Embed Size (px)
Citation preview

JOURNAL OF CLINICAL MICROBIOLOGY, Sept. 1991, p. 1928-1933 Vol. 29, No. 90095-1137/91/091928-06$02.00/0Copyright C) 1991, American Society for Microbiology
Isolation and Continuous Culture of Neorickettsia helminthoecain a Macrophage Cell Linet
YASUKO RIKIHISA,l* HAROLD STILLS,2 AND GARY ZIMMERMAN3Department of Veterinary Pathobiology' and Department of Veterinary Preventive Medicine,2 College of Veterinary
Medicine, The Ohio State University, Columbus, Ohio 43210, and College of Veterinary Medicine,Oregon State University, Corvallis, Oregon 973313
Received 28 January 1991/Accepted 11 June 1991
Experimental evidence is presented supporting the development of a system for the isolation and propagationof a Neorickettsia sp. in a continuous canine macrophage cell line (DH82). To isolate a Neorickettsia sp.pathogenic to the canine species, three naive dogs were fed metacercaria-encysted kidneys of salmon caught ina river where infection of metacercariae with Neorickettsia helminthoeca has been circumstantially known fordecades. Clinically, the classic course of salmon poisoning disease developed in all of the dogs. Parasitemiabegan on day 8 to 11 postinfection, when the dogs developed a febrile peak, and continued until euthanasia. Atnecropsy, characteristic gross and microscopic lesions of the disease were present. A Neorickettsia sp. was alsoisolated from liver and spleen samples of these animals. The isolates have been continuously propagated andpassed in DH82 cells for more than 6 months. Electron microscopic examination confirmed that the rickettsialorganisms multiplied in the membrane-bound compartment of DH82 cells and that they morphologicallyclosely resembled rickettsia belonging to the genus Ehrlichia. An indirect fluorescent antibody test usingNeorickettsia organisms cultured in DH82 cells showed that all dogs seroconverted 13 to 15 days postinfection.Finally, inoculation of the cell-cultured Neorickettsia organisms into a naive dog reproduced clinically typicalsalmon poisoning disease which was of greater severity and had a more rapid time course than that in the dogsfrom which the original isolation was made. On the basis of the clinical and pathologic responses of the dogsin our study, we believe that virulent N. helminthoeca was isolated and cultured in a continuous cell line.
Neorickettsia helminthoeca is a minute, intracytoplasmic,rickettsial organism found in macrophages of Canidae withsalmon poisoning disease. The organism varies from coccoidto crescentic, is nonmotile, stains readily with Giemsa stain,and is gram negative (5, 8, 11). N. helminthoeca is classifiedin the tribe Ehrlichieae in the latest Bergey's Manual ofSystematic Bacteriology (11). The disease was recognized inthe early 19th century by white settlers in the PacificNorthwest and named salmon poisoning disease becausedogs became sick after eating salmon (8).N. helminthoeca is a rickettsia parasitic to a fluke, Nano-
phyetus salmincola, which has a complicated life cycleinvolving both snails (Oxytrema silicula) and salmonid fishas intermediate hosts and carnivores as the final host.Infection of dogs with N. helminthoeca occurs followingingestion of salmonid fish which harbor metacercariae con-taining Neorickettsia organisms (5, 8). N. helminthoeca isthe first and only obligatory helminth-borne pathogen thathas been discovered in the bacterial field (8). Nanophyetussalmincola harbors Neorickettsia organisms throughout itslife cycle stages from egg to adult (5, 8, 11). The evidence forresidence of N. helminthoeca in Nanophyetus salmincola is,however, based entirely on transmission experiments, andthe organism has not been directly confirmed by eitherculture or microscopic evaluation in any developmentalstages of Nanophyetus salmincola.Salmon poisoning disease in dogs is characterized by
fever, depression, anorexia, weight loss, dehydration, vom-iting, and watery, often bloody diarrhea. Mortality exceeds
* Corresponding author.t Oregon Agricultural Experiment Station technical paper no.
