Embed Size (px)
Citation preview

The EMBO Journal vol.7 no.4 pp.1031 -1039, 1988
Isolation of chicken major histocompatibility complexclass 11 (B-L) chain sequences: comparison withmammalian chains and expression in lymphoid organs
Yves Bourlet', Ghislaine Behar,Franpois Guillemot, Nancy Frechin,Alain Billault, Anne-Marie Chausse,Rima Zoorob and Charles AuffrayInstitut d'Embryologie du CNRS et du College de France, 49 bisAvenue de la Belle Gabrielle, 94130 Nogent sur Mamne, France
'In memoriam
Communicated by N.Le Douarin
By cross-hybridization in low stringency conditions, us-ing a probe derived from an HLA-DQ,B cDNA clone, wehave isolated several chicken genomic DNA clones. Theseclones were mapped to the major histocompatibility com-plex (MIHC) of the chick (B complex) by virtue of theirability to detect restriction enzyme length polymorphismsbetween congenic lines of chicken. Evidence was obtain-ed for the presence of at least three B-L,B genes in thechicken genome. The B-Lj genes are transcribedspecifically in tissues containing cells of the B lymphocyteand myeloid lineages and expressing the B-L antigens.Exons encoding the (31, (2 and transmembrane domainsof a B-L3 chain have been identified with 63, 66 and 62%similarity with the HLA-DQ,B sequence. This first isola-tion of an MHC class II gene outside of the mammalianclass provides insight into the evolution of MHC genesbased on the comparison of avian and mammalian classII 3 chain amino acid and nucleotide sequences.Key words: interspecies cross-hybridization/DNA sequenc-ing/evolution/major histocompatibility complex/Northernblot
IntroductionThe major histocompatibility complex (MHC) was firstdiscovered in mice as a genetic region governing the abilityof transplants of neoplastic or normal tissue to succeed(reviewed in Klein et al., 1983). The second animal speciesin which an MHC was subsequently described is the chicken(Briles et al., 1950; reviewed in Longenecker and Mosmann,1981) in which the B locus was first detected as a bloodgroup system. An equivalent locus was discovered later inman, the HLA complex (reviewed in Dausset, 1981). Similarloci have now been described in all mammalian species ex-amined as well as in amphibians, and evidence points to itsexistence in all vertebrate classes. Most of the immunologicphenomena linked to the murine H-2 and human HLA com-plexes are also associated with the chicken B complex, e.g.resistance to neoplastic diseases (Pazderka et al., 1975; Col-lins et al., 1977), control of T cell-B cell interactions(Toivanen and Toivanen, 1977) and susceptibilty to autoim-mune diseases (Bacon and Rose, 1979).Genes of MHC complexes encode polymorphic cell sur-
face glycoproteins involved in the control of various aspectsof immune responses. The class I antigens (HLA-A, B, Cin man, H-2K, D, L in mice and B-F in chicken) are theclassical transplantation antigens expressed by virtually allnucleated cells that function as restricting elements for cyto-toxic T lymphocytes. The class II or Ta antigens (HLA-DR,DP, DQ in man, H-2, I-E and I-A in mice, B-L in chicken)are defined as the products of immune response genes whichrestrict antigen presentation to T helper lymphocytes. Theyare composed of two glycosylated chains, at (33-34K) and( (28-29K), noncovalently associated at the surface ofB lym-phocytes and cells of the myeloid lineage (macrophages anddendritic cells). In mouse and man, the MHC class II anti-gens have been extensively studied by serological, cellularand biochemical techniques, and the corresponding class IIgenes have been isolated and characterized in detail (review-ed in Hood et al., 1983; Auffray and Strominger, 1986).
It is only recently that polyclonal and monoclonal anti-bodies have permitted the study of the structure, biosynthesisand polymorphism of the chicken B-L antigens (Crone etal., 1981; Ewert et al., 1984; Guillemot et al., 1986). Thesereagents have also been used to study expression of B-L anti-gens during ontogeny (Ewert and Cooper, 1978; Guillemotet al., 1984). With the use of the quail -chicken nucleolarmarker (Le Douarin, 1969) in thymic interspecies transplan-tation schemes, it was possible to show that in the thymustwo cell types of different embryonic origin express theB-L antigens (Guillemot et al., 1984). These are the thymicepithelial cells and the dendritic cells differentiating fromhemopoietic precursors which migrate in the thymic primor-dium in successive waves during embryonic development(Jotereau et al., 1980).This model offers an unique opportunity to dissect the
series of early events which determine the ability to binda functional immune response in the context of self MHCclass H antigens. Moreover, since birds and mammals haveevolved separately for more than 250 million years, a detail-ed study at the molecular level of the chicken B-L antigensshould enhance our knowledge of structure -function rela-tionships of class H antigens.We have started a molecular analysis of the B complex
by attempting to isolate the immune response B-L,B genes.In this paper, we describe a successful approach using anHLA-DQf probe in interspecies cross-hybridization, and weprovide insight into the evolution of the MHC class H genesfrom comparisons of mammalian and avian sequences bothat the amino acid and nucleotide levels.
ResultsIsolation of chicken genomic clones using anHLA-DO! probeAs shown in Figure 1, we have chosen as probe a 496 bpSacl-EcoRI fragment derived from an HLA-DQ,B clone
1031©IRL Press Limited, Oxford, England

G/CI I
I I
I I
SP
Sac EcoR I
I I I I
I I I I
I I II1/2 i iTM I ICP CY
G/C
3UT
100pb
Fig. 1. The 496 bp Sacl-EcoRI DNA fragment was derived from the cDNA clone pll1-l (Larhammar et al., 1982). The GC tails generated duringthe cloning procedure are indicated by hatched boxes. The various parts of the DNA clone are delineated by a vertical dashed line; SP: signalpeptides; ,B1, j2: first and second extracellular domains; CP, TM, CY: connecting peptide, transmembrane domain, cytoplasmic domain;3'UT: 3' untranslated region.
1 2 3 1 2 3
300 1 300-C--1 200
HLA-DQJ probe(:496 bp fragment)
-1- 200
B-LSj' probe(whole X phage)
Fig. 2. Cross-hybridization between human and chicken class 11
chain sequences. Northern blot analysis was performed as described inMaterials and methods with the HLA-DQ/3 probe (left, see Figure 1)or the L6 phage clone (right) isolated with the human probe. Lane 1:mRNA from the human B cell line CA; lane 2: chicken liver mRNA;lane 3: chicken spleen mRNA. Size of hybridizing bands is indicatedin nucleotides.
