Embed Size (px)
DESCRIPTION
Phytomorphology volume 60 issues 1 & 2
Citation preview

SUHEL KHAN et al. – High Frequency Plant Regeneration and Detection of Genetic Variation in Datura metel L. 1Phytomorphology 60 (1 & 2) 2010, 1-8
IntroductionDatura metel L. commonly known as ‘Thorn apple’ or‘Kala Dhatura’ is one of the most important medicinalplants of the family Solanaceae. It is widely distributedover tropical and warm temperate regions of the world.Datura has a very special place in Ayurveda as mentionedin the works of Charaka and Sushruta. The principlealkaloid of Datura is hyoscine (scopolamine), which isused as pre-anaesthetic in surgery and childbirth, inophthalmology and in prevention of motion sickness.It also contains hyoscyamine, meteloidin and atropine.It is also useful in the treatment of several other diseases.
There are several reports on plant regeneration inDatura species via embryogenesis (Tyagi et al., 1981;Sharma et al., 1993). Despite being an importantmedicinal plant there are only a few reports availableon shoot multiplication in Datura metel (Bratati De,2003). Emphasis was given to androgenesis in this plantby some workers (Babbar & Gupta, 1986; Iqbal &
High Frequency Plant Regeneration and Detection of GeneticVariation among Micropropagated Plants of Datura metel L.
Suhel Khan1, Purnima Tyagi1, Sumita Kachhwaha1,2 and S.L. Kothari1,2,*
1 Experimental Morphogenesis and Plant Tissue Culture Laboratory, Department of Botany, University of Rajasthan,Jaipur-302004, India
2 Centre for Converging Technologies (CCT), University of Rajasthan, Jaipur-302004, India
ABSTRACT
High frequency plantlet regeneration was achieved from shoot tip explants of Datura metel cultured on MS mediumsupplemented with BAP (2.2 μM ) + IAA (2.8 μM ). Effect of other growth regulators including 6-benzylaminopurine(BAP), kinetin (Kn), indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), α-naphthaleneacetic acid (NAA) orphenylacetic acid (PAA) on in vitro morphogenesis was also investigated. The highest regeneration response was observedon medium containing 2.2 μM BAP + 2.8 μM IAA where approximately 93% of the cultures responded with an averageshoot number of 24.5±0.7 per explant in 6 weeks time. The best elongation response occurred on MS medium withBAP (0.4 μM) + PAA (0.7 μM) + GA3 (0.8 μM) where 93 % of shoots attained the average height of 5.3±0.3 cm.Rooting was best achieved on medium with 4.9 μM IBA. The plantlets with well-developed shoot and root systemswere acclimatized and successfully established in pots containing garden soil and organic manure. Examination of thegenetic fidelity of the micropropagated plants was done by the randomly amplified polymorphic DNA (RAPD) methodwith 35 primers, out of which 28 could successfully generate the bands. A total of 1274 scorable bands were amplified.Among them, 169 were polymorphic, being 13% of the total bands.
Key words : Datura metel, micropropagation, genetic variation, RAPD, regeneration
Abbreviations: BAP – 6-bezylaminopurine, IAA – Indole-3-acetic acid, NAA – α-naphthaleneacetic acid, PAA –Phenylacetic acid, RAPD – Random Amplified Polymorphic DNA, CTAB – Cetyltrimethyl AmmoniumBromide, NaOCl – Sodium hypochlorite
Author for Correspondence: S.L. Kothari, e-mail: [email protected]
Wijesekara, 2007) after the discovery of androgenichaploids in Datura innoxia (Guha & Maheshwari, 1964,1966). Lack of organized cultivation and incessantexploitation from the natural stands has lead to thedepletion of Datura plants at an alarming pace.Micropropagation will circumvent the problem ofavailability of good planting material in the case ofDatura metel which is also cultivated in some parts ofthe country.
Recently workers have started giving emphasis onthe analysis of genetic integrity of micropropagatedplants. Therefore, techniques such as cytological,isozymes and molecular markers have been employedto detect the variation if any or to confirm the geneticstability of micropropagated plants (Gupta & Varshney,1999). Among these techniques, RAPD is widelyemployed method in the detection of genetic variationsince it has the advantage of being technically simple,quick to perform and requiring only small amount ofDNA (Williams et al., 1990). Usefulness of RAPD

2 PHYTOMORPHOLOGY January–June 2010
analysis in detection of variation in micropropagatedplants has been demonstrated in large array of plants(Nybom, 2004).
The study reported here was aimed to develop anefficient regeneration system via multiple shoot budinduction from shoot tip explants in D. metel. Thegenetic homogeneity and variation of in vitro raisedplants of D. metel were assessed using RAPD analysis.Histological examinations were conducted to determinethe origin of the shoot buds.
Materials and Methods
Explant preparation and culture conditionsHealthy shoot tip explants were collected from matureplants of D. metel grown in Botany Department, RajasthanUniversity Campus, Jaipur. Explants were first rinsedwith 20 % (v/v) Extran (Merck, India) followed by3-4 washings with sterile distilled water followed bysurface-sterilization with 4 % (v/v) NaOCl (Qualigens,India) for 15 min. Traces of sterilant were removed byfour consecutive washings of autoclaved distilled water.The Murashige and Skoog (MS) medium (Murashige& Skoog, 1962) supplemented with 3% (w/v) sucrosewas used for all the experiments. The medium wassolidified with 0.8% (w/v) agar (Qualigens,bacteriological grade), pH adjusted to 5.8 before theaddition of agar and autoclaving at 121°C and 1.2–1.3kg cm2 pressure for 20 min. Three explants were keptin a single flask (100-ml ‘Erlenmeyer’ with 40 ml mediumin each) for 4 weeks. All the cultures were incubatedat 26±1°C with the 16-h light and 8-h dark cycle andthe light intensity of 25 μmol m–2 s–1 provided by coolwhite fluorescent tubes (Philips, India).Induction, proliferation and elongation of shoot budsThe shoot tips were cultured on MS mediumsupplemented with BAP (2.2,4.4,8.9,13.3,22.2 μM) aloneor Kinetin (Kn) (2.3,4.6,9.3,13.9,23.2 μM) alone orBAP in combination with IAA (2.8,5.7,11.4 μM), NAA(2.6,5.3,10.7 μM), PAA (3.6,7.3,14.7 μM) or Kn(2.3,4.6,9.3,13.9,23.2 μM). After four weeks ofincubation, shoot buds induced were kept for proliferationon the same medium. For elongation, shoot clusters wereseparated and kept on MS medium supplemented witheither GA3 (0.2, 0.5, 0.8, 1.4 μM) alone or GA3 (0.2,0.5, 0.8, 1.4 μM) in combination with BAP (0.4, 0.8,1.3 μM) or PAA (0.7, 1.4, 2.2 μM). Each treatmentconsisted of three explants with five replicates. Theexperiment was repeated twice. Observations wererecorded on a weekly basis.
Rooting and acclimatizationElongated shoots (2 cm or above) were excised andtransferred on to rooting medium comprising ½ strengthMS or full strength MS basal medium fortified with IAA(2.8,5.7,11.4 μM) and IBA (2.4,4.9,9.8 μM). Eachtreatment consisted of three explants with five replicates.The experiment was repeated twice. The plantlets withwell developed shoot and root systems were carefullytaken out and washed with tap water to remove agarclinging to roots. These plantlets were then transferredto earthen pots containing a mixture of soil and organicmanure (1:1). Humidity was maintained by coveringthe pots with polythene bags for initial few days oftransfer.HistologyShoot tip explants were fixed at the 5-wk stage byimmersion in formalin–acetic acid–alcohol (5:5:90) for48 h. The fixed material was dehydrated gradually bypassing through a TBA-xylol series (Johansen 1940) andthen infiltrated with liquid paraffin followed by threechanges of paraffin wax (Merck, India). Serial sectionsof 10 μM thickness were cut with a rotary microtome.Sections were fixed on the slides with the help of 4%formalin and Haupt’s adhesive. Slides were kept for 2d in xylene to remove the wax and were passed throughan alcohol series (100, 90, 70, 50, and 30%). These slideswere stained with 1% safranine and 0.5% fast green andagain passed through an alcohol series (30, 50, 70, 90,and 100%). The slides were mounted in DPX mountant(Ranbaxy, Haryana, India) and analyzed under a lightmicroscope.
DNA isolationDNA was extracted from fresh leaves taken fromrandomly selected regenerated plants by thecetyltrimethyl ammonium bromide (CTAB) method(Doyle and Doyle 1990). The mother plant from whichshoot tip explants were taken was selected as control.Approximately, 1gm of fresh leaves was ground topowder in liquid nitrogen using a mortar and pestle. Theground powder was transferred to a 25 ml tube with 5ml of CTAB buffer: 10% (w/v) CTAB, 5 M NaCl, 50mM EDTA, 1 M Tris-HCl pH 8.0, and 0.2% (v/v)β-mercaptoethanol. The homogenate was incubated at60°C for 2 h, extracted with an equal volume ofchloroform: iso-amyl alcohol (24:1). DNA concentrationwas estimated by using Nano drop spectrophotometer(ND 1000, Nano Drop Technologies, USA).

SUHEL KHAN et al. – High Frequency Plant Regeneration and Detection of Genetic Variation in Datura metel L. 3
PCR amplificationTwenty six arbitrary 10-base primers (OperonTechnologies Inc., Alameda, California) were used forPolymerase Chain Reaction (PCR). Each 25 μL ofreaction mixture contained 2.0 μL of 1.25 mM each ofdNTPs, 20 ng of the primer, 1× Taq polymerase buffer,0.5 U of Taq DNA polymerase (Genei, India) and 40ng of genomic DNA. DNA amplification was performedin a DNA Thermocycler (Corbett Research, Australia)programmed for 40 cycles: 1st cycle of 5 min at 95°C,1 min at 37°C and 2 min at 72°C; then 39 cycles eachof 1 min at 95°C, 1 min at 37°C, 2 min at 72°C followedby one final extension cycle of 7 min at 72°C. Amplifiedproducts were electrophoresed in a 1.2% (w/v) agarose(Sigma, USA) gels with 1× TBE buffer, stained withethidium bromide, and photographed under ultraviolet(UV) light using the BioRad gel documentation system.The size of the amplification products was estimatedfor a 100-bp ladder (M B I. Fermentas Inc.). All thereactions were repeated at least thrice.
Amplified DNA markers were scored as present orabsent both in the regenerated and the mother plants.
Statistical analysisData were subjected to one-way analysis of variance(ANOVA) by Fischer’s least significant difference(P=0.05, Gomez and Gomez 1984).
ResultsShoot tips of D. metel were treated with different hormonalregimes incorporated in MS medium. The resultsindicated (Table 1) that multiple shoot bud formationoccurred in all the treatments but the number variedsignificantly for each treatment. It ranged from 2 to11on MS medium fortified with BAP (2.2-8.9 μM). Kinetin(Kn) showed poorer response producing only 2-3 shootbuds. BAP was more efficient than Kn in shootregeneration but shoot bud length reached only up to1.5 cm in 6 weeks. In order to get better elongation,shoot tips were cultured on MS medium with BAP incombination with Kn and different auxins. BAP (2.2-8.9 μM) incorporated in the medium with Kn (2.3 μM)enhanced the response up to 16-17 shoot buds perexplant. These shoot buds were stout but failed toelongate further. NAA (2.6-5.3 μM) in combination with
Table 1. PGR optimization for shoot bud regeneration from shoot tip explants of D. metel.Culture period: 6 weeks
Growth Regulators (μM) Regeneration No. of shoot buds/BAP Kn IAA NAA PAA response (%) explant (mean±S.e.)2.2 73 10.7±1.6a
4.4 66 6.2±1.4b
8.9 53 4.0±0.9c
2.3 67 2.6±0.5d
4.6 46 1.5±0.5e
2.2 2.3 80 16.5±0.7f
4.4 2.3 60 9.1±0.7g
8.9 2.3 46 7.7±0.4h
2.2 2.8 93 24.5±0.7i
4.4 2.8 73 16.8±0.9j
8.9 2.8 66 9.9±0.9k
2.2 5.7 60 6.6±0.7l
4.4 5.7 73 10.9±0.8a
8.9 5.7 46 6.1±0.8b
2.2 2.6 53 6.6±0.9l
4.4 2.6 53 9.0±0.9g
8.9 2.6 60 14.0±0.8m
13.3 2.6 73 17.8±0.9n
2.2 5.3 53 6.5±0.5l
4.4 5.3 60 7.2±0.8p
8.9 5.3 66 8.9±0.9g
2.2 3.6 46 4.5±0.5q
4.4 3.6 73 7.5±0.9h
8.9 3.6 53 4.8±0.6t
Means in the column followed by different letters are significantly different at P = 0.05Each experiment comprised of 5 replicates and were repeated twice

4 PHYTOMORPHOLOGY January–June 2010
BAP (2.2-13.3 μM) produced up to 17 shoot buds perexplant. The combinations of PAA (3.6 μM) with BAP(2.2-8.9 μM) although was moderate in shoot induction(4-7 shoot buds) but resultant shoots were longer thanon any other combinations. Although shoot multiplicationwas observed on all the combinations of BAP with Kn,NAA and PAA, highest number (24-25) was obtainedon combination of BAP (2.2 μM) and IAA (2.8 μM)(Fig. 1A,B).
Taking lead from these results, induction of shootbuds was achieved on MS+BAP (2.2-8.9 μM) and theseshoot bud clusters were then transferred to MS+BAP(0.4-1.3 μM)+PAA (0.7-2.2 μM) for elongation.Elongation was observed on above said combinationsbut was achieved at significant level when thiscombination was added with GA3 (0.5-0.8 μM) althoughthe shoots formed were thin and slender. The bestelongation response occurred on MS mediumsupplemented with BAP (0.4 μM) + PAA (0.7 μM) +GA3 (0.8 μM) where 93% of shoots attained the heightof 5 cm or above (Fig. 1C) (Table 2).
The elongated shoots when separated and sub-cultured on full-strength MS medium, showed 46% ofrooting with poor vigor. Shoot inoculation on half-
strength MS medium or MS medium supplemented withIAA (2.8-11.4 μM), improved the percentage of rootingremarkably, along with enhancement in root length (3-4 cm). The roots were thinner, exhibited callus at thebase and hindered survival of plants under fieldconditions. Shoots kept on MS medium fortified withIBA (2.4-9.8 μM) showed maximum response (100%)(Table 3) (Fig. 1D). Here 10-15 thick, long roots with5-6 cm length were observed. The regenerated plantletswere acclimatized and successfully transplanted ex vitroand reared in pots (Fig. 1E).
RAPD studies were initiated for evaluation of clonalfidelity of mother plant and randomly selected tissueculture raised plants (TC). In the present study, of the35 primers used, only 28 generated RAPD markers. 17primers generated monomorphic bands across all theTC-raised plants including mother plant (Dm). However,11 primers were able to identify polymorphism in motherand tissue culture (TC) raised plants. The product rangedin size from 200-3000 bp. Primer OPA-12 generatedonly one band while maximum band reproducibility wasobserved with OPP-12 which induces 11.8 bands perplant. Highest number of polymorphic bands was obtainedwith primer OPE-12 while lowest polymorphism wasdetected with OPB-4. The TC-4 (tissue cultured raisedfourth plant) was true clone as it showed 100% similaritywith Dm (mother plant). TC-1, TC-2, TC-3, TC-5 plantswere close to mother plant by 93.4%. Group of TC-8and TC-9 were 93% similar to the mother plant. Minimumsimilarity was shown by TC-6 and TC-7 by 88%.
The shoot tips after 3- week of inoculation showedinduction of new shoot buds. At this time these werefixed in FAA for histological observations. The apicaldome enlarged and axillary multiplication from it wasobserved (Fig. 1F). However, lower portion of shoot tipenlarged and high growth of parenchymatous cells wasobserved. At the periphery of the parenchymatous callusmass meristemoids were actively engaged in formationof de novo shoot buds (Fig. 1G). Since histologicalobservations clearly indicated the formation of axillaryas well as de novo shoot buds, it was decided to ascertaintheir clonal status.DiscussionThere is critical and urgent need for sustainablemultiplication of over exploited medicinal plant Daturametel. In the present study we have aimed to developa protocol for fast multiplication of D. metel. We couldproduce 25 plantlets in 10 weeks time starting from asingle shoot tip. BAP was more potent in the induction
Table 2. Elongation of shoot buds upon sub-culturing on MSmedium supplemented with different plant growthregulators.
Growth regulator/nutrient (μm) Elongation Shoot length (cm)BAP PAA GA3 response (%) (mean±S.E.)
0.5 53 2.3±0.2a
0.8 66 2.5±0.1b
0.4 0.7 80 2.9±0.2c
0.8 1.4 60 2.8±0.1d
1.3 2.2 66 2.4±0.1e
0.4 0.5 66 2.7±0.2f
0.4 0.8 73 3.1±0.1g
0.8 0.5 60 2.4±0.2h
0.8 0.8 60 2.6±0.3i
1.3 0.8 66 2.4±0.1e
0.4 0.7 0.5 80 3.7±0.1j
0.4 0.7 0.8 93 5.3±0.3k
0.8 0.7 0.8 80 2.9±0.2c
1.3 0.7 0.8 73 2.5±0.2b
0.4 1.4 0.8 66 2.3±0.2a
0.8 1.4 0.8 80 3.1±0.3g
1.3 1.4 0.8 60 3.0±0.2l
0.4 2.2 0.8 66 2.3±0.0a
0.8 2.2 0.8 60 2.2±0.2x
1.3 2.2 0.8 53 2.0±0.2z
Means in the column followed by different letters are significantlydifferent at P = 0.05Each experiment comprised of 5 replicates and were repeated twice