9445.
90% in untreated cases (2, 5, 8). Intestinal hemorrhage andinflammation are the most characteristic lesions and aremore severe than those caused by the fluke alone (2, 5, 8).Generalized lymph node enlargement due to marked infiltra-tion of macrophages is accompanied by severe depletion ofsmall lymphocytes and a loss of germinal centers (2, 4, 5, 8).Splenic follicular central hemorrhage and necrosis and oblit-eration of thymic architecture by macrophage infiltration arecommon (4, 5). Coccoid bodies are found in the macro-phages in most of the lymph nodes by both light and electronmicroscopy (4, 5, 8). Although the disease can be transmit-ted by injection of the blood or lymph node homogenatefrom infected to susceptible dogs, the organism has neverbeen seen in blood smears (8).N. helminthoeca has not been grown in ordinary bacteri-
ologic media, yolk sac, fibroblasts, or small laboratoryrodents. Only two studies are known that describe in vitroculture of the Neorickettsia sp. (1, 3). The organism wasisolated from leukocyte-enriched plasma of experimentallyinfected dogs at the acute stage of infection. The organismwas maintained for up to 2 months by weekly passage inprimary canine blood monocyte cultures in 2 ml of L-15medium containing 40% canine serum (3) or in Eagle'sminimal essential medium containing 20% canine serum and0.1 mM L-glutamine (1). To the authors' knowledge, how-ever, Neorickettsia cultures do not currently exist in anylaboratory and the organism has never been cultured in acontinuous cell line. Difficulties in culturing a sufficientquantity of this organism by using primary canine bloodmonocyte cultures and media containing canine serum havetherefore limited studies on salmon poisoning disease andNeorickettsia species to the clinical, gross, and histopatho-logic levels. The antigenic, biochemical, molecular aspectsof Neorickettsia spp. and the disease pathogenesis are
1928
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

ISOLATION AND CONTINUOUS CULTURE OF N. HELMINTHOECA 1929
unknown. Furthermore, there are currently no antigensreadily available to serodiagnose Neorickettsia infection.The purpose of this study was to isolate and culture
Neorickettsia organisms in an established canine macro-phage cell line. Since no method for detection of Neorick-ettsia infection in the fluke has been described and ourpurpose was to isolate virulent N. helminthoeca, we feddogs with metacercaria-encysted salmon caught in a riverwhere infection of metacercariae with Neorickettsia spp.had been circumstantially known for decades.
This study reports the isolation and cultivation of Neorick-ettsia organisms in a continuous cell line from the blood,liver, and spleen of dogs exhibiting classical clinical signs ofsalmon poisoning disease and characteristic gross and mi-croscopic lesions at necropsy. The time course and severityof parasitemia and antibody titers are for the first timedocumented. Lastly, the reproduction of the disease byusing cell-cultured Neorickettsia organisms was docu-mented.
MATERIALS AND METHODS
Dogs. Four conditioned adult female mongrel dogs werepurchased from Butler Farms, Clyde, N.Y. Salmon poison-ing disease has never been reported east of the Rockies orcontinental divide; therefore, dogs originating from NewYork could be considered free of previous exposure to theagent and thus appropriate experimental subjects. Thesedogs had been vaccinated for canine distemper, infectiouscanine hepatitis, leptospira, parainfluenza, parvovirus, ca-nine adenovirus II, rabies, and bordetella. The dogs hadtested negative for Brucella canis and Dirofilaria immitis andwere treated with a series of antihelminthics consisting ofivermectin (Merck & Co., Inc., Rahway, N.J.), pyrantelpamoate (Strongid-T; Pfizer, Inc., New York, N.Y.), andpraziquantel (Droncit; Haver; Mobay Corp., Shawnee,Kans.) 1 to 2 months before use.