(Larhammar et al., 1982). We searched for hybridizationconditions in which this human probe would allow detec-tion of an homologous chicken sequence. The DQ,B probelabeled at high specific activity by nick-translation was
hybridized to a Northern blot in low stringency conditions(see Materials and methods), and a series of bands were
detected in chicken spleen mRNA. Among them, a band at1200 nt, not detected in these conditions in chicken livermRNA, which migrated slightly faster than the mature1300 nt human DQ3 mRNA (Figure 2), was a good can-
didate for a chicken class II transcript. It seems likely thatthe three other bands represent 18S and 28S ribosomal RNAand breakdown products of 28S RNA. Very similar resultswere obtained after washing in 0.2 x SSC at 350C.We then used the DQ,3 probe to screen chicken genomic
DNA libraries in conditions simnilar to those described above.We screened 106 phage plaques from two independentlibraries (Dodgson et al., 1979; Ballivet et al., 1983). Weobtained 19 clones from the first library and 11 clones fromthe second which hybridized on duplicate filters and which
were purified through successive rounds of screening. Inorder to verify that the isolated clones contained genes thathad the expected characteristics of MHC class II genes, we
used labeled whole phage to reprobe the same Northern blotthat had been hybridized earlier with the DQf probe. The1200 nt band hybridized strongly with the chicken probe inchicken spleen mRNA, and faintly in chicken liver mRNA,whereas the 1300 nt HLA-DQ,B mRNA was detected as a
weak band (Figure 2).
Mapping to the chicken B complexThe sequence hybridizing to the HLA-DQf probe in one
clone from the XL47 library (L6) was isolated as a 3.2 kbHindlll DNA fragment. This fragment, referred to as thep14 probe (see Figure 4), was used to probe Southern blotsof genomic DNA from homozygous inbred and congeniclines of chicken. The p14 probe detected multiple DNA frag-ments of various intensities, some of which were poly-morphic in PvuII and BamHI digests (Figure 3). In contrastEcoRI and HindIII yielded only conserved patterns (notshown). Among the polymorphic bands, some were specificfor the B4, B12 or B14 haplotypes and others for groupsof two or three of them. The B4 and B12 haplotypes studiedare those of the CC and CB congenic lines which have dif-ferent B complexes on an identical genetic background. Thusthe ability of the p14 probe to detect polymorphisms bet-ween these lines readily indicates that it does hybridize tosequences located within the B complex. Similar conclusionscan be drawn from an extensive analysis of several in-dividuals of different chicken lines using multiple restric-tion enzymes (A.M.Chausse and C.Auffray, unpublishedresults).
In order to estimate the number of B-Lj3 genes, we haveused a shorter probe, p234, corresponding only to the 32domain (see below and Figure 4). When hybridized to B4and B12 genomic DNA digested with PvuII, p234 revealsa subpattern of four of the bands detected with the p14 pro-be (Figure 3). Three of these bands common to the B4 andB12 haplotypes are also found in individual clones isolatedfrom the genomic libraries of unknown haplotypes and thusrepresent three independent B-L, genes. The fourth PvuIIband detected with p234 is polymorphic and represents allelicforms of a fourth gene in the B4 and B12 haplotypes. Someof the isolated clones such as C6 (Figure 3) have a differentPvuII band, but whether it corresponds to a fourth gene or
a truncated form of one of the three conserved fragmentsremains to be determined.
1032
Y.Bourlet et al.
aVIXyd
I I-1
I 5 I v PM IIIIII
I I
II

Chicken MHC class 11 (genes
p234 probe
Pvu E
p14 probe
Pvu II BamH I
L20 L6 C2 C6C1OC2OB4 B12 B4 B12B14B19 B4B12B14B19
*- 9.5
qw
w
wl
mw
-0-- 4.4
-.*- 2.3a -o 2
Fig. 3. Enumeration of the B-L,B genes and mapping to the B complex. Southern blot analysis of chicken genomic DNA (right) and phage clones
(left) was performed as described in Materials and methods. L20, L6: clones isolated from the XL47 library; C2, C6, CIO, C20: clones isolated
from the XCharon 4A library. The B haplotypes of the genomic digests are indicated for each lane.
Structure of a chicken B-L,B geneThe 3.2 kb HindIII DNA fragment from the L6 clone was
further subcloned and analysed in detail by DNA sequenc-
ing. The sequence obtained together with its direct com-
parison with the HLA-DQ( sequence is shown in Figure4. Clearly regions of sequence similiarity where no gaps hadto be introduced were distinguishable. They correspond totypical (31, (2 and transmembrane exons containing an open
reading frame which can be connected through splicing us-
ing the typical signals which are found at their limits. It isworth noting that these intron/exon limits interrupt the codingframe between the first and second bases as is observed inall MHC genes and other members of the immunoglobulinsupergene family.Although we have sequenced 860 bp upstream of the (31
exon (Figure 4A and data not shown), it was not possibleto detect the signal sequence exon which might be toodivergent when compared to the HLA-DQ(3 or other classII chain sequences. This is not too surprising because thisregion is very divergent between isotypes in a given species(Tonnelle et al., 1985). Alternatively this exon may belocated outside of the clone analyzed. Exons encoding thecytoplasmic domain and 3' untranslated region were iden-tified by comparison to a cDNA clone corresponding to a
transcript of another B-L(3 gene in the B12 haplotype (Figure4; R.Zoorob and C.Auffray, unpublished data). The fourintrons separating the five exons were found to be very short
(108, 83, 86 and 86 nt) when compared to their homologuesin human and murine class II chain genes. For example,in the HLA-DQ( gene, they are 2889, 515, 484 and 610 ntlong respectively (Boss and Strominger, 1984). The nucleo-tide sequence alignment between the B-L( gene and theHLA-DQ( sequence, part of which was used to clone itreveals that the extent of conservation is approximately thesame in the three exons, ranging from 62-66% of sharedresidues (Figure 4 and Table I).
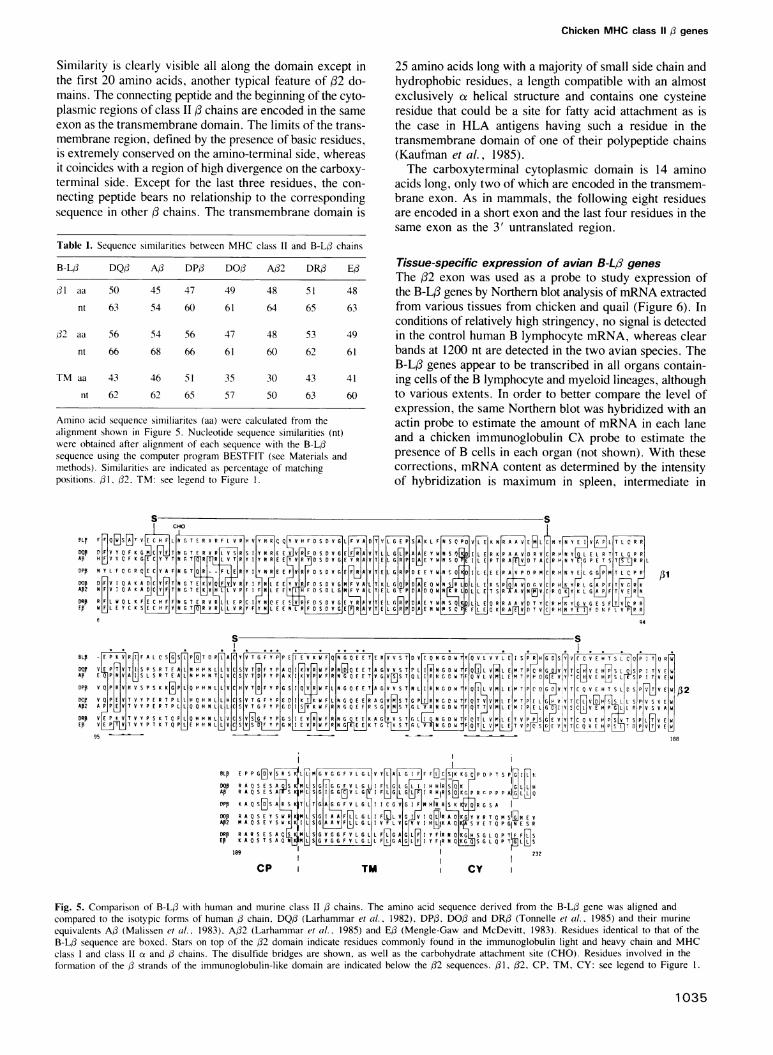
The B-L/3 chainThe predicted amino acid sequence of the chicken B-L3 chainwas compared to that of human and murine class H chains(Figure 5). The chicken sequence has all the characteristicsof a class chain. The (31 exon encodes amino acids 6-94and contains conserved residues for N-glycosylation (NGTat positions 19-21) and intradomain disulfide bond forma-tion (cysteines 15 and 79) as well as several clusters of con-served residues (Figures 4 and 5). The (32 exon (residues96-188) is one codon shorter in the B-L(3 gene, probablydue to a single mutation in the AG dinucleotide at the endof the polypyridine track in the preceeding intron followedby the use of a downstream AG allowing in-phase mRNAprocessing. The (32 domain is therefore 92 amino acids longwith two conserved cysteines involved in the formation ofthe 55 amino acid intrachain disulfide loop that is a typicalfeature of members of the immunoglobulin superfamily.