SUHEL KHAN et al. – High Frequency Plant Regeneration and Detection of Genetic Variation in Datura metel L. 5
Table 3. In vitro rooting of shoot buds on MS medium containing various concentrations of auxins.
MS + Auxin (μM) Shoots rooted (%) Root number (range) Root Length (mean± S.E.) (cm) Root morphology½ MS 100 7-10 4.0±0.3a Thin, LongMS 46 3-4 2.4±0.5b Fragile, ShortMS + IAA
2.8 73 4-5 3.4±0.4a Thin, Short, callus at base5.7 100 7-10 4.0±0.1a Thin, Long, callus at base11.4 86 8-10 3.6±0.2a Thin, Short, callus at base
MS + IBA2.4 93 8-10 3.9±0.3a Thick, Short4.9 100 10-15 5.6±0.6c Thick, Long9.8 93 8-10 4.1±0.1a Thick, Long
Means in the column followed by same letters are not significantly different at P = 0.05Each experiment comprised of 5 replicates and were repeated twice
of shoot buds as observed by many workers earlier(Tyagi & Kothari, 2001). High frequency shoot-budregeneration from shoot tip explants of Datura meteloccurred on BAP and IAA supplemented medium as hasalso been reported in several other plants (Kothari &Chandra, 1984, Hossain et al., 1994). Shoot elongationoccurred on MS medium supplemented with BAP, PAAand GA3. Combination of BAP along with GA3 couldnot develop healthy shoots as these were thin and slender.Elongation of shoot buds was best achieved when mediumwith BAP and GA3 was also added with PAA. Small& Moris (1990) also used PAA for promoting elongationof shoot buds in Phaseolus vulgaris. Husain et al. (1999)reported positive effect of PAA in producing normal,elongated shoots in Capsicum annuum. Dhaka & Kothari(2002) observed improved bud elongation and plantregeneration in Helianthus annuus using PAA. Elongatedshoots were transferred to MS medium augmented withIBA (4. 9 μM). IBA has proved to be effective rootinghormone and is continuously used by many workers.(Pradhan et al., 1998; Ndoye et al., 2003).
Micropropagation through axillary buds or anyorganized meristems is generally considered to be a lowrisk method for genetic instability (Pierik, 1991), becausethe organized meristems are more resistant to geneticchanges as compared to unorganized callus under invitro conditions (Shenoy & Vasil, 1992). However, thereare many reports on the incidence of somaclonalvariations among various micropropagated plants (Rani& Raina, 2000; Gimenez et al., 2001; Rady, 2006;Gagliardi et al., 2007). In the present investigation,variation of 13% from the mother plant was observed.This variation can be attributed to shoot buds whichregenerated through axillary multiplication as well ascallus mediated regeneration. Unorganized callus forregeneration has a higher tendency for genetic changes
Table 4. Genetic variation in the tissue culture raised plantlets ofD. metel using RAPD markers
Primer Nucleotide Average number Sizecode sequence of bands per range of
5’ to 3’ primer per plant fragments (bp)OPA-01 CAGGCCCTTC 5.8 200-1000OPA-02 TGCCGAGCTG 3.8 200-900OPA-04 AATCGGGCTG 4.0 500-1400OPA-05 AGGGGTCTTG 4.8 300-1600OPA-11 CAATCGCCGT 3.6 700-2000OPA-12 TCGGCGATAG 1.0 1700OPA-13 CAGCACCCAC 5.0 300-1800OPA-14 TCTGTGCTGG 4.0 250-1100OPA-15 TTCCGAACCC 5.0 300-2400OPA-16 AGCCAGCGAA 3.7 700-3000OPA-17 GACCGCTTGT 8.0 250-1700OPA-18 AGGTGACCGT 3.0 700-1000OPA-19 CAAACGTCGG 2.5 350-1100OPA-20 GTTGCGATCC 3.1 200-1400OPB-04 GGACTGGAGT 4.4 250-1100OPE-12 TTATCGCCCC 6.7 600-2000OPF-01 ACGGATCCTG 3 465-1624OPF-02 GAGGATCCCT 4 537-1520OPF-03 CCTGATCACC 3 1140-2150OPF-04 GGTGATCAGG 3 1060-2880OPF-05 CCGAATTCCC 5 726-2200OPF-08 GGGATATCGG 5 284-1208OPF-09 CCAAGCTTCC 2 650-1150OPF-10 GGAAGCTTGG 6 500-1800OPF-20 GGTCTAGAGG 5 760-2000OPK-19 CACAGGCGGA 7.2 550-2000OPP-12 AAGGGCGAGT 11.8 300-1500OPS-13 GTCGTTCCTG 4.0 500-1100PCR amplification was repeated twice
which has been proved by RAPD analysis in some otherplants (Piola et al., 1999).
Surprisingly it has been observed that only initiatingexplant and the axillary or adventitious mode ofregeneration are not the only determining factors for thegenetic integrity. In vitro stress including physiological

6 PHYTOMORPHOLOGY January–June 2010
Fig.1: Morphogenic response of shoot tip explants of Datura metel on MS medium supplemented with various growth regulators.(A) Induction of shoot buds on MS + BAP (2.2μm) +IAA (2.8 μm); (B) Proliferation of shoot buds on MS+ BAP (2.2μm) +IAA (2.8 μm)(C) Elongation of shoot buds on MS + BAP (0.4μm) + PAA (0.7μm) + GA3 (0.8μm); (D) Rooting of in vitro regenerated shoots on MS+ IBA (4.9μm); (E) Field transferred plant; (F) Section showing direct differentiation of shoot buds; (G) Indirect differentiation of shootbuds

SUHEL KHAN et al. – High Frequency Plant Regeneration and Detection of Genetic Variation in Datura metel L. 7
and chemical conditions may result in mutational changes.Genetic fidelity has been ascertained by many workersemploying molecular markers like RAPD, ISSR etc.There are reports of similar genetic status of tissueculture progeny and mother plant (Singh et al., 2002;Carvalho et al., 2004; Ryynanen & Aronen 2005). Thepolymorphism in amplification products could resultfrom changes in either the sequence of the primer bindingsite or changes which alter the size or prevent thesuccessful amplification of a target DNA (eg. insertions,deletions, inversions). In our study, the variation wasnot confined to a few individuals but spread throughall the micropropagated plants just similar to the studyin Piper longum (Parani et al., 1997). Variations weredetected in plantlets derived from axillary buds (Sonejiet al., 2002) and direct adventitious shoot formation(Kumar et al., 1999; Virscek-Marn et al., 1999).Thereare other reports also where polymorphism among themicropropagated plants was examined. Santos et al.(2008) reported 21 to 42% polymorphism in Ananascomosus var. bracteatus, confirming the occurrence ofvariation during the micropropagation process.
Plant tissue culture is an alternative mean forcommercial production of large number of plant speciesincluding many medicinal plants. We report for thefirst time, multiplication of D. metel from shoot tip,obtained from field grown plants. This paper reportsa highly reproducible protocol for mass multiplicationof Datura metel. By this method unlimited plant materialcan consistently be obtained through out the year.Uninterrupted supply of plant material can be usedfor future pharmacological, physiological and
biochemical studies. The variation obtained in tissuecultured plants emphasizes further investigation atmolecular level so that off-types can be discarded andonly the genetically stable plants could be transplanted.AcknowledgementResearch fellowship to Suhel Khan awarded byUniversity of Rajasthan, Jaipur is gratefully acknow-ledged.
ReferencesBabbar, S.B. & Gupta, S.C. 1986. Chemicals affecting the androgenic
response of Datura metel: Glutamine, Glutamic acid, Serine andInositol. Beitrage Zur Biologie Der Pflanze, 60: 459-466.
Bratati, De 2003. Steroidal compounds from In vitro regeneratedshoots of Datura metel, Fitoterapia, 74: 14-17.
Carvalho. L., Goulão, L., Oliveira, C., Gonçalves, J.C. & Amâncio,S. 2004. RAPD assessment for identification of clonal identityand genetic stability of in vitro propagated chestnut hybrids. PlantCell Tissue Organ Culture, 77: 23-27.
Dhaka, N. & Kothari, S.L. 2002. Phenylacetic acid improves budelongation and plant regeneration in Helianthus annuus (L).Plant Cell Reports, 21: 29-34.
Doyle, J.J. & Doyle, J.L. 1990. Isolation of plant DNA from freshtissue, Focus, 12: 13-15.
Gagliardi, R.F., Hanai, L.R., Pacheco, G., Oliveira, C.A., Carneiro,L.A., Valls, J.F.M., Mansur, E. & Vieira, M.L.C. 2007. Assessmentof Genetic Stability among In vitro Plants of Arachis retusa usingRAPD and AFLP Markers for Germplasm Preservation. Journalof Integrative Plant Biology, 49: 307-312.
Gimenez, C., Garcia, E.D., Enrech, N.X.D. & Blanca, I. 2001.Somaclonal variation in banana: cytogenetic and molecularcharacterization of somaclonal variant CIEN BTA-03. In vitroCellular & Developmental Biology – Plant, 37: 217-222.
Gomez, K.A. & Gomez, A.A.1984. Statistical Procedures forAgriculture Research. John Wiley & Sons, New York.pp 7-83.
Fig. 2: Agarose gel electrophoresis of RAPD fragments obtained from primer OPK-19 and OPE-12 showing polymorphic bandsLane 1 – Molecular marker (M); Lane 2 – Field grown mother plant of D. metel (Dm); Lane3 11 – Tissue culture raised plantlets
A B

8 PHYTOMORPHOLOGY January–June 2010
Guha, S. & Maheshwari, S.C. 1964. In vitro production of embryosfrom anthers of Datura. Nature (Lond.), 204: 497.
Guha, S. & Maheshwari, S.C. 1966. Cell division and differentiationof embryos in the pollen grains of Datura in vitro, Nature (Lond.),212: 97-98.
Gupta, P.K. & Varshney, R.K. 1999. Molecular markers for geneticfidelity during micropropagation and conservation. CurrentScience, 76: 1308-1310.
Hossain, M., Islam, R., Karim, M.R., Joarder, O.I. & Biswas, B.K.1994 Regeneration of plantlets from in vitro cultured cotyledonsof Aegle marmelos Corr. (Rutaceae). Scientia Horticulture, 57:315-321.
Husain, S., Jain, A. & Kothari, S.L. 1999. Phenylacetic acid improvesIn vitro plant regeneration efficiency in Capsicum annuum L.Plant Cell Reports, 19: 64-68.
Iqbal, M.C.M.. & Wijesekara, K.B. 2007. A brief temperature pulseenhances the competency of microspores for androgenesis inDatura metel. Plant Cell Tissue Organ Culture, 89: 141-149.
Johansen, D.A. 1940. Plant Microtechnique, 1st edn.McGraw Hill,New York.
Kothari, S.L. & Chandra, N. 1984. In vitro propagation of Africanmarigold. Horticulture Science, 19: 703-705.
Kumar, M.B., Barker, R.E. & Reed, B.M. 1999. Morphological andmolecular analysis of genetic stability in micropropagatedFragaria x Ananassa cv. Pocahontas. In vitro CellularDevelopmental Biology - Plant, 35: 254-258.
Murashige, T. & Skoog, F.A. 1962. Revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiologia Plantarum,15: 473-497.
Ndoye, M., Diallo, I. & Dia, Y.K.G. 2003. In vitro multiplication ofthe semi arid forest tree, Balanites aegyptiaca (L.) Del. AfricanJournal of Biotechnology, 2: 421-424.
Nybom, H. 2004. Comparision of different nuclear DNA markers forestimating intraspecific genetic diversity in plants. MolecularEcology, 13: 1143-1155.
Piola, F., Rohr, R. & Heizmann, P. 1999. Rapid detection of geneticvariation within and among In vitro propagated cedar (Cedruslibani Loudon) clones. Plant Science, 141: 159-163.
Pradhan, C., Kar, S., Pattnaik, S. & Chand, P.K. 1998. Propagationof Dalbergia sissoo Roxb. through In vitro shoot proliferationfrom cotyledonary nodes. Plant Cell Reports, 18: 122-126.
Rady, M.R. 2006. In vitro culture of Gypsophila paniculata L. andrandom amplified polymorphic DNA analysis of the propagatedplants. Biologia Plantarum, 50: 507-513.
Rani, V. & Raina, S.N. 1998. Genetic analysis of enhanced axillarybranching derived Eucalyptus tereticornis Smith andE.camaldulensis Dehn. Plants. Plant Cell Reports, 17: 236-242.
Rani, V. & Raina, S.N. 2000. Genetic fidelity of organized meristem-derived micropropagated plants: A critical reappraisal. In vitroCellular Developmental Biology – Plant, 36: 319-330.
Ryynanen, L. & Aronen, T. 2005. Genome fidelity during short-andlong-term tissue culture and differentially cryostored meristemsof silver birch (Betula pendula) Plant Cell Tissue Organ Culture,83: 21-32.
Santos, M.D.M., Buso, G.C.S. & Torres, A.C. 2008. Evaluation ofgenetic variability in micropropagated propagules of ornamentalpineapple [Ananas comosus var. bracteatus (Lindley) Coppensand Leal] using RAPD markers. Genetics and Molecular Research,7(4): 1097-1105.
Sharma, V.K., Jethwani, V. & Kothari, S.L. 1993. Embryogenesis insuspension cultures of Datura innoxia Mill. Plant Cell Reports,12: 581-584.
Singh,A. Negi, M.S., Moses, V.K., Venkateswarlu, B., Srivastava,P.S. & Lakshmikumaran, M. 2002. Molecular analysis ofmicropropagated neem plants using AFLP markers for ascertainingclonal fidelity. In vitro Cellular Developement Biology - Plant,38: 519-524.
Small, D.K. & Moris, D.A. 1990. Promotion of elongation and acidinvertase activity in Phaseolus vulgaris L. internode segments byphenylacetic acid. Plant Growth Regulation, 9: 329-340.
Soneji, J.R., Rao, P.S. & Mhatre, M. 2000. Suitability of RAPD foranalyzing spined and spineless variant regenerants of Pineapple(Ananas comosus L., Merr.) Plant Molecular Biology Reporter,20: 307a-307i.
Tyagi, A.K., Rashid, A. & Maheshwari, S.C. 1981. Promotive effectof polyvinyl polypyrrolidone on pollen embryogenesis in Daturainnoxia. Physiologia Plantarum, 53: 405-406.
Tyagi, P. & Kothari, S.L. 2001. Continuous shoot production formicropropagation of Capparis decidua - A tree of arid agroforestrysystem. Journal of Indian Botanical Society, 80: 5-8.
Virscek-Marn, M., Bohanec, B. & Javornik, B. 1999. Adventitiousshoot regeneration from apple leaves – optimization of theprotocol and assessment of genetic variation among regenerants.Phyton, 39: 61-70.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A. & Tingey,S.V. 1990. DNA polymorphisms amplified by arbitrary primersare useful as genetic markers. Nucleic Acids Research, 18:6531-6535.

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 9Phytomorphology 60 (1 & 2) 2010, 9-19
IntroductionSahni (1948) instituted a new group of Jurassicgymnosperms ‘the Pentoxyleae’ on the basis ofinvestigations carried out by Rao (1943) and Srivastava(1937, 1944, 1945) on silicified premineralized materialcollected from Nipania in the Rajmahal Hills by Prof.Sahni and his students in 1932. Rao (1943) studied themorphology and anatomy of the leaf Taeniopterisspatulata McCleland, while Srivastava (1944, 1945)described the anatomy of four new taxa i.e. stems –Pentoxylon sahnii, Nipanioxylon guptai and seed bearingfructifications – Carnoconites compactus (originalC. compactum) and C. laxum. Vishnu-Mittre (1953)established a new taxon Sahnia nipaniensis for the malefructification of the Pentoxyleae. Since then a largenumber of descriptions, interpretations and mis-interpretations have been published on the material ofthe Pentoxyleae from the Rajmahal Hills (Bose et al.,1984; Sharma, 1969a,b, 1973a,b, 1979, 1989, 1996,2001, 2003; Sharma et al., 1987, 2001; Srivastava &Banerji, 2000; Suthar & Sharma, 1988; Suthar et al.,1987). Harris (1962, 1982) reported the existence ofpentoxylean plants in New Zealand, while Dauglas(1969), Drinnan and Chambers (1985) and White (1981)described the presence of the Pentoxyleae in AustralianMesozoic sediments. Cesari et al. (1998) have alsoreported the existence of probable pentoxylean plantsfrom the Antarctica, which have leaves like Taeniopterisand seed cones resembling Carnoconites (Fig. 3F).Sharma (2003) expressed doubt on pentoxylean affinityof the Antarctica material. Many botanists have alsoattempted hypothetical reconstructions of the organs,
Present Status of the Pentoxyleae – the Mesozoic GymnospermsB.D. Sharma1, D.R. Bohra2, O.P. Suthar3 and R. Harsh4
1 Kathmandi, Narnaul-123001, India, 2 Department of Botany, B.N. College, Udaipur-33001, India3 Department of Botany, Government College, Jaisalmer-345001, India
4 Department of Botany, M.S. Girls College, Bikaner-334001, India
ABSTRACT
Morphology and anatomy of the petrified specimens and slides of the ‘Pentoxyleae’ from the Rajmahal Hills examplifymany interesting characters and throw light on the phylogeny of this group of Mesozoic gymnosperms. Many hypotheticalreconstructions and misinterpretations are discussed. Some new and modified reconstructions are also proposed.Pentoxyleae is an indigenous group of plants which differ markedly from the specimens described from places outsideIndia like New Zealand, Australia and Antarctica.
Key words: Pentoxyleae, Rajmahal hills, modified, interpretations, reconstructions
e.g. male and seed bearing fructifications of thepentoxylean plants (Bose et al., 1985; Crane, 1985,1988; Srivastava & Banerji, 2000; Stewart & Rothwell,1993; Suthar & Sharma, 1988; Taylor, 1988; Taylor &Taylor, 1993). Similarly, many views have been expressedon the systematics and phylogeny of the Pentoxyleaewithout proper study of morphology, anatomy of organsand homology (Meeuse, 1961; Stewart, 1976; Taylor &Taylor, 1993).
The present paper is based on the study of a largenumber of specimens collected from different localitiesin the Rajmahal Hill e.g. Nipania, Sonajori and Amarjolaand several hundred slides prepared from them.Interestingly no differences could be observed inmorphology and anatomy of specimens collected fromdifferent Indian localities, whereas, the materials of thePentoxylales collected from New Zealand, Australia andAntarctica (Cesari et al., 1998; Drinnan & Chambers,1985; Harris, 1962, 1982; White, 1981) show widevariations in shape and size from that of the RajmahalHills. Material of the foreign countries is preservedeither as impressions or incrustations and as such theiranatomy remains unknown i.e. absence of the characteron the basis of which the fossils were grouped by Sahni(1948) into a new assemblage ‘the Pentoxyleae’. Presentstatus of the Pentoxyleae regarding structure, systematicsand phylogeny are discussed in this paper.
Material & MethodsThe Pentoxyleae was instituted on the basis of the studyof silicified material collected from Nipania, a localitysituated 5 Km North-West of the village Amarapara

