível de
metilação com diferenças estatísticas significantes (média= 4,35%) em trabalhadores expostos ao
pó de madeira comparados aos não expostos na função atual (Tabela 8). Dessa maneira, foi
possível estabelecer uma relação estatisticamente significante entre os níveis de metilação do
gene MLH1 e a exposição aos agentes concreto e pó de madeira (Tabelas 7 e 8).
Tabela 7 - Nível de metilação do gene MLH1 analisados de acordo com agentes ocupacionais no ambiente de trabalho em empregos anteriores.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 20 3,61(1,49) 4,13 0 - 5,28 0,69 Sim 36 4,00(1,46) 3,79 0 - 8,47
Concreto Não 30 3,42(1,45) 3,50 0 - 5,46 0,03* Sim 26 4,38(1,35) 4,15 2,39 - 8,47
Madeira Não 33 3,50(1,46) 3,60 0 - 5,50 0,06 Sim 23 4,39(1,35) 4,43 2,63 - 8,47
Fuligem Não 36 4,00(1,23) 4,03 0 - 6,16 0,24 Sim 20 3,62(1,84) 3,52 0 - 8,47
HPA's Não 36 4,06(1,40) 4,03 0 - 8,47 0,22 Sim 20 3,51(1,56) 3,51 0 - 6,16
Ultravioleta Não 6 4,49(0,70) 4,49 3,36 - 5,46 0,15
Sim 48 3,77(1,55) 3,72 0 - 8,47
* Significância estatística ᵃ Mann Whitney para análise estatística
Tabela 8 - Nível de metilação do gene MLH1 analisados de acordo com agentes expostos no ambiente de trabalho atual.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 15 3,62(1,40) 3,71 0 - 5,55 0,49 Sim 40 3,96(1,52) 4,13 0 - 8,47
Concreto Não 10 3,87(0,89) 3,90 2,70 - 4,94 0,80 Sim 46 3,86(1,58) 3,95 0 - 8,47
Madeira Não 32 3,54(1,70) 3,38 0 - 8,47 0,02* Sim 23 4,35(0,97) 4,43 2,22 - 6,16
Sílica Não 36 3,85(1,65) 4,03 0 - 8,47 0,86 Sim 20 3,90(1,14) 3,66 2,22 - 6,16
Ultravioleta Não 7 3,31(1,72) 3,36 0 - 5,28 0,42
Sim 49 3,94(1,44) 4,01 0 - 8,47
* Significância estatística ᵃ Mann Whitney para análise estatística
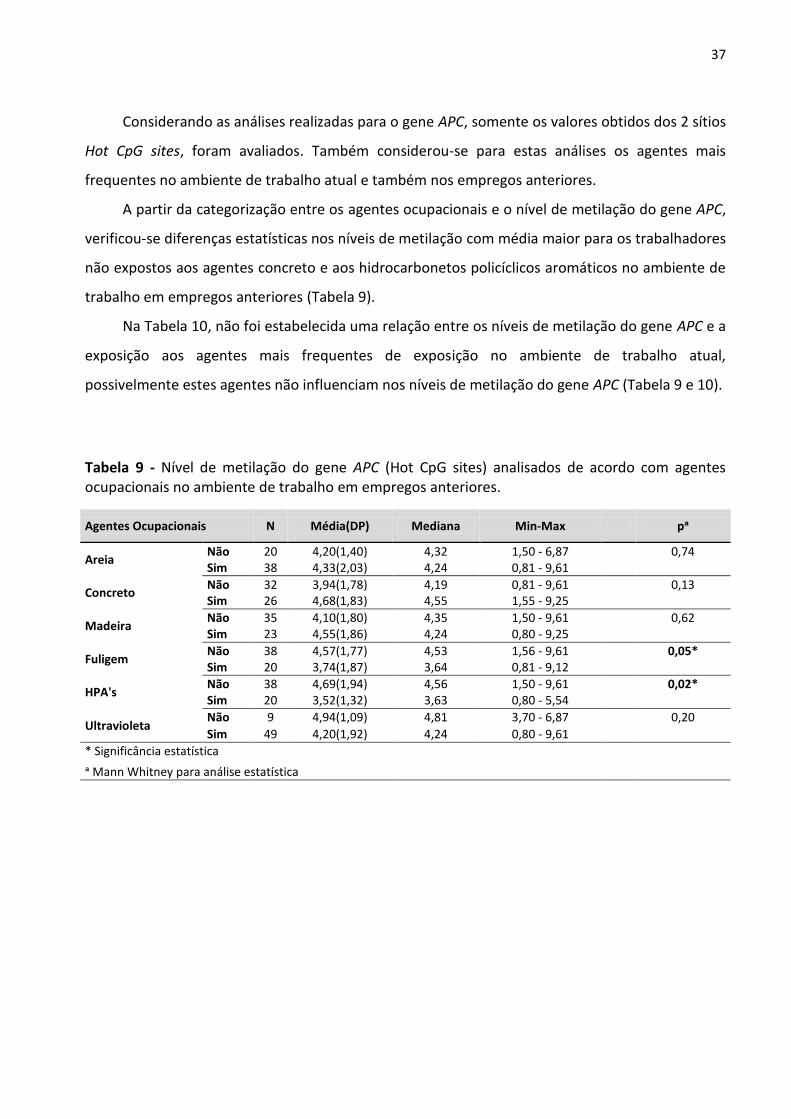
37
Considerando as análises realizadas para o gene APC, somente os valores obtidos dos 2 sítios
Hot CpG sites, foram avaliados. Também considerou-se para estas análises os agentes mais
frequentes no ambiente de trabalho atual e também nos empregos anteriores.
A partir da categorização entre os agentes ocupacionais e o nível de metilação do gene APC,
verificou-se diferenças estatísticas nos níveis de metilação com média maior para os trabalhadores
não expostos aos agentes concreto e aos hidrocarbonetos policíclicos aromáticos no ambiente de
trabalho em empregos anteriores (Tabela 9).
Na Tabela 10, não foi estabelecida uma relação entre os níveis de metilação do gene APC e a
exposição aos agentes mais frequentes de exposição no ambiente de trabalho atual,
possivelmente estes agentes não influenciam nos níveis de metilação do gene APC (Tabela 9 e 10).
Tabela 9 - Nível de metilação do gene APC (Hot CpG sites) analisados de acordo com agentes ocupacionais no ambiente de trabalho em empregos anteriores.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 20 4,20(1,40) 4,32 1,50 - 6,87 0,74 Sim 38 4,33(2,03) 4,24 0,81 - 9,61
Concreto Não 32 3,94(1,78) 4,19 0,81 - 9,61 0,13 Sim 26 4,68(1,83) 4,55 1,55 - 9,25
Madeira Não 35 4,10(1,80) 4,35 1,50 - 9,61 0,62 Sim 23 4,55(1,86) 4,24 0,80 - 9,25
Fuligem Não 38 4,57(1,77) 4,53 1,56 - 9,61 0,05* Sim 20 3,74(1,87) 3,64 0,81 - 9,12
HPA's Não 38 4,69(1,94) 4,56 1,50 - 9,61 0,02* Sim 20 3,52(1,32) 3,63 0,80 - 5,54
Ultravioleta Não 9 4,94(1,09) 4,81 3,70 - 6,87 0,20
Sim 49 4,20(1,92) 4,24 0,80 - 9,61
* Significância estatística
ᵃ Mann Whitney para análise estatística
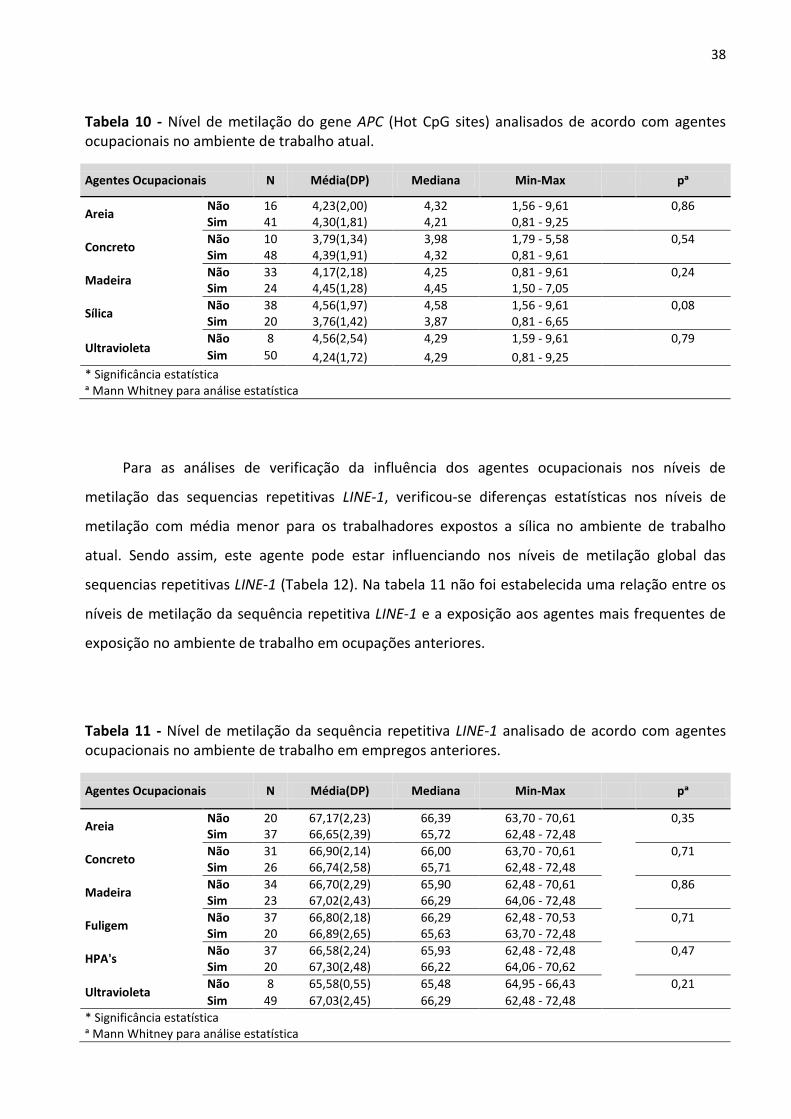
38
Tabela 10 - Nível de metilação do gene APC (Hot CpG sites) analisados de acordo com agentes ocupacionais no ambiente de trabalho atual.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 16 4,23(2,00) 4,32 1,56 - 9,61 0,86 Sim 41 4,30(1,81) 4,21 0,81 - 9,25
Concreto Não 10 3,79(1,34) 3,98 1,79 - 5,58 0,54 Sim 48 4,39(1,91) 4,32 0,81 - 9,61
Madeira Não 33 4,17(2,18) 4,25 0,81 - 9,61 0,24 Sim 24 4,45(1,28) 4,45 1,50 - 7,05
Sílica Não 38 4,56(1,97) 4,58 1,56 - 9,61 0,08 Sim 20 3,76(1,42) 3,87 0,81 - 6,65
Ultravioleta Não 8 4,56(2,54) 4,29 1,59 - 9,61 0,79
Sim 50 4,24(1,72) 4,29 0,81 - 9,25
* Significância estatística ᵃ Mann Whitney para análise estatística
Para as análises de verificação da influência dos agentes ocupacionais nos níveis de
metilação das sequencias repetitivas LINE-1, verificou-se diferenças estatísticas nos níveis de
metilação com média menor para os trabalhadores expostos a sílica no ambiente de trabalho
atual. Sendo assim, este agente pode estar influenciando nos níveis de metilação global das
sequencias repetitivas LINE-1 (Tabela 12). Na tabela 11 não foi estabelecida uma relação entre os
níveis de metilação da sequência repetitiva LINE-1 e a exposição aos agentes mais frequentes de
exposição no ambiente de trabalho em ocupações anteriores.
Tabela 11 - Nível de metilação da sequência repetitiva LINE-1 analisado de acordo com agentes ocupacionais no ambiente de trabalho em empregos anteriores.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 20 67,17(2,23) 66,39 63,70 - 70,61 0,35 Sim 37 66,65(2,39) 65,72 62,48 - 72,48
Concreto Não 31 66,90(2,14) 66,00 63,70 - 70,61 0,71 Sim 26 66,74(2,58) 65,71 62,48 - 72,48
Madeira Não 34 66,70(2,29) 65,90 62,48 - 70,61 0,86 Sim 23 67,02(2,43) 66,29 64,06 - 72,48
Fuligem Não 37 66,80(2,18) 66,29 62,48 - 70,53 0,71 Sim 20 66,89(2,65) 65,63 63,70 - 72,48
HPA's Não 37 66,58(2,24) 65,93 62,48 - 72,48 0,47 Sim 20 67,30(2,48) 66,22 64,06 - 70,62
Ultravioleta Não 8 65,58(0,55) 65,48 64,95 - 66,43 0,21
Sim 49 67,03(2,45) 66,29 62,48 - 72,48
* Significância estatística ᵃ Mann Whitney para análise estatística
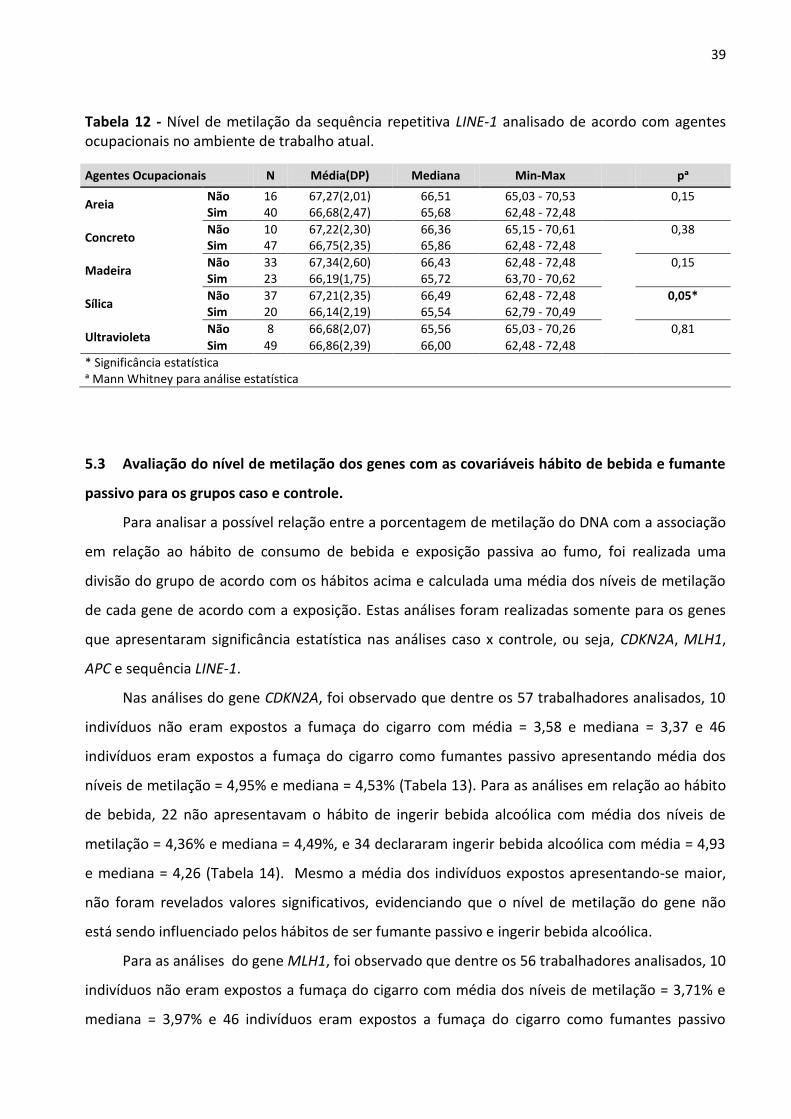
39
Tabela 12 - Nível de metilação da sequência repetitiva LINE-1 analisado de acordo com agentes ocupacionais no ambiente de trabalho atual.
Agentes Ocupacionais N Média(DP) Mediana Min-Max pᵃ
Areia Não 16 67,27(2,01) 66,51 65,03 - 70,53 0,15 Sim 40 66,68(2,47) 65,68 62,48 - 72,48
Concreto Não 10 67,22(2,30) 66,36 65,15 - 70,61 0,38 Sim 47 66,75(2,35) 65,86 62,48 - 72,48
Madeira Não 33 67,34(2,60) 66,43 62,48 - 72,48 0,15 Sim 23 66,19(1,75) 65,72 63,70 - 70,62
Sílica Não 37 67,21(2,35) 66,49 62,48 - 72,48 0,05* Sim 20 66,14(2,19) 65,54 62,79 - 70,49
Ultravioleta Não 8 66,68(2,07) 65,56 65,03 - 70,26 0,81
Sim 49 66,86(2,39) 66,00 62,48 - 72,48
* Significância estatística ᵃ Mann Whitney para análise estatística
5.3 Avaliação do nível de metilação dos genes com as covariáveis hábito de bebida e fumante
passivo para os grupos caso e controle.
Para analisar a possível relação entre a porcentagem de metilação do DNA com a associação
em relação ao hábito de consumo de bebida e exposição passiva ao fumo, foi realizada uma
divisão do grupo de acordo com os hábitos acima e calculada uma média dos níveis de metilação
de cada gene de acordo com a exposição. Estas análises foram realizadas somente para os genes
que apresentaram significância estatística nas análises caso x controle, ou seja, CDKN2A, MLH1,
APC e sequência LINE-1.
Nas análises do gene CDKN2A, foi observado que dentre os 57 trabalhadores analisados, 10
indivíduos não eram expostos a fumaça do cigarro com média = 3,58 e mediana = 3,37 e 46
indivíduos eram expostos a fumaça do cigarro como fumantes passivo apresentando média dos
níveis de metilação = 4,95% e mediana = 4,53% (Tabela 13). Para as análises em relação ao hábito
de bebida, 22 não apresentavam o hábito de ingerir bebida alcoólica com média dos níveis de
metilação = 4,36% e mediana = 4,49%, e 34 declararam ingerir bebida alcoólica com média = 4,93
e mediana = 4,26 (Tabela 14). Mesmo a média dos indivíduos expostos apresentando-se maior,
não foram revelados valores significativos, evidenciando que o nível de metilação do gene não
está sendo influenciado pelos hábitos de ser fumante passivo e ingerir bebida alcoólica.
Para as análises do gene MLH1, foi observado que dentre os 56 trabalhadores analisados, 10
indivíduos não eram expostos a fumaça do cigarro com média dos níveis de metilação = 3,71% e
mediana = 3,97% e 46 indivíduos eram expostos a fumaça do cigarro como fumantes passivo

