Embed Size (px)
Citation preview

FLORIDA(ISSN 0015-4040)
ENTOMOLOGIST(An International Journal for the Americas)
Volume 67, No.2
TABLE OF CONTENTS
June, 1984
67th Annual Meeting of the Florida Entomological Society i
68th Annual Meeting-First Announcement ii
............. • '4
BENSCHOTER, C. A., AND P. C. WITHERELIr--Lethal Effects of SuboptimalTemperatures on Immature Stages of Anastrepha suspensa _.____ 189
ALI, A., D. M. SAUERMAN, AND J. K. NAYAR-Pathogenicity of Indus-trial Formulations of Bacillus thuringiensis serovar. israelensisto Larvae of Some Culicine Mosquitoes in the Laboratory ._ 193
BOCZEK, J., AND R. DAVIS-New Species of Eriophyid Mites (Acari:Eriophyidae) ._____________ 198
JONES, V. P., AND J. G. MORsE-A Synthesis of Temperature DependentDevelopmental Studies with the Citrus Red Mite, Panonychuscitri (McGregor) (Acari: Tetranychidae) . ...._. . .__ 213
GAGNE, R. J., AND G. M. BEAVERS-Contarinia spp. (Diptera: Cecidomyiidae) from Shoots of Slash Pine (Pinus elliotti Engelm.)with the Description of a New Species Injurious to Needles 221
LLOYD, J. E.-Evolution of a Firefly Flash Code .___________________________________ 228
LANDOLT, P. J.-Reproductive Maturation and Premating Period of thePapaya Fruit Fly, Toxotrypana curvicauda (Diptera: Teph-ritidae) . .__. . .___ 240
LIN, J., AND J. D. HARPER-Isolation and Culture of Entomophthoragammae, a Fungal Parasite of Noctuid Larvae _. . . ._.. 245
ZOEBISCH, T. G., D. G. SCHUSTER, AND J. P. GILREATH-Liriomyza tri-folii: Oviposition and Development in Foliage of Tomato andCommon Weed Hosts . . ... 250
WITHERELL, P. C.-Methyl Bromide Fumigation as a Quarantine Treatment for Latania Scale, Hemiberlesia lataniae (Homoptera:Diaspididae) ---.--.-..-- ._._._. .._._. __. .____________ 254
BROWER, J. H., AND L. D. CLINE-Response of Trichogramma pretiosumand T. evanescens to Whitelight, Blacklight or No-Light SuctionTraps .. . .__ ---------------________________________________ 262
ELVIN, M. K., AND P. E. SLODERBECK-A Key to Nymphs of SelectedSpecies of Nabidae (Hemiptera) in the Southeastern USA 269
Continued on Back Cover
Published by The Florida Entomological Society

FLORIDA ENTOMOLOGICAL SOCIETY
OFFICERS FOR 1983-84
President .. .__ . .. . . .... .__ .. C. W. McCoyPresident-Elect .__ . .__. _. ._ .. __ ._ __ . M. L. Wright, Jr.Vice-President . . .__ ._. ._ ...._.._....__.__. . . ._ .... .. . J. A. ReinertSecretary . .__ _.._.. _. __ .. .. .._. __. ._.. . . D. F. WilliamsTreasurer .. __.__.... .__ .__ _.. ._....._..._..__.. ._... .._.... ...._ A. C. Knapp
J. R. CassaniJ.L. Knapp
D. C. HerzogOther Members of the Executive Committee _... K. Lee
C. A. MorrisW. L. Peters
C. A. Musgrave Sutherland
PUBLICATIONS COMMITTEE
Editor ..._.. __ .. .__...._._ ..._._._ _....._.. ..__..__. . .__. C. A. Musgrave SutherlandAssociate Editors _. .._ __.__. . . . ._. . . .. . D. C. Herzog
O. Sosa, Jr.M. D. Hubbard
J. R. McLaughlinJ. B. Heppner
H. V. Weems, Jr.Business Manager _ _ _.._ __.._.. _._._._..__ .. A. C. Knapp
FLORIDA ENTOMOLOGIST is issued quarterly-March, June, September,and December. Subscription price to non-members is $20.00 per year inadvance, $5.00 per copy. Membership in the Florida Entomological Society,including subscription to Florida Entomologist, is $15 per year for regularmembership and $5 per year for students. Inquires regarding membershipand subscriptions should be addressed to the Business Manager, P. O. Box7326, Winter Haven, FL 33883-7326. Florida Entomologist is entered assecond class matter at the Post Office in DeLeon Springs and Gainesville, FL.
Authors should consult "Instructions to Authors" on the inside cover ofall recent issues while preparing manuscripts or notes. When submitting apaper or note to the Editor, please send the original manuscript, originalfigures and tables, and 3 copies of the entire paper. Include an abstract andtitle in Spanish, if possible. Upon receipt, manuscripts and notes are acknowledged by the Editor and assigned to an appropriate Associate Editorwho will make every effort to recruit peer reviewers not employed by thesame agency or institution as the authors (s). Reviews from individualsworking out-of-state or in nearby countries (e.g. Canada, Mexico, and others)will be obtained where possible.
Manuscripts and other editorial matter should be sent to the Editor,C. A. Musgrave Sutherland, 4849 Del Rey Blvd., Las Cruces, NM 88001.
This issue mailed June 30, 1984

THE 67th ANNUAL MEETING OF THEFLORIDA ENTOMOLOGICAL SOCIETY
The Florida Entomological Society will hold its 67th Annual meeting on24-27 July 1984 at the Holiday Inn, 6515 International Drive, Orlando FL32809; telephone-1- (305) -351-3500. Room rates will be $58.00, for single,double, triple, or quadruple.
Questions concerning the local arrangements should be directed to:
FREDERICK L. PETITT, ChairmanLocal Arrangements CommitteeFlorida Entomological SocietyWalt Disney World-Epcot Center-The LandP.O. Box 40Lake Buena Vista, Florida 32830 USAPhone: 1- (305) -827-7256
To present a paper, the tear out sheet must be postmarked and sent nolater than 15 MAY 1984, to:
JAMES A. REINERT, Program ChairmanFt. Lauderdale Research and Education CenterUniversity of Florida3205 S.W. College AvenueFt. Lauderdale, Florida 33314 USA
Eight minutes will be allotted for presentation of oral papers, with 2minutes for discussion. In addition, there will be a separate session formembers who may elect to present a Project (or Poster) Exhibit.
The 3 oral student papers judged to be the best on content and deliverywill be awarded monetary prizes during the meeting. Student authors mustbe Florida Entomological Society Members and must be registered for themeeting. Awards will be $125.00, 75.00 and 50.00.
The 3 student display presentations judged to be the best on content andpreparation will also be awarded monetary prizes during the meeting.Studentt authors must be Florida Entomological Society Members and mustbe registered for the meeting. Awards will be $125.00, 75.00 and 50.00.
Registration Schedule1 for Annual Meeting:
Preregistration Registration On Site
Full & Sustaining MembersStudent not in Student ContestStudent in Student ContestEach Extra Banquet Ticket
1 Each fee includes one banquet ticket.
$35.0018.0013.0010.00
$40.0020.0015.0010.00

68th ANNUAL MEETING-FLORIDA ENTOMOLOGICAL SOCIETY
First Announcement
In 1985, the 68th annual meeting of the Florida Entomological Societywill be held at the Ocho Rios Sheraton Hotel, Ocho Rios, Jamaica, W.I.Meeting dates are from Monday, 5 August to Thursday 8 August 1985. TheOcho Rios Sheraton is located on Mallards Beach on the north coast ofJamaica and is a complete resort facility offering excellent meeting facilitiesand day and night entertainment.
Emmer Travel, Inc. of Gainesville, FL has been designated the officialtravel agency of these meetings. They will make all of your travel arrangements including airline tickets and hotel reservations. The land/hotel package will be $145.00 per person (double occupancy) or $260.00 per person(single occupancy). This price includes:
*4 days/3 nights hotel accommodations*all Jamaican room taxes*all service charges on the rooms*porter service at the hotel*welcome rum swizzle on arrival at the hotel*transportation from/to the Montego Bay airport
Children under 17 stay FREE at the hotel when they stay in the roomwith 2 adults. Those under 17 must pay $22.00 for round-trip transportationfrom/to Montego Bay. Persons wishing to stay after the meetings will pay$70.00 per room per night, including taxes and service charges. Childrenunder 17 staying with adults after the meetings are not charged.
A $50.00 per person deposit will secure your reservations. The balancewill be due by 15 June'1985. All costs are fully refundable if the reservationis cancelled prior to 21 June 1985. Thereafter, a cancellation charge of $75.00per person on the land/hotel package will be levied.
Information on airfare rates, airline schedules and meeting registrationfees will be forthcoming soon.
To make reservations, send check for $50.00 to Emmer Travel, Inc., 2801SW Archer Road, Gainesville, FL 32608. Include your name, address, andr hone number. For further information, call Emmer Travel, Inc. toll free at(800) -342-2223 (Florida residents) or (800) -874-8487 (nationwide) or(904) -377-1222 (in Gainesville).
FIRST CALL FOR PAPERS
Submitted papers for the 68th annual meetings of the FES should be 8minutes long. Time alloted will be 10 minutes: 8 minutes for presentationand 2 minutes for discussion. Use the following tear sheet to submit a papertitle.

D Oral PresentationD Project Exhibit Session
(Formerly Poster Session)D Student Paper
DEADLINE: 15 May 1985
Return to : James A. ReinertFt. Lauderdale Research &
Education CenterUniversity of Florida3205 S.W. College AvenueFt. Lauderdale, FL 33314
1985 Meeting in Ochos Rios, Jamaica
Author's Name _
Title of Paper _
Affiliation and Address _
of the First
(Presenting) Author
Time Required for Presentation (Max. 10 min.)
Abstract: Must be Provided. Do not use more than 75 words.
SuggestionforEveningBullSession ~_~~ _

SLIDE POLICY FOR ANNUAL MEETINGS
The following slide policy will govern slide presentations at the AnnualMeetings. Only Kodak Carousel projectors for 2 x 2 slides will be available.However motion picture projectors will be available by special request tothe Local Arrangements Chairman prior to the date of the meeting.
Authors should keep slides simple, concise, and uncluttered with no morethan 7 lines of type on a rectange 2 units high by 3 units wide. All printedinformation should be readable to an audience of 300 persons.
A previewing room will be designated for author's use. A projectionistwill be available in the previewing room at least one hour before each session.Authors are expected to give the projectionist their slides in the previewingroom prior to each session. Slides will be returned to the authors after eachsession in the meeting room.
Authors are expected to organize their slides in proper order in theirpersonal standard Kodak Carousel slide tray (no substitution, please). Onlya few slide trays will be available in the previewing room from the projectionist for hardship cases. Slides in the tray should be in correct order starting with slot #1 of the tray and positioned correctly (position of slides to gointo tray: 1. upside down, and 2. lettering readable from this position upsidedown and from right to left). A piece of masking tape should be placed onthe slide tray by the author and the following information should be writtenon the tape: 1. author's name, 2. session date, and 3. presentation time.

Benschoter & Witherell: Suboptimal Temperatures 189
LETHAL EFFECTS OF SUBOPTIMAL TEMPERATURESON IMMATURE STAGES OF ANASTREPHA SUSPENSA
C. A. BENSCHOTER AND P. C. WTTHERELLl
Subtropical Horticulture Research UnitAgricultural Research Service, S & E, USDA
Miami, FL 33158 USA
ABSTRACT
Immature stages of Anastrepha suspensa (Loew) (Diptera: Tephritidae)were exposed for variable periods of time to temperatures ranging from 1.7to 15.6°C. Lethal effects varied inversely with temperature. Regressionequations were calculated and LT50 and LT95 values (lethal time for 50%and 95% mortality) are presented. Data points for all stages showed highlinearity when percentage mortalities were converted to probits and plottedagainst log-days (exposure time), providing correlation coefficients withinthe range of 0.934-0.999. Susceptibility to cold decreased with the age of theeggs at the temperatures and exposure periods tested. Less than 50% ofmature larvae were killed from exposure to 15.6°C for 28 days, but 7.2°C for7 days produced 99.8% mortality. Exposure to 15.6°C had essentially nolethal effect on pupae and 12.8°C for 28 days killed only 45.4% of them.Two-day-old pupae were more sensitive than older groups exposed to 4.4°C.By comparison, mortality of 2-day-old pupae exposed to this temperature for3 days was 99.8% while mortality of 10-day-old pupae exposed for 15 dayswas 99.9%. The order of susceptibility (LT95 ) of immature stages of A.S1lspensa exposed to 7.2°C was larvae> eggs> pupae.
RESUMEN
Estadios inmaduros de Anastrepha suspensa (Loew) (Diptera: Tephritidae) fueron sometidos durante diferente peri6dos de tiempo a temperaturas de 1.7 a 15.6°C. El efecto letal vari6 inversamente a la temperatura.Se calculo la ecuacion de regresi6n y los valores de TL50 y TL95 (tiempoletal para mortalidades de 50 y 95%). Todos los estadios mostraron altalinealidad cuando los porcentages de mortalidad se pasaron a probits y serepresentaron en gratica coeficientes de correlaci6n de rango de 0.934 a 0.999.La susceptibilidad al frio disminuy6 con la edad de los huevos para losperiodos de tiempo y exposicion probados. Menos de un 50% de larvasmaduras murieron al exponerlas a 15.6°C durante 28 dias, pero una exposicion a 7.2°C durante 7 dias produjo una mortalidad de 99.8%. La exposicion de las pupas a 15.6°C no tuvo efecto letal y a 12.8°C durante 28dias solo mato el 45.4%. Las pupas de dos dias de edad fueron mas sensiblesque el grupo de mayor edad expuestos a 4.4°C. En comparacion la mortalidadde pupas de dos dias de edad expuestas a esta temperatura durante tresdias fue de 99.8%, mientras que la mortalidad de pupas de 10 dias de edadexpuestas durante 15 dias fue de 99.9%. En orden de suceptibilidad al frio(TL95 ) de estadios inmaduros de A. suspensa expuestos a 7.2°C fue en ordendescendiente: larva, huevo, pupa.
Data concerning the lethal effects of suboptimal temperatures on theCaribbean fruit fly, Anastrepha suspensa (Loew) are lacking. Prescott and
lMethods Development Station, USDA, APHIS, PPQ, 13601 Old Cutler Road, Miami, FL33158 USA.

190 Florida Entomologist 67 (2) June, 1984
Baranowski (1971) determined the optimal temperatures for developmentof immature stages of this species over the range of 10-35°C.
The USDA-APHIS-PPQ Treatment Manual (1983) lists a cold treatmentfor fruit infested by Anastrepha species other than the Mexican fruit fly,Anastrepha ludens (Loew). This all-inclusive group presumably includes theCaribbean fruit fly even though there is no published information on thisspecies.
This paper investigates the efficacy of using low temperature to controlA. suspensa and reports the results of exposing immature stages of this flyto constant temperatures ranging from 1.7°-15.6°C.
MATERIALS AND METHODS
All insect stages used in these experiments were taken from the laboratory colony.
Eggs were collected from 1 to 2-wk-old laboratory stock flies. The numbers of eggs used in each test lot were estimated volumetrically with aneyedropper, then placed in random batches on small squares of wet blottingpaper. Each blotter was placed in a petri dish which was enclosed in a plasticbag to retain moisture. After cold exposure the blotter containing the eggswas placed on a small sponge which elevated the eggs above water in thebottom of a petri dish. The water was used to trap hatched larvae whichcrawled off the blotter. Dilute (0.03%) sodium benzoate solution was usedas a mold inhibitor in the moist environment required for eggs. Afterincubation at 25-27°C and 85+% RH for 4 days, larvae and unhatched eggswere counted with a dissecting microscope to determine the number of eggstreated and percent hatch. Eggs (2, 8, and 24 h old) were held at 1.7°C for1-10 days, at 4.4°C for 3-15 days, and at 7.2°C for 6-18 days, 3 replicates pertreatment.
Mature larvae (8 days old) were washed from laboratory diet and placedin lots of about 200 (measured volumetrically) into 29.6-ml (8 oz) waxedpaper cups containing moist vermiculite (6 ml H 2 0 per 50 ml vermiculite).Six small holes were punched in the lids of the cups to allow air exchange.
Larvae were exposed to 7.2, 10.0, 12.8 and 15.6°C for 7, 14, 21 and 28days, 8 replicates per treatment (except 3 replicates at 7.2°C). Larvae thatwere able to pupate and emerge as adult flies were counted as survivors.
Pupa selected randomly from a single lot were measured volumetricallyinto test aliquots of 175-200 and placed in wax paper cups described forlarvae. At 3, 6, and 10 days of age, they were exposed (3 replicates) to 7.2,10,12.8 and 15.6°C for 7, 14,21 and 28 days. In separate tests, pupae 2, 4, 6,8 and 10 days old were exposed about 300 per test unit (5 replicates) to4.4°C for 3, 7, 11 and 15 days. Survival was based on the number of adultflies that emerged following treatment.
Data were corrected for natural mortality in controls by Abbott's formula(1925). Then, percent mortality values were transformed to probits and exposure periods (days) to logarithms. The transformed data were used tocalculate time mortality regression equations and LT50 and LT95 values. Thelinear regression equation took the form of Y = a + bx where Y = probitmortality, A = y intercept, b = slope, and x = log days. Visual examinationof the plotted data (not presented) revealed that, in some instances, thelowest data points did not align with higher points which otherwise showed
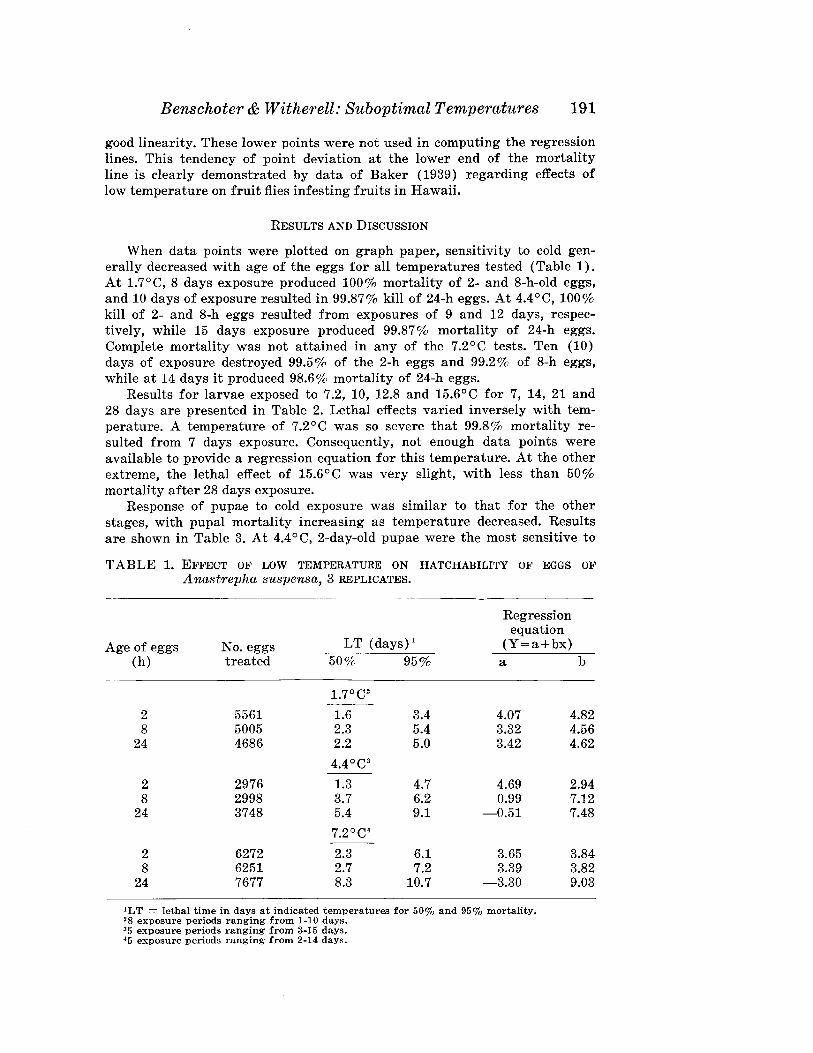
Benschoter & Witherell: Suboptimal Temperatures 191
good linearity. These lower points were not used in computing the regressionlines. This tendency of point deviation at the lower end of the mortalityline is clearly demonstrated by data of Baker (1939) regarding effects oflow temperature on fruit flies infesting fruits in Hawaii.
RESULTS AND DISCUSSION
When data points were plotted on graph paper, sensitivity to cold generally decreased with age of the eggs for all temperatures tested (Table 1).At 107°C, 8 days exposure produced 100% mortality of 2- and 8-h-old eggs,and 10 days of exposure resulted in 99.87% kill of 24-h eggs. At 4.4°C, 100%kill of 2- and 8-h eggs resulted from exposures of 9 and 12 days, respectively, while 15 days exposure produced 99.87% mortality of 24-h eggs.Complete mortality was not attained in any of the 7.2°C tests. Ten (10)days of exposure destroyed 99.5% of the 2-h eggs and 99.2% of 8-h eggs,while at 14 days it produced 98.6% mortality of 24-h eggs.
Results for larvae exposed to 7.2, 10, 12.8 and 15.6°C for 7, 14, 21 and28 days are presented in Table 2. Lethal effects varied inversely with temperature. A temperature of 7.2°C was so severe that 99.8% mortality resulted from 7 days exposure. Consequently, not enough data points wereavailable to provide a regression equation for this temperature. At the otherextreme, the lethal effect of 15.6°C was very slight, with less than 50%mortality after 28 days exposure.
Response of pupae to cold exposure was similar to that for the otherstages, with pupal mortality increasing as temperature decreased. Resultsare shown in Table 3. At 4.4°C, 2-day-old pupae were the most sensitive to
TABLE 1. EFFECT OF LOW TEMPERATURE ON HATCHABILITY OF EGGS OFA nastrepha suspensa, 3 REPLICATES.
Age of eggs(h)
No. eggstreated
LT (days)'
50% 95%
Regressionequation
(Y=a+bx)a b
3.45.45.0
4.76.29.1
2 55618 5005
24 4686
2 29768 2998
24 3748
2 62728 6251
24 7677
107°C'
1.62.32.2
4.4°C3
1.33.75.4
7.2°C4
2.3 6.12.7 7.28.3 10.7
4.07 4.823.32 4.563.42 4.62
4.69 2.940.99 7.12
-0.51 7.48
3.65 3.843.39 3.82
-3.30 9.03
'LT = lethal time in days at indicated temperatures for 50% and 95% mortality.28 exposure periods ranging from 1-10 days.35 exposure periods ranging from 3-15 days.45 exposure periods ranging from 2-14 days.
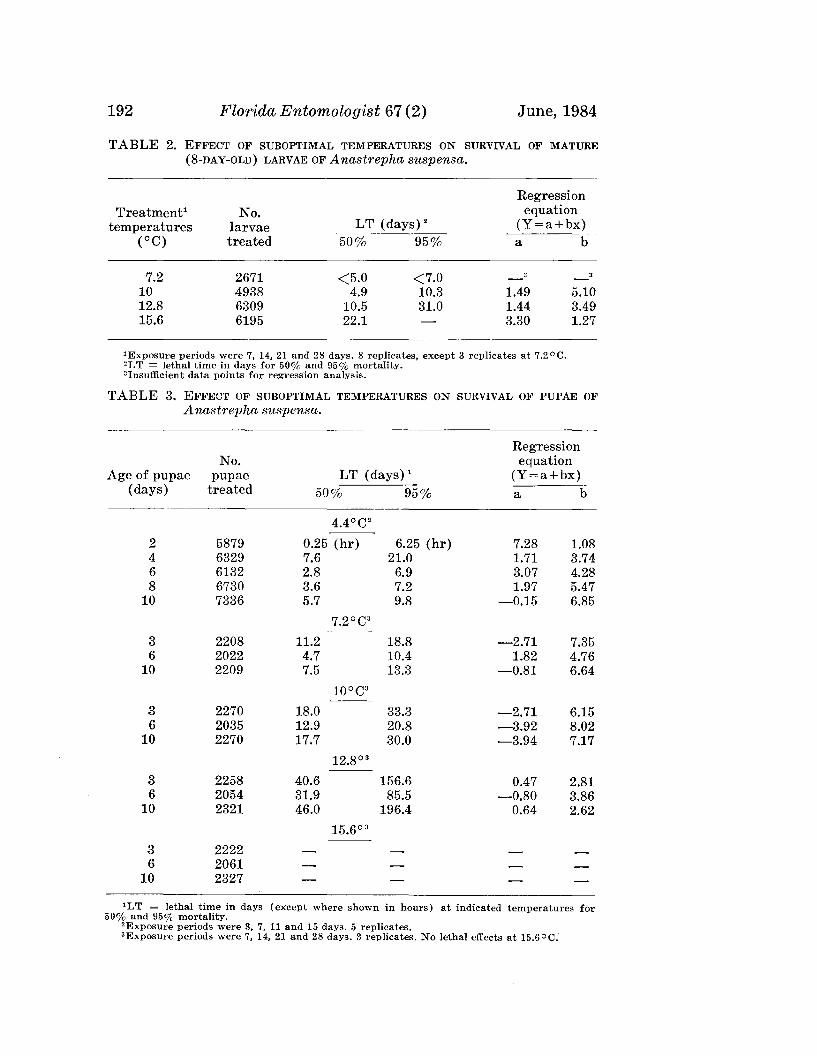
192 Florida Entomologist 67 (2) June, 1984
TABLE 2. EFFECT OF SUBOPTIMAL TEMPERATURES ON SURVIVAL OF MATURE(8-DAY-OLD) LARVAE OF Anastrepha suspensa.
RegressionTreatment1 No. equation
temperatures larvae LT (days)" (Y=a+bx)(OC) treated 50% 95% a b
7.2 2671 <5.0 <7.0 - 3 - 3
10 4938 4.9 10.3 1.49 5.1012.8 6309 10.5 31.0 1.44 3.4915.6 6195 22.1 3.30 1.27
1 Exposure periods were 7, 14, 21 and 28 days. 8 replicates, except 3 replicates at 7.2 ° C."LT = lethal time in days for 50% and 95% mortality.3Insufficient data points for regression analysis.
TABLE 3. EFFECT OF SUBOPTIMAL TEMPERATURES ON SURVIVAL OF PUPAE OFAnastrepha suspensa.
RegressionNo. equation
Age of pupae pupae LT (days) 1 (Y=a+bx)(days) treated 50% 95% a b
4.4°C"
2 5879 0.25 (hr) 6.25 (hr) 7.28 1.084 6329 7.6 21.0 1.71 3.746 6132 2.8 6.9 3.07 4.288 6730 3.6 7.2 1.97 5.47
10 7336 5.7 9.8 -0.15 6.85
7.2°C3
3 2208 11.2 18.8 -2.71 7.356 2022 4.7 lOA 1.82 4.76
10 2209 7.5 13.3 -0.81 6.64
10°C3
3 2270 18.0 33.3 -2.71 6.156 2035 12.9 20.8 -3.92 8.02
10 2270 17.7 30.0 -3.94 7.17
12.8°3
3 2258 40.6 156.6 0.47 2.816 2054 31.9 85.5 -0.80 3.86
10 2321 46.0 196.4 0.64 2.62
15.6°3
3 22226 2061
10 2327
1LT = lethal time in days (except where shown in hours) at indicated temperatures for50% and 95% mortality.
"Exposure periods were 3, 7, 11 and 15 days. 5 replicates.3Exposure periods were 7, 14, 21 and 28 days. 3 replicates. No lethal effects at 15.6 0 C.

Benschoter & Witherell: Suboptimal Temperatures 193
cold, their longevity being measured in hours rather than days. Comparingresults for pupae in the other age groups, 6-day-old pupae were uniformlymore susceptible to cold than younger or older individuals at all temperaturestested. Mortality of pupae in the 12.8°C tests was very moderate with lessthan 50% kill of 6-day old pupae exposed for 28 days. Response of pupaeat 15.6°C was negligible and was rated as "no effect".
Results of exposing immature stages of A. suspensa to suboptimal temperatures indicated that temperatures of 12.8°C or higher would have littlepractical value as a control measure against this insect. The order ofsusceptibility (LT95 ) of immature stages of A. suspensa exposed to 7.2°Cwas larvae> eggs> pupae.
The data points in all of the tests had high correlation coefficients (0.9340.999) indicating a high degree of linearity. Because of this, the regressionequations may be used to estimate exposure peribds (at a given temperature)for any desired level of mortality, or to predict mortalities at other (lower)temperatures not actually tested.
REFERENCES CITED
ABBOTT, W. S. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Ent. 18: 265-7.
BAKER, A. C. 1939. The basis for treatment of products where fruit flies areinvolved as a condition for entry into the United States. USDA Circ.No. 551. 7 p.
PRESCOTT, J. A. III, AND R. M. BARANOWSKI. 1971. Effects of temperature onthe immature stages of Anastrepha suspensa (Diptera: Tephritidae) .Florida Ent. 54: 297-303.
USDA-APHIS-PPQ Treatment Manual. 1983. Sec. VI, T107: 23-25.
.. . • • • • • • • • • • •PATHOGENICITY OF INDUSTRIAL FORMULATIONS OF
BACILLUS THURINGIENSIS SEROVAR. ISRAELENSISTO LARVAE OF SOME CULlCINE MOSQUITOESl
IN THE LABORATORY
ARSHAD ALIUniversity of Florida, IFAS, Agricultural Research and Education Center,
P. O. Box 909, Sanford, FL 32771 USAAND
DONALD M. SAUERMAN AND JAI K. NAYARFlorida Medical Entomology Laboratory
200 9th Street, S.E.Vero Beach, FL 32962 USA
ABSTRACT
In the laboratory, Teknar®, a flowable concentrate formulation, andBactimos® and Vectobac®, 2 wettable powder formulations of Bacillusthuringiensis serovar israelensis (Bti) were tested against laboratory main-
lDiptera: Culicidae.

Benschoter & Witherell: Suboptimal Temperatures 193
cold, their longevity being measured in hours rather than days. Comparingresults for pupae in the other age groups, 6-day-old pupae were uniformlymore susceptible to cold than younger or older individuals at all temperaturestested. Mortality of pupae in the 12.8°C tests was very moderate with lessthan 50% kill of 6-day old pupae exposed for 28 days. Response of pupaeat 15.6°C was negligible and was rated as "no effect".
Results of exposing immature stages of A. suspensa to suboptimal temperatures indicated that temperatures of 12.8°C or higher would have littlepractical value as a control measure against this insect. The order ofsusceptibility (LT95 ) of immature stages of A. suspensa exposed to 7.2°Cwas larvae> eggs> pupae.
The data points in all of the tests had high correlation coefficients (0.9340.999) indicating a high degree of linearity. Because of this, the regressionequations may be used to estimate exposure peribds (at a given temperature)for any desired level of mortality, or to predict mortalities at other (lower)temperatures not actually tested.
REFERENCES CITED
ABBOTT, W. S. 1925. A method of computing the effectiveness of an insecticide. J. Econ. Ent. 18: 265-7.
BAKER, A. C. 1939. The basis for treatment of products where fruit flies areinvolved as a condition for entry into the United States. USDA Circ.No. 551. 7 p.
PRESCOTT, J. A. III, AND R. M. BARANOWSKI. 1971. Effects of temperature onthe immature stages of Anastrepha suspensa (Diptera: Tephritidae) .Florida Ent. 54: 297-303.
USDA-APHIS-PPQ Treatment Manual. 1983. Sec. VI, T107: 23-25.
.. . • • • • • • • • • • •PATHOGENICITY OF INDUSTRIAL FORMULATIONS OF
BACILLUS THURINGIENSIS SEROVAR. ISRAELENSISTO LARVAE OF SOME CULlCINE MOSQUITOESl
IN THE LABORATORY
ARSHAD ALIUniversity of Florida, IFAS, Agricultural Research and Education Center,
P. O. Box 909, Sanford, FL 32771 USAAND
DONALD M. SAUERMAN AND JAI K. NAYARFlorida Medical Entomology Laboratory
200 9th Street, S.E.Vero Beach, FL 32962 USA
ABSTRACT
In the laboratory, Teknar®, a flowable concentrate formulation, andBactimos® and Vectobac®, 2 wettable powder formulations of Bacillusthuringiensis serovar israelensis (Bti) were tested against laboratory main-
lDiptera: Culicidae.

194 Florida Entomologist 67 (2) June, 1984
tained late 3rd and early 4th instars of Aedes taeniorhynchus (Wiedemann),Anopheles albimanus Wiedemann, Culex nigripalpus Theobald, Cx. restuansTheobald, Cx. salinarius Coquillett, Cx. quinquefasciatus Say, and Wyeomyiavanduzeei Dyar and Knab.
Aedes taeniorhynchus was susceptible (LC9o = 0.114 to 0.772 ppm) tothe test formulations. Culex nigripalpus was generally the most susceptibleto each formulation, followed by Cx. salinarius, Cx. quinquefasciatus, and Cx.'restuans. The 3 formulations of Bti were relatively ineffective against An.albimanus and Wy. vanduzeei. Considering the relative potencies of the testformulations of Bti, Bactimos® invariably was the most effective formulationand Teknar® the least against the species tested under laboratory conditions.
RESUMEN
En el laboratorio, Teknar®, una formulaci6n fluida concentrada, yBactimos® y Vectobac®, 2 formulaciones humedecibles de polvo de Bacillusthuringiensis serovar israelensis (Bti) fueron probadas contra tarde terceroy temprano cuarto estadio mantenidos en ellaboratorio de Aedes taeniorhynchus (Wiedemann), Anopheles albimanus Wiedemann, Culex nigripalpusTheobald, Cx. restuans Theobald, Cx. salinarius Coquillett, Cx. quinquefasciatus Say, and Wyeomia vanduzeei Dyar and Knab.
Aedes taeniorhynchus fue susceptible (LC9o = 0.114 a 0.772 ppm) a lasformulaciones probadas. Culex nigripalpus fue generalmente la mas susceptible a cada formulaci6n, seguida por Cx. salinarius, Cx. quinquefasciatus, y Cx. restuans. Las 3 formulaciones de Bti fueron relativamenteineficientes contra An. albimanus y Wy. vanduzeei. Considerando la relativapotencia de las formulaciones de Bti probadas, Bactimos® invariablementefue la formulaci6n mas efectiva, y Teknar® la menos efectiva contra lasespecies probadas bajo condiciones de laboratorio.
A particularly potent isolate of the entomogenous bacterium, Bacillusthuringiensis Berliner, 'was discovered by Goldberg and Margalit (1977) insoil of sewage lagoons in Israel. This isolate, corresponding to a new serotype H-14, was designated the variety israelensis by de Barjac (1978). Inthe past 6 years, B. thuringiensis serovar.israelensis (Bti) has been testedin the laboratory and field in different parts of the world against at least 70mosquito species (Anonymous 1982) and has proven to be a highly effectivemosquito larvicide. Some commercial producers have developed a variety ofincreasingly toxic and potent formulations of this biocide for use againstmosquitoes and some other aquatic pest and vector insects. At present, several wettable powder (WP), fiowable concentrate (FC), granular (G), andslow or sustained release formulations of the microbial insecticide are available and more are being formulated for testing and eventual marketing.Some of these formulations are currently registered for use against mosquitoes and simuliid black flies in the United States.
This paper reports laboratory evaluations of 3 industrial formulations ofBti against 7 species of mosquitoes maintained in the laboratory.
MATERIALS AND METHODS
The formulations evaluated were 2 WPs, Bactimos® (Biochem Products,Montchanin, DE) and Vectobac® (Abbott Laboratories, North Chicago, IL),

Ali et al.: Pathogenicity of Bt israelensis 195
.and a FC, Teknar® (Sandoz, Inc., San Diego, CA). The reported potenciesof Bactimos® and Vectobac®, respectively, were 3500 Aedes aegypti (AA)IUfmg, and 2000 AA IUfmg, while Teknar® contained 1500 AA IU fmg.
For laboratory assays, a mixture of late 3rd and early 4th instars ofAedes taeniorhynchus (Wiedemann), Anopheles albimanus Weidemann,Culex nigripalpus Theobald, Cx. restuans Theobald, Cx. salinarius Coquillett,Cx. quinquefasciatus Say, and Wyeomyia vanduzeei Dyar and Knab wereused. These species were maintained at the Florida Medical EntomologyLaboratory at Vero Beach, Florida.
The procedures of bioassays were generally the same as described byMulla et al. (1982). Twenty larvae were placed in a 120-ml disposable cupcontaining 100 ml of tap water (pH 6.8 ± 0.2). Distilled water (pH 6.9 ±0.2) was used for Wy. vanduzeei because of the high larval mortality of thisspecies occurring in tap water (Nayar et al. 1979). The Bti formulationswere suspended in distilled water by using a commercial blender for 1 to 2min to make a 1% stock suspension (wtfvol) of each formulation. Thesuspension was maintained with a magnetic stirrer for making subsequentserial dilutions and transfers for treatments. Each formulation was testedon 3 or more different occasions. Four or 5 triplicate concentrations of aformulation were applied each time and 3 cups were left untreated as controls. Stock suspensions and their serial dilutions were freshly prepared oneach occasion. After a 24-h exposure period in a controlled temperature (27± 1°C) holding room, larval mortality was recorded. The corrected mortality of a species at different concentrations of a formulation was subjected tolog-probit regression analysis.
RESULTS AND DISCUSSION
Larvae of Ae. taeniorhynchus were susceptible (LC90 = 0.114 to 0.772ppm) to the 3 test formulations of Bti (Table 1). The activity of theseformulations against Ae. taeniorhynchus did not correspond with their relative potencies. For example, Bactimos® was 2.33X more potent than Teknar®and 1.75X more potent than Vectobac®, but showed 6.77X and 2.98X betteractivity against Ae. taeniorhynchus than Teknar® and Vectobac®, respectively. Similarly, Vectobac® was 1.33X more potent than Teknar® butshowed 2.27X better activity than Teknar® against Ae. taeniarhynchus.
Among the species of Culex, Cx. nigripalpus was most susceptible to Bti,followed by Cx. salinarius, Cx. quinquefasciatus, and Cx. restuans. The LC
90values of Bactimos® against these Culex species ranged from 0.117 to 0.552ppm, while those of Vectobac® and Teknar® varied from 0.234 to 1.192 ppmand 0.265 to 1.721 ppm, respectively.
All 3 formulations of Bti were relatively ineffective against An. albimanuswith LC90 values ranging from 2.683 ppm (Bactimos®) to 11.446 ppm (Vectobac®). Similarly, Wy. vanduzeei also remained relatively unaffected; theLC90 value of Teknar® against this species was 4.706 ppm.
There is no previous laboratory work on the activity of Bti against larvaeof Cx. nigripalpus, Cx. salinarius, and Wy. vanduzeei. However, the levels oflarvicidal activity of the 3 formulations of Bti against Ae. taeniorhynchusand ex. quinquefasciatus observed in this study are generally similar to theactivity of comparable Bti formulations tested against the same species inearlier investigations (Ali et al. 1981, Mulla et al. 1982, Purcell 1981, Van
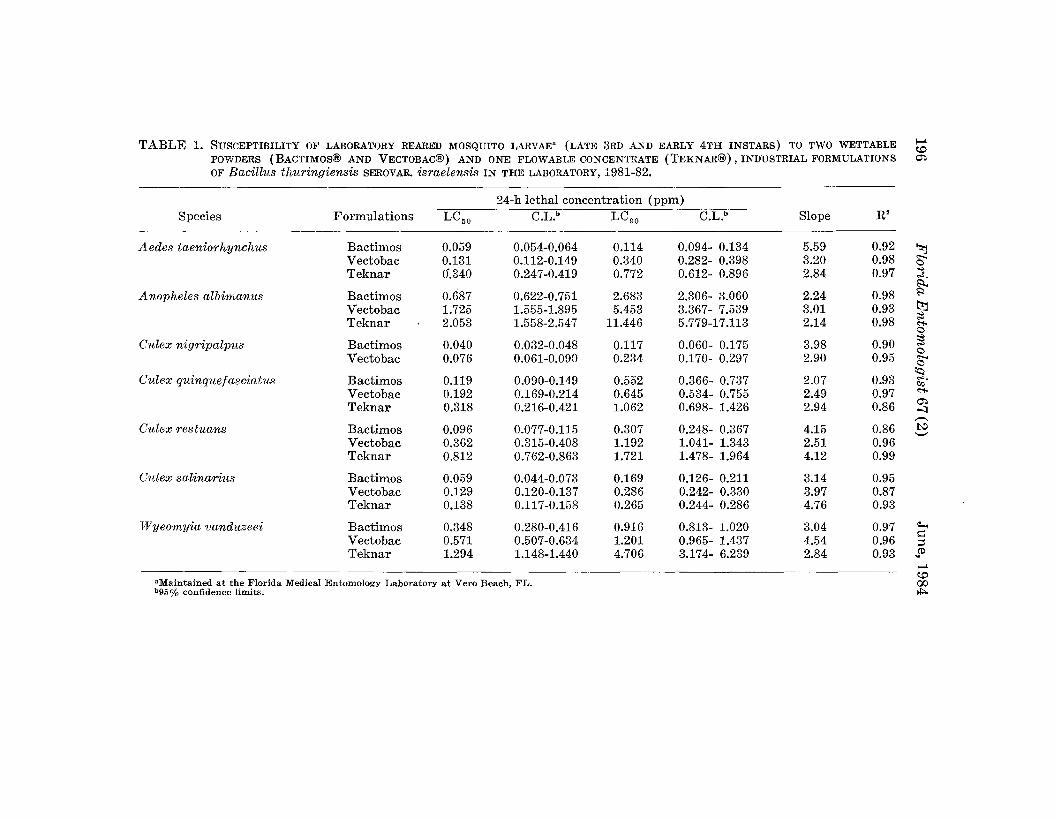
TABLE 1. SUSCEPTIBILITY OF LABORATORY REARED MOSQUITO LARVAE" (LATE 3RD AND EARLY 4TH INSTARS) TO TWO WETTABLE f-"~
POWDERS (BACTIMOS® AND VECTOBAC®) AND ONE FLOWABLE CONCENTRATE (TEKNAR®), INDUSTRIAL FORMULATIONS 0")
OF Bacillus thuringiensis SEROVAR. israelensis IN THE LABORATORY, 1981-82.
24-h lethal concentration (ppm)Species Formulations LC50 C.L.b LC BO C.L.b Slope R2
.Iiedes taeniorhynchus Bactimos 0.059 0.054-0.064 0.114 0.094- 0.134 5.59 0.92 ~Vectobac 0.131 0.112-0.149 0.340 0.282- 0.398 3.20 0.98 "....
CTeknar <J.340 0.247-0.419 0.772 0.612- 0.896 2.84 0.97 :;j.
:;:l...
Anopheles albimanus Bactimos 0.687 0.622-0.751 2.683 2.306- 3.060 2.24 0.98 S<
Vectobac 1.725 1.555-1.895 5.453 3.367- 7.539 3.01 0.93 tt:JTeknar 2.053 1.558-2.547 11.446 5.779-17.113 2.14 0.98 ::s
<:"i-cCulex nigripalpus Bactimos 0.040 0.032-0.048 0.117 0.060- 0.175 3.98 0.90 ~
cVectobac 0.076 0.061-0.090 0.234 0.170- 0.297 2.90 0.95 "....c
Culex quinquefasciatus~
Bactimos 0.119 0.090-0.149 0.552 0.366- 0.737 2.07 0.93 .,...C/;>
Vectobac 0.192 0.169-0.214 0.645 0.534- 0.755 2.49 0.97 <:"i-
Teknar 0.318 0.216-0.421 1.062 0.698- 1.426 2.94 0.860")-.;'J..-..
Culex restuans Bactimos 0.096 0.077-0.115 0.307 0.248- 0.367 4.15 0.86 !':l--Vectobac 0.362 0.315-0.408 1.192 1.041- 1.343 2.51 0.96Teknar 0.812 0.762-0.863 1.721 1.478- 1.964 4.12 0.99
Culex salinarius Bactimos 0.059 0.044-0.073 0.169 0.126- 0.211 3.14 0.95Vectobac 0.129 0.120-0.137 0.286 0.242- 0.330 3.97 0.87Teknar 0.138 0.117-0.158 0.265 0.244- 0.286 4.76 0.93
Wyeomyia vanduzeei Bactimos 0.348 0.280-0.416 0.916 0.813- 1.020 3.04 0.97 ~
Vectobac 0.571 0.507-0.634 1.201 0.965- 1.437 4.54 0.96 s::l::i
Teknar 1.294 1.148-1.440 4.706 3.174- 6.239 2.84 0.93 SOf-"~
"Maintained at the Florida Medical Entomology Laboratory at Vero Beach, FL. 00b95% confidence limits. Jo!:>.

Ali et al.: Pathogenicity of Bt israelensis 197
Essen and Hembree 1980) . The LC90 value of the 2 species for any comparable formulation was <1.0 ppm (Mulla et al. 1982).
It is obvious that An. albimanus and Wy. vanduzeei were relatively lesssusceptible to Bti under laboratory conditions. While Wy. vanduzeei mayhave inherent tolerance to Bti, the tolerance of Anopheles is probably due toits surface feeding habit (Mulla et al. 1982, Nugud and White 1982). Floating formulations of Bti might be more effective against Anopheles larvae.
From this study, it is evident that Bti offers a good potential for the control of Ae. taeniorhynchus, Cx. nigripalpus, Cx. quinquefasciatus, Cx.salinarius, and Cx. restuans. Among the formulations tested under laboratoryconditions, Bactimos® invariably was the most effective material againstthese species and Teknar® the least when their relative potencies are takeninto account.
Florida Agricultural Experiment Stations Journal Series No. 4777.
REFERENCES CITED
ALI, A., R. D. BAGGS, AND J. P. STEWART. 1981. Susceptibility of someFlorida chironomids and mosquitoes to various formulations ofBacillus thuringiensis serovar. israelensis. J. Econ. Ent. 74: 672-7.
ANONYMOUS. 1982. Data sheet on the biological control agent Bacillusthuringiensis serotype H-14 (de Barjac 1978). WHO/VBC/79.750,Rev. 1,46 p.
DE BARJAC, H. 1978. Une nouvelle variete de Bacillus thuringiensis trestoxique pour les moustiques: B. thuringiensis var. israelensis serotype14. C. R. Acad. Sci. (Paris) 286D: 797-800.
GOLDBERG, L. J., AND J. MARGALIT. 1977. A bacterial spore demonstratingrapid larvicidal activity against Anopheles sergentii, Uranotaeniaunguiculata, Culex univitattus, Aedes aegypti, and Culex pipiens.Mosq. News 37: 355-8.
MULLA, M. S., B. A. FEDERICI, AND H. A. DARWAZEH. 1982. Larvicidalefficacy of Bacillus thuringiensis serotype H-14 against stagnant watermosquitoes and its effects on nontarget organisms. Environ. Ent. 11:788-95.
NAYAR, J. K., P. A. PIERCE, AND J. S. HAEGER. 1979. Autogeny in Wyomyiavanduzeei in Florida. Entomologia expo appl. 25: 311-6.
NUGUD, A., AND G. WHITE. 1982. Evaluation of Bacillus thuringiensis serotype H-14 formulations as larvicides for Anopheles arabiensis (speciesB of the An. gambiae complex). Mosq. News 42: 36-40.
PURCELL, B. 1981. Effects of Bacillus thuringiensis var. israelensis on Aedestaeniorhynchus and some nontarget organisms in the salt marsh. Mosq.News 41: 476-84.
VAN ESSEN, F. W., AND S. C. HEMBREE. 1980. Laboratory bioassay ofBacillus thuringiensis against all instars of Aedes aegypti and Aedestaeniorhynchus larvae. Mosq. News 40: 424-31.

198 Florida Entomologist 67 (2) June, 1984
NEW SPECIES OF ERIOPHYID MITES(ACARI: ERIOPHYOIDEA)
JAN BOCZEKl AND ROBERT DAVISStored-Product Insects Research and Development Laboratory
Agricultural Research Service, USDASavannah, Georgia 31403 USA
ABSTRACT
Nine species of eriophyid mites are described, 3 from Poland: Aceria,ma,lva,cea,rum n.sp., Aculus ma,lvae n.sp., and Epitrimerus tanaceti n.sp.; 5from Brazil: Aculus pita,ngae n.sp., A. ca,ulifiorus n.sp., A. ca,ta,ppae n.sp.,A. sola,ni n.sp., and Phyllocoptes casea,ria,e n.sp.; and, 1 from Colombia:Ca,la,Ca,ruB guerreroi n.sp.
RESUMEN
Son descritas nueve especies de acaros eriofidos, tres de las cuales fueroncolectadas en Polonia: Aceria malvacearum n. sp.; Aculus malva,e n. sp.;Epitrimerus ta,na,ceti n. sp.; cinco en Brasil: Aculus pitanga,e n. sp.; A.caulifiorus n. sp.; A. ca,tappa,e n. sp.; A. solani n. sp. y Phyllocoptes ca,seariaen. sp.; y una en Colombia: Calaca,rus guerreroi n. sp.
The eriophyids (Eriophyoidea) are undoubtedly the most numerous of theplant feeding mites although, at present, only 1859 species are known (Daviset al., 1982).
Nine new species of eriophyid mites are described in this paper, 3 fromPoland: Aceria, ma,lvacearum n.sp., Aculus malvae n.sp., and Epitrimerustana,ceti n.sp.; 5 from Brazil: Aculus pitrangae n.sp., A. ca,ulifiorus n.sp.,A. ca,ta,ppae n.sp., A. solani n.sp., and Phyllocoptes caseariae n.sp.; and, 1from Colombia: Ca,lacarus guerreroi n.sp.
Type materials are deposited at the Department of Applied Entomology,Agricultural University of Warsaw, Warsaw, Poland.
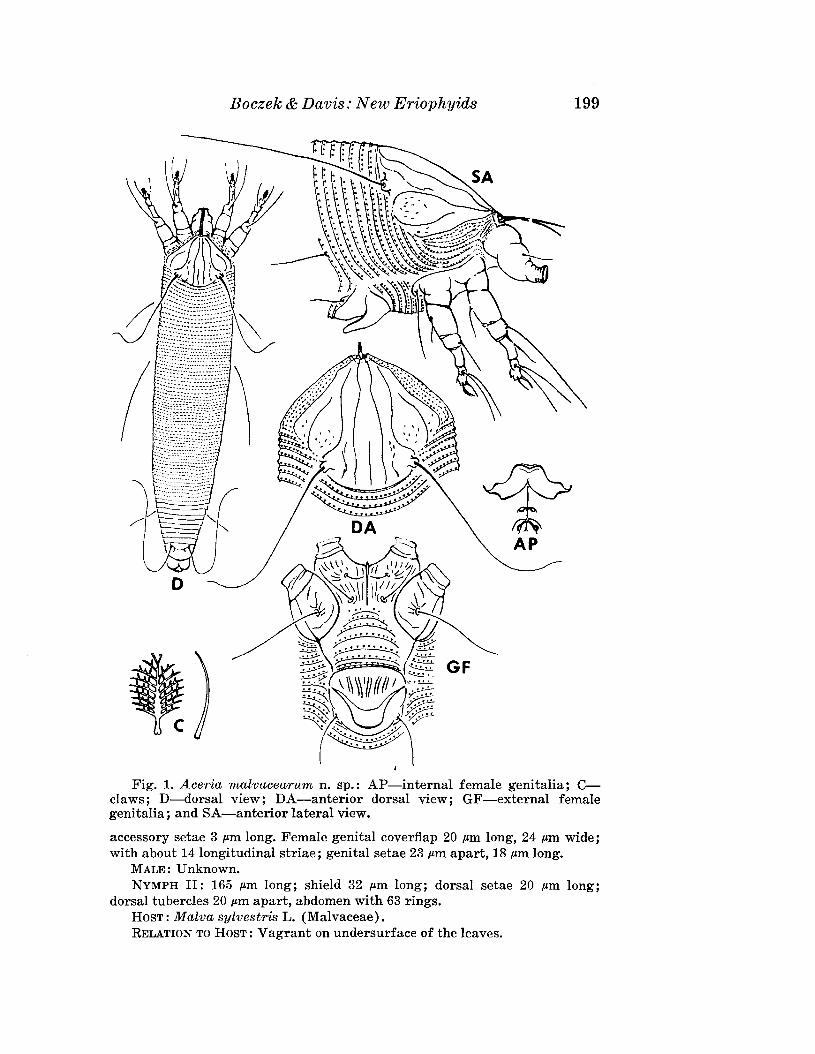
Aceria malvaceMum Boczek and Davis, NEW SPECIES(Fig. 1)
FEMALE: 258 p,m (range of 8 specimens 204-301 p,m) long; 56 p,m wide;wormlike; yellowish. Rostrum 19 p,m long; rostral seta 10 p,m long; chelicerae18 p,m long. Dorsal shield 38 p,m long, 40 p,m wide, without lobe over rostrum,with short median, 1 admedian line and 3 submedian lines on each side.Dorsal tubercles on rear shield margin; 26 p,m apart; with dorsal setae 44 p,m
long, reclinate and diverging. Foreleg 66 p,m long, tibia 8 p,m long; claw10 p,m long; feather-claw 9 p,m long, 8-9 rayed. Hindleg 50 p,m long; hind,coxal setae 28 p,m apart. First pair of coxae with longitudinal ornamentation. Thanosome with about 65 microtuberculate rings; microtubercles triangular. Lateral setae 40 p,m long, on sternite 13; 1st ventral setae 56 p,m
long, on sternite 26; 2nd ventral setae 30 p,m long; caudal setae 41 p,m long;
'Present Address: Department of Applied Entomology, Agricultural University of Warsaw.Nowoursynowska 166. Poland.
Boczek & Davis : New Eriophyids
~1AP
199
Fig. 1. Aceria malvacearum n. sp.: AP-internal female genitalia; Cclaws; D-dorsal view; DA-anterior dorsal view; GF-external femalegenitalia; and SA-anterior lateral view.
accessory setae 3 p,m long. Female genital coverflap 20 p,m long, 24 p,m wide;with about 14 longitudinal striae; genital setae 23 p,m apart, 18 p,m long.
MALE: Unknown.NYMPH II: 165 p,m long; shield 32 .am long; dorsal setae 20 p,m long;
dorsal tubercles 20 .am apart, abdomen with 63 rings.HOST: Malva sylvestris L. (Malvaceae).RELATION TO HOST: Vagrant on undersurface of the leaves.