10 PHYTOMORPHOLOGY January–June 2010
(approach Pakur-Amarapara road) in the Santhal Pargana(Jharkhand). The authors could collect (between 1962-2006) sufficient material from this locality. The seniorauthor (BDS) recently visited (August, 2006) the localityNipania and noticed drastic changes in the topographyof the area and could locate the site with great difficulty.The fossiliferous sediments are either used in constructionof walls or in making an approach road to the villageNipania. Amarjola is another locality which has yieldedpetrified pieces of stems of different diameters, shortshoots with leaf bases and the leaves of Nipaniophyllumraoi (Sharma, 1969, 1973a,b). Fossils are found hereembedded in a ferrugineous sandy Hill and are takenout by digging the sediment. Since the material is fragile,it is cooked in Canada balsam prior to sectioning witha wire bandsaw. The topography of Amarjola is alsochanged partly due to construction of a road betweenAlubhera and Pakur for the transport of coal and secondlythe Forest Department has planted a large number oftrees on the Amarjola Hill. As such, digging and collectionof fossils have become very difficult at Amarjola. Thethird locality is Sonajori situated 4 Km West of PakurRailway Station (Sharma & Bohra, 1976). The fossilsare silicified and are found in quarry No. 4. The quarryis now abondoned and fossiliferous stones are buriedunder debris and sand to a depth of 1-2 feet or moreand not easy to make collections of the fossiliferousmaterial. Slides were prepared by the usual method ofcutting, grinding and polishing techniques and mountedin dilute Canada balsam.
ObservationSystematic position of the Pentoxyleae is Cycadophyta,Pentoxylopsida, Pentoxylales, Pentoxyleae (Pant, 2002).The organ genera and species included in the Pentoxyleaenow are:-Stems –Pentoxylon sahnii Srivastava, 1944, 1945Nipanioxylon guptai Srivastava, 1944, 1945Guptioxylon amarjolense Sharma, 1969aGuptioxylon endocentrica Sharma, 1972Purioxylon jurassica Sharma, 1972aLeaves –Nipaniophyllum raoi Sahni, 1948Nipaniophyllum hirsutum Vishu-Mittre, 1957Nipaniophyllum anomozamoides Sharma, 1975Nipaniophyllum hobsonii Bose et al. 1985Taeniopteris spatulata Harris, 1982Taeniopteris draintreei Douglas, 1965; Drinnan andChambers, 1985
Pentoxylon australis White, 1981Taeniopteris sp. Cesari et al. 1998
Male fructification –Sahnia nipaniensis Vishnu-Mittre, 1953Sahnia laxiphora Osborn et al. 1991
Seed bearing cones –Carnoconites compactus Srivastava, 1945Carnoconites laxum Srivastava, 1945 (C. rajmahalensisBose et al., 1984)Carnoconites australica White, 1981Carnoconites llambiasii Cesari et al. 1998
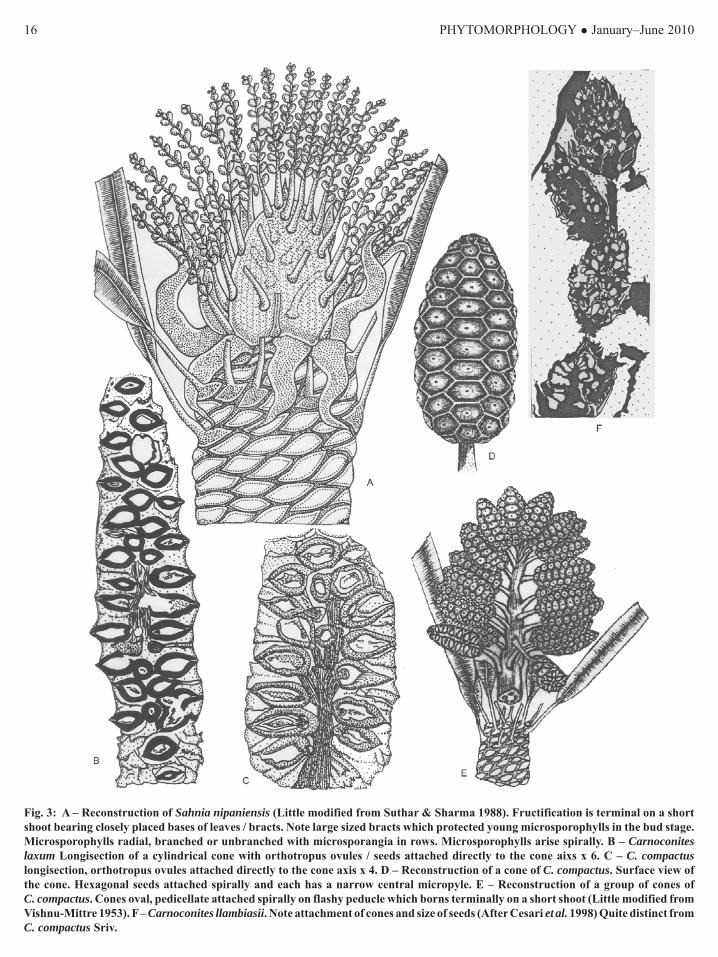
StemsPentoxylon sahnii Srivastava (1945): Hundreds ofspecimens from Amarjola and many chert pieces fromNipania and Sonajori bearing Pentoxylon sahnii arepresent with the authors. Specimens from Amarjolameasure 1.5-12.5 x 0.5-5.2 cm. branched or unbranched(branching is not dichotomous as suggested by Srivastava& Banerji, (2000) in their reconstruction - Fig. 2F).Markings of the detached short shoots are seen frequentlyon thick stems; surface smooth or transverse wrinklesare seen in the periderm (Fig. 1A). Short shoots arevariable in morphology depending on their functions.A vegetative short shoot is densely covered with rhomboidor semi-lunar leaf bases (Fig. 1A). Shoots which terminateinto male fructifications have leaf bases with tufts ofhairs on abaxial sides (Vishnu-Mittre, 1953). Seed bearingcones are produced on simple or branched pedicleswhich are terminal on a fleshy peduncle. The peduncleis terminal on a short shoot (Sahni, 1948, Figs. 39, 40,Suthar et al., 1988, Figs. 2-5) with leaf / bract bases(Fig. 3E). The thin shoots (Sharma, 1973) have distantlyplaced leaf bases, thus the plant had a multimorphicshoot system i.e. vegetative shoots, thin shoots andshoots terminating into male and female fructifications.
Cross section through stem (Fig. 1B) shows a distinctangular pith and a wide cortex. Both have scleroticpatches (Sharma, 1973, Fig. 1). In some of the stemsin addition to the outer periderm layer, an inner layerof periderm surrounding the steles is also visible (Sharma,1974, Fig. 5, 2001, Fig. 4). Steles 5-8, either uniformor of little variable shapes (Fig. 1B), each with a crushedprimary xylem and distinct, well developed secondaryxylem, either equal on both inner and outer sides or betterdeveloped on the inner side. The xylem of the outer sidedivides and the pieces shift to the cortex to make corticalbundles leaf traces (Sharma, 1973). The crushed primaryxylem does not show differentiation of protoxylem and

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 11
Fig. 1: (A-D) A. Reconstruction of a plant of Pentoxylon sahnii bearing leaves on short shoots as well as on thin branches. Both maleand seed bearing fructifications are terminal on short shoots. (Plant monoecious or dioecious ?). B. Cross section stem with 5 endocentricsteles x 4. C. A portion of stele with radially arranged tracheids of secondary wood and secondary phloem consisting of tangential rowsof sieve cells and phloem parenchyma x 40. D. Nipanioxylon guptai. Cross section stem with 8 steles in between the outer and the pithperiderm layers. Steles are exocentric, secondary wood compact with growth rings (Partially diagrammatic) x 6.

12 PHYTOMORPHOLOGY January–June 2010
as such its position (exarch/mesarch/endarch) cannot bedecided. The secondary xylem is compact anddifferentiated into growth rings (Fig. 1B). The tracheidsare squarish and arranged in radial rows (Fig. 1C). Atangential longisection through the wood shows short(1-8 cells high), uniseriate rays, while in radiallongisection uniseriate contiguous bordered pits arevisible on tracheids and one or two large circular pitsin a cross field (Sahni, 1948; Srivastava, 1945).
In many slides secondary phloem is seen wellpreserved surrounding the wood (Fig. 1C). It is 4-6layered and consists of sieve cells and the parenchymaarranged in tangential layers (Sharma, 1973, Fig. 5,Sharma & Bohra, 1973, Fig. 10).
The vegetative short shoots are of variable sizesranging from 0.8 cm. to 9.1 cm. in length and 0.35 to2.2 cm in thickness (Sharma, 1975; Sharma et al., 2001).Leaf bases are closely placed in spiral and show variationsin shape and size (Sharma, et al., 2001, Fig. 5) probablydepending on the age. The anatomy is also variable indifferent types of short shoots (Sharma, 1973, 1974a,b,1979, 1980). The bundles may be circular or little ellipticalwith a distinct primary xylem (Sharma, 1973a, 1979).The secondary xylem may or may not have differentiationof growth rings. In some of the short shoots the stelesare curved lunar shape (Fig. 2A) with xylem of onlycentripetal side (Srivastava, 1945; Sahni, 1948; Vishnu-Mittre, 1957). Leaf traces originate from the lateral sidesof the primary xylem and 3-5 traces enter a leaf base(Sharma, 1973a). In the peduncle of a seed bearing cone,5 or 6 narrow curved steles are present surrounding thepith. Each pedicle receives 5-7 traces (Sahni, 1948;Suthar et al., 1988). Thus the number of steles remainmore or less constant i.e. 5-7 or 8 in all the types ofshoots and in the stem. This cannot be explained on thebasis of the hypothesis of Stewart (1976) who believedthat by fission of a monostele, 5-7 steles of the stemof Pentoxylon were produced.Nipanioxylon guptai Srivastava (1945): It is anincompletely worked out stem found in the Nipaniachert. Srivastava (1937, 1945) described the presenceof unequally developed compact secondary wood in thesteles. Sahni (1948) suggested that the description wasincomplete and defective and described it as a youngshoot system. Vishnu-Mittre (1957) suggestedNipanioxylon a distinct genus on the basis of anatomyof a stem piece which was a conifer wood (Bose et al.,1985). The authors have in their collection three crosssections of shoots from the Nipania chert which resemble
in gross anatomy with N. guptai Sriv. The cross sectionis 12.5 to 15.4 mm in diameter with periderm layers inthe periphery of cortex and the pith. The ground tissueis parenchymatous with small patches of sclerenchyma.There are 7-8 steles surrounding the pith, which varyin shape and size i.e. circular to elliptical to obovate (Fig.1D). Primary xylem is crushed while the secondaryxylem is compact with radially arranged tracheids. Thesecondary xylem is either only on outer side or unequalon outer and inner sides or equally developed on bothsides. Growth rings faintly visible; phloemundifferentiated. In an oblique section through the steleuniseriate contiguous bordered pits are visible on radialwalls of tracheids identical to that of Pentoxylon sahnii.Further details are yet to be studied.Guptioxylon Sharma (1969): The genus was institutedfor a stem bearing pentoxylean anatomy, i.e. 4-6 stelesof variable shapes and sizes having compact secondarywood and crushed primary xylem make the vascularsystem. Pith and cortex wide, parenchymatous and havepatches of sclerenchyma as seen in Pentoxylon. But, thepith has medullary vascular bundles – a character notnoticed in any specimen of the Pentoxylon. Stem surfacesmooth with a periderm layer. Two species are knownof this genus i.e. G. amarjolense (Sharma, 1969a) andG. endocentrica (Sharma, 1972). Both the species werecollected from Amarjola and are preserved in an identicalplan.Guptioxylom amarjolense Sharma (1969a): There arefour circular to irregular shaped main steles with crushedprimary xylem and compact secondary xylemdifferentiated into growth rings. Medullary and corticalbundles vary in shape and size, exarch, mesarch orendarch. These bundles originate from the main stelesas a result of fission and detachment (Sharma, 1969a,1974). In the pith there is a small circular body witha layer of periderm like cells; origin and morphologyof this body remain unknown.G. endocentrica Sharma (1972): There are 5-6 endocentricsteles surrounding a large pith. Sclerotic patches arepresent both in cortex and pith similar to that of Pentoxylonsahnii. The secondary xylem of centrifugal side iscomparatively less developed than that of the centripetalside. It may break up into pieces which pass into cortexand make cortical bundles. The pith bundles originatefrom the centripetal portions of main steles and pass deepinto the pith. The fate of medullary bundles is unknown.The tracheids have typical pentoxylean pitting on theirradial walls.

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 13
Fig. 2: A – Pentoxylon sahnii. Cross section short shoot with sclerotic patches in the ground tissue and five narrow, elongated curvedsteles surrounding a wide pith x 16. B – Nipaniophyllum raoi. Cross section midrib with diploxylic bundles arranged in a row. Centripetalxylem (cpx) is adaxial and triangular. centrifugal xylem (cfx) is visible only in one bundle consisting of curved rows of cells, abaxial tocpx. x 24. C – N. raoi. Abaxial epidermis with scattered stomata in between veins x 60. D – Epidermis enlarged. Showing sinuous walledepidermal cells and haplocheilic stomata x 400. E – Sahnia nipaniensis. Longisection showing central cylindrical receptacle and spirallyborn radial microsporophylls with microsporangia in rows x 12. F – Reconstruction (a portion only) of a plant of Pentoxylon sahniisuggested by Srivastava and Banerji (2000). Note dichotomous branching and absence of short shoots. Leaves are produced terminallyon normal shoots.

14 PHYTOMORPHOLOGY January–June 2010
Guptioxylon is a distinct genus characterized byexcentric to concentric steles, each having a crushedprimary xylem, well developed secondary xylem. Corticalbundles originate from centrifugal side xylem whilemedullary bundles from centripetal side xylem. Corticaland medullary bundles are of various shapes.Purioxylon Sharma (1972a): The genus was institutedin honour of Prof. V. Puri of Meerut University, Meerut(India). The material was collected from Amarjola. Thestem is characterized by the presence of a distinct peridermlayer in the periphery of pith. The vascular zone is madeup of many collateral, conjoint and endarch bundles ina ring outside pith periderm layer. The cortical bundlesare of variable shapes and sizes and in morphology i.e.exarch, endarch or mesarch (Sharma, 1972a, 1974).Each has a compact wood with radially arranged tracheidsidentical to that of the Pentoxylon. Type of branching,associated leaves and the fertile organs are yet to bediscovered.
LeavesNipaniophyllum raoi Sahni (1948): Leaves strap shapedor linear with round to obtuse apices, sometimes lanceolatewith an obtuse apex, sub petiolate with a distinct midrib,lateral venation parallel having dichotomies at all levels.A cross section of the leaf shows 3-7 diploxylic bundlesin a row or an arch in the midrib (Sahni, 1948; Sharma,1982; Sharma & Bohra, 1977). Centripetal xylem istriangular and multi-cellular while centrifugal xylem iseither in two small patches one on either side of theprotoxylem or in an arc abaxial to the protoxylem(Fig. 2B). The protoxylem is a portion of centripetalxylem and its position is mesarch i.e. between cpx andcfx. Each bundle is surrounding by a distinct sheath.Sharma (1982) reported the presence of transfusion cellsin association with the sheath. Mesophyll isundifferentiated consisting of irregular cells. Vein bundlesare visible in the mesophyll. The lower and upperepidermises are quite distinct from the mesophyll cells.Upper epidermal cells are elongated and thick walked.Stomata are absent on this surface. The lower epidermisis made up of sinuous walled cells and is differentiatedinto vein cells and in between veins cells. Stomata areirregularly oriented in between veins (Fig. 2C). Rao(1943) and Sahni (1948) considered stomatasyndetocheilic while Vishnu-Mittre (1957), Sharma(1969, 1982) and Bose et al. (1985) described stomatahaplocheilic (Fig. 2D). Diploxylic bundles in the midriband haplocheilic stomata suggest cycadean affinity ofthe leaf of Pentoxyleae.
Vishnu-Mittre (1957) established a new species ofNipaniophyllum, N. hirsutum on the basis of study ofsections passing through the surface of lamina andobserved the presence of hairs / trichomes on theepidermis. It was probably a rare condition and noneof the later workers has observed this character in theleaves of Nipaniophyllum (Sharma & Bohra, 1977; Boseet al., 1985; Srivastava & Banerji, 2000). In N.anomozamoides Sharma (1975) incised margin of thelamina is identical to that of the bennettitalean taxonAnomozamites (Sharma 1969b). The material is a silicifiedchert from Nipania but the anatomy of this species isyet to be studied. N. hobsonii Bose et al. (1985) is basedon minor morphological differences in shape and sizefrom N. raoi. Sharma (2001) and the present authorsfeel that the creation of a new species N. hobsonii needsreconsideration. Similarly, until anatomical differencesfrom N. raoi are proved N. anomozamoides Sharma andN. hirsutum Vishnu-Mittre be treated as doubtful speciesor may be merged into N. raoi i.e. only one species existsin Nipania i.e. N. raoi Sahni (1948).
The specimens of leaves from New Zealand,Queensland and Antarctica have been collected eitheras impressions or incrustations and as such their anatomyremains unknown. The basic character of the pentoxyleanleaf is occurrence of 3-7 diploxylic bundles in a lineor an arc in the midrib. This character has not yet beenseen in any Taeniopteris leaves known from the countriesexcluding India. The authors feel that establishingcorrelation of the cycadian leaf Taeniopteris with thePentoxyleae is probably not justified. The leaves collectedfrom Antarctica are much narrower than the typicalleaves of N. raoi and correlation with Nipaniophyllumis hypothetical (Sharma, 2003).
FructificationsSahnia nipaniensis Vishnu-Mittre (1953): It is amicrosporangiate fructification seen in thin sectionsprepared through the Nipania chert. The fructificationis produced terminally on a short shoot covered withspirally arranged and closely placed leaf bases. Densegrowth of hairs is seen on the abaxial sides of leaf bases.The fructification consists of a number of radial branchedmicrosprophylls present surrounding a raised receptacle.These are united by their bases forming a cup shapedstructure. Microsporangia are solitary and in verticalrows on the microsporophylls (Vishnu-Mittre, 1953).There are published a number of interpretations on thestructure of Sahnia. Osborn et al. (1991) also showedorigin of microsporangia in rows on the entire length