1033
kb

Y.Bourlet et al.
A
(3 (2 TMCY3'UTcL I U .. 4 ~~~~~~p14
200bpp234
F F Q81B 1 CGCTGGGGGGGCCGAGACCGCGrGGCTTGGCGCAGGACGCCTAGTCCCCCGA CA, TTT TTC CAG 115
DON8 41 AG GAT TTC GTG 51D F V
W S A T V E C H F L N G T E R VAR F L V RNH V Y N NR CAY V116 TGG AGT GCT ACA GTT GAG TGC CAC TTC CTC AAC GGC ACC GAG CGG GTG AGG TTT CTG GTG AGG CAC GTC TAC AAC CGG CAG CAG TAC GTG 205
52 TAC CAG TTT AAG GGC ATG TGC TAC TTC ACC AAC GAG ACA GAG CGC GTG CGT CTT GTG AGC AGA AGC ATC TAT AGC CGA GAA GAG GTC GTG 141V QF K GM CVY FT N G T E V R LV SNR S I AN E E V V
N F D 5 D V G L F V A D T V L G F P S A K L F N S Q P D V L206 CA T A G GAT GTT GGT CTC TTT GTG GCC GAT ACA GTC CTG GGA GAG CCT TCT GCT AAA ETC TTC AAC AGC CAG CEG GAC GTG CTG 295
)1 142 COGC TTC GAC AGC GAC GTG GGG GAG TTC CGG GCG GTG ACG ETG CTG GGG CTG GET GCC GCC GAG TAC TGG APE AGC CAG AAG GAC ATC CTG 231RF D S O VAG E F N A A T L LAG L F A A E A W N S Q K II L
E K N N A A V E M L C G Y N YA E I A A P L T L Q N N296 GAO MG MA AGG GCT GCA GTG GAA ATG CTC TGC AAC TAC AAC TAC GAG ATAGTG GCC CCT TTG ACG CTG CAG AGG AGA A GTGAGTTCACGTG 387
232 GAG AGG AAA CGG GCG GCG GTG GAC AGG GTG TGC AGA CAC AAC TAC CAG TTG GAG CTC CGC ACG ACC TTG CAG CGG CGA G 310ER F K A AM V N A EC H NA Q L E L NT TLRN NR
ERP K AN388 TGCCAGAGCTTGTGACTGGGCACAGCGTGATGAGCGACGGGTGTTACAGCCTGAGAGETCT CAACCTGTCGTCGCTACTTECCCCCCCCCGGAG AG EC AAG GTG AGG ATE 501
311 TAG AG EGG AGA GTG AGC ATE 330A RP T A T
F AL Q S G SLAY T O NL CY C A T F YRP E l E A K502 TTC AG CTG GAG TCG GEC TEE EGA CGE CEM AGC GAC EGG ETA ACT TG TAC GTG ACG GCE TTE TAC CEG CEG GAG ATE GAG GTGAMG TAG 591232331 TCGGCA TCC AGG ACA GAG AGE ETC ME GAE GAGCA ACET ETAGTC TAG TG GTAG ACA GT TTCG TATCCA GC CGAG ATCE A GTE EGG TAG 420
S P 5 R T E A L Nl H H N L L C S A T F A P AR I K V N E
F Q N A Q E E T E R A A S T V I C N A D0 W TKVA L A I592 TTC CAG AACGGAG CAG GAG GA TGACAGAG CGCGTA GTG TCC ACG GAGC GTA AT GCAG AAC GAG OAT TAG AG TAG GAG CTG EAG GTG AGTG ET 681
421 T CGG AAT GAG CAA GAG GAG AGA GET EGGC TT GTG TEE AGE GEE CTT ATT AGE AAT GOT GAG TAG AGE TTC GA ATEC TT GTG ATE ETA 510F R N D C E E T A GAV VA T P L I ON 00W T F 0118 ML
E S ONE D SAG Q A E H TI L Q Q R I T C O W682 GATC AGC CG CAG CG GGCC0 GGA CAGE TAT GTG TG CGAG GTE GAG GAG AGE AGE ET GAG CAG EEC ATC ACCG GAG AC GG GTAAGEGGEC 773
511 GAA ATG AGT GCG GAG GOT GA GAC ATC TAC AC TAGC CGAG GTG GAGTCACG CGEAG GTC GAG AGC GCCATE AGE ETA GAG TGGC 592E M T P Q N A D A A T C H A E N P S L Qn P I T VE 0
E P RGR VS A N S K L174 TCG CAGGCTGTGEAGTGGhGAGAGOATGGGAGCGTGGCGTTCAGAA TCCCGATGCTCETTCETGTCCCTGCAGAGA CGGG GATE ACG TAG AG AGG AGE MG ETG CTG 081CP ** ** ** * ****** * * * ** *** ** ***
59 C AG GET GA TCT AAA TCT GECCAG AGE MGO ATG CTG 627NAR C SE S A C S K M L
M O V E G F LG L Y L A L 0 1 F F F L E O K K882 ATAGAG GTCAGO GGA TTTATG ETA GAGG GT GTE TAG GTT GGA ETA GAG ATC TTGC TT TTC CTAG TC AGT APA AAA G GTGTGTGTATGGGCCAG 974
TM * *** * ** * ** **I*** * ** *** ** ****** ** ***** ** * ** **** ********* ** _
620 AGT AGE AlT GGA AGE CTT GIG ETA GAGCTA ATC TTC CTC GAG GTG GAGC CT ATC ATC CAT GAGC AG AGT GAG MAA E 703S A A OG F A L E L I F L OG L L I NH P QS K
C P 0 P T S P
BLB 975 GCGGTCTTGGTTGGGGAGGAGGGGTGTGGTGAAGCTGTGACCGTGCTGACCTCTCTTGCTGCATTTCAG AG CM CCGT GAT C GAAC CAGTCCA G GTAATGTCTCATTE 1082812 4+ + ++ + + ++ + +++ +
C GGE GGC CEG GTCE AG G GCT GCCA AGGOR V A A AR
O IL N *1083 CCCAGTGACATTACTGTACTCAGAGGAGCGCTAGGGGC TGTGGTCCTTATTTCTCCTCTTTCTCTTAG AG GOTC CTG TEC AGCTAAGAAATGGTGAGTGGTTGCTCCAC 1194
CY ++ ++ +++ ++ ++++ ++++++ + + .. ... ...AG PTA GTG MTTAGCTGCTGCCFEGGCGAGCCGCTGTACG M L N *
1195 CAGCATGTCCAGCACTCCCCGGCCCTGAGCTTTGAGTTTTCTTTGGGGTTTCACCGTGTTCCT ... GCTGCCACTGCAGAGCCTGCTCTGTCCTTCTC. .AATAAGTACTCCTGAG 13063UT . ... ++... .. ..... . + + + + ++ + +++ ++ ++++, ++
CCGCACCCCCCGC . .TCTCCGGCCGTCGCC .TCGGCTCTCCCTCGGGCTGCCACCGCGTCCGTTGGAGATGTCGCCACGATGCACGCTTCGTCCCCATCCTAATAAACGCGCTGACC
1307 ATGTAGCACAAGAAGTGAAGGAAGC12CAGTGGCTTAAATGGGGGTTCAAGG4TCATGCTAGTGGGTCACCAGTAGAGAAACAGTGGGGCACGTTGTGGTAGAAACGGGTATACTGGG1424TTTG