200 Florida Entomologist 67 (2) June, 1984
TYPE MATERIAL. Holotype: female on slide, Poland, Warsaw-Lazienki,22-VI-1982, D. Zalewska. Paratypes (7): females on slides, same data asholotype.
This species is close to Aceria elacanthi Keifer and Can be distinguishedby the shield pattern, size and location of the dorsal tubercles and the shapeof the featherclaw. The dorsal shield A. elacanthi is subtriangular, with abroken median line and one pair of admedian lines; the dorsal tubercles arelarge, situated slightly ahead of rear shield margin; the featherclaw is4-rayed. In the new species dorsal shield is rhomboidal, with a completemedian line and 3 pairs of admedian lines; the dorsal tubercles are small,and on rear shield margin; the featherclaw is 8-9 rayed.
Aculus malvae Roczek and Davis, NEW SPECIES(Fig. 2)
FEMALE: 186 ~m (range of 12 specimens 180-203 ~m) long; 82 ~m wide;fusiform; yellowish-white. Rostrum 24 ,urn long; curved down; rostral seta6 ~m long. Shield 49 ~m long; 75 ~m wide, with broad anteriorly roundedlobe over rostrum, with minute granules; shield pattern of only 2 admedianlines curving laterad at both ends. Dorsal tubercles on rear margin of shield;45 ~m (32-51 ~m) apart; dorsal setae 18 ~m long, reclinate and diverging.Foreleg 57 pm long, with granulated tibiae and tarsi j tibia 11 /lm long, claw6 ~m long, knobbed featherclaw 5-rayed, 5 ~m long. Hindleg 54 ~m long, tibia9 #LID long. Both pairs of coxae ornamented. Hind coxal setae 26 pm apart.Thanosome with about 40 dorsal rings, generally microtuberculate; and about78 sternites with mictrotubercIes. Tergal microtubercles elliptical, ventralones almost rounded. Lateral setae 32 pm long, on sternite 56. Telosome with5 rings; setae 30 pID long; microtubercles elongated; caudal setae 47 pm long;accessory setae 2 pm long. Female genital coverflap 12 .urn long, 24 pm widewith 12 longitudinal striae; genital setae 17 ,urn apart, 20.am long.
MALE: 179 ~m long; shield 48 ~m long; J(enitalia 23 ~m wide.NYMPH II: 154 ~m long, shield 43 ~m long; dorsal setae 12 ~m long,
30 pm apart; abdomen with 54 microtuberculate tergites.HOST: Malva sylvestris L. (Malvaceae).RELATION TO HOST: Vagrant on undersurface of the leaves.TYPE MATERIAL. Holotype: Female on slide, Poland, Warsaw-Lazienki,
22-VI-1982, D. Zalewska. Paratypes (6): females (5) and male (1) on slides,same data as holotype.
This species is close to Aculus fockeui (Nalepa and Troussart) and canbe distinguished by the presence of shield spines, the shape of femalegenitalia and the featherclaw. "In A. fockeui the shield lobe has 2 distinctspines; the female genitalia has 16-18 longitudinal striae; and the featherclaw is 4-rayed. In the new species the shield lobe is without spines; thefemale genitalia has 12 striae; and the featherclaw is 5-rayed.
Epitrimerus tanaceti Boczek and Davis, NEW SPECIES(Fig. 3)
FEMALE: 172 ~m (range of 11 specimens 172-203 ~m) long; 63 ~m wide;straw-yellow; fusiform. Rostrum 17 ,urn long; chelicerae 20 flm long. Dorsalshield 50 ~m long; 56 ~m wide. Shield smooth, only laterally with some

Boczek & Davis: New Eriophyids 201
Fig. 2. Aculus mal'L'ae n. sp.: AP-internal female genitalia; C-clawsjDA-anterior dorsal view; GF-external female genitalia; GM-externalmale genitalia; S-lateral view; and SA-anterior lateral view.
granulation. Dorsal tubercles on rear margin of shield; 20 .urn apart, withdorsal setae 8 .urn long, pointed vertically and curved centrally. Foreleg 45 .urnlong; tibia 7 .urn long; claw 6 .urn long, knobbed; featherclaw 4-rayed, 5 ,urnlong. Hindleg 40 ,urn long. Both pairs of coxae with linear troughs. Hindcoxal setae 23 ,urn apart. Thanosome with 36 (36-41) tergites and about70 sternites. First 3 tergites covered with delicate elongated microtubercles,

202 Florida Entomologist 67 (2)
.~AP
June, 1984
Fig. 3. Epitrimerus tanaceti n. sp.: AP-internal female genitalia; Cclaws; D-dorsal view; DA-anterior dorsal view; ES-lateral view oftergite-sternite juncture; GF-external female genitalia; and SA-anteriorlateral view.
further tergites have such microtubercles only in central and lateral partsforming dorsally 3 longitudinal bands. Lateral setae 20 J.!m long on sternite14; 1st ventral setae 46 J.!m long, on sternite 31; 2nd ventral setae 14 J.!mlong, on sternite 49. Telosome with 5 microtuberculate rings and 22 J.!m longsetae. Caudal setae 22 J.!m long; accessory setae 3 J.!m long. Female genital

Boczek & Davis : New Eriophyids 203
coverfiap 13 Itm long, 24 Itm wide, with 12 longitudinal striae; genital setae14 Itm apart, 25 Itm long.
MALE: Unknown.NYMPH II: 105 Itm long; with microtuberculate rings.HOST: Tanacetum vulgare L. (Compositae).RELATION TO HOST: Vagrant on undersurface of the leaves.TYPE MATERIAL. Holotype: Female on slide, Poland, Warsaw-Powsin
Park, 13-VIII-1982, D. Zalewska. Paratypes (3): Females on slides, samedata as holotype.
This species is close to Epitrimerus jaceae Liro and can be distinguishedby the shape and pattern of the dorsal shield and the tergites. In E. jaceaethe dorsal shield is longer than wide, with a few broken admedian lines; thetergites are smooth. In the new species, the dorsal shield is as long as widewith one short admedian line and some granulations laterally; the tergiteshave microtubercles in central and lateral regions forming a central longitudinal band.
Aculus pitangae Boczek and Davis, NEW SPECIES(Fig. 4)
FEMALE: 185 Itm (range of 23 specimens 166-191Itm) long; 60 Itm wide;beige; fusiform. Rostrum 18 Itm long; rostral seta 6 Itm long. Shield subtriangular, with lobe over rostrum, 42 Itm (36-44 Itm) long, 65 Itm wide.Shield pattern with 2 distinct admedians, submedians forming a ridge whichparallels shield margin, beginning and ending at the base of dorsal tubercles.Dorsal tubercles on rear margin of shield: 41 Itm apart, dorsal setae 12 Itmlong, reclinate and slightly diverging. Foreleg 48 Itm long; genu 5 Itm longwith setae 22 Itm long; tibia 7 Itm long, with seta 2 Itm long. Claw 6 Itm long,knobbed; featherclaw 4-rayed, 6 Itm long. Hindleg 42 Itm long, genu 4 Itmlong, tibia 5 Itm long. Coxae ornamented, with small lines; hind coxae withdots; with setae 22 Itm apart. Thanosome with 29 (26-34) tergites and 63sternites. Tergites with 4 longitudinal bands of microtubercles reachingtelosome. Sternites with oval, minute microtubercles. Lateral setae 12 Itmlong, on sternite 15; 1st ventral setae 44 Itm long, on sternite 30; 2nd ventralsetae 11ltm long, on sternite 48. Telosome with 6 rings and 16 Itm long setae.Caudal setae 60 Itm long; accessory setae absent. Genital coverfiap 11 Itm(10-12 Itm) long; 18 Itm (16-19 Itm) wide; with 12 longitudinal striae; genitalsetae 12 Itm apart, 12 Itm long.
MALE: Unknown.HOST: Eugenia unifiora L. (Myrtaceae).RELATION TO HOST: Vagrants on undersurface of the leaves.TYPE MATERIAL: Female on slide, Brazil, Piracicaba, Sao Paulo, 19-IX
1980, J. Boczek. Paratypes (22): Females on slides, same data as holotype.This species is close to Aculops eugeniae Keifer (1977) described from
the same host plant from Florida. However, it differs in the shield pattern,in the tegrite microtubercles, and from the n. sp. in its relationship to thehost. In A. eugeniae the shield has a broken median and 4 curved admedianlines; the tergites have microtubercles their entire length; and the mitecauses bead galls on the upper surface of leaves. The new species has a dorsalshield with 2 pairs of admedian lines only; the tergites have microtubercles
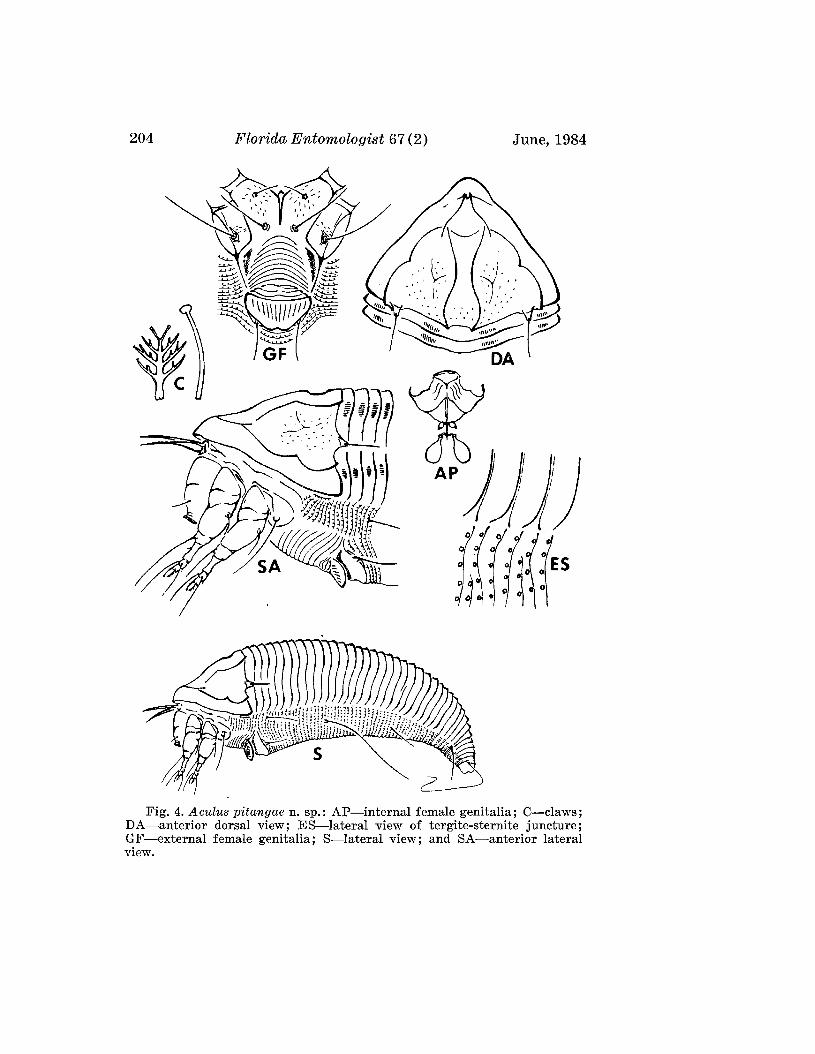
204 FloridrL Entomologist 67 (2) June, 1984
Fig. 4. Aculus pitangae n. sp.: AP-internal female genitalia; C-claws;DA-anterior dorsal view; ES-lateral view of tergite-sternite juncture;GF-external female genitalia; S-lateral view; and SA-anterior lateralview.
Boczek & Davis: New Eriophyids 205
forming 3 longitudinal bands; and they are vagrants on undersurface of theleaves not causing any apparent damage.
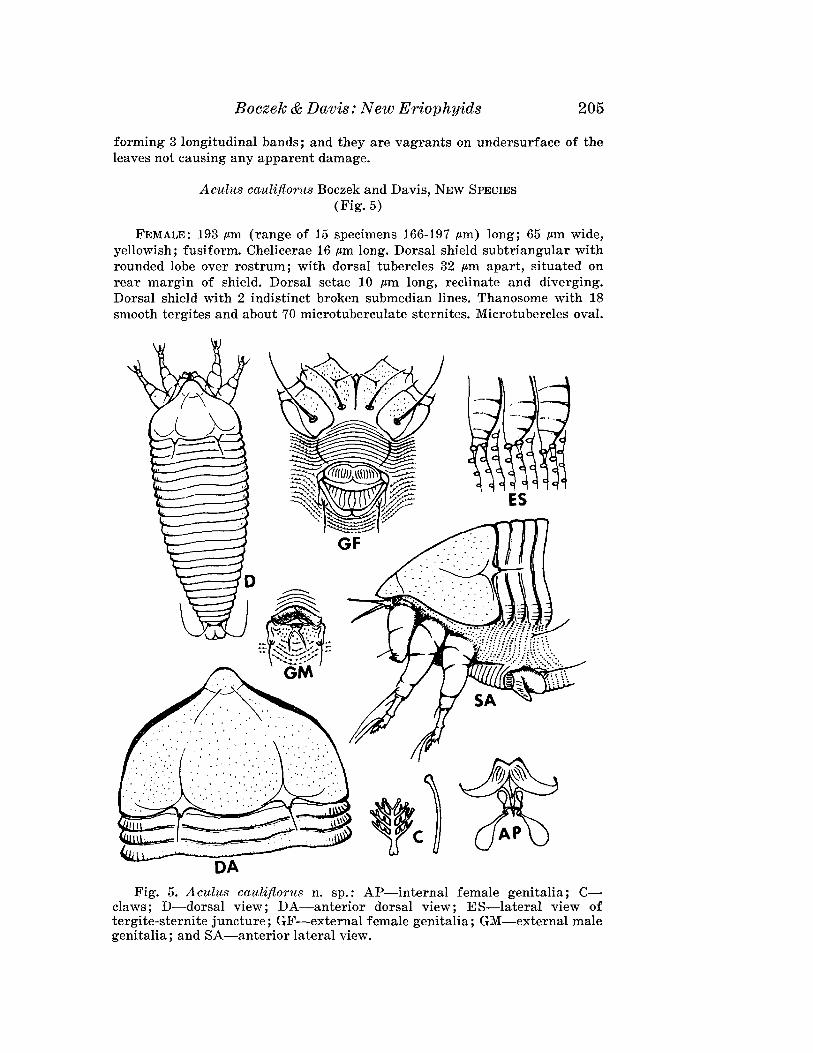
Aculus cauliflorus Boczek and Davis, NEW SPECIES(Fig. 5)
FEMALE: 193.um (range of 15 specimens 166-197 .urn) long; 65 .urn wide,yellowish; fusiform. Chelicerae 16 .urn long. Dorsal shield subtriangular withrounded lobe over rostrum; with dorsal tubercles 32 .urn apart, situated onrear margin of shield. Dorsal setae 10 pm long, reclinate and diverging.Dorsal shield with 2 indistinct broken submedian lines. Thanosome with 18smooth tergites and about 70 microtuberculate sternites. Microtubercles oval.
Fig. 5. Aculus cauliflorus n. sp.: AP-internal female genitalia; Cclaws; D-dorsal view; DA-anterior dorsal view; ES-Iateral view oftergite-sternite juncture; GF-external female genitalia; GM-external malegenitalia; and SA-anterior lateral view.

206 Florida Entomologist 67 (2) June, 1~84
Lateral setae 27 /.tm long on sternite 12; 1st ventral 56 pm long, on sternik30; 2nd ventral 8 I'm long on sternite 49. Caudal setae 48 I'm long; accessorysetae 4 pm long. Forelegs 50 ,am long; genu 6 .am long with seta 19 .am long;tibia 9 I'm long with seta 4 I'm long; claw 6 I'm long, knobbed; featherclaw4-rayed. Hindleg 45 I'm long; genu 6 I'm long with seta 6 I'm long. Telosomewith 5 rings and 20 I'm long setae. Female genital coverflap 16 I'm (16-20 I'm)long, 22 I'm (20-24 I'm) wide, with 12 longitudinal striae; genital setae 16 I'mapart, 12 I'm long. Coxae slightly granulated; hind coxal setae 23 I'm apart.
MALE: 154 I'm long; shield 40 I'm long; dorsal setae 6 I'm long; abdomenwith 23 tergites; genitalia 15 I'm wide; genital setae 14 I'm apart.
HOST: AcnistU8 cauliflorus Schott. (Solanaceae).RELATION TO HOST: Vagrants on undersurface of the leaves.TYPE MATERIAL. Holotype: Female on slide, Brazil, Piracicaba, Sao Paulo,
19-IX-1980, J. Boczek. Paratypes (14): Females (13) and male (1) on slides,same data as holotype.
This species is close to Aculus schlechtendali (Nalepa) and can be distinguished by the shape and the pattern of dorsal shield, presence of microtubercles on tergites and the shape of female genital coverflap. In A.schlechtendali the dorsal shield is semicircular, with median and someadmedian lines j the tergites are smooth; and the female genital cQverflaphas striae in one rank. In the new species the dorsal shield is subtriangularwith one pair of broken admedian lines; the tergites have microtubercIeslaterally; and the female genital coverflap has striae in 2 separate ranks.
Aculu8 catappae Boczek and Davis, NEW SPECIES(Fig. 6)
FEMALE: 166 I'm (range of 15 specimens 151-196 I'm) long; 62 I'm wide,amber; fusiform. Rostrum 20 .urn long, with seta 7 /LID long; chelicerae 16 jtm
long. Shield 41 I'm (32-42 I'm) long, 48 ~m wide. Dorsal shield with medianline fragmented and largely indistinct on posterior 1/3 of shield. Admedianlines complete in most specimens 'and forming a circular pattern on posterior1/3 of shield. First and 2nd submedian lines slightly fragmented but distinguishable from anterior margin to dorsal tubercles. Additional submedianlines fragmented and presenting appearance of a hairy pattern on shieldmargins. Dorsal tubercles 27 I'm apart, on rear margin of shield; dorsalsetae 12 I'm long, pointed reclinate and diverging. Foreleg 54 I'm long; genu6 I'm long with seta 21 I'm long; tibia 10 I'm long with seta 5 I'm long; claw7 I'm long, knobbed featherclaw 7-rayed. Hindleg 51 I'm long. Thanosomewith 52 (39-54) tergites and about 80 sternites. Both tergites and sternitesmicrotuberculate; microtubercles elliptical. Lateral setae 22 ,urn long, onsternite 13; 1st ventral setae 64 I'm long, on sternite 27; 2nd ventral setae16 I'm long, on sternite 48. Telosorne with 6 rings and 32 I'm long setae.Caudal setae 80 ,um long; accessory setae 4 jtm long. Female genital coverflap 13 I'm (10-13 I'm) long, 21 I'm (16-24 I'm) wide, with about 12 longitudinal ribs; genital seta 15 I'm apart, 20 I'm long.
MALE: 151 I'm long; shield 36 I'm long.HOST: Terminalia catappa L. (Combretaceae)RELATION TO HOST: Vagrant On undersurface of the leaves.TyPE MATERIAL. Holotype: Female on slide, Brazil, Piracicaba, Sao Paulo,
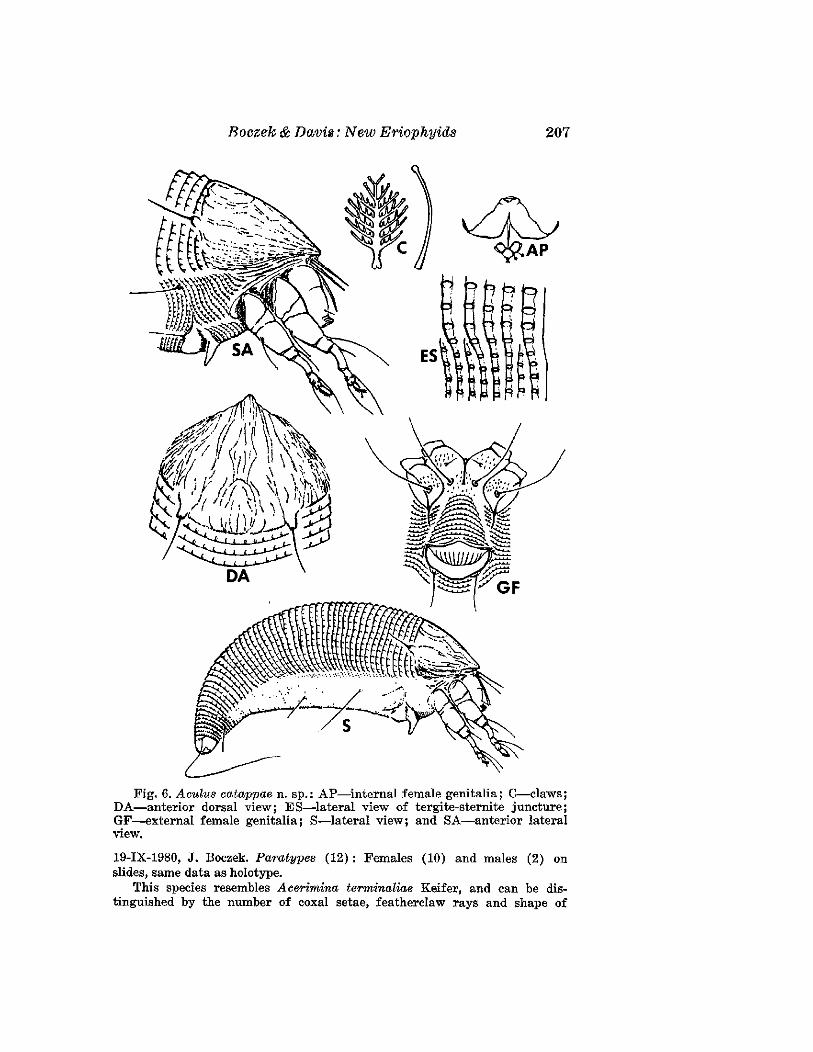
Roczek & Davis: New Eriophyids 207
Fig. 6. Aculus catappae n. sp.: AP-internal female genitalia; C-claws;DA-anterior dorsal view; ES-lateral view of tergite-sternite juncture;GF--external female genitalia; S-lateral view; and SA-anterior lateralview.
19-IX-1980, J. Boezek. Paratypes (12): Females (10) and males (2) onslides, same data as holotype.
This species resembles Acerimina terminaliae Keifer, and can be distinguished by the number of eoxal setae, featherelaw rays and shape of

208 Florida Entomologist 67 (2) June, 1984
genital coverflap. In A. terminaliae there is only one pair of forecoxal setae;the featherclaw is 6-rayed; and the female genital coverflap has diagonallylateral lines basally and is somewhat lobed laterally. In the new species thereare 2 pairs of forecoxal setae; the featherclaw is 7-rayed; and the genitalcoverflap has striae of even length, and no lateral lobes.
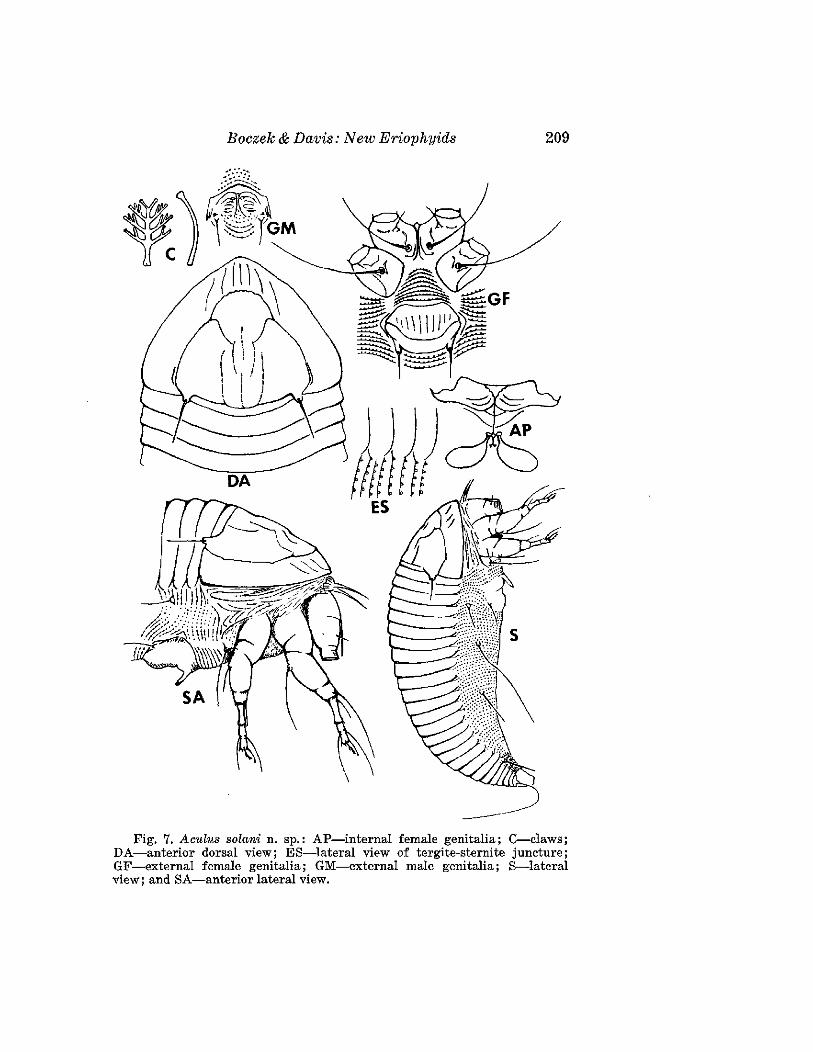
Aculus solani Boczek and Davis, NEW SPECIES(Fig. 7)
FEMALE: 231 ~m (range of 21 specimens 170-237 ~m) long; 56 ~m wide,amber; spindleforrn. Chelicerae 15 ,am long. Dorsal shield subtriangular, 42~m (40-44 ~m) long, with median line visible as 2 short fragments on posterior 1/2 of shield. Medial, admedial, and submedial lines complete onanterior 1/4 of shield. A ridge formed by portions of the submedial lines,beginning and ending at the base of the dorsal tubercles, parallels the shieldmargin. Dorsal tubercles 40 .am apart, on rear margin of shield; dorsal setae14,um long; reclinate and diverging. Forelegs 36 .am longj femur 10 ~m long;tibia 10 ~m long with setae 7 ~m long; claw 8 ~m long, slightly knobbed;featherclaw 6 ~m long, 4 rayed. Hind legs 32 ~m long. Forecoxae with 1stsetae 12 ~m long; 2nd setae 32 ~m long; hindcoxal setae 47 I'm long.Thanosome with 22 (22-24) smooth tergites and about 64 sternites. Sterniteswith oval microtubercles. Lateral setae 20 jLm long, on sternite 6; 1st lateralsetae 85 ~m long, on sternite 26; 2nd lateral 22 ~m long, on sternite 42.Telosome with 5 microtuberculate rings and setae 30 ~m long. Female genitalcoverflap 18 p.m long, 24 ,urn wide, with 16 furrows; genital setae 22 ,urnapart, 13 ~m long.
MALE: 200 p.m long; shield 44 /Lllllong, wiLh dvr;:;al seLae 14 ",m lung, with28 tergites. Male genitalia 16 ~m wide.
HOST: Solanum nigrum L. (Solanaceae).RELATION TO HOST: Vagrant on both surfaces of the leaves.TYPE MATERIAL. H o1otype: Female on slide, Brazil, Piracicaba, Sao Paulo,
18-IX-1980, J. Boczek. Paratypes (13): Females (11) and males (2) onslides, same data as holotype.
This species is close to Aculus rhamnivagrans (Keifer), and can be distinguished by the shield shape and the internal female genitalia. In A.rhamnivagrans the shield is subtriangular with a long and wide lobe overrostrum; the spermathecae are circular. In the new species the shield issemicircular with a very broad, short lobe over rostrum; the spermathecaeare oval.
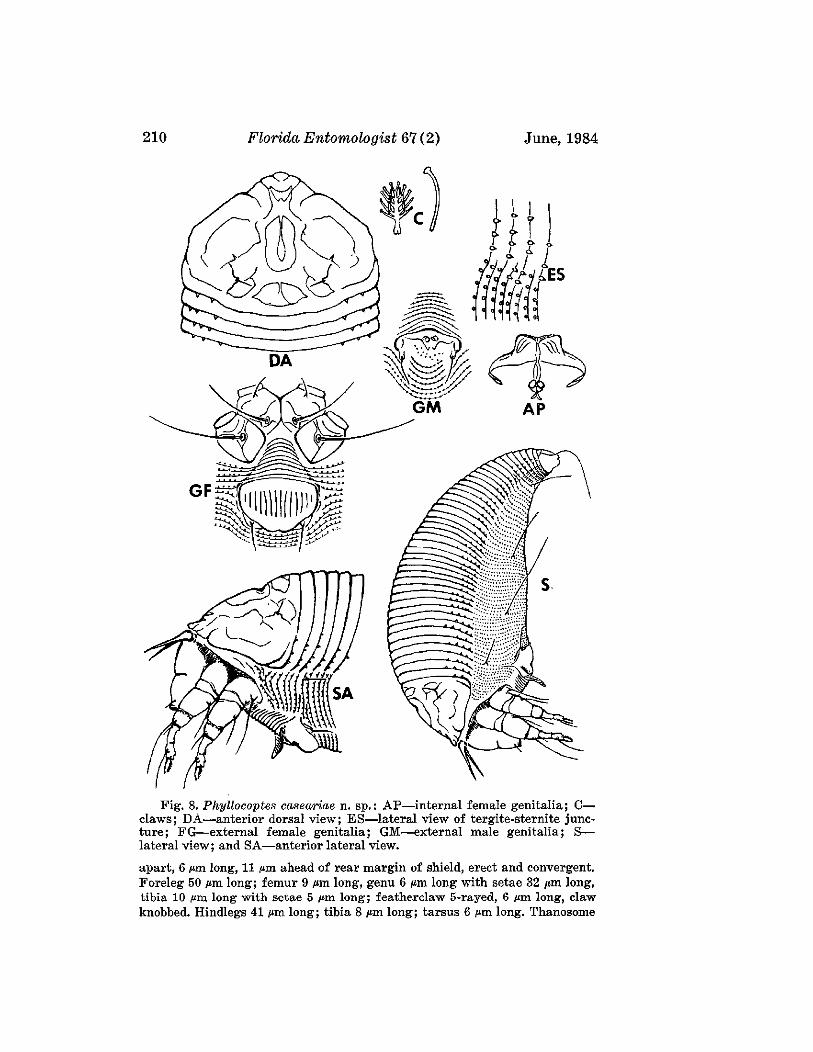
Phyllocoptes caseariae Boczek and Davis, NEW SPECIES(Fig. 8)
FEMALE: 190 ~m (range of 16 specimens 180-220 ~m) long; 67 ~m wide;72 I'm thick; amber; spindleform. Rostrum 24 ~m long, curved downwardobliquely. Chelicerae 17 ~m long. Shield 45 I'm (44-48 ~m) long, with shortlobe over rostrum; shield oval, surface with two declivities along medianline at anterior of shield and near its midpoint. Admedial lines visible surrounding both declivities. Submedial lines extending from anterior marginto base of dorsal tubercles. Overall surface appearance is that of severalfractured irregular placed rectangles. Dorsal tubercles 30 (29-32 ~m)
Boczek & Davis: New Eriophyids 209
J) )#!ff {f
ES
DA
Fig. 7. Aculus solani n. sp.: AP-internal female genitalia; C-claws;DA-anterior dorsal view; ES-lateral view of tergite-sternite juncture;GF-external female genitalia; GM-external male genitalia; S-lateralview; and SA-anterior lateral view.
210 Florida Entomologist 67 (2) June, 1984
Fig. 8. Phyllocoptes caseariae n. sp.: AP-internal female genitalia; Cclaws; DA-anterior dorsal view; ES-Iateral view of tergite-sternite juncture; FG-external female genitalia; GM-external male genitalia; Slateral view; and SA-anterior lateral view.
apart, 6 ~m long, 11 ~m ahead of rear margin of shield, erect and convergent.Foreleg 50 I'm long; femur 9 I'm long, genu 6 I'm long with setae 32 I'm long,tibia 10 ,um long with setae 5 ,urn long; featherclaw 5·rayed, 6 p:m long, clawknobbed. Hindlegs 41 I'm long; tibia 8 I'm long; tarsus 6 I'm long. Thanosome

Boczek & Davis: New Eriophyids 211
with 35 (31-36) broader tergites and about 59 sternites. Lateral ends oftergites with spiny microtubercles and sternites with rounded microtubercles.Lateral setae 12 ,urn long on sternite 5; 1st ventral seta 44 ,urn long onsternite 37. Telosome with 5 rings and 20 ,urn long setae. Caudal setae about64 ,urn long, accessory setae absent. Female genital coverflap 18 ,urn (18-24,urn) wide, with 14-16 elongate furrows; setae 22 ,urn apart, 12 ,urn long.
MALE: 152 ,urn long; dorsal shield 40 ,urn long; dorsal setae 5 ,urn long,24 ,urn apart; 8 ,urn from rear margin of shield; chelicerae 14 ,urn long; malegenitalia 18 ,urn wide, genital setae 7 ,urn long.
NYMPH II: 140 ,urn long; 56 ,urn thick; shield 30 ,uum long; dorsal setae4 ,urn long; dorsal tubercles 9 ,urn apart, with 40 rings with spiny microtubercles.
HOST: Casearia sylvestris Sw. (Flacourtiaceae).RELATION TO HOST: Vagrants on undersurface of leaf.TYPE MATERIAL. H olotype: Female on slide, Brazil, Piracicaba, Sao Paulo,
19-IX-1980, J. Boczek. Paratypes (14) : Females (13) and male (1) on slides,same data as holotype.
This species is close to Phyllocoptes abaenus Keifer and can be distinguished by the shield shape, the shape of microtubercles of the tergites andby the number of featherclaw rays. In P. abaenus the shield has a long broadlobe; the sternites have oval microtubercules their entire length; and thefeatherclaw is 4-rayed. In the new species the shield is almost semicircular;the sternites have spiny microtubercles only laterally; and the featherclaw is5-rayed. This is the first eriophyid mite described from the plant familyFlacourtiaceae.
Calacarus guerreroi Boczek and Davis, NEW SPECIES(Fig. 9)
FEMALE: 196,um (range of 30 specimens 166-196 ,urn) long; 76 ,urn wide;straw yellow; fusiform. Rostrum 36 ,urn long; seta 20 ,urn long. Chelicerae40 ,urn long. Dorsal shield 50 ,urn long; 72 ,urn wide. Shield pattern in form ofnetwork of ridges and granulation laterally. Shield lobe over rostrum wide,short, rounded anteriorly. Dorsal tubercles and setae absent. Foreleg 66 ,urnlong, genu 5 ,urn long with seta 36 ,urn long; tibia 15 ,urn long, with seta 2 ,urnlong. Claw 6,um long, knobbed featherclaw 6 ,urn long, 4-rayed. Hindleg 58 ,urnlong; genu 4 ,urn long; tibia 12 ,urn long. Thanosome with 54 (49-64) tergitesand about 97 sternites. Tergites forming central ridge and two adcentralridges with wax points. Microtubercles present on all sternites and usuallyon some proximal tergites laterally. Telosome of 8 rings and setae 21 ,urnlong; caudal setae 98 ,urn long; accessory setae absent. Telosomal microtubercles elongated. Lateral setae 36 ,urn long, on sternite 19; 1st ventralsetae 40 ,urn long, on sternite 47; 2nd ventral setae 36 ,urn long, on sternite80. Female genital coverflap 22 ,urn long, 28 ,urn wide; epigynium with about12 longitudinal striae; genital setae 18 ,urn apart, 12 ,urn long.
MALE: 154 ,urn long; with dorsal shield 56 ,urn long; genitalia 18 ,urn wide.HOST: Manihot esculenta Crantz (Euphorbiaceae) (Cassava, Tapioca).RELATION TO HOST: On the uppersurface of leaf; they prefer the basal
leaves of the plant. Infested leaves curl upward, and gradually shrink.TYPE MATERIAL. H olotype: Female on slide, Colombia, Cali, 2-VIII-1982,
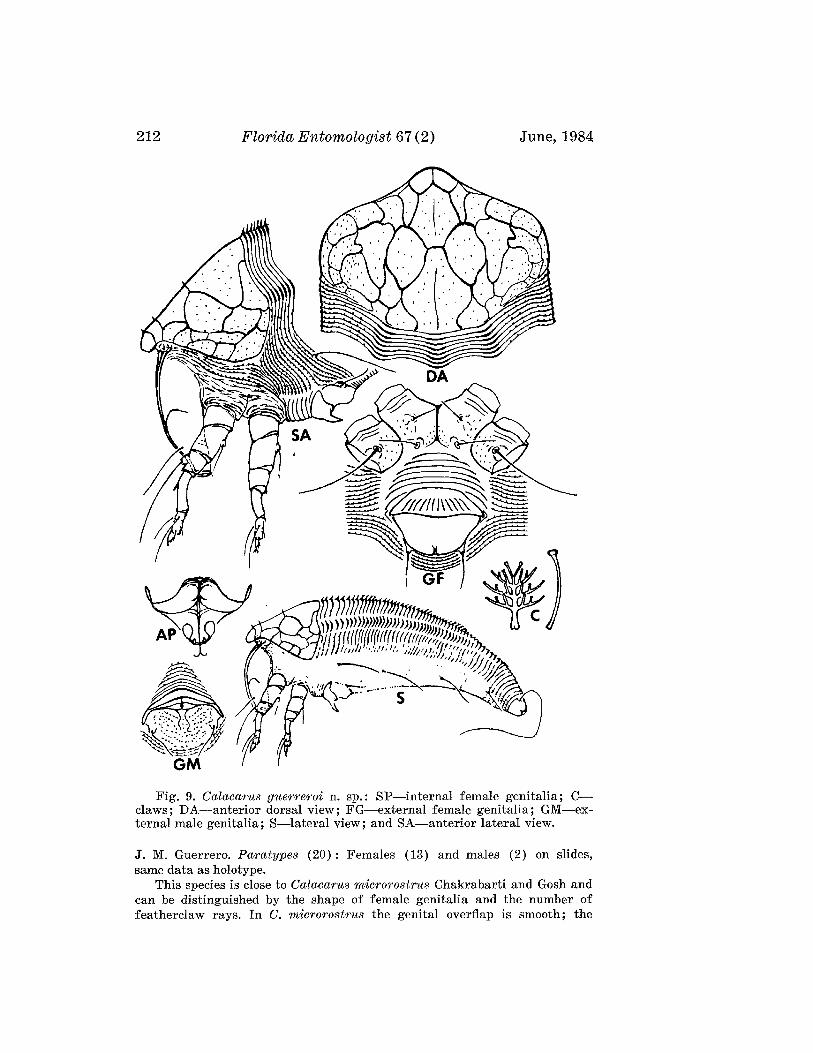
212 Florida Entomologist 67 (2) June, 1984
Fig. 9. Calacarus guerreroi n. sp.: SP-internal female genitalia; Cclaws; DA-anterior dorsal view; FG-external female genitalia; GM-external male genitalia; S-lateral view; and SA-anterior lateral view.
J. M. Guerrero. Paratypes (20): Females (13) and males (2) on slides,same data as holotype.
This species is close to Calacarus microrostrus Chakrabarti and Gosh andcan be distinguished by the shape of female genitalia and the number offeatherclaw rays. In C. microrostrus the genital overflap is smooth; the

Boczek & Davis: New Eriophyids 213
featherclaw is 7-rayed. In the new species the genital coverflap has 12 longitudinal striae; the featherclaw is 4-rayed.
We are pleased to name this mite for Mr. Jose Maria Guerrero, CentroInternacional de Agricultura Tropical, Apartado Aereo 6713, Cali, Colombia.This is the first species of eriophyid mite found on cassava (Bellotti &Schoonhoven 1978), an economical subsistence crop plant grown in about 90developing countries.
ACKNOWLEDGEMENT
We would like to acknowledge the assistance of Dr. C. H. W. Flechtman,ESALQ Zoologia, Piracicaba SP, Brazil, in collecting the Brazilian mites;and of Miss Danuta Zalewska, laboratory assistant, Department of AppliedEntomology, Agricultural University of Warsaw, Warsaw, Poland, for drawing the mites.
REFERENCES CITED
BELLOTTI, A., AND A. VAN SCHOONHOVEN. 1978. Mite and insect pests ofcassava. Ann. Rev. Ent. 23: 39-67.
DAVIS, R., C. H. W. FLECHTMAN, J. BOCZEK, AND H. BARKE. 1982. Catalogueof eriophyid mites (Acari: Eriophyoidea). Agric. Univ. WarsawPress, Warsaw, Poland. 254 p.
KEIFER, H. H. 1977. Eriophyid studies C-13. California Dept. of Agric., Jan.20,1977.24 p.
.. . • • • • • • • • • • . ~
A SYNTHESIS OF TEMPERATURE DEPENDENTDEVELOPMENTAL STUDIES WITH THE
CITRUS RED MITE, PANONYCHUS CITRI (McGREGOR)(ACARI: TETRANYCHIDAE)
V. P. JONES AND J. G. MORSEDept. of Entomology
University of CaliforniaRiverside, CA 92521 USA
ABSTRACT
A literature review revealed 12 temperature-humidity studies performedon the citrus red mite, Panonychus citri (McGregor) (Acari: Tetranychidae). An analysis of these studies using a heat unit scale indicated that theywere in surprisingly close agreement especially in view of the different hostplants and experimental techniques utilized. Theoretical thresholds for development, degree days necessary to complete development, and temperaturedependent mortality for the egg, immature and adult female stages arepresented. This synthesis implies that further research with the citrus redmite is needed in the area of variable temperature developmental rates,fecundity and mortality rates.
RESUMEN
Una revision de la bibliografia revelo 12 estudios de temperatura-

Boczek & Davis: New Eriophyids 213
featherclaw is 7-rayed. In the new species the genital coverflap has 12 longitudinal striae; the featherclaw is 4-rayed.
We are pleased to name this mite for Mr. Jose Maria Guerrero, CentroInternacional de Agricultura Tropical, Apartado Aereo 6713, Cali, Colombia.This is the first species of eriophyid mite found on cassava (Bellotti &Schoonhoven 1978), an economical subsistence crop plant grown in about 90developing countries.
ACKNOWLEDGEMENT
We would like to acknowledge the assistance of Dr. C. H. W. Flechtman,ESALQ Zoologia, Piracicaba SP, Brazil, in collecting the Brazilian mites;and of Miss Danuta Zalewska, laboratory assistant, Department of AppliedEntomology, Agricultural University of Warsaw, Warsaw, Poland, for drawing the mites.
REFERENCES CITED
BELLOTTI, A., AND A. VAN SCHOONHOVEN. 1978. Mite and insect pests ofcassava. Ann. Rev. Ent. 23: 39-67.
DAVIS, R., C. H. W. FLECHTMAN, J. BOCZEK, AND H. BARKE. 1982. Catalogueof eriophyid mites (Acari: Eriophyoidea). Agric. Univ. WarsawPress, Warsaw, Poland. 254 p.
KEIFER, H. H. 1977. Eriophyid studies C-13. California Dept. of Agric., Jan.20,1977.24 p.
.. . • • • • • • • • • • . ~
A SYNTHESIS OF TEMPERATURE DEPENDENTDEVELOPMENTAL STUDIES WITH THE
CITRUS RED MITE, PANONYCHUS CITRI (McGREGOR)(ACARI: TETRANYCHIDAE)
V. P. JONES AND J. G. MORSEDept. of Entomology
University of CaliforniaRiverside, CA 92521 USA
ABSTRACT
A literature review revealed 12 temperature-humidity studies performedon the citrus red mite, Panonychus citri (McGregor) (Acari: Tetranychidae). An analysis of these studies using a heat unit scale indicated that theywere in surprisingly close agreement especially in view of the different hostplants and experimental techniques utilized. Theoretical thresholds for development, degree days necessary to complete development, and temperaturedependent mortality for the egg, immature and adult female stages arepresented. This synthesis implies that further research with the citrus redmite is needed in the area of variable temperature developmental rates,fecundity and mortality rates.
RESUMEN
Una revision de la bibliografia revelo 12 estudios de temperatura-

214 Florida Entomologist 67 (2) June, 1984
humedad sobre Panonychu8 citri (McGregor) (Acari: Tetranychidae). Unamilisis de estos estudios usando una unidad de calor de escala indico queestaban en sorprendente acuerdo, especialmente en vista de las diferentesplantas hospederas y tecnicas experimentales utilizadas. Limite teorico dedesarrollo, grados diarios necesarios para completar el desarrollo, mortalidadde los huevos dependiente de la temperatura, y las etapas de los inmadurosy las hembras adultas son presentados. Esta sintesis implica que mas investigaci6n sobre P. citri es necesaria en el area de grados de desarrollospor temperatura variable, fecundidad, y grados de mortalidad.
The citrus red mite, Panonychu8 citri (McGregor), is one of the mostimportant pests of citrus worldwide (Jeppson et al. 1975). In Californiaalone, the citrus red mite was estimated to cause an economic crop loss ofapproximately 15.9 million dollars in 1977 (Buxton 1977). Its importancehas led to a number of studies dealing with the relationship between temperature and the rate of development (Table 1). However, none of thesestudies have defined parameters, such as degree days or developmentalthresholds, which are necessary for temperature dependent modeling efforts.In this review, we provide a summary and analysis of these studies anddefine several modeling parameters from a selected subset of all work done.
MATERIALS AND METHODS
Data were initially grouped into 3 categories which corresponded to theegg, immature, and adult life stages. The data were summarized (Table 2)and experimental methods reviewed. Experiments were eliminated whichutilized sub-optimal host plants, such as papaya (Maity and Chakrabarti1978), or involved temperature regimes which were not sufficiently definedfor this type of analysis (English and Turnipseed 1941). Data from studiesmarked with an * in Ta.ble 2 were not used in the analysis.
Since there was apparently no high temperature mediated decrease indevelopmental rates, data for t4e egg and immature stages were graphed as(developmental time) -1 vs. (temperature). Linear regression was used toobtain least squares estimates of the slope and x-intercept. Degree days fordevelopment and theoretical developmental thresholds were determined as(slope)-l and (x-intercept), respectively (Arnold 1959).
If more than one humidity was tested per temperature, humidities whichproduced the fastest development time were selected. Only data for the femalestage were considered.
RESULTS AND DISCUSSION
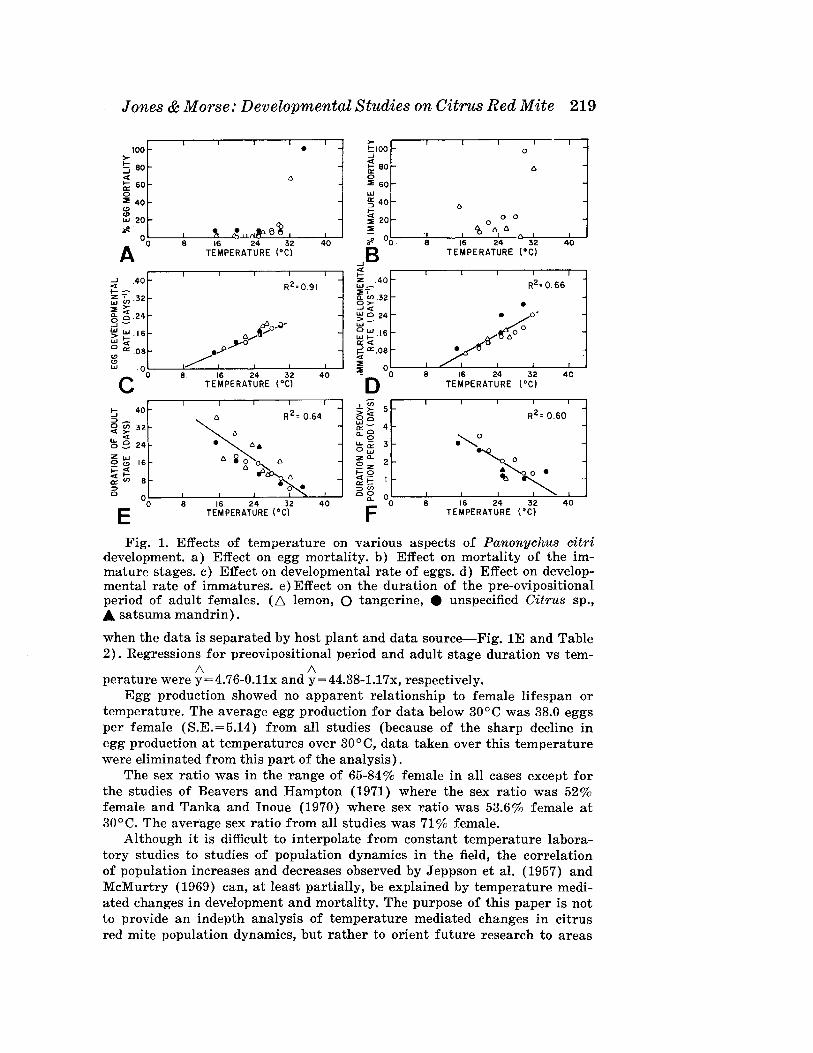
High mortali,ty was found to occur in both the egg and immature stagesat a constant temperature of ca. 30°C (23.7 and 20.6 degree days per day,respectively) (Fig. lA,B). Based primarily on the work of Keetch (1971),Munger (1963), and observations in greenhouse experiments where temperatures peaked in excess of 30°C for short periods without population decrease (Jones 1983), we feel that a sharp increase in mortality occurs onlyafter 1-3 days constant exposure to temperatures over 30°C.
The 5 studies used for the egg development regression were in very goodagreement (R2 of 0.91, Fig. lC). The developmental threshold was found to
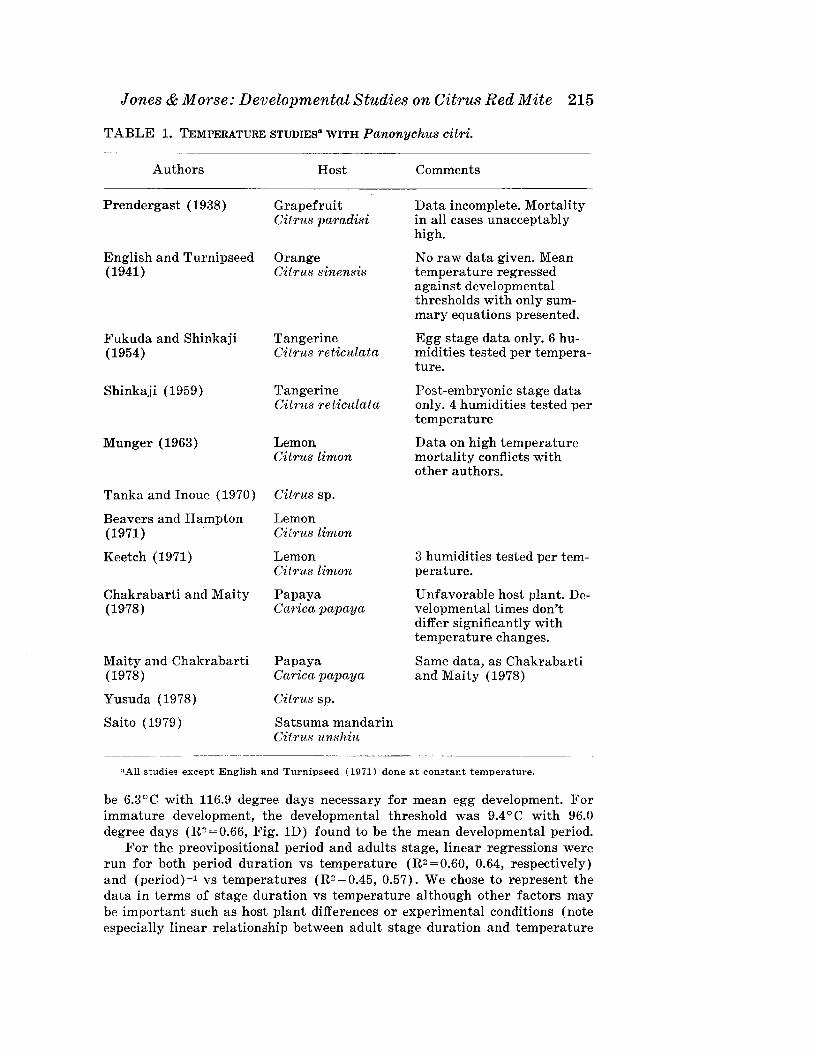
Jones & Morse: Developmental Studies on Citrus Red Mite 215
TABLE 1. TEMPERATURE STUDIES" WITH Panonychus citri.
Authors Host Comments
Prendergast (1938) Grapefruit Data incomplete. MortalityCitrus paradisi in all cases unacceptably
high.
English and Turnipseed Orange No raw data given. Mean(1941) Citrus sinensis temperature regressed
against developmentalthresholds with only sum-mary equations presented.
Fukuda and Shinkaji Tangerine Egg stage data only. 6 hu-(1954) Citrus reticulata midities tested per tempera-
ture.
Shinkaji (1959) Tangerine Post-embryonic stage dataCitrus reticulata only. 4 humidities tested per
temperature
Munger (1963) Lemon Data on high temperatureCitrus limon mortality conflicts with
other authors.
Tanka and Inoue (1970) Citrus sp.
Beavers and Hampton Lemon(1971) Citrus limon
Keetch (1971) Lemon 3 humidities tested per tem-Citrus limon perature.
Chakrabarti and Maity Papaya Unfavorable host plant. De-(1978) Carica papaya velopmental times don't
differ significantly withtemperature changes.
Maity and Chakrabarti Papaya Same data, as Chakrabarti(1978) Carica papaya and Maity (1978)
Yusuda (1978) Citrus sp.
Saito (1979) Satsuma mandarinCitrus unshiu
nAIl studies except English and Turnipseed (1971) done at constant temperature.
be 6.3°C with 116.9 degree days necessary for mean egg development. Forimmature development, the developmental threshold was 9.4°C with 96.0degree days (R2=0.66, Fig. 1D) found to be the mean developmental period.
For the preovipositional period and adults stage, linear regressions wererun for both period duration vs temperature (R2=0.60, 0.64, respectively)and (period)-l vs temperatures (R2=0.45, 0.57). We chose to represent thedata in terms of stage duration vs temperature although other factors maybe important such as host plant differences or experimental conditions (noteespecially linear relationship between adult stage duration and temperature
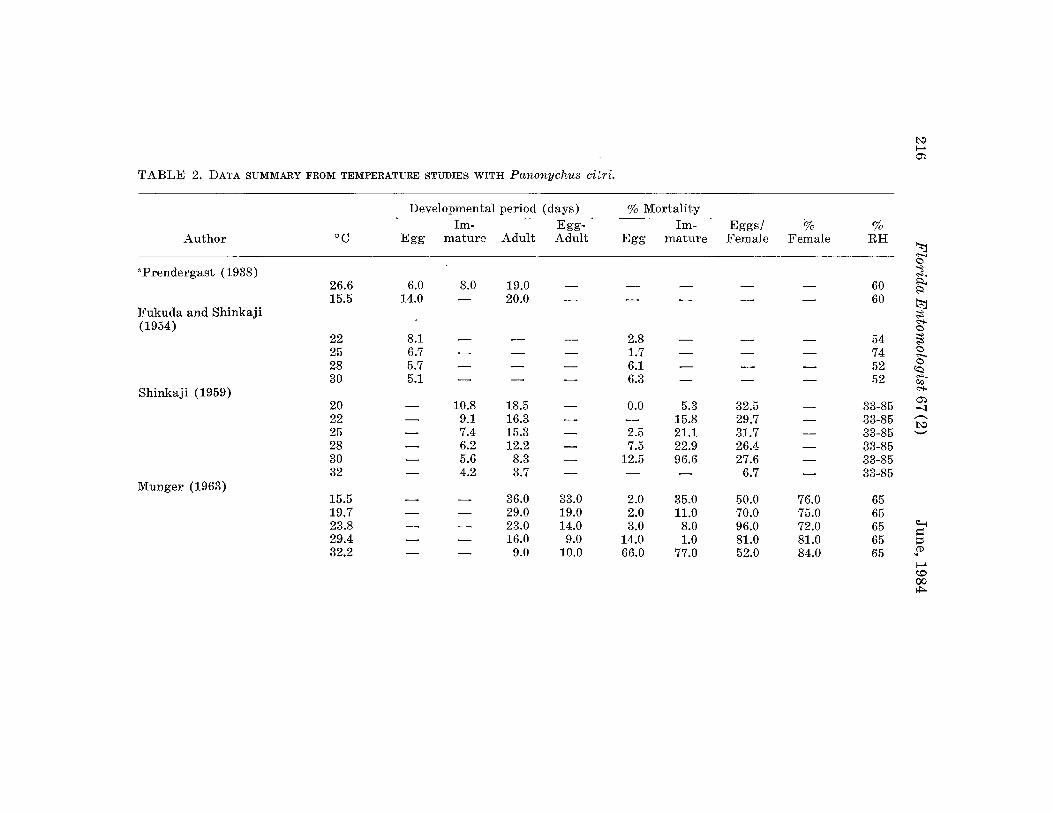
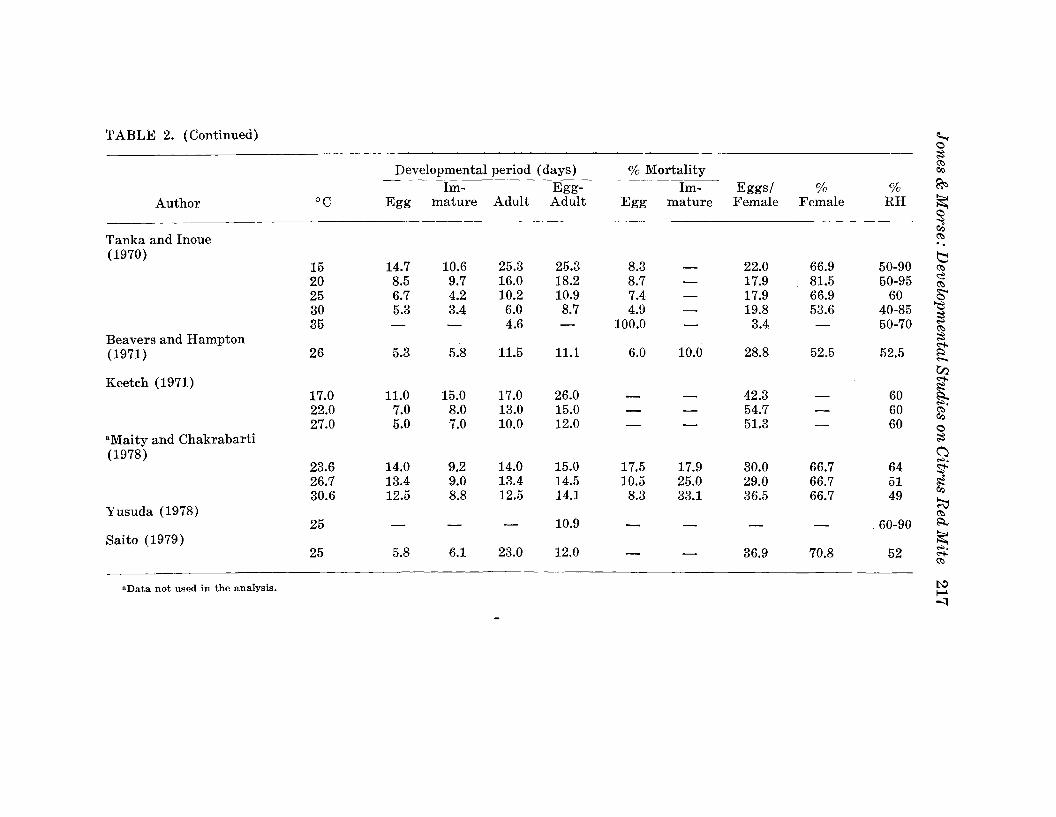
TABLE: 2. (Continued) ~<:>;;Z
Developmental period (days) % Mortalitya:.i:I;)
Im- Egg- Im- Eggs/ % % R'=>
Author °C Egg mature Adult Adult Egg mature Female Female RH ~<:>~
Tanka and Inoue a:.'.
(1970) tl15 14.7 10.6 25.3 25.3 8.3 - 22.0 66.9 50-90 a:.20 8.5 9.7 16.0 18.2 8.7 17.9 81.5 50-95 ~- a:.25 6.7 4.2 10.2 10.9 7.4 17.9 66.9 60 ~- <:>30 5.3 3.4 6.0 8.7 4.9 - 19.8 53.6 40-85 ~
35 - - 4.6 - 100.0 - 3.4 - 50-70 ~a:.
Beavers and Hampton ;;Z~
(1971) 26 5.3 5.8 11.5 11.1 6.0 10.0 28.8 52.5 52.5 ~~
~Keetch (1971) ~
~17.0 11.0 15.0 17.0 26.0 - - 42.3 - 60 ~
22.0 7.0 8.0 13.0 15.0 54.7 60..,.
- - - a:.27.0 5.0 7.0 10.0 12.0 51.3 60
i:I;)
- - - <:>aMaityand Chakrabarti ~
(1978) (J..,.23.6 14.0 9.2 14.0 15.0 17.5 17.9 30.0 66.7 64 ~
26.7 13.4 9.0 13.4 14.5 10.5 25.0 29.0 66.7 51 g30.6 12.5 8.8 12.5 14.1 8.3 33.1 36.5 66.7 49
~Yusuda (1978) a:.25 - - - 10.9 - - - - .60-90 ~
Saito (1979) ~25 5.8 6.1 23.0 12.0 36.9 70.8 52
..,.- - ~
a:.
"Data not used in the analysis. t>:.l~
~
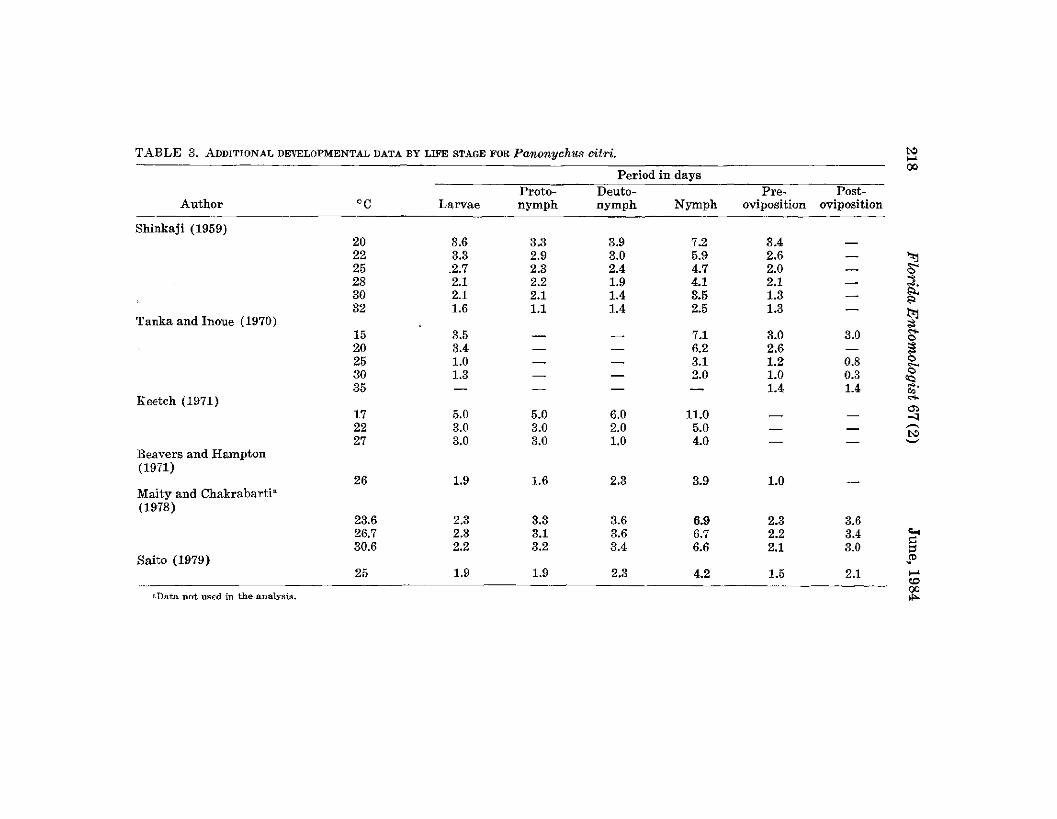
TABLE 3. ADDITIONAL DE''ELOPMENTAL DATA BY LIFE STAGE FOR Panonych'llS citri. t.:>>-'
Period in days00
Proto- Deuto- Pre- Post-Author °C Larvae nymph nymph Nymph oviposition oviposition
Shinkaji (1959)20 3.6 3.3 3.9 7.2 3.422 3.3 2.9 3.0 5.9 2.6 - '""l25 .2.7 2.3 2.4 4.7 2.0 - -028 2.1 2.2 1.9 4.1 2.1 - j.30 2.1 2.1 1.4 3.5 1.3 - ....,32 1.6 1.1 1.4 2.5 1.3 -
~Tanka and Inoue (1970)15 3.5 - - 7.1 3.0 3.0
,....0
20 3.4 - - 6.2 2.6 - ~25 1.0 - - 3.1 1.2 0.8 0-30 1.3 - - 2.0 1.0 0.3 0
'"35 - - - - 1.4 1.4 "'..,Keetch (1971)
,....(j)
17 5.0 5.0 6.0 11.0 - - "'l22 3.0 3.0 2.0 5.0 - - ~
t.:>27 3.0 3.0 1.0 4.0 - - ~
Beavers and Hampton(1971)
26 1.9 1.6 2.3 3.9 1.0Maity and Chakrabarti·(1978)
23.6 2.3 3.3 3.6 6.9 2.3 3.626.7 2.3 3.1 3.6 6.7 2.2 3.4 ....30.6 2.2 3.2 3.4 6.6 2.1 3.0
~::l
Saito (1979) .'"25 1.9 1.9 2.3 4.2 1.5 2.1 >-'
'"aData not used in the analysis.00
"'"
Jones & Morse: Developmental Studies on Citrus Red Mite 219
>-t:100 °~
~ 80 00260wg:; 40 0I-
:i 20 ° °°~ 'b o 0
0!1 00. 8 16 24 32 40
~BTEMPERATURE (OC)
«I-z .40 R2 , 0.66w_27<>. <J) .320>- •~«
~~24 • 0'
w
a--~~~.160:«~ "'.08 ~e'2
0~ 0 8 16 24 32 40
0 TEMPERATURE (OC)
,v;5>~ R2 , 0.600 0w_40: 0<>'0
~o... - 30'" ., °Wz<>.
2S?z "'""'-0 ° •1-0«- I ~~",t::::><J)00 0
<>. 0 8 16 24 32 40
F TEMPERATURE toC)
•
16 24 32 40TEMPERATURE (OCl
8
100>-S 80«~ 60o2 40
'"'"w 20
1ft " • ,,~(\ '8o0~---'8~-~16--"b.J..I.qJ24?=--'~3f:.2---'4~0----'
A TEMPERATURE (OC)
E
!J 40:::>-~ g; 32
'c5 e24
~ ~ 161-«
~ t;; 8:::>o
;i .40I- -~~.322>-g; ~24~ ~ ...~O-Q-~ ~.16 OlJW « ~O/~ '" .08 • .p
~ 0 ~--7'~_---:'~--:'-:----:::':----::-'::--'
CO 8 16 24 32 40
TEMPERATURE (OC)
Fig. 1. Effects of temperature on various aspects of Panonychus citridevelopment. a) Effect on egg mortality. b) Effect on mortality of the immature stages. c) Effect on developmental rate of eggs. d) Effect on developmental rate of immatures. e) Effect on the duration of the pre-ovipositionalperiod of adult females. (6 lemon, 0 tangerine, • unspecified Citrus sp.,A satsuma mandrin).
when the data is separated by host plant and data source-Fig. IE and Table2). Regressions for preovipositional period and adult stage duration vs tem-
/\ /\perature were y=4.76-0.llx and y=44.38-1.17x, respectively.
Egg production showed no apparent relationship to female lifespan ortemperature. The average egg production for data below 30°C was 38.0 eggsper female (S.E.=5.14) from all studies (because of the sharp decline inegg production at temperatures over 30°C, data taken over this temperaturewere eliminated from this part of the analysis).
The sex ratio was in the range of 65-84% female in all cases except forthe studies of Beavers and Hampton (1971) where the sex ratio was 52%female and Tanka and Inoue (1970) where sex ratio was 53.6% female at30° C. The average sex ratio from all studies was 71 % female.
Although it is difficult to interpolate from constant temperature laboratory studies to studies of population dynamics in the field, the correlationof population increases and decreases observed by Jeppson et al. (1957) andMcMurtry (1969) can, at least partially, be explained by temperature mediated changes in development and mortality. The purpose of this paper is notto provide an indepth analysis of temperature mediated changes in citrusred mite population dynamics, but rather to orient future research to areas

220 Florida Entomologist 67 (2) June, 1984
where data necessary for a modeling effort aimed at the citrus red mite isinadequate.
Our analysis has shown that temperature dependent studies from aroundthe world are in surprisingly close agreement and that adequate data areavailable on constant temperature developmental rates of the citrus red mite.Based on these results, future research projects are needed in the followingareas: (1) comparison of constant temperature developmental rates withvariable temperature rates, (2) determination of accurate mortality dataunder variable temperature regime, especially high temperature mortality,.and (3) influence of host plant nutrition on mite development.
ACKNOWLEDGMENTS
The authors would like to thank Mr. Arata Iseki for translating severalof the articles written in Japanese.
REFERENCES CITED
ARNOLD, C. Y. 1959. The determination and signiflcance of the base temperature in a linear heat unit system. American Soc. Hort. Sci. 74:430-45.
BEAVERS, J. B., AND R. B. HAMPTON. 1971. Growth, development, and matingbehavior of the citrus red mite (Acarina: Tetranychidae). Ann. Ent.Soc. America 64: 804-6.
BUXTON, G. M. 1977. Estimated damage and crop loss caused by insect andmite pests. California Dept. of Food and Agric. 12 p.
CHAKRABARTI, S., AND S. P. MAlTY. 1978. Effect of temperature and relativehumidity on the life cycle of Panonychus citri (McGregor) (Acarina:Tetranychidae). Sci. and Culture 44: 233-4.
ENGLISH, L. L., AND G. F. TURNIPSEED. 1941. The influence of temperatureand season on the citrus red mite (Paratetmnychus aitri). J. Agr.Res. 62: 65-77.,
FUKUDA, J., AND N. SHINKAJI. 1954. Experimental studies on the influenceof temperature and rela.tive humidity upon the development of citrusred mite. (Metatetmnychus aitri McGregor). (1) On the influence oftemperature and relative humidity upon the development of the eggs.Bull. Tokai-Kinki Agric. Exp. Sta. (Horticulture). 2: 160-71.
JEPPSON, L. R., C. A. FLESCHNER, M. J. JESSER, AND J. O. COMPLIN. 1957.Influence of season and weather on citrus red mite populations onlemons in southern California. J. Econ. Ent. 50: 293-300.
JEPPSON, L. R., H. H. KEIFER, AND E. W. BAKER. 1975. Mites injurious toeconomic plants. Univ. of California Press, Berkeley. 614 p.
JONES, V. P. 1983. Pest resurgence of, and sampling plans for, the citrusred mite, Panonychus citri (McGregor) (Acari: Tetranychidae).Ph.D. Dissertation. Univ. of Calif., Riverside. 104 p.
KEETCH, D. P, 1971. Ecology of the citrus red mite, Panonychus aitri(McGregor), (Acarina: Tetranychidae) in South Africa. 2. The influence of temperature and relative humidity on the development andlife cycle. J. Ent. Soc. South Africa 34: 103-8.
MAlTY, S. P., AND S. CHAKRABARTI. 1978. Biological studies on Panonychusaitri (Acari: Tetranychidae). Indian J. Acar. 2: 55-9.
McMuRTRY, J. A. 1969. Biological control of citrus red mite in California.Proc. 1st International Citrus Symposium 2: 855-62.
MUNGER, F. 1963. Factors affecting growth and multiplication of the citrusred mite, Panonychus aitri. Ann. Ent. Soc. America 56: 867-74.