40
apresentando média de metilação = 3,90% e mediana = 3,88% (Tabela 13). Quando foi verificado
em relação ao hábito de bebida, 22 não apresentavam o hábito de ingerir bebida alcoólica com
média = 3,84 e mediana = 4,09, e 34 declararam ingerir bebida alcoólica com média = 3,88% e
mediana = 3,82% (Tabela 14). Mesmo a média dos indivíduos expostos apresentando-se maior, a
análise estatística não foi significativa, evidenciando que o nível de metilação do gene não está
sendo influenciado diante do hábito de bebida e exposição a fumaça do cigarro.
Na análise do gene APC (somente os Hot CpG sites foram considerados), foi observado que
dentre os 58 trabalhadores analisados, 10 indivíduos não eram expostos a fumaça do cigarro
apresentando média dos níveis de metilação = 4,17% e mediana = 4,56% e 48 indivíduos foram
expostos a fumaça do cigarro como fumantes passivo apresentando média = 4,31% e mediana =
4,24% (Tabela 13). Para as análises com o hábito de bebida, 23 não apresentavam o hábito de
ingerir bebida alcoólica com média = 4,35% e mediana = 4,19%, e 35 declararam ingerir bebida
alcoólica com média dos níveis de metilação = 4,24% e mediana = 4,30% (Tabela 14). Mesmo a
média dos indivíduos expostos apresentando-se maior, não foram evidenciados valores
estatisticamente significativos, dessa forma, possivelmente o nível de metilação do gene não está
influenciado com os hábito de bebida e de exposição como fumante passivo.
Para a análise da sequência repetitiva LINE-1, foi observado que dentre os 57 trabalhadores
analisados, 10 indivíduos não eram expostos a fumaça do cigarro com média dos níveis de
metilação = 65,81% e mediana = 65,37% e 46 indivíduos eram expostos a fumaça do cigarro como
fumantes passivo apresentando média dos níveis de metilação = 66,98% e mediana = 66,15%
(Tabela 13). Para as análises em relação ao hábito de bebida, 22 não apresentavam o hábito de
ingerir bebida alcoólica com média dos níveis de metilação = 66,33% e mediana = 65,79%, e 34
declararam ingerir bebida alcoólica com média = 67,06% e mediana = 65,97% (Tabela 14). Mesmo
a média dos indivíduos expostos apresentando-se maior, não se obteve significância estatística,
evidenciando que o nível de metilação do elemento repetitivo não está sendo influenciado com os
hábito de bebida e exposição a fumaça do cigarro.
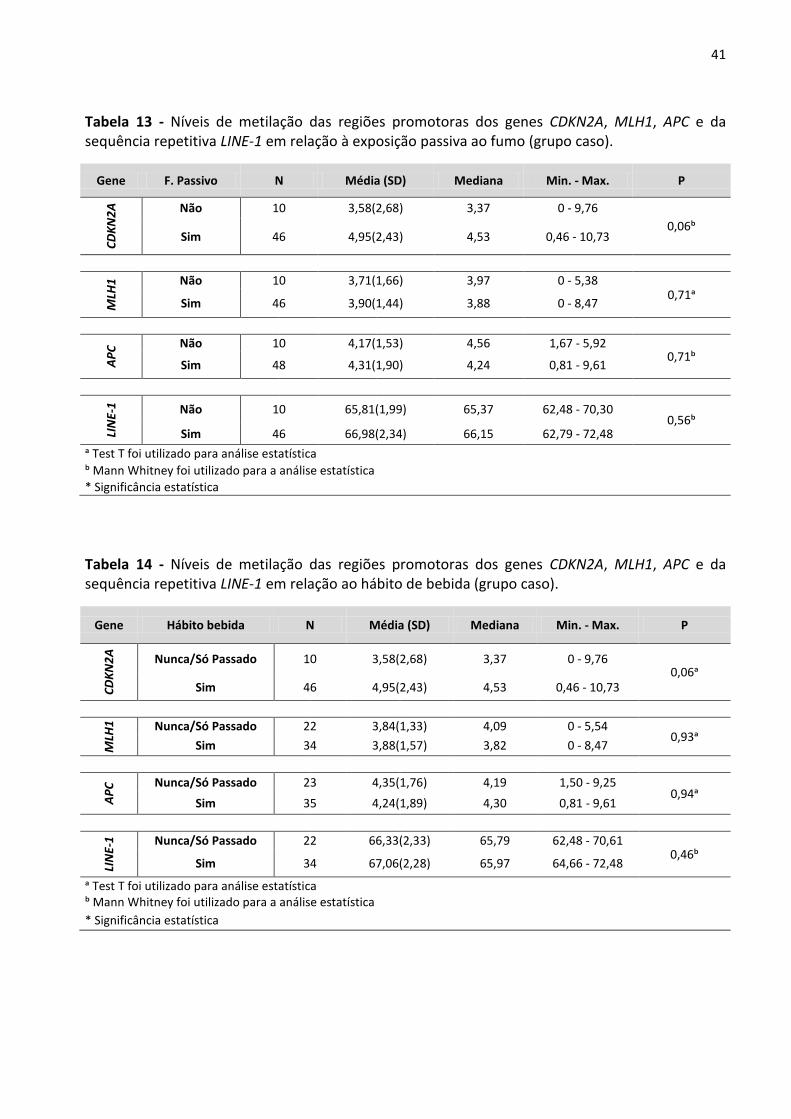
41
Tabela 13 - Níveis de metilação das regiões promotoras dos genes CDKN2A, MLH1, APC e da sequência repetitiva LINE-1 em relação à exposição passiva ao fumo (grupo caso).
Gene F. Passivo N Média (SD) Mediana Min. - Max. P
CD
KN
2A
Não 10 3,58(2,68) 3,37 0 - 9,76
0,06ᵇ Sim 46 4,95(2,43) 4,53 0,46 - 10,73
MLH
1 Não 10 3,71(1,66) 3,97 0 - 5,38
0,71ᵃ Sim 46 3,90(1,44) 3,88 0 - 8,47
AP
C Não 10 4,17(1,53) 4,56 1,67 - 5,92
0,71ᵇ Sim 48 4,31(1,90) 4,24 0,81 - 9,61
LIN
E-1
Não 10 65,81(1,99) 65,37 62,48 - 70,30 0,56ᵇ
Sim 46 66,98(2,34) 66,15 62,79 - 72,48
ᵃ Test T foi utilizado para análise estatística
ᵇ Mann Whitney foi utilizado para a análise estatística
* Significância estatística
Tabela 14 - Níveis de metilação das regiões promotoras dos genes CDKN2A, MLH1, APC e da sequência repetitiva LINE-1 em relação ao hábito de bebida (grupo caso).
Gene Hábito bebida N Média (SD) Mediana Min. - Max. P
CD
KN
2A
Nunca/Só Passado 10 3,58(2,68) 3,37 0 - 9,76 0,06ᵃ
Sim 46 4,95(2,43) 4,53 0,46 - 10,73
MLH
1
Nunca/Só Passado 22 3,84(1,33) 4,09 0 - 5,54 0,93ᵃ
Sim 34 3,88(1,57) 3,82 0 - 8,47
AP
C Nunca/Só Passado 23 4,35(1,76) 4,19 1,50 - 9,25
0,94ᵃ Sim 35 4,24(1,89) 4,30 0,81 - 9,61
LIN
E-1
Nunca/Só Passado 22 66,33(2,33) 65,79 62,48 - 70,61 0,46ᵇ
Sim 34 67,06(2,28) 65,97 64,66 - 72,48
ᵃ Test T foi utilizado para análise estatística ᵇ Mann Whitney foi utilizado para a análise estatística
* Significância estatística