Fig. 4. Structure and sequence of the B-Lfl gene. (A) The structure of the pl4 clone. Exons are indicated by black boxes. The strategy used forsequencing is shown with heavy arrows for Maxam-Gilbert and light arrows for Sanger determinations. The open box corresponds to the phagearm. (B) Partial sequence of pl4 and comparison with HLA-DQI3. Only the three identified exons and their flanking sequences are shown (see text).The DNA sequences (B-L,3 on top, DQ,8 below) have been aligned and matching nucleotides are indicated by stars with the longest stretch in eachexon boxed. The GT and AG dinucleotides at the intron/exon limits are underlined. The coding sequences have been translated using the single-letteramino acid code. Numbering starts at 1 in the B-Lf sequence shown and follows that of the cDNA insert for the HLA-DQJ3 sequence (Larhammaret al., 1982). fl1, fl2, CP, TM, CY: see legend to Figure 1. At the bottom, the B-L3 sequence is aligned with that derived from a B-L3 cDNAclone from the B12 haplotype, with matches indicated by '+' signs. CY, 3'UT: see legend to Figure 1. The stop codon is indicated by a star.
1034
Chicken MHC class 11 d genes
Similarity is clearly visible all along the domain except inthe first 20 amino acids, another typical feature of 32 do-mains. The connecting peptide and the beginning of the cyto-plasmic regions of class II f chains are encoded in the sameexon as the transmembrane domain. The limits of the trans-membrane region, defined by the presence of basic residues,is extremely conserved on the amino-terminal side, whereasit coincides with a region of high divergence on the carboxy-terminal side. Except for the last three residues, the con-necting peptide bears no relationship to the correspondingsequence in other 3 chains. The transmembrane domain is
Table I. Sequence similarities between MHC class II and B-L/ chains
B-LO DQO AO DP,B DO/ A/2 DR,B Ed
d1 aa 50 45 47 49 48 51 48
nt 63 54 60 61 64 65 63
32 aa 56 54 56 47 48 53 49
nt 66 68 66 61 60 62 61
TM aa 43 46 51 35 30 43 41
nt 62 62 65 57 50 63 60
Amino acid sequence similiarites (aa) were calculated from thealignment shown in Figure 5. Nucleotide sequence similarities (nt)were obtained after alignment of each sequence with the B-L/sequence using the computer program BESTFIT (see Materials andmethods). Similarities are indicated as percentage of matchingpositions. /1, 32. TM: see legend to Figure 1.
25 amino acids long with a majority of small side chain andhydrophobic residues, a length compatible with an almostexclusively ax helical structure and contains one cysteineresidue that could be a site for fatty acid attachment as isthe case in HLA antigens having such a residue in thetransmembrane domain of one of their polypeptide chains(Kaufman et al., 1985).The carboxyterminal cytoplasmic domain is 14 amino
acids long, only two of which are encoded in the transmem-brane exon. As in mammals, the following eight residuesare encoded in a short exon and the last four residues in thesame exon as the 3' untranslated region.
Tissue-specific expression of avian B-LA genesThe 32 exon was used as a probe to study expression ofthe B-L, genes by Northem blot analysis of mRNA extractedfrom various tissues from chicken and quail (Figure 6). Inconditions of relatively high stringency, no signal is detectedin the control human B lymphocyte mRNA, whereas clearbands at 1200 nt are detected in the two avian species. TheB-L3 genes appear to be transcribed in all organs contain-ing cells of the B lymphocyte and myeloid lineages, althoughto various extents. In order to better compare the level ofexpression, the same Northern blot was hybridized with anactin probe to estimate the amount of mRNA in each laneand a chicken immunoglobulin CX probe to estimate thepresence of B cells in each organ (not shown). With thesecorrections, mRNA content as determined by the intensityof hybridization is maximum in spleen, intermediate in
CHO
PL F [WISAE T V H F LN G T R V R F R H V Vy N Q Y V H F D S D V GIL F VA D T VL G E P S K L F N S C PD V E K N R A A V EHI+RC N N V ] T L C R R
D5P 0 F V Y Q F K GM C YLF TA G T E R P L V S R S I Y N R E E VR F D S D V G E A V T L L G L P AAI E Y W N S[ DI L E R K P A A V D R V C H N E L R T T L Q P RAP H F V V C F K G E Y Y T N G TOR R L V T P Y Y N R E E V R D V G E Y R A V T E L G R P D E Y W Y S F E I L E P T R A E V D T A C N Y G P E T S T[fR R LDPF N Y L F C, PR E C Y A F N G TR-FEL R Y I Y N R E E FV R|F D S D V G EF A V T E L G R P D E E Y W N Q|0KDDI L E E E P A|V P D P M C R H|N Y E|L G G PM T L C P P 1DOPDAFV IQ A K A IC TFNG T EKQ V R F I F NL E EYV RIF D S DV GMM FVLA L T K L GQ P D A E Q W N S LDIL L E R S AAI D V C P HK YRL GAPIF TAH K