Gagne & Beavers: Contarinia from Slash Pine Shoots 221
PRENDERGAST, D. 1938. Studies of the biology of the citrus red mite,Paratetranychus citri (MeG.). Ph.D. Dissertation, Univ. California,Riverside. 65 p.
SAITO, Y. 1979. Comparative studies on life histories of three species ofspider mites (Acarina: Tetranychidae). App. Ent. Zool. 14: 83-94.
SHINKAJI, N. 1959. Experimental studies on the temperature and relativehumidity influencing the development of the citrus red mite (Panonychus [Metatranychus] citri) (McGregor). (2) On the influence oftemperature and relative humidity on the post-embryonic development,adult life span, and egg-laying potency of the citrus red mite. Bull.Hort. Div. Tokai-Kinki Agr. Exp. Sta. 5: 129-42.
TANKA, M., AND K. INOUE. 1970. Fundamental studies on the utilization ofnatural enemies in citrus groves in Japan. II. The method of prediction of outbreaks of the citrus red mite, Panonychus citri (MeG.).Bull. Hort. Res. Sta., Japan, Ser. D. 6: 1-40.
YUSUDA, M. 1978. Reproduction and factors influencing dispersal of thecitrus red mite, Panonychus citri (McGregor) (Acarina: Tetranychidae) under laboratory conditions. Japanese J. Appl. Ent. Zool. 22:12-7.
• • • • • • • • • • • • •CONTARINIA SPP. (DIPTERA: CECIDOMYIIDAE)
FROM SHOOTS OF SLASH PINE (PINUS ELLIOTTIIENGELM.) WITH THE DESCRIPTION OF A
NEW SPECIES INJURIOUS TO NEEDLES
RAYMOND J. GAGNESystematic Entomology Laboratory, IIBIII, Agricultural Research Service
USDA, c/o U.S. National Museum of Natural HistoryWashington, DC 20560
ANDGREGORY M. BEAVERS3103 McCarty Hall
University of FloridaGainesville, FL 32611
ABSTRACT
A new species, Contarinia acuta Gagne (Diptera: Cecidomyiidae), whichdamages developing needles of slash pine, Pinus elliottii Engelm., in Florida,is described. It is distinguished from other North American species ofContarinia infesting pine, and diagnostic illustrations are provided for thelarvae of the new species and those of 3 other, unnamed species of Contariniaassociated with needles of slash pine.
RESUMEN
Una nueva especie, Contarinia acuta Gagne (Diptera: Cecidomyiidae),que dana las hojas en desarrollo del pino Pinus elliottii Engelm. en la Florida, es descrita. Es distinguida de otras especies Norteamericanas de Contarinia que infestan pinos, e ilustraciones diagn6sticas son proveidas paralarvas de la nueva especie y de otras 3 especies sin nombrar de Contariniaasociadas con las hojas del pino P. elliottii.

Gagne & Beavers: Contarinia from Slash Pine Shoots 221
PRENDERGAST, D. 1938. Studies of the biology of the citrus red mite,Paratetranychus citri (MeG.). Ph.D. Dissertation, Univ. California,Riverside. 65 p.
SAITO, Y. 1979. Comparative studies on life histories of three species ofspider mites (Acarina: Tetranychidae). App. Ent. Zool. 14: 83-94.
SHINKAJI, N. 1959. Experimental studies on the temperature and relativehumidity influencing the development of the citrus red mite (Panonychus [Metatranychus] citri) (McGregor). (2) On the influence oftemperature and relative humidity on the post-embryonic development,adult life span, and egg-laying potency of the citrus red mite. Bull.Hort. Div. Tokai-Kinki Agr. Exp. Sta. 5: 129-42.
TANKA, M., AND K. INOUE. 1970. Fundamental studies on the utilization ofnatural enemies in citrus groves in Japan. II. The method of prediction of outbreaks of the citrus red mite, Panonychus citri (MeG.).Bull. Hort. Res. Sta., Japan, Ser. D. 6: 1-40.
YUSUDA, M. 1978. Reproduction and factors influencing dispersal of thecitrus red mite, Panonychus citri (McGregor) (Acarina: Tetranychidae) under laboratory conditions. Japanese J. Appl. Ent. Zool. 22:12-7.
• • • • • • • • • • • • •CONTARINIA SPP. (DIPTERA: CECIDOMYIIDAE)
FROM SHOOTS OF SLASH PINE (PINUS ELLIOTTIIENGELM.) WITH THE DESCRIPTION OF A
NEW SPECIES INJURIOUS TO NEEDLES
RAYMOND J. GAGNESystematic Entomology Laboratory, IIBIII, Agricultural Research Service
USDA, c/o U.S. National Museum of Natural HistoryWashington, DC 20560
ANDGREGORY M. BEAVERS3103 McCarty Hall
University of FloridaGainesville, FL 32611
ABSTRACT
A new species, Contarinia acuta Gagne (Diptera: Cecidomyiidae), whichdamages developing needles of slash pine, Pinus elliottii Engelm., in Florida,is described. It is distinguished from other North American species ofContarinia infesting pine, and diagnostic illustrations are provided for thelarvae of the new species and those of 3 other, unnamed species of Contariniaassociated with needles of slash pine.
RESUMEN
Una nueva especie, Contarinia acuta Gagne (Diptera: Cecidomyiidae),que dana las hojas en desarrollo del pino Pinus elliottii Engelm. en la Florida, es descrita. Es distinguida de otras especies Norteamericanas de Contarinia que infestan pinos, e ilustraciones diagn6sticas son proveidas paralarvas de la nueva especie y de otras 3 especies sin nombrar de Contariniaasociadas con las hojas del pino P. elliottii.

222 Florida Entomologist 67 (2) June, 1984
During research into the cause of needle damage on slash pine, Pinuselliottii Engelm., 4 distinct species of Contarinia were found associated withnew shoots. A characteristic needle damage is caused by Contarinia acutaGagne n. sp. described in this paper. Larvae of the other 3 species are lesscommon than C. acuta and their particular role on the needles is still unknown. They were not reared to the adult stage, which is advisable beforethey can be named, but they are briefly described with illustrations so thatthey can be distinguished in future studies.
Characteristic brown lesions (ca. 4 mm long) are formed on needles as aresult of the feeding of C. acuta larvae. Feeding occurs inside the fasciclesheath of basally elongating needles (Fig. 1). After needle elongation hasbeen completed, the needles become dry and brittle at the lesion site andfrequently break (Fig. 2). Mature larvae drop to the ground and pupate inthe surface litter. In northern Florida, larvae have been collected as early asmid-February, with 6 generations having been recorded in 1 year and up to4 between May and September (Beavers, in preparation).
Contarinia acuta Gagne, NEW SPECIESFig. 3-12
ADULT. Head. Eyes large, about 5 facets long at vertex, the facetscircular, closely approximated except at midheight of eye where they lieslightly farther apart. Occiput rounded, without peak. Frontoclypeal setae8-10. Labella hemispherical in frontal view with 4-5 setae. Palpus 4-segmented. Male antennal flagellomere 3 (Fig. 3) binodal, bicircumfilar, thecircumfilar loops not attaining the next distal node. Female antennal flagellomeres 1-3 as in Fig. 4; circumfilar loops slightly bowed (Fig. 5).
Thorax. Mesoscutal row with sparse setae in mostly single row interspersed with scales. Mesanepisternum bare. Mesepimeron with 5 setae. Winglength male, 158-164 mm (162, avg. of 5), female, 173-200 mm (190 avg. of5); RS slightly bowed apically, joining C behind wing apex. C broken, thewing margin indented at juncture with R5. Claws barely shorter thanempodia.
Male abdomen (Fig. 8). Tergites 1-7 rectangular with basal pair oftrichoid sensilla, a caudal, single row of sparse setae, 2-4 lateral setae, ando scales; tergite 8 weakly sclerotized caudally but with at least 1-2 caudalsetae laterally, 0 lateral setae, 2 basal trichoid sensilla. Sternites 2-6 rectangular with pair of basal, closely approximated trichoid sensilla, a mostlysingle, caudal row of setae, and mixed setae and setiform scales groupednear midlength; sternite 7 as for preceding except caudal row mostlydouble; sternite 8 with midlength and caudal groups of setae continuous oncaudal half of sclerite, and basal trichoid sensilla not as closely approximatedas on sternites 2-7. Genitalia (Fig. 9 (dorsal) -10 (ventral): cerci short,broadly rounded with 2 caudoventral setae; hypoproct not deeply divided,the lobes broad, with 2 long setae. Aedeagus longer than hypoproct, verynarrow, attenuate, and pointed at apex; gonocoxal apodeme broad; gonopodstout; gonostylus broadest near midlength, tapering to apex, setulosethroughout.
Female abdomen (Fig. 6). Tergites 1-7 and sternites 2-7 as in male butsetae more numerous except for fewer laterals on tergites. Tergite 7 about.14 length distal section of ovipositor. Tergite 8 longer than 7 with pair of

Gagne & Beavers: Contarinia from Slash Pine Shoots 223
Fig. 1-2. 1) Larva of C. acuta feeding on adaxial surface of needle removed from fascicle. 2) Characteristic damage of C. acuta.
224 Florida Entomologist 67 (2)
4 5
June, 1984
6
Fig. 3-7. Contarinia acuta: 3, 3rd () antennal flagellomere; 4, 1st through3rd ~ flagellomeres; 5, 3rd ~ flagellomere; 6, ~ postabdomen; 7, ~ cerci(dorsolateral) .
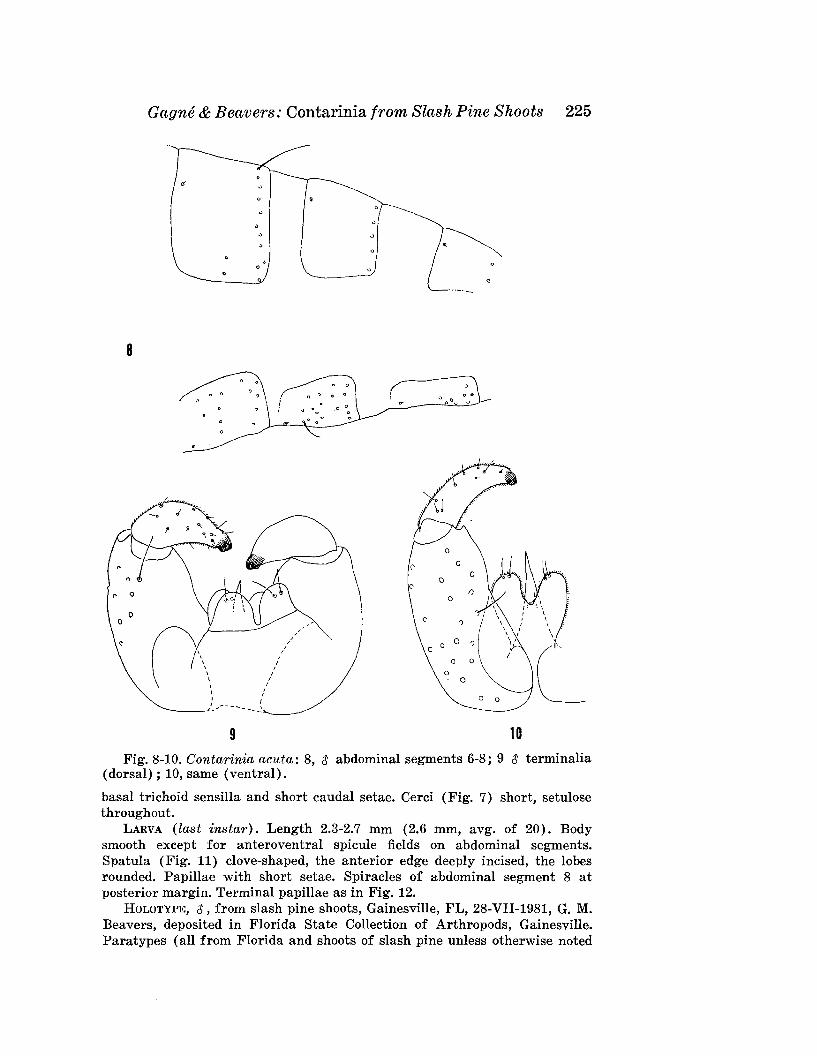
Gagne & Beavers: Contarinia from Slash Pine Shoots 225
o
8
" 0
oo
//,,
;I
II
II
II
I
-_\_-~
9
() "} 0 0
o 0.. 0
o •o 0
o v
COO ?
o 0
oo
10Fig. 8-10. Contarinia acuta: 8, (; abdominal segments 6-8; 9 (; terminalia
(dorsal); 10, same (ventral).
basal trichoid sensilla and short caudal setae. Cerci (Fig. 7) short, setulosethroughout.
LARVA (last instar). Length 2.3-2.7 mm (2.6 mm, avg. of 20). Bodysmooth except for anteroventral spicule fields on abdominal segments.Spatula (Fig. 11) clove-shaped, the anterior edge deeply incised, the lobesrounded. Papillae with short setae. Spiracles of abdominal segment 8 atposterior margin. Terminal papillae as in Fig. 12.
HOLOTYPE, (;, from slash pine shoots, Gainesville, FL, 28-VII-1981, G. M.Beavers, deposited in Florida State Collection of Arthropods, Gainesville.Paratypes (all from Florida and shoots of slash pine unless otherwise noted

226 Florida Entomologist 67 (2) June, 1984
and deposited in Florida State Collection and U.S. National Museum ofNatural History; GMB - G. M. Beavers; JLF ~ J. L. Foltz; RCW ~ R. C.Wilkinson): 6 ~, 1 ~, 3 pupal exuvia, Gainesville, 13-VIII to 2-XI-81, GMB;3 ~ from pitch canker diseased terminals, 30-VII-1974, Flagler Co., RCW;3 ~, emerged from soil under slash pine, Newberry, 17-VIII-1979, RCW &JLF; 5 pupae, Alachua Co., 16-VII-1982, GMB; 9 larvae, Trenton, 30 to31-VII-1979, RCW & JLF; 3 larvae, pitfall traps, Newberry, 5-IX-1979,RCW & JLF; 2 larvae, Newberry, 30-VII-1979, RCW & JLF; 3 larvae,Gainesville, 26-VII-1979, RCW & JLF; 6 larvae, Gainesville, 7-VII-1981,GMB; 7 larvae, pitch canker diseased terminals, Flagler Co., 27-VIII-1975,RCW; 10 larvae, Flagler Co., 30-VII-1974; and 12 larvae from slash pineneedles, Alachua Co., 28-VII-1981, GMB.
Contarinia acuta is so-named for its extremely fine-pointed aedeagus.Adults will readily key to Contarinia in Gagne (1981), and it is the onlyspecies in that genus with such an attenuate aedeagus. It is otherwise distinguished from all other North American Contarinia on pine by the almostequally long claws and empodia (all other cecidomyiids associated with pinecharacteristically have much longer empodia than claws), the wide malehypoproct, and the completely setulose female cerci. Contarinia baeri (Prell)is the only other Contarinia known on pine in eastern North America, butthat species has long empodia relative to the claws, differently-shaped maleand female terminalia (unpub.), and its larva has each of the corniformterminal papillae at the end of a long caudal process (Skuhravy 1973).
The larva of C. aeuta can be separated from those of the other 3 speciesof Contarinia associated with it on slash pine needles with the help of Fig.11-8. In other Contarinia such differences urc diagnostic and will presumably be so here also. Contarinia sp. A. (Fig. 13-4) is distinguished by afield of short spines almost surrounding the terminal corniform papillae. Inaddition, its spatula is least incised anteriorly of all 4 species. Contarinia sp.E. (Fig. 15-6) is rather similar to C. acuta except that the spatula is not asdeeply incised, resulting in slightly differently shaped anterior lobes. Contarinia sp. C. (Fig. 17-8) ha13 almost connate terminal corniform papillaeand its spatula is the largest of the 4 species. Larvae of Thecodiplosis alsohave somewhat approximated corniform terminal papillae also, but withoutassociated adults we cannot definitely say to which of the 2 genera species Cbelongs. For the present the larvae of species C are assigned to Contariniasensu lato.
Specimens of the 3 unnamed species of Contarinia from slash pine arerecorded as follows:
Contarinia sp. A.: 2 larvae, slash pine needle, Newberry, FL, 5-X-1981,GMB; 3 larvae, pitfall trap, Alachua Co., FL, 15-VII-1981, GMB; 1 larva,slash pine needle, Alachua Co., FL, 1-IX-1982, GMB.
Contarinia sp. E.: 2 larvae, pitfall traps, Newberry, FL, 25-1-1980,R.C.W. & J.L.F.; 2 larvae, slash pine needles, DeLand, FL, 7-X-1982, GMB.
Conturinia sp. C.: 2 larvae, pitfall traps, Newberry, FL, 25-1-1980,R.C.W. & J.L.F.; 1 larva, slash pine needle, Alachua Co., FL, 17-1X-1982,GMB; 4 larvae, Perry, FL, 2-III-1977, R.C.W.
We are grateful to Drs. John L. Foltz and Robert C. Wilkinson, Department of Entomology and Nematology, University of Florida, for initiatingthis study, providing research facilities, and submitting specimens foridentification.
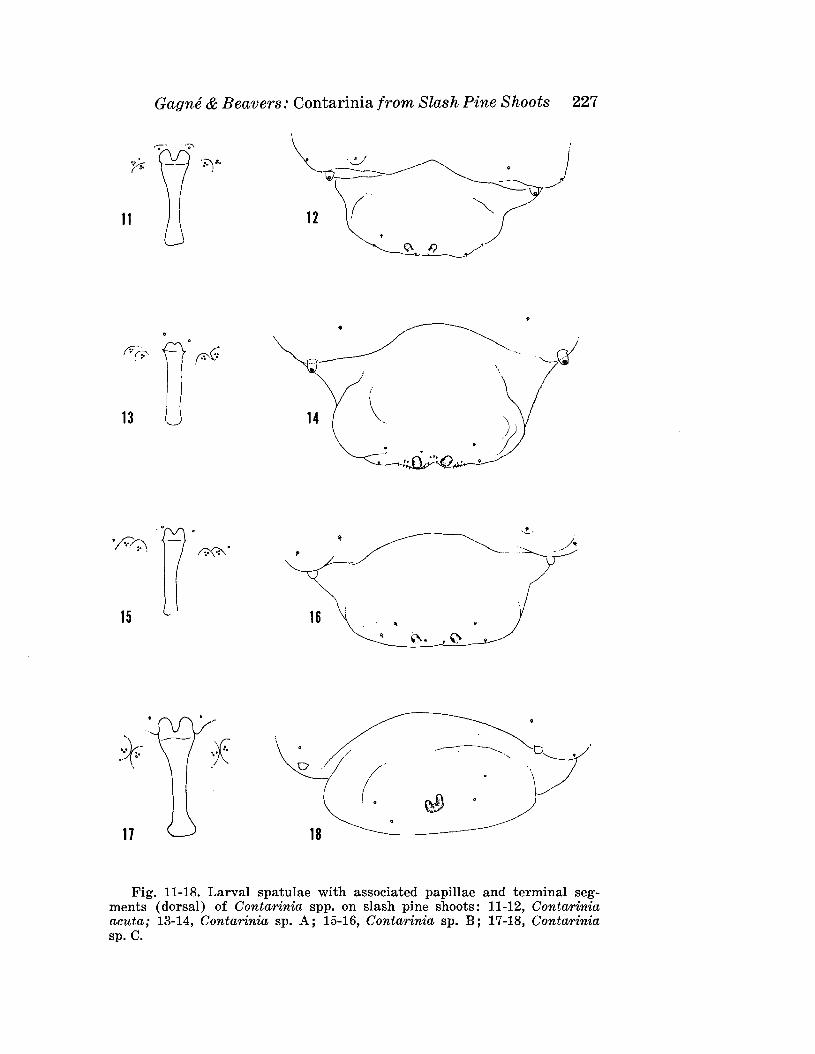
Gagne & Beavers: Contarinia from Slash Pine Shoots 227
11
.::] :.
•\
13
_r..'( ...~~.
~.
.n..''~":~
.. .
15
;:x:?\ •
16
~- )( ~.. , :. ',' :. .--------....... D., ,.
(! . ~
17 18
Fig. 11-18. Larval spatulae with associated papillae and terminal segments (dorsal) of Contarinia spp. on slash pine shoots: 11-12, Contariniaacuta; 13-14, Contarinia sp. A; 15-16, Contarinia sp. B; 17-18, Contariniasp. C.

228 Florida Entomologist 67 (2) June, 1984
REFERENCES CITED
BEAVERS, G. M. In preparation. Bionomics of four species of Contarinia(Diptera: Cecidomyiidae) on slash pine needles. Univ. Florida,Gainesville, M.S. Thesis.
GAGNE, R. J. 1981. Cecidomyiidae. Pages 257-92 In McAlpine, J. F. et al.Manual of Nearctic Diptera. Vol. 1. Research Branch AgricultureCanada Monograph No. 27. vi & 674 p.
SKUHRAVY, V. 1973. "Needle blight" and "needle droop" on Pinus sylvestrisL. in Europe and P. resinosa Ait. in North America (Diptera:Cecidomyiidae). Z. Angew. Ent. 72: 421-8.
• • • • • • • • •• • • •EVOLUTION OF A FIREFLY FLASH CODE
JAMES E. LLOYDDepartment of Entomology and Nematology
University of FloridaGainesville, FL 32611 USA
ABSTRACT
The flash code of Photinus ignitus (male flashes once, female delayslong and then flashes) appears to have evolved from a code like that ofPhotinus macdermotti (male flashes twice, female delays briefly and flashes)by the omission of the second flash of the male pattern and the "connection"of the historic between-flash interval of the male pattern and the femaleshort delay, thus producing the female long delay. Signaling variations andsituations necessary for, and that might have led to, such a change in thehypothetical ancestor occur in today's macdermotti.
RESUMEN
El c6digo de luces de Photjnus ignius (macho: 1 luz; hembra: pausalarga, luz) aparentamente ha evolucionado de un c6digo parecido a el dePhotinus macdermotti (macho: 2 luces; hembra: pausa corta, luz). Haomitido la segunda luz del patron del macho y ha unido el intervalo historicoentre luces del patron del macho con la pausa corta de la hembra, asi produciendo la pausa larga de la hembra. Variaciones de senales y situacionesque permitiria y podria producir tal cambio en el antecesor hipotetico seobserva en macdermotti en poblaciones actuales.
Firefly flashing behavior does not fossilize, but through a comparison ofrelated living species we can see the sorts of changes that occur in evolution,and often get some notion of how and why these changes came about. Sucha comparative approach may suggest specific observations and experimentsthat should be made, and focus attention on overlooked but significant detailsof the behavior of living species. I have made observations on and experimented with the flashing behavior of several species in the Photinus consanguineus group (Green 1956, Lloyd 1969) and on species with which theyhave pertinent interactions. I can suggest a reasonable scheme for theevolution of the code of Photinus ignitus from a precursor like the code of


228 Florida Entomologist 67 (2) June, 1984
REFERENCES CITED
BEAVERS, G. M. In preparation. Bionomics of four species of Contarinia(Diptera: Cecidomyiidae) on slash pine needles. Univ. Florida,Gainesville, M.S. Thesis.
GAGNE, R. J. 1981. Cecidomyiidae. Pages 257-92 In McAlpine, J. F. et al.Manual of Nearctic Diptera. Vol. 1. Research Branch AgricultureCanada Monograph No. 27. vi & 674 p.
SKUHRAVY, V. 1973. "Needle blight" and "needle droop" on Pinus sylvestrisL. in Europe and P. resinosa Ait. in North America (Diptera:Cecidomyiidae). Z. Angew. Ent. 72: 421-8.
• • • • • • • • •• • • •EVOLUTION OF A FIREFLY FLASH CODE
JAMES E. LLOYDDepartment of Entomology and Nematology
University of FloridaGainesville, FL 32611 USA
ABSTRACT
The flash code of Photinus ignitus (male flashes once, female delayslong and then flashes) appears to have evolved from a code like that ofPhotinus macdermotti (male flashes twice, female delays briefly and flashes)by the omission of the second flash of the male pattern and the "connection"of the historic between-flash interval of the male pattern and the femaleshort delay, thus producing the female long delay. Signaling variations andsituations necessary for, and that might have led to, such a change in thehypothetical ancestor occur in today's macdermotti.
RESUMEN
El c6digo de luces de Photjnus ignius (macho: 1 luz; hembra: pausalarga, luz) aparentamente ha evolucionado de un c6digo parecido a el dePhotinus macdermotti (macho: 2 luces; hembra: pausa corta, luz). Haomitido la segunda luz del patron del macho y ha unido el intervalo historicoentre luces del patron del macho con la pausa corta de la hembra, asi produciendo la pausa larga de la hembra. Variaciones de senales y situacionesque permitiria y podria producir tal cambio en el antecesor hipotetico seobserva en macdermotti en poblaciones actuales.
Firefly flashing behavior does not fossilize, but through a comparison ofrelated living species we can see the sorts of changes that occur in evolution,and often get some notion of how and why these changes came about. Sucha comparative approach may suggest specific observations and experimentsthat should be made, and focus attention on overlooked but significant detailsof the behavior of living species. I have made observations on and experimented with the flashing behavior of several species in the Photinus consanguineus group (Green 1956, Lloyd 1969) and on species with which theyhave pertinent interactions. I can suggest a reasonable scheme for theevolution of the code of Photinus ignitus from a precursor like the code of

Lloyd: Firefly Flash Code 229
Photinus macdermotti-ignitus males emit a single flash and their femalesdelay several seconds before flashing their answer; macdermotti males emit2 flashes and their females delay about 1 sec after the second flash beforeresponding (Fig. la, b; Lloyd 1966, 1969).
There are at least 9 Nearctic species in the consanguineus group ofPhotinus and probably more remain to be discovered (Lloyd 1969, p. 38; andunpublished). The flash patterns of mate-seeking males of 6 of the speciesusually or always, depending upon the species, are composed of 2-5 flashes,and those of 2 species are single flashes (Fig. la-g). Another is diurnal andhas given up flash communication for pheromones (Lloyd 1973a, 1977). Themost reasonable and conservative assumption as to the signal of the mostrecent common ancestor of the group is that its males had a multi-flashpattern. This is because most of the known living species in the group, theonly available sample to draw inferences from, have such patterns, and thereis no compelling evidence to the contrary. The 1-flash patterns of ignitusand aquilonius are thus derivative, as is the loss of adult luminosity inindictus. The apparent closest relatives and virtual sibling species (seeLloyd 1969) of ignitus, the (l-flash) firefly whose code evolution I seek toexplain, are: consanguineus, greeni, macdermotti, and an unnamed species,"Barber's photinus." In the codes, of these species, male flash patterns arecomposed of 2 flashes, and female flash answers are delayed about 1 s afterthe second (P2) male flash. The intervals between the 2 flashes in the malepatterns vary among the 4 species (Fig. 1b-e), and females will not answerpaired flashes of inappropriate timing (Lloyd 1966, 1969, unpublished; Buckand Buck 1972) .
If the P2 (second) flash of macdermotti's pattern is omitted the resulting(male-female) code is nearly that of ignitus (cf. Fig. 1a and 1b,h). Butwhy would the females of some ancestral "macdermotti" population, whosemales emitted 2-flash patterns, have become responsive to 1-flash patterns?Would there not have been strong selection pressure in the contexts of reproductive isolation and sexual selection to avoid answering incorrect orinadequate patterns? If females accepted 1-flash patterns could they be expected to delay 1 s beyond where the missing P2 flash would have been?; ormight they delay 1 s after P1? Why should males of this ancestor have begun to emit 1-flash rather than 2-flash patterns? Field observations and labexperiments provide some tentative answers.
METHODS AND MATERIALS
Field observations were made at numerous, diverse localities in easternU.S., mostly in Florida. Field and lab flashing behavior was electronicallyrecorded as described by Lloyd (1973b). Lab experiments used females fromGainesville, FL (Oct.-Nov. 1979), kept on a 16L :8D photoperiod with thedark phase beginning at 2 PM local time, about 30 min before experimentsbegan. Females were held in individual shell vials (8 dram, airtight), corked,with a 5-mm cube of fresh apple to maintain humidity and provide waterand possible nourishment. Artificial flash patterns were produced with anincandescent bulb shining along a 3-cm long, 6.7-mm-diam., covered luciterod whose exposed end had been filed and rounded to produce even illumination. The flasher was aimed at subjects (in their vials) from distances of30-50 cm, and moved a few centimeters from time to time when responses
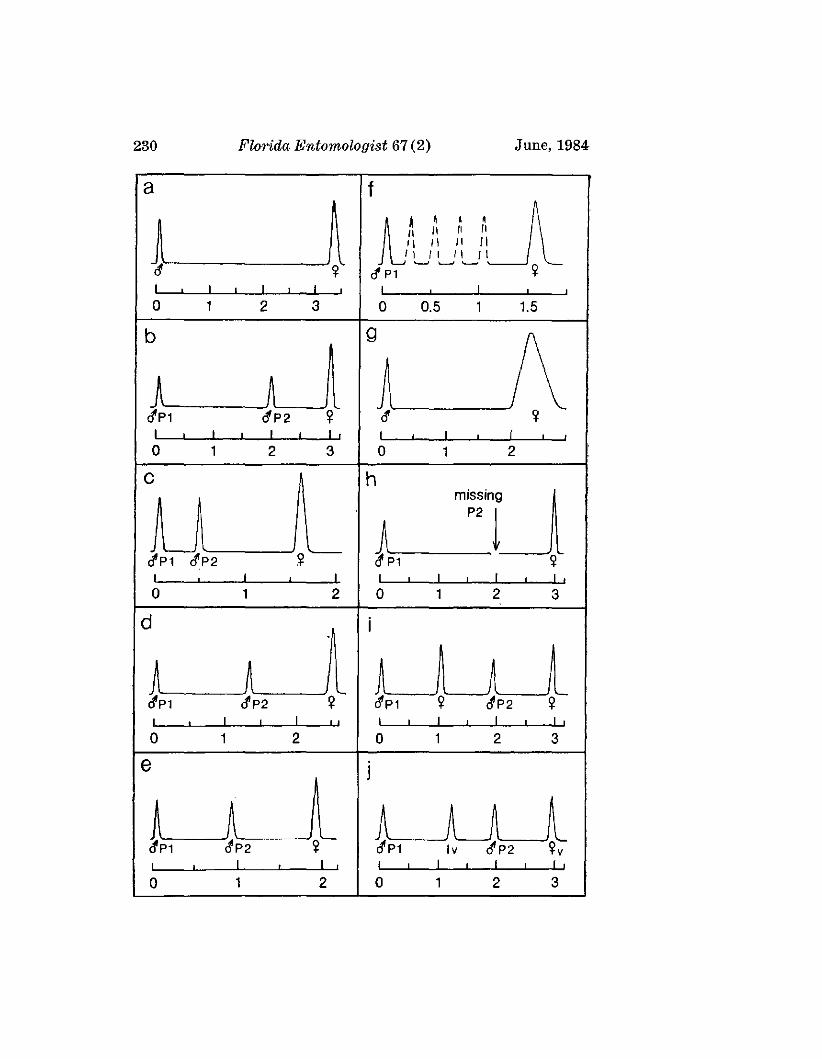
230 Florida Entomologist 67 (2) June, 1984
a f
~ ~ 1LA ,: i ,:.JtII II II IIII 11 II II
d'P1 ~ ~ ~ ~
I I I I I J I I I I I I I
0 1 2 3 0 0.5 1 1.5
b g
1 A ~ Jd'0'P1 d'P2 ~ 'i'I I I I I I I , I I I I ( I ,0 1 2 3 0 1 2
C h
"'. 'i''--- ~m~:lJ
d'P1 d'P2 d' P1 'i'I I I J I I I I I I I I ,
0 1 2 0 1 2 3
d
~i
fi A A ~ A ld'P1 d'P2 ~ O'P1 ~ 0'P2 ~
I , I I I I! I I I I I I I ,
0 1 2 0 1 2 3
e J
~A A&'P1 &'P2 d'P1 Iv d'P2 ~vI I I r I r ! I I I I I I ,
0 1 2 0 1 2 3

Lloyd: Firefly Flash Code 231
waned. Simulations of the 2-flash macdermotti pattern were made uniform(± <1 %) by a stable electronic timing generator (Alton Electronics) whoseflash interval was verified and monitored by the electronic recording systemwith which many of the experiments were recorded. Some experiments weretimed with a split-timer, digital stopwatch (Siliconix ET110). The flashinterval between P1 and P2 was approximately that measured over the pastseveral years in free flying macdermotti males (Fig. 2). Flash duration wasea. 0.1 s. Test females (n=13) were collected as adults by their responses topenlight simulations of their males' pattern and probably included bothvirgins and nonvirgins; eggs in dissected females numbered: 1, 4, 6, 16, 16,47, 70, and 78, suggesting that some had already laid eggs. Wing (1982) hasfound through field study of marked individuals that females may mate atleast 3 times, and will attract males and copulate on consecutive evenings.
Experimental protocol was as follows: individual females were presentedwith series of 5 (designated A-E) flash patterns at 5-10 s intervals; 10 serieswere presented in succession with 1 min of darkness interposed betweenseries. Patterns in each series were either properly-timed (for ambient temp.,Fig. 2) macdermotti patterns, or experimental, single flashes. The positionof the 13, 1-flash (=X) presentations in the various series from 1-10 are asfollows: series 1, no X; 2, X at E; 3, no X; 4, X at Ej 5, X at Ej 6, X at Dand E j 7, X at E j 8, X at D and E; 9, X at C and E; 10, X at C, D, and E.There were 410 series presented, and in addition, a few longer series ofpatterns.
RESULTS
Responsiveness to Experiment Flashes. When presented single flashes atthe end of and within series of 2-flash patterns timed like those of males oftheir species, macdermotti females often answered, and, as anticipated, almost always at a long delay (see below). There was a range of responselevels: one female answered only 6% (3/49) of the 1-flash patterns butanswered 98% (169/173) of the preliminary and interspersed 2-flash (macdermotti) patterns. At the other extreme one female answered 71% of the1-flash patterns (36/51; 2-flash patterns = 95% 139/146), and another89 % (34/38; 2-flash patterns = 100% 113/113). (Totals in the precedingintroductory summary include data from a few modified procedures not included in the remainder of this report.) No overall increased or reduction in
Fig. 1. Flash codes of species in the Photinus consanguineus group (a-g),and other flash interactions. Flash forms drawn from chart traces of electronic recordings and, along with time intervals, are adjusted to about 23° C.Horizontal axis, time in sec; vertical axis, relative intensity. Variations inheight among flashes not significant. Codes of: (a) P. ignitus, (b) P. macdermotti, (c) P. consanguineus, (d) P. greeni, (e) "Barber's Photinus," (f)P. lineellus, indicating variation occurring in a male's pattern, (g) P.aquilonius. (h) An ignitus-like code is produced when a macdermotti femaleanswers a single flash (compare with 'a'). (i) P. macdermotti female mistakenly answered after P1 j note identical delays after P1 and P2. (j) Falseinjection (Iv) and false female response (Q v) of Photuris versicolor femaleaggressive mimic, a predator) j note different delays to P1 and P2 (see text,Lloyd 1981 and in prep).
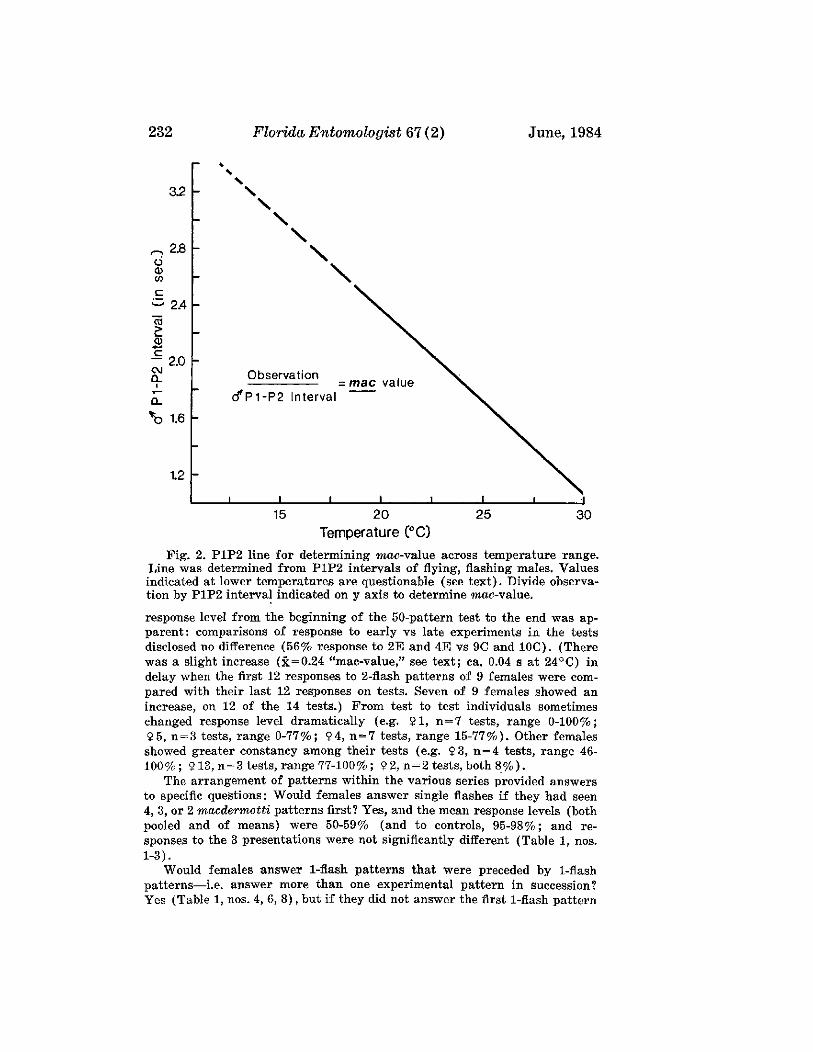
232 Florida Entomologist 67 (2) June, 1984
Observation = mac valued"P1-P2 Interval --
•,,"-
""-"-
""
3.2
~ 2.8<.iQ)(f)
c;::; 2.4c;;
~ 2.0(\J
C-,~
C-
'b 1.6
1.2
15 20Temperature (oG)
25 30
Fig. 2. PIP2 line for determining mac~value across temperature range.Line was determined from PIP2 intervals of flying, flashing males. Valuesindicated at lower temperatures are questionable (see text). Divide observa~
tion by PIP2 interva~ indicated on y axis to determine mac-value.
response level from the beginning of the 50-pattern test to the end waS apparent: comparisons of response to early VB late experiments in the testsdisclosed no difference (56% response to 2E and 4E vs 9C and 10C). (Therewas a slight increase (i=O.24 "mac-value," see text; ca. 0.04 s at 24°C) indelay when the first 12 responses to 2-flash patterns of 9 females were compared with their last 12 responses on tests. Seven of 9 females showed anincrease, on 12 of the 14 tests.) From test to test individuals sometimeschanged response level dramatically (e.g. ~ 1, n~7 tests, range 0-100%;~5, n~3 tests, range 0-77%; ~4, n-7 tests, range 15-77%). Other femalesshowed greater constancy among their tests (e.g. ~ 3, n=4 tests, range 46100%; ~ 13, n~3 tests, range 77-100%; ~ 2, n~2 tests, both 8%).
The arrangement of patterns within the various series provided answersto specific questions: Would females answer single flashes if they had seen4,3, or 2 macdermotti patterns first? Yes, and the mean response levels (bothpooled and of means) were 50-59% (and to controls, 95-98%; and responses to the 3 presentations were not significantly different (Table 1, nos.1-3).
Would females answer l-flash patterns that were preceded by 1-flashpatterns-Leo answer more than one experimental pattern in succession?Yes (Table 1, nos. 4, 6, 8), but if they did not answer the first 1-flash pattern
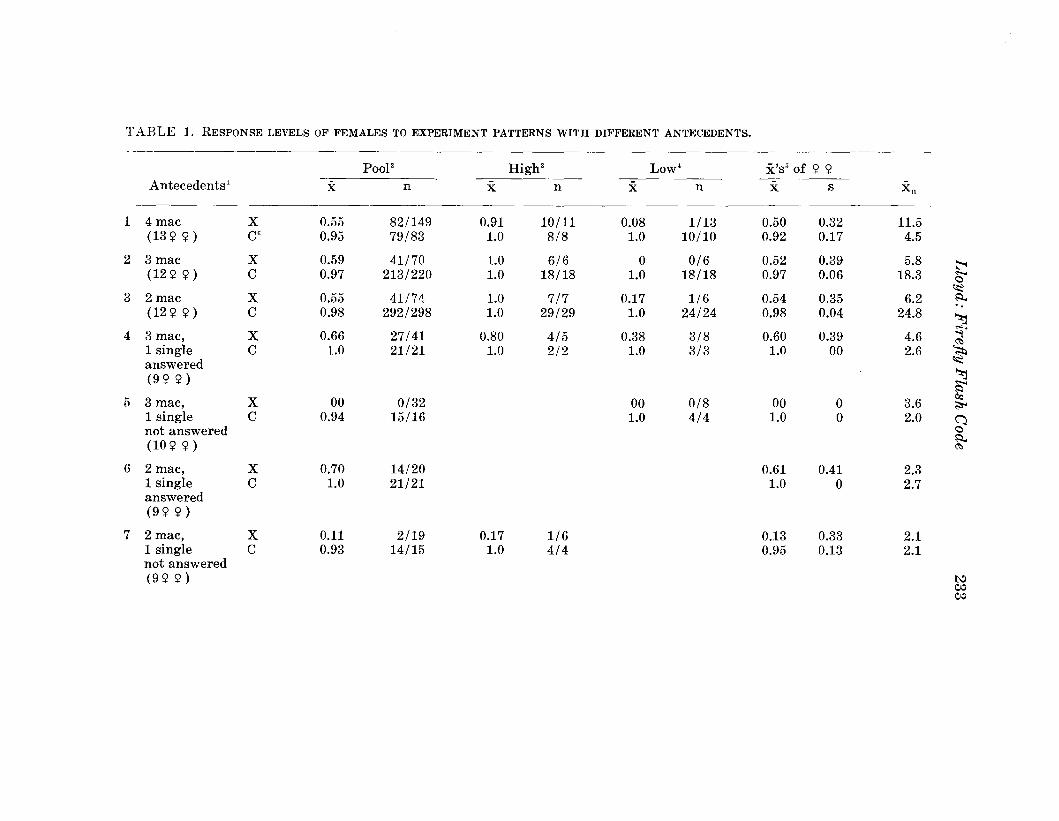
TABLE 1. RESPONSE LEVELS OF FEMALES TO EXPERIMENT PATTERNS WITH DIFFERENT ANTECEDENTS.
Pool2 High3 Low4 X'S5 of c;? c;?
Antecedents! X n X n X n X s Xu
1 4 mac X 0.55 82/149 0.91 10/11 0.08 1/13 0.50 0.32 11.5(13 c;? c;? ) C· 0.95 79/83 1.0 8/8 1.0 10/10 0.92 0.17 4.5
2 3 mac X 0.59 41/70 1.0 6/6 0 0/6 0.52 0.39 5.8 ~(12 c;? c;?) C 0.97 213/220 1.0 18/18 1.0 18/18 0.97 0.06 18.3 .,....
<:>~
3 2 mac X 0.55 41/74 1.0 7/7 0.17 1/6 0.54 0.35 6.2 R..'.(12 c;? c;? ) C 0.98 292/298 1.0 29/29 1.0 24/24 0.98 0.04 24.8~.....
4 3 mac, X 0.66 27/41 0.80 4/5 0.38 3/8 0.60 0.39 4.6 ~(':)
1 single C 1.0 21/21 1.0 2/2 1.0 3/3 1.0 00 2.6 ~
answered ~
(9 c;? c;? ) ~
~5 3 mac, X 00 0/32 00 0/8 00 0 3.6 ;;:r-o1 single C 0.94 15/16 1.0 4/4 1.0 0 2.0 C'Jnot answered <:>
R..(10 c;? c;? ) (':)
6 2 mac, X 0.70 14/20 0.61 0.41 2.31 single C 1.0 21/21 1.0 0 2.7answered(9 c;? c;? )
7 2 mac, X 0.11 2/19 0.17 1/6 0.13 0.33 2.11 single C 0.93 14/15 1.0 4/4 0.95 0.13 2.1not answered(9 c;? c;? ) l\:l
CI:iCI:i
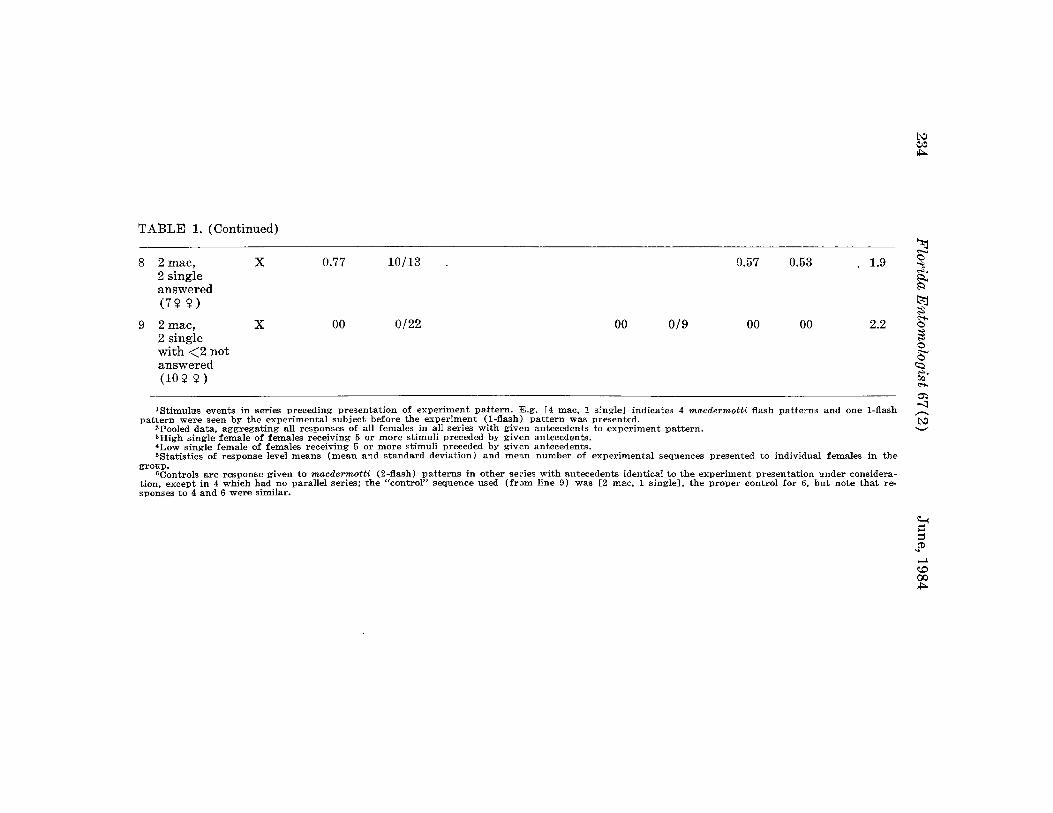
'Stimulus events in series preceding presentation of experiment pattern. E.g. [4 mac, 1 single] indicates 4 macdermotti flash patterns and one I-flashpattern were seen by the experimental subject before the experiment (l-flash) pattern was presenkd.
2Pooled data, aggregating all responses of all females in all series with given antecedents to experiment pattern."High single female of females receiving 5 or more stimuli preceded by given antecedents.'Low single female of females receiving 5 or more stimuli preceded by given antecedents."Statistics of response level means (mean and standard deviation) and mean number of experimental sequences presented to individual females in the
group••Controls are response given to macdermotti (2-flash) patterns in other series with antecedents identical to the experiment presentation under considera
tion, except in 4 which had no parallel series; the "control" sequence used (fr.>m line 9) was [2 mac, 1 single], the proper control for 6, but note that responses to 4 and 6 were similar.
'rABL:8 1. (Continued)
8 2 mac,2 singleanswered(7~ ~)
9 2 mac,2 singlewith <2 notanswered(10 ~ ~ )
x
x
0.77
00
10/13
0/22 00 0/9
0.57
00
0.53
00
1.9
2.2
~~H::-
~"-C""i:.,...R..:;:l
~""'"C
~C"-C(Q~.
""'"cr;,~,-,t-.?'-"
~~I:l~etl
f-4~00H::-

Lloyd: Firefly Flru;h Code 235
they were not likely to answer another one following it (Table 1, nos. 5, 7, 9).There was not a significant difference in response level among 3, compared,antecedent conditions (involving 2 vs 3 macdermotti patterns and 1 vs 2 1flash patterns; Table 1, nos. 4, 6,8). Would females answer more than threeI-flash patterns in succession? Yes; 5 of the 13 females answered 3 or moreconsecutive I-flash patterns. The limit of female responsiveness to patternsin succession was "probed" beyond the measure encorporated in the protocol(series 10, C-E) by sometimes continuing to present I-flash patterns after10E, at 5-10 s intervals. High values for continuous answering of these(extra) single-flash patterns for different females were 15, 14, 6, 5, and 4.The 2 high females with 15 and 14 responses, in other post 10E series answered 13 and 10, respectively. One was given 146 patterns after the 50pattern procedure. Each of the 16 times she stopped responding to I-flashpatterns, she immediately answered a 2-flash pattern and 1-5 subsequentI-flash patterns.
Response levels of females on individual tests and in total, during theirexperimental careers and including as well as excluding zero test scores, wereexamined for correlations with: days since capture, days till death, numberof eggs present at death (""'" at capture), and number of tests given. Only"days until death" showed any significant correlation, and this was improved by excluding zero-score tests and 2 very high scores that occurred oneday before death in 2 individuals (r = 0.63; Fig. 3). Female response toI-flash patterns gradually dropped between 12 and 4 days before death (y =0.28 + .06x), and then appears to drop sharply. If there actually is a sharpbreak in the response curve at ca. 3 days it would explain and incorporateseveral low and zero scores.
Timing of Female Responses. Female macdermotti flashed answers toapproximately 1800 I-flash and 2-flash stimulus patterns during this study.In all but 26 their answers followed the P2 position, and roughly by 1 swhether the P2 flash was present or not; thus, when timed from PI the delayranged around 3 s (.:;;:,d9°C, Fig. 1h). Delays varied predictably with temperature, as do other firefly flash parameters, including the delay of ignitusfemales. In order to combine and compare measurements (of delays, etc.)made at different temperatures, delays were divided by the male P1P2 interval measured at various temperatures in free flying males over the pastseveral years (Fig. 2). The resulting number is referred to as a mac-value(=mv; Lloyd 1981).
A total of 789 delays from 11 females were measured. The mean femaledelay to I-flash patterns (n=120) was about 3% longer than that to 2flash patterns (n = 669; 1.33 vs 1.29 mv), and this bias direction occurred in8 of the 11 females. This difference is not significant if the responses of allfemales (n = 11) are compared (t = 1.50, DF = 20). However, if the responsesof only the 8 are compared, the difference (1.37 vs 1.30 mv; 5% longer) issignificant between 0.01 and 0.001 (t=3.76, DF=14). The mean femaledelays to 2-flash patterns that were presented immediately before I-flashpatterns were like those given to all other 2-flash patterns (before: i = 1.29,8=0.044; all: i=1.29, 8=0.039; t=O.O, DF=20). The delays elicited by 2flash patterns immediately following I-flash (experimental) patterns werelike those after all other 2-flash patterns (after: i = 1.30, 8 = 0.046; all:i=1.29, 8=0.039; t=0.553, DF=20).
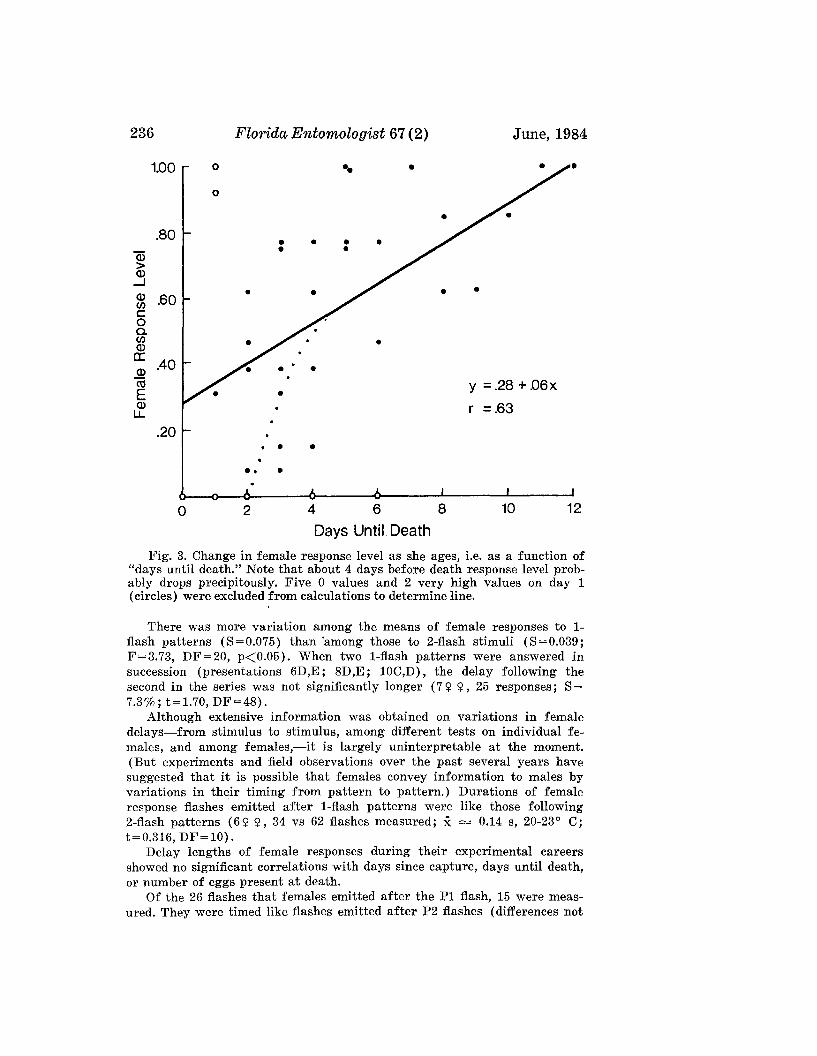
236 Florida Entomologist 67 (2) June, 1984
too
Q)>Q)-IQ)enc:oCoenQ)c:Q)
Ci5EQ)
LL
.80
.60
.40
o
o
•
••
••
•
•
•
..
•• •
•
•
•
•
•
•
y =.28 + .06x
r =.63
.20•• •
•• •
o 2 468
Days Until. Death10 12
Fig. 3. Change in female response level as she ages, i.e. as a function of"days until death." Note that about 4 days before death response level probably drops precipitously. Five °values and 2 very high values on day 1(circles) were excluded from calculations to determine line.
There was more variation among the means of female responses to 1flash patterns (8=0.075) than 'among those to 2-flash stimuli (8=0.039;F=3.73, DF=20, p<0.05). When two 1-flash patterns were answered insuccession (presentations 6D,E; 8D,E; 10C,D), the delay following thesecond in the series was not significantly longer (7 ~ ~, 25 responses; 8 =7.3%; t=1.70, DF=48).
Although extensive information was obtained on variations in femaledelays-from stimulus to stimulus, among different tests on individual females, and among females,-it is largely uninterpretable at the moment.(But experiments and field observations over the past several years havesuggested that it is possible that females convey information to males byvariations in thE;ir timing from pattern to pattern.) Durations of femaleresponse flashes emitted after 1-flash patterns were like those following2-flash patterns (6~ ~, 34 vs 62 flashes measured; X "'=' 0.14 s, 20-23° C;t=O.316, DF=10).
Delay lengths of female responses during their experimental careersshowed no significant correlations with days since capture, days until death,or number of eggs present at death.
Of the 26 flashes that females emitted after the P1 flash, 15 were measured. They were timed like flashes emitted after P2 flashes (differences not