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 15
of microsporophyll. Whereas, Bose et al. (1985) andTaylor, (1988) have described and figured in thereconstruction of Sahnia, origin of microsporangia interminal groups of 3-5 on small lateral branches in thedistal portion of radial, branched (Bose, et al. 1985) orunbranched (Taylor, 1988) microsporophylls. Such akind of attachment of microsporangia is neither visiblein the type specimen (slide) available at the Birbal SahniInstitute of Palaeobotany, Lucknow (India) nor in thetwo longisections (now preserved at the conservatoryof B.S.I.P., Lucknow) studied and described (Fig. 2E)by Suthar and Sharma (1988) and Sharma (1996, 2001).Slightly modified reconstruction of Sahnia nipaniensisis given here. According to this figure (Fig. 3A) theyoung male fructification was covered by large sizedbracts. These bracts spread out on maturation of thefructification. The microsporophylls were produced inspiral on an elongated receptacle and not in a whorl assuggested by Vishnu-Mittre’s (1953).Carnoconites Srivastava (1944, 1945): The genus wasinstituted for seed bearing cones and two species wereidentified i.e. C. compactus and C. laxum. Sahni (1948)and Vishnu-Mittre (1953) added further observations onthese cones.C. compactus: It is a small stalked globose to oval bodybearing a large number of seeds. In surface view thehexagonal seeds are seen arranged in vertical rows(Fig. 3D), actually the seeds are arranged spirally. Alongisection shows a central axis on which orthotropusovules / seeds are attached in a close spiral (Fig. 3C).Each ovule has a thick integument differentiated intofleshy sarcotesta, stony sclerotesta and a thin endotesta.The innermost layer is visible only in few ovules (Sharma,1989, 2001, Fig. 15). The vascular supply ends at thechalazal i.e. integumentary layers are non-vascularized(Sharma 1996). Inner portion of sarcotesta adjacent tosclerotesta bears idioblasts, solitary or in radial groupsof 2 or 3. Nucellus is free from integument except thenucellar pad. Pollen chamber is absent. In some seedsmegagametophyte is preserved bearing 1 to 4 embryos(Sharma, 1989; Srivastava, 1945; Sahni, 1948; Vishnu-Mittre, 1953). Bose et al. (1985) described the seedsdicotyledonous but the authors have not yet seen thepreservation of cotyledons in any seed.C. laxum Srivastava (C. rajmahalensis Bose et al., 1984):It is an elongated cylindrical, branched or unbranchedseed bearing cone (Fig. 3B). Vishnu-Mittre (1953)observed a tribranched cone of C. laxum. Seeds / ovulesare orthotropus and attached directly to cone axis
(Fig. 3B). The ovules are comparatively smaller in sizethan those of C. compactus but built on identical plani.e. integument is thick, heterogeneous and non-vascularized. However, idioblasts are not seen in theinner portion of sarcotesta. Nucellus is free fromintegument and a distinct nucllar pad is formed.Megagametophyte is preserved in some ovules but neitherarchegonia nor the embryos could be seen in any ovule/seed.
Regarding the manner of attachment of cones to thepeduncle and the number of cones on a short shoot isa matter of debate among the palaeobotanists. Sahni(1948) showed 8-12 cones in a loose bunch, whileVishnu-Mittre (1953) figured 14-18 cones on a cylindricalpeduncle. Crane (1985) made a reconstruction showinghundreds of seed bearing cones on a narrow, fleshy shortshoot. Bose et al. (1985) followed Crane and made thereconstruction bearing hundred of pedicillate seed bearingcones on a peduncle which in turn was borne terminallyon a short shoot. Srivastava and Banerji (2000) however,reduced the number of cones in their reconstruction ofC. compactus. Sharma (2001) did not agree with Crane(1985), Bose et al. (1985) and Srivasatava and Banerji(2000) and considered Sahni’s (1948) opinion the bestbecause this is based on facts and visible in slides andnot merely an imagination. In our reinvestigation ofslides and specimens we observed that the seed coneof C. campactus is oval in shape (not globose as suggestedby Sahni (1948)) with an obtuse to round apex. In surfaceview hexagonal seeds are seen in spiral and each bearsa central narrow micropyle (Fig. 3D). There are 16-20pedicillate cones in C. compactus produced in a spiralon a fleshy peduncle. This peduncle in turn was borneterminally on a short shoot (Fig. 3E).
DiscussionThe Pentoxyleae was established on the basis of uniqueanatomical characteristics viz., multistelic vasculature,crushed primary xylem, pycnoxylic wood with excentricdevelopment and distinct growth rings; short anduniseriate rays; tracheids with uni or biseriate contiguousbordered pits on radial walls and 1-2 large and circularpits in cross field (Sahni, 1948). Both pith and cortexhave patches of sclerenchyma/sclerotic cells. Mucilagecanals are absent (Sharma, 2006). Shoot systemmultimorphic. Leaf simple and spatulate, lateral veinsparallel with dichotomies at all levels. Midrib has 3-7diploxylic bundles in an arc or row. Stomata irregularlyoriented and haplocheilic. A number of traces enter aleaf base. Both male and seed bearing fructifications are
16 PHYTOMORPHOLOGY January–June 2010
Fig. 3: A – Reconstruction of Sahnia nipaniensis (Little modified from Suthar & Sharma 1988). Fructification is terminal on a shortshoot bearing closely placed bases of leaves / bracts. Note large sized bracts which protected young microsporophylls in the bud stage.Microsporophylls radial, branched or unbranched with microsporangia in rows. Microsporophylls arise spirally. B – Carnoconiteslaxum Longisection of a cylindrical cone with orthotropus ovules / seeds attached directly to the cone aixs x 6. C – C. compactuslongisection, orthotropus ovules attached directly to the cone axis x 4. D – Reconstruction of a cone of C. compactus. Surface view ofthe cone. Hexagonal seeds attached spirally and each has a narrow central micropyle. E – Reconstruction of a group of cones ofC. compactus. Cones oval, pedicellate attached spirally on flashy peducle which borns terminally on a short shoot (Little modified fromVishnu-Mittre 1953). F – Carnoconites llambiasii. Note attachment of cones and size of seeds (After Cesari et al. 1998) Quite distinct fromC. compactus Sriv.

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 17
terminal on short shoots. Male cone consists of radialmicrosporophylls with solitary microsporangia in rows.Seeds/ovules are attached directly to cone axis,orthotropus, integument thick heterogeneous and non-vascularized and nucellus free without pollen chamber.
Stewart (1976) did not agree with the polystelicvasculature in the Pentoxyleae and described divisionor fission of a monostele into 5 or 6 bundles. The authorshave collected hundreds of specimens of Pentoxylonstem and short shoots in cherts from Nipania and Sonajori,and individual stem pieces from Amarjola, and preparedseveral hundred slides from them. However, division orfission of a monostele into 5 or 6 steles of regular shapesand sizes was not observed in any of them. This is aconstant character of stem, and in all kinds of shortshoots, and we feel that reconsideration of Stewart’sconcept is therefore suggested. Similarly correlation ofanatomy of Pentoxylon with that of Pandanaceae(angiosperm) by Meeuse (1961) is also an imaginativehypothesis.
In Nipanioxylon guptai there are 7-8 steles presentsurrounding a large pith. In Pentoxylon secondary woodis more developed towards the pith whereas, inNipanioxylon it may be reverse or the steles are concentricwith crushed primary xylem. The secondary wood iscompact like that of Pentoxylon. Sahni (1948) consideredNipanioxylon to be a shoot system of Pentoxylon andnot a separate taxon. There is no doubt that Vishnu-Mittre’s (1957) specimen of Nipanioxylon was a coniferwood. Bose et al. (1985), Sharma (1996, 2001) and thepresent investigation favour Srivastava (1944, 1945) inconsidering Nipanioxylon a distinct genus. However,further investigations are required and search should becontinued for the collection of better preserved specimensof Nipanioxylon from Nipania.
Guptioxylon is certainly a pentoxylean stem distinctfrom Pentoxylon and Nipanioxylon in having medullarybundles and in peculiar structure of cortical bundles(Sharma, 1969a, 1972, 1974). In G. amarjolense, themain four steles are well developed but slightly irregularin shape. On the other hand in G. endocentrica theseare of comparatively regular shapes and arrangedsurrounding the pith as in Pentoxylon (Sharma, 1972).The present investigation favours Sharma’s opinion(1996, 2001) in deriving Pentoxylon anatomy fromMedullosa like stem through G. amarjolense andG. endocentrica. Purioxylon Sharma (1972a) is a distinctgenus having the characters of cycads on the one handand Pentoxyleae on the other hand. The vascular cylinder
is made up of many collateral and conjoint bundles likethe cycads whereas, the cortical bundles have compactsecondary xylem identical to the cortical bundles of thePentoxyleae.
Nipanophyllum raoi Sahni (including N. hirsutumVishnu-Mittre, N. anomozamoides Sharma and N.hobsonii Bose et al.) resembles Taeniopteris spatulatain external morphology. However, differs in anatomy,which decides the pentoxylean affinity i.e. presence of3-7 diploxylic bundles in an arc or a row in the midrib.The specimens of leaves described from New Zealand,Queensland (Australia) and Antarctica are known onlyas impressions or incrustations as such, the anatomy ofmidrib remains unknown and thus their cycadean affinitycannot be ruled out.
The male fructification Sahnia nipaniensis iscomparatively rare in occurrence in the Nipania chert.The validity of a reconstruction is based on the descriptionof the slides and specimens of the organ or organs.Vishnu-Mittre (1953) showed solitary microsporangiain linear rows on the microsporophylls whereas, Boseet al. (1985) on the other hand showed the attachmentof microsporangia in bunches on small lateral shoots.Taylor (1988) also figured attachment of microsporangiain bunches on short lateral shoots. Osborn (1988) andSuthar and Sharma (1988) agreed with the descriptionof Vishnu-Mittre (1953) and suggested reconstructionsof male fructifications. The present reconstruction isslightly modified of after Suthar and Sharma (1988,Fig. 2). These changes are made on the basis of studyof additional three slides of the male fructificationavailable at the conservatory of B.S.L.P., Lucknow ofthe male fructification. We could neither see attachmentof microsporangia in bunches nor on small lateral shoots.The presence of large sized bracts are also made in thereconstruction (Fig. 3A). These bracts protected/coveredyoung microsporangia during bud stage. In a medianlongisection (Suthar & Sharma, 1988, Fig. 2) two largebracts, one on either side are visible.
A number of reconstructions are proposed for theseed bearing fructification Carnoconites compactus(Sahni, 1948; Vishnu-Mittre, 1953; Bose, et al., 1985;Crane, 1985; Srivastava & Banerji, 2006). Sharma (2001)has given critical remarks on all these fructifications andfavoured Sahni’s interpretation (Sahni, 1948, Fig. 46).We do not know if some body has a slide bearinghundreds of seed cones in a bunch. Atleast neither wehave nor available at the Birbal Sahni Institute ofPalaeobotany, Lucknow. Maximum of 14-20 cones are

18 PHYTOMORPHOLOGY January–June 2010
seen in a bunch on a peduncle. Canoconites llambiasiiCesari et al. (1998) from Antarctica (Fig. 3F) neitherresembles in shape and size of seeds nor in the mannerof attachment of cones to the peduncle with C. compactus.The transfer of C. laxum to C. rajmahalensis Boseet al., (1984) is also not justified (Sharma, 2001) andneeds reconsideration.
The Pentoxyleae includes unique type offructifications both male and the seed bearing. The malehas spirally attached, radial, branched and unbranchedmicrosporphylls bearing balloon shaped microsporangiain rows. This fructification as such can neither be relatedto cycads nor the Bennettitales nor any other group ofextinct or extant plants. Cladistic investigations relatePentoxyleae with the bennettitalean clade which is alsorelated with the flowering plants and Gnetophyta (Crane,1985, 1988; Doyle & Donoghue, 1986). How muchuseful is this cladistic approach? We do not know. Sinceneither in morphology nor in anatomy the Pentoxyleaeresembles the Bennettitales, Gnatales and the floweringplants. The pentoxylean plants do show relationship withthe medullosean pteridosperms, cycads, Ginkgoales andthe conifers. It is a synthetic group of extinct Mesozoicplants and needs further investigations.
Literature citedBose, M.N., Pal, P.K. & Harris, T.M. 1984. Carnoconites
rajmahalensis (Wieland) Comb. Nov. from the Jurassic ofRajmahal Hills, India. Palaeobotanist, 32: 368-369.
Bose, M.N., Pal, P.K. & Harris, T.M. 1985. The Pentoxylon plant.Philosophical Transactions of Royal Society London, 110B:77-108.
Cesari, S.N., Parica, C.A., Remesal, M.B. & Salani, F.M. 1998. Firstevidence of Pentoxylales in Antarctica. Cretaceous Research, 19:733-743.
Crane, P.R. 1985. Phylogenetic relationships in seed plants. Cladistics,1: 329-348.
Crane, P.R. 1988. Major clades and relationships in the “Higher”gymnosperms. In Beck, C.B. (ed.) Origin and Evolution ofGymnosperms, Columbia Univ. Press, New York. pp.218-272.
Douglas, J.G. 1969. The Mesozoic Flora of Victoria. Memoirs ofGeological Survey Victoria, 28: 1-310.
Doyle, A. & Donoghue, M.J. 1986. Seed plant phylogeny and theorigin of angiosperms. An experimental cladistic approach.Botanical Review, 52: 321-431.
Drinnan, A.N. & Chambers, T.C. 1985. A reassessment of Taeniopterisdaintreei from the Victorian Early Cretaceous, a member of thePentoxyleae and a significant Gondwana plant, Australian J.Botany, 33: 89-100.
Harris, T.M. 1962. The occurrence of the fructification Carnoconitesin New Zealand. Philosophical Transactions of Royal Society,New Zealand, 1: 17-27.
Harris, T.M. 1982. Fossils from New Zealand ascribed in thePentoxylon plant, Phyta-studies on living and fossil plants (Pantcomm. vol.) Allahabad. pp. 91-103.
Meeuse, A.D.J. 1961. The Pentoxyleae and the origin ofmonocotyledons, Proc. K. ned. Akad. Wet. Ser. C, 64: 545-559.
Osborn, J.M., Taylor, T.N. & Crane, P.R. 1991. The ultra structure ofSahnia pollen (Pentoxyleae). American Journal of Botany, 78:1560-1569.
Pant, D.D. 2002. Gymanosperms, Cycas and Cycadales BSIPmonograph No. 4. Birbal Sahni Institute of Palaeobotany Lucknow.India.
Rao, A.R. 1943. The structure and affinities of Taeniopteris spatulataMc Cl. Proceedings of National Academy of Sciences India, 13:335-355.
Sahni, B. 1948. The Pentoxyleae-a new group of Jurassic gymno-sperms from the Rajmahal Hills of India. Botanical Gazette,110: 47-80.
Sharma, B.D. 1969c. On Pentoxyleae remains from Amarjola in theRajmahal Hills, India. Ameghiniana, 6: 50-56.
Sharma, B.D. 1969a. Guptioxylon amarjolense gen. et sp. nov. fromAmarjola in the Rajmahal Hills. Palaeotographica Abt,B 126 (4-6): 145-153.
Sharma, B.D. 1969b. On some fossil cycadean fronds from India.Bulletin of Botanical Survey of India. 11: 115-119.
Sharma, B.D. 1972. Guptioxylon endocentrica sp. nov. du Jurassic d’Amarjola dans les Rajmahal Hills (inde). Bulletin of SocietyLinnaeus Lyon, 41: 114-120.
Sharma, B.D. 1972a. Purioxylon Jurassic gen. et sp. nov. fromAmarjola in the Rajmahal Hills, India. Advances in PlantMorphology (Puri Comm vol): pp. 233-242.
Sharma, B.D. 1973b. Further observations on Pentoxylon sahnii Sriv.from the Jurassic of Amarjola in the Rajmahal Hills, India.Palaeobotanist, 20: 216-220.
Sharma, B.D. 1973a. On the anatomy of dwarf shoot of Pentoxylonsahnii Sriv. collected from Amarjola, Rajmahal Hill, India. ActaPalaeobotanica, 14: 195-206.
Sharma, B.D. 1974b. Pentoxylon and allied fossil woods fromAmarjola in the Rajmahal Hill, India. Bulletin of Natural ScienceMuseum, Tokyo, 17: 75-86.
Sharma, B.D. 1974a. Observations on branching in Pentoxylon sahnii.Bulletin of Natural Science Museum, Tokyo, 17: 315-324.
Sharma, B.D. 1975. Further observations on the fossil flora of Nipaniain the Rajmahal Hills, India. Ameghiniana, 12: 329-336.
Sharma, B.D. 1979. Further observations on the dwarf shoot ofPentoxylon sahnii Sriv. collected from the Jurassic of RajmahalHills India. Acta Palaeobotanica, 20: 129-136.
Sharma, B.D. 1980. Further observations on branching in Pentoxylonsahnii Sriv. Indian Journal of Earth Sciences, 7: 100-102.
Sharma, B.D. 1982. Studies on the transfusion cells in the petrifiedleaves of Ptilophyllum and Nipanioxylon from the Rajmahal Hills,India. Palaeobotanist, 30: 181-184.
Sharma, B.D. 1989. Possible occurrence of polyembryony inPentoxyleae. Phytomorphy, 39: 199-201.
Sharma, B.D. 1996. The Pentoxyleae-an overview. Palaeobotanist,45: 50-56.