AP2 N F V I Q A K A2JC TN G T E KY L AV P Fl F NIL E H F D S D 16G MN A L T E L G P DAD Q W N LI E T S AVR NEJA CR SKY K GAP F TVERDRB RFL W Q L K FE C H FIFN T E R VRLLE P C I Q E E F D S DV G E Y RPA V T E LKISRP D[AjE Y W N SK DJL L E Q R R A A VID T Y C R H YLG E S F T V RE WhFL E Y C K S E C H F YN G T VR R Y F L E E N L R F D S D V G E[3RL V T E G R P D A E N W S C PE F L E Q K R A AD T V C R H N YV F N F V PHH
94
s s
BLF E P K V RJI F A L C S [GI S LP T D P 1ACYV TG F Y PP E V K WFK QN G E ETS ERK V V STA D V I Q N G D W TY V L V V L E ISP GDS V C H T S L C Q PI TSQ RHD5P EP T AT IS P S R T F AIL|N H HN LIVCDSV TD F Y P A QI KVRW FP NESQK E T A GAV V S TP L II RN G D W F SQjIL VAN L ENM T PQR G D V VYT C V E HPs L SIP ITV EWAP E P N VI S L S R T E A YLNP N T LVC|S|V TD F Y P A KSI K V PWFKRYP G Q F E TAV G V fS TSQ L IIP N G C W TF|Q V L VIM|L EM T PHPSGSE V YT C V E HPs LSIK SPI TV EAWDPP V Q5 P RVA S P S K Kj P LQ H N L ALVC|HVTC F Y PG SI0 ARW FLINSETAPGV T N II RN FOIL AM M T PC Q5G V YVTC Q V E H T S L|C SPFV TA A 32
DCp VQA P|EAVT V Y P E R T P L|LH Q H N L|LHC|S
ATG F Y P6
01 KSI K W FLP N G Q E EKR A GDVA S LIP AVG TF ST M EM T PE SY TC
L SIPV S EAB2 AYPPEVT V Y P E P T PLPLCQQ H N LIL|L|C|SV TG F Y P|G K W FR|K G Q EFPRSGPVVASTG LVRN G D WTFLQDTTVVMCLE|M ILPELAKYPGSIICIVE H P P|PV S V A|WDRB V| P K V|T V Y P S K T C F|L|Q H H N L L|V|C S| Y P| SI E VW FRN G g E E|K A G V V S T|G LLA G D W T|F|Q|T| v|tL E|T v P_ S|E V|YT|C Q V E IPPSL T S PIL TV EIW|ED V EP WK T MPLOHEHNLLSSASFYP|IEARAFR EKKTGA TGLV NGDATTLVMITVPSDE VYTIC Q V EHKPSVTSLTVW
95 188
BLB E P P G HR S L MG V G G F V L G LIV YAL L G F F Ff1C_SIK KG P D P T S P [N]
DQE R A Q S E S MISSML SGIG G F V L G L F|L G|LG I H HIR SNK IGLHAP R A Q S E S AS KM L S G'IG G[3V L GEI F LGLGG LI R HiR,R PHr P P P Al GLL QOPP K A Q SnDS A S TI SLTGAG G F V L G L I C G V G I F NAIR R S K R S A
~~HA]
F IML~ TiVODOP R A QS E Y S ARKMLSSGIRA L G L I FQlL V G I IR KG YV R T Q M INAP2 M A QS E Y S WKK L S G AAYIFL L G L IVFLAG VAJIK A S|KS V E T Q P GIN E S RDRF R A R S E S A CLKM LI S|G V G G F V L G L L FnLG A G LM I Y FIR N QKG H S G L Q P TIF F L SEF K A Q S T S A Q ALLSIGVG G F LIP G AL IY FIR N Q KGS G L Q P T 5ELLS
189 232
cP TM CY
Fig. 5. Comparison of B-L/ with human and murine class II /3 chains. The amino acid sequence derived from the B-L/3 gene was aligned andcompared to the isotypic forms of human / chain. DQ/ (Larhammar et al.. 1982), DP/. DO,B and DR/ (Tonnelle et al., 1985) and their murineequivalents AO3 (Malissen et al., 1983), A/2 (Larhammar et al.. 1985) and Ed (Mengle-Gaw and McDevitt, 1983). Residues identical to that of theB-L/ sequence are boxed. Stars on top of the 32 domain indicate residues commonly found in the immunoglobulin light and heavy chain and MHCclass I and class 11 ce and /3 chains. The disulfide bridges are shown, as well as the carbohydrate attachment site (CHO). Residues involved in theformation of the d strands of the immunoglobulin-like domain are indicated below the /32 sequences. /31, /32, CP, TM, CY: see legend to Figure 1.
1035

Y.Bourlet et al.
,ts< - n i < 6 S r. = ._ . _ __ _, _ _ . __. i_ . _ , __ ....... _, _ ,., .. ... ,, . ... _,, _ .= _ .... ...... .. .'9$-;!
_T
.7 > ;fo.
_ Tof
w..:
...
a. -
'. ._.'; Ld .,' 'aS
\5. 1.4,,,: S_
*ati_ :
= .S=V :s rrO* S_=t, qop
J.
Fig. 6. Tissue specific transcription of the B-L13 genes. Northern blot analysis was performed as described in Materials and methods using the B-Lfl2 domain probe p234, and 10 yg mRNA from the various sources indicated.
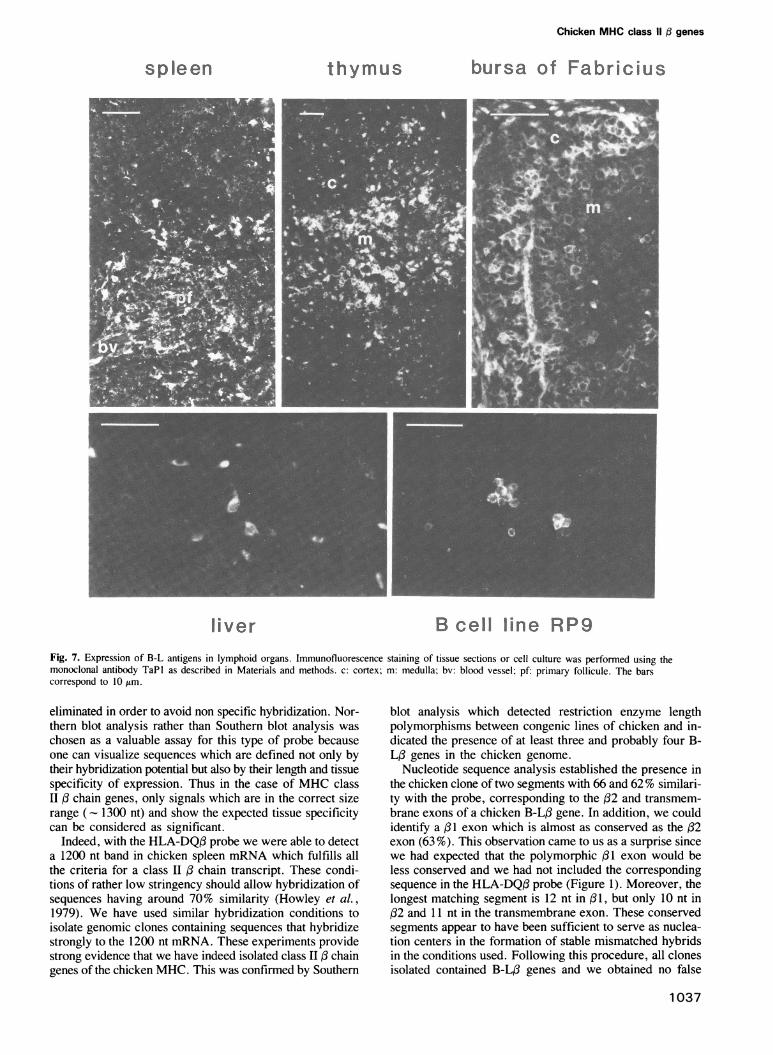
thymus and bursa of Fabricius, and low in liver, whereasno signal can be detected in quail cerebellum. This is alsoin good agreement with the number of cells and the intensi-ty of immunofluorescence staining detected with the anti B-L monoclonal antibody TaPl in each organ or cell type(Figure 7). TaPl labels strongly myeloid (macrophage-like)cells in all tissues analyzed but these cells are found in greaternumbers in the spleen around the blood vessels and in thethymic medulla than in liver or bursa of Fabricius. In thethymic cortex, epithelial cells are stained weakly whereasthymic lymphocytes are negative. In contrast, immature lym-phocytes located in the medulla of bursal follicules are label-ed weakly, whereas more mature cells of the B lineage foundin the cortex of bursal follicules and in splenic primaryfollicules express higher amounts of B-L antigens. Thus B-L,B genes are transcribed and expressed with the same tissuespecificity as mammalian class II: genes.