Lloyd: Firefly Flash Code 237
significant, t=O.16, DF=23). Particular attention was given to this timingbecause signal mimicking predators and competing males of this species inject "extra" flashes between P1 and P2 of an approaching male (Lloyd1979, 1981). It was conceivable that female macdermotti flashes occurringafter P1 could have some connection with the injected flashes of males andpredators. This does not seem to be the case (Fig. 1, cf i with j ).
DISCUSSION
The experiments demonstrate that female macdermotti will answer 1flash patterns after they have seen 2-flash patterns characteristic of theirspecies, and that some females, or females sometimes, will answer severalin succession.
These experiments also demonstrate that female macdermotti answersingle flashes at long delays, thus producing a flash interaction similar tothat of ignitus (Fig. 1a,h), the species whose flash code is of evolutionaryinterest here. Since vegetation can sometimes obscure one flash of a male2-flash pattern, it seems reasonable that females would sometimes answerincomplete patterns, thus helping the approach of a male they have alreadyidentified as a member of their species.
But why should females rarely (26/ """,1800) treat a single stimulus flashas P1 and not P2? I believe part of the answer is that firefly females havea timing mechanism that measures flash pattern intervals (repetition cycles).This could explain why competing males of Photinus pyralis that hover andflash together near a responsive female do not flash immediately after afemale response, but instead wait a few seconds and then flash, thus, in nearsynchrony (Maurer 1968, Lloyd 1979). Such a timer could also have somerole in a more complex coding used to detect the predaceous mimicries ofPhoturis females (Lloyd 1983).
The time-measuring mechanisms of macdermotti females of primary importance here are those that measure the interval of the 2 flashes in themales' 2-flash pattern and the delay after P2. To recognize males of herspecies, a macdermotti female measures the P1-P2 interval. If this intervalis appropriate, and P2 is seen during the critical "window," a timer countsdown to trigger the response flash at the appropriate time. P. macdermottifemales that answer a 1-flash pattern at the long delay have, in one simplemodel, coupled the 2 timers. Additional experiments should permit more to besaid about the factors that influence "decision making" at the junction ofthese timers.
Why would males of ignitus' ancestor have begun to emit 1-flash ratherthan 2-flash patterns? (Note that their females were already responsive to1-flash patterns after initial contact and identification.) At first they wouldhave emitted both 1-flash and 2-flash patterns, depending upon circumstances. There are 3 situations in which 1-flash patterns would have beenadaptive: (1) The typical species pattern was altered to avoid attractingother males that would have competed for the female. At first this couldhave been an advantage, but eventually, in evolutionary time, it would havebeen countered by competitor recognition of the "secret code" approach, andfemales should have encouraged the counter (sons would get "genes" forthe alternative mate-finding tactic).
(2) Males might omit P2 upon close approach to a responding light to

238 Florida Entomologist 67 (2) June, 1984
cause a signal-mimicking predator to reveal her identity. The omission ofP2 would probably cause Photuris females to stop answering or to answerincorrectly either because: their neural mechanisms for answering after theP2 flash were not like those of macdermotti females, whose machinery measures intervals between P1 and P2, and Photuris could not time from P1; orbecause they "computed" the single flash as that of another species theypreyed upon and answered in its code. Experiments with Photuris versicolorfemales (Florida), predators of macdermotti, indicate that they answer 1flash patterns with a short «1 sec) delay, not a long one (Lloyd 1981, unpublished) .
(3) Photuris females attack flashing males in the air, using the flashes toaim their attacks, and they sometimes switch to this predatory tactic fromsignal mimicry, when approaching males hesitate (Lloyd and Wing 1983).A 2-flash ignitus ancestor might have omitted P2 on close approach to a female, or even during searching, because it provided a target for predatorsthat approachd P1, during the P1P2 interval, and struck at P2. Severalfactors could have been responsible for finally fixing the 1-flash pattern inignitus, but after viewing aerial attack behavior by various Photuris speciesand noting its efficiency and frequency of occurrence at certain times, andthe effects it seems to have on macdermotti behavior (Wing 1982, Lloyd andWing 1983), I believe it could well have been the critical factor in fixingignitus' 1-flash pattern. Complete acceptance of 1-flash patterns by femaleswould follow the change in males if the risks to males were very great.
EPILOGUE
The high level of response to 1-flash patterns found in some females wasnot anticipated. On numerous occasions over the past several years femalesin the field and lab had been presented with single flashes and none hadflashed in response. Therefore, the experimental protocol used in this studywas not designed to determine whether females would answer 1-flash patterns without prior 2-flash stimulation. This was tested on females takenfrom the same locality as those used above, in the spring (n=5) and fall(n=3) of 1983. They were presented 15 single flashes in 3 groups of 5 (inthe protocol described above), then an increasing number of normal (2flash) patterns was interspersed in subsequent groups of the tests. Responselevel to normal patterns was 0.78, lower than in previous testing in whichnormal patterns were more numerous (cf. 0.93-1.0, Table 1). Only 5 of 661single flashes were answered at a time delay that indicated females judgedthem to be P1's. These responses occurred after normal patterns had beenpresented and not when females had seen only single flashes. Thus, somepriming by the normal (species-typical) male pattern is necessary to elicitlong-delayed responses to single flashes. Of more interest is the fact that 3females answered a total of 50 1-flash patterns at a time delay that indicatedthat they judged them to be P2's. This is being studied.
ACKNOWLEDGMENTS
I thank John Alcock, John Sivinski, Tom Walker, Steve Wing, and ananonymous referee for commenting on the manuscript; Ngo Dong for dissecting the fireflies; Ray Littel for statistical consultation; Laura Reep for

Lloyd: Firefly Flash Code 239
drawing the figures; Jack Schuster for composing the Resumen; and BarbaraHollien for typing the manuscript. This study was supported by my Department and N.S.F. Grant DEB-7921744. Florida Agricultural Experiment Station Journal Series No. 4527.
REFERENCES CITED
BUCK, J. B., AND E. BUCK. 1972. Photic signaling in the firefly Photinusgreeni. BioI. Bull. 142: 195-205.
GREEN, J. W. 1956. Revision of the Nearctic species of Photinus (Lampyridae: Coleoptera). Proc. California Acad. Sci. 28: 561-613.
LLOYD, J. E. 1966. Studies on the flash communication of Photinus fireflies.Univ. Michigan Mus. Zool. Misc. ColI. 130: 1-95.
---. 1969. Flashes, behavior and additional species of Nearctic Photinusfireflies (Coleoptera: Lampyridae). Coleop. Bull. 23: 29-40.
---. 1973a. Chemical communication in fireflies. Environ. Ent. 1: 265-6.---. 1973b. Fireflies of Melanesia; bioluminescense, mating behavior, and
synchronous flashing. Environ. Ent. 2: 991-1008.---.. 1977. Bioluminsecence and communication. Pages 164-83 in T. A.
Sebeok, ed. How animals communicate. Indiana Univ. Press, Bloomington.
---,. 1979. Sexual selection in luminescent beetles. Pages 393-42 in M. andA. Blum, eds. Sexual selection and reproductive competition in insects. Academic Press, New York.
---,. 1981. Firefly mate-rivals mimic their predators and vice versa.Nature 290: 498-500.
---. 1983. Bioluminescence and communication in insects. Annu. Rev.Ent. 28: 131-60.
---, AND S. R. WING. 1983. Nocturnal aerial predation of fireflies bylight-seeking fireflies. Science 222: 634-5.
MAURER, U. M. 1968. Some parameters of photic signaling important tosexual species recognition in the firefly Photinus pyralis. M.S. Thesis,SUNY, Stony Brook, New York.
WING, S. R. 1982. The reproductive ecologies of three species of fireflies.M.S. Thesis, Univ. of Florida, Gainesville.

240 Florida Entomologist 67 (2) June, 1984
REPRODUCTIVE MATURATION AND PREMATINGPERIOD OF THE PAPAYA FRUIT FLY,
TOXOTRYPANA CURVICAUDA(DIPTERA: TEPHRITIDAE)
PETER J. LANDOLT'
Subtropical Horticulture Research Unit, USDA, ARS13601 Old Cutler RoadMiami, FL 33158 USA
ABSTRACT
Female papaya fruit flies, Toxotrypana curvicauda Gerstaecker, obtainedfrom field-infested papayas, Carica papaya L., required 5 to 6 days to develop mature ovaries comparable in size to those of Dvipositing field-collectedfemales. Younger females, with ovaries less than 0.25 em long, were absentfrom papaya groves, sJ.1ggesting that female papaya fruit flies do not searchout host plants until mature. In mating tests females mated when 5 days oldor older, while males mated at any age (one to 6 days after emergence) although mating rates were less with one-day-old males.
RESUMEN
Hembras de Ia mosca de Ia papaya, Toxotrypana curvicauda Gerstaecker,obtenidas de papayas, Carica papay L., infestadas en el campo necesitaron5 6 6 diRs para desarrollar ovarios maduros parecidos en tamafio a aquellosde las hembras adultas ponedoras de huevos coleccionadas en el campo. NoSe encontraron en el campo hembras mas j6venes, con ovarios de menos dede 0.25 cm, de longitud, sugiriendo esto que la hembra de la mosca de lapapaya no husea planta hospedera hasta au madurez. En pruebas de acoplamiento las hembras se acoplaron al alcanzar 5 dias de edad 0 mas, mientrasque los machos se acoplaron a cualquier edad (de uno a 6 dias despues deemerger) aunque los niveles de acoplamiento fueron menores en machos deun dia de edad.
Tephritid fruit flies generally are reproductively immature upon emergence as adults and require a maturation period before mating and oviposition occur. Laboratory-reared Anastrepha suspensu (Loew) begin mating at10 days of age (Mazomenos et al. 1977), coinciding with maximum development of male pheromone glands (Nation 1974) and attraction of femalesto males (Nation 1972). Similar premating periods are 3 to 4 days inCeratitis capitata (Wiedemann) (Mourikis 1965), 12 to 14 days in Dacustryoni (Froggatt) (Fletcher 1969), and 7 to 8 days in Rhagoletis pomonella(Walsh) (Prokopy et al. 1972). Many tephritids require a proteinaceousfood source (usually honeydew) during this period for development to sexualmaturity (Bateman 1972).
It was reported by Landolt and Hendrichs (1983) that female papayafruit flies, Toxotrypana curvicauda Gerstaecker, observed in field cages matedand oviposited only when 6 days old or older, suggesting an obligatory pre-
lInsect Attractants. Behavior & Basic Biology Research Laboratory, USDA, ARS, P.O. Box14565, Gainesville, FL 82604 USA.

Landolt: Toxotrypana curvicauda 241
mating period. This paper describes the growth of the ovaries of femalepapaya fruit flies after emergence and the mating propensity of males andfemales of different ages. The primary purpose was to determine if such anobligatory pre-mating period exists in the papaya fruit fly and how long itlasts.
MATERIALS AND METHODS
Flies used in laboratory tests were obtained from infested papaya fruits,Carica papaya L., collected in a commercial grove in Dade Co., Florida fromFebruary through July 1982. Mature larvae from these fruit pupated inmoist soil placed 7 cm deep in 8 oz wax-cartons. These cartons of soil withpupae were kept at 26-28°C and 60-80% RH until adult emergence. Emergedflies were sorted by sex daily (at 8-9 AM EST) and placed in (24 cm) 3
screen cages with sugar cubes and a wet sponge.Fifteen female papaya fruit flies of each age from 1 to 8 days old (1 day
= 6 to 30 hold) were dissected to measure ovarian development. Dissectionswere made under 80% ethanol using a Bausch and Lomb binocular microscope. All measurements were made at lOx with the aid of an ocular micrometer.
Flies tested for mating propensity were placed in 8 oz wax-coated cartonswith screen on both top and bottom, with 1 pair of flies per carton. Testswere initiated at 1 :30-2 :00 PM EST and pairs were observed for matingevery 0.5 h thereafter until 4 :00 PM. Mating observed in field cages washighest at this time of day (Landolt and Hendrichs 1983). All combinationsof ages, from 1 to 6 days old, were tested for mating propensity with 40pairs (4 groups of 10 pairs) per age combination. Groups of flies in whichmating occurred were not used in subsequent mating tests.
To obtain an estimate of the normal maximum size of papaya fruit flyovaries, 72 adult females were netted in a papaya grove during March 1982.These were dissected and the ovaries measured to determine the normal rangeof ovary size in a field population. Females collected while ovipositing(n=32) were considered mature and were evaluated separately to determinethe size of ovaries in reproductively mature females.
RESULTS
Emerging female papaya fruit flies had undeveloped ovaries withoutmature eggs. The average length and width of ovaries of 1-day-old flies were1.25 and 0.44 mm respectively (Fig. 1), while the maximum egg lengthaverage 0.23 mm (Fig. 2). In most flies at this age the abdomen was filledwith fat body as clusters of fat body cells. The length and width of ovariesand length of eggs increased nearly linearly for 6 days after emergence,decreasing thereafter. The average length and width of ovaries peaked at3.14 and 0.75 mm respectively and the maximum length of eggs was 1.3 mmin females 6 days old. The fat body of 3 to 4-day-old flies was restricted tothe posterior area of the abdomen but had disappeared in 6 to 7-day-oldfemales in which the ovaries had reached maximum size.
Ovipositing female papaya fruit flies collected in the commercial papayagrove had ovaries with an average length of 3.1 mm (SE = 0.3) and widthof 0.9 mm (SE = 0.1), comparable to that of 5 to 6-day-old flies in the labo-
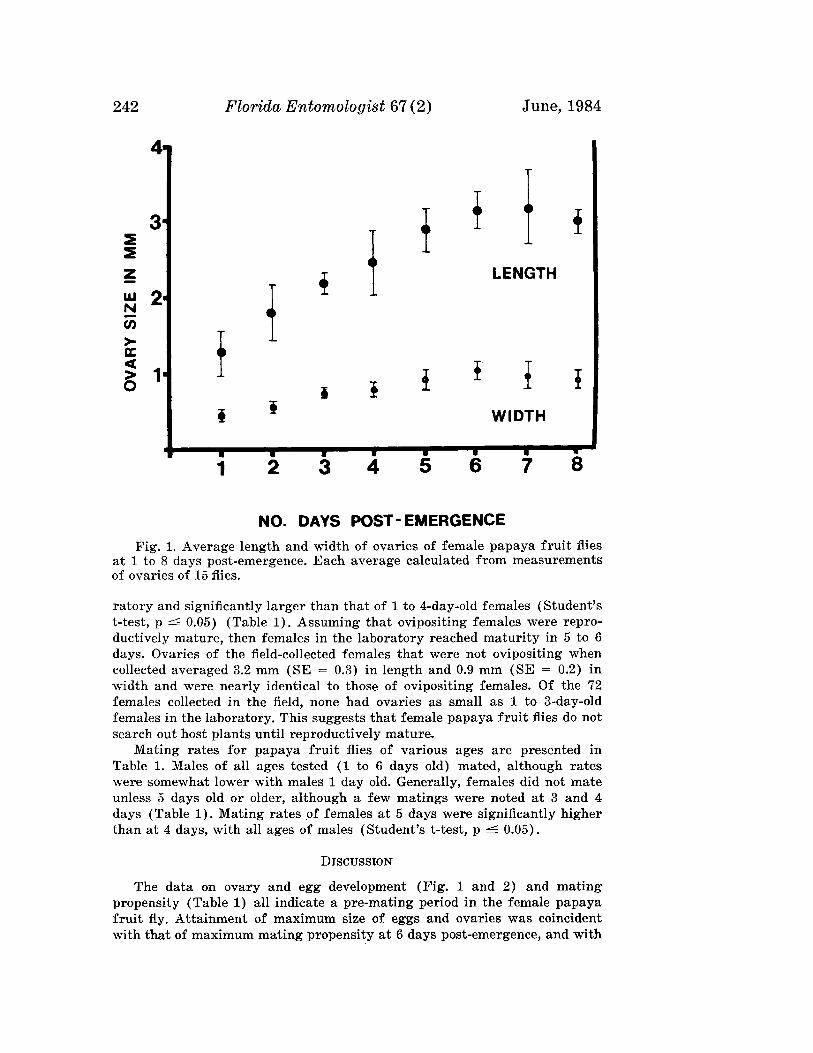
242 Florida Entomologist 67 (2) June, 1984
LENGTH
WIDTH
f
IIi
~~
z-w 2N-en>0:oct~ 1
1 2 3 4 5 6 7 8
NO. DAYS POST- EMERGENCE
Fig. 1. Average length and width of ovaries of female papaya fruit fliesat 1 to 8 days post-emergence. Each average calculated from measurementsof ovaries of 15 flies.
ratory and significantly larger than that of 1 to 4-day-old females (Student'st-test, p ~ 0.05) (Table 1). Assuming that ovipositing females were reproductively mature, then females in the laboratory reached maturity in 5 to 6days. Ovaries of the field-collected females that were not ovipositing whencollected averaged 3.2 mm (SE = 0.3) in length and 0.9 mm (SE = 0.2) inwidth and were nearly identical to those of ovipositing females. Of the 72females collected in the field, none had ovaries as small as 1 to 3-day-oldfemales in the laboratory. This suggests that female papaya fruit flies do notsearch out host plants until reproductively mature.
Mating rates for papaya fruit flies of various ages are presented inTable 1. Males of all ages tested (1 to 6 days old) mated, although rateswere somewhat lower with males 1 day old. Generally, females did not mateunless 5 days old or older, although a few matings were noted at 3 and 4days (Table 1). Mating rates of females at 5 days were significantly higherthan at 4 days, with all ages of males (Student's t-test, p ~ 0.05).
DISCUSSION
The data on ovary and egg development (Fig. 1 and 2) and matingpropensity (Table 1) all indicate a pre-mating period in the female papayafruit fly. Attainment of maximum size of eggs and ovaries was coincidentwith that of maximum mating propensity at 6 days post-emergence, and with
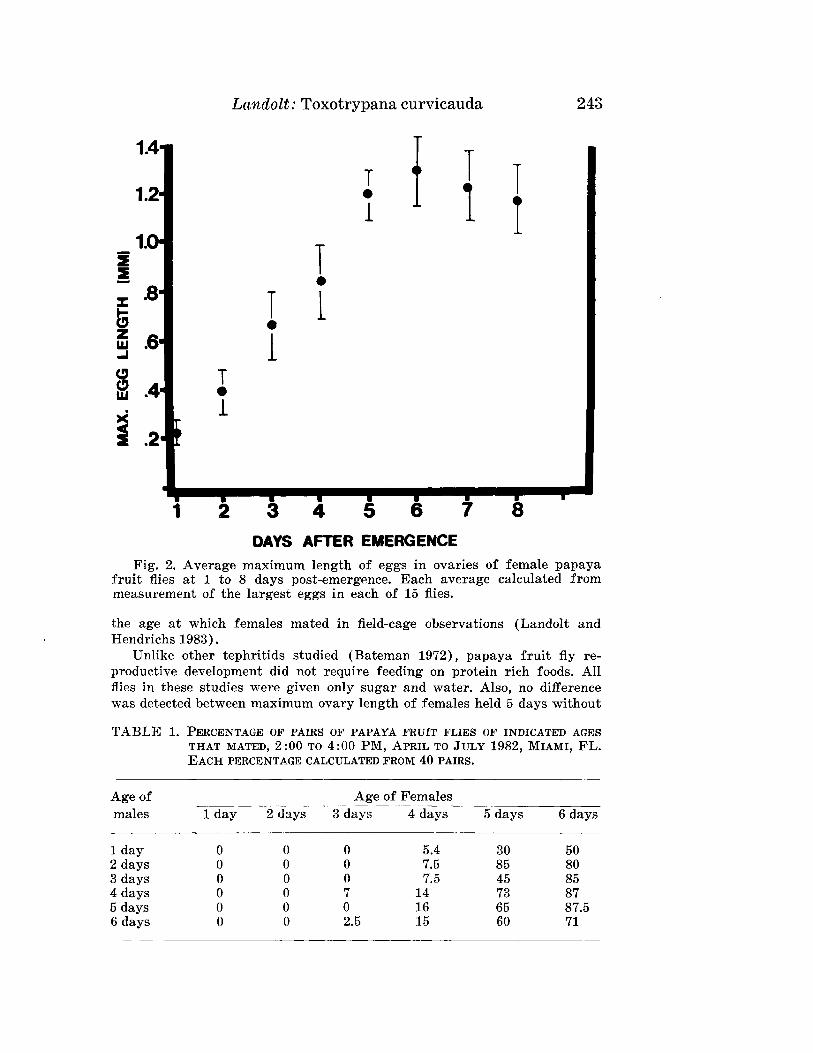
Landolt: Toxotrypana curvicauda 243
::z::t;ZW...I
T•1
T•1
T•1
T•1
f T1 T
I
1 2 3 4 5 6
DAYS AFTER EMERGENCE
Fig. 2. Average maximum length of eggs in ovaries of female papayafruit flies at 1 to 8 days post-emergence. Each average calculated frommeasurement of the largest eggs in each of 15 flies.
the age at which females mated in field-cage observations (Landolt andHendrichs 1983).
Unlike other tephritids studied (Bateman 1972), papaya fruit fly reproductive development did not require feeding on protein rich foods. Allflies in these studies were given only sugar and water. Also, no differencewas detected between maximum ovary length of females held 5 days without
TABLE 1. PERCENTAGE OF PAIRS OF PAPAYA FRUIT FLIES OF INDICATED AGESTHAT MATED, 2:00 TO 4:00 PM, APRIL TO JULY 1982, MIAMI, FL.EACH PERCENTAGE CALCULATED FROM 40 PAIRS.
Age of Age of Femalesmales 1 day 2 days 3 days 4 days 5 days 6 days
1 day 0 0 0 5.4 30 502 days 0 0 0 7.5 85 803 days 0 0 0 7.5 45 854 days 0 0
,.,14 73 87I
5 days 0 0 0 16 65 87.56 days 0 0 2.5 15 60 71

244 Florida. Entomologist 67 (2) June, 1984
a protein source (2.8 ± 0.6 mm, n=25) or with yeast hydrolysate paste (2.84± 0.4 mm, n=25). Apparently, enough reserves are stored during the larvalstages to permit reproductive development without such feeding. This maybe related to young larvae feeding on the more nutritious seed embryoswithin the fruit (Mason 1922) rather than on fruit pulp as do otherfrugiverous tephritid larvae.
No premating period was evident in male papaya fruit flies. Toxotrypanacurvicauda apparently is the first tephritid species for which such a sharpdifference in reproductive maturation between the sexes has been demonstrated. Maximum male mating propensity was reached by the second dayafter emergence, coinciding with the age at which my personal observationsindicate that males begin puffing (indicative of pheromone release).
ACKNOWLEDGMENTS
Technical assistance was provided by L. Lichtenstein and B. Brown.R. Nelson kindly allowed use of his papaya grove. Spanish translation of theabstract was provided by Alicia Arner and Maria Nieto.
REFERENCES CITED
BATEMAN, M. A. 1972. The ecology of fruit flies. Ann. Rev. Ent. 17: 493-518.FLETCHER, B. S. 1969. The structure and function of the sex pheromone
glands of the male Queensland fruit fly, Dacus tryoni. J. InsectPhysiol. 15: 1309-22.
LANDOLT, P. J., AND J. HENDRICHS. 1983. Behavior of the papaya fruit fly,Toxotrypana curvicauda Gerstaecker (Diptera: Tephritidae). AnnalsEnt. Soc. America 76: 413-7.
MASON, S. C. 1922. Biology of the papaya fruit fly, Toxotrypana curvicauda,in Florida. USDA Bull. 1081: 1-9.
MAZOMENOS, B., J. L. NATION, W. J. COLEMAN, K. C. DENNIS, ANDR. ESPONDA. 1977. Reproduction in Caribbean fruit flies: Comparison between a laboratory strain and a wild strain. Florida Ent. 60:139-44.
MOURIKIS, P. A. 1965. Data concerning the development of the immaturestages of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann)(Diptera: Tephritidae) in different host fruits and on artificial mediaunder laboratory conditions. Ann. Inst. Phytopathol., Benaki, 7 (N.S.) :59-105.
NATION, J. L. 1972. Courtship behavior and evidence for a sex attractant inthe male Caribbean fruit fly, Anastrepha suspensa. Ann. Ent. Soc.America 1364-7.
---. 1974. The structure and development of two sex specific glands inmale Caribbean fruit flies. Ann. Ent. Soc. America 67: 731-4.
PROKOPY, R. J., E. W. BENNETT, AND G. L. BUSH. 1972. Mating behavior inRhagoletis pomonella. II. Temporal organization. Canadian Ent. 104:97-104.

Lin & Harper: Entomophthora gammae Culture 245
ISOLATION AND CULTURE OFENTOMOPHTHORA GAMMAE,
A FUNGAL PARASITE OF NOCTUID LARVAE
JING-SHYAN LIN AND JAMES D. HARPER
Department of Zoology-EntomologyAlabama Agricultural Experiment Station
Auburn University, AL 36849 USA
ABSTRACT
Entomophthora gammae (Weiser) was isolated by allowing conidia frominfected soybean looper, Pseudoplusia includens (Walker), larvae to showeronto egg yolk agar. Fungal and bacterial contaminants were always present,but at 20°C, E. gammae grew fast enough to allow the subculture of hyphalbodies to clean medium. Once isolated, the fungus was cultured on Czapeksolution agar, potato dextrose agar, Sabouraud dextrose agar, andSabouraud maltose agar, all containing 0.2% yeast extract, and on Grace'sinsect tissue culture medium. It did not grow on Wolf's medium, potatoagar or potato dextrose agar without yeast extract. The fungus grew mostrapidly in culture when maintained at 25°C and 100% RH. Conidia on glassslides held at 25°C germinated only at 100% RH. A gradual increase inconidium size occurred over a 10-month period of continuous culture. Attempts to infect larvae with the cultured fungus were unsuccessful.
RESUMEN
Entomophthora gammae (Weiser) fue aislado permitiendo que lasconidias provenientes de las larvas del medidor de la soya, Pseudoplusiaincludens (Walker) se dispersaran sobre agar de la yema de huevo degallina. Hongos y bacterias contaminantes estuvieron siempre presentes,pero a 20°C, E. gammae creci6 10 suficientemente rapido para permitir lasubcultura de cuerpos hifales para clarificar el medio. Una vez aislado, elhongo fue cultivado en una soluci6n de agar Czapek, papa dextrosa agar,Sabouraud dextrosa agar y Sabouraud maltosa agar, todos los medios contenian 0.2% de extracto de levadura y sobre el medio Grace para cultivo detejidos de insectos. No hubieron crecimientos sobre el medio Wolf, papa agaro papa dextrosa agar cuando no se agreg6 extracto de levadura. El crecimiento maximo y la germinaci6n de las esporas ocurri6 a los 25 ° C y con un100% de humedad relativa. Un incremento gradual del tamafio de lasconidias ocurri6 durante los 10 meses de cultivos continuos. Intentos parainfestar larvas con el hongo cultivado no tuvieron exito.
Several species of the noctuid subfamily Plusiinae are susceptible toinfection by Entomophthora (Tarichium) gammae (Weiser). Harper andCarner (1973) reported this pathogen from soybean looper, Pseudoplusiaincludens (Walker), and cabbage looper, Trichoplusia ni (Hubner), andWeiser (1965) reported it from Plusia gamma (L.) in Europe. It is a widespread pathogen of P. includens in the southeastern United States wherethis pest attacks soybean (Harper et al. 1983). Several studies on thispathogen have defined environmental conditions which are related to itsability to induce and maintain epizootics (Newman and Carner 1974,1975a,b). This report presents results of work on the isolation and culture

246 Florida Entomologist 67 (2) June, 1984
of E. gammae, on defining conditions for in vitro growth of the fungus, andon laboratory infectivity studies.
METHODS AND MATERIALS
ISOLATION
Fourth- and fifth-instar P. includens larvae were collected on soybeansin fields near Tallassee, Alabama. At the time of collection, an epizootic ofE. gammae was in progress. Larvae were placed in individual 30-ml cupscontaining meridic diet (Shorey and Hale 1965) and returned to the laboratory. Larvae infected with E. gammae typically died in late afternoonand rapidly turned an abnormal pale yellow color. Dead larvae were tapedto the inside of the lids of 20 x 100 mm petri dishes containing either eggyolk agar (EYA), potato agar, potato dextrose agar (PDA), Sabouraudmaltose agar, Sabouraud dextrose agar, or Wolf's medium (Wolf 1951).Plates of each medium were held at 20 and 25°C. By the following morning,conidiophores and conidia had been produced, the latter having showeredonto the medium surface below.
Once Entomophthora could be identified microscopically on the plates,it was transferred to fresh medium using a sterile inoculating needle. Bytransferring at this stage, contaminants, which were always numerous,were left behind. Several passages were sometimes necessary to completelyeliminate all contaminants.
CULTURE
Once isolated, small quantities of hyphae were transferred with sterileinoculating loops to the centers of plates containing various media or intotubes of liquid media and maintained at 20 or 25°C. Transfers were madeevery 14 days to maintain actively growing cultures. Cultures were alsoheld at 10°C and transferred every 2 months without loss of viability.
GROWrH AND GERMINATION
Plates of EYA and PDA plus 0.2% yeast extract were inoculated withE. gammae as described above and incubated in temperature cabinets maintained at 5 ± 1 degree intervals from 5 to 35°C. Colony size was determineddaily for 13 days by placing each dish over a cool light source and recordingthe average of two diameter measurements taken at right angles to eachother.
Seven petri dishes containing pure sporulating cultures of E. gammaewere inverted over seven EYA plates and allowed to shower for 3-4 hours.Inoculated plates were covered and maintained at the temperatures used inthe growth rate studies. Plates were examined daily under a microscope todetermine the percent germination of 100 conidia.
Humidity chambers were established using glass desiccator jars containing saturated salt solutions to provide the following humidity levels(Solomon 1951); <1% (CaCI, without water), 30% (CaCI, • 6H,O), 50%[CO (NO,) , • 4H,O], 70% (NH.CI + KNO,) , 90% (ZnSO•• 7H,O), 100%(distilled water). Humidity in each chamber was checked with a humidity

Lin & Harper: Entomophthora gammae Culture 247
gauge (Bacharach Industrial Instrument Co.), allowing sufficient time afterclosure for the humidity level to stabilize.
Sporulating cultures of E. gammae were inverted over glass slides for3-4 h, and the slides were introduced into the chambers. All chambers wereheld at room temperature (ca. 25°C) under fluorescent lighting. Slides wereremoved from the chambers at ca. 5-h intervals for 1 day and again at 57 h.Percent germination was determined at each interval from the first 100conidia encountered while scanning the central area of the slide.
INFECTIVITY STUDIES
Field collected cadavers exhibiting symptoms of E. gammae infectionwere suspended over individual healthy, laboratory-reared P. includenslarvae held inside petri dishes. Seven larvae were successfully exposed inthis manner. Ten larvae were exposed to conidia showering from each of 3separate, inverted culture plates which had been maintained in the laboratory through 17 transfers over a I-year period. After 4-6 hours exposure, alllarvae were transferred to individual I-ounce cups containing diet and wereheld at either 90 or 100% RH until they pupated or died.
RESULTS
ISOLATION
Although several media supported growth of E. gammae, EYA held at20°C was the only medium-temperature combination from which a pureculture could be initiated. The inoculation technique introduced many bacterial and fungal contaminants which at 25°C overgrew the plates beforeE. gammae colonies could become established. These contaminants grewmore slowly at 20°C on EYA, and E. gammae colonies were able to developvegetatively and could be transferred to clean plates before being overgrown. After the transfer of hyphal bodies, the mycelium grew vegetativelyfor 2-3 days; then hyphal bodies produced conidia which were ejected.Those landing on fresh medium germinated, produced germ tubes whichelongated, thickened and grew vegetatively, repeating the above cycle. Thisprocess continued as a series of expanding concentric rings of growth untilthe petri dish limited further growth.
CULTURE
Hyphal bodies transferred to media other than EYA exhibited varyinggrowth responses. Potato agar, Wolf's medium, and PDA did not supportgrowth. A very weak growth was obtained in Czapek solution containing0.2% yeast extract. PDA + 0.2% yeast extract supported good growth,while Sabouraud dextrose agar and Sabouraud maltose agar, both containing 0.2% yeast extract, supported even greater growth. Greatest growthoccurred on EYA containing 0.2% yeast extract and on Grace's insect tissueculture medium. On EYA held at 25°C, E. gammae, following the growthcycle described above, covered a 10 cm plate within 1 week. On Grace'smedium, the inoculum floated to the surface and grew as a white, compactmass.
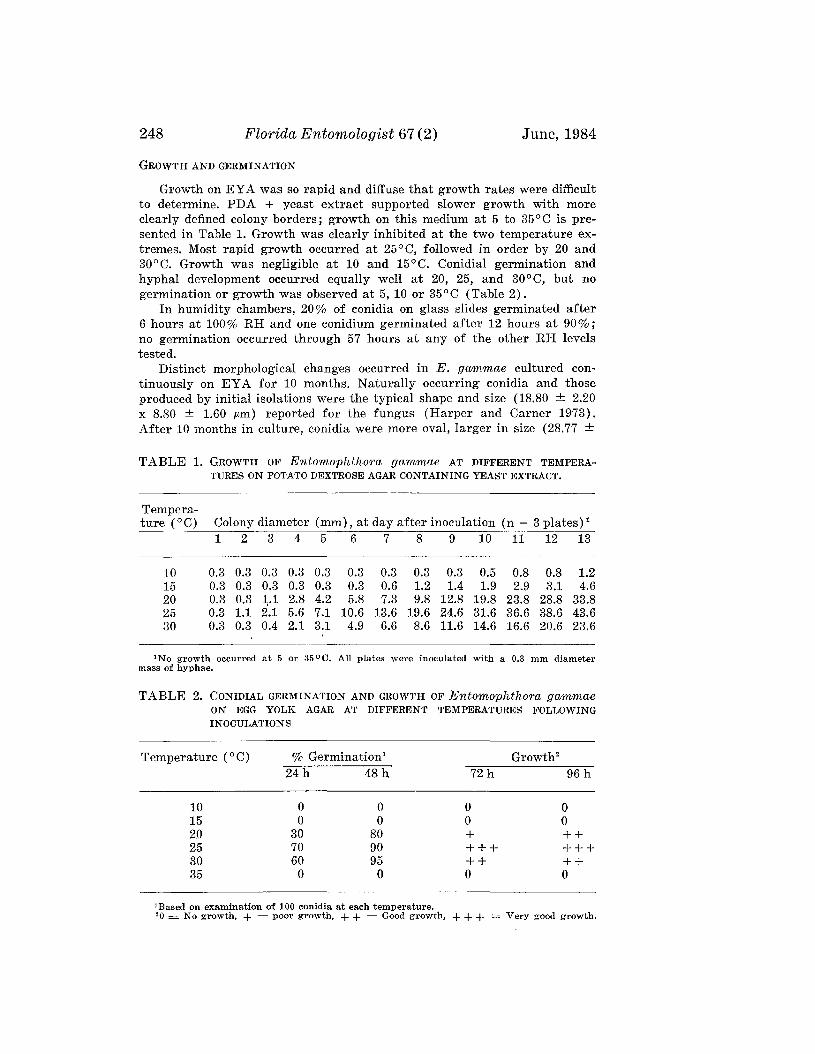
248 Florida Entomologist 67 (2) June, 1984
GROWTH AND GERMINATION
Growth on EYA was so rapid and diffuse that growth rates were difficultto determine. PDA + yeast extract supported slower growth with moreclearly defined colony borders; growth on this medium at 5 to 35°C is presented in Table 1. Growth was clearly inhibited at the two temperature extremes. Most rapid growth occurred at 25°C, followed in order by 20 and30°C. Growth was negligible at 10 and 15°C. Conidial germination andhyphal development occurred equally well at 20, 25, and 30°C, but nogermination or growth was observed at 5, 10 or 35°C (Table 2).
In humidity chambers, 20% of conidia on glass slides germinated after6 hours at 100% RH and one conidium germinated after 12 hours at 90%;no germination occurred through 57 hours at any of the other RH levelstested.
Distinct morphological changes occurred in E. gammae cultured continuously on EYA for 10 months. Naturally occurring conidia and thoseproduced by initial isolations were the typical shape and size (18.80 ± 2.20x 8.80 ± 1.60 ,urn) reported for the fungus (Harper and Carner 1973).After 10 months in culture, conidia were more oval, larger in size (28.77 ±
TABLE 1. GROWTH OF Entomophthora gammae AT DIFFERENT TEMPERA-TURES ON POTATO DEXTROSE AGAR CONTAINING YEAST EXTRACT.
Tempera-ture (OC) Colony diameter (mm), at day after inoculation (n = 3 plates) 1
1 2 3 4 5 6 7 8 9 10 11 12 13
10 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.5 0.8 0.8 1.215 0.3 0.3 0.3 0.3 0.3 0.3 0.6 1.2 1.4 1.9 2.9 3.1 4.620 0.3 0.3 ~.1 2.8 4.2 5.8 7.3 9.8 12.8 19.8 23.8 28.8 33.825 0.3 1.1 2.1 5.6 7.1 10.6 13.6 19.6 24.6 31.6 36.6 38.6 43.630 0.3 0.3 0.4 2.1 3.1 4.9 6.6 8.6 11.6 14.6 16.6 20.6 23.6
'No growth occurred at 5 or 35°C. All plates were inoculated with a 0.3 mm diametermass of hyphae.
TABLE 2. CONIDIAL GERMINATION AND GROWTH OF Entomophthora gammaeON EGG YOLK AGAR AT DIFFERENT TEMPERATURES FOLLOWINGINOCULATIONS
Temperature (OC) % Germination' Growth'24 h 48h 72 h 96 h
10 0 a 0 015 a a a 020 30 80 + ++25 70 90 +++ +++30 60 95 ++ ++35 a 0 a a
'Based on examination of 100 conidia at each temperature.'0 = No growth, + = poor growth, + + = Good growth, + + + = Very good growth.

Lin & Harper: Entomophthora gammae Culture 249
4.34 x 19.85 ± 4.65 /lm), and contained larger and more numerous oilglobules. In culture, conidiophores were difficult to find. Conidia werenormally formed and ejected from the tips of the stocky hyphal bodies. Themembrane or gelatinous layer which surrounds the naturally occurringconidium is often quite thick in relation to conidium size (Harper andCarner 1973). After continuous culture, this layer became very thin.
Repeated attempts to infect P. includens larvae with conidia from cultures of E. gammae were unsuccessful, and only one of 7 larvae became infected after exposure to freshly sporulating, field collected larvae. The infected larva died after 7 days, became mumonified, retained its shape, andbecame glossy black in color. On dissection, the cadaver was found to befilled with typical E. gammae resting spores as described by Harper andCarner (1973).
DISCUSSION
There is considerable information on optimal conditions for growth anddevelopment of many species in the Entomophthoraceae. Members of theEntomophthoraceae vary considerably in their nutritive requirements.Coagulated egg yolk or media containing egg yolk has proven satisfactoryfor culturing many species (see review by King and Humber, 1981). EYAwas clearly the optimal solid growth medium for E. gammae in this study.Once isolated, the fungus grew on a number of standard mycological mediaif supplemented with yeast extract. While resting spores are commonlyproduced by E. gammae under field conditions (Harper and Carner 1973,Newman and Carner 1975b), they were never produced on any mediumunder any of the conditions utilized in this study.
Temperature requirements for growth of the Entomophthoraceae varyconsiderably (Gustafsson 1965, Hall and Bell, 1960, 1961). In this study,the optimal temperature for growth of E. gammae in vitro was 25°C orslightly lower. Spore germination on medium also appeared to be optimal inthe same temperature range, which agrees with an earlier report by Newman and Carner (1975a), who found that sporulation by E. gammae oncadavers was maximal at 21 and 26.7°C.
The procedures used to determine optimal humidity conditions for sporegermination served only as an indication of this requirement, as the response was very low. However, it is clear that germination occurred only ator near conditions of 100% humidity, which is also in agreement with results of Newman and Carner (1975a). They found no germination ofconidia of this species at 90% RH, only 1% at 95% RH, and 95% at 100%RH.
Lack of infectivity by the cultured fungus may have resulted from theenvironmental conditions or techniques employed, or from a loss of infectiousness following repeated subculturing. The low level of infectivity usingfresh cadavers as the inoculum source suggests that the inoculation technique was inadequate. However, the gradual change in spore morphologywith continued culture indicates that a selection process was occurring,possibly for an organism that was better adapted for saprophytic growthat the expense of pathogenicity. This phenomenon would reinforce findingswith other pathogens which have shown the importance of maintainingminimal passage stock cultures to insure maintenance of desireable char-

250 Florida Entomologist 67 (2) June, 1984
acteristics in cultures used for experimental or production purposes.Alabama Agricultural Station Journal No. 15-83449.
REFERENCES CITED
GUSTAFSSON, M. 1965. On species of the genus Entomophthora Fres. inSweden II. Cultivation and physiology. Lantbr. Hogsk. Annlr. 31:405-57.
HALL, I. M., AND J. V. BELL. 1960. The effect of temperature on someentomophthoraceous fungi. J. Insect Pathol. 2: 247-53.
---. 1961. Further studies on the effect of temperature on the growthof some entomophthoraceous fungi. J. Insect Pathol. 3: 289-96.
HARPER, J. D., AND G. R. CARNER. 1973. Incidence of Entomophthora sp. andother natural control agents in populations of Pseudoplusia includensand Trichoplusia ni. J. Invertebr. Pathol. 22: 80-5.
HARPER, J. D., M. SHEPARD, AND R. M. MCPHERSON. 1983. Geographical andseasonal occurrence of parasites, predators and entomopathogens.Pages 7-19 In "Natural Enemies of Arthropod Pests in Soybeans".H. R. Petri, ed. Southern Coop. Ser. Bull. 285. 90 p.
KING, D. S., AND R. A. HUMBER. 1981. Identification of the Entomophthorales. Pages 107-127 In "Microbial Control of Pest and PlantDiseases 1970-1980". H. D. Burges, ed. Academic Press, London.
NEWMAN, G. G., AND G. R. CARNER. 1974. Diel periodicity of Entomophthoragammae in the soybean looper. Environ. Ent. 3: 888-90.
---. 1975a. Environmental factors affecting conidial sporulation andgermination of Entomophthora gammae. Environ. Ent. 4: 615-8.
---. 1975b. Factors affecting the spore form of Entomophthora gammae.J. Invertebr. Pathol. 26: 29-34.
SHOREY, H. H., AND R. L. HALE. 1965. Mass-rearing of the larvae of ninenoctuid species on a simple artificial medium. J. Econ. Ent. 58: 522-4.
SOLOMON, M. E. 1951. Control of humidity with potassium hydroxide, sulfuric acid, or other solutions. Bull. Ent. Res. 42: 543-53.
WEISER, J. 1965. Not~s on two new species of the genus Tarichium Cohn(Entomophthoraceae). Ceska Myko. 19: 201-4.
WOLF, F. T. 1951. The cultivation of two species of Entomophthora onsynthetic media. Bull. Torrey Bot. Club 78: 211-20.
• • • • • • • • • • • . . ..LIRIOMYZA TRIFOLII:
OVIPOSITION AND DEVELOPMENT IN FOLIAGEOF TOMATO AND COMMON WEED HOSTS
T. G. ZOEBISCH, D. J. SCHUSTER AND J. P. GILREATHAgricultural Research & Education Center
5007-60th Street EastBradenton, FL 34203 USA
ABSTRACT
Oviposition and development of Liriomyza trifolii (Burgess) werestudied in the laboratory in foliage of tomato, Lycopersicon esculentumMill. cv. 'Walter,' a common nightshade, Solanum nodifiorum Jacq., common beggar-tick, Bidens alba (L.) DC., and downy groundcherry, Physalispubescens L. Liriomyza trifolii oviposited successfully in foliage of all plant

250 Florida Entomologist 67 (2) June, 1984
acteristics in cultures used for experimental or production purposes.Alabama Agricultural Station Journal No. 15-83449.
REFERENCES CITED
GUSTAFSSON, M. 1965. On species of the genus Entomophthora Fres. inSweden II. Cultivation and physiology. Lantbr. Hogsk. Annlr. 31:405-57.
HALL, I. M., AND J. V. BELL. 1960. The effect of temperature on someentomophthoraceous fungi. J. Insect Pathol. 2: 247-53.
---. 1961. Further studies on the effect of temperature on the growthof some entomophthoraceous fungi. J. Insect Pathol. 3: 289-96.
HARPER, J. D., AND G. R. CARNER. 1973. Incidence of Entomophthora sp. andother natural control agents in populations of Pseudoplusia includensand Trichoplusia ni. J. Invertebr. Pathol. 22: 80-5.
HARPER, J. D., M. SHEPARD, AND R. M. MCPHERSON. 1983. Geographical andseasonal occurrence of parasites, predators and entomopathogens.Pages 7-19 In "Natural Enemies of Arthropod Pests in Soybeans".H. R. Petri, ed. Southern Coop. Ser. Bull. 285. 90 p.
KING, D. S., AND R. A. HUMBER. 1981. Identification of the Entomophthorales. Pages 107-127 In "Microbial Control of Pest and PlantDiseases 1970-1980". H. D. Burges, ed. Academic Press, London.
NEWMAN, G. G., AND G. R. CARNER. 1974. Diel periodicity of Entomophthoragammae in the soybean looper. Environ. Ent. 3: 888-90.
---. 1975a. Environmental factors affecting conidial sporulation andgermination of Entomophthora gammae. Environ. Ent. 4: 615-8.
---. 1975b. Factors affecting the spore form of Entomophthora gammae.J. Invertebr. Pathol. 26: 29-34.
SHOREY, H. H., AND R. L. HALE. 1965. Mass-rearing of the larvae of ninenoctuid species on a simple artificial medium. J. Econ. Ent. 58: 522-4.
SOLOMON, M. E. 1951. Control of humidity with potassium hydroxide, sulfuric acid, or other solutions. Bull. Ent. Res. 42: 543-53.
WEISER, J. 1965. Not~s on two new species of the genus Tarichium Cohn(Entomophthoraceae). Ceska Myko. 19: 201-4.
WOLF, F. T. 1951. The cultivation of two species of Entomophthora onsynthetic media. Bull. Torrey Bot. Club 78: 211-20.
• • • • • • • • • • • . . ..LIRIOMYZA TRIFOLII:
OVIPOSITION AND DEVELOPMENT IN FOLIAGEOF TOMATO AND COMMON WEED HOSTS
T. G. ZOEBISCH, D. J. SCHUSTER AND J. P. GILREATHAgricultural Research & Education Center
5007-60th Street EastBradenton, FL 34203 USA
ABSTRACT
Oviposition and development of Liriomyza trifolii (Burgess) werestudied in the laboratory in foliage of tomato, Lycopersicon esculentumMill. cv. 'Walter,' a common nightshade, Solanum nodifiorum Jacq., common beggar-tick, Bidens alba (L.) DC., and downy groundcherry, Physalispubescens L. Liriomyza trifolii oviposited successfully in foliage of all plant

Zoebisch et al.: Liriomyza trifolii 251
species. However, females exposed simultaneously to all plant species deposited more eggs per cm2 of tomato foliage than per cm2 of foliage of theother plant species. Differences among plant species in percent egg hatch,percent pupation or pupal weight of L. trifolii were not significant. Larvaepupated most rapidly from foliage of nightshade.
RESUMEN
Se estudi6 en el laboratorio la oviposici6n y el desarrollo de Liriomyzatrifolii (Burgess) en follaje de tomate, Lycopersicon esculentum Mill.variedad 'Walter,' Solanum nodifiorum Jacq., Bidens alba (L.) y Physalispubescens L. Liriomyza t1'ifolii oviposit6 con buen exito en el follaje de todaslas especies antes mencionadas. Sin embargo, las hembras expuestas simultaneamente a todas las especies de plantas depositaron mas huevecillos porcm2 en el follaje de tomate que en las demas especies de plantas. No seencontraron diferencias significativas entre las especies de plantas conrespecto a la eclosi6n, porciento de pupaci6n y peso pupal de L. trifolii. Laslarvas que provenian del follaje de Solanum nodifiorum puparon masrapidamente.
Liriomyza spp. leafminers are considered important pests of tomatoalthough evidence indicates they are secondary in nature (Oatman andKennedy 1976, Johnson et al. 1980). Nevertheless, in Florida, if populationsare not controlled, up to 70% by weight of the 5th harvest may be lost(Schuster, unpublished data).
An understanding of the role of alternative host plants in the population dynamics of the pest is important for the management of any pest.Alternative host plants may serve as sources of epidemic numbers of pestsor as sources of low levels of pests which may initiate and maintain infestations in crop plants.
The most abundant leafminer attacking tomato on the west coast ofFlorida at the present time is L. trifolii (Burgess) (Schuster, unpublisheddata). The role of alternative host plants in the population dynamics ofthis pest is not known. The host range of L. trifolii has been summarizedby Stegmaier (1966), Spencer (1973), and Spencer and Stegmaier (1973).However, no records are available regarding the relative suitability of weedhosts for L. trifolii.
As a first step in elucidating the role of weeds in L. trifolii populationdynamics, we compared oviposition, egg hatch and larval development ofL. trifolii in foliage of tomato, eggplant and selected weeds. Nightshade,downy groundcherry and common beggar-tick were selected for evaluationbecause 80-95% of the Liriomyza larvae observed on weeds on the perimeters of commercial tomato fields on the west coast of Florida during thespring of 1982 occurred on these plants (Schuster, unpublished data). Pigweed was included as a less favorable host (as determined by field observations) .
MATERIALS AND METHODS
Experiments were conducted in a room at ca. 27°C and 12 h photoperiod.Liriomyza trifolii adults for all experiments were obtained from a colonythat had been maintained for at least 3 generations on Phaseolus limensi.<r

252 Florida Entomologist 67 (2) June, 1984
var. limenanus L. H. Bailey cv. 'Henderson' bush bean by techniques similarto those of Ketzler and Price (1982). Four to 6 week old seedlings of tomato(Lycopersicon esculentum Mill., cv. 'Walter'), a common nightshade(Solanum nodiflorum Jacq.), downy groundcherry (Physalis pubescens L.),common beggar-tick (Bidens alba (L.) DC), pigweed (Amaranthus viridisL.) and eggplant (S. melongena L. cv. 'Florida Market') in 10-cm-diam.pots were used to evaluate the ovipositional response of flies. The first 4species were used in the experiments to evaluate development. Plants of similar age were used in each replication.
Percentage data were transformed by arcsine V%/100 prior to statisticalanalysis but are presented in the original scale. Two-way analyses of variance were performed on all data and, if significant F values (P = 0.05) forplant species effects were detected, Duncan's new multiple range test wasused to compare means.
To study ovipositional preference, 1 seedling of each of the 6 plantspecies was randomly assigned to a position in a circle within a 61 x 61 x 61cm cage covered with nylon organdy in a greenhouse. About 100 L. trifoliiadults were released into the cage for 24-48 h after which the numbers ofeggs deposited were counted with the aid of a dissecting microscope. Leafarea for each plant was measured to the nearest 0.1 cm2 with a LI-COR®Model LI-3000 area meter (Lambda Instruments Corp., Lincoln NB). Theadults were fed honey 24 h prior to testing and during the test were provided with a 9-cm diam. filter paper dipped in honey. The experiment wasreplicated 9 times.
To evaluate the impact of plant host on oviposition (in the absence ofalternative plant choices) and development of immature stages, clip cageswere constructed (Beegle et al. 1981) so that 4.5 cm2 of leaf surface couldbe enclosed. One male and 2 female L. trifolii were confined 24 h on theupper surface of the leaflet with a clip cage supported so that the plane ofthe leaflet was horizontal. One clip cage was used on each of 2 leaflets ofabout the same age from the middle of each plant. The adults were fedwith honey at least 24 h prior to testing to enhance oviposition. Two dropsof honey were also placed on the inside of each cage. Eggs were counted oneach leaflet and were marked as they hatched. To prevent overcrowding, thefirst 3 to 5 larvae per leaflet were allowed to survive while all others werekilled with a dissecting needle. In order to obtain puparia, the leaflets werecovered with polyethylene bags with small ventilation punctures. Pupariawere weighed to the nearest 0.01 mg with a semi-microbalance. Twelveplants were used per plant species for the oviposition, egg hatch and pupalweight measurements and 6 were used to determine the time to pupation.
RESULTS AND DISCUSSION
When L. trifolii females were permitted a choice among the plant spedes, significantly more eggs were deposited in foliage of tomato, nightshadeand eggplant than in foliage of the other plant species (Table 1). No eggswere deposited in foliage of pigweed. Because the leaf sizes varied amongthe plant species, egg counts were converted to a per cm2 basis. On an areabasis, significantly more eggs were observed in tomato foliage than infoliage of the other plant species. More eggs were deposited per cm2 ofnightshade foliage than per cm2 of downy groundcherry or pigweed foliage.
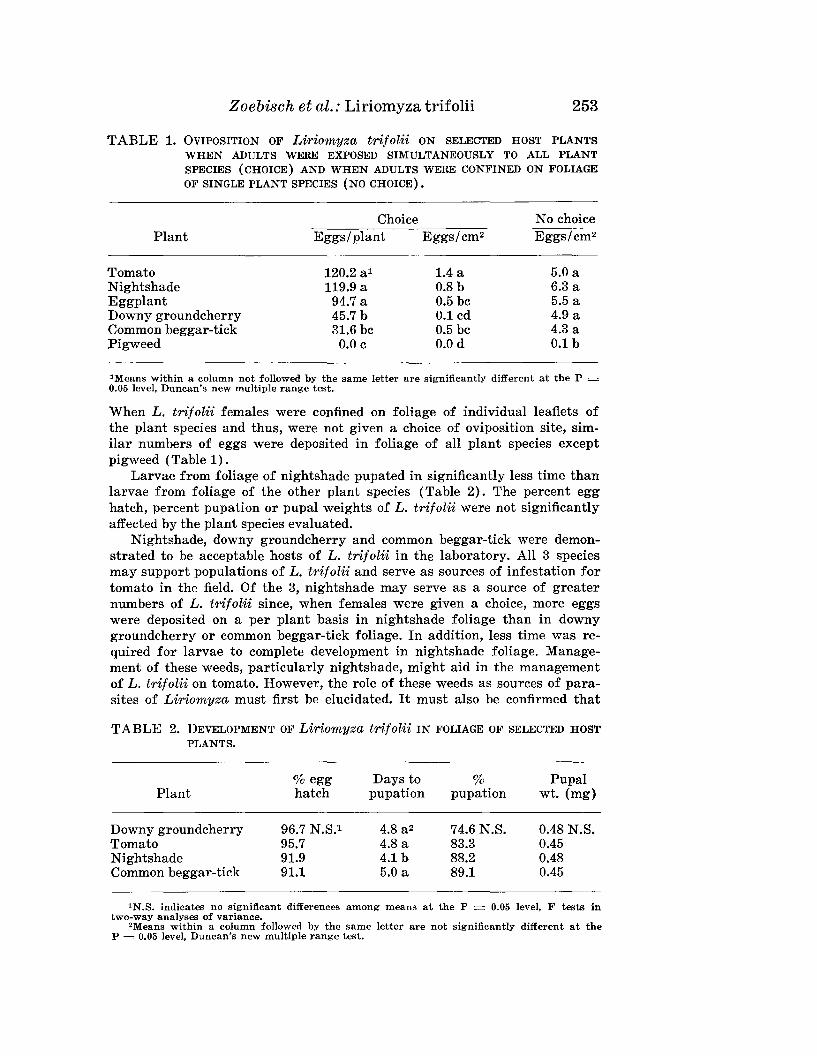
Z oebisch et al.: Liriomyza trifolii 253
TABLE 1. OVIPOSITION OF Liriamyza trifalii ON SELECTED HOST PLANTSWHEN ADULTS WERE EXPOSED SIMULTANEOUSLY TO ALL PLANTSPECIES (CHOICE) AND WHEN ADULTS WERE CONFINED ON FOLIAGEOF SINGLE PLANT SPECIES (NO CHOICE).
PlantChoice
Eggs/plant Eggs/cm2
No choiceEggs/cm2
TomatoNightshadeEggplantDowny groundcherryCommon beggar-tickPigweed
120.2 a l
119.9 a94.7 a45.7b31.6 bc
0.0 c
1.4 a0.8 b0.5 bc0.1 cd0.5 bc0.0 d
5.0 a6.3 a5.5 a4.9 a4.3 a0.1 b
'Means within a column not followed by the same letter are significantly different at the P =0.05 level, Duncan's new multiple range test.
When L. trifalii females were confined on foliage of individual leaflets ofthe plant species and thus, were not given a choice of oviposition site, similar numbers of eggs were deposited in foliage of all plant species exceptpigweed (Table 1).
Larvae from foliage of nightshade pupated in significantly less time thanlarvae from foliage of the other plant species (Table 2). The percent egghatch, percent pupation or pupal weights of L. trifolii were not significantlyaffected by the plant species evaluated.
Nightshade, downy groundcherry and common beggar-tick were demonstrated to be acceptable hosts of L. trifolii in the laboratory. All 3 speciesmay support populations of L. trifolii and serve as sources of infestation fortomato in the field. Of the 3, nightshade may serve as a source of greaternumbers of L. trifalii since, when females were given a choice, more eggswere deposited on a per plant basis in nightshade foliage than in downygroundcherry or common beggar-tick foliage. In addition, less time was required for larvae to complete development in nightshade foliage. Management of these weeds, particularly nightshade, might aid in the managementof L. trifalii on tomato. However, the role of these weeds as sources of parasites of Liriamyza must first be elucidated. It must also be confirmed that
TABLE 2. DEVELOPMENT OF Liriamyza trifolii IN FOLIAGE OF SELECTED HOSTPLANTS.
% egg Days to % PupalPlant hatch pupation pupation wt. (mg)
Downy groundcherry 96.7 N.S.l 4.8 a 2 74.6 N.S. 0.48 N.S.Tomato 95.7 4.8 a 83.3 0.45Nightshade 91.9 4.1 b 88.2 0.48Common beggar-tick 91.1 5.0 a 89.1 0.45
'N.S. indicates no significant differences among means at the P = 0.05 level, F tests intwo-way analyses of variance.
2Means within a column followed by the same letter are not significantly different at theP = 0.05 level, Duncan's new multiple range test.