B.D. SHARMA et al. – Present Status of the Pentoxyleae – the Mesozoic Gymnosperms 19
Sharma, B.D. 2001. Misinterpretations about the “Pentoxyleae” – aMesozoic gymnospermous group of plants. Palaeobotanist, 50:255-265.
Sharma, B.D. 2003. Pentoxyleae in Antarctica? Needs areconsideration. Geophytology, 31: 103-104.
Sharma, B.D. & Bohra, D.R. 1976. A new assemblage of fossil plantsfrom the Jurassic of Rajmahal Hills, India, Geobios (France) 9:111-123.
Sharma, B.D. & Bohra, D.R. 1977. Structure of phloem in someplants of Bennettitales and Pentoxylales collected from theRajmahal Hills, India, Geophytology, 7: 214-216.
Sharma, B.D., Bohra, D.R. & Suthar, O.P. 1987. The Phylogeny ofPentoxyleae. Facena, 7: 5-14.
Sharma, B.D., Bohra, D.R. & Suthar, O.P. 2001. Some interestingplant fossils from the Mesozoic rocks of the Rajmahal Hills,India, Palaeabotanist, 50: 207-212.
Sharma, B.D., Bohra, D.R. & Suthar, O.P. 2006. Mucilage canals inthe bennettitalean remains from the Rajmahal Hills, Jharkhand,India, Geophytology, 36: 47-52.
Srivastava, B.P. 1937. Studies on some silicified remains from theRajmahal series of India. Proc. 24th Indian Sci. Congr. Hyderabad,273-274 (Abstract).
Srivastava, B.P. 1944. Silicified Plant remains from the RajmahalHills. Palaeobotany in India 5, Proc. Natn Acad Sci. India, 14:73-76.
Srivastava, B.P. 1945. Silicified plant remains from the RajmahalHills, India, Proc. Natn. Acad. Sci., India, 15: 185-211.
Srivastava, S.C. & Banerji, J. 2000. Pentoxylon Plant: a reconstructionand interpretations, Plant Cell Biol & Devpt, 13: 11-18.
Stewart, W.N. 1976. Primary xylem and the Pteropsida, Prof. ACSeward Mem. Lecturer 1-13. B.S.I.P., Lucknow, India.
Stewart, W.N. & Rothwell, G.W. 1993. Palaeobotany and the Evolutionof Plants. Cambridge Univ. Press, New York.
Suthar, O.P. & Sharma, B.D. 1988. A new interpretation of the structureof Sahnia nipaniensis Vishnu-Mittre from the Rajmahal Hills,India, Palaeobtanist, 37: 90-93.
Suthar, O.P., Sharma, B.D. & Bohra, D.R. 1988. Record of an additionalshoot system in Pentoxylon sahnii Sriv. from the Rajmahal Hills,India, J. Earth Sci., 15: 75-78.
Taylor, T.N. 1988. Pollen and pollen organs of fossil gymnosperms.Phylogeny and reproductive biology, In Back CB (ed), Originand Evolution of Gymnosperms, Columbia Univ. Press, NewYork.
Taylor, T.N. & Taylor. E.L. 1993. The Biology and evolution of fossilplants. Prentice Hall Inc., New York.
Vishnu-Mittre, 1953. A male flower of the Pentoxyleae with remarkson the structure of the female cones of the group, Palaeobotanist,2: 75-84.
Vishnu-Mittre, 1957. Studies on the fossil flora of Nipania (RajmahalSeries), India-Pentoxyleae, Palaeobotanist, 6: 31-45.
White, M.E. 1981. Revision of the Tal brager fish bed flora (Jurassic)of New South Wales, Records Aust. Mus., 33: 695-721.

A. RAJANIKANTH et al. – An Integrated Inquiry of Early Cretaceous Flora, Palar Basin, India 21Phytomorphology 60 (1 & 2) 2010, 21-28
IntroductionThe east coast of India encompasses a number ofunconnected outcrops often referred ‘CoastalGondwanas’. These continental rock sequences withthin marine intercalations were given ‘Upper Gondwana’status, which include co-equivalent, paralic and lagoonalMesozoic sediments distributed in detached outcropsparallel to the shoreline. These sediments distributed indifferent basins/grabens - Cauvery, Palar, Krishna-Godavari, Pranhita-Godavari and Mahanadi arecharacterized by Ptilophyllum flora (Venkatachala, 1977;Bose et al., 1990; Rajanikanth et al., 2000). The geologyand stratigraphy of different sedimentary basins of eastcoast are relatively well known (Foote, 1873; Oldham,1893; King, 1958; Sastry et al., 1981; Dutta et al., 1983;Lal et al., 2009; Kumaraguru et al., 1994; Vaidyanadhan& Ramakrishanan, 2008).
The Palar Basin is one of the seventeen riverbasins in Tamil Nadu. It covers an area of about 18,300sq km extending to Andhra Pradesh and Karnataka. ThePalar Basin basement is composed of an Archeanmetamorphic complex overlain by the Gondwanasediments - Fluvio-glacial deposits of Early Permian
An Integrated Inquiry of Early Cretaceous Flora, Palar Basin, IndiaA. Rajanikanth1, Anil Agarwal1 and A. Stephen2*
1 Birbal Sahni Institute of Palaeobotany, 53 University Road, Lucknow-226007, India2 French Institute of Pondicherry, 11, St. Louis Street, PB 33, Pondicherry-605001, India
ABSTRACT
The Palar Basin is one of the 17 river basins that exist in India. The basin embodies an Archean metamorphic complexoverlain by the Gondwana sediments represented by the Lower Gondwana sequence (Fluvio-glacial deposits of EarlyPermian) with a northeast-southwest trend. The Sriperumbudur Formation, named after Sriperumbudur town in SouthIndia, near Chennai constitutes a part of Early Cretaceous outcrops along the east coast and characterized by marineintercalations in between the fresh water Upper Gondwana sequence. The Sriperumbudur Formation is exposed in thePalar Basin is about 600 m thick and characterized by the splintery, gray and greenish shales containing dark-gray gypseousclay and interbedded sandstone, ironstone and limestone. The palaeovegetational diversity and phytogeographic distributionof the Sriperumbudur flora are discussed in comparison with various coeval floras of Indian peninsula along with theother Gondwanan regions. The palynofloral assemblage comprises spore/pollen assignable to bryophytes, pteridophytesand gymnosperms. The plant megafossil evidences when viewed in conjunction with faunal and palyno floral evidencesindicate an Early Cretaceous age. The palaeogeographic analysis suggests occurrence of post-gondwanan cosmopolitanflora (pre-angiosperms) in the east coastal regions during Early Cretaceous times and comparable with other gondwanancontinental floras.
Key words: Early Cretaceous, biodiversity, palaeovegetation, palaeoclimate, Sriperumbudur, Palar Basin, India
Author for correspondence e-mail: [email protected]
(Lower Gondwana). The Upper Gondwana sequencewas represented by the Sriperumbudur beds (EarlyCretaceous) characterized by marine intercalations. Thesucceeding sequence deposited under littoral to nearshore fluvial conditions (Avadi and Satyaveduformations). The Tertiary sequence is corresponded bythe Cuddalore Sandstone. This in turn is overlain by theKanchivaram Gravel and Pliestocene laterites andconglomerates of fluvial nature. On the top lie theHolocene alluvial sands and clays. The early CretaceousSriperumbudur Formation is characterized by arenaceousand argillaceous rock units comprising splintery greenshale, clays and sandstones with ironstone intercalationsand on conformably overlying either the Precambrianbasement or Precambrian boulder beds and green shales.The beds contain marine intercalations (Murthy & Sastry,1961). Their lithologic suites and fossil fauna aresuggestive of deposition under shallow and brackishconditions, probably close to the shoreline (Sastry et al.,1974). The present communication presents a holisticinquiry of plant evidences known from the EarlyCretaceous of Palar Basin incorporating the recoveredtaxa (see Figs. 1 & 2, Tables 2 & 3).

22 PHYTOMORPHOLOGY January-June 2010
General GeologyThe Palar Basin is one of the seventeen river basins inTamil Nadu. It covers an area of 18,300 sq. km. extendingto Andhra Pradesh and Karnataka. The Palar is a seasonalriver and for the most part of the year it is dry. The PalarBasin (Sastri et al., 1974; Rangaraju et al., 1993; Vairavan,1993; Kumaraguru, 1991, 1992) basement is composedof an Archean metamorphic complex overlain by theGondwana sediments represented by the LowerGondwana sequence (Fluvio-glacial deposits of EarlyPermian). The Upper Gondwana sequence wasrepresented by the Sriperumbudur beds (EarlyCretaceous) characterized by marine intercalations. Thesucceeding sequence deposited under littoral to nearshore fluvial conditions (Satyavedu beds). The Tertiarysequence is corresponded by the Cuddalore Sandstone.This in turn is overlain by the Kanchivaram Gravel andPliestocene laterites and conglomerates of fluvial nature.The Sriperumbudur Formation is characterized byarenaceous and argillaceous rock units comprisingsplintery green shale, clays and sandstones with ironstoneintercalations and on conformably overlying either thePrecambrian basement or Precambrian boulder beds andgreen shales. The beds contain marine intercalations(Murthy & Sastry, 1961). Their lithologic suites andfossil fauna are suggestive of deposition under shallowand brackish conditions, probably close to the shoreline(Sastry et al., 1974).
Table 1. General Stratigraphic Sequence of Palar Basin
Age Gross LithologyHolocene
Pleistocene
Mio-Pliocene
Palaeogene
Upper Cretaceous
Early Cretaceous
Permian
Archean
Fossil assemblages of Sriperumbudur FormationThe mega plant fossils were mostly preserved in the formof fossil leaves represented by the species of Cladophlebis(Pteridophytes), Dictyzamites, Taeniopteris,Pterophyllum (Cycadophytes), Araucarites, Conites,(Coniferales), Ginkgoites (Ginkgoales) and petrifiedwood fossils belonging to conifers (Fiestmantel, 1879;Seward & Sahni, 1920; Sahni, 1928, 1931;Suryanarayana, 1954, 1956). Several species ofpycynoxylic wood belonging to the conifers were alsoreported from the upper Gondwana sediments of theSriperumbudur Formation, Palar Basin in Tamil Nadu,India. These include Cupressinoxylon coromandelinum,Mesembrioxylon sp. (Sahni, 1931), M. thirumangalense(Suryanarayana, 1953), Araucarioxylon giftii,Araucarioxylon rajivii (Jeyasingh & Kumarasamy,1994a), Araucarioxylon mosurense (Jeyasingh &Kumarasamy, 1995). Besides, Pityospermum Nathorsttoo was recorded from the Sriperumbudur Formation(Jeyasingh & Kumarasamy, 1994b). The floralassemblage in general is dominated by Conifers followedby Cycadophytes and ferns. Pteridosperms andGinkgoales are poorly represented. Foote (1868)compared Sriperumbudur megaflora with that ofRajmahal. A Jurassic affinity to this flora was alsosuggested (Fiestmantel, 1879). Besides extensive workon surface and subsurface sequences of this Formationhave yielded rich palyno assemblage characterized bythe forms - Aequitriradites, Coptospora, Cooksonites,Foraminisporis, Staplinnisporites, Sestrosporites,Ornamentifera, Klukisporites, Impardecispora,Cicatrisporites, Undulatisporites, Coronatisporia,Polycingulatisporites, Taurocusporites, Crybelosporites,Murospora and Micrcachrydites (Ramanujam &Srisailam, 1974; Varma & Ramanujam, 1984).Sriperumbudur palynoflora shows significantresemblance with the Early Cretaceous palynoflora fromCauvery and Krishna Godavari basins. The palynofloraknown from both the surface and subsurface is suggestiveof Early Cretaceous age (Varma & Ramanujam, 1984;Ramanujam & Srisailam, 1974; Ramanujam & Varma,1977, 1981). An Early Cretaceous fauna in the form ofammonites Pascoites crassus and forams - Pelosinacomplaneta, Haplophragmoides concave, H. footei, H.indicus, Bathysiphon cf. taurinensis, Ammodiscuscretaceous, Lituotuba sp. and Spiroplectammina indicawas also recorded (Murthy & Sastry, 1961).
The present paper deals with leaf flora ofPtilophyllum, Elatocladus, Pagiophyllum and associated
Blown sands adjoining the sea coast and*river alluvium/ marine and estuarinealluviumLaterites and lateritic conglomeratescontaining sandstones, pebblesCuddalore sandstones, conglomeratic andclayey sandstones, yellowish purple andwhite variagations, contains quartz pebblesBlack clay stones and bands of thinsandstonesSatyavedu beds, coarse bolders beds,conglomerates, compact fine grainedsandstonesSriperumbudur beds, splintery gray andgreenish shales, dark clays, partly gypseous,inter bedded with sandstones and thin bandsof ironstone and limestone (UpperGondwana)Boulders beds and greenish shales correlatedto Talchir shale of Lower GondwanaIgneous and metamorphic basement
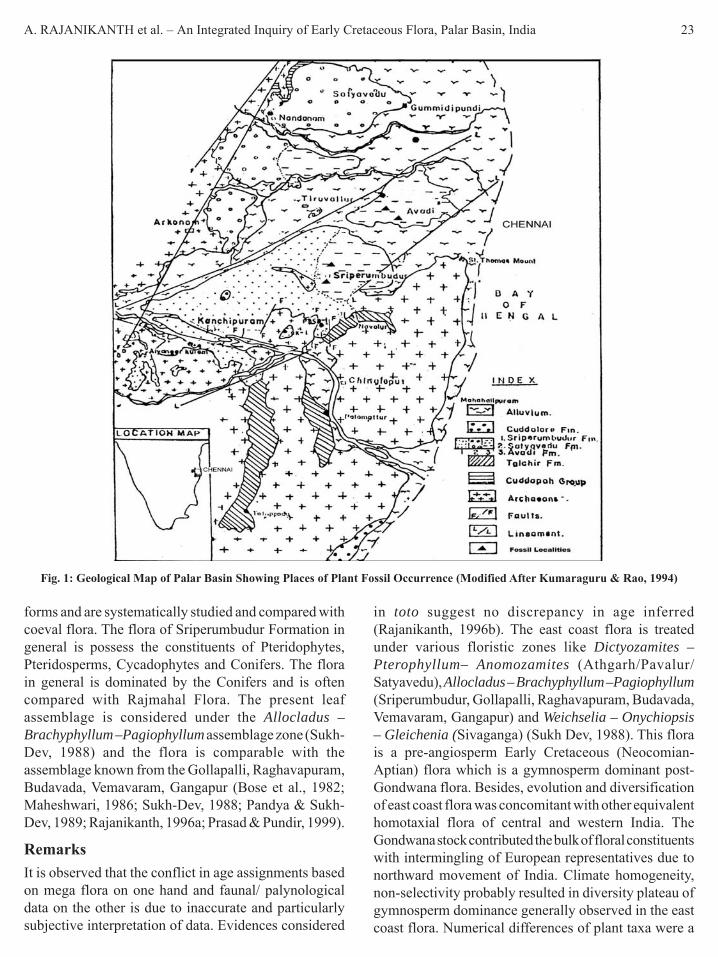
A. RAJANIKANTH et al. – An Integrated Inquiry of Early Cretaceous Flora, Palar Basin, India 23
forms and are systematically studied and compared withcoeval flora. The flora of Sriperumbudur Formation ingeneral is possess the constituents of Pteridophytes,Pteridosperms, Cycadophytes and Conifers. The florain general is dominated by the Conifers and is oftencompared with Rajmahal Flora. The present leafassemblage is considered under the Allocladus –Brachyphyllum –Pagiophyllum assemblage zone (Sukh-Dev, 1988) and the flora is comparable with theassemblage known from the Gollapalli, Raghavapuram,Budavada, Vemavaram, Gangapur (Bose et al., 1982;Maheshwari, 1986; Sukh-Dev, 1988; Pandya & Sukh-Dev, 1989; Rajanikanth, 1996a; Prasad & Pundir, 1999).
RemarksIt is observed that the conflict in age assignments basedon mega flora on one hand and faunal/ palynologicaldata on the other is due to inaccurate and particularlysubjective interpretation of data. Evidences considered
in toto suggest no discrepancy in age inferred(Rajanikanth, 1996b). The east coast flora is treatedunder various floristic zones like Dictyozamites –Pterophyllum– Anomozamites (Athgarh/Pavalur/Satyavedu), Allocladus – Brachyphyllum –Pagiophyllum(Sriperumbudur, Gollapalli, Raghavapuram, Budavada,Vemavaram, Gangapur) and Weichselia – Onychiopsis– Gleichenia (Sivaganga) (Sukh Dev, 1988). This florais a pre-angiosperm Early Cretaceous (Neocomian-Aptian) flora which is a gymnosperm dominant post-Gondwana flora. Besides, evolution and diversificationof east coast flora was concomitant with other equivalenthomotaxial flora of central and western India. TheGondwana stock contributed the bulk of floral constituentswith intermingling of European representatives due tonorthward movement of India. Climate homogeneity,non-selectivity probably resulted in diversity plateau ofgymnosperm dominance generally observed in the eastcoast flora. Numerical differences of plant taxa were a
Fig. 1: Geological Map of Palar Basin Showing Places of Plant Fossil Occurrence (Modified After Kumaraguru & Rao, 1994)