Source of infectious Neorickettsia organisms. Five 2- to 3-lb(ca. 0.1- to 1-kg) salmon from the Roaring River State FishHatchery, near Lacomb, Oreg., were obtained through theOregon Department of Fisheries and Wildlife. In westernOregon, most salmonid fish of this size category are parasi-tized by Nanophyetus salmincola and carry the etiologicagent of the disease. Fluke metacercariae, presumed to beNanophyetus salmincola, were visually identified (x 160magnification) in kidney tissue prior to shipping the fish onice, by express mail, to the primary author's laboratory. Thekidneys were removed from the fish upon arrival, and thepresence of viable metacercariae containing moving juvenileflukes within the cysts was again confirmed. Selection ofkidney tissue was based on tendency of the metacercariae tobe concentrated in kidney tissue of fish, the ease of recov-ering this tissue, and the willingness of dogs to ingest thisorgan.
Infection of dogs. Three dogs were fed approximately oneand a half fluke-infested salmon kidneys per dog. Rectaltemperature, pulse rate, respiratory rate, appetite, attitude(depression), and consistency of feces (diarrhea with orwithout blood) were recorded daily. Approximately 10 ml ofheparinized blood was obtained from the cephalic vein everyother day to isolate neorickettsia from the blood. The dogswere euthanatized at the acute stage of infection, and thetissue samples from the mesenteric lymph node, liver, andspleen were aseptically resected for neorickettsia isolation.Tissue specimens from the major organs were fixed in 10%neutral buffered formalin and processed for light micro-
scopic observation after hematoxylin and eosin staining.Lymph node samples were prepared for electron micros-copy.
Isolation and culture of Neorickettsia organisms. Heparin-ized blood samples (approximately 10 ml) from each dogwere overlaid on 10 ml of Histopaque 1077 (Sigma ChemicalCo., St. Louis, Mo.) and centrifuged at 800 x g for 15 min atroom temperature. The interface containing leukocytes wascollected and centrifuged at 1,000 x g for 5 min. The pelletwas suspended in Hanks balanced salt solution and washedby centrifugation two further times. These dog peripheralblood leukocytes were overlaid on a semiconfluent mono-layer of canine macrophage DH82 cells (12) in Dulbecco'smodified minimum essential medium (GIBCO Laboratories,Grand Island, N.Y.) containing 10% fetal bovine serum(Flow Laboratories, Inc., McLean, Va.) and 2 mM L-glu-tamine (GIBCO) and cultured in 5% C02-95% air at 37°C.The original seed of DH82 cells had been kindly provided byM. Wellman at The Ohio State University, and a relativelymore adherent and larger cell population was selected afterseveral passages in the senior author's laboratory beforebeing used for this study. The liver, spleen, and mesentericlymph node samples were immediately soaked in Hanksbalanced salt solution after they were removed. Thesetissues were smashed with the blunt end of a 10-ml syringethrough a Cellector (Thomas Scientific, Swedesboro, N.J.)to release the cells. The cell suspension was overlaid onHistopaque 1077, and the interface was collected, washed,overlaid on a monolayer of DH82 cells and cultured at 37°Cin 5% C02-95% air. DH82 cells were dissociated from thebottom of the flask by tapping the flask, and an aliquot (about0.2 ml) was centrifuged onto a glass slide in a Cytospin 2cytocentrifuge (Shandon, Inc., Pittsburgh, Pa.) every 2 days.The cells were stained with Diff-Quik stain (Baxter ScientificProducts, Obetz, Ohio) and examined for the presence ofcoccobacillary bodies by light microscopy. Positive cultureswere confirmed by indirect fluorescent antibody (IFA) label-ing using known positive canine anti-N. helminthoeca se-rum. These cultures were weekly passed for up to 1 monthand 6 months for microorganism-negative and microorgan-ism-positive cultures, respectively.IFA tests of the dog sera. The cells infected with Neorick-
ettsia organisms (90% infection) were suspended in Hanksbalanced salt solution and were air dried and acetone fixedonto Teflon (E. I. du Pont de Nemours & Co., Inc.,Wilmington, Del.)-coated 12-well slides (Cell-Line Associ-ates, Newfield, N.J.) at 104 cells per well. Serial twofolddilutions of test sera in phosphate-buffered saline, pH 7.4(PBS), starting at a 1:20 dilution, were made in 96-wellmicrotiter plates, and a 100-,ul volume and 10-,lI volume ofeach dilution were applied to a well of antigen-coated IFAslides. The slides were incubated in a humidified 37°Cincubator for 30 min and rinsed three times in PBS contain-ing 0.002% Tween 20. After the slides were blotted to neardryness, 10 pAl of fluorescein-conjugated goat anti-dog immu-noglobulin (United States Biochemical Corp., Cleveland,Ohio) diluted at 1:200 in PBS was added to each well. Theslides were incubated, washed, and counterstained withEvans blue (Sigma Chemical Co.), and a small drop ofmounting fluid (70% glycerol in PBS) was placed in eachwell. Coverslips were added, and the slides were examinedwith a fluorescence microscope.