254 Florida Entomologist 67 (2) June, 1984
L. trifolii reared from foliage of the weed species are not host-conditionedand will oviposit and develop on tomato.
ACKNOWLEDGMENT
We wish to thank Dr. David Han for identification of the weed speciesutilized in these experiments.
Florida Agricultural Experiment Stations Journal Series No. 4675.
REFERENCES CITED
BEEGLE, C. C., H. T. DULMAGE, D. A. WOLFENBARGER, AND E. MARTINEZ.1981. Persistance of Bacillus thuringiensis Berliner insecticidal activity on cotton foliage. Environ. Ent. 10: 400-1.
JOHNSON, M. W., E. R. OATMAN, AND J. A. WYMAN. 1980. Effects of insecticides on populations of the vegetable leafminer and associatedparasites on summer pole tomatoes. J. Econ. Ent. 73: 61-6.
!{ETZLER, L. D., AND J. F. PRICE. 1982. Methods for growers to evaluateeffects of their cultural practices on Liriomyzu, trifolii leafminers ina simple laboratory. Proc. Florida State Hort. Soc. 95: 162-4.
OATMAN, E. R., AND G. G. KENNEDY. 1976. Methomyl induced outbreak ofLiriomyza sativae on tomato. J. Econ. Ent. 69: 667-8.
SPENCER, K. A. 1973. Agromyzidae (Diptera) of Economic Importance.W. Junk B.V., The Hague. 418 p.
SPENCER, K. A., AND C. E. STEGMAIER, JR. 1973. Arthoropods of Floridaand Neighboring Lands, Vol. 7. Agromyzidae of Florida with a supplement on species from the Caribbean. Florida Dept. Agric. Consumer Serv., Div. Plant Industry, Bureau Ent. Contribution No. In.205 p.
STEGMAIER, C. E., JR. 1966. Host plants and parasites of Liriomyza trifoliiin Florida (Diptera: Agromyzidae) . Florida Ent. 49: 75-80.
• • • ., . • • • • • • • •METHYL BROMIDE FUMIGATION AS A
QUARANTINE TREATMENT FOR LATANIA SCALE,HEMIBERLESIA LATANIAE
(HOMOPTERA: DIASPIDIDAE)
PETER C. WITHERELLMiami Methods Development Station USDA, APHIS, PPQ
Miami, FL 33158 USA
ABSTRACT
Fumigation tests using methyl bromide at normal atmospheric pressureshowed that 16 glm' for 2% h at 22.8°C or above, or 32 g/m3 for 2% hat18.3 to 22.2°C was sufficient as a quarantine treatment against latania scale,Hemiberlesia lataniae on nursery stock. For use on avocado fruits, recommendations are for 24 g/m3 for 2% h at 22.8°C or above, and 32 glm' for3 h at 18.3 to 22.2°C. Microscopic examination of fumigated and controlscales showed that certain body movements, particularly pharyngeal pulsation, is a much more reliable indicator of scale viability than the merepresence of body fluids. Following fumigation, body fluids required up to31 days to dehydrate completely, under humid conditions.

254 Florida Entomologist 67 (2) June, 1984
L. tri/olii reared from foliage of the weed species are not host-conditionedand will oviposit and develop on tomato.
ACKNOWLEDGMENT
We wish to thank Dr. David Han for identification of the weed speciesutilized in these experiments.
Florida Agricultural Experiment Stations Journal Series No. 4675.
REFERENCES CITED
BEEGLE, C. C., H. T. DULMAGE, D. A. WOLFENBARGER, AND E. MARTINEZ.1981. Persistance of Bacillus thuringiensis Berliner insecticidal activity on cotton foliage. Environ. Ent. 10: 400-1.
JOHNSON, M. W., E. R. OATMAN, AND J. A. WYMAN. 1980. Effects of insecticides on populations of the vegetable leafminer and associatedparasites on summer pole tomatoes. J. Econ. Ent. 73: 61-6.
!{ETZLER, L. D., AND J. F. PRICE. 1982. Methods for growers to evaluateeffects of their cultural practices on Liriomyzu, trifolii leafminers ina simple laboratory. Proc. Florida State Hort. Soc. 95: 162-4.
OATMAN, E. R., AND G. G. KENNEDY. 1976. Methomyl induced outbreak ofLiriomyza sativae on tomato. J. Econ. Ent. 69: 667-8.
SPENCER, K. A. 1973. Agromyzidae (Diptera) of Economic Importance.W. Junk B.V., The Hague. 418 p.
SPENCER, K. A., AND C. E. STEGMAIER, JR. 1973. Arthoropods of Floridaand Neighboring Lands, Vol. 7. Agromyzidae of Florida with a supplement on species from the Caribbean. Florida Dept. Agric. Consumer Serv., Div. Plant Industry, Bureau Ent. Contribution No. In.205 p.
STEGMAIER, C. E., JR. 1966. Host plants and parasites of Liriomyza tri/oliiin Florida (Diptera: Agromyzidae) . Florida Ent. 49: 75-80.
• • • ., . • • • • • • • •METHYL BROMIDE FUMIGATION AS A
QUARANTINE TREATMENT FOR LATANIA SCALE,HEMIBERLESIA LATANIAE
(HOMOPTERA: DIASPIDIDAE)
PETER C. WITHERELLMiami Methods Development Station USDA, APHIS, PPQ
Miami, FL 33158 USA
ABSTRACT
Fumigation tests using methyl bromide at normal atmospheric pressureshowed that 16 glm' for 2% h at 22.8°C or above, or 32 g/m3 for 2% hat18.3 to 22.2°C was sufficient as a quarantine treatment against latania scale,Hemiberlesia lataniae on nursery stock. For use on avocado fruits, recommendations are for 24 g/m3 for 2% h at 22.8°C or above, and 32 glm' for3 h at 18.3 to 22.2°C. Microscopic examination of fumigated and controlscales showed that certain body movements, particularly pharyngeal pulsation, is a much more reliable indicator of scale viability than the merepresence of body fluids. Following fumigation, body fluids required up to31 days to dehydrate completely, under humid conditions.

Witherell: Fumigation for Latania Scale 255
RESUMEN
Pruebas de fumigaci6n con bromura de metilo a una presion atmosfericanormal demostraron que 16 gjm3 por 21;2 horas a 22.8°C 0 mas, 0 32 g/m'Spor 2Y2 horas de 18.3 a 22.2°C fue suficiente como un tratamiento decuarantena contTa la escama Iatania, Hemiberrlesia lalaniae en viverosde plantas. Para nsar en frutas de aguacate, las recomendaciones son 24g{m3 por 2Y, horas a 22.8°C 0 mas, y 32 g{m3 por 3 horas de 18.3 a 22.2°C.Examenes microsc6picos de escamas fumigadas y el control demonstraroDque movimientos del cuerpo, especialmente pulso faringeo, eS un indicadormejor de Ia viabilidad de la escama que la mera presencia de los fhiidoscorporales. La perdida total de fluidos corporales tomo hasta 31 dias posttratamiento en condiciones humedas despues de fumigarse.
Latania scale, Hemiberlesia lataniae (Signoret) is widespread throughout tropical and subtropical areas, and is also found in greenhouses intemperate climates (Dekle 1976, Hamon 1979, Nakahara 1982). The speciesis polyphagous, and is known to infest more than 300 plant species in Florida (Dekle 1976). It is a serious pest of palms and other ornamentals,particularly ponytail palm or elephant-foot tree (Beaucarnea recurvataLem.) , Australian pine (Casuarina ecraiseti/olia) , loquat (Eriobotryajaponica) , and rose (Rosa spp.). Leaves, stems, and fruits may all bedamaged as a result of the insects sucking plant juices.
H. lataniae was best described by Ferris (1938). The armor is usuallyround (female) or oval (male), but may be irregular in shape when several scales overlap or crowd together. The armor is strongly convex and 1.5to 2 mm in diameter when mature. Exuviae are large, pale brown, centrallylocated to sub-centrally, and are surrounded by a dirty white area. Whenthe armor is carefully removed with a pin, the soft, lemon-yellow, flattened,sac-like body is revealed. The body is attached to the host by thread-likestylets that arise near the middle of the body, and extend as long or longerthan the body. Legs are absent. in all secondary stages except 1st instarnymphs (crawlers).
H. lataniae is quarantined in some foreign countries. This often presentsa barrier to the free flow of international commerce. During the recent out~
breaks of Mediterranean fruit fly (Ceratitis capitata (Wied.» in California, avocado fruits originating from outside regulated areas of that statewere shipped to Japan and elsewhere. Shipments received a combined treatment of methyl bromide (MB) plus refrigeration as a precaution (i.e., MB@ 32 g{m3 for 2'h h at 21.1°C or above, followed by 7 days of storage at7.2°C or below).'
Subsequently, inspectors of the Japanese Ministry of Agriculture, Fisheries and Forestry (MAFF), reported live latania scales in 3 out of 5 seacontainers of California avocados, which had received the USDA's precautionary treatment against Mediterranean fruit fly. The Japanese authorities re-treated these fruits with hydrogen cyanide (HON). The California avocado industry then requested a fumigation schedule be developedwhich would allow acceptance of certified fruit. A predeparture treatment
lEquivalent to 32 oz./1000 ft. 3 . This is Treatment Schedule T 102 (a) (1) of the USDAPlant Protection and Quarantine Programs Treatment Manual (Anon. 1981).

256 Florida Entomologist 67 (2) June, 1984
would eliminate the necessity and cost of an additional treatment upon arrival in Japan, and would give the avocados a longer shelf-life.
To determine scale viability Japanese inspectors puncture or crush thescale, and if body fluids are present, the scale is considered to be alive.(This is the so-called "squash test".) Determining scale mortality on commodities receiving quarantine fumigation treatment presents a special problem. Several factors are involved, including time elapse between treatmentand inspection and temperature and RH during in-transit storage. Moreover, the waxy scale covering may prevent large female scales from desiccating, so that when the last scales have dehydrated, the commodity hasexceeded its marketable shelf-life.
A more time-consuming but more accurate method of assessing scaleviability is the so-called "pump test"; an unpublished technique developedat the University of California, Riverside. Naked specimens are examinedmicroscopically for vital signs, viz., slight body movements, particularly thepumping activity of the pharynx. The experiment reported here was designed to clarify the question of treatment efficacy by microscopically examining fumigated scales.
METHODS AND MATERIALS
Attempts to obtain scale-infested avocado fruits from packing houses inFlorida and California were unsuccessful. Moreover, since avocados are nota favored host of this insect (D. Fiskaali, California Dept. of Food & Agric.,pers. comm.), securing sufficient numbers of infested fruits presented aformidable problem. Therefore, a more favored host was obtained, viz.,ponytail palm or elephant-foot tree, Beaucarnea recurvata Lem. (Liliaceae).In addition, unpublished results of MB fumigations of latania scale, by theCalifornia Dept. of Food & Agriculture in 1936 through 1948, were obtained and analysed.
Twenty moderate-to-heavily infested potted ponytail palms (ca. 46 emtall) were purchased locally, a,nd scales were microscopically examined toconfirm that some scales were alive. Plants were stored over night at thefumigation temperatures, in the room equipped with fumigation chambers.The following morning, plants were divided into treatment groups (lor 2plants each). There were approximately equal numbers of scale insects ineach treatment group. Fumigation trials were conducted, using the following test schedules (MB at normal atmospheric pressure) :
(A) 16 g/m3 for 2% h; 22.8-23.9 and 18.3°C temperatures(B) 32 g/m3 for 2% h; 22.8-23.9 and 18.3°C temperatures(C) 48 g/m3 for 2% h; 22.8-23.9 and 18.3°C temperatures(D) 32 g/m3 for 4 h; 24.4-25 and 20°C temperatures2
(E) Combination of MB fumigation plus cold treatment: 32 g/m3 for 2%h; 24.4-25 and 20°C temperatures, followed by 7 days in storage at4.4-7.2°C3
(F) Controls (no treatment)
2This is Treatment Schedule T 105 (a) (l) of the USDA Plant Protection and QuarantinePrograms Treatment Manual (Anon. 1981), except that temperature during treatment must'be 21.1°0 or above. (Recommended for avocados.)
"This is Treatment Schedule T 102 (a) (1) of the USDA Plant Protection and QuarantinePrograms Treatment Manual (Anon. 1981), except that the temperature during treatmentmust be 21.100 or above; during storage, 7.200 or below. (Recommended for avocados.)

Witherell: Fumigation for Latania Scale 257
ME gas concentrations within the fumigation chambers were monitoredduring the course of the treatments by using a Gow-Mac gas analyser.4
Following treatment, plants were isolated from each other and placed ina greenhouse, except for plants receiving Treatment E. The latter wereplaced in a walk-in cold room (4.4-7.2°C, lights on). RH in the two locations ranged from 75 to 90%. Plants were watered (roots only) twiceweekly, immediately following examination of scales.
A representative sample of 100 scales per treatment was microscopicallyexamined twice a week post-treatment over a 31-day period to document therate of desiccation and mortality. All stages were examined, except eggs.
During inspection, one or more leaves were removed from the plant, andexamined under a binoccular, dissecting microscope (15X), using a 30 Wspotlight. Plants receiving Treatment E (fumigation plus refrigeration)were exposed to room temperature before they were inspected. High magnification (45 or 90X) was used to give a more detailed view. The scale cover(armor) was teased off with a pin to reveal the yellow, sac-like body beneath. The large body of mature females often had a cluster of yellow eggsandlor crawlers immediately adjacent to it. If alive, the female would contract, expand, undulate, or twist when g~ntly prodded with a blunt object.If no movement was detected, the insect was teased from the substrate, andturned upsidedown. The triangular-shaped base of the mouthparts was examined for pulsating or pumping action in the pharynx of live specimens.Often, also the threadlike stylet tube moved.
Scales examined were classified into 3 categories:(D) Dead (dry, with no body juices)(L) Live (showing pulsation or other body movements)(M) Moribund (body juices still intact, at least partially, but body show
ing no movement when prodded)The latter category also included scales in various stages of decomposition,as well as those which were parasitized.
RESULTS AND DISCUSSION
Early unpublished work by the California Dept. of Food & Agricultureon ME fumigation for quarantine control of latania scale is summarized inTable 1. All treatments were reported as 100% effective. Steinweden (1948),however, reported poor control with ME at 32 g/m3 at 15.5°C against thisscale on avocados (2-h fumigation).
In tests conducted in Miami, FL, the ponytail plants tolerated the treatments well, except for Treatment D at 24.4-25°C, which resulted in excessive browning of the outer whorls of leaves ("tip burn"). Latania scalesshowed a definite preference for the upper leaf surface of ponytail palm,and congregated in the leafaxils, where the tissue is succulent.
Thoroughness of treatment, as indicated by periodic monitoring of gasconcentration during the course of fumigation, can be expressed in terms ofa "CT product" (mean concentration of ME gas (g/m3 ) X time in hours).The CT products of Treatments A through F were calculated as follows:tests at 22.8-25°C, 38.1, 71.9, 105, 113.2, 81.9, and 0, respectively, and testsat 18.3-20°C, 35, 73.8, 106.9, 126.7, 83.1, and 0, respectively.
4Manufactured by Gow-Mac Instrument Co., Bridgewater, NJ (Model No. 20-350).
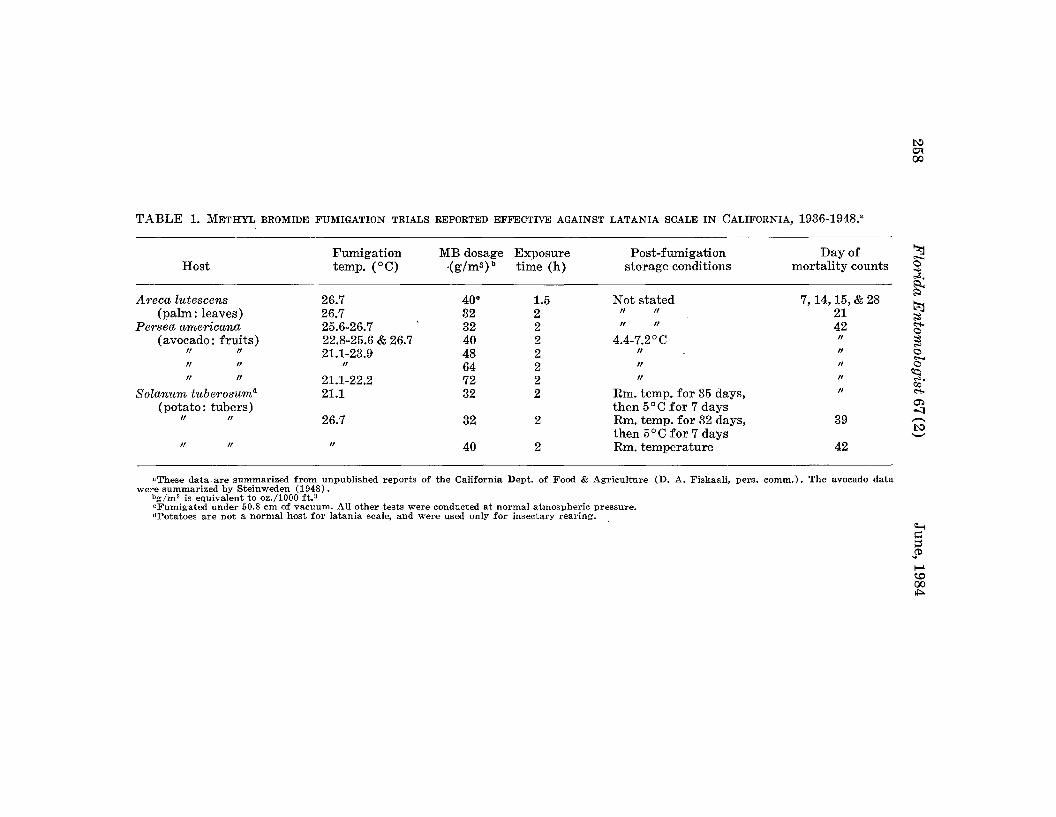
TABLE 1. METHYL BROMIDE FUMIGATION TRIALS REPORTED EFFECTIVE AGAINST LATANIA SCALE IN CALIFORNIA, 1936-1948.a
t-:l0100
Fumigation MB dosage Exposure Post-fumigation Day of ~-Host temp. (OC) -(g/m3 ) b time (h) storage conditions mortality counts c;]
Areca lutescens 26.7 40· 1.5 Not stated 7, 14, 15, & 28~t::j
(palm: leaves) 26.7 32 2 1/ " 21 ;2Persea americana 25.6-26.7 32 2 " " 42 ......c
(avocado: fruits) 22.8-25.6 & 26.7 40 2 4.4-7.2°C " ~" " 21.1-23.9 48 2 1/ 1/ C-1/ 1/ 1/ 64 2 1/ 1/ C
CCl1/ 1/ 21.1-22.2 72 2 " 1/ ""'.~
Solanum tuberosumd 21.1 32 2 Rm. temp. for 35 days, 1/ ......(potato: tubers) then 5°C for 7 days 0':>
-::J
" 1/ 26.7 32 2 Rm. temp. for 32 days, 39 ---then 5°C for 7 days t-:l'-'
1/ " " 40 2 Rm. temperature 42
aThese data are summarized from unpublished reports of the California Dept. of Food & Agriculture (D. A. Fiskaali, pers. comm.). The avocado datawere summarized by Steinweden (1948).
bg/m' is equivalent to oz./1000 ft.'CFumigated under 50.8 em of vacuum. All other tests were conducted at normal atmospheric pressure.dPotatoes are not a normal host for latania scale, and were used only for insectary rearing.
~s:::::s~(t)
~
~00~

Witherell: Fumigation for Latania Scale 259
The efficacy of Treatments A through F is shown in Table 2. At thedosages used, ME fumigation was 100% effective at 22.8-25°C. Good resultseven at the low dosage of 16 g/m3 for 2lh h (Treatment A) confirms thevalidity of earlier data from California (Table 1). At cooler temperatures(18.3-20°C), fumigation was still effective, except in Treatment A (16 g/m3
for 2lh h), where 4-25% (x = 13.1%) survival occurred. Similar scalesurvival was recorded in the controls (Treatment F), i.e., 1-26% (x =12.4% survival).
Eody decomposition and moisture loss occurred over an extended periodof time under the humid conditions prevalent in Florida, and at the highRH at which fresh fruits and vegetables are best stored. At 31 days posttreatment a few insects retained some body moisture. Among the controlsexamination revealed 32 to 99% (x = 73.5%) of the scales were dead (D);1 to 62% (x = 22.4%) were alive (L); and 0 to 12% (x = 3.5%) weremoribund (M). For all practical purposes scales classified as "moribund"were dead. There was no "delayed-action mortality," when lack of movement rather than lack of moisture was used as the criterion of death. Afew fumigated scales recorded as "moribund" contained a wasp larva, whichappeared to be dead.
As the scales decomposed and lost moisture, their bodies changed froma bright lemon-yellow color, bloated and flexible, to an amber, moist, tostraw colored and dehydrated. Immature scales, especially, desiccatedquickly, became dry, brittle, and dark brown in color. Fully mature femalescales dehydrated more slowly than others. Occasionally the armor of deadscales sloughed off prior to inspection. Also, some of the dead scales examined contained an opening made by thrips or emerging wasps. In controls, live thrips were occasionally found within hollowed-out scales.Whether these thrips were predators or merely scavengers is unknown.
Color and consistency provide good evidence of mortality, but should notbe the only consideration when examining freshly fumigated scales. Theprogression of color changes was somewhat slower among scales which hadreceived Treatment E. This is probably a result of the preservative effectof refrigeration. Inasmuch as these scales retained their lemon-yellow colorlonger, warming them and checking for slight movement was deemedespecially important for this group. No movement, however, was ever observed.
CONCLUSIONS AND RECOMMENDATIONS
The results of this study lead to the following conclusions and recommendations:
1.) The "squash test" for determining viability of armored scales in thefield is not an adequate test for fumigated specimens. An accurate assessment of treatment efficacy can be made only by microscopic examination todetect slight body movements and pharyngeal pulsations.
(2) During fumigation, the dosage, length of exposure, and temperatureof the commodity are factors critical to the success of treatment. Higherdosages of a fumigant are required at lower temperatures, in order to effecta complete kill, because of reduced metabolism in the insects (Monro 1969,Anon. 1981). Data from California (D. Fiskaali, unpubl, Steinweden 1948)illustrated that ME at a dosage of 32 g/m3 for 2 h was completely effective
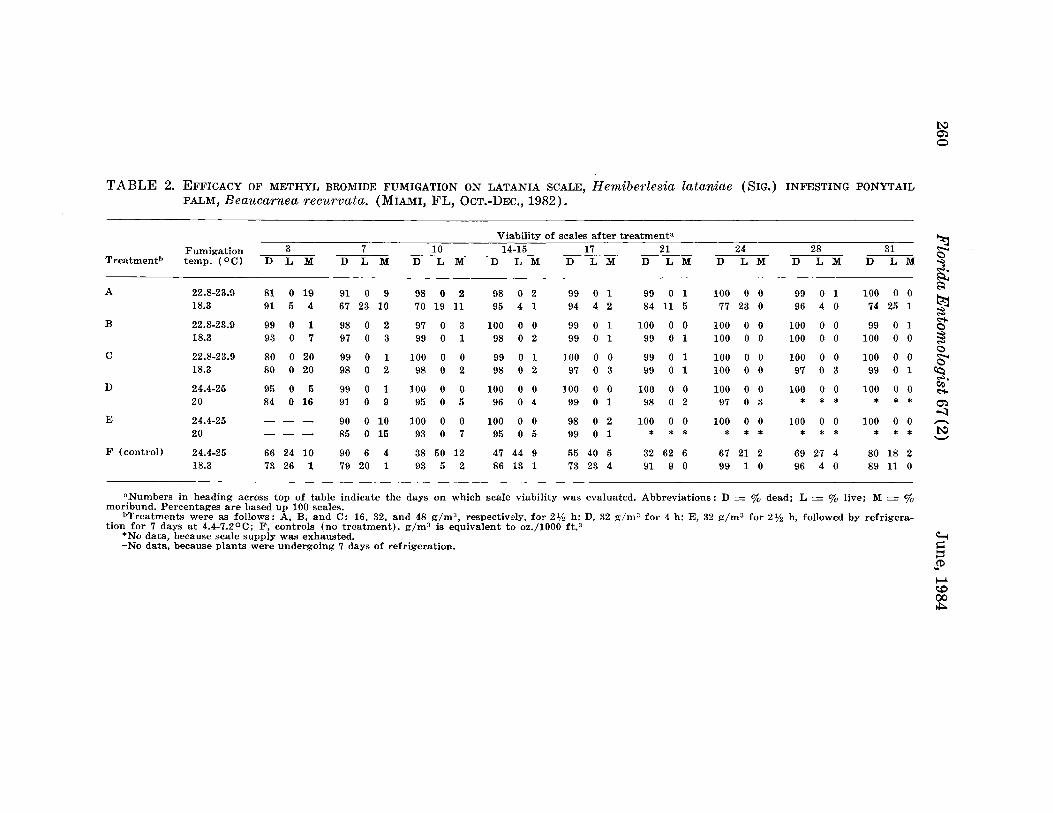
t\JO':l0
TABLE 2. EFFICACY OF METHYL BROMIDE FUMIGATION ON LATANIA SCALE, Hemibe1'Zesia Zataniae (SIG.) INFESTING PONYTAIL
PALM, Beaucarnea recurvata. (MIAMI, FL, OCT.-DEC., 1982).
Viability of scales after treatmenta ':l:jFumigation 3 7 10 14-15 17 21 24 28 31 ......
CTreatmentb temp. (OC) D L M D L M D L M D L M D L M D L M D L M D L M D L M ;;]~
A 22.8-23.9 81 o 19 91 0 9 98 0 2 98 o 2 99 o 1 99 o 1 100 o 0 99 o 1 100 o 0~
18.3 91 5 4 67 23 10 70 19 11 95 4 1 94 4 2 84 11 5 77 23 0 96 4 0 74 25 1 ~B 22.8-23.9 99 0 1 98 0 2 97 0 3 100 0 0 99 0 1 100 0 0 100 0 0 100 o 0 99 0 1 <:"i-
C18.3 93 0 7 97 0 3 99 0 1 98 o 2 99 o 1 99 o 1 100 0 0 100 o 0 100 0 0 ~
CC
22.8-23.9 80 o 20 99 0 1 100 0 0 99 0 1 100 0 0 99 0 1 100 0 0 100 0 0 100 0 0 C'18.3 80 o 20 98 0 2 98 0 2 98 o 2 97 o 3 99 o 1 100 o 0 97 0 3 99 0 1 ~.,.,.
D 24.4-25 95 0 5 99 0 1 100 0 0 100 o 0 100 0 0 100 0 0 100 o 0 100 o 0 100 o 0 ~<:"i-
20 84 o 16 91 0 9 95 0 5 96 o 4 99 0 1 98 o 2 97 o 3 * * * * * * O':l
E-:J
24.4-25 - -- 90 o 10 100 0 0 100 0 0 98 o 2 100 0 0 100 o 0 100 0 0 100 0 0 ---20 - -- 85 0 15 93 0 7 95 0 5 99 0 1 * * * * * * * * * * * * t\J--F (control) 24.4-25 66 24 10 90 6 4 38 50 12 47 44 9 55 40 5 32 62 6 67 21 2 69 27 4 80 18 218.3 73 26 1 79 20 1 93 5 2 86 13 1 73 23 4 91 9 0 99 1 0 96 4 0 89 11 0
aNumbers in heading across top of table indicate the days on which scale viability was evaluated. Abbreviations: D = % dead; L = % live; M = %moribund. Percentages are based up 100 scales.
"Treatments were as follows: A. B, and C: 16, 32, and 48 g/m" respectivdy, for 2% h; D. 32 g/m" for 4 h; E, 32 g/m" for 2 1h h, followed by refrigera-tion for 7 days at 4.4-7.2 o C; F, controls (no treatment). g/m" is equivalent to oz./1000 ft."
<:..j*No data, because scale supply was exhausted.-No data, because plants were undergoing 7 days of refrigeration. ~
::l,!i>~
e.o00Il::o-

Witherell: Fumigation for Latania Scale 261
at 21.1°C, but killed only 28.3% of the latania scales at 15.6°C. Based onthe results of the present study, the following are the minimum schedulesthat can be recommended as effective against latania scale on ornamentals(MB at normal atmospheric pressure) :
16 g/m3 for 2lh h; 22.8°C or above.32 g/m3 for 2lh h; 18.3 to 22.2°C.
Air circulation should be provided so that temperatures throughout thechamber load are relatively uniform, and never below 18.3°C.
(3) If the scale-infested commodity being fumigated is avocado, oneshould bear in mind that these fruits, like many others, tend to absorb MBgas (P. C. Witherell, unpub. data). Gas loss is even greater if the fruitsare packed in corrugated fiberboard cartons or under tarpaulin. Accordingly,fumigation schedules of slightly higher dosage or longer exposure time thanaforementioned would be advisable. The following schedules, therefore, arerecommended for use against latania scales on avocado (MB at normalatmospheric pressure) :
24 g/m3 for 2lh h; 22.8°C or above.32 g/m3 for 3 hj 18.3 to 22.2°C.
Fruit load in the chamber or under tarpaulin should be 70% or less, calculated by the height of the chamber or enclosure. Treatments D and E, at21.1°C or above-both of which are prescribed for use on avocados againsteggs and larvae of fruit flies (Anon. 1981)-are also 100% effective againstlatania scale. These schedules, however, may cause some phytotoxic damageto the fruit. Tolerance of avocado to fumigation depends, to some extent,on the variety of fruit. Varieties exhibit a wide range of tolerance to MB(Witherell et al. 1982). Normal fruit rots, particularly anthracnose, develop faster after MB fumigation, since the ripening process is acceleratedby 2 to 4 days. Vigorous brushing of fruits to remove scales is also possible,but probably impractical for large shipments.
ACKNOWLEDGMENTS
I wish to thank Mr. Kenneth D. Havel (USDA, APHIS, PPQ, Hoboken,NJ), Miss Alicia Arner (USDA, ARS, Miami, FL), and Mr. Don A. Fiskaali(California Dept. of Food & Agric.) for their help with this manuscript. Ialso thank Mr. David Lowe (formerly with Florida DACS, DPI) for providing a source of infested material.
POST SCRIPT: In a recent meeting between USDA and Japanese MAFFofficials in Tokyo, the data in this paper were presented. Agreement wasreached that Japanese inspectors will use microscopic examination to determine scale insect mortality in fumigated commodities.
REFERENCES CITED
ANONYMOUS. 1981. Plant Protection and Quarantine Programs TreatmentManual (as revised). Sect. VI, Series T-lOO, pp. 1-29. USDA, APHIS,PPQ, Hyattsville, MD.
DEKLE, G. W. 1976. Page 71 In: Florida Armored Scale Insects. Vol. 3of: Arthropods of Florida and Neighboring Land Areas. FloridaDACS, DPI, Gainesville, FL.

262 Florida Entomologist 67 (2) June, 1984
FERRIS, F. G. 1938. Atlas of Acale Insects of North America. Vol. 2, Ser.Nos. 137-268. California: Stanford Univ. Press.
HAMON, A. B. 1979. Latania scale, Hemiberlesia lataniae (Signoret)(Homoptera: Coccoidea: Diaspididae. Ent. Circ. No. 208. FloridaDACS, DPI, Gainesville, FL, 2 p.
MONRO, H. A. U. 1969. Manual of Fumigation for Insect Control. F AOAgric. Stud., No. 79. Rome: FAO, UN, 381 p.
NAKAHARA, S. 1982. Checklist of the Armored Scales (Homoptera:Diaspididae) of the Conterminous United States. USDA, APHIS,PPQ, Beltsville,tMD, 110 p. (p. 42).
STEINWEDEN, J. L. 1948. Commodity treatments. Pages 236-41 In: 29thAnnu. Rep., period ending Dec. 31, 1948. California Dep. Agric. Bull.37.
WITHERELL, P. C., D. H. SPALDING, AND C. A. BENSCHOTER. 1982. Toleranceof Florida avocado cultivars to methyl bromide fumigation treatmentseffective against fruit flies. Proc. Florida State Hort. Soc. 95: 227-9.
• • • • • • • • • • • • •RESPONSE OF TRICHOGRAMMA PRETIOSUM ANDT. EVANESCENSl TO WHITELIGHT, BLACKLIGHT
OR NO-LIGHT SUCTION TRAPS
J. H. BROWER AND L. D. CLINEStored-Product Insects Research and Development Laboratory
Agricultural Research Service, USDASavannah, GA 31403 USA
ABSTRACT
The response of Trichogramma pretiosum Riley and T. evanescensWestwood to blacklight, whitelight, or no-light suction traps in empty 44.7m3 rooms showed that significantly more (P<O.Ol) of both species responded to blacklight than to whitelight or no-light traps. About 40% and60% of the T. evanescens and T. pretiosum, respectively, were caught in theblacklight trap. No significant difference (P>0.05) was found in the response of T. pretiosum to whitelight and no-light traps, while significantlymore (P<0.05) T. evanescens were caught in the whitelight trap than in theno-light trap. Females, especially those of T. evanescens, appeared to bemore responsive than males as more females than males of both specieswere usually caught. The implications of these findings should be consideredin planning releases of Trichogramma for control of storage moths inwarehouses.
RESUMEN
EI comportamiento de Trichograma pretiosum Riley and T. evanescensWestwood hacia trampas de succion de luz negra, luz blanca, 0 trampas sinluz en cuartos vacios de 44.7 m3 , indico que significativamente (P < 0.01)ambas especies respondieron mas a las trampas de luces negras que a lastrampas de luces blancas 0 a las de sin luces. Alrededor de 40% y 60% deT. evanescens y T. pretiosum respectivamente fueron atrapadas en latrampa de luz negra. No se encontro ninguna diferencia significativa (P >
1 Hymenoptera, Trichogrammatidae

262 Florida Entomologist 67 (2) June, 1984
FERRIS, F. G. 1938. Atlas of Acale Insects of North America. Vol. 2, Ser.Nos. 137-268. California: Stanford Univ. Press.
HAMON, A. B. 1979. Latania scale, Hemiberlesia lataniae (Signoret)(Homoptera: Coccoidea: Diaspididae. Ent. Circ. No. 208. FloridaDACS, DPI, Gainesville, FL, 2 p.
MONRO, H. A. U. 1969. Manual of Fumigation for Insect Control. F AOAgric. Stud., No. 79. Rome: FAO, UN, 381 p.
NAKAHARA, S. 1982. Checklist of the Armored Scales (Homoptera:Diaspididae) of the Conterminous United States. USDA, APHIS,PPQ, Beltsville,tMD, 110 p. (p. 42).
STEINWEDEN, J. L. 1948. Commodity treatments. Pages 236-41 In: 29thAnnu. Rep., period ending Dec. 31, 1948. California Dep. Agric. Bull.37.
WITHERELL, P. C., D. H. SPALDING, AND C. A. BENSCHOTER. 1982. Toleranceof Florida avocado cultivars to methyl bromide fumigation treatmentseffective against fruit flies. Proc. Florida State Hort. Soc. 95: 227-9.
• • • • • • • • • • • • •RESPONSE OF TRICHOGRAMMA PRETIOSUM ANDT. EVANESCENSl TO WHITELIGHT, BLACKLIGHT
OR NO-LIGHT SUCTION TRAPS
J. H. BROWER AND L. D. CLINEStored-Product Insects Research and Development Laboratory
Agricultural Research Service, USDASavannah, GA 31403 USA
ABSTRACT
The response of Trichogramma pretiosum Riley and T. evanescensWestwood to blacklight, whitelight, or no-light suction traps in empty 44.7m3 rooms showed that significantly more (P<O.Ol) of both species responded to blacklight than to whitelight or no-light traps. About 40% and60% of the T. evanescens and T. pretiosum, respectively, were caught in theblacklight trap. No significant difference (P>0.05) was found in the response of T. pretiosum to whitelight and no-light traps, while significantlymore (P<0.05) T. evanescens were caught in the whitelight trap than in theno-light trap. Females, especially those of T. evanescens, appeared to bemore responsive than males as more females than males of both specieswere usually caught. The implications of these findings should be consideredin planning releases of Trichogramma for control of storage moths inwarehouses.
RESUMEN
EI comportamiento de Trichograma pretiosum Riley and T. evanescensWestwood hacia trampas de succion de luz negra, luz blanca, 0 trampas sinluz en cuartos vacios de 44.7 m3 , indico que significativamente (P < 0.01)ambas especies respondieron mas a las trampas de luces negras que a lastrampas de luces blancas 0 a las de sin luces. Alrededor de 40% y 60% deT. evanescens y T. pretiosum respectivamente fueron atrapadas en latrampa de luz negra. No se encontro ninguna diferencia significativa (P >
1 Hymenoptera, Trichogrammatidae

Brower & Cline: Light Trapping Trichogramma 263
0.05) en el comportamiento de T. pretiosum hacia trampas con luz blanca 0
sin luz, mientras que significativamente mas (P < 0.05) T. evanescensfueron atrapadas en trampas de luz blanca que en trampas sin luces.Hembras, especialmente aquellas de T. evanescens, parecen responder masque los machos, mientras que mas hembras que machos de ambas especiesfueron usualmente atrapadas. Las implicaciones de este descubrimientodebe de considerarse cuando se planee soltar Trichograma para controlarpolillas en almacenes.
Traditionally pests of stored-food commodities have been controlled bychemical means, but recently the potential of biological control methods hasreceived more attention. Some studies have suggested that various speciesof parasitic Hymenoptera might be useful as control agents for storedproduct Lepidoptera (Corbet and Rotheram 1965, Reinert and King 1971,Takahashi 1973, Press et al. 1977), and it is well known that eggs ofstored-product moths can serve as laboratory hosts for the rearing ofTrichogramma (Alden and Farlinger 1931, Morrison and King 1977). Infact, eggs of the Angoumois grain moth, Sitotroga cerealella (Olivier) 2, areusually used to produce the large numbers of Trichogramma needed forreleases to control field crop pests (Morrison et al. 1976). Thus, Brower(1983) suggested that large numbers of Trichogramma might be releasedperiodically into warehouses to reduce or eliminate populations of storedproduct moth pests. However, the effectiveness of parasites in indoor situations has not been studied extensively and many questions need to beanswered regarding warehouse releases.
Many parasites and predators seem to be lured to and can often be collected at warehouse windows and around lights (Ghani and Sweetman1955), and Trichogramma spp. are known to be positively phototactic(Morrison et al. 1976, Martin 1969). To determine the strength of this attractancy to light we tested the response of 2 species of Trichogramma towhitelight, blacklight, and no-light suction traps.
MATERIAL AND METHODS
EXPOSURE ROOMS
Tests were conducted in two adjacent rooms that were each ca. 44.7 m 3 •
Several modifications had to be made to the rooms to make them tightenough to confine the small test insects: (1) clear plexiglass shields werebuilt around the light fixtures and electrical outlets, (2) 295 p.-mesh Nitex®screens were placed over the heating and return air ducts, (3) special doorswere constructed that could be closed tightly, (4) an anteroom connectingthe rooms was built to reduce the chances of any accidental introduction ofunwanted insects, and (5) all cracks and crevices were caulked and recaulked as needed between replicates.
The test conditions of the rooms were kept as constant as was practical.The minimum temperature was 27* °C; the maximum temperature wasnot controlled, but it never exceeded 31 % 0 C. The fluorescent ceiling lightsautomatically turned on at 0700 h and off at 1900 h which coincided with the
"Lepidoptera. Gelechiidae

264 Florida Entomologist 67 (2) June, 1984
light-dark cycle at which the Trichogramma were reared. The relativehumidity in the rooms fluctuated between 40-60%.
LIGHT TRAPS
The New Jersey mosquito light traps3 used in these tests were identicalexcept for their light sources. The whitelight (WL) trap had a 25-wattclear incandescent bulb; the blacklight (BL) trap had a 32-watt, ca. 0.3 mcircline fluorescent blacklight lamp (GE FC12TI0-BL Rapid Start). Thestandard wire screen funnel was lined with 295 I'-mesh Nitex® screen andthe collection jars contained a small amount of deobase to retain and killthe trapped insects. Both traps were modified so the light source could beturned off with the fan left running, thus creating a no-light, suction trap.
One light trap was suspended from the ceiling of each room in thecorner furthest from the door, so that the center of the light source was ca.0.5 m away from the walls and the ceiling and ca. 2.1 m from the floor. Thetraps ran continuously during the tests and the collection jars were changeddaily.
TEST INSECTS
Two species of Trichogramma were used in this test, T. pretiosum Rileyand T. evanescens Westwood. T. pretio8um cultures were obtained from theCotton Insects Research Laboratory, College Station, TX; and they originated from a field collection in Arkansas. T. evanescens cultures were obtained from the Southern Grain Insects Research Laboratory, Tifton, GA;and they had originated in Poland. Both species Were reared for manygenerations at the Savannah Laboratory on the eggs of the almond moth,Cadra cautella (Walker)', in a controlled environment at 27±214 °C and60±5% RH with a 1~L:12D light cycle. Large numbers of almond motheggs were collected by placing 0- to 2-day-old adults in inverted 8.8-liter jarswith screen bottoms. Eggs were removed after 24 h, sifted to remove debris,and placed in open plastic petri dishes for exposure to adult Trichogramma.The 0- to 24-h-old almond moth eggs were exposed by placing the open petridish in a closed 8.8-liter jar along with an open petri dish containing largenumbers of parasitized almond moth eggs that contained Trichogramma 0
to 24-h before emergence. The almond moth eggs were exposed to Trichogramma adults for 8 h and then removed from the exposure chambers.
One day before adult Trichogramma emergence, parasitized almondmoth eggs were counted into groups of 200, and each group was glued to alabeled 9 cm filter paper disk and placed in a petri dish. Dishes containingeach species were then placed in the center of either the WL or BL room,or were reserved as controls. The parasitized eggs were placed in the roomsat ca. 1400 h. Emergence of control insects held at the aforementioned controlled conditions usually started by 0800 h the following morning.
Trap catches were removed each morning for 6 days and the specimenswere separated by species, sexed, counted; and recorded. The 2 species were
3Manufaetured by Hausherr's Machine Work, R. D. 1, Old Freehold !Wad, Toms River,NJ 08753.
~ Lepidoptera, Pyralidae.
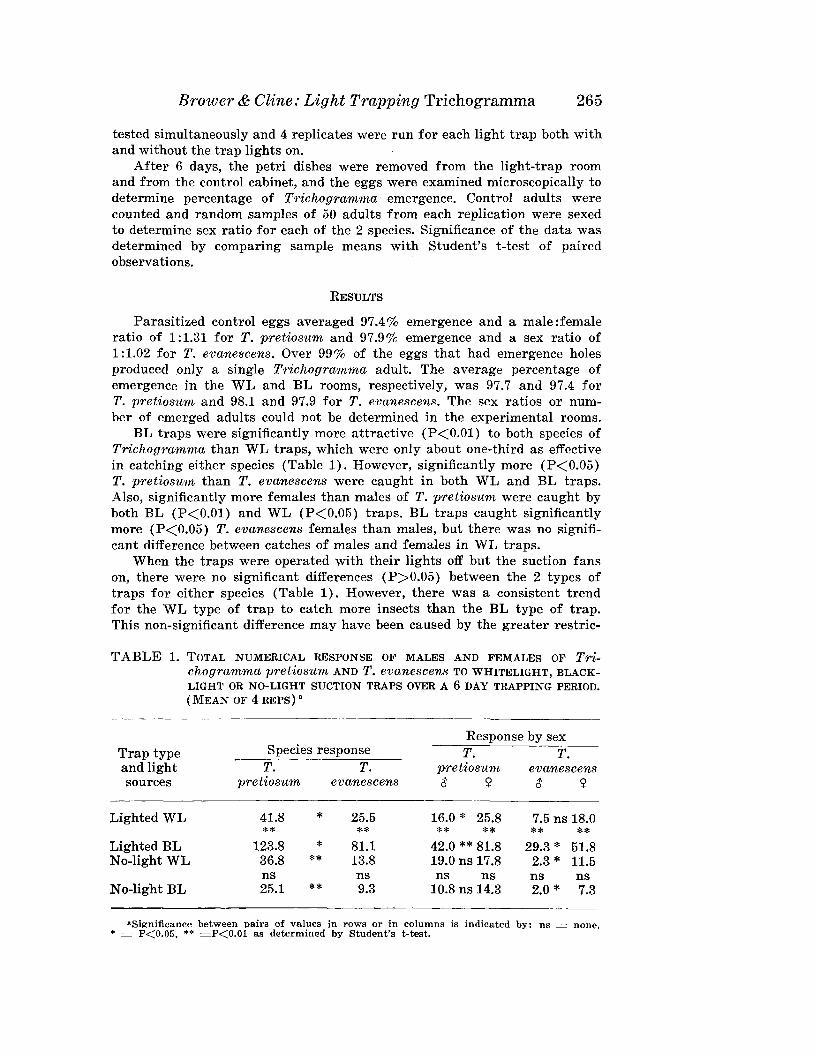
Brower & Cline: Light Trapping Trichogramma 265
tested simultaneously and 4 replicates were run for each light trap both withand without the trap lights on.
After 6 days, the petri dishes were removed from the light-trap roomand from the control cabinet, and the eggs were examined microscopically todetermine percentage of Trichogramma emergence. Control adults werecounted and random samples of 50 adults from each replication were sexedto determine sex ratio for each of the 2 species. Significance of the data wasdetermined by comparing sample means with Student's t-test of pairedobservations.
RESULTS
Parasitized control eggs averaged 97.4% emergence and a male :femaleratio of 1:1.31 for T. pretiosum and 97.9% emergence and a sex ratio of1 :1.02 for T. evanescens. Over 99% of the eggs that had emergence holesproduced only a single Trichogramma adult. The average percentage ofemergence in the WL and BL rooms, respectively, was 97.7 and 97.4 forT. pretiosum and 98.1 and 97.9 for T. evanescens. The sex ratios or number of emerged adults could not be determined in the experimental rooms.
BL traps were significantly more attractive (P<O.Ol) to both species ofTrichogramma than WL traps, which were only about one-third as effectivein catching either species (Table 1). However, significantly more (P<0.05)T. pretiosum than T. evanescens were caught in both WL and BL traps.Also, significantly more females than males of T. pretiosum were caught byboth BL (P<O.Ol) and WL (P<0.05) traps. BL traps caught significantlymore (P<0.05) T. evanescens females than males, but there was no significant difference between catches of males and females in WL traps.
When the traps were operated with their lights off but the suction fanson, there were no significant differences (P>0.05) between the 2 types oftraps for either species (Table 1). However, there was a consistent trendfor the WL type of trap to catch more insects than the BL type of trap.This non-significant difference may have been caused by the greater restric-
TABLE 1. TOTAL NUMERICAL RESPONSE OF MALES AND FEMALES OF Trichogramma pretiosum AND T. evanescens TO WHITELIGHT, BLACKLIGHT OR NO-LIGHT SUCTION TRAPS OVER A 6 DAY TRAPPING PERIOD.(MEAN OF 4 REPS)'
Response by sexTrap type Species response T. T.and light --'1'. T. pretiosum evanescenssources pretiosum evanescens (; <;> (; <;>
Lighted WL 41.8 * 25.5 16.0 * 25.8 7.5 ns 18.0** ** ** ** ** **
Lighted BL 123.8 * 81.1 42.0 ** 81.8 29.3 * 51.8No-light WL 36.8 ** 13.8 19.0 ns 17.8 2.3 * 11.5
ns ns ns ns ns nsNo-light BL 25.1 ** 9.3 10.8 ns 14.3 2.0 * 7.3
'Significance between pairs of values in rows or in columns is indicated by: ns = none.* = P<O.05, ** =P<O.Ol as determined by Student's t-test.

266 Florida Entomologist 67 (2) June, 1984
tion of airflow by the large, circular BL bulb. This pattern of greater catchby the WL type trap was opposite from the pattern found when the traplights were on. Significantly more (P<O.Ol) T. pretiosum than T. evanescenswere caught in both traps. Males and females of T. pretiosum were caughtwith equal frequency in the no-light traps, but significantly more (P<0.05)females than males of T. evanescens were caught in both no-light traps.
The temporal pattern of Trichograma catch was probably influenced byemergence pattern, flight propensity, trap type, and species. For T.pretiosum most catch occurred on the 2nd day (Table 2) in lighted trapswith very little catch on the 3rd and 4th days and none on the 5th or 6thdays. The catch was somewhat delayed in the no-light traps and fewerinsects were trapped. T. evanescens responded most strongly to the WLtrap on the 1st day and catch decreased thereafter (Table 2). Response tothe BL trap and to no-light traps was very similar in this species with thegreatest catch occurring on the 2nd day.
DISCUSSION
General observations of Trichogramma spp. usually show a movementtoward or aggregation near light sources (Martin 1969, Morrison et aI.1976). However, to our knowledge no controlled studies of this phenomenonhave been published until now. This study showed that T. pretiosum andT. evanescens moved several meters to a BL source, but that an incandescentWL source was only minimally attractive. More than 41 % and 63% of theemerged T. evanescens and T. pretiosum adults, respectively, were caughtby the BL traps, but less than 22% of either species was caught by the WLtraps. This observation supports the supposition of Costas (1941) that WLonly increases the activity and dispersion of Trichogramma, but not oriented movement. Previous observations of positive phototactic behavior ofTrichogramma may have resulted from increased movement or attractionto the UV portion of most WL sources. Females of T. pretiosum werecaught significantly more often than males by both BL and WL traps, butthe interpretation of this finding must be tempered by the fact that the sexratio of this species was skewed in favor of females. Equal numbers offemales and males of T. pretiosum were caught in the no-light traps. Females of T. evanescens were caught significantly more often than males in
TABLE 2. PERCENTAGES OF Trichogramma pretiosum AND T. evanescensADULTS CAUGHT IN WHITE LIGHT, BLACK LIGHT AND NO-LIGHTSUCTION TRAPS DURING 5 DAYS OF OPERATION.
T. pretiosum T. evanescensLighted Lighted Combined Lighted Lighted Combined
Day WL BL no light WL BL no light
1 28.7 22.2 16.2 51.0 34.3 38.32 62.3 67.7 48.6 23.2 55.5 51.43 8.4 8.5 32.0 9.8 8.3 8.74 0.6 1.6 3.2 1.0 1.9 1.65 0 0 ° 0 0 °

Brower & Cline: Light Trapping Trichogramma 267
the no-light traps. Females are apparently more active fliers than males ofthis species since the sex ratio was near unity in the controls. Females ofother species of parasitic wasps have also been shown to be very attractedto BL sources (Hagstrum and Sharp 1975, Cline et al. 1983).
A comparison of the response of T. pretiosum with that of T. evanescensshowed some species specific differences. A significant difference (P<0.05),was found between the number of T. evanescens caught by the WL trapwith the light on and the number caught with the light off. In contrast,there was no significant difference (P>0.05) between the numbers of T.pretiosum caught by the WL trap with the light on or off. Of course, because of the strong response to BL, both species were caught significantlymore often by the BL trap than by the no-light trap.
The release of large numbers of insectary-reared Trichogramma intocommodity storages for control of moth populations has been proposed(Brower 1983). Recently, 2 species of Trichogramma have been foundnaturally parasitizing almond moth eggs in peanut storages in Georgia(Brower, in press). In order to maximize the chances of success, the findings reported here indicate that BL traps should not be used concurrentlywith Trichogramma releases. If monitoring of moth populations is essential then pheromone traps or perhaps WL traps could be used. It mightalso be desirable to eliminate or reduce UV sources such as open windows,doors, skylights, etc. to reduce Trichogramma loss to the outside. However,the warehouse should probably not be kept in total darkness because Trichogramma may not find or parasitize their hosts in total darkness (Costas1941, Orphanides and Gonzalez 1970, Ashley et al. 1973). If light traps areto be used to sample for the presence of or to monitor populations of Trichogramma, then BL traps should be selected rather than WL traps because of the greater response to BL.
ACKNOWLEDGEMENTS
The authors are grateful for the help of R. K. Morrison, Cotton InsectsResearch Laboratory, USDA, ARS, P. O. Box DG, College Station, TX77841, who kindly supplied cultures of T. pretiosum and to D. A. Nordlund,Southern Grain Insect Research Laboratory, USDA, ARS, Georgia CoastalPlains Experiment Station, Tifton, GA 31793, who supplied the cultures ofT. evanescens.
Mention of a commercial or proprietary product does not constitute anendorsement by the USDA. Revised for publication 15 February 1984.
REFERENCES CITED
ALDEN, C. H., AND D. F. FARLINGER. 1931. The artificial rearing andcolonization of Trichogramma minutum. J. Econ. Ent. 24: 480-83.
ASHLEY, T. R., D. GONZALEZ, AND T. F. LEIGH. 1973. Reduction in effectiveness of laboratory-reared Trichogramma. Environ. Ent. 2: 1069-73.
BROWER, J. H. 1983. Utilization of stored-product Lepidoptera eggs as hostsby Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). J. Kansas Ent. Soc. 56: 50-4.
BROWER, J. H. 1984. The natural occurrence of the egg parasite, T1'ichogramma, on almond moth eggs in peanut storages in Georgia. J.Georgia Ent. Soc. (In Press).

268 Florida Entomologist 67 (2) June, 1984
CLINE, L. D., B. R. FLAHERTY, AND J. W. PRESS. 1983. Response of selectedparasitoids and predators of stored-product insects to whitelight orblacklight traps. J. Econ. Ent. 76: 2"98-301.
CORBET, S. A., AND S. ROTHERAM. 1965. The life history of the ichneumonidNemeritis (Devorgilla) canescens (Gravenhorst) as a parasite of theMediterranean flour moth, Ephestia (Anagasta) kuehniella Zeller,under laboratory conditions. Proc. R. Ent., Soc. London 40: 67-72.
COSTAS, L. A. 1941. The effect of varying conditions on oviposition by Trichogramma on eggs of Angoumois grain moths. J. Econ. Ent. 34:57-8.
GHANI, M. A., AND H. L. SWEETMAN. 1955. Ecological studies on thegranary weevil parasite, Aplastomorpha calandrae (Howard).Biologia 1: 118-39.
HAGSTRUM, D. W., AND J. E. SHARP. 1975. Population studies on Cadracautella in a citrus pulp warehouse with particular reference todiapause. J. Econ. Ent. 68: 11-4.
MARTIN, F. J. 1969. Searching success of predators in artificial leaf litter.American Midland Natur. 81: 218-27.
MORRISON, R. K., AND E. G. KING. 1977. Mass production of naturalenemies. Pages 183-217. In: Ridgway, R. L., and S. B. Vinson (eds.).Biological control by augmentation of natural enemies. Plenum Press,New York. 480 p.
---, R. E. STINNER, AND R. L. RIDGWAY. 1976. Mass production of Trichogramma pretiosum on eggs of the Angoumois grain moth. Southwestern Ent. 1: 74-80.
ORPHANIDES, F. M., AND D. GONZALEZ. 1970. Importance of light in the biology of Trichogramma pretiosum. Ann. Ent. Soc. America 63: 1734-40.
PRESS, J. W., B. R. FLAHERTY, AND R. T. ARBOGAST. 1977. Interactionsamong N emeritis canescens (Hymenoptera: Ichneumonidae), Braconhebetor (Hymenoptera: Braconidae), and Ephestia cautella (Lepidoptera: Pyralidae). J. Kansas Ent. Soc. 50: 259-62.
REINERT, J. A., AND E. W. KING. 1971. Action of Bracon hebetor Say as aparasite of Plodia interpunctella at controlled densities. Ann. Ent.Soc. America 64:' 1335-40.
TAKAHASHI, F. 1973. An experimental study on the suppression and regulation of the population of Cadra cautella (Walker) (Lepidoptera;Pyralidae) by the action of a parasitic wasp, Nemeritis canescensGravenhorst (Hymenoptera; Ichneumonidae). Mem. CoIl. Agric.,Kyoto Univ. 104: 1-12.

Elvin & Sloderbeck: Key to Nabid Nymphs
A KEY TO NYMPHS OF SELECTED SPECIESOF NABIDAE (HEMIPTERA) IN
THE SOUTHEASTERN USA
M. K. ELVINI AND P. E. SLODERBECK2
Department of Entomology and NematologyUniversity of Florida
Gainesville, FL 32611 USA
269
ABSTRACT
A key is presented for distinguishing nymphs of the 4 most commonlyrecorded species of nabids (Hemiptera: Nabidae) found in row crops in thesoutheastern United States: Nabis roseipennis Reuter, Nabis americoferusCarayon, Tropiconabis capsiformis (Germar), and Hoplistoscelis deceptivus(Harris). Characters used to separate the species include body shape andpatterns of pigmentation on the head, antennae, thorax, and legs.
RESUMEN
Una clave es presentada para distinguir las ninfas de las 4 especiesregistradas mas comunes de nabidos (Hemiptera: Nabidae) encontrada encultivos de surcos en el sur-este de los Estados Unidos: Nabis roseipennisReuter, Nabis americoferus Carayon, Tropiconabis capsiformis (Germar),and Hoplistoscelis deceptivus (Harris). Los caracteres usados para separarlas especies incluyen la configuraci6n del cuerpo y la forma de la pigmentaci6n en la cabeza, antena, t6rax, y las patas.
Nabids (Hemiptera: Nabidae) belonging to the tribe Nabini are someof the most common predators found in row crops in the southeasternUnited States (Dinkins et al. 1970, Shepard et al. 1974, Deitz et al. 1976,and Pitre et al. 1978). The 4 most commonly recorded species are Nabisroseipennis Reuter, N. americoferus Carayon, Tropiconabis capsiformis(Germar), and Hoplistoscelis deceptivus (Harris) (Dinkins et al. 1970,Neal 1974, Deitz et al. 1976, and Pitre et al. 1978). The adults of these 4species of nabids may be identified by using keys found in a monograph onNorth American nabids (Harris 1928) or one of several keys to selectedspecies (Benedict and Cothran 1975, Deitz et al. 1976, Hormchan et al.1976, and Irwin and Shepard 1980). Unfortunately, there are no keysavailable for distinguishing to species the nymphs of these common predators. To amend this situation, we present a key to the nymphs of the 4 mostcommon species of nabids in row crops in the southeastern United States.
FAMILY: NABIDAE
Immature nabids have a long, slender, 4 segmented labium arising fromthe anterior part of the head. The labium is not located in a rostral groovein the gular region. They also have long, slender antennae, apical tarsal
1 Present address: Dept. of Entomology, University of Arkansas, Fayetteville, AR 72701USA.
'Southwest Area Extension Office, Kansas State University, Garden City, KS 67846 USA.