24 PHYTOMORPHOLOGY January-June 2010
Table 2. Plant Mega Fossils of the Palar Basin
Mega plant taxa Sriperum Satya Avadibudur Fm. vedu Fm. Fm.
Cladophlebis sp. + + +C. indica + – –C. whitbyensis + – –Equisetites sp. – + –Thinnfeldia sp. – – –Taeniopteris sp. + + +Taeniopteris Mc Clellandii + – –T. spatulata – + –Anomozamites sp. – + –Anomozamites lindleyanus + – –Pterophyllum sp. + – +Pterophyllum footeanum + – +Ptilophyllum acutifolium. + + +Ptilophyllum cutchense + + +Ptilophyllum sp. + – –Dictyozamites sp. + – –Otozamites.rarinervis + – –Elatocladus plana + – +E. conferta + – –E. tennerrima + – +Pagiophyllum peregrinum + – +Brachyphyllum rhombicum + – –B. rajmahalensis + – –Araucarites. cutchensis + – +A. macropterus + – –A minutus + + +Pityospermum sp. + – –Conites sessiles + – +C.sripermaturensis + – –C. verticillatus + – –Ginkgoites sp. + – +G. crassipes + – –Araucarioxylon giftii + – –Podocarpoxylon parthasarathyi + – –P. tirumangalense + – –Cupressinoxylon coromandelinum + – –Araucarioxylon mosurense + – –Araucarioxylon rajmahalense + – –
Table 3. Plant Micro Fossil Records of Palar Basin
Palyno Taxa Sriperumbudur Fm.BryophytesAequitriradites dubius +Aequitriradites spinulosusAequitriradites verrucosusAequitriradites sp.Cooksonites variabilis +Cooksonites sp.Coptospora cutchense +Coptospora cauverianaCoptospora spForaminisporis sp. +
PteridophytesAppendicisporites sp.Baculatisporites baculatus +Baculatisporites clavaeoides +Baculatisporite comaumensis +Biformaesporites baculosusBiretisporites sp. +Biretisporites potniaei +Biretisporites spectabilisCallispora sp. +Callispora foveolataCallispora potonieiCeratosporites sp. +Ceratosporites couliensisCeratosporites equalis +Cicatricosisporites sp. +Cicatricosisporites apicanalisCicatricosisporites augustusCicatricosisporites australiensisCicatricosisporites goniodontos +Cicatricosisporites hallei +Cicatricosisporites hughesiCicatricosisporites ludbrooki +Cicatricosisporites mohrioidesContignisporites cooksoniiConcavisporites cutchensis +Contignisporites dorsostriatus +Contignisporites fornicatusContignisporites glebulentusContignisporites multimuratusContignisporites problematicusCoptospora cauverianaCoptospora kutchensisConverrucosisporites sp. +Coronatispora sp.Coronatispora perforata +Crassimonoletes sp. +Crassimonoletes surangei +Crybelosporites sp. +Crybelosporites punctatusCyathidites australisCyathidites minorDeltoidospora diaphana +Deltoidospora juncta +Dictyophyllidites venkatachalai +Dictyotosporites complex
result of taphonomic bias influenced by sedimentologicalprocesses dictated by tectonic forces and sea levelinfluence. Plant evidences (Mega-Micro) indicate uplandnear basinal and marginal floral components. The eastcoast flora represents a unique-mixed flora preservedon the passive east coast margins as a result ofsedimentation at the time of initiation of rifting of Indianplate (Yoshida et al., 1992; Ziegler et al., 1983). Thismixed flora is characterized by the constituents ofPtilophyllum flora spread in the Cauvery, Palar, Krishna-Godavari/Pranhita-Godavari and Mahanadi basins.Distribution pattern of this flora in these sedimentary

A. RAJANIKANTH et al. – An Integrated Inquiry of Early Cretaceous Flora, Palar Basin, India 25
Palyno Taxa Sriperumbudur Fm.Distalanulisporites sp.Densoisporites sp.Echinatisporis vembanii +Eoseisporites sp. +ForaminisporitesFoveosporites sp. +Foveotriletes subtriangularisFoveotrietes sp. +Gleicheniidites sp.Gleicheniidites cercinidites +Gleicheniidites senonicus +Impardecispora apiverrucatus +Impardecispora marylandensis +Impardecispora trioreticulosus +Ischyosporites crateris +Ischyosporites punctatusKlukisporites areolatusKlukisporites foveolatusKraeuselisporites sp. +Kraeuselisporites cf. linearisLametatriletes sp. +Leptolepidites sp. +Leptolepidites verrucatus +Lycopodiumsporites austroclavatidites +Lycopodiumsporites eminulus +Lycopodiumsporites sp. cf.trambauensis +Matonosporites crassiangulatusMatonosporites discoidalisMatonosporites phlebopteroidesMatonosporites sahiiMetamonoletes sp.Metamonoletes baradensis +Monolites indicus +Monoletes ingranulatus +Murospora sp. +Murospora cf. mesozoicaNeoraistrickia truncatus +Ornamentifera sp. +Ornamentifera echinataOsmundacidites sp.Osmundacidites singhii +Osmundacidites wellmaniiPilosisporitesPilosisporites sp.Plicifera minutus +Polycingulatisporites sp. +Polycingulatisporites reduncusSchizosporis regulatusSestrosporites sp. +Setrosporites pseudoalveolatusSpheripollenites sp. +Staplinisporites sp. +Staplinisporites caminusStereisporites antiquasporites +Taraucosporites sp. +Taurocusporites segmaentatus
Palyno Taxa Sriperumbudur Fm.Todisporites crassus +Todisporites minorTodisporites rotundiformisTrilites orikkaiense +Trilites tuberculiformis +Trilites verrucosus +Undulatisporites pannuceus +Verrucosisporites sp. +Verrucosisporites rotundus
GymnospermsAlisporites grandis +Alisporites ovalisAlisporites cf. rotundusAraucariacites australis +Baculatisporites clavaeoidesCallialasporites dampieriCallialasporites discoidalisCallialasporites kattivakkaamense +Callialasporites punctatusCallialasporites reticulatusCallialasporites segmentatusCallialasporites trilobatusCedripites cretaceusCedripites nudisClassopollis sp.Classopollis classoidesClassopollis obidosensisCycadopites sp.Cycadopites couperiCycadopites sakrigaliensisDacrycarpites sp. +Dacrycarpites australiensisEchinatispori varispinosusEchinatispori vembaniiGranuloperculatipollis +Granuloperculatipollis sp.Laricoidites indicus +Laricoidites communisMicrocachrydites sp.Microcachryidites antarcticus +Osmundacidites singhiiOsmundacidites wellmaniiPhyllocladidites sp. +Phyllocladidites inchoatusPlatysaccus sp.Platysaccus densusPodocarpidites ellipticusPodocarpidites majorPodocarpidites miniuulcusPodosporites sp.Podosporites tripakshiPsilospora sp.Spheripollenites sp. +Spheripollenites scabratusVitreisporites sp.Vitreisporites pallidus

26 PHYTOMORPHOLOGY January-June 2010
Table 4. Faunal Records of Palar Basin
Taxa Sriperumbudur Satyavedu AvadiOstrea (Bivalve) – – +Pectan (Bivalve) – + +Lima (Bivalve) – – +Inoceramus (Bivalve) – – +Cardium (Bivalve) – – +Peratabelus sp. (Belemnite) – – +Gymnoplites simplex (Ammonites) – – +Pascoeites crassus (Ammonites) + – +Ammobaculites sp. (Foraminifers) + – +Ammodiscus cretaceous(Foraminifers) + + –Ammodiscus sp. (Foraminifers) + – +Bathysiphon cf taurinense(Foraminifers) + – –Bathysiphon sp. (Foraminifers) – – +Haphlophragmoides sp.(Foraminifers) – – +Haphlophragmoides dickinsonii(Foraminifers) + – –Haphlophragmoides concava(Foraminifers) + – –Haphlophragmoides footei(Foraminifers) + + –Haphlophragmoides indicus(Foraminifers) + + –Pelosina complanata(Foraminifers) + – +Lituotuba sp. (Foraminifers) + – –Spiroplectammina indica(Foraminifers) + – –
basins and its implication on evolution, ecology and agecorrelation has been analyzed (Rajanikanth, 1996b, 2007).This mixed flora, a precursor of flowering plants showedlocal variations and most of the forms got extinct ormigrated with the arrival of later new flora. This EarlyCretaceous flora shows a cosmopolitan affinity. It is alsoobserved that most of the Ptilophyllum floral bearingsediments were deposited during the Early Cretaceousand even a continuous succession with the UpperCretaceous marine sediments is encountered in the eastcoast as evident from the sub-surface data from theKrishna-Godavari and Cauvery basins (Acharyya &Lahiri, 1991). Occurrence of marine Albian calcareousalgae above the continental Neocomian-Aptian beds ofCauvery Basin also indicate marine intervention (Misraet al., 2004). Both Lower and Upper Cretaceous marinesediments along the east coast were thus deposited byone and the same marine transgression (Venkatachalaet al., 1993; Rajanikanth et al., 2000). The east coastfloral assemblages are characterized by paucity of leafcuticles and preponderance of spore/pollen, whichindicate a certain distance of transport. Fossil florasshow taphonomically biased distribution governed bydepositional environment. Plant parts in the form ofleaves, woods, axes, spores, cuticles and pollen arepreserved in different depositional milieu. Thus the floraknown from the Sriperumbudur, Avadi and Satyavedulitho units was in essence the same flora that prevailedover the globe during early Cretaceous and representpre-angiosperm post-gondwanan cosmopolitan flora (seeTables 2, 3 & 4).
Fig. 2: (A-F) Plant Fossils From Palar BasinA – Ptilophyllum acutifolium. B – Otozamites sp.. C – Otozamitessp.. D – Ptilophyllum acutifolium. E & F – Elatocladus plana.
AcknowledgementsAR & AA wishes to express their gratitude to Dr. NCMehrotra, Director, BSIP for encouragement andproviding necessary help. AS wishes to thank Dr. DEPJeyasingh, Prof & Head (Retd.), Madras Christian Collegefor his help in preliminary identification of fossil samples.
ReferencesAcharyya, S.K. & Lahiri, T.C. 1991. Cretaceous palaeogeography
of the Indian subcontinent: a review. Cretaceous Research, 12:3-26.
Bose, M.N. 1966. Plant remains from the Rajmahal and Jabalpurseries in the Upper Gondwana of India: In Proc. Symposiumon floristics and stratigraphy of Gondwanaland, Birbal SahniInstitute of Palaeobotany, Lucknow, 143-154.
Bose, M.N., Kutty, T.S. & Maheswari, H.K. 1982. Plant fossilsfrom the Gangapur formation. The Palaeobotanist, 30(2):121-125.

A. RAJANIKANTH et al. – An Integrated Inquiry of Early Cretaceous Flora, Palar Basin, India 27
Bose, M.N., Taylor, E.L. & Taylor, T.N. 1990. Gondwana floras ofIndia and Antarctica a survey and appraisal In: T. N. Taylor &E. L. Taylor (Eds.), Antarctic Paleobiology: its role in thereconstruction of Gondwana. Springer-Verlag, New York, USA,PP. 118-148.
Dutta, N.R., Mitra, N.D. & Bandyopadhyay, S.K. 1983. Recent trendsin the study of Gondwana basins of peninsular and extra-peninsularIndia. Petroleum Asia Journal, 6(4): 159-169.
Fiestmantel, O. 1879. The fossil flora of the upper Gondwana, outlierson the Madras coast. Memoir of Geological Survey of India,Palaeontologia Indica Ser. 2, 1(4): 1191-1224.
Foote, R.B. 1868. On the Distribution of Stone Implements inSouthern India. Quarterly Journal of the Geological Society,24(1-2): 484-495.
Foote, R.B., 1873. On the Geology of parts of Madras and NorthArcot district, Memoir Geological Survey India. Vol. X, Pt. 1, pp.1-732.
Jeyasingh, D.E.P. & Kumarasamy, D. 1994a. Araucarioxylon fromthe Sriperumbudur Formation, Upper Gondwana, Tamil Nadu,India.. Geophytology, 24(1-2): 43-48.
Jeyasingh, D.E.P. & Kumarasamy, D. 1994b. Occurrence ofPityospermum Nathorst in the Sriperumbudur Formation, TamilNadu. Current Science, 67(5): 305.
Jeyasingh, D.E.P. & Kumarasamy, D. 1995. An unusual pycnoxylicwood from a new Upper Gondwana locality in Tamil Nadu, India.Review Palaeobotany and Palynology, 85: 341-350.
Kumaraguru, P. 1991. Stratigraphic drilling in Palar basin Tamilnadu.Records of Geological Survey of India, Vol. 124, Pt. 5, pp. 139-143.
Kumaraguru, P. 1992. Stratigraphic drilling in the gondwanas ofTamilnadu. Records of Geological Survey of India, 125, Pt. 5,pp. 114 and 115.
Kumaraguru, P. & Rao, A.T. 1994. A Reappraisal of the Geology andTectonics of the Palar Basin Sediments, Tamilnadu. In NinthInternational Gondwana Symposium, Hyderabad, Jan. 1994, 2,Rotterdam, Geological Survey of India and Balkema,pp.821-831.
King, L.C. 1958. Basic palaeogeography of Gondwanaland duringthe late Palaeozoic and Mesozoic eras. Quarterly Journal ofGeological Society of London, 114: 47-70.
Lal, N.K., Siawal, A. & Kaul, A.K. 2009. Evolution of East Coast ofIndia- A Plate Tectonic Reconstruction. Journal of GeologicalSociety of India. 73: 249-260.
Maheshwari, H.K. 1986. Thinnfeldia indica Feistmantel and associatedfossils from Tiruchirapalli District, Tamil Nadu. The Palaeo-botanist, 35(1): 12-21.
Misra, P.K., Rajanikanth, A., Jauhri, A.K., Kishore, S. and Singh,S.K. 2004. Albian limestone building algae of Cauvery basin,south India. Current Science, 87: 1516-1518.
Murthy, N.G.K. and Sastry, V.V. 1961. Foraminifera from theSriperumbudur beds near Madras. Indian Minerals, 14: 214-215.
Oldham, R.D. 1893. A manual of the geology of India: stratigraphicaland structural geology 2nd Edn. Govt. of India Calcutta.
Pandya, N. and Sukh-Dev 1989. Fossil flora of Gollapalli formation.The Palaeobotanist, 38: 147-154.
Prasad, B. and Pundir, B.S. 1999. Biostratigraphy of the exposedGondwana and Cretaceous rocks of Krishna- Godavari basin,India. Palaeontological Society of India, 44: 91-117.
Rajanikanth, A. 1996a. Palaeobotany and stratigraphic implicationof Mesozoic Gondwana sediments of Pranhita – Godavari Graben.Proc. 9th Int. Gondwana Symposium, Hyderabad, GondwanaNine 1, 425-439.
Rajanikanth, A. 1996b. Diversification and evolution of EarlyCretaceous east coast flora of India. The Palaeobotanist, 45:121-131.
Rajanikanth, A. 2007. Status of Coastal Gondwana – A Floristicperspective. In: Jayappa, K.S. & A.C. Narayana (eds.) CoastalEnvironments I. K. International Publishing House Pvt. Ltd., NewDelhi, 20: 258-269.
Rajanikanth, A. Venkatachala, B.S. & Ashok Kumar 2000. Geologicalage of the Ptilophyllum flora in India – a critical reassessment.Geological Society of India Memoir, 46: 245-256.
Ramanujam, C.G.K. & Srisailam, K. 1974. Palynology of thecarbonaceous shale from a bore hole at Kattavakkam, nearConjeevaram, Tamil Nadu, India. Pollen Spores, 16: 67-102.
Ramanujam, C.G.K. & Varma, Y.N.R. 1977. Palynological evidencefor the age of Sriperumbudur beds encountered in a borehole atOrikkai near Conjeevaram, Tamil Nadu. Journal of GeologicalSociety of India, 18 (8): 429-435.
Ramanujam, C.G.K. & Varma, Y.N.R. 1981. Hilate spores from theupper Gondwana deposits of Palar basin, Tamil Nadu. ThePalaeobotanist, 28 & 29: 308-315.
Rangaraju, M.K., Agarwal, A. & Prabhakar, K.N. 1993. Tectono-stratigraphy, structural styles, evolutionary model and hydrocarbonprospects of Cauvery and Palar Basins, India. In: Biswas et al.(Eds.), Proc.2nd Seminar on Petroliferous Basins of India,v.1, Indian Petroleum Publishers, Dehradun-248001. India.pp. 371-388.
Sahni, B. 1928. Revision of Indian fossil plants Part I Coniferales (a)impressions and incrustations. Memoir of Geological Survey ofIndia. Palaeontologia Indica, 11: 1-49.
Sahni, B. 1931. Revision of Indian fossil plants. Part II Coniferales(b) petrifactions. Memoir of Geological Survey of IndiaPalaeontologia Indica N.S., 11: 54-124.
Sastry, V.V., Raju, A.T.R., Singh, R.N. & Venkatachala, B.S. 1974.Evolution of the Mesozoic sedimentary basins on the east- coastof India. APEA Journal, 29-41.
Sastry, V.V., Venkatachala, B.S. and Narayanan, V. 1981. The evolutionof east-coast of India. Palaeogeography, Palaeoclimatology,Palaeoecology, 36: 23-54.
Seward, A.C. & Sahni, B. 1920. Indian Gondwana plants - a revision.Memoir of Geological Survey of India, Palaeontologia Indica,7: 1-41.
Suryanarayana, A. 1953. Mesembrioxylon tirumangalense, a newspecies from Sriperumbudur group near Madras. Journal of IndianBotanical Society, 32(4): 159-164.
Suryanarayana, A. 1954. Fossil plants from the Jurassic rocks of theMadras Coast, India. The Palaeobotanist, 3: 87-90.
Suryanarayana, A. 1956. Dadoxylon rajmahalense Sahni from thecoastal Gondwanas of India. The Palaeobotanist, 4: 89-90.