42
5.4 Avaliação do nível de metilação dos genes considerando as covariáveis hábito de bebida,
fumante passivo e ex-fumantes para os grupos caso/controle.
Afim de verificar a influência nos níveis de metilação dos trabalhadores do grupo caso em
relação a serem ex-fumantes, fumantes passivos e consumir bebida alcoólica, para os genes que
apresentaram significância estatística nas análises caso x controle, foi realizada uma análise
seguindo o modelo de regressão linear múltipla, considerando as variáveis citadas a cima. Dessa
forma, foi ajustado um modelo para cada gene analisado, tendo como resposta a porcentagem
média de metilação do DNA.
Nas análises do gene CDKN2A, quando analisou-se as quatro variáveis em conjunto,
observou-se que, apesar de não significativa, o grupo apresenta um valor de significância menor,
sendo p = 0,06, enquanto as outras variáveis ex-fumantes, fumantes passivos e hábito de bebida,
apresentam valores maiores, sendo, p = 0,18, p = 0,70 e p = 0,22, respectivamente. (Tabela 15).
Para as análises do gene MLH1, após a análise das quatro variáveis, observamos que a
variável “grupo” apresenta valores significativos, sendo, p < 0,001 quando analisado em conjunto
com as outras variáveis que não foram significativas. (Tabela 15).
Nas análises do gene APC (Hot CpG sites), foi observado que na correlação entre as variáveis,
o grupo apresenta valores significativos, sendo p = 0,03 e quando consideradas as outras variáveis
os valores não são estatisticamente significantes. (Tabela 15).
Para a análise da sequência repetitiva LINE-1, foi observado que dentre as quatro variáveis, o
grupo apresentou um p = 0,03, sendo estatisticamente significantes em relação as outras
variáveis. Para ex-fumantes observamos p = 0,95, fumante passivo p = 0,38 e hábito de bebida p =
0,18. (Tabela 15).
Baseado no modelo de regressão linear múltipla, constatou-se que os níveis de metilação de
todos os genes sofrem uma influência significativa do grupo (exposição) quando as outras
variáveis são mantidas constantes, isso pode ser verificado na tabela 16. Quando analisou-se os
grupos independentes, mantendo-se as variáveis constantes, verificamos que os genes CDKN2A,
MLH1, APC apresentaram média de porcentagem de metilação maiores no grupo caso, sugerindo,
mais uma vez, que os níveis de metilação desses genes estão influenciados pela exposição dos
trabalhadores. O mesmo ocorre quando verificou-se a sequência repetitiva LINE-1, foi observado
que mantendo-se as variáveis constantes e analisando apenas o grupo, os níveis de metilação no
grupo caso são menores, sugerindo também, que a exposição influência nos níveis de
porcentagem de metilação.
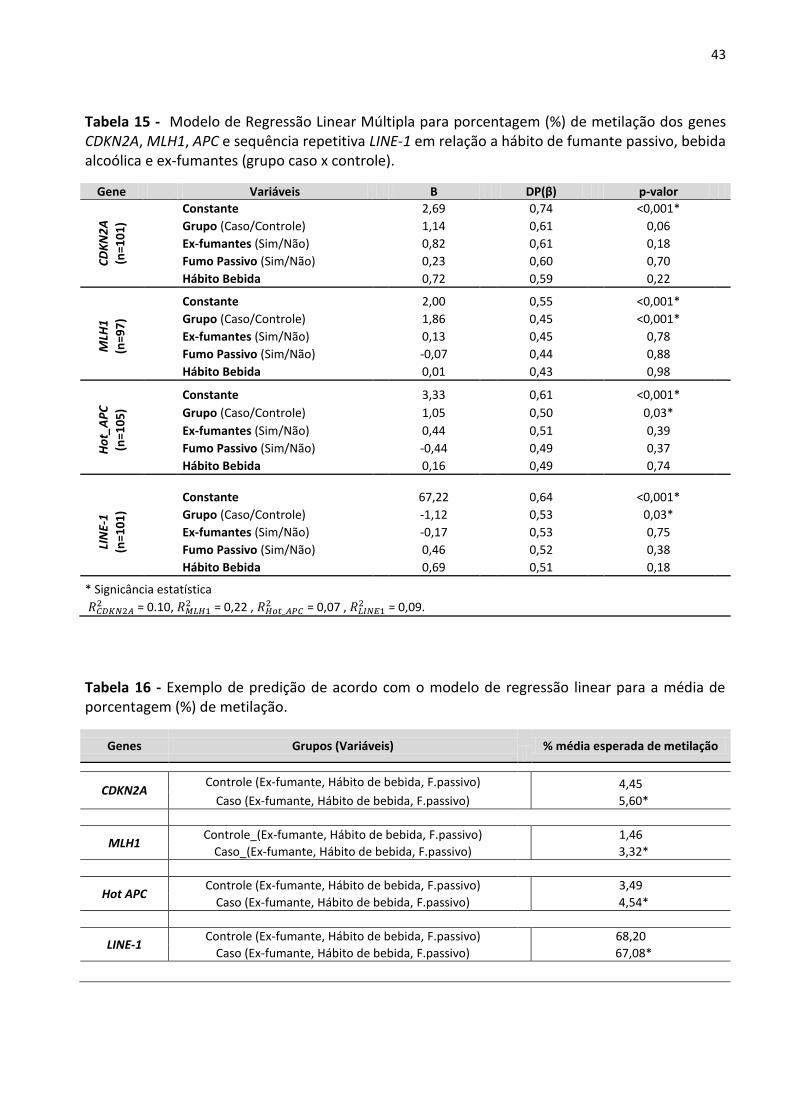
43
Tabela 15 - Modelo de Regressão Linear Múltipla para porcentagem (%) de metilação dos genes CDKN2A, MLH1, APC e sequência repetitiva LINE-1 em relação a hábito de fumante passivo, bebida alcoólica e ex-fumantes (grupo caso x controle).
Gene Variáveis Β DP(β) p-valor
CD
KN
2A
(n=1
01
)
Constante
2,69
0,74
<0,001* Grupo (Caso/Controle)
1,14
0,61
0,06
Ex-fumantes (Sim/Não)
0,82
0,61
0,18 Fumo Passivo (Sim/Não)
0,23
0,60
0,70
Hábito Bebida 0,72 0,59 0,22
MLH
1
(n=9
7)
Constante
2,00
0,55
<0,001* Grupo (Caso/Controle)
1,86
0,45
<0,001*
Ex-fumantes (Sim/Não)
0,13
0,45
0,78 Fumo Passivo (Sim/Não)
-0,07
0,44
0,88
Hábito Bebida 0,01 0,43 0,98
Ho
t_A
PC
(n=1
05
)
Constante
3,33
0,61
<0,001* Grupo (Caso/Controle)
1,05
0,50
0,03*
Ex-fumantes (Sim/Não)
0,44
0,51
0,39 Fumo Passivo (Sim/Não)
-0,44
0,49
0,37
Hábito Bebida 0,16 0,49 0,74
LIN
E-1
(n=1
01
)
Constante
67,22
0,64
<0,001* Grupo (Caso/Controle)
-1,12
0,53
0,03*
Ex-fumantes (Sim/Não)
-0,17
0,53
0,75 Fumo Passivo (Sim/Não)
0,46
0,52
0,38
Hábito Bebida
0,69
0,51
0,18 * Signicância estatística
= 0.10,
= 0,22 , = 0,07 ,
= 0,09.
Tabela 16 - Exemplo de predição de acordo com o modelo de regressão linear para a média de porcentagem (%) de metilação.
Genes Grupos (Variáveis)
% média esperada de metilação
CDKN2A
Controle (Ex-fumante, Hábito de bebida, F.passivo) 4,45
Caso (Ex-fumante, Hábito de bebida, F.passivo) 5,60*
MLH1 Controle_(Ex-fumante, Hábito de bebida, F.passivo) 1,46
Caso_(Ex-fumante, Hábito de bebida, F.passivo) 3,32*
Hot APC Controle (Ex-fumante, Hábito de bebida, F.passivo) 3,49
Caso (Ex-fumante, Hábito de bebida, F.passivo) 4,54*
LINE-1 Controle (Ex-fumante, Hábito de bebida, F.passivo) 68,20
Caso (Ex-fumante, Hábito de bebida, F.passivo) 67,08*