270 Florida Entomologist 67 (2) June, 1984
claws, and usually 3-4 pair of scent glands present on the abdomen (DeCoursey 1971).
SEPARATION OF NYMPHAL INSTARS
The same general characteristics may be used to distinguish to speciesindividuals in each nymphal instar. However, for some nymphal instars,there are unique characteristics that are used to separate the 4 species.Such characteristics are noted in the key where appropriate.
The nymphal instar of a particular immature nabid can be determinedby assessing the amount of wing pad development. Individuals in the 1stnymphal instar show no wing pad development (Fig. la, 2a, 3a, and 4a).By the 2nd nymphal instar, the fore wing pads appear as slightly rounded,lateral extensions of the dorsum of the mesothorax (Fig. 2b, 3b, and 4b).Nymphs in the 3rd instar have obvious wing pads on the dorsum of both themeso- and metathorax (Fig. 2c, 3c, and 4c). Individuals in the 4th nymphalinstar have hind wing pads that cover at least the dorsum of the first abdominal segment and extend over the dorsum of the second abdominal segment (Fig. 2d, 3d, and 4d). By the 5th instar, the hindwing pads extendover 1/3-1/2 of the dorsum of the abdomen (Fig. 2e, 3e, and 4e). Someimmatures of H. deceptivus show little or no wing pad development; however, they can be distinguished from immatures of the other 3 species bytheir distinctive body shape (Fig. 1).
The following key is intended as a field key. Most of the characters usedfor separating the species can be seen with the use of a hand lens; however,some characters may require finer optics. Be aware that this key may notaccount for the polymorphism that occurs within species, and that thedrawings are not to scale.
KEY
1. Body pear shaped with posterior part of abdomen 1-1/2-3times as wide as head a)ld thoracic regions. Body color usuallydark brown to black. Fig 1. _ Hoplistoscelis deceptivu8
1'. Body elongate oval. Posterior part of abdomen usually never
a b d
Fig. 1. H oplistoscelis deceptivus: 1st-4th nymphal instars, a-d.
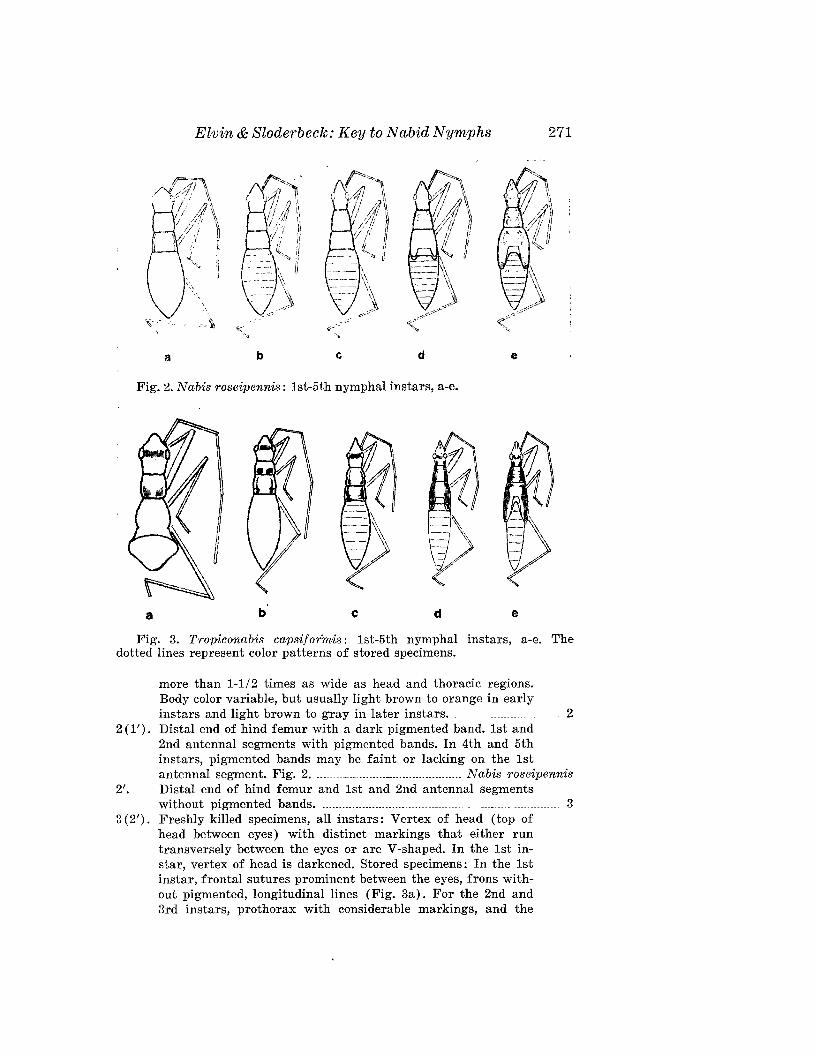
Elvin & Sloderbeck: Key to Nabid Nymphs 271
a b c d e
Fig. 2. Nabis roseipennis: 1st-5th nymphal instars, a-e.
a b c d e
Fig. 3. Tropiconabis capsiformis: 1st-5th nymphal instars, a-e. Thedotted lines represent color patterns of stored specimens.
more than 1-1/2 times as wide as head and thoracic regions.Body color variable, but usually light brown to orange in earlyinstars and light brown to gray in later instars. 2
2 (1'). Distal end of hind femur with a dark pigmented band. 1st and2nd antennal segments with pigmented bands. In 4th and 5thinstars, pigmented bands may be faint or lacking on the 1stantennal segment. Fig. 2. Nabis roseipennis
2'. Distal end of hind femur and 1st and 2nd antennal segmentswithout pigmented bands. 3
3(2'). Freshly killed specimens, all instars: Vertex of head (top ofhead between eyes) with distinct markings that either runtransversely between the eyes or are V-shaped. In the 1st instar, vertex of head is darkened. Stored specimens: In the 1stinstar, frontal sutures prominent between the eyes, frons without pigmented, longitudinal lines (Fig. 3a). For the 2nd and3rd instars, prothorax with considerable markings, and the
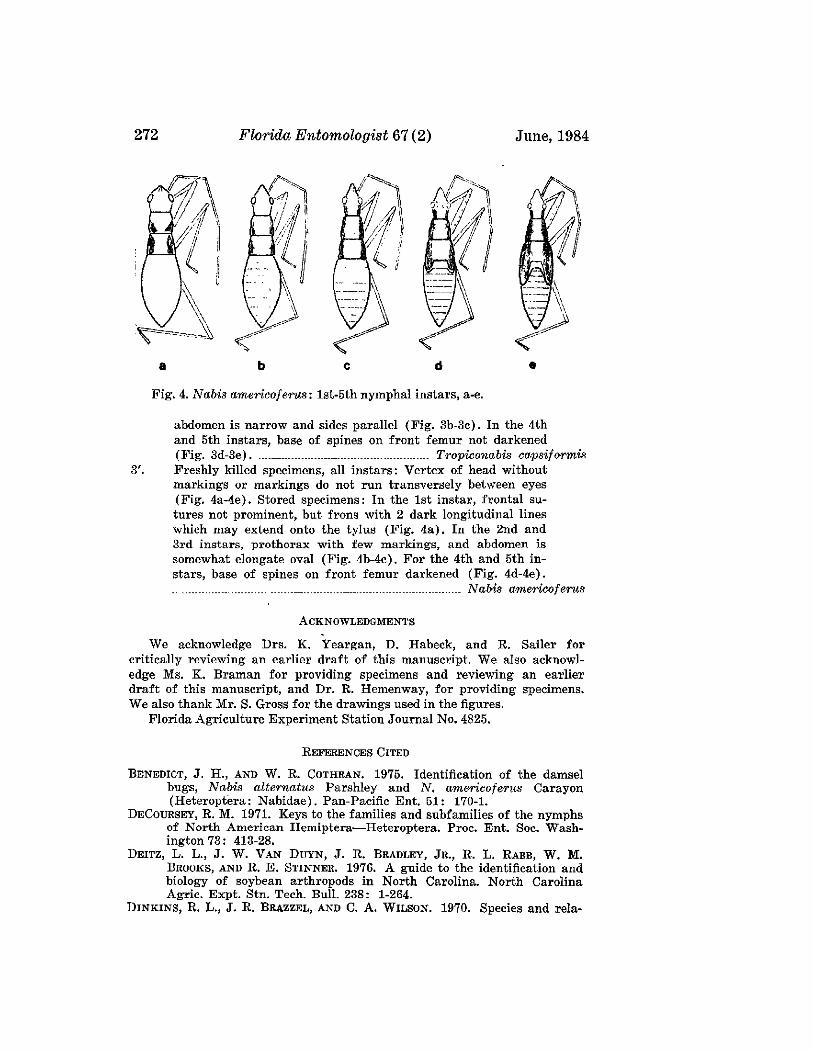
272 FlorilUJ, Entomologist 67 (2) June, 1984
•dcba
Fig. 4. Nabis amerieoferus: 1st-5th nymphal instars, a-e.
abdomen is narrow and sides parallel (Fig. 3b-3e). In the 4thand 5th instars, base of spines on front femur not darkened(Fig. 3d-3e). TropieonabUi eapsiformUi
3'. Freshly killed specimens, all instars: Vertex of head withoutmarkings or markings do not run transversely between eyes(Fig. 4a-4e). Stored specimens: In the 1st instar, frontal sutures not prominent, but frons with 2 dark longitudinal lineswhich may extend onto the tylus (Fig. 4a). In the 2nd and3rd instars, prothorax with few markings, and abdomen issomewhat elongate oval (Fig. 4b-4c). For the 4th and 5th instars, base of spines on front femur darkened (Fig. 4d-4e).
_____________ NabUi amerieoferus
ACKNOWLEDGMENTS
We acknowledge Drs. K. Yeargan, D. Habeck, and R. Sailer forcritically reviewing an earlier draft of this manuscript. We also acknowledge Ms. K. Braman for providing specimens and reviewing an earlierdraft of this manuscript, and Dr. R. Hemenway, for providing specimens.We also thank Mr. S. Gross for the drawings used in the figures.
Florida Agriculture Experiment Station Journal No. 4825.
REFERENCES CITED
BENEDICT, J. H., AND W. R. COTHRAN. 1975. Identification of the damselbugs, Nabis alternatu8 Parshley and N. americoferus Carayon(Heteroptera: Nabidae). Pan-Pacific Ent. 51: 170-1.
DECOURSEY, R. M. 1971. Keys to the families and subfamilies of the nymphsof North American IIemiptera-Heteroptera. Proc. Ent. Soc. Washington 73: 413-28.
DEITZ, L. L., J. W. VAN DUYN, J. R. BRADLEY, JR., R. L. RABB, W. M.BROOKS, AND R. E. STINNER. 1976. A guide to the identification andbiology of soybean arthropods in North Carolina. North CarolinaAgric_ Expt. Stn. Tech_ BulL 238: 1-264.
DINKINS, R. L., J. R. BRAZZEL, AND C. A. WILSON. 1970. Species and rela-

Elvin & Sloderbeck: Key to Nabid Nymphs 273
tive abundance of Chrysopa, Geocoris, and Nabis in Mississippi cottonfields. J. Econ. Ent. 63: 660-661.
HARRIS, H. M. 1928. A monographic study of the Hemipterous familyNabidae as it occurs in North America. Ent. Amer. 9: 1-97.
HORMCHAN, P., L. W. HEPNER, AND M. F. SCHUSTER. 1976. Predaciousdamsel bugs: Identification and distribution of the subfamily Nabinaein Mississippi. Mississippi Agric. For. Expt. Stn. Tech. Bull. 76: 1-4.
IRWIN, M. E., AND M. SHEPARD. 1980. Sampling predaceous Hemiptera onsoybeans. Pages 503-531 In M. Kogan, and D. C. Herzog, eds.Sampling methods in soybean entomology. Springer-Verlag, NewYork. 587 p.
NEAL, T. M. 1974. Predaceous arthropods in Florida soybean agroecosystem. M.S. Thesis. University of Florida, Gainesville, Florida. 194 p.
PITRE, H. N., T. L. HILLHOUSE, M. C. DONAHUE, AND H. C. KINARD. 1978.Beneficial arthropods on soybeans and cotton in different ecosystemsin Mississippi. Mississippi Agric. For. Expt. Stn. Tech. Bull. 90: 1-9.
SHEPARD, M., G. R. CARNER, AND S. G. TURNIPSEED. 1974. Seasonal abundance of predaceous arthropods in soybeans. Environ. Ent. 3: 985-8.
• • • • • • • • • • • • •DAMAGE OF SUNFLOWER BY THE SOUTHERN
ARMYWORM (LEPIDOPTERA: NOCTUIDAE)
EVERETT R. MITCHELLInsect Attractants, Behavior, and Basic Biology Research Laboratory,
Agricultural Research Service, USDA, Gainesville, FL 32604 USA
ABSTRACT
Spodoptera eridania (Cramer) occurred in economic infestations in ca.50% of the 567 ha of sunflower planted in Alachua County, Florida in thefall of 1982. Outbreaks occurred in sunflower having dense stands of pigweed (Amaranthus sp.), a primary host. After consuming pigweed, thesouthern armyworm larvae moved en masse to the sunflower. Differences inseed yields from sunflower heads harvested from plants with 0, 50, and100% defoliation caused by larval feeding were highly significant. Seedyields from plants showing 50 and 100% defoliation were reduced 45 and98%, respectively, compared to plants showing little or no defoliation. Theimpact of native parasites on S. eridania populations in sunflower wasnegligible. Parasites recovered included Opion sp., Chelonus insularisCresson, Meteorus autographae Muesebeck, and tachinids.
RESUMEN
Infestaciones de Spodoptera eridania (Cramer) causando danos economicos, ocurrio en aproximadamente et 50% de las 567 ha de girasolessembrados en el Condado de Alachue de la Florida en el otono de 1982.Brotes de infestacion ocurrioen girasoles teniendo un denso establecimientode A maranthus sp., que es un hospedero principal. Despues de comerse lasplantas de Amaranthus sp., las larvas de S. eridania se movieron en masahacia el girasol. Diferencias en el rendimiento de semillas de girasolescosechadas de plantas con un 0, 50, y 100% de defoliacion causado por laslarvas, fueron altamente significativas. Los rendimientos de semillas deplantas con un 50 y 100% de defoliacion fueron reducidos un 45 y 98%respectivamente cuando se compararon con plantas con poca 0 sin defoliacion.
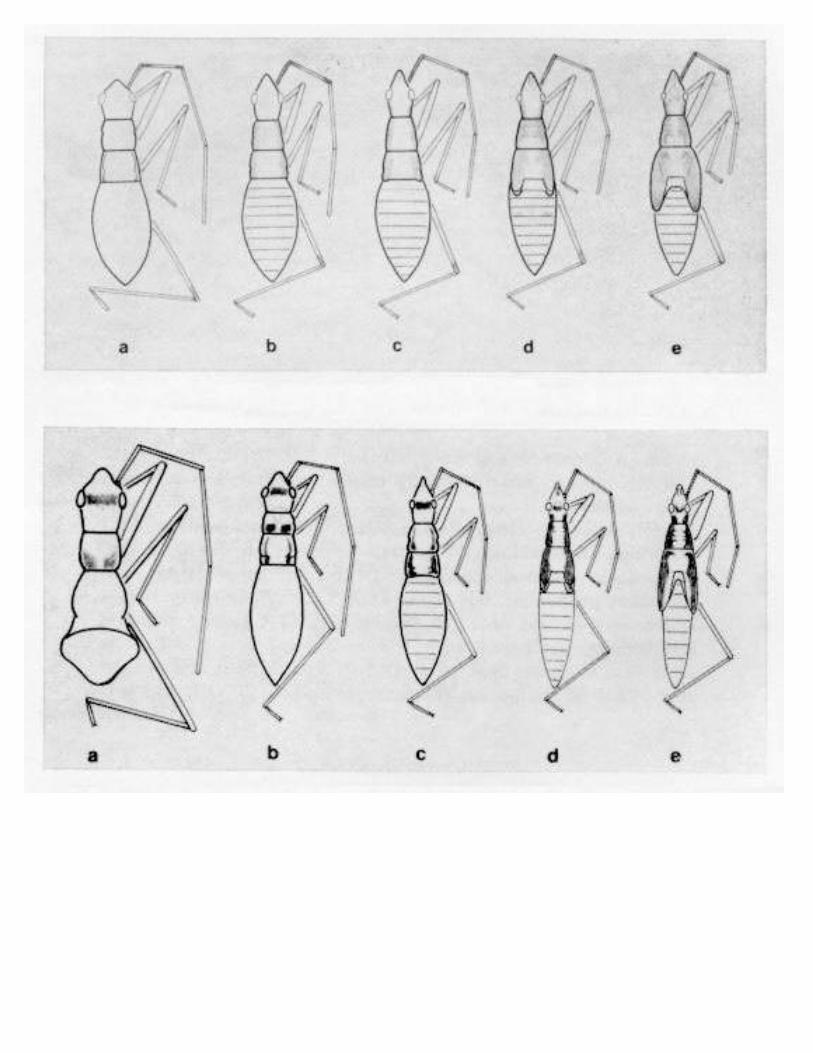
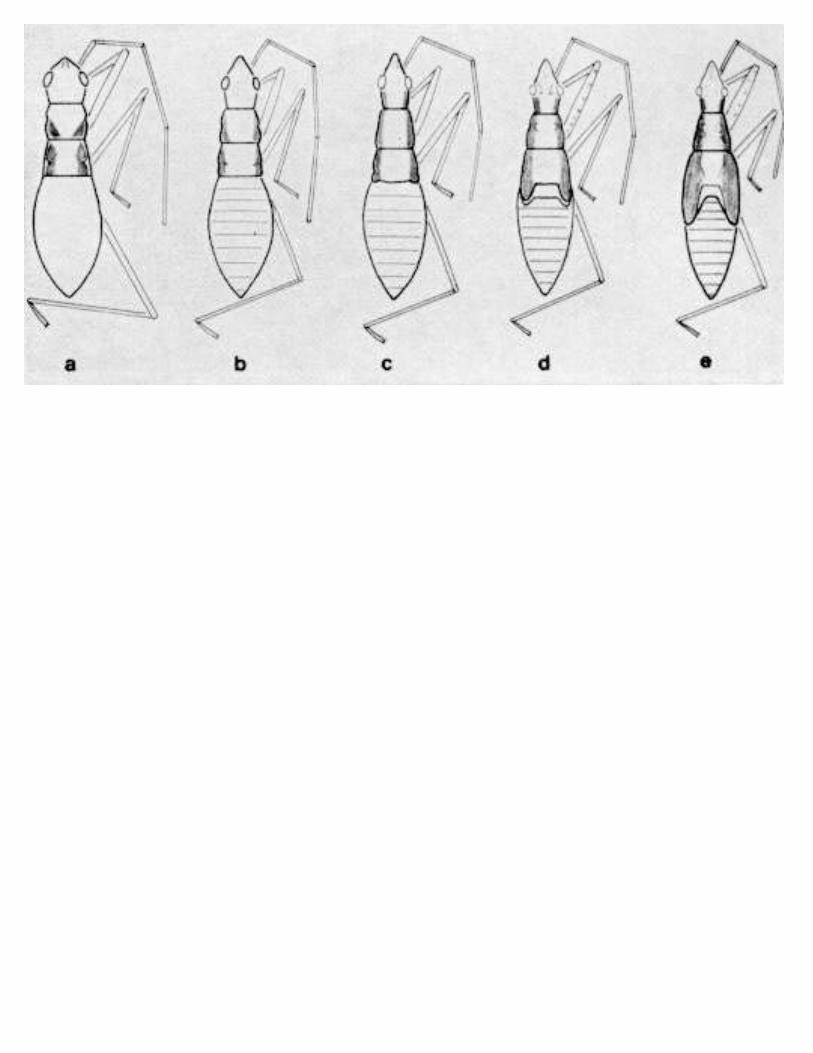

Elvin & Sloderbeck: Key to Nabid Nymphs 273
tive abundance of Chrysopa, Geocoris, and Nabis in Mississippi cottonfields. J. Econ. Ent. 63: 660-661.
HARRIS, H. M. 1928. A monographic study of the Hemipterous familyNabidae as it occurs in North America. Ent. Amer. 9: 1-97.
HORMCHAN, P., L. W. HEPNER, AND M. F. SCHUSTER. 1976. Predaciousdamsel bugs: Identification and distribution of the subfamily Nabinaein Mississippi. Mississippi Agric. For. Expt. Stn. Tech. Bull. 76: 1-4.
IRWIN, M. E., AND M. SHEPARD. 1980. Sampling predaceous Hemiptera onsoybeans. Pages 503-531 In M. Kogan, and D. C. Herzog, eds.Sampling methods in soybean entomology. Springer-Verlag, NewYork. 587 p.
NEAL, T. M. 1974. Predaceous arthropods in Florida soybean agroecosystem. M.S. Thesis. University of Florida, Gainesville, Florida. 194 p.
PITRE, H. N., T. L. HILLHOUSE, M. C. DONAHUE, AND H. C. KINARD. 1978.Beneficial arthropods on soybeans and cotton in different ecosystemsin Mississippi. Mississippi Agric. For. Expt. Stn. Tech. Bull. 90: 1-9.
SHEPARD, M., G. R. CARNER, AND S. G. TURNIPSEED. 1974. Seasonal abundance of predaceous arthropods in soybeans. Environ. Ent. 3: 985-8.
• • • • • • • • • • • • •DAMAGE OF SUNFLOWER BY THE SOUTHERN
ARMYWORM (LEPIDOPTERA: NOCTUIDAE)
EVERETT R. MITCHELLInsect Attractants, Behavior, and Basic Biology Research Laboratory,
Agricultural Research Service, USDA, Gainesville, FL 32604 USA
ABSTRACT
Spodoptera eridania (Cramer) occurred in economic infestations in ca.50% of the 567 ha of sunflower planted in Alachua County, Florida in thefall of 1982. Outbreaks occurred in sunflower having dense stands of pigweed (Amaranthus sp.), a primary host. After consuming pigweed, thesouthern armyworm larvae moved en masse to the sunflower. Differences inseed yields from sunflower heads harvested from plants with 0, 50, and100% defoliation caused by larval feeding were highly significant. Seedyields from plants showing 50 and 100% defoliation were reduced 45 and98%, respectively, compared to plants showing little or no defoliation. Theimpact of native parasites on S. eridania populations in sunflower wasnegligible. Parasites recovered included Opion sp., Chelonus insularisCresson, Meteorus autographae Muesebeck, and tachinids.
RESUMEN
Infestaciones de Spodoptera eridania (Cramer) causando danos economicos, ocurrio en aproximadamente et 50% de las 567 ha de girasolessembrados en el Condado de Alachue de la Florida en el otono de 1982.Brotes de infestacion ocurrioen girasoles teniendo un denso establecimientode A maranthus sp., que es un hospedero principal. Despues de comerse lasplantas de Amaranthus sp., las larvas de S. eridania se movieron en masahacia el girasol. Diferencias en el rendimiento de semillas de girasolescosechadas de plantas con un 0, 50, y 100% de defoliacion causado por laslarvas, fueron altamente significativas. Los rendimientos de semillas deplantas con un 50 y 100% de defoliacion fueron reducidos un 45 y 98%respectivamente cuando se compararon con plantas con poca 0 sin defoliacion.

274 Florida Entomologist 67 (2) June, 1984
EI impacto de parasitos nativos en poblaciones de S. eridania fue neglegible.Los parasitos encontrados incluyeron Opium sp., Chelonus insularis Cresson,Meteorus autographae Muesebeck, y tachinedos.
Increasing production costs, high land prices, and low commodity pricesare causing farmers to seek more effective means of using machinery andland resources in the production of crops such as corn, soybean, andsorghum. Double cropping is gaining favor as one way of spreading fixedcosts over a longer growing season. Double cropping also provides opportunities for more efficient utilization of fertilizer where a second cropuses the residue from the first crop. Production of more than one crop onthe same land each year also reduces the financial risks for growers whomay experience losses in one crop because of inclement weather conditions,insects, or other natural disasters.
Growers in north-central Florida are experimenting with sunflower,Helianthus annuus L., as an alternate crop in double cropping systems.Sunflower is resistant to frost, and yields are high when planted the firsthalf of February in the Gainesville, Florida area. When sunflower is plantedin early February in northern Florida, the crop escapes severe losses bydiseases and insects. Sunflower planted in August also shows promise as alate-season crop in north-central Florida because late plantings usually arenot seriously damaged by mildew or rust, although the crop may haveheavy infestations of the corn earworm, Heliothis zea (Boddie), and thesunflower moth, Homoeosoma electellum (Hulst) (Bailey et al. 1978).
This paper reports outbreaks of the southern armyworm, S. eridania(Cramer), in sunflower in Alachua County, Florida, in October 1982. Thisis the first documented report of this species as a pest of commercial sunflower. The impact of plant defoliation by southern armyworm larvae onsunflower yields also w~s evaluated.
MATE~IALSAND METHODS
Approximately 567 ha of sunflower was grown in Alachua County in1982, 50% of which were infested with southern armyworm larvae. Twofields infested with southern armyworm were selected as study sites. Eachfield was ca. 16 ha and was adjacent to cultivated sunflower on at least oneside. Study fields were separated by ca. 5 km, and were planted to sunflower(cv. 'Dalgren 164') between 25 August (field A) and 28 August (field B).
Initial observations were made on 20 October when plants were ca. 1.75m in height and in full bloom (i.e., ray petals fully exposed). The plantswere already infested with larvae when first examined. Approximately 450larvae were collected from each field, classified as small (..e:::0.8 cm), medium(0.9-3.4 cm), or large (>3.5 cm), and placed individually in a 30 ml sizeplastic cup containing sufficient diet (Leppla et al. 1979) for completelarval development. Cups were checked several times a week, and the fateof each larva recorded-larva died before pupation, pupa died, mothemerged, or parasite emerged.
Degrees of plant defoliation by southern armyworm larvae on sunflowerhead size (1 head/plant on 'Dalgren 164') was determined by measuring thewidth of 10 consecutive mature heads at 10 randomly selected sites in dif-

Mitchell: Southern Armyworm on Sunflower 275
ferent areas of the field. Estimates of defoliation were made within 7 daysafter most (>98%) of the larvae had matul'ed and dropped from the plantsto the soil to pupate. Three defoliation levels were estimated: 0-5%-littleor no feeding damage on any leaves; 50%-nearly all leaves showing signsof heavy feeding with many large holes and some leaves almost totallyeonsumed; 100%-all green leaf tissue consumed with only major leaf veinsand the stalk remaining (Fig. 1). Groups of 10 consecutive plants that mostnearly fit the leaf damage criteria for each of the 3 categories were selectedand marked for evaluation of head size and yield when the plants matured.A total of 100 heads (10 replicates of 10 heads each) were measured forf:lach of the 3 defoliation levels. The effect of plant defoliation on seed yieldwas determined by harvesting 10 consecutive mature heads from the sameareas from which the head measurements were taken. Heads were air dried,hand threshed, and weighed. The data were analyzed using Student's t-test.
RESULTS AND DISCUSSION
Most of the larvae collected from fields A and B on 20 October weremedium-sized. Only a few were classified as small or large (ca. 10% foreach class). Examination of other infested fields in Alachua County revealeda similar larval age, indicating that infestations apparently resulted froma general flight of adults that occurred simultaneously over the entire area.
The southern armyworm is a polyphagous species that attacks a widevariety of weeds and cultivated crops (tomatoes, potatoes, celery, sweetpotato, and various flowering ornamentals) (Kimball 1965). Pigweed,Amaranthus sp. (Amaranthaceae), is a favored host of the southern armyworm (Tingle et al. 1978). The severe armyworm infestations observed instudy fields A and B, as well as elsewhere in the area, were directly associated with pigweed infestations of cultivated fields.
In those areas of the fields where armyworm densities were greatest,
Fig. 1. Defoliation levels used for assessing impact of southern armyworm feeding on sunflower (left to right) : 0-5% damage, 50% damage, and100% damage.

276 Florida Entomologist 67 (2) June, 1984
larvae had consumed all but the stems of pigweed before moving to thesurrounding sunflower. Ten to 25 medium and large larvae/sunflower werecommon throughout the infested areas when first observed. Virtually allsunflower in field A, and an estimated 70% in field B, had a high level ofdefoliation (50-100%).
After larvae consumed the sunflower foliage, they moved to the head.Most head feeding was confined to the soft tissue on the backside andperiphery, but larvae chewing through the backside into the head was rare.Heads were generally not totally destroyed by feeding larvae except whenthe initial attack occurred during the late budding stage.
Sunflower defoliation had a significant effect on head size and on weightof the seed produced (Table 1). Severe defoliation affected seed weightmuch more than head size. Reduction in yield at the 50% defoliation levelreflected a reduced capacity on the part of the sunflower plant to furnishthe nutrients required to fill its seed, as was evident by the large number offully formed but "light" seed. Plants classified as 100% defoliated producedonly small heads with few or no seed.
The impact of native parasites on southern armyworm densities in sunflower was negligible, as is often the case with insect pests occurring in outbreaks. Of the 996 larvae collected and held in the laboratory, 426 producedadults (ca. an equal number of males and females), 159 produced parasites,and 411 died from unknown causes. Parasites recovered included Ophionsp. (16), Chelonus insularis Cresson (6), Meteorus autographae Muesebeck(75), and tachinids (62). Forty-seven larvae yielded 1 fly each; 2 or moreflies were recovered from 15 larvae. Counting only live larvae that producedadults or parasites, ca. 27% of the larvae were parasitized.
In conclusion, the southern armyworm can be a serious pest of sunflowerin fields where pigweed is allowed to grow in the crop. Egg masses or newlyhatched larvae were never found on sunflower in any field in this study,.which indicated that. the southern armyworm probably did not ovipositdirectly on the sunflower. Therefore, the most obvious control measure for
TABLE 1. EFFECTS OF DEFOLIATION BY SOUTHERN ARMYWORM LARVAE ONHEAD SIZE AND YIELD OF SUNFLOWER (CV. 'DALGREN 164').ALACHUA COUNTY, FLORIDA. OCTOBER-NoVEMBER, 1982.
Defoliationlevel (%)
0-550
100
0-550
100
X Headdiam.(mm ± S.E.)l
Field A
149.5 ± 3.2 a117.5±3.7b
Field B
131.7 ± 3.5 a123.5 ± 4.0 a84.1 ± 3.1 b
X Seed wt/head(g ± S.E.)l
31.8 ± 2.0 a8.1 ± 1.1 b
74.6 ± 4.1 a40.7 ± 3.6 b1.2 ± 0.3 c
'Means followed by different letters in the same column differ significantly at the 1%level, Student's t-test.

Mitchell: Southern Armyworm on Sunflower 277
this sunflower pest involves early destruction of pigweed in and aroundfields.
ACKNOWLEDGMENT
I thank Bill Copeland and Ron Hines for their assistance in locatinginfested fields, collecting and rearing larvae, cleaning and weighing sunflower seed, and summarizing data. Revised manuscript received for publication 27 January 1984.
REFERENCES CITED
BAILEY, B. A., E. B. WHITTY, AND V. E. GREEN, JR. 1978. Agronomy facts:Sunflower production in Florida. Florida Coop. Ext. Servo Publ. No.74, University of Florida, Gainesville. 4 p.
KIMBALL, C. P. 1965. Lepidoptera of Florida. Div. of Plant Industry,Florida Dept. of Agric., Gainesville, FL. 363 p.
LEPPLA, N. C., P. V. VAIL, AND J. R. RYE. 1979. Mass rearing and handlingtechniques for the cabbage looper. Pages 57-75 In Proc. Radioisotopesand Radiation in Ent., Training course, FAO/IAEA.
TINGLE, F. C., T. R. ASHLEY, AND E. R. MITCHELL. 1978. Parasites ofSpodoptera exigua, S. eridania [Lep.: Noctuidae] and Herpetogrammabipunetalis [Lep.: Pyralidae] collected from A maranthus hybridus infield corn. Entomophaga 23: 343-7.
• • • •• • • • • • • • •POPULATION FLUCTUATIONS OF THE SCHAUS
SWALLOWTAIL (LEPIDOPTERA: PAPILIONIDAE)ON THE ISLANDS OF BISCAYNE BAY, FLORIDA,
WITH COMMENTS ON THE BAHAMAN SWALLOWTAIL
WILLIAM F. LOFTUS AND JAMES A. KUSHLANNational Park Service, South Florida Research Center
P.O. Box 279, Homestead, FL 33030 USA
ABSTRACT
Censuses of the Schaus swallowtail butterfly (Heraelides (= Papilio)aristodemus poneeanus (Schaus», a federally listed threatened species,were conducted from 1979 to 1982 on islands in Biscayne Bay, southernFlorida. The population size was small during 1979 and 1980, but adultswere widely distributed in suitable habitat. Emergence season was similarin both years, from late April to late June. Numbers of the Schaus swallowtail were lower in 1981, and the emergence period seemed to be retarded,probably due to the very dry winter and spring. Following the wet winterof 1981-1982, the largest numbers of adult Schaus swallowtails since 1972were recorded. It appears that the severity of the winter dry season maydetermine the population size of this species in southern Florida.
In combination with results of previous censuses, these data provide acontinuous eleven-year record of Schaus swallowtail population fluctuationson the Biscayne Bay islands. Human disturbances, especially habitat destruction, have reduced the numbers and range of the Schaus swallowtail, sothe butterflies on these islands presently form the nucleus of its population


Mitchell: Southern Armyworm on Sunflower 277
this sunflower pest involves early destruction of pigweed in and aroundfields.
ACKNOWLEDGMENT
I thank Bill Copeland and Ron Hines for their assistance in locatinginfested fields, collecting and rearing larvae, cleaning and weighing sunflower seed, and summarizing data. Revised manuscript received for publication 27 January 1984.
REFERENCES CITED
BAILEY, B. A., E. B. WHITTY, AND V. E. GREEN, JR. 1978. Agronomy facts:Sunflower production in Florida. Florida Coop. Ext. Servo Publ. No.74, University of Florida, Gainesville. 4 p.
KIMBALL, C. P. 1965. Lepidoptera of Florida. Div. of Plant Industry,Florida Dept. of Agric., Gainesville, FL. 363 p.
LEPPLA, N. C., P. V. VAIL, AND J. R. RYE. 1979. Mass rearing and handlingtechniques for the cabbage looper. Pages 57-75 In Proc. Radioisotopesand Radiation in Ent., Training course, FAO/IAEA.
TINGLE, F. C., T. R. ASHLEY, AND E. R. MITCHELL. 1978. Parasites ofSpodoptera exigua, S. eridania [Lep.: Noctuidae] and Herpetogrammabipunetalis [Lep.: Pyralidae] collected from A maranthus hybridus infield corn. Entomophaga 23: 343-7.
• • • •• • • • • • • • •POPULATION FLUCTUATIONS OF THE SCHAUS
SWALLOWTAIL (LEPIDOPTERA: PAPILIONIDAE)ON THE ISLANDS OF BISCAYNE BAY, FLORIDA,
WITH COMMENTS ON THE BAHAMAN SWALLOWTAIL
WILLIAM F. LOFTUS AND JAMES A. KUSHLANNational Park Service, South Florida Research Center
P.O. Box 279, Homestead, FL 33030 USA
ABSTRACT
Censuses of the Schaus swallowtail butterfly (Heraelides (= Papilio)aristodemus poneeanus (Schaus», a federally listed threatened species,were conducted from 1979 to 1982 on islands in Biscayne Bay, southernFlorida. The population size was small during 1979 and 1980, but adultswere widely distributed in suitable habitat. Emergence season was similarin both years, from late April to late June. Numbers of the Schaus swallowtail were lower in 1981, and the emergence period seemed to be retarded,probably due to the very dry winter and spring. Following the wet winterof 1981-1982, the largest numbers of adult Schaus swallowtails since 1972were recorded. It appears that the severity of the winter dry season maydetermine the population size of this species in southern Florida.
In combination with results of previous censuses, these data provide acontinuous eleven-year record of Schaus swallowtail population fluctuationson the Biscayne Bay islands. Human disturbances, especially habitat destruction, have reduced the numbers and range of the Schaus swallowtail, sothe butterflies on these islands presently form the nucleus of its population

278 Florida Entomologist 67 (2) June, 1984
in the United States. The Bahaman swallowtail (Heraelides (~ Papilio)andraernon bonhotei (Sharpe» was not observed during the censuses, andwe conclude that it is not presently established on the islands we studied.
RESUMEN
Desde 1979 al 1982, en las islas de Biscayne Bay en el sur de la Floridase hizo un censo de la mariposa, Heraclides ( = Papilio) OIf'istodemusponeeanus, la cua! el gobierno federal la ha declarado como una especieamenazada de extincion. EI numoro de la poblacion durante 1979 y 1980fue minima, pero los adultos estaban bien distribuidos en habitaci6n adecuada. La estaci6n de emersi6n fue muy similar durante los dos alios, desdefinales de Abril al final de Junio. Los numeros de la mariposa Schaus fueronmas bajos en 1981 y el perfodo de emersion parece haber sida atrasado,posiblemente debido al invierno y la primavera seea. Luego del inviernohumedo de 1981-1982, se registr6 el mayor numero de la mariposa Schausdesde el 1972. Asi que parece que la severidad de la estaci6n seea del inviernopuede determinar el tamano de la poblacion de esta especie en nuestra areade estudio.
Junto a otros censos estos datos proveen un record continuo de onceanos de las fluctuaci6nes en Ia poblaci6n de la mariposa Schaus en los cayosde la Bahia de Biscayne. Los disturbios humanos1 especialmente la destruccion del ambiente han reducido los numeros y el area geografica de lamariposa Schaus y por 10 tanto las mariposas en estas islas forman elDueleo de la poblaci6n de este especie en los Estado Unidos. La mariposaHeraclides (=Papilio) andraemon bonhoteino Se observ6 durante los censos,y dudamos que est. establecida alii.
The Schaus swallowtail (Heraelides (~ Papilio) aristodernus poneeanus(Schaus» was first described at the turn of this century from the Miamiarea of Dade County in southern Florida (Schaus 1911). It was eliminatedfrom that site as the 'city developed but was rediscovered further south onthe Florida Keys, Monroe County. The species received much attention following the 1935 hurricane because of a report of its extirpation on the keysby the storm (Grimshawe 1940). Henderson (1945a, b) later documentedthe continued existence of the Schaus swallowtail in the Florida Keys. Bythis time, the butterfly had become a glamour species in great demand bycollectors, and, during the next two decades, both Klots (1951) and Kimball(1965) noted that over-collection of this rare species was possible. In 1972,Covell and Rawson (1973) and Brown (1973a, b) independently surveyedthe upper Florida Keys, including the Elliott Key group in Biscayne Bay,Dade Gounty, to the north of the main-line Florida Keys. They found thatthe Schaus swallowtail was well-established on these northern islands whereBrown (1973a) reported seeing 100 adults in a day. From 1973 to 1976,Covell (1977) continued to observe this species on the islands of BiscayneBay but found that it had decreased in numbers from the levels of 1972. Hespeculated that the decrease may have been the result of winter droughtconditions that inhibited new growth of the larval food plants during thoseyears. Limited surveys were also conducted during 1977 and 1978, with fewadults being observed on the keys (J. Tilmant, pers. comm.).
The Bahaman swallowtail (fle1"aclideR (= Papilio) o:ndraemon bonhote:i(Sharpe» has been recorded from southeastern Florida and the Florida

Loftus & Kushlan: Schaus Swallowtail on Biscayne Bay 279
Keys on several occasions during this century. Only on a few occasions hasthe existence of a breeding population been suggested. The collection of afresh adult on Long Key in Monroe County, Florida was used to supportsuch a contention (Kimball 1965), as was a more recent report by Brown(1973a, b) of large numbers of adults, including ovipositing females, on theislands in Biscayne Bay.
The two swallowtail butterflies became the first invertebrates to beincluded on the U. S. List of Endangered and Threatened Species. Theoverlapping range of these species in the United States was thought to belimited to tropical hardwood hammocks of the upper Florida Keys, including those in southern Biscayne Bay, where environmental conditions aresuitable for hammock growth. Such hammocks are normally found on thehighest land in southern Florida and the Florida Keys, usually near thecoast where rapid urban and commercial development has taken place.Habitat destruction has been devastating to the butterflies, especially considering that their population sizes and ranges in Florida appear to havebeen small under natural conditions.
Little biological data existed for these butterflies at the start of ourstudy. Our purpose was to determine the status and distribution of thesespecies on the relatively undisturbed islands in Biscayne Bay, to documentthe season and the duration of adult emergence, to confirm the larval foodplants, and to define those factors that affect the population status of thebutterflies. Preliminary results from 1979-1981 censuses and suggestedmanagement options for the Schaus swallowtail were presented by Loftusand Kushlan (1982). A companion study by Gerold Morrison (1981) examined food plants and larval ecology. Both species of butterflies, originallyplaced in the genus Papilio, have recently been reassigned to the genusH eraclides (Miller and Brown 1981) .
STUDY AREA
We studied two islands in Biscayne Bay, Elliott Key and Old RhodesKey, because of their mature hardwood hammocks in which many swallowtails had been sighted in recent years.
On Elliott Key, we established an approximately 1-km transect south ofBilly's Point, near Petrel Point (Fig. 1). This route crossed the island fromthe ocean to the bay through mature hammock, included a section of thecleared central portion of the island, and looped southward into a hammockon the bay side. The transect included large stands of the host plants forSchaus swallowtail larvae, torchwood (Amyris elemifera) and wild lime(Zanthoxylu1'n fagara) (Rutkowski 1971), and an old grove of key limes(Citrus aurantifolia) , the reported larval host plant of the Bahamanswallowtail (Klots 1951). A similar transect on Old Rhodes Key was anapproximately 1-km loop through a mature hammock on the southern tip ofthe island (Fig. 1). This hammock had an unbroken canopy, except wheretrees had fallen and hosted large stands of torchwood and wild lime trees.
On the surveys in 1980, additional monthly visits were also made to anabandoned key lime grove at the northern tip of Totten Key (Fig. 1). Thelime trees were mixed with encroaching hammock species, resulting in amore open canopy than in the mature hammock.
Rainfall on the keys follows a pattern similar to that of the mainland,
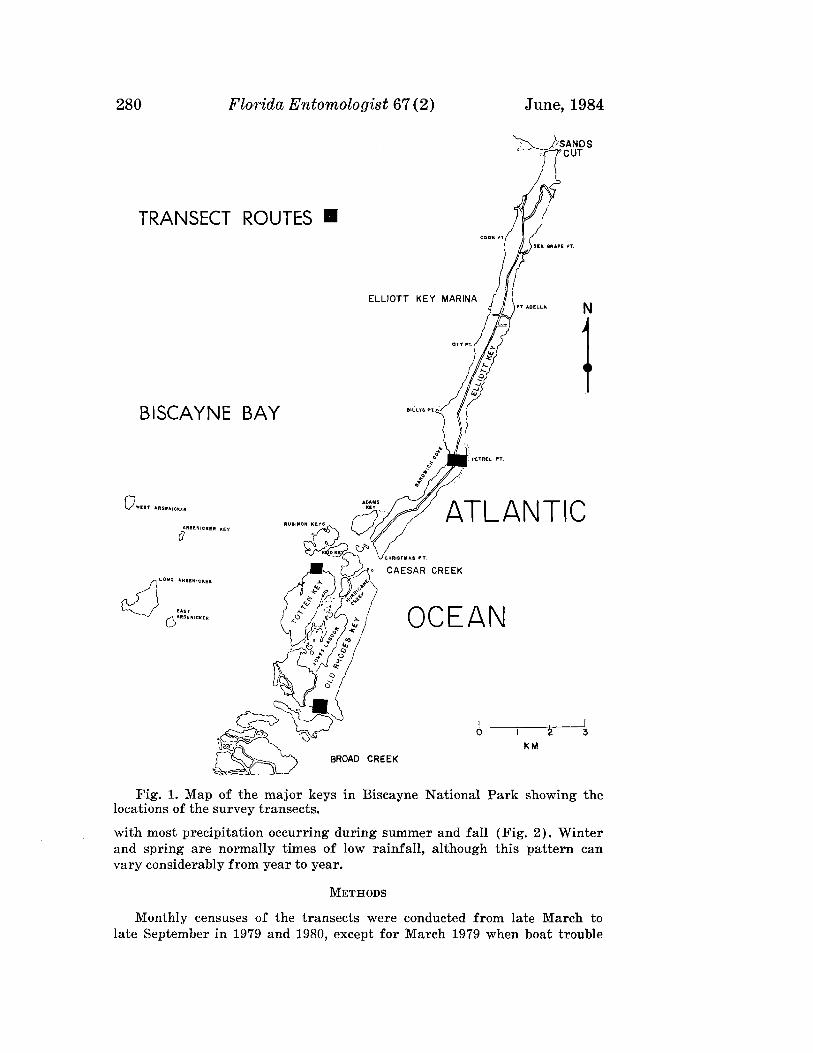
280 Florida Entomologist 67 (2) June, 1984
3
N
t
I ~
KM
OCEAN
o
CAESAR CREEK
ELLIOTT KEY MARINA
TRANSECT ROUTES •
BISCAYNE BAY
D"'[Sf ARSI!~tCkER
~LON' "SEN'eK,.
~ FAnOARS'ENICl(ER
Fig. 1. Map of the major keys in Biscayne National Park showing thelocations of the survey transects.
with most precipitation occurring during summer and fall (Fig. 2). Winterand spring are normally times of low rainfall, although this pattern canvary considerably from year to year.
METHODS
Monthly censuses of the transects were conducted from late March tolate September in 1979 and 1980, except for March 1979 when boat trouble
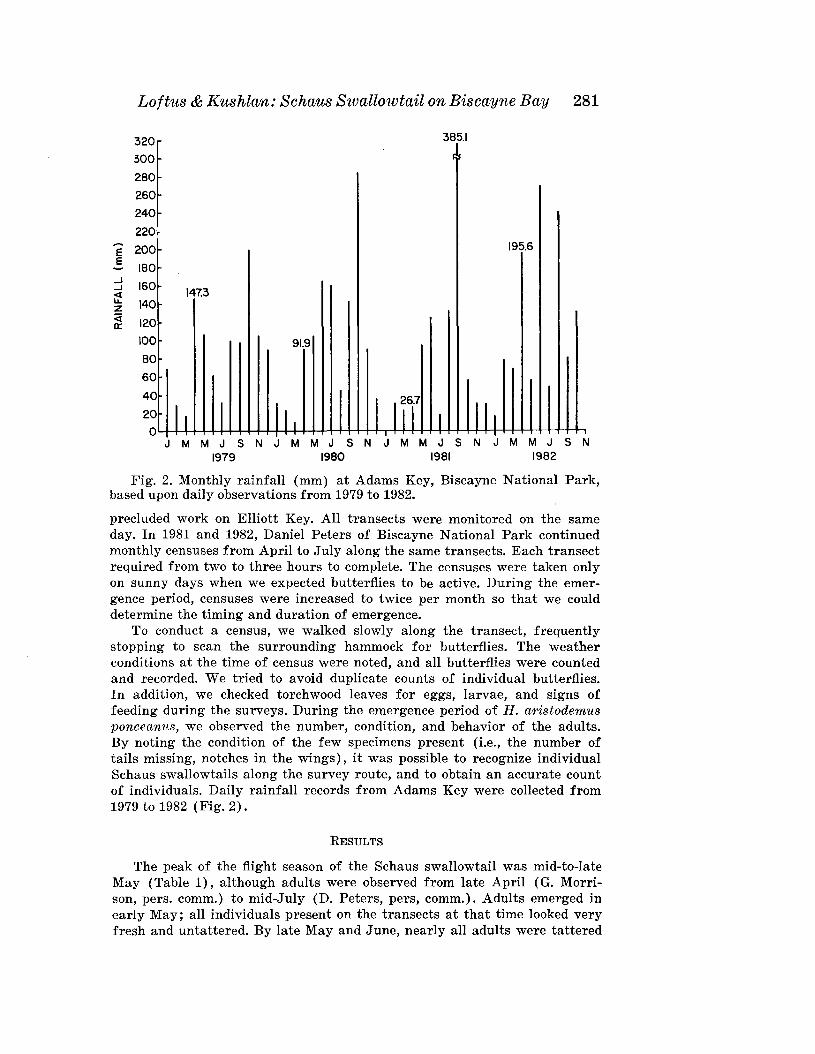
Loftus & Kushlan: Schaus Swallowtail on Biscayne Bay 281
385.1
~
t195.6
147.3
91.9
II I IL IllY I III
320
300
280
260
240
220
E 200E 180
160
140
120
100
80
60
40
20
a
...J
...J
~z<i0::
J M M J S N J M M J S N J M M J S N J M M J S N1979 1980 1981 1982
Fig. 2. Monthly rainfall (mm) at Adams Key, Biscayne National Park,based upon daily observations from 1979 to 1982.
precluded work on Elliott Key. All transects were monitored on the sameday. In 1981 and 1982, Daniel Peters of Biscayne National Park continuedmonthly censuses from April to July along the same transects. Each transectrequired from two to three hours to complete. The censuses were taken onlyon sunny days when we expected butterflies to be active. During the emergence period, censuses were increased to twice per month so that we coulddetermine the timing and duration of emergence.
To conduct a census, we walked slowly along the transect, frequentlystopping to scan the surrounding hammock for butterflies. The weatherconditions at the time of census were noted, and all butterflies were countedand recorded. We tried to avoid duplicate counts of individual butterflies.In addition, we checked torchwood leaves for eggs, larvae, and signs offeeding during the surveys. During the emergence period of H. aristodemusponceanus, we observed the number, condition, and behavior of the adults.By noting the condition of the few specimens present (i.e., the number oftails missing, notches in the wings), it was possible to recognize individualSchaus swallowtails along the survey route, and to obtain an accurate countof individuals. Daily rainfall records from Adams Key were collected from1979 to 1982 (Fig. 2).
RESULTS
The peak of the flight season of the Schaus swallowtail was mid-to-IateMay (Table 1), although adults were observed from late April (G. Morrison, pers. comm.) to mid-July (D. Peters, pers, comm.). Adults emerged inearly May; all individuals present on the transects at that time looked veryfresh and untattered. By late May and June, nearly all adults were tattered
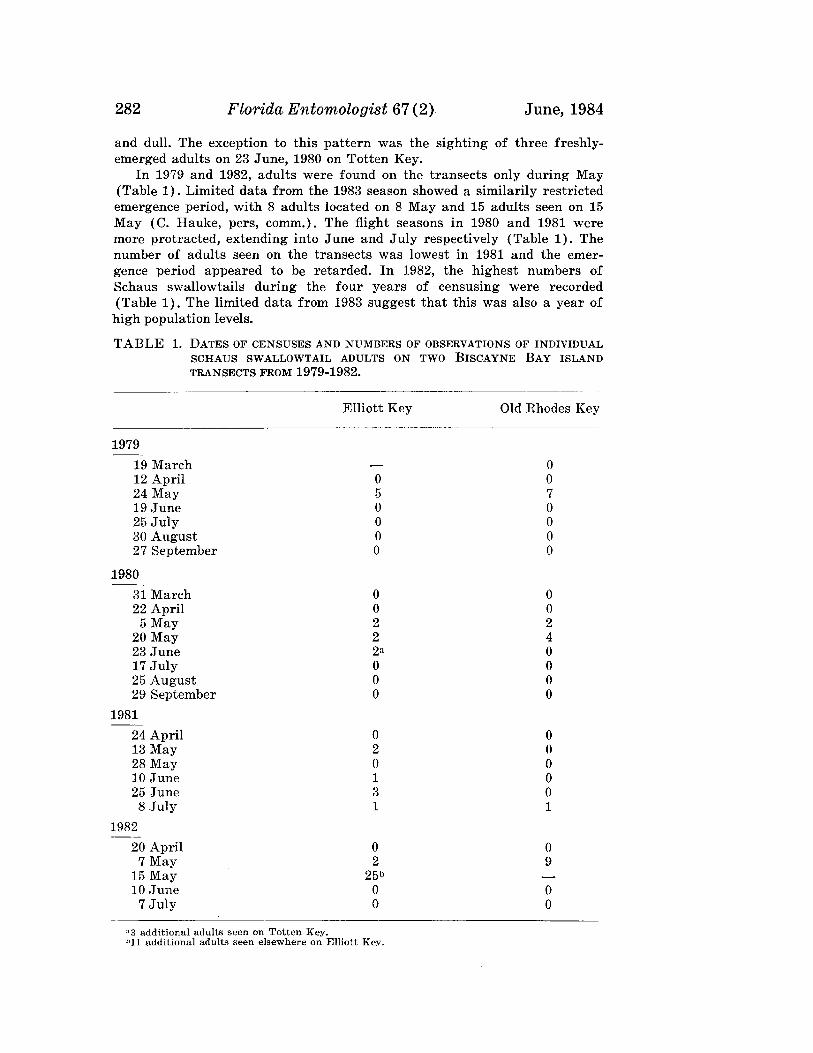
282 Florida Entomologist 67 (2) June, 1984
and dull. The exception to this pattern was the sighting of three freshlyemerged adults on 23 June, 1980 on Totten Key.
In 1979 and 1982, adults were found on the transects only during May(Table 1). Limited data from the 1983 season showed a similarily restrictedemergence period, with 8 adults located on 8 May and 15 adults seen on 15May (C. Hauke, pers, comm.). The flight seasons in 1980 and 1981 weremore protracted, extending into June and July respectively (Table 1). Thenumber of adults seen on the transects was lowest in 1981 and the emergence period appeared to be retarded. In 1982, the highest numbers ofSchaus swallowtails during the four years of censusing were recorded(Table 1). The limited data from 1983 suggest that this was also a year of
high population levels.
TABLE 1. DATES OF CENSUSES AND NUMBERS OF OBSERVATIONS OF INDIVIDUALSCHAUS SWALLOWTAIL ADULTS ON TWO BISCAYNE BAY ISLANDTRANSECTS FROM 1979-1982.
Elliott Key Old Rhodes Key
1979
19 March 012 April 0 024 May 5 719 June 0 025 July 0 030 August 0 027 September 0 0
1980
31 March 0 022 April 0 0
5 May 2 220 May 2 423 June 2(1 017 July 0 025 August 0 029 September 0 0
1981--
24 April 0 013 May 2 028 May () 010 June 1 025 June ;{ 08 July 1 1
1982--
20 April 0 07 May 2 9
15 May 25b
10.June 0 07 July 0 0
"3 additional adults seen on Totten Key.hl1 additional adults seen elsewhere on Elliott Key.