Electron microscopy. Infected DH82 cells and lymph nodespecimens of infected dogs were fixed in a mixture of 2.5%paraformaldehyde, 5% glutaraldehyde, and 0.03% trinitro-phenol in 0.41 M cacodylate buffer (pH 7.4) and postfixed in
VOL. 29, 1991
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

1930 RIKIHISA ET AL.
o 40.6 Dog 10
* Dog 3400
I 2
0~~~~~~~~~~~~~~Dog 2 diarrhea Do0
37.8 _Dog 3 diarrheadjr
0 5 10 15 20Days Post Infection
FIG. 1. Clinical signs of dogs infected with N. helminthoeca. Three dogs were infected with N. helminthoeca by feeding them portionsof Nanophyetus salmincola metacercaria-infested salmon kidneys on day 0. Eu, euthanasia.
1% osmium tetroxide in 1.5% potassium ferrocyanide. Afterblock staining in 1% uranyl acetate in maleate buffer (pH5.2), cells were dehydrated in a graded series of ethanol andpropylene oxide and embedded in Poly/Bed 812 (Poly-sciences, Inc., Warrington, Pa.). Thin sections (60 to 90 nm)were cut and stained with uranyl acetate and lead citrate andexamined with a Philips 300 electron microscope.
Inoculation of a dog with cell-cultured N. helminthoeca. Oneof four naive dogs described above was intravenously inoc-ulated with 2 x 107 N. helminthoeca-infected DH82 cells.Clinical signs, parasitemia, and immunoglobulin antibodytiter were monitored as described above. At necropsy, tissuespecimens of the major organs were fixed for both light andelectron microscopy as described above.
RESULTS
Clinical features of dogs. The clinical features of theexperimentally infected dogs were consistent with thosedescribed previously for salmon poisoning disease (2, 5, 8).All infected dogs became febrile (39.8 to 40.9°C) beginningapproximately 7 days after eating fluke-encysted kidney(postinfection) for a duration of approximately 5 days beforetheir temperatures sharply dropped to below the normallevel (Fig. 1). The dogs developed persistent depression,anorexia, and severe watery diarrhea beginning on days 8(dog 3), 10 (dog 2), and 15 (dog 1) postinfection. Dehydrationand weight loss occurred in all three dogs, and two of thedogs had a thick yellowish conjunctival discharge and serousnasal discharge.Necropsy lesions. The pathologic changes observed in
experimentally infected dogs were compatible with thosepreviously reported (2-4, 8). Briefly, gross necropsy lesionsincluded enlarged lymph nodes, especially mesenteric lymphnodes, and splenomegaly. Microscopic lesions included hy-perplasia of lymph node reticuloendothelial cells with small-lymphocyte depletion. Intestinal lesions were limited tolymphoid hyperplasia, macrophage infiltration of the laminapropria, and the presence of Nanophyetus salmincola in thesmall intestine of two of the dogs. Microscopically, blue- andlilac-stained small cocci were observed in the smears of thehomogenate of the mesenteric lymph nodes of dogs 2 and 3after Diff-Quik staining, as well by IFA labeling and byelectron microscopy. In other tissues stained with Diff-Quik,microorganisms were not identifiable.