44
5.5 Análise do perfil de metilação do DNA nos trabalhadores da construção civil expostos no
1º dia de trabalho da semana (segunda- feira) e 5º dia de trabalho da semana (sexta- feira).
Para análise e comparação do perfil de metilação do DNA desse segundo grupo, foram
coletados sangue dos trabalhadores da construção civil em dois dias diferentes da semana, a
coleta foi realizada do mesmo indivíduo no 1° dia de trabalho da semana e a segunda coleta no 5°
dia de trabalho, a fim de detectar as potenciais diferenças nos níveis de metilação do DNA dos
genes propostos em curto período de tempo de exposição no ambiente de trabalho da construção
civil. Além disso, as análises com este grupo proporcionam a verificação do nível de metilação do
DNA dos indivíduos trabalhando no mesmo local de trabalho sem interferentes de outros
participantes que não tem a exposição e com características socioeconômicas diferentes. Sendo
assim, foram avaliadas o perfil de metilação dos genes MGMT, RASSF1A, APC, MLH1 e CDKN2A e
das sequências repetitivas ALU e LINE-1.
Nas análises das regiões promotoras dos genes MGMT, RASSF1A, APC, MLH1 e do elemento
repetitivo ALU foram analisados 38 indivíduos da construção civil no 1º dia de trabalho da semana
e no 5º dia de trabalho da semana. Para análise de LINE-1 analisou-se 39 trabalhadores da
construção civil. Conforme pode-se observar na Tabela 17 as análises dos genes MGMT, RASSF1A,
APC, MLH1 e sequências ALU e LINE-1 em trabalhadores expostos no 1º e 5º dia da semana, não
foram verificadas diferenças estatisticamente significantes entre os grupos.
Para as análises da região promotora do gene CDKN2A foram analisados 38 indivíduos da
construção civil no 1º dia de trabalho da semana e foi observado uma média = 2,88%, mediana =
2,62% (1,39 – 5,56) e comparado ao 5º dia de trabalho da semana com média = 3,31%, mediana =
3,15 (1,35 – 6,17), verificamos uma diferença estatisticamente significante entre os grupos no 1º e
5º dia de exposição. Nas análises de cada sítio CpG, foram avaliados 7 sítios para o gene CDKN2A,
sendo que 4 sítios apresentaram diferenças estatísticas significantes entre os 1º e 5º dia de coleta,
sendo eles os sítios 1 (p= 0,02), sítio 4 (p=0,06), sítio 5 (p=0,01) e sítio 6 (p=0,008). Também foi
observado a média maior com valores significantes na metilação do DNA no grupo exposto no 5º
dia de exposição (p=0,04), o que possivelmente nos leva a considerar que um curto tempo de
exposição pode alterar o nível de metilação do DNA desses trabalhadores (Tabela 18). Estes dados
confirmam com os achados encontrados nas análises do primeiro grupo (caso x controle), onde a
região promotora de gene CDKN2A apresenta níveis de metilação significantes entre os grupos.
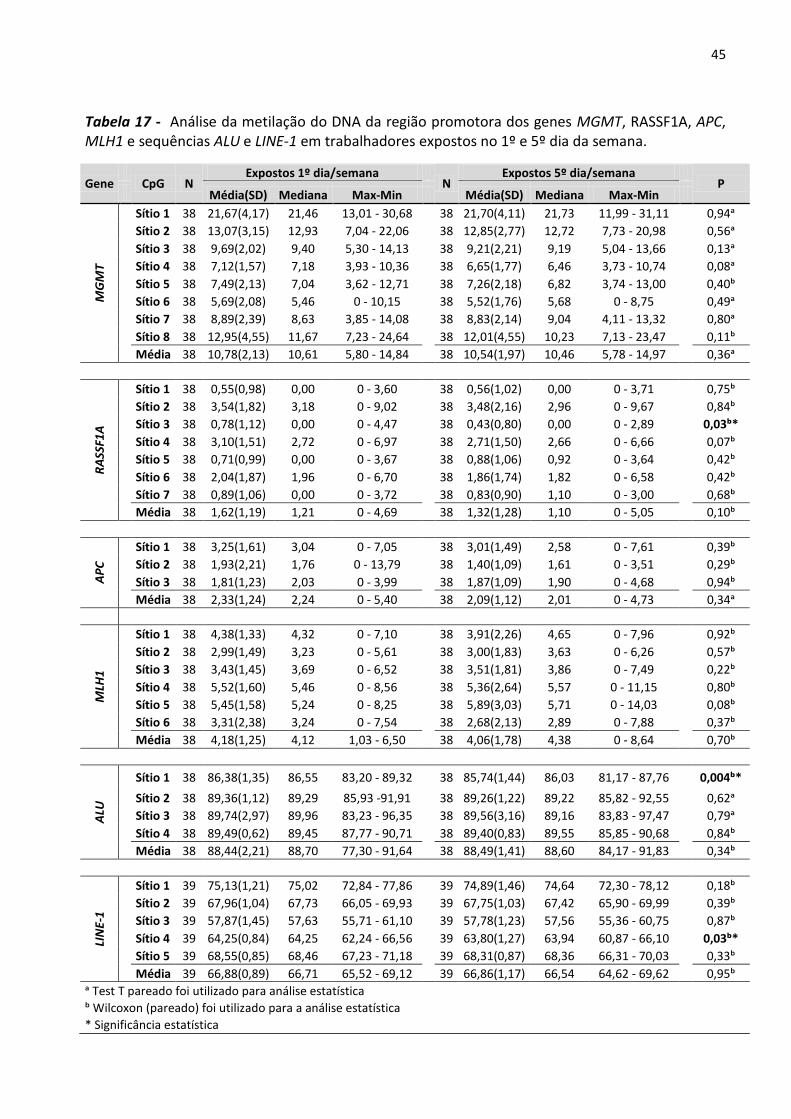
45
Tabela 17 - Análise da metilação do DNA da região promotora dos genes MGMT, RASSF1A, APC, MLH1 e sequências ALU e LINE-1 em trabalhadores expostos no 1º e 5º dia da semana.
Gene
CpG N Expostos 1º dia/semana
N Expostos 5º dia/semana
P Média(SD) Mediana Max-Min Média(SD) Mediana Max-Min
MG
MT
Sítio 1 38 21,67(4,17) 21,46 13,01 - 30,68
38 21,70(4,11) 21,73 11,99 - 31,11
0,94ᵃ
Sítio 2 38 13,07(3,15) 12,93 7,04 - 22,06
38 12,85(2,77) 12,72 7,73 - 20,98
0,56ᵃ
Sítio 3 38 9,69(2,02) 9,40 5,30 - 14,13
38 9,21(2,21) 9,19 5,04 - 13,66
0,13ᵃ
Sítio 4 38 7,12(1,57) 7,18 3,93 - 10,36
38 6,65(1,77) 6,46 3,73 - 10,74
0,08ᵃ
Sítio 5 38 7,49(2,13) 7,04 3,62 - 12,71
38 7,26(2,18) 6,82 3,74 - 13,00
0,40ᵇ
Sítio 6 38 5,69(2,08) 5,46 0 - 10,15
38 5,52(1,76) 5,68 0 - 8,75
0,49ᵃ
Sítio 7 38 8,89(2,39) 8,63 3,85 - 14,08
38 8,83(2,14) 9,04 4,11 - 13,32
0,80ᵃ
Sítio 8 38 12,95(4,55) 11,67 7,23 - 24,64
38 12,01(4,55) 10,23 7,13 - 23,47
0,11ᵇ
Média 38 10,78(2,13) 10,61 5,80 - 14,84 38 10,54(1,97) 10,46 5,78 - 14,97 0,36ᵃ
RA
SSF1
A
Sítio 1 38 0,55(0,98) 0,00 0 - 3,60
38 0,56(1,02) 0,00 0 - 3,71
0,75ᵇ
Sítio 2 38 3,54(1,82) 3,18 0 - 9,02
38 3,48(2,16) 2,96 0 - 9,67
0,84ᵇ
Sítio 3 38 0,78(1,12) 0,00 0 - 4,47
38 0,43(0,80) 0,00 0 - 2,89
0,03ᵇ*
Sítio 4 38 3,10(1,51) 2,72 0 - 6,97
38 2,71(1,50) 2,66 0 - 6,66
0,07ᵇ
Sítio 5 38 0,71(0,99) 0,00 0 - 3,67
38 0,88(1,06) 0,92 0 - 3,64
0,42ᵇ
Sítio 6 38 2,04(1,87) 1,96 0 - 6,70
38 1,86(1,74) 1,82 0 - 6,58
0,42ᵇ
Sítio 7 38 0,89(1,06) 0,00 0 - 3,72
38 0,83(0,90) 1,10 0 - 3,00
0,68ᵇ
Média 38 1,62(1,19) 1,21 0 - 4,69 38 1,32(1,28) 1,10 0 - 5,05 0,10ᵇ
AP
C
Sítio 1 38 3,25(1,61) 3,04 0 - 7,05
38 3,01(1,49) 2,58 0 - 7,61
0,39ᵇ
Sítio 2 38 1,93(2,21) 1,76 0 - 13,79
38 1,40(1,09) 1,61 0 - 3,51
0,29ᵇ
Sítio 3 38 1,81(1,23) 2,03 0 - 3,99
38 1,87(1,09) 1,90 0 - 4,68
0,94ᵇ
Média 38 2,33(1,24) 2,24 0 - 5,40 38 2,09(1,12) 2,01 0 - 4,73 0,34ᵃ
MLH
1
Sítio 1 38 4,38(1,33) 4,32 0 - 7,10
38 3,91(2,26) 4,65 0 - 7,96
0,92ᵇ
Sítio 2 38 2,99(1,49) 3,23 0 - 5,61
38 3,00(1,83) 3,63 0 - 6,26
0,57ᵇ
Sítio 3 38 3,43(1,45) 3,69 0 - 6,52
38 3,51(1,81) 3,86 0 - 7,49
0,22ᵇ
Sítio 4 38 5,52(1,60) 5,46 0 - 8,56
38 5,36(2,64) 5,57 0 - 11,15
0,80ᵇ
Sítio 5 38 5,45(1,58) 5,24 0 - 8,25
38 5,89(3,03) 5,71 0 - 14,03
0,08ᵇ
Sítio 6 38 3,31(2,38) 3,24 0 - 7,54
38 2,68(2,13) 2,89 0 - 7,88
0,37ᵇ
Média 38 4,18(1,25) 4,12 1,03 - 6,50 38 4,06(1,78) 4,38 0 - 8,64 0,70ᵇ
ALU
Sítio 1 38 86,38(1,35) 86,55 83,20 - 89,32
38 85,74(1,44) 86,03 81,17 - 87,76
0,004ᵇ*
Sítio 2 38 89,36(1,12) 89,29 85,93 -91,91
38 89,26(1,22) 89,22 85,82 - 92,55
0,62ᵃ
Sítio 3 38 89,74(2,97) 89,96 83,23 - 96,35
38 89,56(3,16) 89,16 83,83 - 97,47
0,79ᵃ
Sítio 4 38 89,49(0,62) 89,45 87,77 - 90,71
38 89,40(0,83) 89,55 85,85 - 90,68
0,84ᵇ
Média 38 88,44(2,21) 88,70 77,30 - 91,64 38 88,49(1,41) 88,60 84,17 - 91,83 0,34ᵇ
LIN
E-1
Sítio 1 39 75,13(1,21) 75,02 72,84 - 77,86
39 74,89(1,46) 74,64 72,30 - 78,12
0,18ᵇ
Sítio 2 39 67,96(1,04) 67,73 66,05 - 69,93
39 67,75(1,03) 67,42 65,90 - 69,99
0,39ᵇ
Sítio 3 39 57,87(1,45) 57,63 55,71 - 61,10
39 57,78(1,23) 57,56 55,36 - 60,75
0,87ᵇ
Sítio 4 39 64,25(0,84) 64,25 62,24 - 66,56
39 63,80(1,27) 63,94 60,87 - 66,10
0,03ᵇ*
Sítio 5 39 68,55(0,85) 68,46 67,23 - 71,18
39 68,31(0,87) 68,36 66,31 - 70,03
0,33ᵇ
Média 39 66,88(0,89) 66,71 65,52 - 69,12
39 66,86(1,17) 66,54 64,62 - 69,62
0,95ᵇ
ᵃ Test T pareado foi utilizado para análise estatística
ᵇ Wilcoxon (pareado) foi utilizado para a análise estatística
* Significância estatística
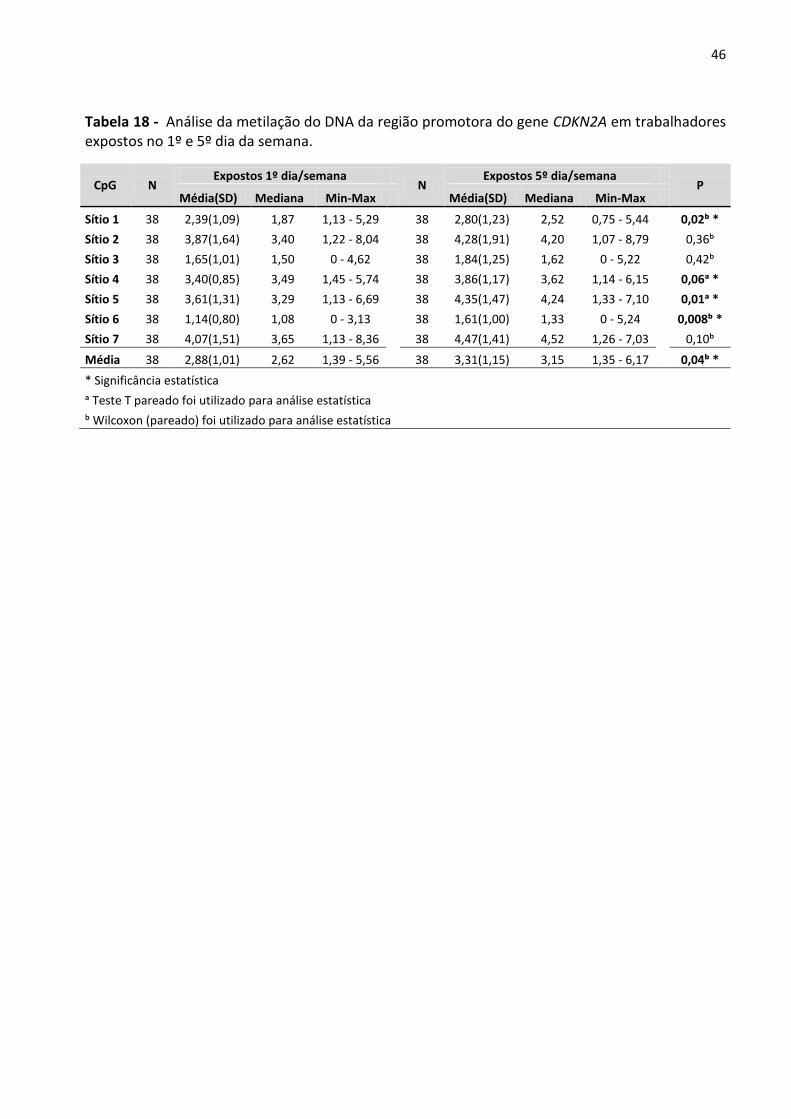
46
Tabela 18 - Análise da metilação do DNA da região promotora do gene CDKN2A em trabalhadores expostos no 1º e 5º dia da semana.
CpG N Expostos 1º dia/semana
N Expostos 5º dia/semana
P Média(SD) Mediana Min-Max Média(SD) Mediana Min-Max
Sítio 1 38 2,39(1,09) 1,87 1,13 - 5,29
38 2,80(1,23) 2,52 0,75 - 5,44
0,02ᵇ *
Sítio 2 38 3,87(1,64) 3,40 1,22 - 8,04
38 4,28(1,91) 4,20 1,07 - 8,79
0,36ᵇ
Sítio 3 38 1,65(1,01) 1,50 0 - 4,62
38 1,84(1,25) 1,62 0 - 5,22
0,42ᵇ
Sítio 4 38 3,40(0,85) 3,49 1,45 - 5,74
38 3,86(1,17) 3,62 1,14 - 6,15
0,06ᵃ *
Sítio 5 38 3,61(1,31) 3,29 1,13 - 6,69
38 4,35(1,47) 4,24 1,33 - 7,10
0,01ᵃ *
Sítio 6 38 1,14(0,80) 1,08 0 - 3,13
38 1,61(1,00) 1,33 0 - 5,24
0,008ᵇ *
Sítio 7 38 4,07(1,51) 3,65 1,13 - 8,36
38 4,47(1,41) 4,52 1,26 - 7,03
0,10ᵇ
Média 38 2,88(1,01) 2,62 1,39 - 5,56 38 3,31(1,15) 3,15 1,35 - 6,17 0,04ᵇ *
* Significância estatística ᵃ Teste T pareado foi utilizado para análise estatística
ᵇ Wilcoxon (pareado) foi utilizado para análise estatística

47
6 DISCUSSÃO
A indústria da construção civil continua a ser uma das principais fontes de emprego nos
países industrializados. Trabalhadores da construção podem ser agrupados em, qualificados, tais
como, pintores, carpinteiros, encanadores, eletricistas e trabalhadores de ferro, e não
qualificados, "operários", que executam uma variedade de tarefas de trabalho em apoio. Tais
trabalhadores são expostos a numerosos agentes físicos e químicos, como o amianto, sílica, outras
poeiras, solventes e outros produtos químicos116. Desde 1990, vários estudos de coorte realizados
em diferentes países encontraram resultados significativos de mortalidade para o câncer de
pulmão entre os trabalhadores da construção civil117. Um grande número de exposições
comumente encontrados no ambiente da indústria da construção têm demonstrado um possível
risco para o desenvolvimento de câncer, incluindo o de pulmão, entretanto, muitos destas
exposições não possuem avaliação para o risco de câncer. A metilação do DNA é uma modificação
epigenética que desempenha um papel essencial no estabelecimento da identidade celular
influenciando na expressão dos genes118. Sabe-se que este processo de metilação do DNA pode
sofrer alterações frente a fatores ambientais, bem como em populações expostas
ocupacionalmente.
No presente estudo nós avaliamos o perfil de metilação do DNA da região promotora dos
genes APC, RASSF1A, CDKN2A, TP53, MGMT, MLH1 e das sequências repetitivas ALU e LINE-1 em
leucócitos de sangue periférico dos trabalhadores da construção civil, utilizando-se o método de
pirosequenciamento que tem a vantagem de outros ensaios por produzir medidas individuais em
mais de um dinucleotídeo CpG, refletindo mais precisamente a metilação do DNA na região
estudada. Portanto, esse método foi escolhido por ser um ensaio quantitativo e qualitativo, e por
também apresentar controle para eficácia da conversão de bissulfito.
Os genes de interesse em nosso estudo estão envolvidos em controle do ciclo celular
(CDKN2A), invasão e metástase (APC), apoptose (TP53), via sinalização RAS (RASSF1A), via de
reparo e erros na replicação (MLH1), enzima de reparo e defesa de efeitos citotóxicos e
carcinogênicos (MGMT) e todos eles tem mostrado os níveis de metilação alterados no sangue de
pacientes com câncer de pulmão82,103. Considerando a primeira análise realizada comparando os
trabalhadores da construção com o grupo controle sem exposição, os níveis de metilação dos
genes MGMT, RASSF1A, TP53 e a sequência ALU, não foram encontrados valores significativos
entre os grupos. Somente os genes CDKN2A, MLH1, APC e a sequência LINE-1 foram significativos

48
nas análises entre o grupo caso x controle. Hossain et al.107 avaliaram os efeitos epigenéticos de
exposição ao arsênio nos genes relacionados a câncer, entre eles, CDKN2A, MLH1 e na sequência
repetitiva LINE-1 em indivíduos que tenham sido expostos através da ingestão de água. Foi
verificado que a exposição ao arsênio, aumenta os níveis de metilação do DNA dos genes CDKN2A
e MLH1 e diminui os níveis de metilação da sequência repetitiva LINE-1. Além disso, neste estudo
com as análises de regressão linear múltipla foi possível uma associação positiva entre a fração de
arsênio dosado na urina e a metilação de p16. Em nossos resultados, não encontramos uma
associação significativa quando correlacionamos certas substâncias presentes no ambiente de
trabalho da construção civil aos níveis de metilação da região promotora de CDKN2A. Por outro
lado, para os níveis de metilação do gene MLH1 foram encontradas associações significativas para
os indivíduos expostos ao concreto e ao pó de madeira.
O gene CDKN2A é um regulador do ciclo celular e hipermetilação deste gene tem sido
encontrada em tumores de cólon, do fígado, da pele, bem como em lesões pré-malignas119-120,121-
122-107. Adicionalmente, a hipermetilação de CDNK2A tem sido retratada não somente em tecidos
tumorais, mas também, em leucócitos do sangue periférico em indivíduos expostos a arsênio,
cromato, tabaco, radônio e ao material particulado contendo HPAs e radiação107-123,124-125-126-82.
Nossos resultados corroboram com estes estudos e foi verificado hipermetilação do gene CDKN2A
em leucócitos do sangue periférico no grupo exposto ao ambiente de trabalho da construção civil
em relação ao grupo controle.
O MLH1 é um componente importante na manutenção de estabilidade genômica e é
membro de um conjunto de genes conhecidos como os genes de reparação de emparelhamentos
incorretos (MMR). A metilação do gene MLH1 é relacionado no desenvolvimento câncer gástrico e
colorretal127-128,129-107. Como descrito anteriormente os níveis de metilação do gene MLH1 foram
significantes para os indivíduos expostos ao arsênio. No trabalho de Wu et al.103 foi analisado a
instabilidade cromossômica, avaliada por micronúcleos, e a metilação da região promotora do
gene MLH1 de trabalhadores expostos ao cloreto de vinila (plástico), onde o gene MLH1 não
apresentou valores significativos para indivíduos expostos e que apresentavam maiores danos
cromossômicos. Contrariamente aos nossos resultados onde os níveis de metilação do gene MLH1
foram significantes nas análises caso x controle.
No supressor tumoral APC foi verificado em nossos resultados a presença dos maiores níveis
de metilação no grupo de trabalhadores da construção civil quando comparado ao grupo controle.
No gene APC ocorre hipermetilação em estágios iniciais da tumorigénese e é um processo