Loftus & Kushlan: Schaus Swallowtail on Biscayne Bay 283
Surveys of the keys in Biscayne National Park during peak emergencetimes revealed that the Schaus swallowtail is widespread. It appears to belimited to hammock and hammock edge habitats. We observed specimens onElliott, Old Rhodes, Adams, and Totten Keys and assume that this speciesis present in suitable habitat throughout the Park.
We rarely observed mating or oviposition, although courtship activitywas common. On 24 May 1979 we watched a Schaus swallowtail oviposit onwild lime (Zanthoxylum). Efforts to locate Schaus swallowtail larvae onnew torchwood leaves were usually unsuccessful, although fresh feedingsigns were often visible. We were able to find and photograph only a fewlarvae during this study. Fresh feeding sign was most apparent in May andearly June, with a few weeks of adult emergence. The few larvae discoveredalong the transects were also located during May.
We never found the Bahaman swallowtail on the keys in Biscayne Bay,in spite of attempts to locate this species. We planned our census times toinclude the times of Brown's (1973a, b) sightings. The transects includedold key lime plantings, and if this butterfly was present, we would have expected to find it there. Dennis Leston (pers. comm.) and Gerold Morrison(pers. comm.) were studying butterflies on the keys at the times of ourcensuses and did not find this species. Bahaman swallowtails were not observed in 1981 or 1982 surveys on these keys (D. Peters, pers. comm.; C. V.Covell, Jr., pers. comm.).
DISCUSSION
BAHAMAN SWALLOWTAIL
The specimen history of the Bahaman swallowtail in Florida has beensporadic. Some records appear dubious and few provide any evidence thatthe butterfly has been established (Klots 1951, Kimball 1965, Florida Gameand Fresh Water Fish Commission 1982). Records of H. andraemon inFlorida appear to be the result of sporadic colonizations during which shortterm breeding colonies may form, only to eventually die out (Miller 1975,Morrison 1981). Morrison (1981) pointed out that current taxonomic interpretation of H. andraemon does not recognize a distinct Florida subspecies.This would indicate that if establishment had ever taken place, it has notbeen of sufficient duration for morphological differentiation to have developed.
The sightings by Brown (1973a) and his students provided the impetusto list the Bahaman swallowtail as a threatened species. They reported thepresence of numerous adults on Elliott Key in 1972, including females thatwere ovipositing on citrus trees. Covell (1976) was collecting on Elliott Keyin 1972, and, although he collected Schaus swallowtails, he did not observethe Bahaman swallowtails. Covell (1977) continued to collect in the parkarea from 1973-1976 but never observed H. andraemon bonhotei. This specieswas not observed during surveys of Elliott Key in 1977-1978 (J. Tilmant,pers. comm.) or during our censuses. We also examined a number of keylime trees for larvae during each month of the 1979-1980 censuses, but wenever found H. andraemon larvae. We believe that the specimens reportedby Brown (1973a, b) failed to establish a breeding colony in Florida.
There appears to be an inconsistency in identification in Brown's papers

284 Florida Entomologist 67 (2) June, 1984
(Brown 1973a: 139, and Brown 1974: 11). Recounting what seems to be thesame experience, Brown (1973a) stated "on one occasion a P. aristodemusponceanus eluded our pursuit and flew safely from the south end of one keyand entered the jungle on the northeast side of an adjacent key in a routethat covered over one-half mile," while in Brown (1974), he stated inidentical wording that H. andraemon bonhotei eluded capture. Both Brown(1973a, 1974) and Covell (1977) use this incident to show that H.aristodemus ponceanus can fly between islands; however, it would seem thatthe identity of the species is unclear.
Based upon the results of our work and of others, we do not believethat sufficient evidence exists to establish the permanent residency of theBahaman swallowtail in the United States. The Bahaman swallowtail doesnot appear to be in danger of extirpation in its native Bahaman Islands,and it has become numerous in Jamaica where it was recently introduced(Tyler 1975). Under the 1978 amendments to the Endangered Species Actregarding invertebrates, this subspecies should not be listed unless it can beshown to be in danger throughout a significant portion of its range. Untilthe existence of established breeding populations in this country can beverified, it must be assumed that the occurrence of this butterfly in theUnited States is sporadic.
SCHAUS SWALLOWTAIL
The Schaus swallowtail is adapted to life within the shady tropical hammocks of the Biscayne Bay keys and upper Florida Keys. Its normal population size appears to be low in all life stages, although the numbers ofadults may follow a cyclical pattern of unknown periodicity. Large numbersof adults were present throughout its known range on the Biscayne Baykeys from 1969-1972 (Brown 1973, Covell and Rawson 1973). The population levels were low during the next nine years but rebounded in 1982 and1983.
In both 1979 and 1980, adults occurred in small numbers along thetransects. The slightly lower numbers observed in 1980 do not necessarilyrepresent a decline but may be an artifact of the census method. Theemergence season in both years extended from late April to June and corresponded well with the times of emergence reported by Covell and Rawson(1973) and Brown (1973).
In 1981, the number of Schaus swallowtails recorded along the transectswas lower than in 1979 and 1980 (Table 1). This decrease may have beenthe result of high larval mortality in 1980 (Morrison 1981) or could havebeen related to the severe spring dry season of 1981. The 1981 data indicatethat spring droughts may affect the butterfly and its larval food plant bydelaying emergence and new leaf production, respectively. On 13 May 1981,torchwood trees were just beginning to produce new leaves and many treesshowed no new leaves at all, while in previous years, most torchwood treeswere fully flushed with new growth by late April. A coincident delay in theemergence of Schaus swallowtail adults occurred in 1981. The first adultswere recorded on 13 May 1981 and not on earlier censuses in late April andearly May. The last sighting of adults occurred in mid-July 1981. The emergence period of the Schaus swallowtail appears to have been delayed byseveral weeks in 1981, coinciding with the delayed flush of new leaves by

Loftus & Kushlan: Schaus Swallowtail on Biscayne Bay 285
torchwood. Sightings in July 1981, several weeks later than the final sightings in 1979 and 1980, suggest that the entire emergence season was retarded in 1981. Other lepidopteran species along the transects showed similar delays in emergence in 1981 when compared with the previous two years(D. Peters, pers. comm.). It appears that the severe spring dry season mayhave been responsible for the retardance of emergence in 1981, but thisrelationship requires much more study.
The number of adult swallowtails increased greatly in 1982 (Table 1).RaiJ1lfall during the winter and spring of 1982 was relatively high (Fig. 2)which may have contributed to the greater survival of individuals. Adultemergence began in early May, but did not extend into June, resulting in avery short flight season. Numbers of other butterfly species on the islandswas also high during 1982 (D. Peters, pers. comm.).
The factors leading to changes in abundance are not well understood, butCovell (1976) suggested that severe droughts may have deleterious effects onthe timing of emergence and numbers of adult Schaus swallowtails. There isevidence that the pupae can overwinter for two years during a severedrought (Covell 1976), a capability which may have been an importantevolutionary factor for this species (Morrison 1981). Droughts occurnaturally in southern Florida and, even during relatively wet years, thereis a period in winter and early spring when rainfall is low (Fig. 1). Itappears that H. aristodemus ponceanus emergence coincides with thebeginning of leaf flush in the larval host plant, torchwood. Our census results indicate that rainfall during the months immediately preceding theusual emergence time may affect both the timing and duration of emergenceand the number of adults that will emerge. This relationship had beenpreviously suggested by Covell (1976), Miller (1975), and was discussed byMorrison (1981). In 1981, total rainfall during the period of Februarythrough April (80.7 mm) was the lowest recorded for the four survey years(Fig. 2), and the number of Schaus swallowtail adults was correspondinglylow on the transects. In addition, the timing of emergence was delayed andthe entire flight season was retarded. Rainfall from February through Aprilof 1982 (337.6 mm) exceeded that of any similar period during the foursurvey years (Fig. 2), and butterfly numbers were also the highest recorded. Similarly, in February through April in 1983, when rainfall totaled396.5 mm, limited censusing data showed high numbers of adults present inMay.
We usually observed fresh herbivory on torchwood leaves within one ortwo weeks after the emergence of adult Schaus swallowtails. The adultspresumably mate and lay eggs soon after emerging. From our lack of results in locating larvae, it appears that few larvae are either produced orsurvive, and those that do survive through several instars often disappearbefore pupating (Morrison 1981). Much of the life cycle of the Schausswallowtail is spent in the pupal stage, in which it can remain in diapausefor up to two years (Grimshawe 1940).
Covell (1977) discussed the possibility of a second emergence of Schausswallowtail adults in late summer. The latest sighting of adults during fouryears of censusing occurred in mid-July 1981, even though censuses werecontinued into August or September. Food for early instar larvae producedby a second emergence would probably be scarce because they feed on new

286 Florida Entomologist 67 (2) June, 1984
leaves which are primarily produced in spring (Morrison 1981). This sourcewould be less available later in the year. It seems more likely that the lateseason records of adults represent sporadic emergences, possibly resultingfrom asynchronous termination of diapause.
Several natural and human factors, such as freezes, insecticides, andcollecting, may affect Schaus swallowtail populations (Covell 1976). Suchfactors are more serious on the upper Florida Keys than on the islands westudied. Collecting, destruction of habitat, and insecticide spraying areminimal on the islands in Biscayne National Park; however, good habitatoutside the park, especially on Key Largo, is constantly being degraded byaerial mosquito spraying or lost through destruction of hammock for development. The amount of collecting outside of the park is unknown, butpoaching inside the park is discouraged by the isolation of the keys and thepatrolling by park rangers during the emergence period.
Freezes do not seem to be as damaging on the islands as they are on themainland. During the hard freeze of 1977, no adverse effects on vegetationor wildlife occurred on the islands in Biscayne Bay because of the moderating effect of the surrounding waters (J. Tilmant, pers. comm.). Hurricanescan severely affect vegetation and insect populations in south Florida(Grimshawe 1940; Craighead 1971). A direct pass of a hurricane over theBiscayne Bay keys could devastate the remaining population of the Schausswallowtail through wind damage, flooding, and reduced availability of thefood plant for several seasons. Such an event could extirpate the Floridasubspecies because its geographical range is now limited, and because populations on the upper Florida Keys from which recolonization might occurhave been severely reduced.
The butterflies on the islands of Biscayne Bay, located in BiscayneNational Park, appear to form the nucleus of the Schaus swallowtail population in Florida at this time. Little evidence of breeding on the majorFlorida Keys outside the park was found in 1980 (Morrison 1981), andrecent development pressures on northern Key Largo will result in thefurther diminution of suitable habitat. The islands in the park thereforerepresent the last undisturbed and unthreatened Florida habitat for thisinsect.
ACKNOWLEDGEMENTS
This survey was sponsored by The National Park Service, South FloridaResearch Center. Biscayne National Park personnel provided logistical support. We are very grateful to James Tilmant, Daniel Peters, Darcy Bailey,and Craig Hauke of Biscayne National Park for their cooperation in thisproject. Mr. Tilmant shared information on butterfly distribution and abundance on the keys in 1977 and 1978. Mr. Peters, Ms. Bailey, and Mr. Haukeaccompanied us on several surveys, provided data from 1981-1983 surveys,and supplied rainfall data. Dr. Charles V. Covell and others kindly offeredinformation on their observations of Schaus swallowtails on the keys. Weespecially thank Gerold Morrison for sharing his data on distribution andlarval mortality during 1980, and Oron Bass for reviewing this paper.
REFERENCES CITED
BROWN, L. N. 1973a. Populations of Papilio andraemon bonhotei Sharpe

Loftus & Kushlan: Schaus Swallowtail on Biscayne Bay 287
and Papilio aristodemus ponceanus Schaus (Papilionidae) in Biscayne National Monument, Florida. J. Lepid. Soc. 27: 136-140.
---,. 1973b. Populations of a new swallowtail butterfly found in theFlorida Keys. Florida Nat. 46(2): 25.
---,. 1974. Haven for rare butterflies. National Parks and Conserv. Mag.July 1974: 10-13.
COVELL, C. V., JR. 1976. The Schaus swallowtail: a threatened subspecies?Insect World Dig. 3: 21-26.
---. 1977. Project Ponceanus and the status of the Schaus swallowtail(Papilio aristodemus ponceanus) in the Florida Keys. Atala 5: 4-6.
---, AND G. W. RAWSON. 1973. Project Ponceanus: A report on firstefforts to survey and preserve the Schaus swallowtail (Papilionidae)in southern Florida. J. Lepid. Soc. 27: 206-210.
CRAIGHEAD, F. C., SR. 1971. The trees of south Florida. Vol. 1. Universityof Miami Press, Coral Gables, Florida.
FLORIDA GAME AND FRESH WATER FISH COMMISSION. 1982. Draft recoveryplan for the Schaus Swallowtail Butterfly (Heraclides (Papilio)aristodemus ponceanus (Schaus» including recommendations forBahaman Swallowtail Butterfly (Heraclides (Papilio) andraemonbonhotei (Sharpe». Report to U.S. Fish and Wildlife Service, Atlanta, Georgia. 46 p.
GRIMSHAWE, F. M. 1940. Place of sorrow: the world's rarest butterfly andMatecumbe Key. Nature Mag. 33: 565-567,611.
HENDERSON, W. F. 1945a. Papilio aristodemus ponceana Schaus (Lepidoptera: Papilionidae). Ent. News 56: 29-32.
---. 1945b. Additional notes on Papilio aristodemus ponceana Schaus(Lepidoptera: Papilionidae). Ent. News. 56: 187-188.
KIMBALL, C. P. 1965. Lepidoptera of Florida. Arthropods of Florida andneighboring land areas. Vol. 1. Fla. Div. Plant Indust., Gainesville,Florida. 363 p.
KLOTS, A. B. 1951. Field guide to the butterflies. Houghton-Mifflin, Boston,Massachusetts. 349 p.
LOFTUS, W. F. AND J. A. KUSHLAN. 1982. The status of the Schaus swallowtail and the Bahama swallowtail butterflies in Biscayne NationalPark. National Park Service, South Florida Research Center ReportM-649, Homestead, Florida. 18 p.
MILLER, L. D. 1975. Threatened status for two butterflies? Field MuseumNat. Hist. Bull: 15-18,22.
---, AND F. M. BROWN. 1981. A catalogue/checklist of the butterflies ofAmerica north of Mexico. Lep. Soc. Mem. 2. 280 p.
MORRISON, G. 1981. Draft recovery plan for the Schaus Swallowtail(Papilio aristodemus ponceanus Schaus), with recommendations concerning the Bahaman Swallowtail (Papilio andraemon bonhoteiSharpe). Final Rept. to Florida Game and Fresh Water Fish Comm.,Tallahassee, .Florida. 23 p.
RUTKOWSKI, F. 1971. Observations on Papilio aristodemus ponceanus(Papilionidae). J. Lepid. Soc. 25: 126-136.
SCHAUS, W. 1911. A new Papilio from Florida, and one from Mexico(Lepid). Ent. News. 22: 438-439.
TYLER, H. A. 1975. The swallowtail butterflies of North America. Naturegraph Pub. Inc., Healdsbury, California. 192 p.

288 Florida Entomologist 67 (2) June, 1984
BUPRESTIDAE (COLEOPTERA) OF THEVIRGIN ISLANDS
MICHAEL A. IVIE AND RICHARD S. MILLER
Department of EntomologyThe Ohio State UniversityColumbus, OH 43210, USA
ABSTRACT
Nine species of Buprestidae are recorded from the Virgin Islands.Acmaeodera gundlachi Fisher, Trigonogya uniformis (Waterhouse), andChrysobothris wolcotti Fisher are recorded for the first time from the VirginIslands. All species are keyed, diagnosed and notes on synonomy, distribution, taxonomic history, and biology are provided. A generic key for larvaeis provided. Polycesta thomae Chevrolat is returned to synonymy withPolycesta porcata (Fabricius).
RESUMEN
Nueve especies de Buprestidae de las Islas Virgenes fueron registradas.Acmaedora gundlachi Fisher, Trigonogya uniformis (Waterhouse), yChrysobothris wolcotti Fisher fueron registradas por primera vez en lasIslas Virgenes. A todas las especies se les hicieron claves, se diagnosticarony se proveeron notas de sinonimos, distribucion, historia taxonomica ybiologia. Una clave generica para las larvas es dada. Polycesta thomaeChevolat es devuelta como sinonimo de Polycesta porcata (Fabricius).
The West Indian Buprestidae were last studied as a whole by Fisher in1925; they are poorly known due in large part to a scattered literature anda general lack of museum specimens. Concerning the Virgin Islands, misidentifications and lack of collecting have led to confusion about faunalcomposition. This study of the Virgin Island Buprestidae was undertakento report recently assembled material, clear up misidentifications, and provide a sound taxonomic framework for field biologists, taxonomists andother interested workers.
Knowledge of larval habits and other biological aspects of the VirginIsland species is largely incomplete. Larvae. of only 2 species included hereare known. Although most buprestids are wood borers in the larval stage,at least one Virgin Island species is suspected of being a leaf miner. Morefield work is needed to complete distributional and life history information.
Discussions of the origin and zoogeography of this group would bepremature, as distributions and phylogeny are poorly known. Heatwole andLevins (1972: 115) gave conclusive evidence that some buprestid larvae cansurvive for long periods in marine flotsam around the Puerto Rican Bank.Thus, larval transport by rafting may play an important role in maintaining the integrity of the more widespread species.
Specimens cited are deposited in various collections, indicated with thefollowing acronyms:
AMNH-American Museum of Natural History, New York, NY.BMNH-British Museum (Natural History), London, England.

Ivie & Miller: Buprestidae of the Virgin Islands 289
CNCI-Canadian National Collection of Insects, Ottawa, Canada.ICCM-Carnegie Museum, Pittsburgh, PA.MAIC-Michael A. Ivie, private collection, Columbus, OH.NMPC-Narodni Museum, Prague, Czekoslovakia.OSUC-Ohio State University Collection of Insects and Spiders, Colum-
bus,OH.PMNH-Peabody Museum of Natural History, New Haven, CT.RSMC-Richard S. Miller, private collection, Columbus, OH.TAMU-Department of Entomology, Texas A & M University, College
Station, TX.UCDC-Department of Entomology, University of California, Davis, CA.UZMD-Zoologisk Museum, Copenhagen, Denmark.VIER-Virgin Islands Ecological Research Station, St. John, V.I.VIES-Virgin Islands Cooperative Extension Service, St. Croix, V.1.VIFW-Virgin Islands Division of Fish and Wildlife, St. Croix, V.1.
KEY TO SPECIES (ADULTS)
1. Lateral edges of first visible abdominal tergite produced forward to contact metepimeron; metacoxal plate expandedmedially 2
1'. Hind coxa extending laterally between first abdominal tergiteand metepimeron 6
2 (1). Tooth on exterior surface of profemur absent, obtuse, orrounded; Length greater than 8 mm __ 3
2'. Tooth on exterior surface of profemur acute; length less than7 mm 4
3 (2). Pronotum with 3 to 5 distinct depressions on each side, lateralmargins obtusely angulate; eyes separated above by approximately the diameter of antennal scape; shining black; elytracostate Chrysobothris tranquebarica (Gmelin)
3'. Pronotum with an indistinct depression on each side behindanterior margin and a distinct one in front of anteriormostpoint of elytron, lateral margins evenly sinuate; eyes separated above by approximately the length of antennal scape;bright metallic green-blue with purplish maculations onelytra; elytra without costae Chrysobothris fisheriana Obenberger
4 (2'). Disk of pronotum with simple punctation; pronotum brightred Chrysobothris thoracica (Fabricius)
4'. Disk of pronotum transversly rugose-punctate; pronotumpurple to black 5
5 (4'). Apex of last visible sternite notched on each side [slight insome males (Fig. 2) J; pronotum purplish _____________________________________________________________________ Chrysobothris wolcotti Fisher
5'. Apex of last visible sternite not notched, truncate to sub-truncate; pronotum greenish black _____________________________ Chrysobothris thomae Kerremans
6 (1'). Elytra with coarse punctures arranged in distinct rows;length greater than 4 mm 7
6'. Elytra with fine punctures scattered on surface; length lessthan 4 mm ----------------------------------------____ --- 8
290 Florida Entomologist 67 (2) June, 1984
q
6
a8
Fig. 1-9. 1) Last visible sternite of Chrysobothris fisheriana Obenberger(sex unknown). 2) Same of C. wolcotti Fisher (female). Same of C. tranquebarica Gmelin 3) male, 4) female. 5) Profemur of C. tranquebarica. 6)Same of C. thoracica (Fabricius). 7) Same of C. fisheriana. 8) Same ofC. wolcotti. 9) Brachys thomae Fisher, habitus.

Ivie & Miller: Buprestidae of the Virgin Islands 291
7(6). Scutellum not visible; tarsal claws dentate; elytra marked withyellow (Fig. 11) Acmaeodera gundlachi Fisher
7'. Scutellum visible; tarsal claws simple; black with metallicreflections ,----------------------------- Polycesta porcata (Fabricius)
8(6'). Head strongly narrowed by antennal cavities; elytron withlongitudinal carina extending from prominent humerus to nearapex (Fig. 9) ; flattened dorsally Brachys thomae Fisher
8'. Head not narrowed by antennal cavities; elytra withoutcarinae (Fig. 10); convex dorsally __ ______________________________________ . Trigonogya uniformis Waterhouse
PROVISIONAL KEY TO GENERA (LARVAE) EXCEPT Trigonogya*(Adapted from Burke 1917)
1. Leaf miners, body wedge-shaped, gradually narrowing fromfirst segment to twelfth _ - - ---- ._______ Brachys
1'. Twig, bark or wood borers; with wide, flattened head; ab-domen distinctly narrower than the thorax . ._______________ 2
2 (1'). Dorsal plate of prothorax marked with an inverted Y, V, orU . . Chrysobothris
2'. Dorsal plate of prothorax bearing only a single median im-pression or groove _... . .. . 3
3(2'). Head with a dark, sunken spot laterad of each mandible; ven-tral plate laterally with longitudinal corrugations ._. Polycesta
3'. Head not as above, ventral plate not corrugated ____ Acmaeodera
Acmaeodera gundlachi Fisher(Fig. 11)
Acmaeodera gundlachi Fisher 1925: 45. Wolcott 1936: 214; 1941: 88; 1951:277. Blackwelder 1944: 307. Ramos 1946: 34. Martorell 1976: 221, 228.
This species is variable in size and color pattern as noted by Fisher(1925: 45). The Anegada specimens are within the range of variation ofthe type series.
DIAGNOSIS: This species can be separated from all others of the familyknown from the Virgin Islands by the absence of a visible scutellum (Fig.11). From other West Indian Acmaeodera it can be recognized by itsslender form, the 4th antennal segment not serrate, and the pronotum widestat base and no wider than the elytra.
BIOLOGY: Specimens have been taken on low vegetation and feeding onthe large yellow flowers of Urechites lutea and on flowers of Lantanainvolvucrata L. Wolcott [1936: 214, 1951: 227 (summarized by Martorell1976: 221, 228)] noted collections from the flowers of red mangrove, Rhizophora mangle L. at Martin Pena and on sugarcane, Saccharum ofjicinarumL. at Gminica, Puerto Rico. The larval habits of this species are -unknown.
DISTRIBUTION: Puerto Rico, Mona, Anegada.TYPE LOCALITY: Gminica, Puerto Rico, holotype in NMNH.MATERIAL EXAMINED: (The number of specimens in each series is in
italics.)
*The larvae of Trigonogya are unknown.
292 Florida Entomologist 67 (2) June, 1984
Fig. 10-11. 10) Trigonogya uniformis (Waterhouse), habitus. 11)Acmaeodera gundlachi Fisher, habitus.
Puerto Rico: Holotype and paratypes from Gminica, Anasco and otherlocalities (NMNH).
Anegada: 20-23-VIII-1980, M. A. Ivie, on flowers of Urechites lutea(L.) Britt. (Apocynaceae) (15 MAIC, 4 UCDC, 1 VIES). 1-Setting Point,09-VIII-21-IX-1975, R. K. Butlin (specimen reported in unpublished "Report of the Cambridge Anegada Expedition", R. P. Dunne and B. E. Brown,1975) (BMNH). 20-Setting Point, 20-30-VIII-1982, on flowers of Lantanainvolucrata L. (Verbenaceae), R. S. Miller; 1-ibid. on flowers of Urechiteslutea (L.) Britt. (Apocynaceae) (16 RSMC, 5 aSUC).
Brachys thomae Fisher(Fig. 9)
Brachys thomae Fisher 1925: 180. Blackwelder 1944: 333.

Ivie & Miller: Buprestidae of the Virgin Islwruls 293
The inclusion of this species in the Virgin Island fauna is questionable.Fisher copied the label data of the unique holotype (La Belle, St. Thomas)and assumed it referred to St. Thomas, Virgin Islands. No La Belle wasencountered by M. A. Ivie in 2 years of residence on St. Thomas, and nosuch locality is listed in the U. S. Board of Geographic Names Gazetteer,1958, for the Virgin Islands. Gazetteers for other areas do show that LaBeLIe or Belle is a common name in the West Indies, occurring on Jamaica,Barbados and Hispaniola. It also occurs in Guyana, Trinidad, and Florida.On both Jamaica and Barbados, a Belle occurs in St. Thomas Parish. Fisher(1924: 4) noted that the buprestid material he borrowed from the CarnegieMuseum was from the Isle of Pines and Jamaica.
The only other record of Brachys from the West Indies is that of B.aerQsa Melsheimer from Jamaica (also common in the United States)(Blackwelder 1944: 332). Until more material of B. thomae becomes available, its true distribution is in doubt.
DIAGNOSIS: This species is distinguished by its small size (3 mm) incombination with its deeply grooved prosternal process and the lateralcarinae on the pronotum and elytra (Fig. 9).
BIOLOGY: Unknown. Other members of this genus are leaf miners in thelarval stage.
DISTRIBUTION: St. Thomas?TYPE LOCALITY: La Belle, St. Thomas (unique holotype in ICCM).MATERIAL EXAMINED:St. Thomas: La Belle (holotype, ICCM).
Chrysobothris fisheriana Obenberger(Fig. 1,7)
Chrysobothris fisheriana Obenberger 1928: 299 rJisheri pI. 6., Fig. 20 (lapsuscalami). Hoscheck 1931: 158. Obenberger 1934: 663. Blackwelder 1944:317.Obenberger's original description carried the name jisheriana in the text
and fisheri on the plate. The label on the type reads "typus, ChrysobothrisFisheri m. Type = Fisheriana m. n. n., det. Dr. Obenberger." Thus, it ap~
pears that Obenberger originally intended that fisheri be the name for thisspecies. Under article 32b of the International Code of Zoological Nomenclature it is up to the first revisor to choose between multiple spellings in anoriginal description. This species has not been included in a revision and thistreatment is certainly not one. However, Chrysobothris fisheri is preoccupied by a TMry (1927) species from Cuba. To choose fisheri for theObenberger species would unnecessarily complicate the nomenclature. Therefore, we have followed previous authors (including Obenberger 1934) inusing fisheria1UL for this species.
The unique type, represents a species quite unlike any other we haveseen from the West Indies. Fearing to destroy information for a futurerevisor, we did not dissect the specimen, and its sex is unknown.
DIAGNOSIS: The shape of the last visible sternite (Fig. 1) will separatethis species from all others in the Virgin Islands, and possibily the WestIndies. The shape of the profemur (Fig. 7) and the brilliant green pronotumfurther distinguish it.
BIOLOGY: Unknown.

294 Florida Entomologist 67 (2) June, 1984
DISTRIBUTION: St. Thomas.TYPE LOCALITY: St. Thomas, holotype in NMPC.MATERIAL EXAMINED:St. Thomas: holotype (NMPC).
Chrysobothris thomae Kerremans
Chrysobothris thomae Kerremans 1899: 337. Fisher 1925: 118. Blackwelder1944: 318.DIAGNOSIS: In addition to the characters in the key, this species can be
distinguished by the presence of significant areas of true red on the elytra.BIOLOGY: Unknown.DISTRIBUTION: St. Thomas.TYPE LOCALITY: St. Thomas, 2 syntypes in BMNH.MATERIAL EXAMINED: No specimens of this species were seen. The key
and diagnosis were based on a series of SEM photos and color transparenciesof the syntypes.
Chrysobothris thoracica (Fabricius)(Fig. 6)
Buprestis thoracica Fabricius 1798: 138.Chrysobothrus thoracica Castelnau and Gory 1837: 96. Fisher 1925: 123.
Blackwelder 1944: 318.The Virgin Island material reported here was compared with a specimen
in the NMNH from Gmlnica, Puerto Rico, determined by W. S. Fisher.Fisher (1925: 125) reported a specimen in the BMNH from St. Thomas.For a complete synonymy of the early literature on this species see Fisher(1925: 125).
DIAGNOSIS: This species can be recognized by an acute spine on theprofemur (Fig. 6) in combination with a red pronotum.
BIOLOGY: The Anegada specimens were beaten from Croton sp. and whitemangrove [Laguncularia racemosa (L.) Gaertn].
DISTRIBUTION: Puerto Rico, St. Thomas, St. John, Anegada, St. Croix.TYPE LOCALITIES: Of thoracica, St. Thomas, 3 cotypes in the UZMD,
(Zimsen 1964: 157).MATERIAL EXAMINED:St. Thomas: I-Mus. Weterm. (UZMD).St. John: 4-June 1826, Stamm. colI. (UZMD).Anegada: I-Flamingo pond, 22-VIII-1980, M. A. Iviej 4-23-VIII-1980,
M. A. Ivie (2 MAIC, 1 RSMC, 1 VIES).St. Croix: 1-Frederiksted, 25-1-1977, J. A. Yntema (NMNH).No specific island: I-Vest Indien, Mus. Hauschild; I-Ind. orient, Mus.
Drews (UZMC).
Chrysobothris tranquebarica (Gmelin)(Fig. 3, 4, 5)
Buprestis impressa Fabricius 1787: 182 (not Fabricius 1775).Buprestis tranquebarica Gmelin 1788: 1932 (replacement name).Chrysobothris tranquebarica, Kerremans 1892: 214. Fisher 1918: 173, 1925:
96, 1942: 198. Snyder 1919: 155. Blackwelder 1944: 318. Paulian 1947:

Ivie & Miller: Buprestidae of the Virgin Islands 295
147. Benoit 1966: 330. Miskimen and Bond 1970: 86. Martorell 1976: 31,48, 110, 142, 221.This economically important species in one of the most common and
widespread buprestids in the West Indies. This has led to a large body ofliterature and several complicated synomymies and homonymies in theearly literature (for an account see Fisher 1918: 176 and 1925: 96). Since1925, the name tranquebarica has been uniformly applied to this species.Therefore, the citations included here are limited to those pre-1925 references of greatest importance and those since 1925 pertinent to the VirginIsland fauna.
The citation of Tranquebar for the type locality of this West Indianspecies is probably due to mislabelling. Fabricius received considerable material through members of the Danish Civil Service and Military (Zimsen1964: 11). Tranquebar, India, was a Danish colony in Fabricius' time, andit is possible that the type reached him through a contact that had servedboth in India and the Danish West Indies (now the U. S. Virgin Islands).Such material could easily have been mislabelled, or simply labelled withthe port from which it was shipped. In any case, the species is restricted tothe West Indies and south Florida.
This species was recorded from St. Thomas by Fisher (1925: 99) basedon a specimen in the BMNH. Miskimen and Bond (1970: 86) reported itfrom St. Croix. The Virgin Island specimens reported here were comparedwith 2 specimens from Puerto Rico (NMNH), determined by G. B. Vogt.
DIAGNOSIS: This species can be recognized by its large size (10 mm),the rounded or obtuse tooth on the profemur (Fig. 5), the angulate pronotalmargins, by usually having 8-10 depressions on the blackish pronotum, andthe shape of the last visible sternite (Fig. 3, 4).
BIOLOGY: The life history was studied by Snyder (1919), who illustratedall life stages of this species [Benoit (1966: 330) further described thelarva]. Snyder reported it as an economic pest of the introduced Australianpine (Casuarina equisetifolia Forst.) and of the red mangrove (Rhizophoramangle L.). Chamberlin (1926: 174) recorded it from buttonwood (Conocarpus erectus L.), a record verified by material from Key Largo, Florida(R. L. Westcott, pers. com.). Martorell (1976: 31, 48, 110, 142, 221) summarized Puerto Rican host records, adding gree-gree (Bucida buceras L.),Eucalyptus sp. and Inga vera Willd. to the above. Miskimen and Bond(1970: 86) reported it from St. Croix in year-round, sub-economic levelson Australian pine. M. A. Ivie reared one specimen from red mangrove anda series from a limb of a fallen tree on St. Thomas (see material examined).Four tachinid flies [Zelia (Minthozelia) sp., det. D. M. Wood, specimens inthe CNCI] were reared from this limb, presumably they were parasitic onC. tranquebarica.
DISTRIBUTION: Florida, Bahamas, Cuba, Hispaniola, Puerto Rico, St.Thomas, St. John, St. Croix, Guadeloupe, Grenada [South American localities for this species (Blackwelder 1944: 318) belong to Halecia impressa(Fabricius, 1775) see Fisher 1918: 175J.
TYPE LOCALITY: "Tranquebariae" [India].MATERIAL EXAMINED:St. Thomas: 2-VI-[1826], Stamm. colI. (UZMD). 1-Est. Nazareth,
National Park HQ, 27-VII-1980, emerged 21-VIII-1980 from red mangrove,

296 Florida Entomologist 67 (2) June, 1984
M. A. lvie. (MAIC). 49-Magens Bay Arboretum, collected as larvae 30VII-1980, emerged 08-20-VIII-1980, M. A. Ivie. (22 MAIC, 10 asuc, 10UCDC, 5 RSMC, 2 VIES). LARVAE: 9-ibid. (MAIC). PUPAE: 1-ibid.(MAIC).
St. John: 1-VIII-1968, A. E. D[ameon] (VIER).
Chrysobothris wolcotti Fisher(Fig. 2, 8)
Chrysobothris wolcotti Fisher 1925: 119. Wolcott 1941: 89, 1951: 279. Blackwelder 1944: 319. Martorell 1976: 153.The St. Thomas specimen reported below was compared with a Puerto
Rican specimen from Mayaguez (AMNH). The shape of the last visiblesternite and the color of the elytra match quite well, although the St.Thomas specimen is significantly smaller than the Puerto Rican one. TheTortola specimen is more within the normal size range.
DIAGNOSIS: This species can be distinquished from the other VirginIsland species by the notched last abdominal sternite (Fig. 2) ; the purplish,transversly rugose pronotum, and the toothed profemur (Fig. 8).
BIOLOGY: Wolcott (1941: 89) reported all life stages in crepe myrtle(Lagerstroemia indica L.) in Puerto Rico. The St. Thomas specimen reported above was found dead in an emergence tunnel in a limb of a deadshrub ca. 2 m tall.
DISTRIBUTION: Puerto Rico, St. Thomas, Tortola.TYPE LOCALITY: Puerto Rico, holotype in NMNH.MATERIAL EXAMINED:St. Thomas: 1-Smith Bay, 08-VIII-1980, ex gallery in dead branch,
M. A. Ivie (fragmented) (MAIC).Tortola: 1-Green Bank, 900 ft., 21-III-1974, C. L. Remington (PMNH) .
. Polycesta porcata (Fabricius)
Buprestis porcata Fabricius 1775: 219.Polycesta porcata, Kerremans" 1906 : 516. Fisher 1925: 19. Wolcott 1941:
88; 1951: 275, Fig. p. 276. Blackwelder 1944: 306. Martorell 1948: 316,1976: 251. Cobos 1981: 71, Fig. 82, 94.
Polycesta thomae Chevrolat 1838: 55. Fisher 1925: 28. Blackwelder 1944:306. Beatty 1944: 137. Ramos 1946: 34. Wolcott 1951: 276. Miskimenand Bond 1970: 87. Martorell 1976: 48. Cobos 1981: 71. RETURNED TOSYNONOMY.
Polycesta nr. depressa, Miskimen and Bond 1970: 87 (not Linnaeus 1771).For references prior to 1925 see Fisher (1925).The nomenclatural status of West Indian Polycesta is unsettled (Cobos
1981: 68) and will remain uncertain until more material is assembled anda complete revision of the genus undertaken. The modern taxonomy of thisgenus requires males for authoritative determination; males can be distinguished from females in this species by a densely setose patch on the 1stvisible abdominal sternum. Unfortunately the male genitalia of many ofthe nominate West Indian taxa remain unstudied.
The first Virgin Island record of Polycesta dates to the description ofP. thomae Chevrolat from a female specimen in 1838. The St. Thomasrecords for P. porcata were based on the synonymy of P. thomae with P.

Ivie & Miller: Buprestidae of the Virgin Islands 297
porcata by Kerremans (1906: 516). Fisher (1925: 27) reinstated thornaeas a valid species, and reported (1925: 28) 2 additional St. Thomas specimens, both females, under that name, one in the BMNH, the other in theAMNH. Beatty (1944: 137) recorded this species from St. Croix (asthornae), as did Miskimen and Bond (1970: 87, as P. nr. depressa, vouchera female, det. "D.M.W." in NMNH). Cobos (1981: 71) expressed uncertainty of the validity of P. thornae, but lacked males to settle the issue.
The material assembled for this study includes several males. Thegenitalia of the Anegadan male matches Cobos' illustration (1981: Fig. 94)of that of typical porcata. This specimen also has the scutellar costae welldeveloped, as used by Fisher to distinguish porcata from thornae. However,2 females collected from the same shrub by R. S. Miller lack this character,though there can be little doubt of their conspecificity. Most other VirginIsland specimens lack scutellar costae. Males exhibit considerable variationin the form of the base of the lateral lobes of the genitalia, though the St.Croix male is somewhat intermediate between the well developed Anegadanspecimen and the lesser so St. Thomas, Saba, and St. John specimens. Thecharacters used by Fisher to reinstate thornae: costae, punctation, bodywidth and color are all variable in our series. Barr (1949) notes variationin all these characters in the North American species. He also notes thatthe genitalia are slightly variable in Polycesta, as is apparent in our materiaL
DIAGNOSIS: This species can be recognized from all others in the VirginIslands by the characters in the key. From other Polycesta, it is best separated by the form of the median lobe of the male genitalia (Cobos 1981:Fig. 94). Habitus illustrations of this species are given by Wolcott (1951:275) and Cobos (1981: Fig. 82).
BIOLOGY: Wolcott (1941: 88) reported a specimen reared from Dominican mahogany [Swietenia rnahagony (L.) Jacq. (Meliacae)]. Ramos (1946:34) collected larvae, pupae and adults from dead stems of Australian pine(Casuarina equisetifolia). Miskimen and Bond (1970: 87) report this species on sea grape Coccoloba uvifera (L.) L. Whether the specimen wasreared from the plant or the adult was collected on it is unmentioned, thelabel data is simply "attacking sea grape". A labelled pin in the NMNHstates "P. thornae in Elaeodendrurn xylocarpurn, 'nothing nut', Sandy CayBeach, British Virgin Islands, VIII 1972, Th. FlavelL" The larvae andpupae reported were found in the bole of a dead shrub, just below thefirst fork. One was reared to adulthood for confirmation of the identification.
DISTRIBUTION: Hispaniola, Mona, Puerto Rico, Vieques, St. Thomas, St.John, Little Saba (V. S. Virgin Islands), Grass Cay (V.S.V.I.), NormanIs. (British Virgin Islands), Sandy Is. ? (B.V.I.), Anegada, St. Croix.
TYPE LOCALITY: Of porcata, "Oriente Forskahl" (mislabeled ?), 2syntypes in VZMD Copenhagen (Zimsen 1964: 153). Of thornae, St.Thomas, holotype in BMNH (Fisher 1925: 28).
MATERIAL EXAMINED:St. Thornas: 6 (1 t )-R. H. Stamm. colI.; 1-0rsted; 1-1854, H. Krebs
(VZMD). 1-VII-1915 [C. Shoemaker] (AMNH). 1-"46-84",. receivedfrom BMNH 1923; 1-24-VII-1965, R. Delgado; 1-20-III-1970, R. Cole(NMNH). 1 (t )-Frenchmans Bay Estate, 01-III-1978, M. A. Ivie; 3(2 t )-Smith Bay, 04-VIII-1980, M. A. Ivie, reared from larvae (1 pair

298 Florida Entomologist 67 (2) June, 1984
MAIC,l (; OSUC). LARVAE: 2-ibid. (MAIC). PUPA: 1-ibid. (MAIC).Little Saba: 1 «; )-24-III-1979, M. A. Ivie; 3-05-VI-1980, C. A.
Jennings (MAIC).Grass Cay: 1-19-IV-1971, R. Wharton (TAMU).St. John: 1-V. 1. National Park, Lameshur Bay, 17-VI-1968, R. M.
(VIER); 1 «(; )-V. 1. National Park, Cruz Bay, 1970, A. E. D[ameon](NMNH).
Norman Is.: 1-03-VI-1979, M. A. and L. L. Ivie (MAIC) (fragmented).A negada : 3 (1 d' )-Setting Point, 21-24-IV-1983, R. S. Miller (1 pair
RSMC, 1 MAIC).St. Croix: 2-Mus. Westerm. (UZMD). 1-Christianstead, rec. 19-XI
1941, H. A. Beatty; 1-Cane Bay, No. 1508, 23-VII-1960, attacking seagrape, Cocolobus sp. [sic]; 1 (d' )-Estate Canaan, elv. 120 m., Ol-VI1980, W. 1. Knaussenberger (NMNH). 1-Golden Grove, CVI, 14-XI-1979,W. 1. Knaussenberger (VIES).
No specific island: 1-Ins. Americ., Mus. Drews; 1-Vestindien, F. Riise;3-Ins. Americ., Mus. S[ehested and] T[lmder] L[und] (UZMD).
Trigonogya uniformis (Waterhouse)(Fig. 10)
Mastogenius uniformis Waterhouse 1896: 105. Fisher 1925: 203. Blackwelder 1944: 34l.
Trigonogya uniformis, Fisher 1949: 35l.We were able to compare the single Anegada specimen with the unique
holotype from Grenada. The Virgin Island specimen differs in the pronotumbeing broader, with the lateral margin extending to the anterior margin,the hypomeron more convex, the transverse depression at the base of theelytra less deep, and in the shape of the clypeus and labrum. With only 2specimens, we were unable to evaluate the possibility of an undescribedspecies vs. intraspecific variation. The sex of both specimens is unknown.Due to the rarity of specimens, it was considered undesirable to dissectthem, to avoid the possibility of destroying data for a future revisor. Untilmore material becomes available from both localities, as well as the intervening 800 km, it seems best to treat them as conspecific.
DIAGNOSIS: This species can easily be separated from other VirginIsland species by its size (2 mm) and its lack of elytral costae (Fig. 10).
BIOLOGY: Unknown. The Al1egada specimen was collected by sweepinglow vegetation. The related genus Mastogenius is usually collected by beating various trees and shrubs, usually on portions containing dead twigs(R. L. Wescott, pers. com.).
DISTRIBUTION: Grenada, Anegada.TYPE LOCALITY: Mount Gay Estate, leeward side, Grenada, West Indies.
Unique holotype in the BMNH.MATERIAL EXAMINED:Anegada: 1-23-VIII-1980, M. A. Ivie (MAIC).Grenada: holotype (BMNH).
ACKNOWLEDGEMENTS
We would like to thank the following people for the loan of material:L. Herman (AMNH), C. M. F. von Hayek (BMNH), G. Ekis (ICCM),

Ivie & Miller: Buprestidae of the Virgin Islands 299
S. BUy (NMPC), J. M. Kingsolver (NMNH), D. G. Furth (PMNH), S. J.Merritt (TAMU), O. Martin (UZMD), W. I. Knaussenberger (VIES),J. Ynetma (VIFW), J. Miller, Virgin Islands National Park, St. Thomas(for material in VIER). C. A. Jennings, Starkville, Mississippi, providedvaluable material. Special thanks to Mrs. E. Peacock (BMNH) for providing an excellent series of SEM photos and color transparencies of typematerial. We are grateful to D. M. Wood (CNCI) for the identification oftachinid parasitoids, and to Dr. W. G. D'Arcy, Missouri Botanical Garden,St. Louis, for identifying photos of floral hosts of Acmaeodera gundlachifrom Anegada. To Dr. C. A. Triplehorn, Ohio State University, and Dr.R. L. Westcott, Oregon Department of Agriculture, Salem, go our thanksfor their helpful reviews of an earlier draft of the manuscript.
REFERENCES CITED
BARR, W. F. 1949. A revision of the species of the genus Polycesta occurring in the United States (Coleoptera, Buprestidae). AmericanMus. Novit. (1432): 1-42.
BEATTY, H. A. 1944. Fauna of St. Croix, Virgin Islands. J. Agric. Univ.Puerto Rico 28: 103-85.
BENOIT, P. 1966. Descriptions of Chrysobothris larvae (Coleoptera:Buprestidae) occurring in the United States and Mexico. CanadianEnt. 98: 324-30.
BLACKWELDER, R. E. 1944. Checklist of the coleopterous insects of Mexico,Central America, the West Indies, and South America. Part 2. UnitedStates Nat. Mus. Bull. 185: 189-341.
BURKE, H. E. 1917. Flat-headed borers affecting forest trees in the UnitedStates. United States Dept. Agric. Bull. 437: 1-8.
CASTELNAU, F. L., AND H. L. GORY. 1837. Historie naturelle et iconographiedes insectes coleopteres, publier par monographies, separees, vol. 1(various pagination). Paris.
CHAMBERLIN, W. J. 1926. The Buprestidae of North America exclusive ofMexico. A Catalog. 289 p. + 1 p. index. Corvallis, OR.
CHEVROLAT, L. A. 1838. Descriptions de trois buprestides et dun superbeCyphus nouveaux. Silbermann's Rev. Ent. 5: 54-5.
COBOS, A. 1981. Ensayo sobre los generos de la subfamilia Polycestinae(Coleoptera, Bl1nrestidae), Parte 2. Eos, Revista Espanola de Entomolgia [1979] 55-56: 23-94.
FABRICIUS, J. C. 1775. Systema Entomologiae. 30 + 832 p. Lipsiae.---. 1787. Mantissa Insectorum 1: 1-348. Hafinae.---. 1798. Supplementum Entomologiae Systematicae. 572 p. Hafniae.FISHER, W. S. 1918. Chrysobothris tranquebarica Gmel. versus impressa
Fabr. (Coleoptera, Buprestidae). Proc. Ent. Soc. Washington 20:173-7.
---. 1925. A revision of the West Indian Coleoptera of the familyBuprestidae. United States Nat. Mus. Proc. 65: 1-207.
---. 1942. A revision of the North American species of buprestidbeetles belonging to the tribe Chrysobothrini. United States Dep.Agric. Misc. Publ. 470: 1-275.
---. 1949. New buprestid beetles from Mexico, Central and SouthAmerica, and the West Indies. Proc. United States Nat. Mus. 99(3240): 327-51.
GMELIN, J. F. 1788 [1790]. Linne's Systema Naturae... 13 ed. 1: 1517-2224.HEATWOLE, H., AND R. LEVINS. 1972. Biogeography of the Puerto Rican
Bank: Flotsam transport of terrestrial animals. Ecology 53: 112-7.

300 Florida Entomologist 67 (2) June, 1984
HOSCHECK, A. B. 1931. Beitrage zur kenntnis der Buprestiden (Col.) 3.Mitt. Zoo!' Mus. Berlin 17: 133-64.
KERREMANS, C. 1892. Catalogus synonymicus des buprestides descrits de1785 a 1890. Mem. Soc. Entomol. Belgique 1: 1-304.
---. 1899. Contribution it l'etude de la faune intertropicale americaine.Buprestides. Ann. Soc. Ent. Belgique 43: 329-67.
---,. 1906. Monographie des buprestides. 1. 534 p. Paris.LINNAEUS, C. 1771. Pages 521-52 In: Mantissa Plantarum II. Holmiae.MARTORELL, L. F. 1948 [1945]. A survey of the forest insects of Puerto Rico.
Part 1. J. Agric. Univ. Puerto Rico 29: 69-354.---. 1976. An Annotated Food Plant Catolog of the Insects of Puerto
Rico. Agricultural Experiment Station, University of Puerto Rico.303 p.
MISKIMEN, G. W., AND R. M. BOND. 1970. The insect fauna of St. Croix,U. S. Virgin Islands. New York Acad. Sci. Scientific Survey of PortoRico and the Virgin Islands 13: 1-117.
OBENBERGER, J. 1928. Opuscula buprestologica 1. Arch. Naturgesch. 92:225-350.
---,. 1934. Buprestidae 3. Coleopterum Catalogus, Pars 132. 12: 569-781.PAULIAN, R. 1947. Famille des Buprestidae. Pages 140-53 In Fleutiaux
et al. Coleopteres des Antilles. Faune de I'Empire Fran~ais 7.RAMOS, J. A. 1946. The insects of Mona Island. J. Agric. Univ. Puerto
Rico 30: 1-74.SNYDER, T. E. 1919. Injury to casuarina trees in Southern Florida by the
mangrove borer. J. Agric. Research 16: 155-63.THERY, A. 1927. Buprestides nouveaux des Antilles. Ann. Soc. Ent. France
96: 31-6.WATERHOUSE, C. O. 1896. Observations on some Buprestidae from the West
Indies and other localities. Ann. Mag. Nat. Hist. Series 6. 18: 104-7.WOLCOTT, G. N. 1936. Insectae Borinquenses: a revised annotated check
list of the insects of Puerto Rico. J. Agric. Univ. Puerto Rico 20:1-600.
---. 1941. Supplement to Insectae Boringquenses. J. Agric. Univ. PuertoRico 25: 33-158.
---,. 1951 [1948]. The insects of Puerto Rico. Coleoptera. J. Agric. Univ.Puerto Rico 32: 225-416.
ZIMSEN, E. 1964. The Type Material of 1. C. Fabricius. 656 p. Munksgaard.
.. . . • • • • • • • • ••STRUCTURAL DETAILS OF
THE SCALES OF 28 SPECIES OF MOSQUITOES
KAMEL T. KHALAF AND RAMIZ K. KHALAFLoyola University
New Orleans, LA 70118 USA
ABSTRACT
Light photographs of scales on the wing and body of 28 species ofmosquitoes, which belong to 11 genera, were taken from impressions madeon triafol BN. The shape and striae of the scales were clearly discernibleand therefore could be utilized in routine taxonomy. The type of scalesfound on the wing, as well as on the scutum, varied among the differenttaxa. Those on the abdominal terga were obovate, or intermixed in some

300 Florida Entomologist 67 (2) June, 1984
HOSCHECK, A. B. 1931. Beitrage zur kenntnis der Buprestiden (Col.) 3.Mitt. Zoo!' Mus. Berlin 17: 133-64.
KERREMANS, C. 1892. Catalogus synonymicus des buprestides descrits de1785 a 1890. Mem. Soc. Entomol. Belgique 1: 1-304.
---. 1899. Contribution it l'etude de la faune intertropicale americaine.Buprestides. Ann. Soc. Ent. Belgique 43: 329-67.
---,. 1906. Monographie des buprestides. 1. 534 p. Paris.LINNAEUS, C. 1771. Pages 521-52 In: Mantissa Plantarum II. Holmiae.MARTORELL, L. F. 1948 [1945]. A survey of the forest insects of Puerto Rico.
Part 1. J. Agric. Univ. Puerto Rico 29: 69-354.---. 1976. An Annotated Food Plant Catolog of the Insects of Puerto
Rico. Agricultural Experiment Station, University of Puerto Rico.303 p.
MISKIMEN, G. W., AND R. M. BOND. 1970. The insect fauna of St. Croix,U. S. Virgin Islands. New York Acad. Sci. Scientific Survey of PortoRico and the Virgin Islands 13: 1-117.
OBENBERGER, J. 1928. Opuscula buprestologica 1. Arch. Naturgesch. 92:225-350.
---,. 1934. Buprestidae 3. Coleopterum Catalogus, Pars 132. 12: 569-781.PAULIAN, R. 1947. Famille des Buprestidae. Pages 140-53 In Fleutiaux
et al. Coleopteres des Antilles. Faune de I'Empire Fran~ais 7.RAMOS, J. A. 1946. The insects of Mona Island. J. Agric. Univ. Puerto
Rico 30: 1-74.SNYDER, T. E. 1919. Injury to casuarina trees in Southern Florida by the
mangrove borer. J. Agric. Research 16: 155-63.THERY, A. 1927. Buprestides nouveaux des Antilles. Ann. Soc. Ent. France
96: 31-6.WATERHOUSE, C. O. 1896. Observations on some Buprestidae from the West
Indies and other localities. Ann. Mag. Nat. Hist. Series 6. 18: 104-7.WOLCOTT, G. N. 1936. Insectae Borinquenses: a revised annotated check
list of the insects of Puerto Rico. J. Agric. Univ. Puerto Rico 20:1-600.
---. 1941. Supplement to Insectae Boringquenses. J. Agric. Univ. PuertoRico 25: 33-158.
---,. 1951 [1948]. The insects of Puerto Rico. Coleoptera. J. Agric. Univ.Puerto Rico 32: 225-416.
ZIMSEN, E. 1964. The Type Material of 1. C. Fabricius. 656 p. Munksgaard.
.. . . • • • • • • • • ••STRUCTURAL DETAILS OF
THE SCALES OF 28 SPECIES OF MOSQUITOES
KAMEL T. KHALAF AND RAMIZ K. KHALAFLoyola University
New Orleans, LA 70118 USA
ABSTRACT
Light photographs of scales on the wing and body of 28 species ofmosquitoes, which belong to 11 genera, were taken from impressions madeon triafol BN. The shape and striae of the scales were clearly discernibleand therefore could be utilized in routine taxonomy. The type of scalesfound on the wing, as well as on the scutum, varied among the differenttaxa. Those on the abdominal terga were obovate, or intermixed in some

Khalaf & Khalaf: Scales on Mosquitoes 301
genera with fewer larger scales. The number of striae on any type of scaleswas quite fixed for each species.
RESUMEN
Con el uso de impresi6nes simples en triafol BN junto con microscopiade luz, micrografias de buena resolucion fueron conseguidas de las escamasdel ala y del cuerpo de 28 especies de mosquitos, que pertenecen a 11 generos.Las formas de las escamas y sus estrias eran claramente discernibles conesta tecnica y por consiguiente podran ser utilizados en taxonomia rutinaria.Los tipos de escamas descubiertos en el ala, asi como en el escuto, variaronen differentes taxa. Esas en las tergas abdominales eran obovados, 0 entremezclados en algunos generos con menor numero de escamas mas grandes.La cantidad de estrias en cualquier tipo de escama fue bastante fijo encada especie.
A simple technique for preparing transparent impressions was used tostudy the cuticle surface (Khalaf 1980). The technique gave greater resolution than that obtainable by direct stereoscopic microscope examination.This method was used in the present investigations of the surface of theintegument of mosquitoes.
Theobald (1901, 1905) recognized the taxonomic value of scales ofvarious bodily parts of mosquitoes, and described and named many of theirshapes. He concluded that wing scales were important in the classificationof genera but such taxa are now only ranked as subgenera or mere speciesgroups. Apparently hampered by limited resolution, Theobald's descriptionswere often general and repetitive. Subsequent authors sparingly used scalesto differentiate taxa (Mattingly 1957-1965, Foote and Cook 1959). Christophers (1960) described the scales of Aedes aegypti (Linnaeus). Becausespecimen preparations for light and scanning electron microscopy arelengthy, the structural details of the scales have not been effectively appliedin the taxonomy of Culicidae. Only generalized features of the scales, suchas color, width, and length, have been used. Using our method, details ofscales in various regions of the body and wing can be easily distinguished,and hence can readi~ybe used in systematic analysis.
MATERIALS AND METHODS
Details of the method were described previously (Khalaf 1980). Impressions of scales were made on triafol BN (Mobay Chemical Corporation,Plastic and Coatings Division, Pittsburgh, PA) rectangles which weremoistened by a drop of acetone. After removing the specimen, the impressionwas examined and photographed under high magnification (X600).
In the present investigation, scales on the upper surfaces of the wing,scutum, and abdominal terga of females of 28 species were studied (Table1). Such scales from males of 9 species, as marked in Table 1, were similarlystudied.
Impressions were taken from 1 or 2 specimens of each species. Onlystriae of scales that produced an entire and flattened impression werecounted.
The scales varied in the different regions of the wing. This is also true
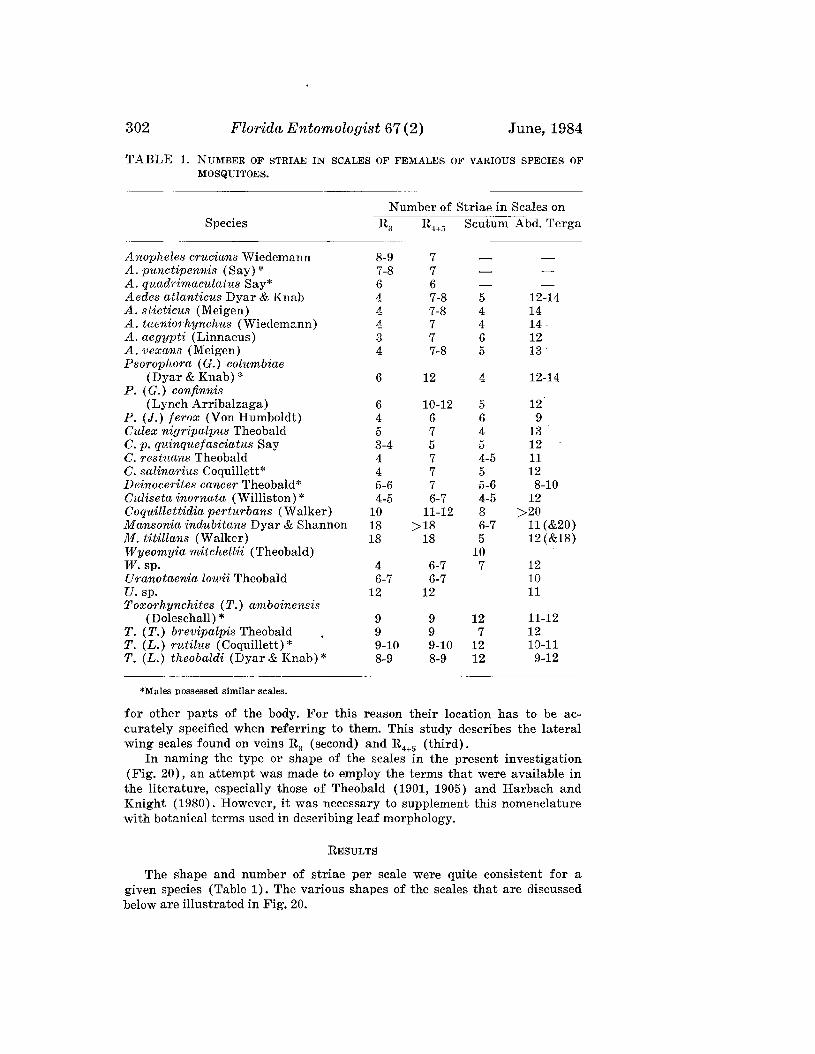
302 Florida Entomologist 67 (2) June, 1984
TABLE 1. NUMBER OF STRIAE IN SCALES OF FEMALES OF VARIOUS SPECIES OFMOSQUITOES.
SpeciesNumber of Striae in Scales on
R g R 4+5 Scutum Abd. Terga
Anopheles crucians WiedemannA. punctipennis (Say) ,;,A. quad1"imaculaius Say*Aedes atlanticus Dyar & KnabA. sticticus (Meigen)A. tcwniorhynchus (Wiedemann)A. aegypti (Linnaeus)A. vexans (Meigen)Psorophora (G.) columbiae
(Dyar & Knab) *P. (G.) confinnis
(Lynch Arribalzaga)P. (J.) ferox (Von Humboldt)Culex nigripalpus TheobaldC. p. quinquefasciatus SayC. restuans TheobaldC. salinarius Coquillett*Deinocerites cancer Theobald*Culiseta inornata (Williston)'~
Coquillettidia perturbans (Walker)Mansonia indubitans Dyar & ShannonM. titillans (Walker)lVyeomyia mitchellii (Theobald)lV.sp.Vranotaenia lowii TheobaldV.sp.Toxorhynchites (T.) amboinensis
(Doleschall) *T. (T.) brevipalpis TheobaldT. (L.) rutilus (Coquillett)*T. (L.) theobaldi (Dyar & Knab) *
*Males possessed similar scales.
8-9 77-8 76 64 7-84 7-84 73 74 7-8
6 12
6 10-124 65 73-4 54 74 75-6 74-5 6-7
10 11-1218 >1818 18
4 6-76-7 6-7
12 12
9 99 99-10 9-108-9 8-9
54465
4
564[)
4-555-64-586-75
107
127
1212
12-1414141213 .
12-14
129
13121112
8-1012
>2011 (&20)12 (&18)
121011
11-1.21210-11
9-12
for other parts of the body. For this reason their location has to be accurately specified when referring to them. This study describes the lateralwing scales found on veins R g (second) and R 4+5 (third).
In naming the type or shape of the scales in the present investigation(Fig. 20), an attempt was made to employ the terms that were available inthe literature, especially those of Theobald (1901, 1905) and Harbach andKnight (1980). However, it was necessary to supplement this nomenclaturewith botanical terms used in describing leaf morphology.
RESULTS
The shape and number of striae per scale were quite consistent for agiven species (Table 1). The various shapes of the scales that are discussedbelow are illustrated in Fig. 20.