Isolation of Neorickettsia organisms. Neorickettsia organ-isms were consistently isolated and continuously propagatedin DH82 cells from the blood monocytes of all three dogs.Light microscopic examination of Diff-Quik-stained bloodfilms consistently failed to detect Neorickettsia organisms inmonocytes. If we define parasitemia as positive isolation andculture of the organism from the blood by using this culturesystem within 30 days of in vitro culture, two dogs becameparasitemic on day 8 postinfection, with the remaining dogbecoming parasitemic on day 11 postinfection (Fig. 2).Detectable parasitemia persisted until euthanasia. Incuba-tion periods before the DH82 cell cultures were identified aspositive decreased as the disease progressed.
Cocultivation of monocyte-rich fractions of lymph nodes,spleen, and liver samples with DH82 cells resulted inNeorickettsia isolation. Once isolated, Neorickettsia organ-isms from the liver and spleen but not from the lymph nodecontinuously multiplied in DH82 cells (Table 1). No viruses,other bacteria, or parasites were detected in any cell-cul-tured specimens or light and electron microscope specimens.Morphology of Neorickettsia organisms in culture. Neorick-
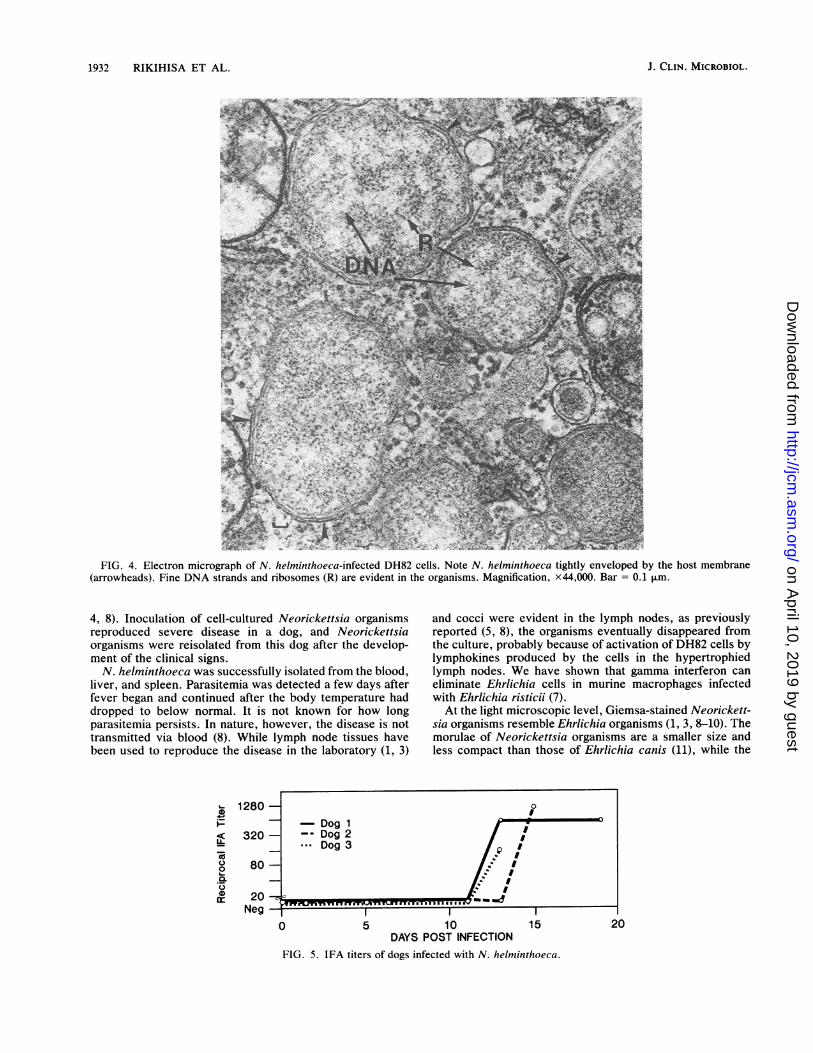
ettsia organisms were distributed in DH82 cells as individualcocci as well as morulae of various sizes, and more than 100organisms were often found per cell (Fig. 3). The presence ofintact organisms was confirmed by electron microscopy. Byelectron microscopy, the organisms were very tightly envel-oped by the host membrane (Fig. 4). Two layers (outer andinner) of membrane, ribosomes, and fine DNA strands wereevident in the organism (Fig. 4). There was no thickening ofthe inner or outer leaflet of the outer membrane (Fig. 4).IFA titration of dog sera. All three dogs were seronegative
at the time of infection (titer, <1:20) and showed a greater
TABLE 1. Isolation of Neorickettsia organisms from organs ofdogs orally infected with metacercaria
Culture result (day)" for dog no.:Organ
1 2 3
Lymph node - (0) + (0-1) + (0-8)- (after 3)b - (after 10)b
Liver + (after 17) + (after 14) + (after 26)Spleen + (after 10) + (after 7) + (after 23)
' Days in culture after euthanasia; day 0, day of euthanasia.b Neorickettsia organisms died.
J. CLIN. MICROBIOL.