28 PHYTOMORPHOLOGY January-June 2010
Sukh-Dev 1988. Floristic zones in the Mesozoic formations and theirrelative age. The Palaeobotanist, 36: 161-167.
Vaidyanadhan, R. & Ramakrishnan, M. 2008a. Geology of India,v. 1, Geological Society of India, 1-994.
Vaidyanadhan, R. & Ramakrishnan, M. 2008b. Geology of India,v. 2, Geological Society of India, pp. 1-994.
Vairavan, V. 1993. Tectonic history and hydrocarbon prospects ofPalar and Pennar Basins. In: Biswas et al. (Eds.), Proc. 2nd Seminaron Petroliferous Basins of India, pp. 389-396.
Varma, Y.N.R. & Ramanujam, C.G.K. 1984. Palynology of someupper Gondwana deposits of Palar basin, Tamil Nadu, India.Palaeontographica, B, 190: 37-86.
Venkatachala, B.S. 1977. Fossil floral assemblages in the east coastGondwana -a critical review. Journal of Geological Society ofIndia, 18: 378-397.
Venkatachala, B.S., Rajanikanth, A. & Maheswari, H.K. 1993. TheGondwana super group. Gondwana. Geological Magazine, SpecialVolume, Proceedings of Birbal Sahni Centenary Symposium,Nagpur; 80-92.
Yoshida, M., Funaki, M. & Vitannage, P.W. 1992. Proterozoic toMesozoic East Gondwana. The juxtaposition of India, Sri Lankaand Antarctic Tectonica, 11: 381-391.
Ziegler, A.M. Scotese, C.R. & Barrett, S.F. 1983. Mesozoic andCenozoic paleogeographic maps In: Brosche P. and SundermannJ. (eds.) Tidal Friction and the Earth’s Rotation II. Springer Berlin,pp. 240-252.

REKHA J. SALUNKE et al. – Foliar Architecture as Biomarker for Authentication of Carissa L. 29Phytomorphology 60 (1 & 2) 2010, 29-32
IntroductionUtility of Karonda as medicine has been well reportedin Indian system of medicine. ‘Karmadra’ has beenmentioned as one of the species in ‘Hruadya gana’ ofCharak
. M 0 > . M 0
> $ 2 ? A
0 . 0 M & 5 C
M 7 > . M 2 >
-. M 2 5 G $ 8
A 5 2 , & 0 & >
! ? . . > $ A 2
A Xç > ( @ $ ?
& 6 G . > ( ? 9
C & M / > ( ? -
5 ( M $ ? || $ ?
$ A 7 M : 7
> / 5 0 M : || g f
|| (Charak sutrastan adhaya - 4/10)
The ripe fruits have cooling, digestive properties andare used as cardio tonic, antiscorbutic, appetizer forsubsiding ‘pitta’, and is useful in anorexia, burningsensation, skin diseases particularly scabies and pruritus;unripe fruits are used in ‘vata’, and its preparation isgood for gum disorders; root bark is used for skin rash,
Foliar Architecture as Biomarker for Authentication of‘Karamarda’ (Carissa L.)
Rekha J. Salunke* and Vinaya S. GhateAgharkar Research Institute, Department of Botany, Pune-411004, India.
ABSTRACT
‘Karamarda’ has been mentioned as one of the species in ‘Hruadya gana’ of Charak, Ayurvedic system of medicine.Different species locally known by the names Karonda or karondi are used as ‘Karamarda’. Preliminary macro-characterization indicates potential of leaves as key characters for authentication of species under use. Four differentspecies viz. Carissa congesta Wight, Carissa carandas L, Carissa opaca Stapf ex Haines and Carissa spinarum L.have been characterized using leaf architecture methods. Species specific variations have been recorded with respectto shape, leaf tip and base in general and venation pattern in particular. Mid vein of all the species is strong. Secondaryveins in C. opaca are week and do not form the network as seen in other species. Arrangement of secondary veinsand loop formation towards margins in other species are unique and set apart the species. Authentication key basedon these variations has been proposed.
Key words: Leaf morphology, venation pattern, identification, Carissa species.
Author for Correspondence: Rekha J. Salunke, e-mail: [email protected], [email protected]
scabies and snake bite; root is used for wound healingin animals; leaves are used in cough and fever (Trivedi,et al., 1663; Kirtikar & Basu, 1984; Deshpande, et al,1989; Anonymous 1992; Anonymous 1993). Use ofdifferent plant parts of Karonda has also been recordedin ethnobotany (Jain, 1991).
Authentication of ‘Karmarda’ or ‘Karonda’ at specieslevel in Ayurveda as well as in traditional system is veryvague. Shloka indicating the species identity are notfound in Ayurvedic literature. Different names such asKrushnapak-phala, Kshira-phena, Samlapushpa, Sushen,Karamla, karmardak, Avighna, Mani-marda, Pak-krushna, Dindim, Krushna-phala indicating morpho-characters or properties are found in Dravyaguna Vidnyan(Deshpande, et al., 1989, Gogate, 2008). They however,are inadequate to identify Carissa at the species level.Species are also identified by the terms like ‘Jangalikaronda’ or ‘Kadu Karonda’. Jangali means wild. AllIndian Carissa species are wild in nature except for C.carandas. Similarly, Kadu i.e. bitter is a taste variantof fruit – Genotype, found in almost all the species.Recently 22 distinct types of fruits have been identifiedwith number of elite types based on fruit morphology(Ghate et al., 1999). Fruit based description such asspecies with big fruit is Karonda and with small fruitsis Karondi therefore, may mislead the species identity.
Under the project on comparative pharmacognosy
This content inHindi is not legiblebecause of fontcompatibility.
Kindly send afterscanning
we will composeand set right

30 PHYTOMORPHOLOGY January-June 2010
of medicinally important Carissa species preliminarymacro-characterization indicate potential of leaves askey characters for authentication of species under use.Detailed leaf architecture studies therefore, have beencarried out and the results have been described in presentpaper.
Materials and MethodsTen mature leaves of each species have been collectedrandomly from the different plants in order to covervariation in morphology and venation patterns. Theleaves have been fixed and processed for leaf architectureusing method described by Maqdoom and Prabhakar,(2003). The KOH treated leaves have been treated withH2O2, a clearing agent, to separate the debris and clearthe veins. These have been thoroughly washed withwater, stained with safranin and dehydrated with alcoholgrades and permanent slides have been prepared followingArnot (1959).
Leaf architecture has been described after (Hickey,1973; Dilcher, 1974; Prabhakar & Anna Mani, 1996).Vein islet numbers have been measured using proceduresgiven by Trease and Evans (1983). The comparative datahas been compiled in Tables 1, 2, 3 & 4.
Results and DiscussionCarissa L. (Karaunda/ Karavanda) is a genus of morethan 60 species distributed mostly in warm parts ofAfrica, Australia and Asia (Anonymous, 1895-1990).Hooker (1882) described five species of Carissa L. from
Table 1. Morphology of leaf.
Sr.no. Name of the species Lamina Form Base Apex1 C. congesta Texture-coriaceous-glabrous; Margin-entire; Venation- Elliptic Acute Acute
conspicuous on lower side2 C. carandas Texture-coriaceous-glabrous; Margin-entire; Venation- Oblong Rounded Rounded/
conspicuous on lower side Retuse3 C. spinarum Texture- coriaceous with wolly surface; Venation- Lanceolate Acute Mucronate
conspicuous on lower side; Margin-entire cuneate4 C. opaca Texture-coriaceous-glabrous; Venation- conspicuous Elliptic Shortly Mucronate
on lower side; Margin – entire attenuate
Table 2. Venation patterns and characters of primary and secondary veins
Median primary Secondary veinSr.No Name of the species Venation type
Course Size Number Position Angle of divergence Course1 C. congesta PNCB S WE 3 A AN AC2 C. carandas PNCB S ST 6 AS AM AC3 C. spinarum PNCB S WE 5 AS AM AC4 C. opaca PNCR S WE 3 OS AN UCPNCB-pinnate-camptodromous-brochidodromous; PNCR- pinnate-camptodromous-reticulodromous; S-straight; WE-week; ST-stout; A-alternate;AS-alternate to subopposite; OS-opposite to subopposite; AN-acute narrow; AM-acute moderate; AC-abruptly curved; UC-uniformly curved.
the Indian subcontinent with special note that the speciesin the area are very difficult to define and may probablybe reduced to one or two variable types. Haines (1919)described 5 Indian species with number of varieties ineach. There are however, number overlaps in the classifiedvariables. The variability and ambiguity has been alsopointed out in many regional and location specific floristicworks (Cooke, 1903-1908; Talbot, 1911; Santapau, 1960).Very recently 22 distinct types have been identified byARI, Pune with number of elite types on the basis ofexo-morphic fruit characters (Ghate et al., 1999).
Leaves of genus Carissa in general are opposite,petiolate, lamina symmetric, coriaceous with entiremargin. Species specific variables are found in apex,base, texture and venation pattern. Leaf shape is ellipticin C. congesta and C. opaca, oblong in C. carandas andlanceolate in C. spinarum. Base is acute in C. congesta,acute cuneate in C. spinarum, round in C. carandas, andshortly attenuate in C. opaca. Apex is mucronate in C.spinarum and C. opaca, acute in C. congesta and roundedor retuse in C. carandas. Texture is coriaceous withwoolly surface in C. spinarum and coriaceous-glabrousin C. congesta, C. carandas and C. opaca.
Venation is pinnate-camptodromous-brochi-dodromous in C. congesta, C. carandas, C. spinarumand pinnate-camptodromous-reticulodromous in C.opaca. Median primary vein is stout in C. carandas ascompared to C. congesta, C. spinarum and C. opaca.Position of secondary vein is alternate in C. congesta;alternate to sub opposite in C. carandas and C. spinarum;

REKHA J. SALUNKE et al. – Foliar Architecture as Biomarker for Authentication of Carissa L. 31
and opposite to sub opposite in C. opaca. Angle ofdivergence of secondary vein is acute moderate in C.carandas and C. spinarum and acute narrow in C.congesta and C. opaca. Behavior of loop forming branchesof secondary veins is acute to obtuse angled in C.carandas and C. spinarum; acute in C. congesta whereasthe loops are absent in C. opaca. Intersecondary veinsare simple in C. opaca were as they are composite inC. congesta, C. carandas and C. spinarum. In C. congesta,C. carandas and C. spinarum the tertiary veins arealternate and opposite and its relationship to mid-veinis approximately at right angle to oblique decreasingdownward. They are absent in C. opaca. The vein isletnumber is maximum in C. spinarum and lowest in C.carandas. The net work in C. opaca is week but showsvein islet number 35. This may be because of the spreadof secondary and tertiary veins in given area. The leafarchitecture in all species is unique particularly withreference to secondary veins and loop formations andset-apart the species. Authentication key based on thesecharacters has been formulated as follows.
Key1a. Leaf venation pinnate-camptodromous-
brochidodromous C. opaca1b. Leaf venation pinnate-camptodromous-
reticulodromous 22a. Position secondary vein alternate C. congesta2b. Position secondary vein alternate
to sub opposite 33a. Primary vein thick stout C. carandas3b. Primary vein thin week C. spinarum
AcknowledgementThe authors are thankful to the Director and In-chargeBotany Group ARI, Pune for providing facilities for thiswork. They are grateful to BSI Western Circle, Pune forallowing herbarium consultation for confirmation ofspecies.
Literature CitedAnonymous, 1895-1990. Index Kewensis: Plantarum
Phanerogamarum I: 441; suppls. I : 83; II : 44; III : 36;IV : 39; VII : 40; VIII : 41; IX : 53; XII : 29; XIII: 24;XV : 26; XVI : 57. KOELTZ SCIENTIFIC BOOKS, D-6240,Keinigstein West Germany.
Anonymous, 1992. (Rev.Ed). The Wealth of India: A dictionary ofIndian Raw materials and Industrial Products. CSIR Publications,New Delhi. Vol 3: 294-299.
Anonymous, 1993. Indian Medicinal Plants (Compendium of 500species) Arya Vaidya Shala, Kottakkal. Vol I: 350-353.
Arnot, H.J. 1959. Leaf clearing. Turtex News 37: 192-194.Cooke, 1903-1908. The Flora of the Presidency of Bombay
Reprinted in 1967 Botanical Survey of India, Calcutta. Vol. 2:186-188.
Deshpande, A.P., Javalaker, R.R. & Ranade, S. 1989. DravyagunVidyanan. Anamol Prakashan, Pune-411002, Part I & II: 488.
Dilcher, L. 1974. Approaches to the Identification of AngiospermLeaf Remains. Botanical Review, 40:1-157.
Ghate, V.S, Kulkarni, D.K. & Upadhey, A.S. 1999. Screening ofnatural diversity in karvanda (Carissa L.): commerciallypotential wild fruit in Maharashtra. Indian Jonral of Plant. Genet.Resources, 12(1): 10-15, 1999.
Gogate, V.M. Dravyagunvidnyan. First edition 2008. Vaidya MitraPrakashan, Pune 411030. 318.
Haines, 1919. Indian species of Carissa. Indian forester Vol XLV:357-388.
Table 3. Characters of secondary veins.
Sr. No Name of the speciesSecondary veins Intersecondary veins
No Distance(cm) Behavior of loop forming No Distance(cm) Typebranches
1 C. congesta 4 0.5-1.5 Acute 3 0.4-1.4 Composite2 C. carandas 6 0.5-1.0 Acute to Obtuse angle 6 0.7-0.9 Composite3 C. spinarum 5 0.9-1.2 Acute to Obtuse angle 11 0.2-1 Composite4 C. opaca 2 0.5-0.7 Loops absent 7 0.1-0.9 Simple
Table 4. Characters of the tertiary veins of leaf lamina and vein islet number.
Tertiary veins Relationshipof midvein Arrangement Vein Islet numbersSr.no Name of the species Angle of origin Course1 C. congesta RR FRSN ARODD ALOP 292 C. carandas RR FR ARODD ALOP 253 C. spinarum OO FR ARODD ALOP 454 C. opaca – – – – 35RR-right: right angle; OO-obtuse: obtuse angle; FRSN-forked: sinous; FR-forked; ARODD-approximately at right angle to oblique decreasingdownward; ALOP-alternate and opposite.

32 PHYTOMORPHOLOGY January-June 2010
Hickey, L.J. 1973. Classification of the architecture of dicotyledonsleaves. American Journal of Botani. 60: 17-33.
Hooker, J.D. 1882. The Flora of British India III: 630-632. London.Jain, S.K. 1991. Dictionary of Indian folk medicines and
Ethnobotany. Carissa: 45. Deep Publications, New Delhi.Kirtikar, K.R. & Basu, B.D. 1984. Indian Medicinal Plant. Vol II:
1546-1549.Maqdoom, M. & Prabhakar, M. 2003. Foliar architecture of
Asclepiadaceae in relation to taxonomy, Geophytology, 31(1& 2): 59-70.
Prabhakar, M. & Anna Mani, B. 1996. Foliar architecture inidentification of the flora. In: Frontiers in Plant Science, I.A.Khan (Ed.) pp. 1006-1026. Nawab Shah Alam Khan. C.P.G.S& Res. Hyderabad.
Santapau, H. 1960. Revised. The Flora of Khandala on WesternGhats of India. In: Records of Botanical Survey of India XVI(1): 129-130.
Talbot, W.A. Forest Flora of the Bombay Presidency and Sind.Poona. I Vol 1909, II vol. 1911 (1909 & 1911).
Trease, G.E. & Evans, W.C. 1983. Pharmacognosy. Alden Press,Oxford.
Trivedi, K., Garg, D., Agraval, J. & Gard, D. (Editors) 1663.Dhanvantari: Vanaushadhi Vishaheanke Vol 37(2): 164-166.