Khalaf & Khalaf: Scales on Mosquitoes 303
Wing: several kinds of scales were found on veins Rs and R 4+5 •
1-0blanceolate (widest subapically, and with short, pointed apex):Anopheles (Fig. 1).
2-Spatulate (rounded apically): Deinocerites (Fig. 4), Uranotaenia(Fig. 5) (approaching weak asymmetrical), Toxorhynchites (Lynchiella) (Fig. 9). In Toxorhynchites (Toxorhynchites) the scaleswere shorter (obovate) (Fig. 8).
3-Asymmetrical (one side of the apex more or less produced): Mansoniini (Fig. 6, 7).
Fig. 1-5. Light micrographs of replicas of the wings of female mosquitoes: 1, Anopheles punctipennis. 2, Aedes taeniorhynchus, vein Rs (left)and R,j,+5 (right). 3, Psorophora confinnis, vein Rs left. 4, Deinoceritescancer, vein Rs left. 5, Uranotaenia sp., veing Rs' All X 600.
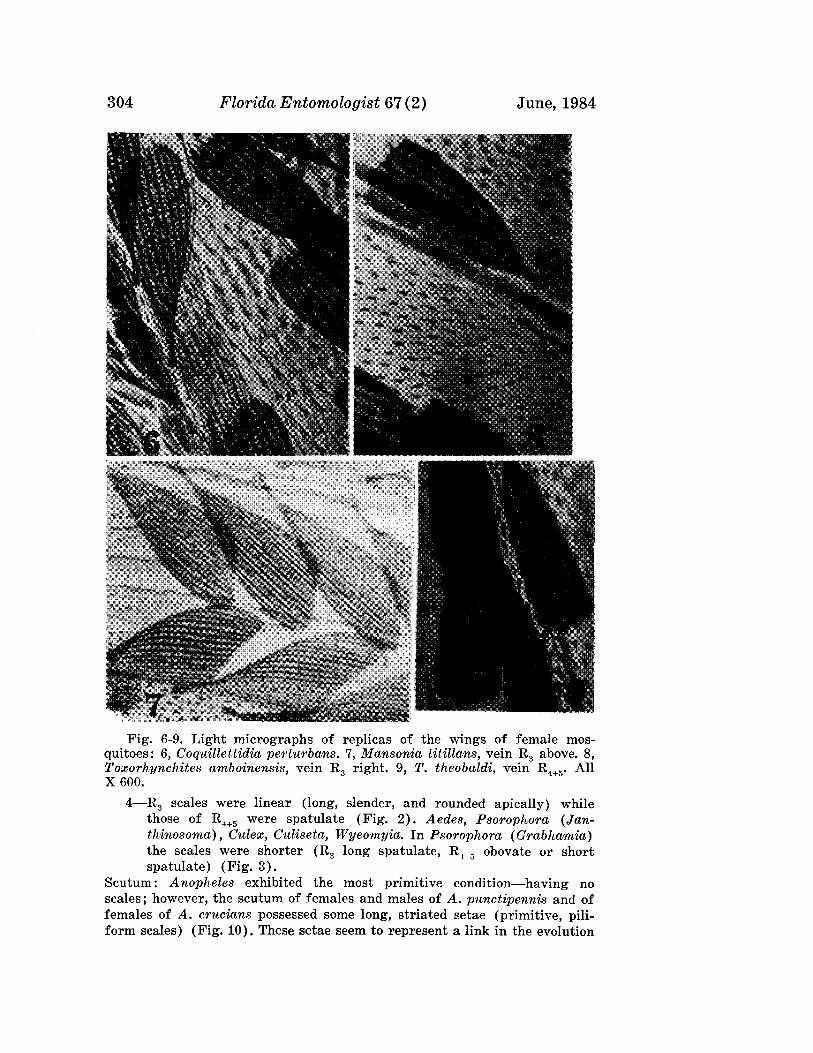
304 Florida Entomologist 67 (2) June, 1984
Fig. 6-9. Light micrographs of replicas of the wings of female mosquitoes: 6, Coquillettidia pwrturbans. 7, Mansonia titillans, vein R3 above. 8,Toxorhynchites amboinensis, vein R 3 right. 9, T. theobaldi, vein R4+5 • AllX 600.
4-R3 scales were linear (long, slender, and rounded apically) whilethose of R4+5 were spatulate (Fig. 2). Aedes, Psorophora (Janthinosoma), Culex, Culiseta, Wyeomyia. In Psorophora (Grabhamia)the scales were shorter (R3 long spatulate, R4+5 obovate or shortspatulate) (Fig. 3).
Scutum: Anopheles exhibited the most primitive condition-having noscales; however, the scutum of females and males of A. punctipennis and offemales of A. crucians possessed some long, striated setae (primitive, piliform scales) (Fig. 10). These setae seem to represent a link in the evolution
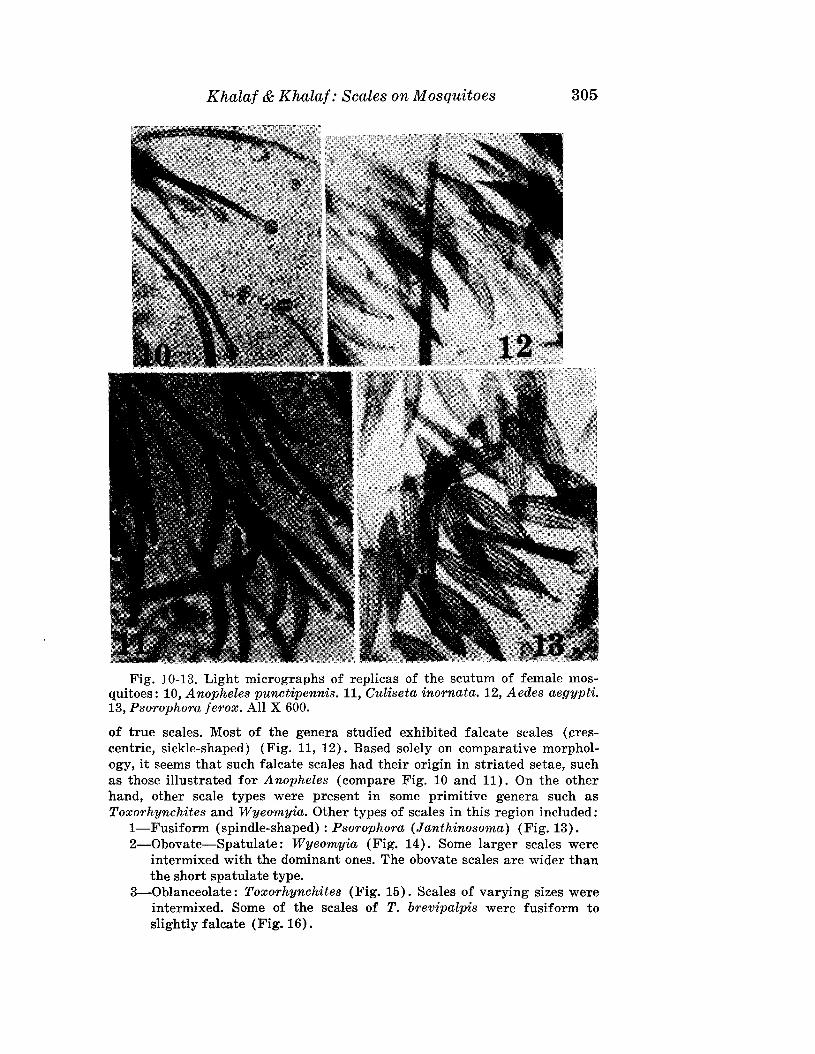
Khalaf & Khalaf: Scales on Mosquitoes 305
Fig. 10-13. Light micrographs of replicas of the scutum of female mosquitoes: 10, Anopheles punctipennis. 11, Culiseta inornata. 12, Aedes aegypti.13, Psorophora ferox. All X 600.
of true scales. Most of the genera studied exhibited falcate scales (~res
centric, sickle-shaped) (Fig. 11, 12). Based solely on comparative morphology, it seems that such falcate scales had their origin in striated setae, suchas those illustrated for Anopheles (compare Fig. 10 and 11). On the otherhand, other scale types were present in some primitive genera such asToxorhynchites and Wyeomyia. Other types of scales in this region included:
1-Fusiform (spindle-shaped) : Psorophora (Janthinosoma) (Fig. 13).2-0bovate-Spatulate: Wyeomyia (Fig. 14). Some larger scales were
intermixed with the dominant ones. The obovate scales are wider thanthe short spatulate type.
3-0blanceolate: Toxorhynchites (Fig. 15). Scales of varying sizes wereintermixed. Some of the scales of T. brevipalpis were fusiform toslightly falcate (Fig. 16).

306 Florida Entomologist 67 (2) June, 1984
Fig 14-16. Light micrographs of replicas of the scutum of mosquitoes:14, Wyeomyia mitchelii female. 15, Toxorhynchites theobaldi male. 16, T.brevipalpis female. All X 600.
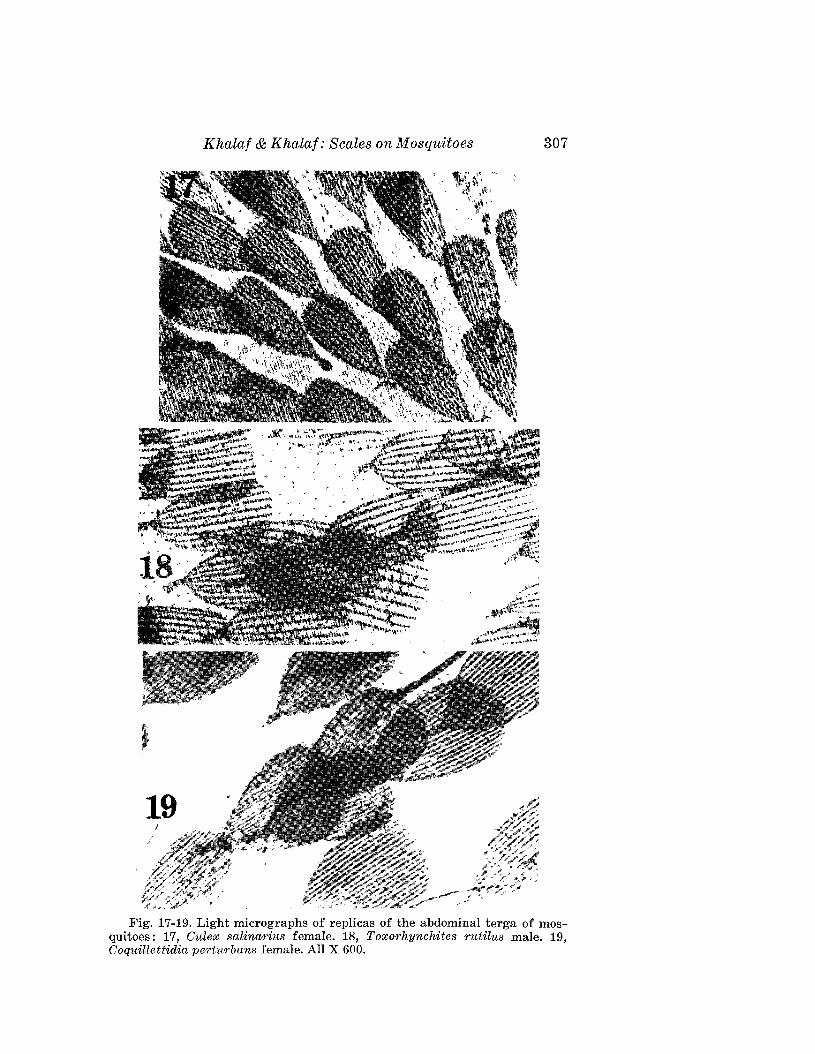
19"
Khalaf & Khalaf: Scales on Mosquitoes 307
Fig. 17-19. Light micrographs of replicas of the abdominal terga of mosquitoes: 17, Culex salinarius female. 18, Toxorhynchites rutilus male. 19,Coquillettidia perturbans female. All X 600.
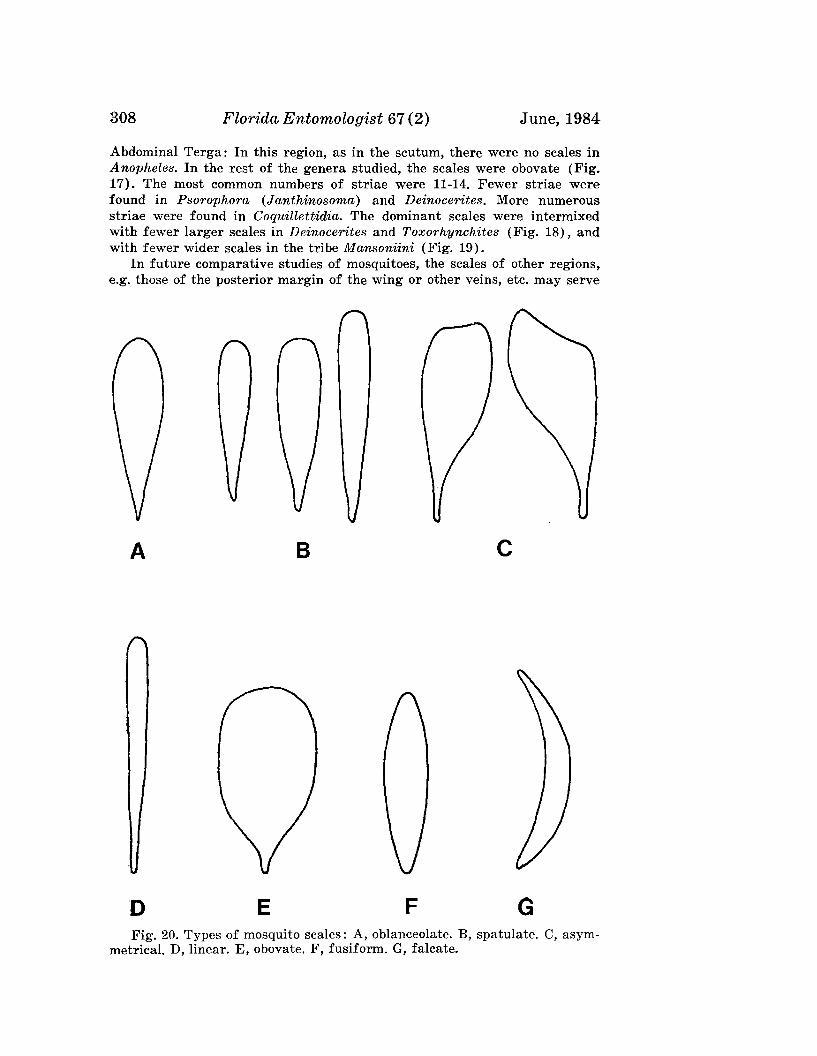
308 Florida Entomologist 67 (2) June, 1984
Abdominal Terga: In this region, as in the scutum, there were no scales inAnopheles. In the rest of the genera studied, the scales were obovate (Fig.17). The most common numbers of striae were 11-14. Fewer striae werefound in Psorophora (Janthinosoma) and Deinocerites. More numerousstriae were found in Coquillettidia. The dominant scales were intermixedwith fewer larger scales in Deinocerites and Toxorhynchites (Fig. 18), andwith fewer wider scales in the tribe M ansoniini (Fig. 19).
In future comparative studies of mosquitoes, the scales of other regions,e.g. those of the posterior margin of the wing or other veins, etc. may serve
A B c
o E F GFig. 20. Types of mosquito scales: A, oblanceolate. B, spatulate. C, asym
metrical. D, linear. E, obovate. F, fusiform. G, falcate.

Khalaf & Khalaf: Scales on Mosquitoes 309
taxonomy effectively. Keys of adult mosquitoes still involve clusters of species, e.g. Anopheles crucians Wied., A. georgianus King, and A. bradleyiKing, that cannot be differentiated from each other. Species of the groupcan be reared from determined larvae; a comparative survey of scales ofall the areas and veins of these reared adults can then be done which mightprove rewarding in differentiating members of the complex.
ACKNOWLEDGMENT
The mosquitoes were received through the courtesy of Dr. Joseph C.Cooney and Mr. Eugene Pickard of Tennessee Valley Authority, Fisheriesand Aquatic Ecology Branch; Mr. George Carmichael, Dr. Michael K.Carroll, and Mr. Stephen R. Sackett of New Orleans Mosquito Control; Mr.Leonard W. Trager, Jr. of Jefferson Parish Mosquito Control; and FloridaMedical Entomology Laboratory at Vero Beach. This investigation receivedsupport from the Academic Grant Fund of Loyola University.
REFERENCES CITED
CHRISTOPHERS, S. R. 1960. Aedes aegypti (L.) the yellow fever mosquito.,University Press, Cambridge. 738 p.
FOOTE, R. H., AND D. R. COOK. 1959. Mosquitoes of medical importance.Agriculture Handbook No. 152. ARS, USDA, Washington. 158 p.
HARBACH, R. E., AND K. L. KNIGHT. 1980. Taxonomists' glossary of mosquito anatomy. Plexus Publishing, Inc. 415 p.
KHALAF, K. T. 1980. Micromorphology of beetle elytra, using simple replicas. Florida Ent. 63 (3): 307-40.
MATTINGLY, P. F. 1957-1965. The culicine mosquitoes of the Indomalayanarea. British Museum (Natural History), London.
THEOBALD, F. V. 1901. The classification of mosquitoes. J. Trop. Med. 4:229-35.
---. 1905. Genera Insectorum, Diptera, Fam. Culicidae. Fas. 26. Belgium. 50 p.
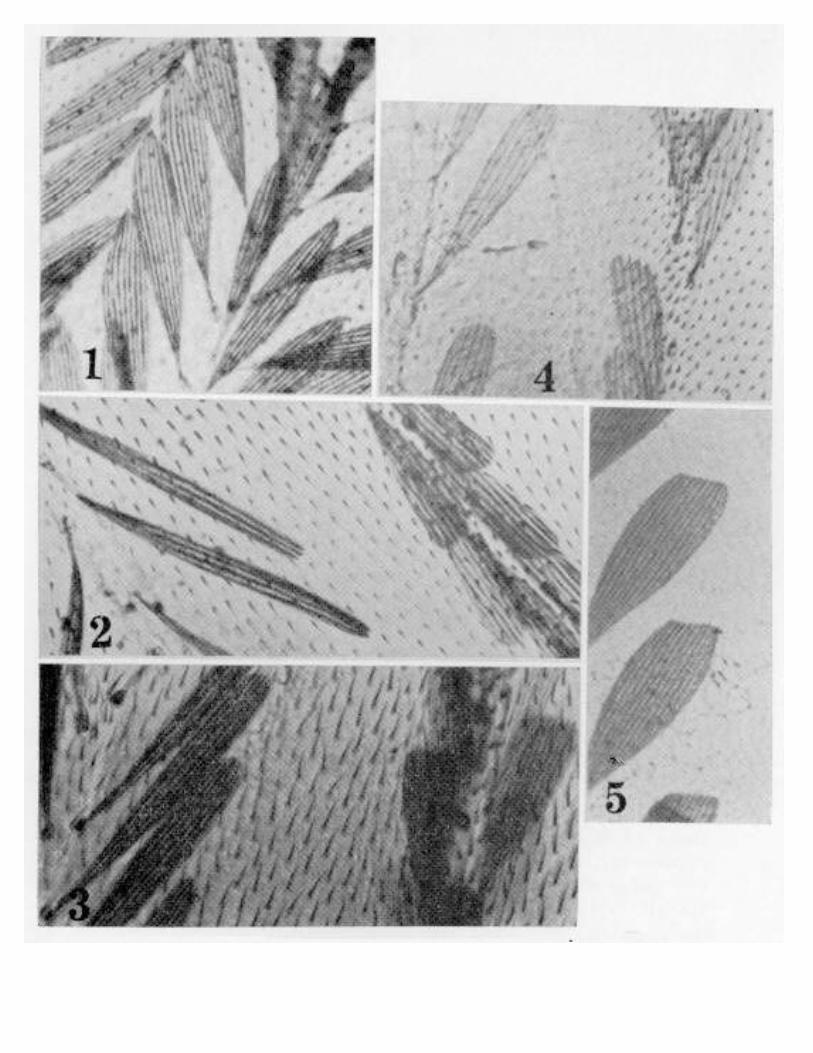

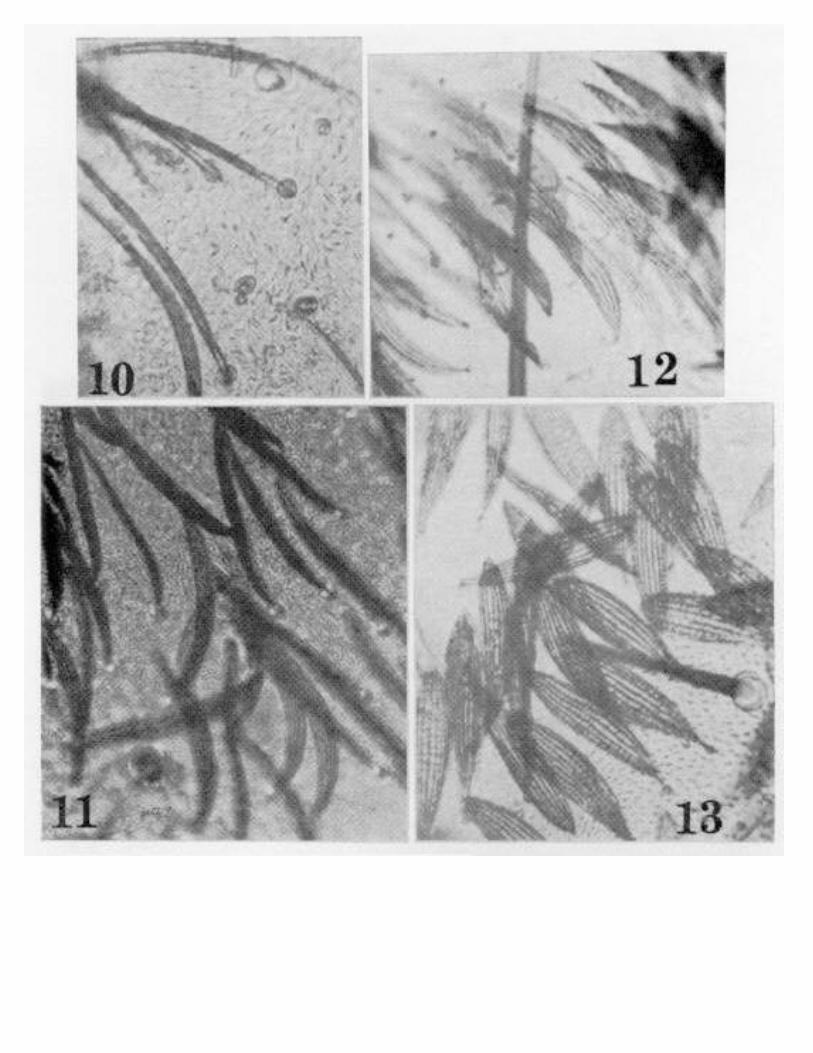
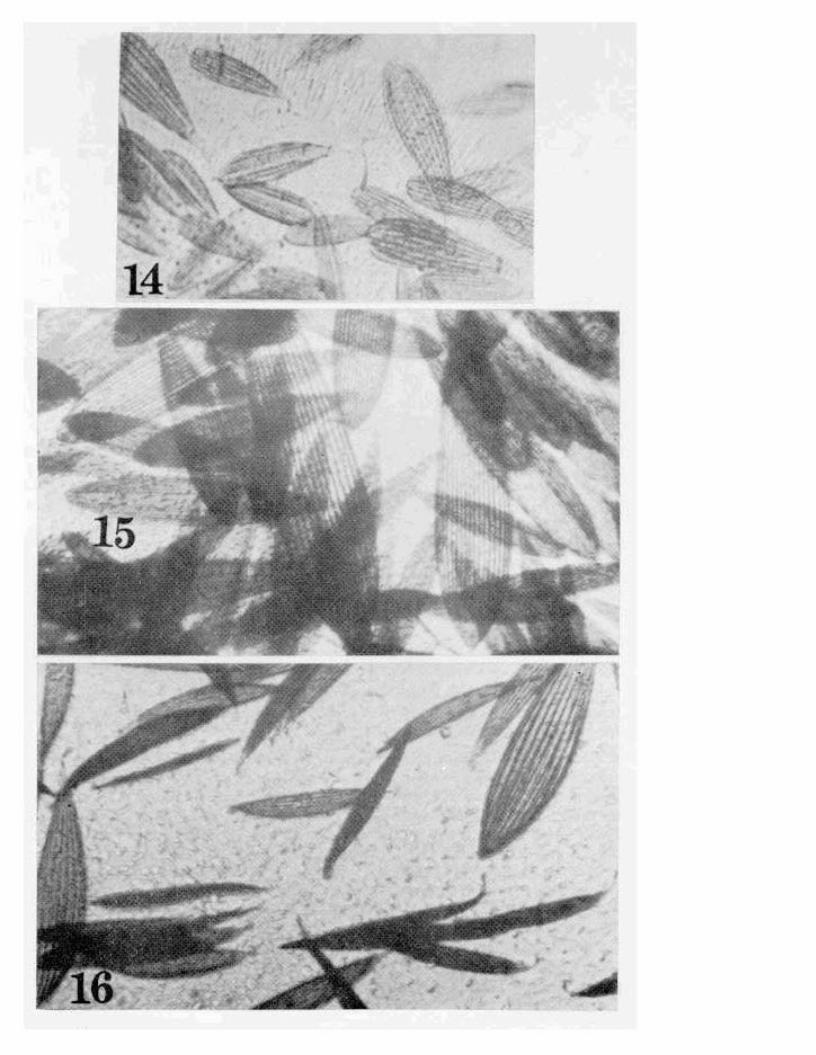
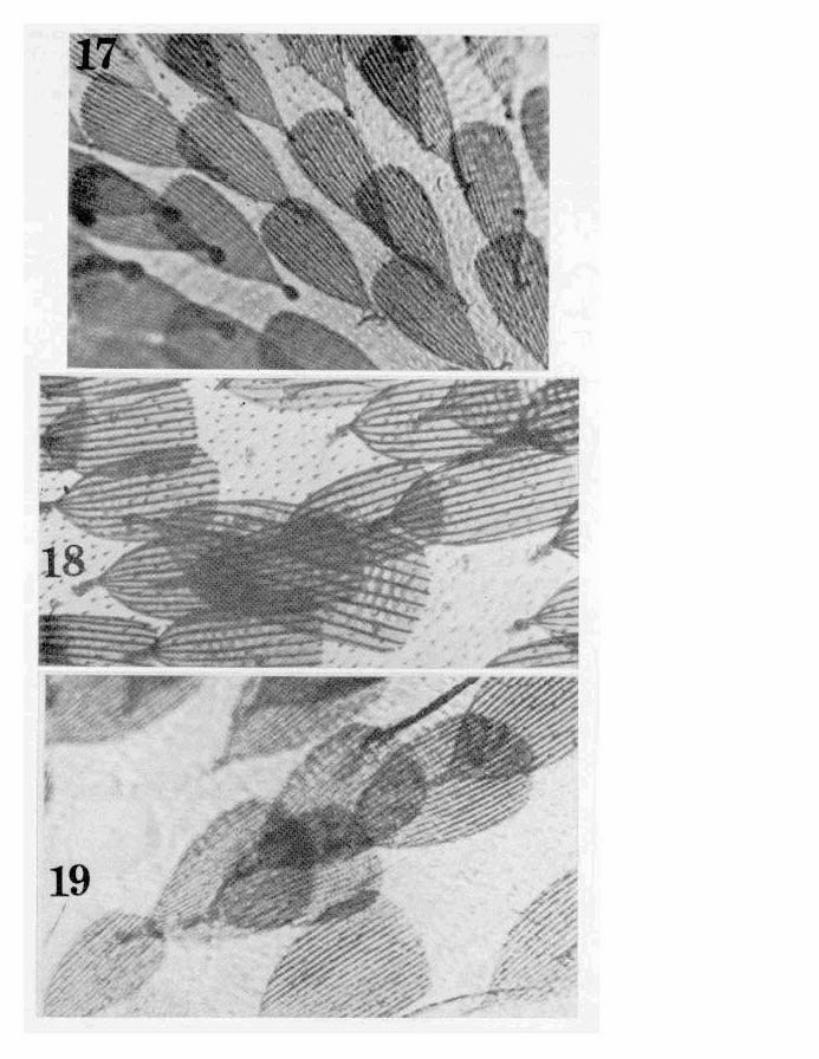

310 Florida Entomologist 67 (2)
SCIENTIFIC NOTES
STRUCTURAL ABNORMALITIES INTWO WORKER YELLOWJACKETS
(HYMENOPTERA: VESPIDAE)
KENNETH G. ROSSl
Department of EntomologyCornell University
Ithaca, NY 14853 USA
June, 1984
Developmental abnormalities among social wasps of the subfamilyVespinae have been infrequently reported (Edwards 1980). These mostcommonly involve irregularities in segmentation of the gaster (tergumsplitting or fusion) (Chapman 1917, Duncan 1939, Janet 1903, Maneval1932), but irregular segmentation of an antenna has also been recorded(Crevecoeur and De Walsche 1929). While collecting workers from 35Vespula colonies during the summer and fall of 1982 in central New Yorkstate, I di£covered 2 workers with external morphological abnormalities.
The first wasp was taken from a colony of V. fiavopilosa Jacobson on 17August. The head of this worker was deformed on one side and the gasterwas irregularly segmented (Fig. 1a & b). The left side of the head wascompressed dorso-ventrally in the region of the left compound eye whichwas expanded laterally. Gastral terga I and II were apparently normal,while tergum III was split at the midline. Terga IV and V were incompleteand partially fused; tergum VI appeared normal. The ventral segmentationof the gaster and the sting apparatus appeared normal so that this waspwas unlikely to be gynandromorphic. This worker was collected from withinthe colony and the ~ings showed no sign of wear. Ninety other workerscollected from this colony and 320 workers collected from 3 other conspecificcolonies appeared normal in al! respects.
A worker of V. vulgaris (L.) with an abnormal third gastral tergumwas collected on 17 September (Fig. 1c). The sclerite was split medially andthe right portion was partially fused to tergum IV. The wasp was normalin all other respects. This wasp was captured in a foraging trap placed overthe colony entrance (Schmidt et aI. 1978), thus she was capable of flyingand may have been an active forager. None of 39 other foragers examinedfrom this colony, or 657 workers from 7 other conspecific colonies exhibitedany obvious morphological abnormalities.
Given that the 35 Vespula colonies collected during this survey constitutea representative sample, the discovery of markedly aberrant individuals in2 of the colonies suggests that structural abnormalities in these wasps maybe more common than previously suspected. Extensive surveys of workerswithin colonies have not often been undertaken, and deformed workers maybe confined to the nest interior because of diminished flight or orientationcapabilities. The V. vulgaris worker from this report and a foraging workerof V. germanica (F.) with abnormal segmentation of the gaster (Chapman1917) are conspicuous exceptions to this expectation.
'Present address: Department of Entomology, University of Georgia, Athens, GA 30602.
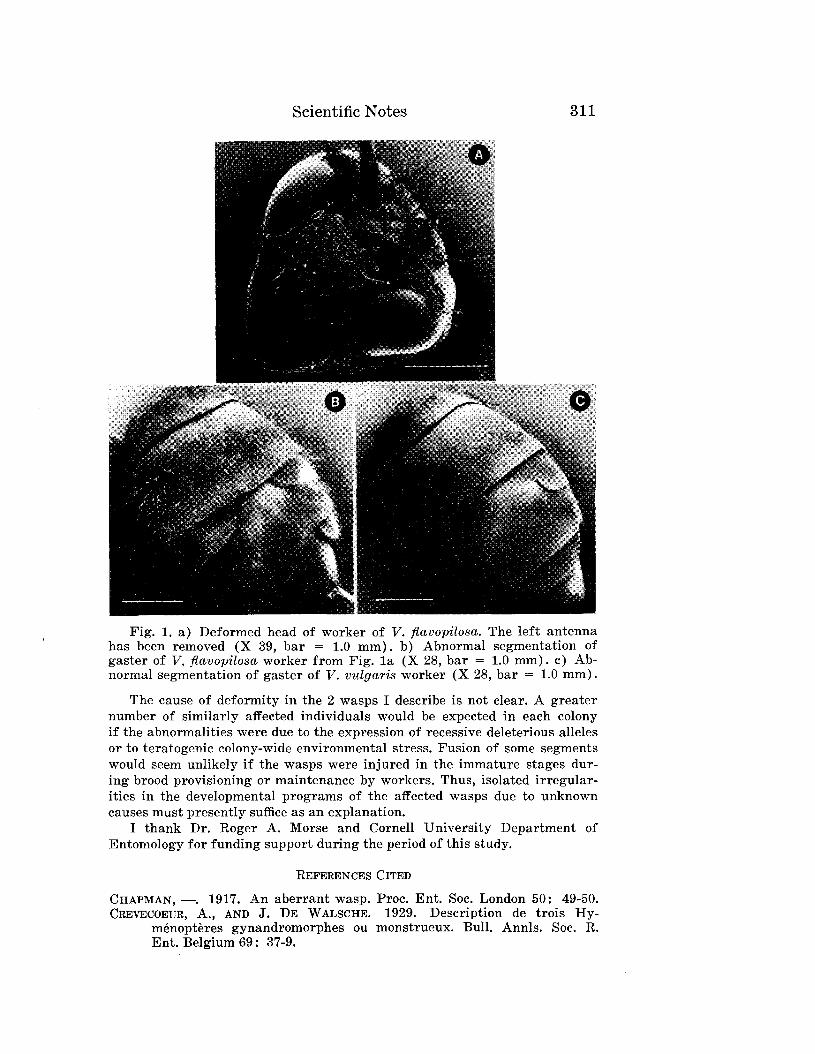
Scientific Notes 311
Fig. 1. a) Deformed head of worker of V. jlavopilosa. The left antennahas been removed (X 39, bar = 1.0 mm). b) Abnormal segmentation ofgaster of V. jlavopilosa worker from Fig. 1a (X 28, bar = 1.0 mm). c) Abnormal segmentation of gaster of V. vulgaris worker (X 28, bar = 1.0 mm).
The cause of deformity in the 2 wasps I describe is not clear. A greaternumber of similarly affected individuals would be expected in each colonyif the abnormalities were due to the expression of recessive deleterious allelesor to teratogenic colony-wide environmental stress. Fusion of some segmentswould seem unlikely if the wasps were injured in the immature stages during brood provisioning or maintenance by workers. Thus, isolated irregularities in the developmental programs of the affected wasps due to unknowncauses must presently suffice as an explanation.
I thank Dr. Roger A. Morse and Cornell University Department ofEntomology for funding support during the period of this study.
REFERENCES CITED
CHAPMAN, -. 1917. An aberrant wasp. Proc. Ent. Soc. London 50: 49-50.CREVECOEUR, A., AND J. DE WALSCHE. 1929. Description de trois Hy
menopteres gynandromorphes ou monstrueux. Bull. AnnIs. Soc. R.Ent. Belgium 69: 37-9.

312 Florida Entomologist 67 (2) June, 1984
DUNCAN, C. D. 1939. A contribution to the biology of North Americanvespine wasps. Stanford Univ. PubIs. BioI. Sci. 8: 1-272.
EDWARDS, R. 1980. Social Wasps, Their Biology and Control. Rentokil,East Grinstead, Britain. 398 p.
JANET, C. 1903. Observations sur les Guepes. Naud, Paris. 85 p.MANEVAL, H. 1932. Notes recueillies sur les Hymenopteres. Ann. Soc. Ent.
France 101: 85-112.SCHMIDT, J. 0., M. S. BLUM, AND R. W. MATTHEWS. 1978. Mass capture of
live yellowjackets: application and results of a capture-survey technique. J. Georgia Ent. Soc. 13: 163-8.
• • • • • • • • • • • • •A NEW TRAP FOR CAPTURING
DIAPREPES ABBREVIATUS(COLEOPTERA: CURCULIONIDAE)
W. J. SCHROEDERU.S. Department of Agriculture-ARS
Orlando, FL 32803 USAAND
1. F. JONESTri-State University
Angola, IN 46703 USA
Diaprepes abbreviatus L. is an important weevil pest of sugarcane,citrus, and many other commercial crops in the Caribbean Basin. It wasfirst reported in Florida by Woodruff (1964). During 1968, 2,630 ha in theinfested area were quarantined. Infestations are now distributed throughout 29,300 ha of commercial citrus in central Florida and 2,428 ha of outdoor nurseries and environs in south Florida. Because there has been noefficient method of detecting D. abbreviatus adult populations, recent research efforts have been directed toward developing a trap and trappingmethods.
Previous trapping studies on this insect in citrus were conducted withlight traps (Beavers et al. 1979) and sticky traps (Beavers, et al. 1982).Light traps were ineffective. Sticky traps are difficult to work with andcannot easily be placed in citrus trees where adult weevil populations aremost abundant. Described here is a durable, lightweight trap without stickysurfaces that is more useful for detection and population survey of D.abbreviatus.
Initially, 2 trap designs (A and B) were studied. The traps were similarin that each was a screen (0.32 cm mesh) trap consisting of a 15-cm-diam x16-cm-high cylindrical holding chamber (cricket cage) atop a 22-cm-diam x20-cm-high screen COIW with openings of 3.6 cm2 and 380 cm2• The two designs were different in that they had modified surfaces under the cone (Fig.1) .
Trap A had wire cloth vanes 38 cm long x 12.5 cm wide attached underthe cone. A 2.7-cm-OD PVC pipe seated in the ground was used to supportthe trap so that the bottom of the vanes touched foliage in the upper canopyof the tree.
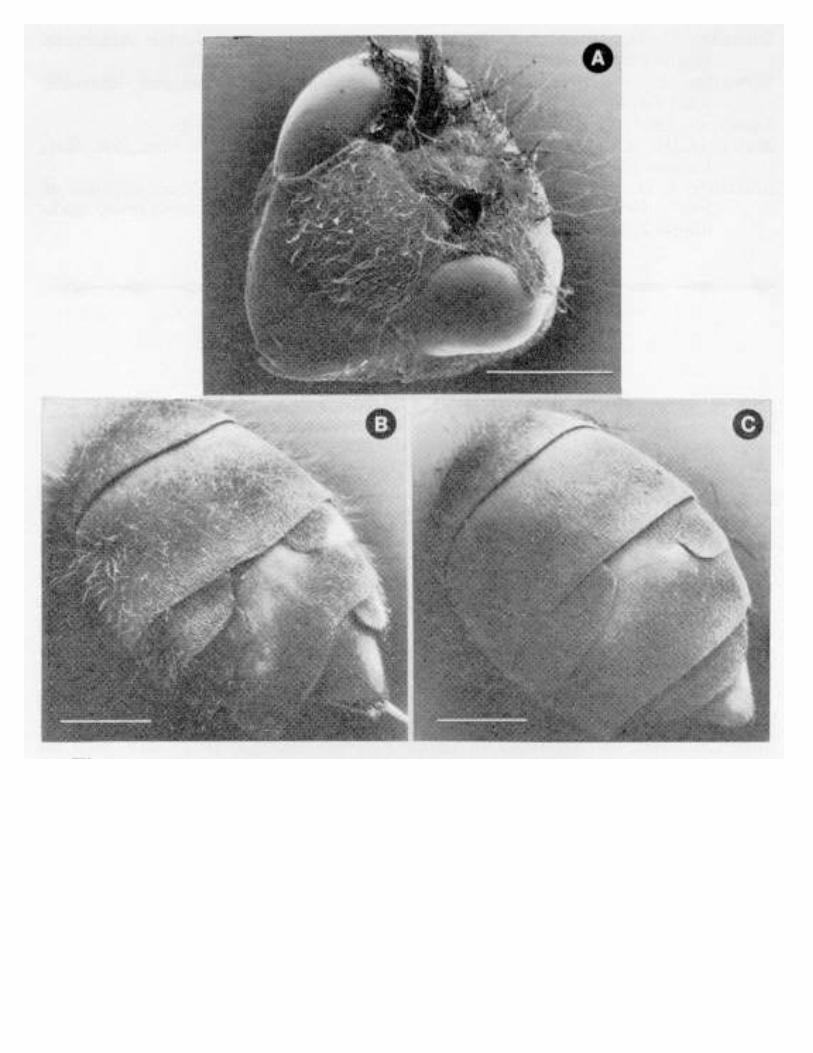

312 Florida Entomologist 67 (2) June, 1984
DUNCAN, C. D. 1939. A contribution to the biology of North Americanvespine wasps. Stanford Univ. PubIs. BioI. Sci. 8: 1-272.
EDWARDS, R. 1980. Social Wasps, Their Biology and Control. Rentokil,East Grinstead, Britain. 398 p.
JANET, C. 1903. Observations sur les Guepes. Naud, Paris. 85 p.MANEVAL, H. 1932. Notes recueillies sur les Hymenopteres. Ann. Soc. Ent.
France 101: 85-112.SCHMIDT, J. 0., M. S. BLUM, AND R. W. MATTHEWS. 1978. Mass capture of
live yellowjackets: application and results of a capture-survey technique. J. Georgia Ent. Soc. 13: 163-8.
• • • • • • • • • • • • •A NEW TRAP FOR CAPTURING
DIAPREPES ABBREVIATUS(COLEOPTERA: CURCULIONIDAE)
W. J. SCHROEDERU.S. Department of Agriculture-ARS
Orlando, FL 32803 USAAND
1. F. JONESTri-State University
Angola, IN 46703 USA
Diaprepes abbreviatus L. is an important weevil pest of sugarcane,citrus, and many other commercial crops in the Caribbean Basin. It wasfirst reported in Florida by Woodruff (1964). During 1968, 2,630 ha in theinfested area were quarantined. Infestations are now distributed throughout 29,300 ha of commercial citrus in central Florida and 2,428 ha of outdoor nurseries and environs in south Florida. Because there has been noefficient method of detecting D. abbreviatus adult populations, recent research efforts have been directed toward developing a trap and trappingmethods.
Previous trapping studies on this insect in citrus were conducted withlight traps (Beavers et al. 1979) and sticky traps (Beavers, et al. 1982).Light traps were ineffective. Sticky traps are difficult to work with andcannot easily be placed in citrus trees where adult weevil populations aremost abundant. Described here is a durable, lightweight trap without stickysurfaces that is more useful for detection and population survey of D.abbreviatus.
Initially, 2 trap designs (A and B) were studied. The traps were similarin that each was a screen (0.32 cm mesh) trap consisting of a 15-cm-diam x16-cm-high cylindrical holding chamber (cricket cage) atop a 22-cm-diam x20-cm-high screen COIW with openings of 3.6 cm2 and 380 cm2• The two designs were different in that they had modified surfaces under the cone (Fig.1) .
Trap A had wire cloth vanes 38 cm long x 12.5 cm wide attached underthe cone. A 2.7-cm-OD PVC pipe seated in the ground was used to supportthe trap so that the bottom of the vanes touched foliage in the upper canopyof the tree.
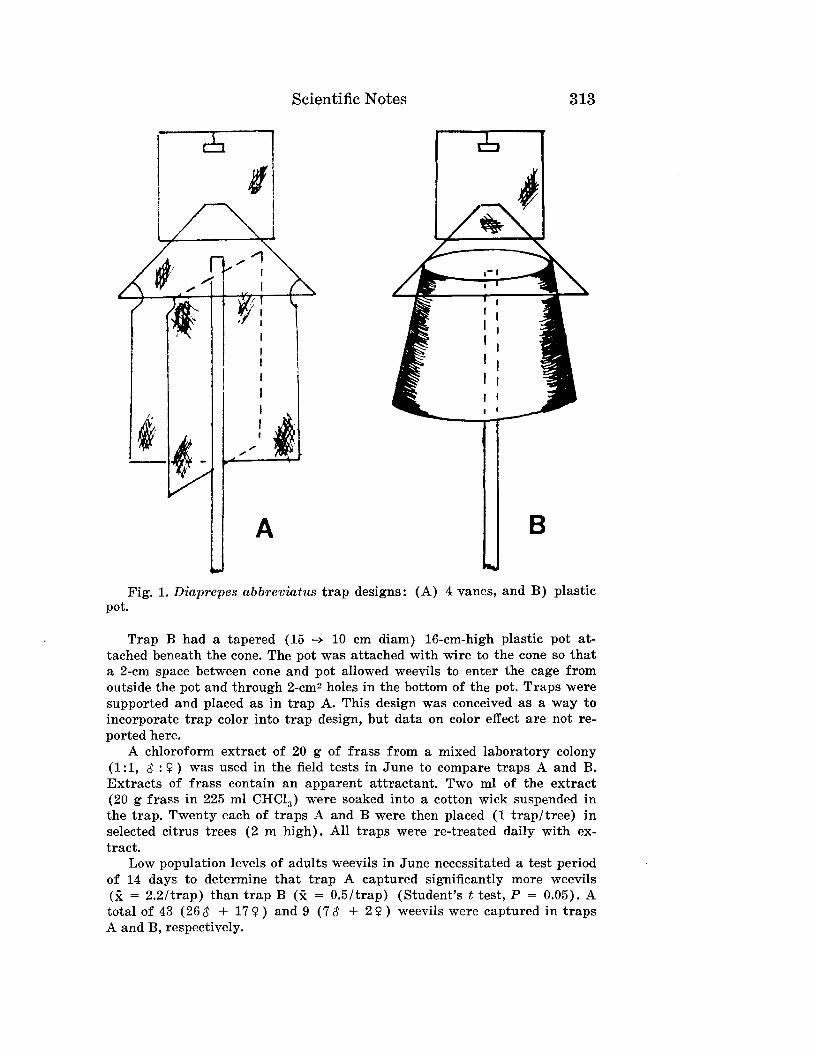
Scientific Notes 313
CS JII
I
A B
Fig. 1. Diaprepes abbreviatus trap designs: (A) 4 vanes, and B) plasticpot.
Trap B had a tapered (15 ~ 10 cm diam) 16-cm-high plastic pot attached beneath the cone. The pot was attached with wire to the cone so thata 2-cm space between cone and pot allowed weevils to enter the cage fromoutside the pot and through 2-cm2 holes in the bottom of the pot. Traps weresupported and placed as in trap A. This design was conceived as a way toincorporate trap color into trap design, but data on color effect are not reported here.
A chloroform extract of 20 g of frass from a mixed laboratory colony(1:1, <!; : <j') was used in the field tests in June to compare traps A and B.Extracts of frass contain an apparent attractant. Two ml of the extract(20 g frass in 225 ml CHClg ) were soaked into a cotton wick suspended inthe trap. Twenty each of traps A and B were then placed (1 trap/tree) inselected citrus trees (2 m high). All traps were re-treated daily with extract.
Low population levels of adults weevils in June necessitated a test periodof 14 days to determine that trap A captured significantly more weevils(x = 2.2/trap) than trap B (x = 0.5/trap) (Student's t test, P = 0.05). Atotal of 43 (26 <!; + 17 <j') and 9 (7 <!; + 2 <j') weevils were captured in trapsA and B, respectively.

314 Florida Entomologist 67 (2) June, 1984
REFERENCES CITED
BEAVERS, J. B., T. P. MCGOVERN, AND V. E. ADLER. 1982. Diaprepes abbreviatus: laboratory and field behavioral and attraetancy studies.Environ. Ent. 11: 436-9.
---, J. M. STANLEY, H. R. AGEE, AND S. A. LOVESTRAND. 1979. Diaprepesabbreviatus response to light traps in field and cage tests. FloridaEnt. 62: 136-9.
WOODRUFF, R. E. 1964. A Perto Rican weevil new to the United States(Coleoptera: Curculionidae). Florida Dept. Agric. Div. Plant Ind.Ent. Circ. 30: 1-20.
• • • • • • • • • •CORRELATIONS OF BURROW CHARACTERISTICSAND BODY SIZE IN BURROWING WOLF SPIDERS
(ARANEAE: LYCOSIDAE)
GARY L. MILLER AND PATRICIA R. MILLERDepartment of Entomology
Mississippi Entomological Museum, Drawer EMMississippi State University, Mississippi State, MS 39762 USA
The wolf spiders Lycosa carolinensis Walckenaer, L. georgicolaWalckenaer and members of the genus Geolycosa (Araneae: Lycosidae) aresimilar in their use of constructed burrows for retreats (Wallace 1942).Some of the characteristics of the burrows constructed by G. domifex (Hancock) and G. godeffroyi (L. Koch) are positively correlated with the size ofthe spider inhabiting the burrow (McQueen 1978, Humphreys 1976, respectively). However, similar relationships are not known for L. carolinensis or L. georgicola. Although these 2 wolf spiders construct burrows,they retain many of the typical lycosid behavioral characteristics, notablythe tendency to wander in search of prey (Kuenzler 1958, pers. observ.).Also only certain instars of L. carolinensis burrow (Shook 1979). Becauseof these life history differences we predicted that burrow characteristics,such as opening diameter, would not be as strongly correlated with bodysize for the 2 Lycosa species as in the Geolycosa. We attempted to verifythis from field collections in Florida and Mississippi and from laboratoryobservations.
We collected specimens of Lycosa carolinensis (Levy county), Geolycosaescambiensis Wallace (Holmes County), G. micanopy Wallace (Alachua,Levy, Marion and Putnam counties), G. ornatipes (Bryant) (Levy andPutnam counties), G. patellonigra Wallace (Clay, Levy, Marion and Putnam counties) and G. turricola (Treat) (Alachua and Santa Rosa counties)in Florida between 5 and 10 December 1983. Specimens of L. georgicola weretaken in Oktibbeha county near Starkville, Mississippi during the summerof 1983. All specimens were taken from burrows, and for each burrow,except those of L. georgicola and the laboratory populations, we measuredthe depth, the largest diameter of the burrow opening, the height of theturret when present (measured from ground level) and the distance acrossthe burrow entrance between the 2 extreme points of the turret (when theturret was present) which we termed the "total burrow diameter". During

314 Florida Entomologist 67 (2) June, 1984
REFERENCES CITED
BEAVERS, J. B., T. P. MCGOVERN, AND V. E. ADLER. 1982. Diaprepes abbreviatus: laboratory and field behavioral and attraetancy studies.Environ. Ent. 11: 436-9.
---, J. M. STANLEY, H. R. AGEE, AND S. A. LOVESTRAND. 1979. Diaprepesabbreviatus response to light traps in field and cage tests. FloridaEnt. 62: 136-9.
WOODRUFF, R. E. 1964. A Perto Rican weevil new to the United States(Coleoptera: Curculionidae). Florida Dept. Agric. Div. Plant Ind.Ent. Circ. 30: 1-20.
• • • • • • • • • •CORRELATIONS OF BURROW CHARACTERISTICSAND BODY SIZE IN BURROWING WOLF SPIDERS
(ARANEAE: LYCOSIDAE)
GARY L. MILLER AND PATRICIA R. MILLERDepartment of Entomology
Mississippi Entomological Museum, Drawer EMMississippi State University, Mississippi State, MS 39762 USA
The wolf spiders Lycosa carolinensis Walckenaer, L. georgicolaWalckenaer and members of the genus Geolycosa (Araneae: Lycosidae) aresimilar in their use of constructed burrows for retreats (Wallace 1942).Some of the characteristics of the burrows constructed by G. domifex (Hancock) and G. godeffroyi (L. Koch) are positively correlated with the size ofthe spider inhabiting the burrow (McQueen 1978, Humphreys 1976, respectively). However, similar relationships are not known for L. carolinensis or L. georgicola. Although these 2 wolf spiders construct burrows,they retain many of the typical lycosid behavioral characteristics, notablythe tendency to wander in search of prey (Kuenzler 1958, pers. observ.).Also only certain instars of L. carolinensis burrow (Shook 1979). Becauseof these life history differences we predicted that burrow characteristics,such as opening diameter, would not be as strongly correlated with bodysize for the 2 Lycosa species as in the Geolycosa. We attempted to verifythis from field collections in Florida and Mississippi and from laboratoryobservations.
We collected specimens of Lycosa carolinensis (Levy county), Geolycosaescambiensis Wallace (Holmes County), G. micanopy Wallace (Alachua,Levy, Marion and Putnam counties), G. ornatipes (Bryant) (Levy andPutnam counties), G. patellonigra Wallace (Clay, Levy, Marion and Putnam counties) and G. turricola (Treat) (Alachua and Santa Rosa counties)in Florida between 5 and 10 December 1983. Specimens of L. georgicola weretaken in Oktibbeha county near Starkville, Mississippi during the summerof 1983. All specimens were taken from burrows, and for each burrow,except those of L. georgicola and the laboratory populations, we measuredthe depth, the largest diameter of the burrow opening, the height of theturret when present (measured from ground level) and the distance acrossthe burrow entrance between the 2 extreme points of the turret (when theturret was present) which we termed the "total burrow diameter". During

Scientific Notes 315
our collecting in Florida, not all excavated burrows were measured. Spidersfrom these burrows were returned to the lab and allowed to construct newburrows in plastic cups containing sand. Inside burrow diameter andcarapace width were measured for these and the data are presented separately (identified as "lab"). We measured only inside burrow diameterfor L. georgicola. The parametric correlation between the natural log of thecarapace width, which we consider a measure of relative age (Hagstrum1971), and burrow characteristics and the corresponding t-value were determined.
The correlation between carapace width and inside burrow diameter forL. georgicola was not significant (Table 1). There was a moderate positivecorrelation between burrow diameter and carapace width for L. carolinensis,but the correlations were substantially stronger for the Geolycosa (Table1). The average difference between carapace width (untransformed) andinside burrow diameter was 10.6, 14.8 and 4.7 mm respectively for L. caroilnensis, L. georgicola and the combined Geolycosa. Only G. escambiensisand G. ornatipes had significant correlation between carapace width and theheight of the turret. For all of the Geolycosa, there were significant correlations between both burrow depth and total burrow diameter and carapacewidth (Table 1). The lab populations of G. micanopy and G. patellonigrahad significant correlation between carapace width and inside burrow diameter.
The higher correlations observed in the Geolycosa are a reflection of thegreater importance of the burrow in the life history of the group. The importance of the burrow to the Geolycosa is well known. Humphreys (1975)described how G. godeffroyi used burrows to reduce mortality resulting fromclimatic conditions and McQueen (1978) reported that G. domifex usedburrows as protection against predation. Shook (1978) indicated that members of his desert population of L. carolinensis also derived some protectiveand thermoregulatory advantage from the burrow. However, his data indicate that the members of the species may not need the protection of theburrow at all instars. Our observations on several populations of L.georgicola near Starkville, Mississippi indicate that the species is not dependent on the burrow during all instars and does not use the burrow forprey capture. Juvenile L. georgicola do not build burrows and adults foragesubstantial distances from their burrows. It is not known whether the burrow provided a thermoregulatory advantage.
The small average difference between the carapace width and burrowdiameter in the Geolycosa is another indication of the restrictive lifestyle ofthe group. Geolycosa do not generally occupy burrows constructed by otherspiders, whereas small L. carolinensis and L. georgicola are often foundhidding in burrows which were presumably constructed by larger membersof the species (pers. observ.).
We appreciate the comments of G. Baker, G. Uetz, and M. Greenstoneon an early draft of this note. We thank W. H. Cross for his continuedsupport of our research.
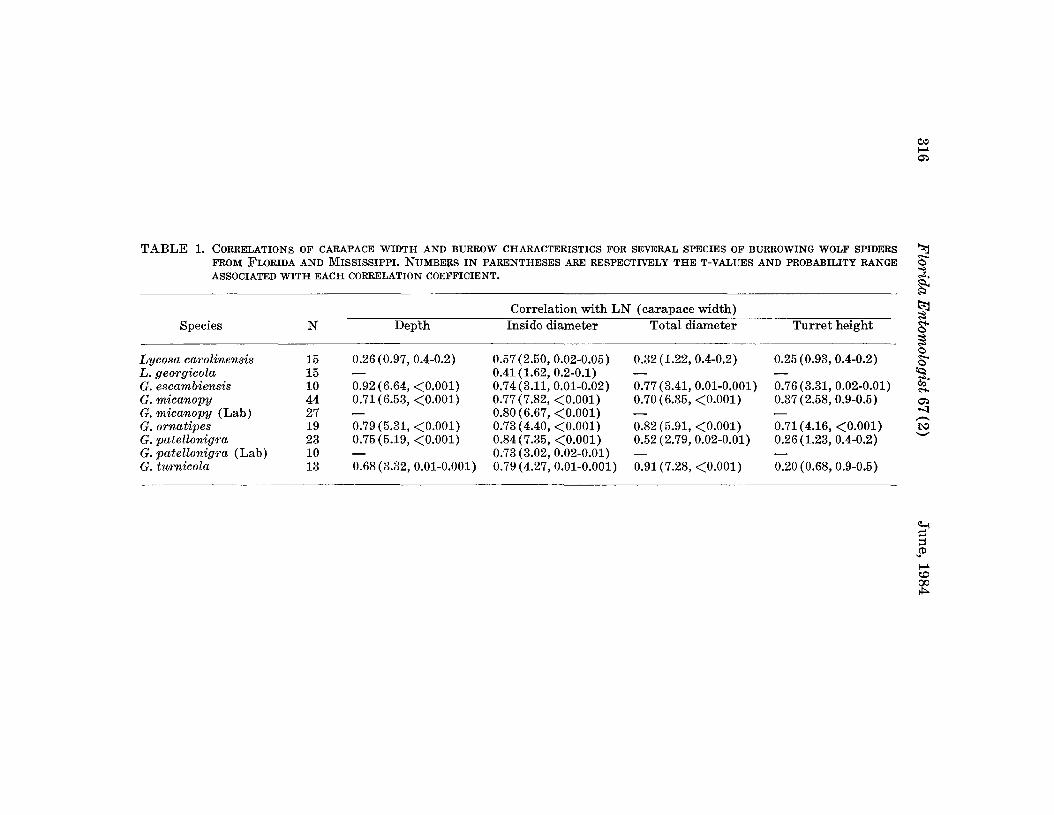
TABLE 1. CORRELATIONS OF CARAPACE WIDTH AND BURROW CHARACTERISTICS FOR SEVERAL SPECIES OF BURROWING WOLF SPIDERSFROM FLORIDA AND MISSISSIPPI. NUMBEltS IN PARENTHESES ARE RESPECTIVELY THE T-VALUES AND PROBABILITY RANGEASSOCIATED WITH EACH CORRELATION COEFFICIENT.
Correlation with LN (carapace width)Species N Depth Inside diameter Total diameter Turret height
Lycosa carolinensis 15 0.26 (0.97,0.4-0.2) 0.57 (2.50,0.02-0.05) 0.32 (1.22, 0.4-0.2) 0.25 (0.93,0.4-0.2)L. georgicola 15 - 0.41 (1.62, 0.2-0.1)G. escambiensis 10 0.92 (6.64, <0.001) 0.74 (3.11,0.01-0.02) 0.77 (3.41,0.01-0.001) 0.76 (3.31, 0.02-0.01)G. micanopy 44 0.71 (6.53, <0.001) 0.77 (7.82, <0.001) 0.70(6.35, <0.001) 0.37 (2.58,0.9-0.5)G. micanopy (Lab) 27 - 0.80 (6.67, <0.001)G.ornatipes 19 0.79 (5.31, <0.001) 0.73 (4.40, <0.001) 0.82 (5.91, <0.001) 0.71(4.16, <0.001)G. patellonigra 23 0.75(5.19, <0.001) 0.84 (7.35, <0.001) 0.52 (2.79, 0.02-0.01) 0.26 (1.23, 0.4-0.2)G. patellonigra (Lab) 10 - 0.73 (3.02,0.02-0.01)G. turnicola 13 0.68 (3.32, 0.01-0.001) 0.79 (4.27,0.01-0.001) 0.91 (7.28, <0.001) 0.20 (0.68,0.9-0.5)
CI:i....0')
~.,...c~
R:~
~~~c0~~..,..,.0')-::J.-~--
~r::I:l~(1)
....1:000H:>-

Scientific Notes 317
REFERENCES CITED
HAGSTRUM, D. W. 1971. Carapace width as a tool for evaluating the rateof development in spiders in the laboratory and field. Ann. Ent. Soc.America 64: 757-60.
HUMPHREYS, W. F. 1975. The influence of burrowing and thermoregulatory behavior on the water relations of Geolycosa godeffroyi(Araneae: Lycosidae) an Australian wolf spider. Oecologia 21: 291
311.---. 1976. Population dynamics of an Australian wolf spider. Oecologia
21: 291-311.KUENLER, E. J. 1958. Niche relations of three species of lycosid spiders.
Ecology 39 (3): 494-500.MCQUEEN, D. J. 1978. Field studies of growth, reproduction, and mortality
in the burrowing wolf spider Geolycosa domifex (Hancock). CanadianJ. Zool. 56: 2037-49.
SHOOK, R. S. 1979. Ecology of the wolf spider, Lycosa carolinensisWalckenaer (Araneae, Lycosidae) in a desert community. J.Arachnol. 6 (1): 53-64.
WALLACE, H. K. 1942. A revision of the burrowing spiders of the genusGeolycosa (Araneae, Lycosidae). American MidI. Nat. 27 (1): 1-61.

318 Florida Entomologist 67 (2)
Errata
June, 1984
ERRATA-THE GENUS fizophora IN FLORIDA. J. A. Slater and R. M.Baranowski. Florida Ent. 66 (4): 416-40.
On page 436 of this manuscript there is an error that is likely to causenomenclatorial confusion on the status of Ozophora 1"eperta and O. inornata.The CORRECT listing of these 2 species is as follows:
Ozophora reperta Blatchley
1954. Ozophora inornata Barber, American Mus. Novit. N. 1682, p. 5-6.NEW SYNONYMY.
The authors recognize inornata as the new junior synonym of Blatchleytsspecies, reperta. The Editor regrets not catching the mistake in time.

Book Announcement 319
ADVANCES AND CHALLENGES IN INSECT REARING. 1984. E. G. King and N. C.Leppla, eds. VSDA-ARS. 306 + xvi pages. 80 illustrations, tables, keys.
Order from: Superintendent of Documents, V.S. Gov't. Printing Office,Washington, D.C. 20402, Stock No. 001-000-04393-5. $8.00, domestic price.
"Advances and Challenges in Insect Rearing" fills the need for an up-to<late publication detailing the methods and materials, the problems and solutions, and the failures and successes of insect rearing as a scientific andeommercial endeavor. Whether you rear only a few insects for entomologicalresearch, larger numbers for pesticide screening or for production of consumer products, or millions for pest-control programs, this handbook willhave information you need on:
• Establishment and maintenance of insect colonies through genetic con-trol
• Diets and containerization• Engineering of rearing facilities and equipment• Control of pathogens and microbial contaminants in the rearing en
vironment• Production, use, and quality testing of many insect species, phyto
phagous and entomophagous, on natural and artificial diets• Management of insect-rearing systems, with emphasis on use of sys
tems analysis and data processing
The handbook is an outgrowth of an international meeting held in 1980by the V.S. Department of Agriculture and the Insect Rearing Group tobring together the leading experts in the field of insect rearing. The 36papers in the book are greatly revised versions of the papers presented atthat meeting.



![Entomological Bulletin - hkentsoc.orghkentsoc.org/bulletin/HKEB6(1)_April_2014.pdf · Hong Kong Entomological Bulletin ... Polistes hebreus [!]; Radoszkowski, 1871, Horae Entomol](https://img.pdfslide.net/doc/110x75/5c051a0709d3f2ff398d19b6/entomological-bulletin-1april2014pdf-hong-kong-entomological-bulletin.jpg)







![Entomological Society of Queensland NEWS BULLETIN1].pdf · Entomological Society of Queensland NEWS BULLETIN Volume 43, ... The ENTOMOLOGICAL SOCIETY OF QUEENSLAND, ... Geoff Thompson…](https://img.pdfslide.net/doc/110x75/5b078ec07f8b9ad1768e5cc7/entomological-society-of-queensland-news-1pdfentomological-society-of-queensland.jpg)

