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

ISOLATION AND CONTINUOUS CULTURE OF N. HELMINTHOECA 1931
5C0° 1000
00
= 20
0._0 oas CDo0zZ 30o
Neg0 5 10 15
Days Post Infection20
FIG. 2. Neorickettsia isolation from the peripheral blood of three dogs infected with N. helminthoeca. Blood was collected every otherday from each dog, and monocyte fractions were overlaid on DH82 cell monolayers. The infectivity of cultured cells was evaluated everyother day after staining cytospin-prepared cells with Diff-Quik. For each blood monocyte culture from each dog, the first day when the culturebecame clearly positive with Neorickettsia infection was recorded. The culture period (days) required for positive Neorickettsia identificationfor each blood collection is indicated on the vertical axis.
than fourfold rise in immunoglobulin G IFA titer (1:160 to1:1,280) at day 13 to 15 postinfection (Fig. 5).Reproduction of the disease with cell-cultured Neorickettsia
organisms. A dog inoculated with cell-cultured N. hel-minthoeca developed severe clinical signs of salmon poison-ing disease. The dog became febrile beginning 3 days afterinoculation. Neorickettsia organisms were reisolated fromthe blood of this dog starting at day 2 postinoculation.Necropsy lesions were consistent with, but much moresevere than, those seen previously, including generalized
lymph adenopathy, hemorrhages in the small intestinal mu-cosa, and hyperemia in the brain.
DISCUSSION
This study demonstrated that N. helminthoeca can beisolated from dogs exhibiting the clinical signs of salmonpoisoning disease and continuously propagated in a caninemacrophage cell line. Clinical signs and gross and histo-pathologic lesions were consistent with previous reports (2,
FIG. 3. Light micrograph of N. helminthoeca-infected DH82 cells. The organisms are seen in the cytoplasm as small morulae (arrows),as individual organisms (arrowheads), and outside the host cells. Diff-Quik stain. Magnification, x1,500. Bar = 10 ,um.
VOL. 29, 1991
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
1932 RIKIHISA ET AL.
FIG. 4. Electron micrograph of N. helminthoeca-infected DH82 cells. Note N. helminthoeca tightly enveloped by the host membrane(arrowheads). Fine DNA strands and ribosomes (R) are evident in the organisms. Magnification, x44,000. Bar = 0.1 .m.