49
estudado em câncer de pulmão82. No estudo desenvolvido por Ali et al.130 foi detectado níveis de
metilação aumentados nos supressores tumorais APC, MLH1 e CDKN2A em pacientes com câncer
de pulmão e com confirmado histórico de exposição ao crômio (trabalhadores da indústria),
comparado a pacientes com câncer de pulmão sem o mesmo histórico de ocupação. Os autores
sugeriram que o câncer de pulmão provocado pela exposição ao crômio é ligado a progressiva
metilação de alguns supressores tumorais. Ding et al.131 verificou que em pulmão de ratos
expostos a mistura de MP do trafego de veículos de acordo com o aumento do número de dias de
exposição, ocorreu o aumento dos níveis de metilação dos genes APC e CDKN2A e diminuição dos
níveis de metilação na sequência repetitiva LINE-1. Nossos resultados também encontraram
aumento da metilação do DNA nestes supressores tumorais e uma diminuição nos níveis de
metilação significativa de LINE-1 nos trabalhadores da construção civil expostos a uma mistura de
agentes no ambiente de trabalho. Estes dados sugerem que o conjunto de agentes no ambiente
de trabalho podem estar relacionados com as alterações nos níveis de metilação do DNA. Além
disso, essa mistura pode estar relacionada a instabilidade genômica, conforme descrito por Ramos
et al.132 que verificou em trabalhadores da construção civil uma maior frequência de micronúcleos
quando comparado a um grupo sem exposição.
Elementos repetitivos LINE-1 são retrotransposons altamente repetidos no genoma e seu
nível de metilação em sítios CpGs podem ser usados como uma medida da metilação global133.
Estudos têm utilizado metilação de LINE-1 como um biomarcador de susceptibilidade à câncer134.
A hipometilação de LINE-1 em leucócitos tem sido associado a um risco aumentado de vários tipos
de câncer135. Salas et al.133, avaliaram indivíduos expostos a trihalometano, uma classe de
subprodutos de desinfecção (solventes), e sugeriram uma associação positiva entre os níveis de
metilação de LINE-1 e a exposição a esse solvente.
No presente estudo nós avaliamos o perfil de metilação do DNA da região promotora dos
genes APC, RASSF1A, CDKN2A, TP53, MGMT, MLH1 e das sequências repetitivas ALU e LINE-1 em
leucócitos de sangue periférico dos trabalhadores da construção civil no primeiro dia da semana
de trabalho e no quinto dia. Somente obtivemos significância estatística nesta análise para o gene
CDKN2A, que apresentou maiores níveis de metilação no quinto dia de exposição. Para estas
análises optamos por um curto período de tempo de exposição dos trabalhadores ao ambiente de
trabalho da construção civil. Além disso, as análises com este grupo proporcionam a verificação do
nível de metilação do DNA dos indivíduos trabalhando no mesmo local de trabalho sem
interferentes de outros participantes que não tem a exposição ou com características

50
socioeconômicas diferentes. Hou et al.82 demonstraram em trabalhadores de fundição expostos
ao material particulado rico em partículas metálicas, que os níveis de metilação dos genes APC e
CDKN2A foram significativamente maiores no quarto dia de exposição ao ambiente de trabalho
quando comparado com o primeiro dia da semana (baseline). Existem evidências em seres
humanos entre as rápidas alterações na metilação do DNA frente as exposições ambientais, como
já discutido acima. Tais alterações podem persistir ao longo do tempo, até mesmo na ausência das
condições que as estabeleceram e acumular-se em resposta à exposição contínua.
Byun et al.112 realizaram um estudo de estabilidade temporal de vários marcadores
epigenéticos, em trabalhadores expostos ao MP, avaliando o 1° e 4° dia de trabalho da semana.
Foi demonstrado que a estabilidade do perfil de metilação do DNA é dependente do marcador
testado e também associado com as características das sequencias analisadas. Ainda neste estudo
APC, CDKN2A e CDH13 demonstraram pequenas diferenças estatisticamente significantes entre o
1° e o 4° dia. Nossos resultados foram encontrados somente alterações nos níveis de metilação do
gene CDKN2A, sugerindo que no ambiente da construção civil foi possível detectar alterações no
processo de metilação em curto período de tempo. Além disso, nossos resultados não podem ser
aplicados a mudanças progressivas e lentas na metilação do DNA, tais como, as associadas com o
envelhecimento112. A realização de estudos com maior número de tempos ao longo da exposição
podem fornecer achados mais consistentes sobre alterações acumulativas de longo prazo.
Neste trabalho nossos resultados foram inconsistentes para a grande maioria dos sítios CpG
analisados, sendo as diferenças encontradas em sítios isolados para cada gene, estes foram
chamados de Hot CpG sites. Estes resultados sugerem que existem hot spots, ou seja, CpG que
respondem aos estímulos tóxicos ambientais. Yang et al.136 avaliaram em trabalhadores expostos
aos HPAs que em 35 sítios CpG analisados na região promotora do gene CDKN2A foi verificado que
22 sítios CpG foram hipermetilados. Estes sítios estavam localizados em uma região a
aproximadamente a 230 pb do sitio de iniciação do gene e os autores sugerem que estes sítios é
que estão participando da regulação da expressão genica.
No estudo realizado por He et al.137, foi avaliado os níveis de metilação do gene RASSF1A no
sangue periférico de indivíduos expostos aos hidrocarbonetos policíclicos aromáticos. Foram
avaliados 87 sítios CpG e somente 5 foram hipermetilados nos trabalhadores expostos e estes
foram designados como hot CpG sites. Além disso, foi confirmado a hipermetilação destes sítios e
a diminuição da expressão de RASF1A in vitro em linfócitos humano e na linhagem celular de
pulmão HBE expostas a HPAs, dessa forma, confirmando a participação destes hot CpG sites na

51
regulação da transcrição. Sendo assim, a identificação de hot CpG sites para determinados genes
podem ser cruciais para afinidade de ligação de fatores de transcrição. Dessa forma, para os genes
em nosso trabalho que demonstraram diferenças estatísticas significantes e que foi possível
identificar os hot CpG sites, foi realizada uma análise de busca de motifs nas sequencias analisadas
e verificou-se vários sítios ligantes de fatores de transcrição, como podemos observar no anexo F.
Como mencionado acima em nossas observações, nossos resultados demonstram que a
detecção dos níveis de metilação do DNA em genes supressores tumorais, bem como, em
elementos repetitivos LINE-1, em leucócitos do sangue periférico, podem ser um valoroso
indicador de especificas exposições e efeitos adversos a saúde. Além disso, a identificação de
níveis alterados de metilação do DNA podem ser utilizados como potenciais biomarcadores para
detecção de alterações em indivíduos expostos ocupacionalmente. Estudos futuros devem ser
realizados para comprovar nossos achados. Análises in vitro, utilizando cultura de linfócitos e
linhagem celular de pulmão expostas ao MP captado no ambiente da construção civil, podem
elucidar os nossos achados sobre o perfil de metilação dos genes CDKN2A, APC, MLH1 e também
comprovar a influência na regulação da expressão destes genes. Ainda pretendemos avaliar o
perfil de metilação do DNA destes genes em pacientes com câncer de pulmão e com reconhecido
histórico de ocupação por longo período de tempo no ambiente da construção civil, e compara-los
com indivíduos com câncer sem este histórico. Dessa forma, relacionando o papel destes
resultados na carcinogênese.

52
7 CONCLUSÕES
Com base em nossos dados podemos concluir que:
1) Foi possível pela aplicação do questionário, caracterizar o ambiente da construção civil e
correlacionar estes dados com os níveis de metilação do DNA.
2) Diante dos nossos achados, não encontramos alterações nos níveis de metilação na
região promotora dos genes TP53, MGMT, RASSF1A e da sequência repetitiva ALU.
3) Foi possível verificar alterações nos níveis de metilação dos genes CDKN2A, MLH1, APC e
da sequência repetitiva LINE-1 em trabalhadores da construção civil comparados ao
grupo controle.
4) Além disso, foi encontrado alterações no perfil de metilação da região promotora do
gene CDKN2A no grupo de trabalhadores da construção civil durante curto tempo de
exposição.
5) Nossos resultados sugerem que o ambiente da construção civil pode influenciar no perfil
de metilação de genes envolvidos no processo de carcinogênese e também pode levar à
alterações na metilação global, proporcionando instabilidade genômica.

53
REFERÊNCIAS
1 Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer J Int Cancer. 2015; 136: E359–386.
2 Facina T. Resenha Estimativa 2014. Incidência de Câncer no Brasil. 2014; : 63.
3 World Health Organization. National cancer control programmes: policies and managerial guidelines. 2002.http://www.who.int/cancer/media/ en/408.pdf.
4 INCA. INCA. As profissões e o Câncer. 2012; 2012: 18–20.
5 Fritschi L, Driscoll T. Cancer due to occupation in Australia. Aust N Z J Public Health. 2006; 30: 213–219.
6 INCA. Diretrizes para a vigilância do câncer relacionado ao trabalho. MINISTÉRIO DA SAÚDE Instituto Nacional de Câncer José Alencar Gomes da Silva (INCA). 2012; : 187.
7 INCA. INCA. Fatores Ocupacionais. Rio Jan. 2015.http://www1.inca.gov.br/conteudo_view.asp?ID=17.
8 Ezzati M, Lopez AD. Estimates of global mortality attributable to smoking in 2000. Lancet. 2003; 362: 847–852.
9 Tyczynski JE, Bray F, Parkin DM. Lung cancer in Europe in 2000: epidemiology, prevention, and early detection. Lancet Oncol. 2003; 4: 45–55.
10 Olsson AC, Gustavsson P, Zaridze D, Mukeriya A, Szeszenia-Dabrowska N, Rudnai P et al. Lung cancer risk attributable to occupational exposures in a multicenter case-control study in Central and Eastern Europe. J Occup Environ Med Am Coll Occup Environ Med. 2011; 53: 1262–1267.
11 Driscoll T, Nelson DI, Steenland K, Leigh J, Concha-Barrientos M, Fingerhut M et al. The global burden of disease due to occupational carcinogens. Am J Ind Med. 2005; 48: 419–431.
12 Malhotra J, Sartori S, Brennan P, Zaridze D, Szeszenia-Dabrowska N, Świątkowska B et al. Effect of occupational exposures on lung cancer susceptibility: a study of gene-environment interaction analysis. Cancer Epidemiol Biomark Prev Publ Am Assoc Cancer Res Cosponsored Am Soc Prev Oncol. 2015; 24: 570–579.
13 Júnior CA da SF. A Saúde do Trabalhador no Maranhão - Uma visão atual e proposta de atuação. 1999.
14 Hashim D, Boffetta P. Occupational and environmental exposures and cancers in developing countries. Ann Glob Health. 2014; 80: 393–411.
15 Ziech D, Franco R, Pappa A, Malamou-Mitsi V, Georgakila S, Georgakilas AG et al. The role of epigenetics in environmental and occupational carcinogenesis. Chem Biol Interact. 2010; 188: 340–349.
16 IBGE. Pesquisa Anual da Indústria da Construção. 2012; 19: 1–98.
17 SNIC. SINDICATO NACIONAL DA INDÚSTRIA DO CIMENTO. 2013; : 40.

54
18 Consonni D, De Matteis S, Pesatori AC, Bertazzi PA, Olsson AC, Kromhout H et al. Lung cancer risk among bricklayers in a pooled analysis of case-control studies. Int J Cancer J Int Cancer. 2014; 136: 360–371.
19 INCA. Vigilância do Câncer relacionado ao trabalho e ao ambiente/Instituto Nacional do Câncer. Rev Atual. 2010; : 66.
20 Sellappa S, Prathyumnan S, Balachandar V. DNA damage induction and repair inhibition among building construction workers in South India. Asian Pac J Cancer Prev APJCP. 2010; 11: 875–880.
21 Bezerra PN, Vasconcelos AGA, Cavalcante LLA, Marques VB de V, Nogueira TNAG, Holanda MA. Hard metal lung disease in an oil industry worker. ras ne o i a a o o ras ne o Tisilogia. 2009; 35: 1254–1258.
22 Donaldson K, Poland CA. Nanotoxicology: new insights into nanotubes. Nat Nanotechnol. 2009; 4: 708–710.
23 Ryman-Rasmussen JP, Tewksbury EW, Moss OR, Cesta MF, Wong BA, Bonner JC. Inhaled multiwalled carbon nanotubes potentiate airway fibrosis in murine allergic asthma. Am J Respir Cell Mol Biol. 2009; 40: 349–358.
24 Yang M. A current global view of environmental and occupational cancers. J Environ Sci Health Part C Environ Carcinog Ecotoxicol Rev. 2012; 29: 223–249.
25 Menegozzo S, Comba P, Ferrante D, De Santis M, Gorini G, Izzo F et al. Mortality study in an asbestos cement factory in Naples, Italy. Ann DellIstituto Super Sanità. 2011; 47: 296–304.
26 Gómez MG, Castañeda R, López VG, Vidal MM, Villanueva V, Espinosa ME et al. Evaluation of the national health surveillance program of workers previously exposed to asbestos in Spain (2008). Gac Sanit SESPAS. 2012; 26: 45–50.
27 Straif K. The burden of occupational cancer. Occup Environ Med. 2008; 65: 787–788.
28 Yang P, MA J, Zhang B. Cancer Epidemiology, Biomarkers & Prevention. 2011.
29 Brown T, Darnton A, Fortunato L, Rushton L. Occupational Cancer Burden Study Group. Occupational cancer in Britain. Respiratory cancer sites: larynx, lung and mesothelioma. Br J Cancer. 2012; 107: 56–70.
30 Barbieri PG, Mirabelli D, Somigliana A, Cavone D, Merler E. Asbestos fibre burden in the lungs of patients with mesothelioma who lived near asbestos-cement factories. Ann Occup Hyg. 2012; 56: 660–670.
31 Consonni D, De Matteis S, Pesatori AC, Bertazzi PA, Olsson AC, Kromhout H et al. Lung cancer risk among bricklayers in a pooled analysis of case-control studies. Int J Cancer J Int Cancer. 2015; 136: 360–371.
32 Kukkonen MK, Hämäläinen S, Kaleva S, Vehmas T, Huuskonen MS, Oksa P et al. Genetic susceptibility to asbestos-related fibrotic pleuropulmonary changes. Eur Respir J. 2011; 38: 672–678.
33 Dianzani I, Gibello L, Biava A, Giordano M, Bertolotti M, Betti M et al. Polymorphisms in DNA repair genes as risk factors for asbestos-related malignant mesothelioma in a general population study. Mutat Res. 2006; 599: 124–134.