DiscussionThe successful cloning of the human and mouse MHC genesaccomplished in the past years has provided new tools tostudy the structure and function of MHC antigens and hasallowed a detailed study of the chromosomal region con-taining this complex. Thus genetic markers defined withgreat precision are now being used in association with thosedefined by biochemical or cellular assays in attempts tounravel the genetic basis for MHC-linked immune regula-tion. The ability of an organism to develop immune defenses
in the context of its own MHC antigens is learned duringthe embryonic life, but we know very little at present onthe succession of molecular and cellular interactions whichare critical in this process. One possible approach to iden-tify the structure -functional relationships of MHC antigensis to reconstruct the evolutionary history of the MHC bydescribing the MHC genes, their products and functions inclasses of vertebrates which have evolved separately fof longperiods of time.We have been interested in the role ofMHC class II anti-
gens during ontogeny in the chicken embryo and we havetherefore investigated the possibility of using HLA class IIgene probes to characterize their chicken equivalents, theB-L genes. With the accumulation of DNA and protein se-quences, most often derived from that ofcDNA or genomicclones, for various isotypes of a and ,B chains in mouse andman, it became clear that the class II ,Bchains are somewhatmore conserved between isotypes in a given species or bet-ween species than ce chains. Thus we assumed that an HLAclass II : chain sequence would represent a good candidateprobe for detection of chicken sequences in interspeciescross-hybridization experiments. The precise design of theprobe derived from an HLA-DQ,B cDNA clone was directedby the following considerations: the probe should containthe segment encoding the parts of the ( chain with the highestdegree of sequence conservation, that is the (32 immuno-globulin-like domain and the core of the transmembrane do-main. The AT-rich 3' untranslated region and the GC-richregions generated during the cloning procedure should be
1036
Chicken MHC class ll 3 genes
thymus bursa of Fabricius
liver B cell line RP9Fig. 7. Expression of B-L antigens in lymphoid organs. Immunofluorescence staining of tissue sections or cell culture was performed using themonoclonal antibody TaPI as described in Materials and methods. c: cortex; m: medulla; bv: blood vessel; pf: primary follicule. The barscorrespond to 10lm.
eliminated in order to avoid non specific hybridization. Nor-thern blot analysis rather than Southern blot analysis waschosen as a valuable assay for this type of probe becauseone can visualize sequences which are defined not only bytheir hybridization potential but also by their length and tissuespecificity of expression. Thus in the case of MHC classII , chain genes, only signals which are in the correct sizerange (- 1300 nt) and show the expected tissue specificitycan be considered as significant.
Indeed, with the HLA-DQ,B probe we were able to detecta 1200 nt band in chicken spleen mRNA which fulfills allthe criteria for a class II ( chain transcript. These condi-tions of rather low stringency should allow hybridization ofsequences having around 70% similarity (Howley et al.,1979). We have used similar hybridization conditions toisolate genomic clones containing sequences that hybridizestrongly to the 1200 nt mRNA. These experiments providestrong evidence that we have indeed isolated class II ( chaingenes of the chicken MHC. This was confirmed by Southem
blot analysis which detected restriction enzyme lengthpolymorphisms between congenic lines of chicken and in-dicated the presence of at least three and probably four B-L,B genes in the chicken genome.
Nucleotide sequence analysis established the presence inthe chicken clone of two segments with 66 and 62% similari-ty with the probe, corresponding to the ,(2 and transmem-brane exons of a chicken B-L( gene. In addition, we couldidentify a (31 exon which is almost as conserved as the (2exon (63 %). This observation came to us as a surprise sincewe had expected that the polymorphic (31 exon would beless conserved and we had not included the correspondingsequence in the HLA-DQ( probe (Figure 1). Moreover, thelongest matching segment is 12 nt in (1, but only 10 nt in(2 and 11 nt in the transmembrane exon. These conservedsegments appear to have been sufficient to serve as nuclea-tion centers in the formation of stable mismatched hybridsin the conditions used. Following this procedure, all clonesisolated contained B-L( genes and we obtained no false
1037
spleen
I

Y.Bourlet et al.
positives. Thus the experimental protocol used in this studyis both sensitive and specific. However, in similar condi-tions we were unable to obtain any positive clones using anHLA-DQa probe in several attempts. This might be due tothe fact that MHC class II ae chains have diverged more ex-tensively than / chains (as pointed out above).We have compared the B-L/ chain sequence with all
known mammalian class II chains both at the amino acidand nucleotide levels (Figure 5 and Table I). In the ex-tracellular domains, the B-L3 sequence has 46-56% aminoacid sequence similarity and 55-68 % nucleotide sequencesimilarity with its mammalian homologues. This suggeststhat the series of duplications that led to the present isotypesof class 1 chains had already taken place at the time whenbirds and mammals diverged from a common ancestoraround 250 million years ago. The transmembrane exon isoverall more divergent, but this reflects the lack of conser-vation in the connecting peptide, whereas the hydrophobiccore of the transmembrane segment is as conserved as theextracellular domains. Since the B-L,B gene sequenced isequally similar to the various mammalian sequences, it canbe anticipated that any of the human or murine probes wouldhave been suitable for isolation of the B-L/ genes. It is thuspossible that the three genes isolated in this study representa full complement of chicken class II: chain genes in theMHC haplotypes from which the genomic libraries werederived, with at least one member of different isotypicfamilies. Analysis of these additional genes will provide astructural basis to answer these questions.Although the overall homology between the chicken and
mammalian sequences is smaller than that obtained in intraor interspecies comparison in the mammalian class, both thenature and the location of the conserved residues are similar.This is well illustrated by the fact that similarity reaches 71 %when one takes into account the amino acids in the /2 do-main of the B-L/3 sequence which have physical and chemicalproperties similar to the most common amino acids foundin the other sequences (for example Ser/Thr at 106, Ala/Valat 116, Lys/Arg at 130). Among the 48 positions in the /32domain, where most / chains share the same amino acids,25 are also common with other members of the immuno-globulin gene superfamily (Figure 5). These include the ma-jority of the hydrophobic residues found at altemate positionsand involved in the formation of /3 strands which are theessential building blocks of the prototype three-dimensionalimmunoglobulin domain fold.