4, 8). Inoculation of cell-cultured Neorickettsia organismsreproduced severe disease in a dog, and Neorickettsiaorganisms were reisolated from this dog after the develop-ment of the clinical signs.N. helminthoeca was successfully isolated from the blood,
liver, and spleen. Parasitemia was detected a few days afterfever began and continued after the body temperature haddropped to below normal. It is not known for how longparasitemia persists. In nature, however, the disease is nottransmitted via blood (8). While lymph node tissues havebeen used to reproduce the disease in the laboratory (1, 3)
L-
a.'00
1280
320
80
20Neg
and cocci were evident in the lymph nodes, as previouslyreported (5, 8), the organisms eventually disappeared fromthe culture, probably because of activation of DH82 cells bylymphokines produced by the cells in the hypertrophiedlymph nodes. We have shown that gamma interferon caneliminate Ehrlichia cells in murine macrophages infectedwith Ehrlichia risticii (7).At the light microscopic level, Giemsa-stained Neorickett-
sia organisms resemble Ehrlichia organisms (1, 3, 8-10). Themorulae of Neorickettsia organisms are a smaller size andless compact than those of Ehrlichia canis (11), while the
0 5 10 15 20DAYS POST INFECTION
FIG. 5. IFA titers of dogs infected with N. helminthoeca.
J. CLIN. MICROBIOL.
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

ISOLATION AND CONTINUOUS CULTURE OF N. HELMINTHOECA 1933
individual forms are similar to those of E. risticii (9, 10). Alack of antigenic cross-reactivity, however, has been previ-ously reported between a Neorickettsia sp. and Ehrlichiasennetsu (6) and E. canis (1) by direct fluorescent antibodytesting. These results should be reexamined with tissue-cultured Neorickettsia organisms and specific antisera with ahigher titer, since one study used infected lymph node tissueas the antigen and antisera with a titer of less than 1:32 (6).In the other study, the antibody titer was not reported (usedat 1:3 dilution) (1).By using tissue-cultured Neorickettsia organisms as the
antigen, an IFA technique was developed and temporaldevelopment of immunoglobulin G antibody against aNeorickettsia sp. was determined. The dogs seroconverted13 to 15 days after infection. Thus, Neorickettsia organismscultured in DH82 cells may be useful for serodiagnosis andserological determination of the disease distribution.
Cell-cultured Neorickettsia organisms induced diseasewith much greater severity than that caused by the naturallyinfected fluke. Very interestingly, intravenous inoculation ofNeorickettsia organisms induced severe hemorrhagesthroughout the small intestine. Thus, the intestinal lesionsare produced not because of the natural portal of entry ofNeorickettsia organisms, i.e., the oral route, but probablybecause of the intestinal tissue trophism of Neorickettsiaorganisms.
In addition to N. helminthoeca, Neorickettsia elokomin-ica, or Elokomin fluke fever agent, has been describedpreviously (3-5). Both organisms have been cultured inprimary dog blood monocytes by continuous passages for upto 2 months (3), and no morphological differences have beennoted (3) at either the light or electron microscopic level.The organisms were not cross protective in dogs (4), but bya direct fluorescent antibody test, slight cross-reaction wasseen by using antisera at a 1:1 dilution (6). Without moredata to judge by, some researchers believe that N. eloko-minica is an isolate or a strain of N. helminthoeca (5).Elokomin fluke fever was originally reported in dogs alongthe Elokomin River in the state of Washington (5). Clinicalsigns of N. elokominica in dogs are much milder than thoseof N. helminthoeca (4, 5). With N. helminthoeca infections,dogs develop a fever with more of a temperature spike,whereas Elokomin fluke fever produces a longer plateau-type fever (5). The lymph node pathology is different be-tween two Ehrlichia species. Follicles are active in N.elokominica infection, but in N. helminthoeca infection,follicles disappear accompanied by histiocytosis (4). On thebasis of the severe clinical signs in our four dogs, sharppeak-type fever, lymph node histopathology, and the factthat the salmon had been caught in an area in the state ofOregon where Elokomin fluke fever disease has not beenrecognized, we believe that the organism we cultured isN. helminthoeca, not Elokomin fluke fever agent. Furtherisolations and studies at the antigenic and genetic levels mayclarify the still-elusive agent, N. elokominica, in the future.The success in propagation in continuous cell line could lead
to clarification of the phylogenetic relationship of Neorick-ettsia spp. to other members of the tribe Ehrlichieae and thefamily Rickettsiaceae in the future. In a practical aspect, inthe future a vaccine may be developed for salmon poisoningdisease by using a tissue-cultured organism.