55
34 Arthey S, Clarke VA. Suntanning and sun protection: a review of the psychological literature. Soc Sci Med 1982. 1995; 40: 265–274.
35 Ness AR, Frankel SJ, Gunnell DJ, Smith GD. Are we really dying for a tan? BMJ. 1999; 319: 114–116.
36 Wei H, Zhao C, Heymsfield S, Zhu S. Actinic skin damage and mortality - the first national health and nutrition examination survey epidemiologic follow-up stud. Plos One. 2011; 6: 01–09.
37 Gallagher RP, Lee TK, Bajdik CD, Borugian M. Ultraviolet radiation. Chronic Dis Can. 2010; 29 Suppl 1: 51–68.
38 Holick MF. Vitamin D deficiency. N Engl J Med. 2007; 357: 266–281.
39 Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993; 75: 843–854.
40 Bulur I, Boyuk E, Saracoglu ZN, Arik D. Perianal Basal cell carcinoma. Case Rep Dermatol. 2015; 7: 25–28.
41 Lim Y-H, Kim H, Kim JH, Bae S, Park HY, Hong Y-C. Air pollution and symptoms of depression in elderly adults. Environ Health Perspect. 2012; 120: 1023–1028.
42 Kampa M, Castanas E. Human health effects of air pollution. Environ Pollut Barking Essex 1987. 2008; 151: 362–367.
43 Resende F. Poluição atmosférica por emissão de material particulado: avaliação e controle nos canteiros de obras de edificios. 2007.
44 Araújo V, Cardoso F. Redução de impactos ambientais do canteiro de obras. Projeto Finep Habitações + sustentáveis. 2006; : 15.
45 Li Y-G, Gao X. Epidemiologic studies of particulate matter and lung cancer. Chin J Cancer. 2014; 33: 376–380.
46 Vizcaya-Ruiz A. Characterization and in vitro biological effects of concentrated particulate matter from Mexico City. Atmos Environ. 2006; 40: 583–592.
47 Senthilkumar S, Manju A, Muthuselvam P, Shalini D, Indhumathi V, Kalaiselvi K et al. Characterization and genotoxicity evaluation of particulate matter collected from industrial atmosphere in Tamil Nadu state, India. J Hazard Mater. 2014; 274: 392–398.
48 Solomon P, Sioutas C. Continuous and semicontinuous monitoring techniques for particulate matter mass and chemical components: a synthesis of findings from EPA’s Particulate Matter Supersites Program and related studies. J Air Waste Manag Assoc. 2008; 58: 164–195.
49 Schlesinger RB. The health impact of common inorganic components of fine particulate matter (PM2.5) in ambient air: a critical review. Inhal Toxicol. 2007; 19: 811–832.
50 De Oliveira Alves N, Matos Loureiro AL, Dos Santos FC, Nascimento KH, Dallacort R, de Castro Vasconcellos P et al. Genotoxicity and composition of particulate matter from biomass burning in the eastern Brazilian Amazon region. Ecotoxicol Environ Saf. 2011; 74: 1427–1433.

56
51 Pope CA. Epidemiology of fine particulate air pollution and human health: biologic mechanisms and who’s at risk? Environ Health Perspect. 2000; 108 Suppl 4: 713–723.
52 Hamra GB, Guha N, Cohen A, Laden F, Raaschou-Nielsen O, Samet JM et al. Outdoor particulate matter exposure and lung cancer: a systematic review and meta-analysis. Environ Health Perspect. 2014; 122: 906–911.
53 Delfino RJ, Staimer N, Tjoa T, Arhami M, Polidori A, Gillen DL et al. Associations of primary and secondary organic aerosols with airway and systemic inflammation in an elderly panel cohort. Epidemiol Camb Mass. 2010; 21: 892–902.
54 Ghio AJ, Carraway MS, Madden MC. Composition of air pollution particles and oxidative stress in cells, tissues, and living systems. J Toxicol Environ Health B Crit Rev. 2012; 15: 1–21.
55 Sava F, Carlsten C. Respiratory health effects of ambient air pollution: an update. Clin Chest Med. 2012; 33: 759–769.
56 Soberanes S, Gonzalez A, Urich D, Chiarella SE, Radigan KA, Osornio-Vargas A et al. Particulate matter Air Pollution induces hypermethylation of the p16 promoter Via a mitochondrial ROS-JNK-DNMT1 pathway. Sci Rep. 2012; 2: 275.
57 Dagher Z, Garçon G, Gosset P, Ledoux F, Surpateanu G, Courcot D et al. Pro-inflammatory effects of Dunkerque city air pollution particulate matter 2.5 in human epithelial lung cells (L132) in culture. J Appl Toxicol JAT. 2005; 25: 166–175.
58 Torres-Ramos YD, Montoya-Estrada A, Guzman-Grenfell AM, Mancilla-Ramirez J, Cardenas-Gonzalez B, Blanco-Jimenez S et al. Urban PM2.5 induces ROS generation and RBC damage in COPD patients. Front Biosci Elite Ed. 2011; 3: 808–817.
59 Zhao J, Gao Z, Tian Z, Xie Y, Xin F, Jiang R et al. The biological effects of individual-level PM(2.5) exposure on systemic immunity and inflammatory response in traffic policemen. Occup Environ Med. 2013; 70: 426–431.
60 Brait M, Sidransky D. Cancer epigenetics: above and beyond. Toxicol Mech Methods. 2011; 21: 275–288.
61 Sharma S, Kelly TK, Jones PA. Epigenetics in cancer. Carcinogenesis. 2010; 31: 27–36.
62 Mansuy IM, Mohanna S. Epigenetics and the human brain: where nurture meets nature. Cerebrum Dana Forum Brain Sci. 2011; 2011: 8.
63 Pogribny IP, Rusyn I. Environmental toxicants, epigenetics, and cancer. Adv Exp Med Biol. 2013; 754: 215–232.
64 Jenuwein T, Allis CD. Translating the histone code. Science. 2001; 293: 1074–1080.
65 Chen Z, Riggs AD. DNA methylation and demethylation in mammals. J Biol Chem. 2011; 286: 18347–18353.
66 Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004; 116: 281–297.
67 Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. 2014; 20: 460–469.

57
68 Marino ALF, Evangelista AF, Vieira RAC, Macedo T, Kerr LM, Abrahão-Machado LF et al. MicroRNA expression as risk biomarker of breast cancer metastasis: a pilot retrospective case-cohort study. BMC Cancer. 2014; 14: 739.
69 Elamin BK, Callegari E, Gramantieri L, Sabbioni S, Negrini M. MicroRNA response to environmental mutagens in liver. Mutat Res. 2011; 717: 67–76.
70 Vrijens K, Bollati V, Nawrot TS. MicroRNAs as Potential Signatures of Environmental Exposure or Effect: A Systematic Review. Environ Health Perspect. 2015; 123: 399–411.
71 Umer M, Herceg Z. Deciphering the epigenetic code: an overview of DNA methylation analysis methods. Antioxid Redox Signal. 2013; 18: 1972–1986.
72 Miousse IR, Chalbot M-CG, Aykin-Burns N, Wang X, Basnakian A, Kavouras IG et al. Epigenetic alterations induced by ambient particulate matter in mouse macrophages. Environ Mol Mutagen. 2014; 55: 428–435.
73 Lorincz AT. Cancer diagnostic classifiers based on quantitative DNA methylation. Expert Rev Mol Diagn. 2014; 14: 293–305.
74 Stirzaker C, Taberlay PC, Statham AL, Clark SJ. Mining cancer methylomes: prospects and challenges. Trends Genet TIG. 2014; 30: 75–84.
75 Bird A. The essentials of DNA methylation. Cell. 1992; 70: 5–8.
76 Tang J, Fang F, Miller DF, Pilrose JM, Matei D, Huang TH-M et al. Global DNA methylation profiling technologies and the ovarian cancer methylome. Methods Mol Biol Clifton NJ. 2015; 1238: 653–675.
77 Wang Y, Leung FCC. An evaluation of new criteria for CpG islands in the human genome as gene markers. Bioinforma Oxf Engl. 2004; 20: 1170–1177.
78 Irizarry RA, Ladd-Acosta C, Wen B, Wu Z, Montano C, Onyango P et al. The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores. Nat Genet. 2009; 41: 178–186.
79 Doi A, Park I-H, Wen B, Murakami P, Aryee MJ, Irizarry R et al. Differential methylation of tissue- and cancer-specific CpG island shores distinguishes human induced pluripotent stem cells, embryonic stem cells and fibroblasts. Nat Genet. 2009; 41: 1350–1353.
80 Wilson AS, Power BE, Molloy PL. DNA hypomethylation and human diseases. Biochim Biophys Acta. 2007; 1775: 138–162.
81 Baccarelli A, Bollati V. Epigenetics and environmental chemicals. Curr Opin Pediatr. 2009; 21: 243–251.
82 Hou L, Zhang X, Tarantini L, Nordio F, Bonzini M, Angelici L et al. Ambient PM exposure and DNA methylation in tumor suppressor genes: a cross-sectional study. Part Fibre Toxicol. 2011; 8: 25.
83 Yegnasubramanian S, Haffner MC, Zhang Y, Gurel B, Cornish TC, Wu Z et al. DNA hypomethylation arises later in prostate cancer progression than CpG island hypermethylation and contributes to metastatic tumor heterogeneity. Cancer Res. 2008; 68: 8954–8967.
84 Yang AS, Estécio MRH, Doshi K, Kondo Y, Tajara EH, Issa J-PJ. A simple method for estimating global DNA methylation using bisulfite PCR of repetitive DNA elements. Nucleic Acids Res. 2004; 32: e38.

58
85 Román AC, González-Rico FJ, Fernández-Salguero PM. B1-SINE retrotransposons: Establishing genomic insulatory networks. Mob Genet Elem. 2011; 1: 66–70.
86 Cowley M, Oakey RJ. Transposable elements re-wire and fine-tune the transcriptome. PLoS Genet. 2013; 9: e1003234.
87 Wu H, Chen Y, Liang J, Shi B, Wu G, Zhang Y et al. Hypomethylation-linked activation of PAX2 mediates tamoxifen-stimulated endometrial carcinogenesis. Nature. 2005; 438: 981–987.
88 Han JS, Szak ST, Boeke JD. Transcriptional disruption by the L1 retrotransposon and implications for mammalian transcriptomes. Nature. 2004; 429: 268–274.
89 Brennan K, Flanagan JM. Is there a link between genome-wide hypomethylation in blood and cancer risk? Cancer Prev Res Phila Pa. 2012; 5: 1345–1357.
90 Veeck J, Esteller M. Breast cancer epigenetics: from DNA methylation to microRNAs. J Mammary Gland Biol Neoplasia. 2010; 15: 5–17.
91 Demetriou CA, Vineis P. Carcinogenicity of ambient air pollution: use of biomarkers, lessons learnt and future directions. J Thorac Dis. 2015; 7: 67–95.
92 Baccarelli A, Wright RO, Bollati V, Tarantini L, Litonjua AA, Suh HH et al. Rapid DNA methylation changes after exposure to traffic particles. Am J Respir Crit Care Med. 2009; 179: 572–578.
93 Pavanello S, Pesatori A-C, Dioni L, Hoxha M, Bollati V, Siwinska E et al. Shorter telomere length in peripheral blood lymphocytes of workers exposed to polycyclic aromatic hydrocarbons. Carcinogenesis. 2010; 31: 216–221.
94 Belinsky SA. Gene-promoter hypermethylation as a biomarker in lung cancer. Nat Rev Cancer. 2004; 4: 707–717.
95 Anglim PP, Alonzo TA, Laird-Offringa IA. DNA methylation-based biomarkers for early detection of non-small cell lung cancer: an update. Mol Cancer. 2008; 7: 81.
96 Merlo A, Herman JG, Mao L, Lee DJ, Gabrielson E, Burger PC et al. 5’ CpG island methylation is associated with transcriptional silencing of the tumour suppressor p16/CDKN2/MTS1 in human cancers. Nat Med. 1995; 1: 686–692.
97 Umemura S, Fujimoto N, Hiraki A, Gemba K, Takigawa N, Fujiwara K et al. Aberrant promoter hypermethylation in serum DNA from patients with silicosis. Carcinogenesis. 2008; 29: 1845–1849.
98 Belinsky SA. Silencing of genes by promoter hypermethylation: key event in rodent and human lung cancer. Carcinogenesis. 2005; 26: 1481–1487.
99 Yauk C, Polyzos A, Rowan-Carroll A, Somers CM, Godschalk RW, Van Schooten FJ et al. Germ-line mutations, DNA damage, and global hypermethylation in mice exposed to particulate air pollution in an urban/industrial location. Proc Natl Acad Sci U S A. 2008; 105: 605–610.
100 Huang T, Chen X, Hong Q, Deng Z, Ma H, Xin Y et al. Meta-analyses of gene methylation and smoking behavior in non-small cell lung cancer patients. Sci Rep. 2015; 5: 8897.