In addition to the distinctive features of a class II a chain,there are highly conserved segments in each domain. Thelongest of these segments in the polymorphic /31 domain isthe VHFDSDVG sequence (38 -45). It corresponds to theregion where similarity with the fibronectin cell-attachmentsite has been noted previously (Auffray and Novotny, 1986),leading to the proposal that it represents the non-polymorphicbinding site of the class II /3 chain interacting with the CD4T-cell differentiation antigen (Auffray, 1986). The pentapep-tide NGDWT (150-154) and a stretch of 9 amino acids,GGVGGFVLGL (202-210), appear as structural invariantsignatures of the 32 and transmembrane domains, respec-tively.We have investigated the pattern of transcription of B-L/3
genes in various tissues by Northern blot analysis using the/2 domain as probe. The faint signal detected in liver isrelated to the presence of macrophage-like cells expressingB-L antigens (Figure 7), probably analogous to the passenger
1038
macrophages and Kupffer cells of human liver (Lauten-schlager et al., 1984). The high level of expression seen inthe thymus reflects the fact that this organ contains a largenumber of cortical epithelial cells expressing B-L antigensrather faintly and that most medullary dendritic cells expressB-L antigens at a high level (Guillemot et al., 1984 andFigure 7). In contrast, the signal detected in the bursa ofFabricius is surprisingly low considering the large numberof bursal lymphocytes which stain with an anti-B-L mono-clonal antibody when analyzed in suspension. However, thisstaining is restricted to the mature B lymphocytes and themedulla of the follicules is labeled only weakly. This resultmight also be explained by the fact that we have used 8-week-old animals in which involution of the bursa is in progress,or it could be related to a different turnover and level ofB-L/ mRNA in this organ. At this stage of the analysis, itis not possible to determine how many and which of the B-L/ genes are transcribed and translated. The B-L/ geneanalyzed has an exon/intron structure similar to the mam-malian class II / genes, except for the very small size ofthe introns. Because we have not identified the regions ofthis gene containing the signal peptide exon and the promotersequences, further experiments will be necessary to establishthat it is an expressed gene.
Since the B-L/ probe also hybridizes to quail mRNA, itwill be possible to isolate the corresponding genes and todetermine the degree of similarity of class II antigens in twoavian species in which interspecies intra-embryonic graftsare tolerated and are used extensively to study ontogeny.In addition, chicken class II gene probes will help definingnew markers for the genetic analysis of the B complex us-ing restriction enzyme polymorphisms and chromosomewalking procedures. Hopefully it will be possible tomanipulate these genes during the embryonic stages in orderto better understand their role in the establishment of a func-tional immune system.
Materials and methodsThe HLA-DQ/3 probeA 496 bp Sac -EcoRI DNA fragment from the HLA-DQ,B cDNA clonepll13-I (Larhammar et al., 1982) was subcloned into pUC12 (Vieira andMessing, 1982). Plasmid DNA was prepared from an overnight saturatedEscherichia coli culture by alkali/SDS extraction (Bimboim and Doly, 1979),followed by banding in two successive CsCI/ethidium bromide gradients,cut with EcoRI + Sacl to completion and the 496 bp fragment was isolatedafter electrophoresis on a 5% polyacrylamide gel by electroelution andDEAE-cellulose chromatography. The isolated fragment was labeled bynick-translation to a specific activity of 2-4 x 10 c.p.m./ltg (Rigby etal., 1977).
Screening of chicken genomic DNA librariesA X Charon 4A genomic library (Dodgson et al., 1979) and a XL47 genomiclibrary (Ballivet et al., 1983) were screened with the HLA-DQf3 probe onduplicate filters essentially as described (Maniatis et al., 1982). Nitrocellulosefilters (Millipore HAHY) were prewashed 2 h at 42'C in 50 mM Tris,pH 8.0, 1 mM EDTA, 0.1% SDS, prehybridized 4 h at 420C in 50% form-amide, 5 x SSPE, 0.1% SDS, I jug/ml poly(A), 1 Ag/ml poly(C),100 1g/ml denatured salmon sperm DNA. The nick-translated probe(2-4 x l08 c.p.m./g) was then added to the same buffer and hybridiza-tion carried out 24 h at 42°C and 24 h at 37°C. The filters were washed2 x 1 h at 45C and 2 x 1 h at 55°C in 2 x SSC, 0.1% SDS, and ex-posed to X-ray films (Kodak) for 3-7 days with two intensifying screens(Dupont Liplus). Positive clones were purified by rescreening under thesame hybridization conditions.
Mapping, subcloning and DNA sequencingPhage DNA was prepared as described (Maniatis et al., 1982). A restric-tion map of the recombinant phage was determined using the oligo cos/partial

Chicken MHC class 11 , genes
digest approach (Rachwitz et al., 1984) and a 3.2 kb HindIII DNA frag-ment hybridizing to the HLA-DQ1 probe was identified by Southern blotanalysis using Genescreen (NEN) nylon membrane (Southern et al., 1975).This DNA fragment was subcloned into pUC9 (Vieira and Messing, 1982),yielding the subclone p14. Restriction enzyme and Southern blot analysisindicated that the HLA-DQ,B probe hybridized to two internal PstI fragmentsfrom p14, 400 and 250 bp in length. The latter, corresponding to the ,B2exon, was subcloned in pUC9, and is referred to as the p234 clone.DNA sequencing of p14, and subclones thereof, was performed using
the chemical degradation procedure (Maxam and Gilbert, 1980) by label-ing 3' ends with the Klenow fragment of DNA polymerase I and ax 32P_labeled deoxynucleotides, and by the dideoxy chain termination method(Sanger et al., 1977) using [a-35S]dATP and modified T7 DNA polymerase(Sequenase, US Biochemicals) after subcloning the p14 insert in Ml3mpl8.17mer oligonucleotides used as sequencing primers were synthesized onan Applied Biosystems DNA synthesizer. Nucleotide sequences were com-piled on a micro computer using the PCGENE software package (Genofit).Alignments were performed using the program BESTFIT using theBISANCE facility (CITI2, Paris).
Southern blot analysisHigh mol. wt DNA was extracted from red blood cells of B4, B12, B14,B19 chickens with proteinase K and SDS (Blin and Stafford, 1976), digestedto completion with PvuII or BamHI, electrophoresed on 0.6% agarose gelsand blotted on nylon membranes (Amersham) as described (Southern, 1975).
Northern blot analysisLiver, spleen, thymus, bursa of Fabricius were dissected from 8-week-oldWhite Leghorn chickens (Gallus gallus) and japanese quails (Cotumnix cotur-nix japonica) and frozen in liquid nitrogen. The chicken RP9 cell line wasmaintained in culture in DMEM supplemented with 10% fetal calf serumand 1 % chicken serum in a humidified atmosphere containing 5% CO2-Cells were collected by centrifugation and washed in phosphate buffer saline.The final pellet was frozen in liquid nitrogen. RNA was extracted usingthe LiCl/urea procedure (Auffray and Rougeon, 1980). Poly(A)+ RNA waspurified by two cycles of oligo(dT)cellulose chromatography, 10 yg wasdenatured with glyoxal, electrophoresed on a 1.1 % agarose gel (McMasterand Carmichael, 1977) and transferred to a nylon membrane (NENGenescreen). The Northern blot was prehybridized for 4 h at 420C in 50 mMNa phosphate, pH 6.5, 50% formamide, 5 x SSC, 1 x Denhardt,250 ug/ml sonicated salmon sperm DNA. Hybridization was performed inthe same buffer at 37°C for the HLA-DQ13 probe, and at 50°C for the p234or the whole L6 phage (see above). The probes were labeled by nick-translation at 2-4 x 108 c.p.m./4g. The final wash was in 2 x SSC,0.1% SDS at 550C (HLA-DQ,B) or in 0.1 x SSC, 0.1% SDS at 500C(p234, L6 phage).