ACKNOWLEDGMENTS
We thank James Laird and Holly Ferrell for technical support inculturing Neorickettsia organisms and Saverit Capuano and ThomasPhilips for technical help in collecting blood from the dogs andkeeping clinical records. We also appreciate the technical assistanceof Kathy Dailey in sample collecting at necropsy.
This study was supported in part by the State of Ohio CanineResearch Grant. Oregon efforts were supported in part by fundsfrom the Northwest Canine Research Foundation.
REFERENCES1. Brown, J. L., D. L. Huxsoll, M. Ristic, and P. K. Hildebrandt.
1972. In vitro cultivation of Neorickettsia helminthoeca, thecausative agent of salmon poisoning disease. Am. J. Vet. Res.33:1695-1700.
2. Cordy, D. R., and J. R. Gorham. 1950. The pathology andetiology of salmon disease in the dog and fox. Am. J. Pathol.26:617-637.
3. Frank, D. W., T. C. McGuire, and J. R. Gorham. 1974.Cultivation of two species of Neorickettsia in canine mono-cytes. J. Infect. Dis. 129:257-262.
4. Frank, D. W., T. C. McGuire, J. R. Gorham, and R. K. Farrell.1974. Lymphoreticular lesions of canine neorickettsiosis. J.Infect. Dis. 129:163-171.
5. Gorham, J. R., and W. J. Foreyt. 1984. Salmon poisoningdisease, p. 538-544. In C. E. Greene (ed.), Clinical microbiologyand infectious disease of the dog and cat. The W. B. SaundersCo., Philadelphia.
6. Kitao, T., R. K. Farrell, and T. Fukuda. 1973. Differentiation ofsalmon poisoning disease and Elokomin fluke fever: fluorescentantibody studies with Rickettsia sennetsu. Am. J. Vet. Res.34:927-928.
7. Park, J., and Y. Rikihisa. 1990. Inhibition of Ehrlichia risticii inmurine peritoneal macrophage by calcium ionophore, gamma-interferon, and concanavalin A, abstr. 1335, p. A496. ProgramAbstr. 74th Annu. Meet. Fed. Am. Soc. Exp. Biol. Federationof American Societies for Experimental Biology, Washington,D.C.
8. Philip, C. B., W. J. Hadlow, and L. E. Hughes. 1954. Studies onsalmon poisoning disease of canines. 1. The rickettsial relation-ships and pathogenicity of Neorickettsia helminthoeca. Exp.Parasitol. 3:336-350.
9. Rikihisa, Y., and B. D. Perry. 1985. Causative ehrlichial organ-isms in Potomac horse fever. Infect. Immun. 49:513-517.
10. Rikihisa, Y., B. D. Perry, and D. 0. Cordes. 1985. Ultrastruc-tural study of ehrlichial organisms in the large colon of poniesinfected with Potomac horse fever. Infect. Immun. 49:505-512.
11. Ristic, M., and D. Huxsoll. 1984. Tribe II. Ehrlichieae, p.704-711. In N. R. Krieg and J. G. Holt (ed.), Bergey's manualof systematic bacteriology, vol. 1. The Williams & Wilkins Co.,Baltimore.
12. Wellman, M. L., S. Krakowka, R. M. Jacobs, and G. J. Kociba.1988. A macrophage-monocyte cell line from a dog with malig-nant histiocytosis. In Vitro Cell. Dev. Biol. 24:223-229.
VOL. 29, 1991
on April 10, 2019 by guest
http://jcm.asm
.org/D
ownloaded from





