59
101 Fleisher AS, Esteller M, Tamura G, Rashid A, Stine OC, Yin J et al. Hypermethylation of the hMLH1 gene promoter is associated with microsatellite instability in early human gastric neoplasia. Oncogene. 2001; 20: 329–335.
102 Watts GS, Pieper RO, Costello JF, Peng YM, Dalton WS, Futscher BW. Methylation of discrete regions of the O6-methylguanine DNA methyltransferase (MGMT) CpG island is associated with heterochromatinization of the MGMT transcription start site and silencing of the gene. Mol Cell Biol. 1997; 17: 5612–5619.
103 Wu F, Liu J, Qiu Y-L, Wang W, Zhu S-M, Sun P et al. Correlation of chromosome damage and promoter methylation status of the DNA repair genes MGMT and hMLH1 in Chinese vinyl chloride monomer (VCM)-exposed workers. Int J Occup Med Environ Health. 2013; 26: 173–182.
104 Esteller M, Catasus L, Matias-Guiu X, Mutter GL, Prat J, Baylin SB et al. hMLH1 promoter hypermethylation is an early event in human endometrial tumorigenesis. Am J Pathol. 1999; 155: 1767–1772.
105 Lahue RS, Au KG, Modrich P. DNA mismatch correction in a defined system. Science. 1989; 245: 160–164.
106 Duan H, He Z, Ma J, Zhang B, Sheng Z, Bin P et al. Global and MGMT promoter hypomethylation independently associated with genomic instability of lymphocytes in subjects exposed to high-dose polycyclic aromatic hydrocarbon. Arch Toxicol. 2013; 87: 2013–2022.
107 Hossain MB, Vahter M, Concha G, Broberg K. Environmental arsenic exposure and DNA methylation of the tumor suppressor gene p16 and the DNA repair gene MLH1: effect of arsenic metabolism and genotype. Met Integr Biometal Sci. 2012; 4: 1167–1175.
108 Guo L, Byun H-M, Zhong J, Motta V, Barupal J, Zheng Y et al. Effects of short-term exposure to inhalable particulate matter on DNA methylation of tandem repeats. Environ Mol Mutagen. 2014; 55: 322–335.
109 Kato T, Suzuki K, Okada S, Kamiyama H, Maeda T, Saito M et al. Aberrant methylation of PSD disturbs Rac1-mediated immune responses governing neutrophil chemotaxis and apoptosis in ulcerative colitis-associated carcinogenesis. Int J Oncol. 2012; 40: 942–950.
110 IBGE. Pesquisa Anual da Indústria da Construção. 2013; 23: 88.
111 Qiagen. kit Epitech Bisulfite kit. www.qiagen.com. (accessed 10 Nov2015).
112 Byun H-M, Nordio F, Coull BA, Tarantini L, Hou L, Bonzini M et al. Temporal stability of epigenetic markers: sequence characteristics and predictors of short-term DNA methylation variations. PloS One. 2012; 7: e39220.
113 Ahmadian A, Ehn M, Hober S. Pyrosequencing: history, biochemistry and future. Clin Chim Acta Int J Clin Chem. 2006; 363: 83–94.
114 Normando D, Tjäderhane L, Quintão C. A escolha do teste estatístico - um tutorial em forma de apresentação em PowerPoint*. 2010; 15: 101–106.
115 Mathelier A, Zhao X, Zhang AW, Parcy F, Worsley-Hunt R, Arenillas DJ et al. JASPAR 2014: an extensively expanded and updated open-access database of transcription factor binding profiles. Nucleic Acids Res. 2014; 42: D142–147.

60
116 Lacourt A, Pintos J, Lavoué J, Richardson L, Siemiatycki J. Lung cancer risk among workers in the construction industry: results from two case-control studies in Montreal. BMC Public Health. 2015; 15: 941.
117 Dement JM, Ringen K, Welch LS, Bingham E, Quinn P. Mortality of older construction and craft workers employed at Department of Energy (DOE) nuclear sites. Am J Ind Med. 2009; 52: 671–682.
118 Nordlund J, Bäcklin CL, Zachariadis V, Cavelier L, Dahlberg J, Öfverholm I et al. DNA methylation-based subtype prediction for pediatric acute lymphoblastic leukemia. Clin Epigenetics. 2015; 7: 11.
119 Deng D, Liu Z, Du Y. Epigenetic alterations as cancer diagnostic, prognostic, and predictive biomarkers. Adv Genet. 2010; 71: 125–176.
120 Jicai Z, Zongtao Y, Zongtao Y, Jun L, Jun L, Haiping L et al. Persistent infection of hepatitis B virus is involved in high rate of p16 methylation in hepatocellular carcinoma. Mol Carcinog. 2006; 45: 530–536.
121 Kukitsu T, Takayama T, Miyanishi K, Nobuoka A, Katsuki S, Sato Y et al. Aberrant crypt foci as precursors of the dysplasia-carcinoma sequence in patients with ulcerative colitis. Clin Cancer Res Off J Am Assoc Cancer Res. 2008; 14: 48–54.
122 Licchesi JDF, Westra WH, Hooker CM, Herman JG. Promoter hypermethylation of hallmark cancer genes in atypical adenomatous hyperplasia of the lung. Clin Cancer Res Off J Am Assoc Cancer Res. 2008; 14: 2570–2578.
123 Divine KK, Pulling LC, Marron-Terada PG, Liechty KC, Kang T, Schwartz AG et al. Multiplicity of abnormal promoter methylation in lung adenocarcinomas from smokers and never smokers. Int J Cancer J Int Cancer. 2005; 114: 400–405.
124 Kondo K, Takahashi Y, Hirose Y, Nagao T, Tsuyuguchi M, Hashimoto M et al. The reduced expression and aberrant methylation of p16(INK4a) in chromate workers with lung cancer. Lung Cancer Amst Neth. 2006; 53: 295–302.
125 Zhang A-H, Bin H-H, Pan X-L, Xi X-G. Analysis of p16 gene mutation, deletion and methylation in patients with arseniasis produced by indoor unventilated-stove coal usage in Guizhou, China. J Toxicol Environ Health A. 2007; 70: 970–975.
126 Su S, Jin Y, Zhang W, Yang L, Shen Y, Cao Y et al. Aberrant promoter methylation of p16(INK4a) and O(6)-methylguanine-DNA methyltransferase genes in workers at a Chinese uranium mine. J Occup Health. 2006; 48: 261–266.
127 Stone JE, Petes TD. Analysis of the proteins involved in the in vivo repair of base-base mismatches and four-base loops formed during meiotic recombination in the yeast Saccharomyces cerevisiae. Genetics. 2006; 173: 1223–1239.
128 Wheeler JM, Loukola A, Aaltonen LA, Mortensen NJ, Bodmer WF. The role of hypermethylation of the hMLH1 promoter region in HNPCC versus MSI+ sporadic colorectal cancers. J Med Genet. 2000; 37: 588–592.
129 Imai K, Yamamoto H. Carcinogenesis and microsatellite instability: the interrelationship between genetics and epigenetics. Carcinogenesis. 2008; 29: 673–680.

61
130 Ali AHK, Kondo K, Namura T, Senba Y, Takizawa H, Nakagawa Y et al. Aberrant DNA methylation of some tumor suppressor genes in lung cancers from workers with chromate exposure. Mol Carcinog. 2011; 50: 89–99.
131 Ding R, Jin Y, Liu X, Zhu Z, Zhang Y, Wang T et al. Characteristics of DNA methylation changes induced by traffic-related air pollution. Mutat Res Genet Toxicol Environ Mutagen. 2016; 796: 46–53.
132 Ramos MA, Cury F de P, Scapulatempo Neto C, Marques MMC, Silveira HCS. Micronucleus evaluation of exfoliated buccal epithelial cells using liquid-based cytology preparation. Acta Cytol. 2014; 58: 582–588.
133 Salas LA, Villanueva CM, Tajuddin SM, Amaral AFS, Fernandez AF, Moore LE et al. LINE-1 methylation in granulocyte DNA and trihalomethane exposure is associated with bladder cancer risk. Epigenetics Off J DNA Methylation Soc. 2014; 9: 1532–1539.
134 Terry MB, Delgado-Cruzata L, Vin-Raviv N, Wu HC, Santella RM. DNA methylation in white blood cells: association with risk factors in epidemiologic studies. Epigenetics Off J DNA Methylation Soc. 2011; 6: 828–837.
135 Andreotti G, Karami S, Pfeiffer RM, Hurwitz L, Liao LM, Weinstein SJ et al. LINE1 methylation levels associated with increased bladder cancer risk in pre-diagnostic blood DNA among US (PLCO) and European (ATBC) cohort study participants. Epigenetics. 2014; 9: 404–415.
136 Yang P, Ma J, Zhang B, Duan H, He Z, Zeng J et al. CpG site-specific hypermethylation of p16INK4α in peripheral blood lymphocytes of PAH-exposed workers. Cancer Epidemiol Biomark Prev Publ Am Assoc Cancer Res Cosponsored Am Soc Prev Oncol. 2012; 21: 182–190.
137 Zhini H, Huawei D, Biao Z. CpG site-specific RASSF1a hypermethylation is associated with occupational PAH exposure and genomic instability. 2015. doi:DOI: 10.1039/c5tx00013k.