Immunofluorescence analysisIndirect immunofluorescent staining of 1-week-old chicken spleen, thymus,bursa of Fabricius and liver was performed on fixed tissue sections, whereaslabeling of the RP9 cell line was done on living cells as previously describ-ed (Guillemot et al., 1984). Reagents used were the anti-chicken B-L mono-clonal antibody TaPl (Guillemot et al., 1986) and a fluoresceinated goatanti-mouse immunoglobulin serum (Nordic).
MaterialsEnzymes and chemicals were obtained from Appligene, Amersham, Biolabs,Boehringer, BRL, Genofit, Pharmacia and Sigma. Radiochemicals werefrom Amersham.
Acknowledgements
We thank Drs Dan Larhammar and Per Peterson (Uppsala) for the pIIB- 1plasmid, Dr Marc Ballivet (Geneva) for the XL47 genomic DNA library,Dr Jean-Claude Weill (Paris) for the chicken immunoglobulin CX probe,Drs Dominique Piatier-Tonneau and Angelica Keller (Nogent) for the giftof human and quail mRNA, Caroline Knipp and Louis-Noel Gastinel foroligonucleotide synthesis, members of our laboratory for helpful discus-sions, Dr Nicole Le Douarin for encouragement, Marie-FrancoiseMeunier, Sophie Gournet and Yann Rantier for typing, artwork andphotography. This work was supported by grants from Centre National de
ReferencesAuffray,C. (1986) C.R. Acad. Sci. (Paris), 302, 287-292.Auffray,C. and Novotny,J. (1986) Human Immunol., 15, 381-390.Auffray,C. and Strominger,J.L. (1986) Adv. Hum. Genet., 15, 197 -247.Auffray,C. and Rougeon,F. (1980) Eur. J. Biochem., 107, 303-313.Bacon,L.D. and Rose,N.R. (1979) Proc. Natl. Acad. Sci. USA, 76,
1435- 1437.Ballivet,M., Nef,P., Stalder,R. and Fulpius,B. (1983) Mol. Neurobiol., 48,
83-87.Birnboim,H.C. and Doly,J. (1979) Nucleic Acids Res., 7, 1513-1524.Blin,N. and Stafford,W. (1976) Nucleic Acids Res., 3, 2303-2308.Boss,J.M. and Strominger,J.L. (1984) Proc. Natl. Acad. Sci. USA, 81,
5199-5203.Briles,W.E., McGibbon,W.H. et al. (1950) Genetics, 35, 633-652.Collins,W.M., Briles,W.E. and Irwin,M.R. (1977) Immunogenetics, 5,
333 -343.Crone,M., Jensenius,J.C. et al. (1981) Scand, J. Immunol., 14, 591-597.Dausset,J. (1981) Science, 213, 1469-1474.Dodgson,J.B., Strommer,J. and Engel,J.D. (1979) Cell, 17, 879-887.Ewert,D.L. and Cooper,M.D. (1978) Immunogenetics, 7, 521-535.Ewert,D.L., Munchus,M.S., Chen,C.L. and Cooper,M. (1984) J. Im-
munol., 132, 2524-2530.Guillemot,F.P., Oliver,P.D., Peault,B. and Le Douarin,N. (1984)J. Exp.
Med., 160, 1803-1819.Guillemot,F., Turmel,P., Charron,D. and Le Douarin,N. (1986) J. Im-
munol., 137, 1251-1257.Hood,L., Steinmetz,M. and Malissen,B. (1983) Annu. Rev. Immunol., 1,529-568.
Howley,P.M., Israel,M.A., Law,M.F. and Martin,M.A. (1979) J. Biol.Chem., 254, 4876-4883.
Jotereau,F.V., Houssaint,E. and Le Douarin,N. (1980) Eur. J. Immunol.,10, 620-627.
Kaufman,J.F., Krangel,M.S. and Strominger,J.L. (1984) J. Biol. Chem.,259, 7230-7238.
Klein,J., Figueroa,F. and Nagy,Z.A. (1983) Annu. Rev. Immunol., 1,119- 142.
Larhammar,D., Schenning,L., Gustafson,K., Wiman,K., Claesson,L.,Rask,L. and Peterson,P.A. (1982) Proc. Natl. Acad. Sci. USA, 79,3687-3691.
Larhammar,D., Hanmmerling,U., Rask,L. and Peterson,P.A. (1985) J. Biol.Chem., 260, 14111-14119.
Lautenschlager,I., Taskinen,E., Inkinen,K., Lehto,V.P., Virtanen,I. andHayry,P. (1984) Cell. Immunol., 85, 191-200.
Le Douarin,N. (1969) Bull. Biol. Fr. Belg., 103, 435-452.Longenecker,B.M. and Mosmann,T. (1981) Immunogenetics, 13, 1-23.Malissen,M., Hunkapiller,T. and Hood,L. (1983) Science, 211, 750-753.Maniatis,T., Fritsch,E.F. and Sambrook,J. (1982) Molecular Cloning: a
Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Maxam,A.M. and Gilbert,W. (1980) Methods Enzymol., 65, 499-560.McMaster,G.K. and Carmichael,G.G. (1977) Proc. Natl. Acad. Sci. USA,
74, 4835-4838.Mengle-Gaw,L. and McDevitt,H.O. (1983) Proc. Natl. Acad. Sci. USA,
80, 7621-7625.Pazderka,F., Longenecker,B.M., Law,G.R.J. and Ruth,R.F. (1975) Im-
munogenetics, 2, 93-100.Rachwitz,H.R., Zehetner,G., Frischauf,A.M. and Lehrach,H. (1984) Gene,
30, 195-200.Rigby,P.W.J., Dieckermann,M., Rhodes,C. and Berg,P. (1977) J. Mol.
Biol., 113, 237-251.Sanger,F., Nicklen,S. and Coulson,A.R. (1977) Proc. Natl. Acad. Sci. USA,
74, 5463-5467.Southern,E. (1975) J. Mol. Biol., 98, 503-518.Toivanen,A. and Toivanen,P. (1977) J. Immunol., 118, 431-436.Tonnelle,C., De Mars,R. and Long,E.O. (1985) EMBO J., 4, 2839-2848.Vieira,J. and Messing,J. (1982) Gene, 19, 259-268.
Received on August 18, 1986; revised on January 18, 1988
la Recherche Scientifique, Institut National de la Sante et de la RechercheMedicale, Ministere de la Recherche et de la Technologie and Associationpour la Recherche contre le Cancer to C.A.
1039





