62
Anexo A – Questionário sócio-demográfico aplicado aos grupos

63

64

65

66
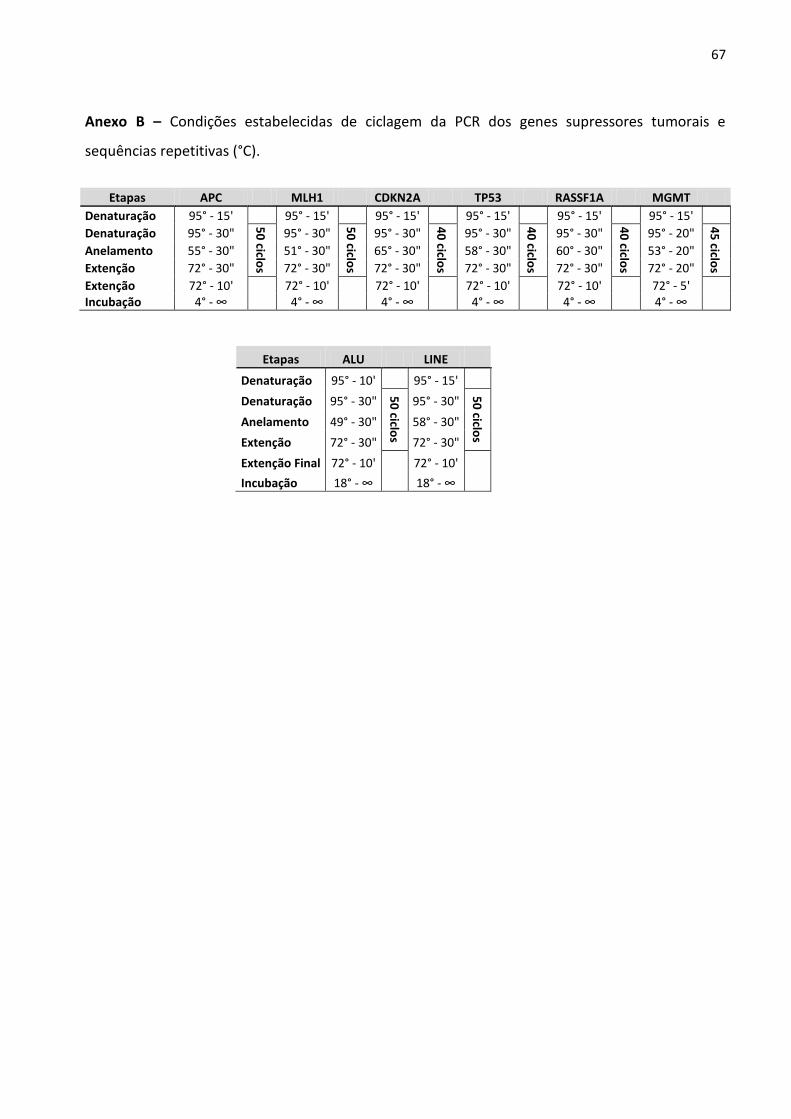
67
Anexo B – Condições estabelecidas de ciclagem da PCR dos genes supressores tumorais e
sequências repetitivas (°C).
Etapas APC MLH1 CDKN2A TP53 RASSF1A MGMT
Denaturação 95° - 15' 95° - 15' 95° - 15' 95° - 15' 95° - 15' 95° - 15'
Denaturação 95° - 30"
50
ciclos
95° - 30"
50
ciclos
95° - 30"
40
ciclos
95° - 30"
40
ciclos
95° - 30"
40
ciclos
95° - 20"
45
ciclos
Anelamento 55° - 30" 51° - 30" 65° - 30" 58° - 30" 60° - 30" 53° - 20"
Extenção 72° - 30" 72° - 30" 72° - 30" 72° - 30" 72° - 30" 72° - 20"
Extenção 72° - 10' 72° - 10' 72° - 10' 72° - 10' 72° - 10' 72° - 5' Incubação 4° - ∞ 4° - ∞ 4° - ∞ 4° - ∞ 4° - ∞ 4° - ∞
Etapas ALU LINE
Denaturação 95° - 10' 95° - 15'
Denaturação 95° - 30" 5
0 ciclo
s 95° - 30"
50
ciclos
Anelamento 49° - 30" 58° - 30"
Extenção 72° - 30" 72° - 30"
Extenção Final 72° - 10' 72° - 10'
Incubação 18° - ∞ 18° - ∞
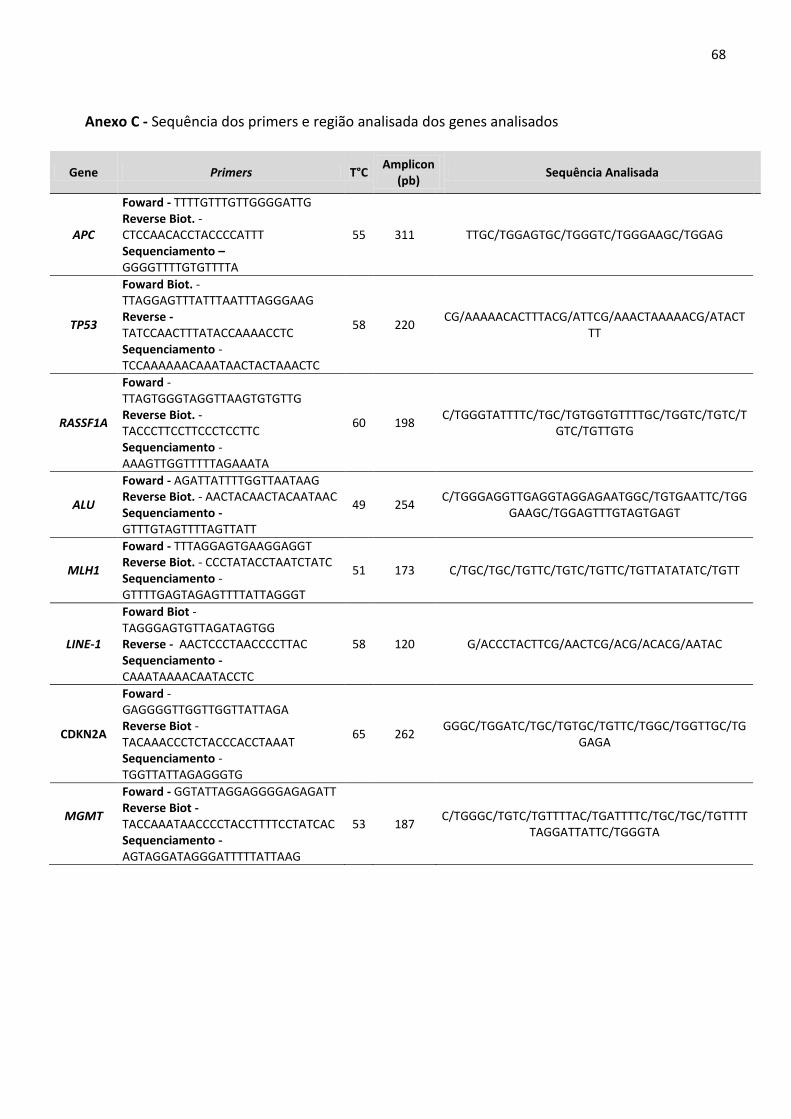
68
Anexo C - Sequência dos primers e região analisada dos genes analisados
Gene Primers T°C Amplicon
(pb) Sequência Analisada
APC
Foward - TTTTGTTTGTTGGGGATTG Reverse Biot. - CTCCAACACCTACCCCATTT Sequenciamento – GGGGTTTTGTGTTTTA
55 311 TTGC/TGGAGTGC/TGGGTC/TGGGAAGC/TGGAG
TP53
Foward Biot. - TTAGGAGTTTATTTAATTTAGGGAAG Reverse - TATCCAACTTTATACCAAAACCTC Sequenciamento - TCCAAAAAACAAATAACTACTAAACTC
58 220 CG/AAAAACACTTTACG/ATTCG/AAACTAAAAACG/ATACT
TT
RASSF1A
Foward - TTAGTGGGTAGGTTAAGTGTGTTG Reverse Biot. - TACCCTTCCTTCCCTCCTTC Sequenciamento - AAAGTTGGTTTTTAGAAATA
60 198 C/TGGGTATTTTC/TGC/TGTGGTGTTTTGC/TGGTC/TGTC/T
GTC/TGTTGTG
ALU
Foward - AGATTATTTTGGTTAATAAG Reverse Biot. - AACTACAACTACAATAAC Sequenciamento - GTTTGTAGTTTTAGTTATT
49 254 C/TGGGAGGTTGAGGTAGGAGAATGGC/TGTGAATTC/TGG
GAAGC/TGGAGTTTGTAGTGAGT
MLH1
Foward - TTTAGGAGTGAAGGAGGT Reverse Biot. - CCCTATACCTAATCTATC Sequenciamento -GTTTTGAGTAGAGTTTTATTAGGGT
51 173 C/TGC/TGC/TGTTC/TGTC/TGTTC/TGTTATATATC/TGTT
LINE-1
Foward Biot - TAGGGAGTGTTAGATAGTGG Reverse - AACTCCCTAACCCCTTAC Sequenciamento - CAAATAAAACAATACCTC
58 120 G/ACCCTACTTCG/AACTCG/ACG/ACACG/AATAC
CDKN2A
Foward - GAGGGGTTGGTTGGTTATTAGA Reverse Biot - TACAAACCCTCTACCCACCTAAAT Sequenciamento - TGGTTATTAGAGGGTG
65 262 GGGC/TGGATC/TGC/TGTGC/TGTTC/TGGC/TGGTTGC/TG
GAGA
MGMT
Foward - GGTATTAGGAGGGGAGAGATT Reverse Biot - TACCAAATAACCCCTACCTTTTCCTATCAC Sequenciamento - AGTAGGATAGGGATTTTTATTAAG
53 187 C/TGGGC/TGTC/TGTTTTAC/TGATTTTC/TGC/TGC/TGTTTT
TAGGATTATTC/TGGGTA

69
Anexo D – Sequência genômica dos genes de acordo com o banco de dados da UCSC
Gene APC
Gene TP53
Gene RASSF1A

70
Gene CDKN2A
Gene MGMT
Gene MLH1
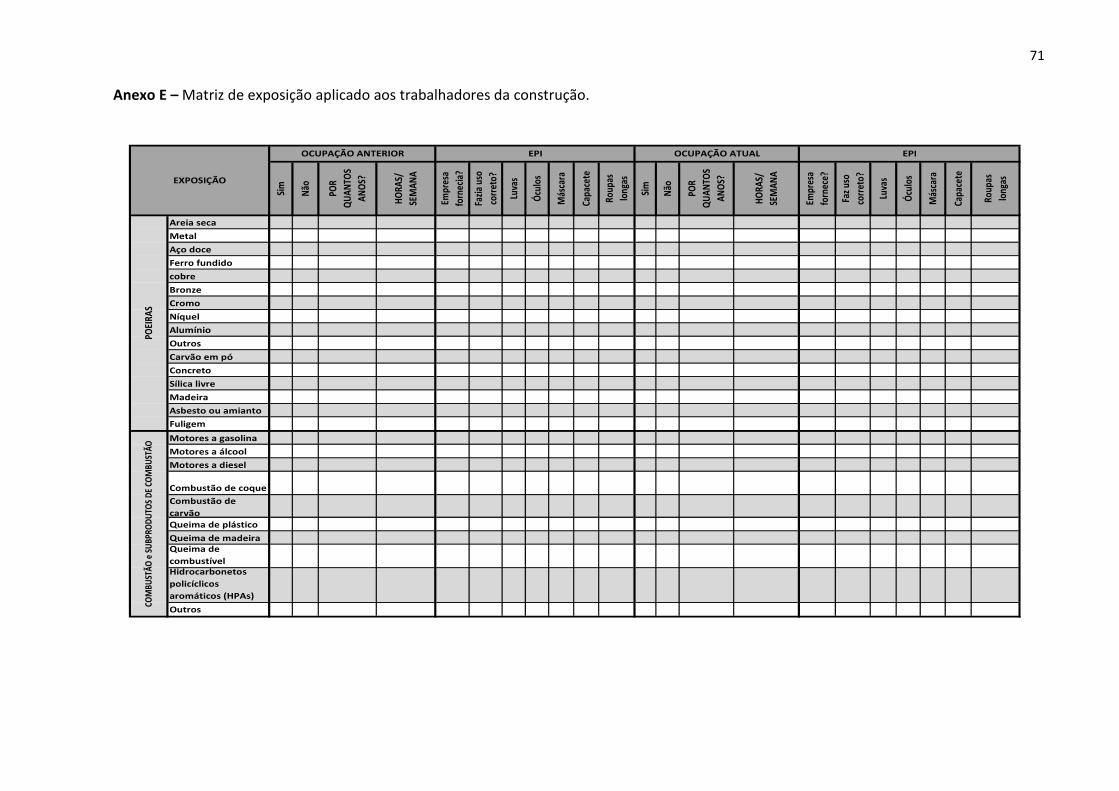
71
Anexo E – Matriz de exposição aplicado aos trabalhadores da construção.
Areia seca
Metal
Aço doce
Ferro fundido
cobre
Bronze
Cromo
Níquel
Alumínio
Outros
Carvão em pó
Concreto
Sílica livre
Madeira
Asbesto ou amianto
Fuligem
Motores a gasolina
Motores a álcool
Motores a diesel
Combustão de coque
Combustão de
carvão
Queima de plástico
Queima de madeiraQueima de
combustívelHidrocarbonetos
policíclicos
aromáticos (HPAs)
Outros
HO
RAS/
SEM
AN
A
EXPOSIÇÃO
Luva
s
Más
cara
Capa
cete
Roup
as
long
as
POEI
RAS
COM
BUSTÃ
O e
SU
BPRO
DU
TOS
DE
COM
BUSTÃO
Sim
Não
OCUPAÇÃO ANTERIOR
POR
QU
AN
TOS
AN
OS?
HO
RAS/
SEM
AN
A
Capa
cete
Roup
as
long
as
OCUPAÇÃO ATUAL
Não
EPI
Empr
esa
forn
ecia
?
Fazi
a us
o
corr
eto?
Ócu
los
EPI
Empr
esa
forn
ece?
Faz
uso
corr
eto?
Luva
s
Ócu
los
Más
cara
Sim
POR
QU
AN
TOS
AN
OS?
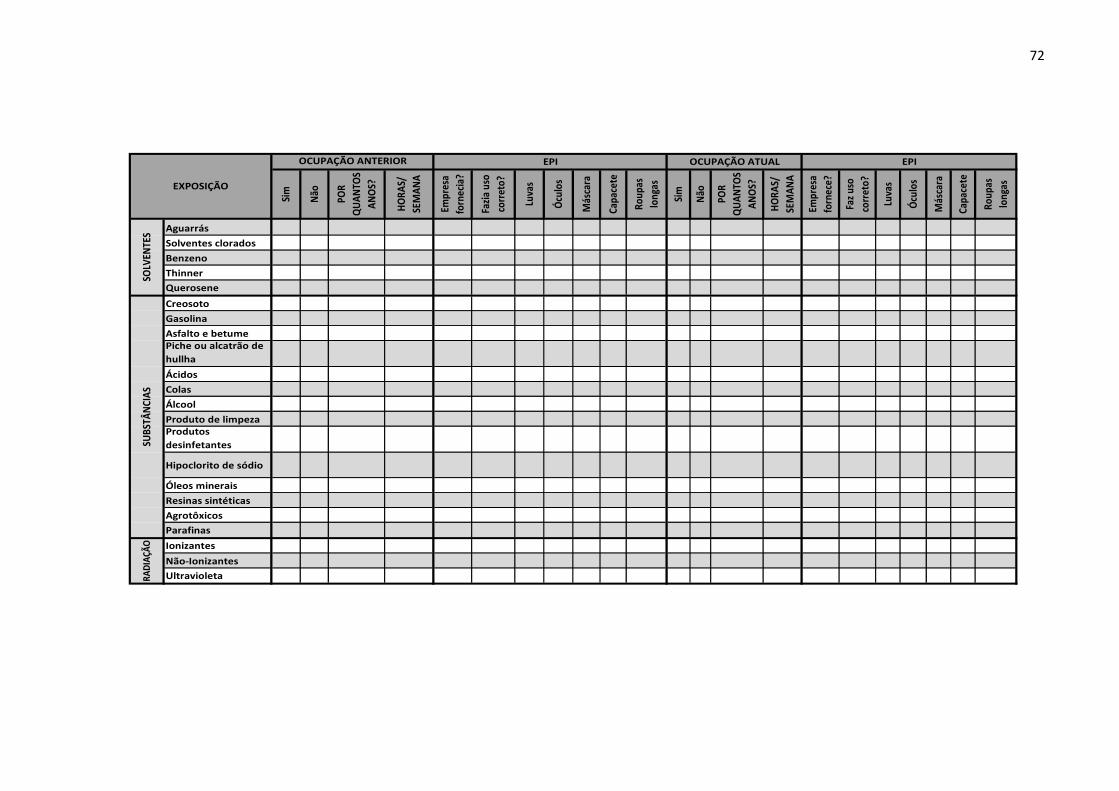
72
Aguarrás
Solventes clorados
Benzeno
Thinner
Querosene
Creosoto
Gasolina
Asfalto e betumePiche ou alcatrão de
hullha
Ácidos
Colas
Álcool
Produto de limpezaProdutos
desinfetantes
Hipoclorito de sódio
Óleos minerais
Resinas sintéticas
Agrotôxicos
Parafinas
Ionizantes
Não-Ionizantes
Ultravioleta
Sim
Não
POR
QU
AN
TOS
AN
OS?
HO
RA
S/
SEM
AN
A
Empr
esa
forn
ecia
?
Más
cara
Capa
cete
Rou
pas
long
as
Sim
Não
POR
QU
AN
TOS
AN
OS?
HO
RA
S/
SEM
AN
A
Empr
esa
forn
ece?
Faz
uso
corr
eto?
RAD
IAÇÃ
OSO
LVEN
TES
SUBS
TÂN
CIA
S
Luva
s
Ócu
los
Fazi
a us
o
corr
eto?
Luva
s
Ócu
los
Más
cara
Capa
cete
Rou
pas
long
asEXPOSIÇÃO
OCUPAÇÃO ANTERIOR EPI OCUPAÇÃO ATUAL EPI

73
Anexo F – Parecer consubstanciado do CEP

74

75

76

77

78

79

80
Anexo G – Análise dos motifs das regiões promotoras dos genes CDKN2A, MLH1 e APC
utilizando o banco de dados JASPAR.
Gene APC

81
Gene MLH1

82
Gene CDKN2A





