K. SRI RAMA MURTHY et al. – Effect of Cytokinins and Auxins on Flowering of Ceropegia sp. 33Phytomorphology 60 (1 & 3) 2010, 33-37
IntroductionCeropegia L. (Asclepiadaceae) is a genus of climbersand erect herbs that are distributed in tropical andsubtropical Asia, Africa, and Australia (Anonymous,1992). The flowers of all Ceropegia species aremorphologically unique, having a long corolla tube atthe distal end, which dissects out into five corolla lobesthat are connate at their apices, thus creating five smallwindows. These colorful flowers form an attractive cagefor insects. The genus Ceropegia is used as an ornamentalin the Europe and United States (Hodgkiss, 2004;Reynolds, 2006). Although none of the Indian Ceropegiaspecies has yet been domesticated for this purpose,plants of the Indian subcontinent are under threat becauseof either habitat degradation or over utilization of theirtubers. Tuberous roots of many Ceropegia species areedible (Anonymous, 1992). The root tubers are theofficinal parts known to contain an alkaloid called“Ceropegin” (Nadkarni, 1976). Bitterness of the tuberswas eliminated by boiling or roasting and then consumed(Mabberley, 1987). The Ceropegia spiralis root tubersknown to contain starch, sugars, gum, albuminoids, fats,crude fiber and valuable constituents in many traditionalIndian Ayurvedic drug preparations that are active againstmany diseases especially diarrhea, dysentery. The starchytubers are used as a nutritive tonic (Kirtikar & Basu,
Effect of Cytokinins and Auxins on In Vitro Flowering ofEndangered Ceropegia spiralis Wight and C. pusilla Wight & Arn1
K. Sri Rama Murthy* and R. KondamudiSchool of Conservation Biology and Plant Biotechnology, Department of Biotechnology, Montessori Mahila Kalasala,
Vijayawada-520010, Andhra Pradesh, India
ABSTRACT
Nodal and nodal TCLs explants of endangered Ceropegia spiralis and C. pusilla were cultured on Murashige & Skoogmedia (full, half and quarter strengths) supplied with sucrose, cytokinins and auxins alone and in combinations. Floweringwas achieved after 5-10 weeks of culture. In C. spiralis, the maximum flowering (84%) was observed on ½ MS mediumsupplemented with NAA 10.74 μM/l, 3% sucrose and in C. pusilla, the maximum flowering (80%) was recorded on½ MS containing IBA 0.0123 μM/l, with 3% sucrose. Although these flowers are smaller in size, the in vitro flowerswere morphologically comparable with that of in vivo derived flowers.
Key words: Asclepiadaceae, Ceropegia spiralis, Ceropegia pusilla, in vitro flowering
Abbreviations: BAP – 6-benzyle aminopurine, Kn – Kinetin, IBA – Indole-3-butyric acid, IAA – Indole-3-aceticacid, NAA – á-naphthalene acetic acid, 2,4-D – 2,4-diclorophenoxy acetic acid, MS – Medium-Mirashige & Skoog’s medium, PGRs – Plant growth regulators, TCLs – Thin cell layers.
*Author for correspondence: K. Sri Rama Murthy, e-mail: [email protected]
1935; Reddy et al. 2006; Chopra et al. 1956). In thisgenus C. bulbosa and C. candelabrum also have medicinalproperties (Jain & Defillips, 1991). Ceropegia speciesfrom Peninsular India have been designated as rare(Nayar & Sastry, 1987; Walter & Gillet, 1998) and theirexistence has become restricted to remote pockets in theHimalayas and the Western Ghats, two biodiversityhotspots. Unfortunately, the genus Ceropegia has nowbeen added to the list of Indian endangered plants (BSI,2002). For its conservation, large scale propagation,especially in vitro, has long been encouraged (Walter& Gillet, 1998). The reproductive physiology of theseendemic species must be studied in order to designcapable breeding and conservation strategies. So, farthere was no report on in vitro studies of these twoendangered Ceropegia species. In the presentinvestigation in vitro flowering was studied.
Materials and MethodsIn the midst of July and August (2007-08) the Ceropegiaspiralis Wight (Nimmati gadda) and C. pusilla Wight& Arn. (Churning stic) were collected from Akashagangaof Tirumala hills and the later was collected from Shevaroyhill ranges of Eastern Ghats, Tamilnadu, India. Thevoucher specimens were deposited in the herbarium,department of biotechnology, Montessori Mahila

34 PHYTOMORPHOLOGY January-June 2010
Kalasala, Vijayawada, Andhra Pradesh, India. The nodescontaining axillary buds were washed in the running tapwater followed by tween 20 (5% v/v for 4 min) afterwords cycloheximide and streptomycin (0.3% for 10min) surface disinfectant HgCl2 (0.1% w/v for 2 min,freshly prepared) later repeated washes were done indouble distilled water. The sterilized segments were thenwashed thoroughly with sterilized distilled water, cutinto appropriate sizes, and cultured on nutrient medium.The in vitro grown plant nodes and nodal thin cell layerexplants (TCLs) were excised to induce in vitro flowering.The employed TCLs were excised from the nodal andinternodal meristematic regions. About 6-7 TCLs (3from each side) were taken from either side of the node.MS medium (Murashige & Skoog, 1962) which wassolidified with agar 0.9% (w/v) HiMedia LaboratoriesPvt. Ltd. Mumbai and supplemented with different growthregulators (BA, Kinetin, NAA, IAA, IBA) at differentconcentrations either alone or in combinations. In thepresent investigation all the media were autoclaved at121°C and 15 lbs pressure for 20 min after adjustmentof the pH to 5.7±0.2 with 1N NaOH and 1N HCl. Allthe cultures were maintained at 24±2°C under 16hrphotoperiod with 3000 lux light intensity using fluorescentlights (Philips India Ltd.). The explants were inoculatedin 250 ml tissue culture bottles and 25×150 mm testtubes. Twenty cultures were raised for each treatmentand all experiments were repeated thrice. Here in this
investigation, the TCL explants also employed to inducethe flowering. Welldeveloped shoots were inoculated onthe MS media supplemented with different compositionsand combinations containing auxins alone for rootingand the plantlets were observed for flowering too. Hence,the plantlets were transferred to low light intensity asit favors the flowering. Here to induce flowering, chillingeffect was used as it favors the flowering.
ResultsFlower induction varied depending on the species andthe compositions of the media. The explants node / TCLsshowed their response within 7 days. These explantsproduce vigorously growing adventitious shoots after15 days of incubation. Each inflorescence consists of4 or 5 flower buds. The tissue culture of C. spiralis andC. pusilla not only favored rapid multiplication but alsocharacterized by seasonal in vitro flowering that was insynchrony with that of plants growing in the wild.
In this investigation, the media supplemented withauxins alone had the capacity to support numerousflower bud induction in C. spiralis, which did not bloom.Instead, they fall. When the ½ MS medium supplementedwith NAA 10.74 μM/l induced 10.14±0.37 buds (84 %)whereas, the medium augmented with IBA 49.2 μM/linduced 16.19±0.49 buds (50 %) but no anthesis wasobserved (Table 1). The fully developed andmorphologically similar flowers were observed on the
Table 1. Effect of different auxins on in vitro flowering in Ceropegia spiralis
Conc. of the PGR in μM/l Media strength Sucrose % Flowers mean ± SD Response %IBA IAA NAA49.2 ½ MS 6% 16.19±0.49a 504.92 ½ MS 3% 2.21±0.34d 339.84 ½ MS 3% 5.18 ±0.27c 5049.2 ½ MS 3% 3.10±0.19d 33
5.77 ½ MS 3% 1.14±0.17d 1611.54 ½ MS 3% 2.11±0.16d 3346.16 ½ MS 3% 1.16±0.23d 16
5.37 ½ MS 3% 1.07±0.19d 1610.74 ½ MS 3% 10.14±0.37b 8453.7 ½ MS 3% 2.04±0.36d 33
4.92 26.85 ½ MS 6% 1.16±0.44d 169.84 21.48 ½ MS 6% 1.13 ±0.26d 1614.76 16.11 ½ MS 6% 3.06±0.18d 3319.68 10.74 ½ MS 6% 4.98±0.18c 5024.6 5.37 ½ MS 6% 3.00±0.10d 33Data indicate mean ± standard deviation following by the same letter was not significantly different by the Tukey-Kramer multiple comparisonstest at 0.05 % probability. Twenty replicates were used per treatment experiments were repeated thrice.

K. SRI RAMA MURTHY et al. – Effect of Cytokinins and Auxins on Flowering of Ceropegia sp. 35
medium augmented with IAA 11.54 μM/l but they areless (2.11±0.16) in number with 33% within a periodof 35 days. The ½ MS medium supplemented with IBA19.68 μM/l + NAA 10.74 μM/l with 6 % sucrose wasfound optimum for both flowering and rooting (Table1),the synergistic effect was much adoptable than the PGRsalone.
In C. pusilla, the flowering was best reported onMS media supplemented with deferent combinationsand concentrations of cytokinins and auxins either aloneor in combination. The induction of the flower buds isas same as C. spiralis, but the buds failed to bloom. Whenthe flowers formed are less in number on a shoot theyare able to bloom. For the induction of more numberof flowers IBA 0.0123 μM/l (15.04±0.08) and IAA0.02885 μM/l (20.04±0.07) and IAA 11.54 μM/l(15.03±0.06) were employed (Table 2).
After the auxins it was the combination of cytokininsand auxins which played a significant role in the floweringof C. pusilla (Table 2). On these media flowers werebloomed and retained on the plant for the 3 days. Theflowering was observed throughout the year, irrespectiveof flowering season. The flower buds attained full bloomonly when the cultures were transferred to low light
intensity regime 2000 lux within a period of 4 days. Itwas observed that, under 16 / 8 hr light / dark cycle someof the buds failed to bloom and died off. This could havebeen due to fluctuations in the floral signal receptivityof shoot buds of those species. The plant growth regulatorsinduce morphologically normal in vitro flowers. Thesein vitro flowers are similar to that of in vivo flowers.It should be noted that neither floral fertility nor pollenviability were tested in this current series of experiments.The TCLs of C. spiralis and C. pusilla were culturedon MS medium supplemented with cytokinin and auxinswere able to induce more number of flowers. Whencompared with nodes, nodal TCLs induce more numberof flowers.
The whole investigation for rooting was carried outin the ½ MS medium with different percentages ofsucrose. It was noticed that, the plants in 6% sucroseare able to form more number of flowers, but they didnot opened. The shoots were able to form the aerial tuberswhich bulged a bit due to the storage of starch in theirinternodes. In conclusion, the in vitro flowering of C.spiralis showed no favorable response for 6% sucrose.But the plants growing in 3% sucrose media are steadywith their flowering. Whereas in C. pusilla, varied
Table 2. Influence of cytokinins and auxins in vitro flowering of Ceropegia pusilla
Conc. of PGR (μM/l) Media strength Sucrose % Flowers ResponseBAP Kn 2,4-D IBA IAA mean ± SE %
13.32 1.13 MS 3 3.01 ± 0.02d 4013.32 0.577 MS 3 2.04 ± 0.03d 3313.32 1.44 MS 3 7.00 ± 0.03c 5013.32 5.77 MS 3 2.06 ± 0.03d 3313.32 0.49 MS 3 2.06 ± 0.03d 3313.32 1.23 MS 3 4.97 ± 0.05d 4513.32 2.46 MS 3 4.97 ± 0.02d 4513.32 4.92 MS 3 10.09 ± 0.03b 5013.32 MS 3 15.02 ± 0.05a 80
18.16 MS 3 4.04 ± 0.02d 3045.4 MS 3 7.05 ± 0.02c 45
0.246 ¼ MS 6 5.13 ± 0.04c 500.123 ½ MS 6 5.15 ± 0.05c 500.246 ½ MS 6 5.01 ± 0.01c 500.0123 ½ MS 3 15.04 ± 0.08a 80
0.02885 ½ MS 3 20.04 ± 0.07a 600.1154 ½ MS 3 6.12 ± 0.05c 50
0.01442 ½ MS 3 0.99 ± 0.00d 2011.54 MS 3 15.03 ± 0.06a 55
Data indicate mean ± standard error (SE) of the mean following by the same letter was not significantly different by the Tukey-Kramer multiplecomparisons test at 0.05 % probability. Twenty replicates were used per treatment experiments were repeated thrice.

36 PHYTOMORPHOLOGY January-June 2010
response is being noticed in 3% sucrose augmentedmedia. The variations are due to the varyingconcentrations of the PGR and their combinations. Steadyresponse was noticed in 6% sucrose supplied mediairrespective of the strength of the media.
DiscussionIn both the cases well developed shoots were transferredto the MS medium supplemented with auxins i.e. IBA,IAA, NAA initiated in vitro flowering along with rooting,similar results were also reported in Ceropegia bulbosavar. bulbosa (Britto et al., 2003), Basilicum polystachyon(Amutha et al., 2008), Cichorium intybus (Nandagopal& Ranjithakumari 2006). Even though majority of theflowers were observed on the rooting media, only somecombinations were favorable for the enlargement of theflowers in C. spiralis in the present study.
Initially all the plants are growing in the mediumcontaining Kn 23.712 μM/l. Later the plants weresubcultured to high concentrations of auxins alone showsincreased the flowering frequency. It is due to the shockcaused by sudden dose change of PGR. Many factorslike carbohydrates, PGR, light and PH of the mediumare playing a major role in flowering (Heylon & Vendrig,
1998). Patil (1998) used spermine in the culture mediumto induce the flowering in C. jainii, C. bulbosa varbulbosa and C. bulbosa var lushii. Silver nitrate andcobalt chloride also favors in vitro flowering in Capsicumfrutescens (Sharma et al., 2008).
The increased amount of the sucrose in the mediumhad enhancing effect on the flowering in six Ceropegiaspecies like C. bulbosa var bulbosa, C. hirsuta, C. lawii,C. maccannii, C. oculata, and C. sahyadrica along withBAP (Nair et al., 2007). Different cytokinins have differenteffects on in vitro flowering has also been made byMeeks-Wagner et al. (1989), who showed that inNicotiana tabaccum, Kn promoted flower formation,whereas, zeatin promoted leafy shoot formation. Strongeffect of cytokinin, sucrose and photoperiod on in vitroflowering in embryogenic cultures was observed inKinnow mandarin (Singh et al., 2006). In Sinapis albathe effect of cytokinins and sucrose on the developmentof shoots, roots and in vitro flowering from the TCLexplants was studied (Havelange et al., 2000).
For the domestication of these aesthetic andornamental Ceropegia, we must focus on novel variations,like elimination of seasonality of flowering and mustadd some commercial values to these plants by makingthem transgenic. By producing these species routinely,floriculturists can add a new flower in the bouquet, withthe help of low cost tissue culture techniques.
AcknowledgementsThe authors are thankful to the Council of Scientific andIndustrial Research (CSIR), New Delhi, India forproviding the financial assistance.
Literature CitedAmutha, R., Jawahar, M. & Ravi Paul, S. 2008. Plant regeneration
and in vitro flowering from shoot tip of Basilicum polystachyon(L.) Moench - An important medicinal plant, Journal ofAgricultural Technology, 4: 117-123.
A nonymous 1992. Ceropegia Linn. (Asclepiadaceae), The Wealthof India, 3: CSIR, New Delhi, India. pp 448-449.
Botanical Survey of India 2002. Studies on Rare and EndangeredSpecies. http://www.envfor,nic.in/bsi/research.html
Britto, S.J., Natarajan, E. & Arockiasamy, D.I. 2003. In vitroflowering and shoot multiplication from nodal explants ofCeropegia bulbosa Roxb. var. bulbosa, Taiwania, 48: 106-111.
Chopra, R.N., Nayar, S.L. & Chopra, I.C. 1956. Glossary of IndianMedicinal Plants, CSIR, New Delhi.
Havelange, A., Lejeune, P. & Bernier, G. 2000. Sucrose/cytokininand auxins on flower neoformation in thin cell layers of Nicotianatabaccum L. Plant Cell Physiology, 29: 665-671.
Fig.1: In vitro flowering of Ceropegia spiralis and C. pusillaA. Ceropegia spiralis after 5 weeks of culture on ½ MS mediumsupplemented with 3% sucrose and IAA 11.54 μM.B. Ceropegia pusilla after 6 weeks of culture on ½ MS mediumsupplemented with 3% sucrose and IBA 0.0123 μM.
A B
KindlyconfirmFigs. A & BLabel

K. SRI RAMA MURTHY et al. – Effect of Cytokinins and Auxins on Flowering of Ceropegia sp. 37
Heylon, C. & Vendrig, J.C. 1998. Influence of different cytokininsand auxins on flower formation in thin cell layers of Nikotianatabaccum L., Plant Cell Physiology, 29: 665-671.
Hodgkiss, R.J. 2004. http://www.succulent-plant.com/ceropg.htmlJain, S.K. & Defillips, R.A. 1991. Asclepiadacae. In: Medicinal
Plants of India. 1: Algonac, India.Kirtikar, K.R. & Basu, B.D. 1935. Indian Medicinal Plants, 3: pp
(1638) M/s Bishen Singh Mahendrapal, New Delhi, India.Mabberely, D.J. 1987. The Plant Book. Cambridge University Press,
Cambridge Pp.114-115.Meeks-Wagner, D.R., Dennis, E.S., Tran Thanh Van, K. & Peacock,
W.J. 1989. Tobacco genes expressed during in vitro floralinitiation and their expressed during normal plant development,Plant Cell, 1: 25-35.
Murashige, T. & Skoog, F. 1962. A revised medium for rapid growthand bioassays with tobacco tissue cultures, Physiol. Plant. 15:473-497.
Nadkarni, K.M. 1976. Indian Materia Medica, 1: Pp. (303-304)Popular Prakasha, Bombay, India.
Nair, A.K., Naik, D.D. & Subhash, P.S. 2007. High frequency invitro flowering in six species of Ceropegia. Journal of PlantBiology, 50: 347-377.
Nandagopal, S. & Ranjithakumari, B.D. 2006. Adenine sulphateinduced high frequency shoot organogenesis in callus and in
vitro flowering of Cichorium intybus L. cv. focus - a potentmedicinal plant. Acta Agriculture Slovenica, 87: 415-425.
Nayar, M.P. & Sastry, A.R.K. 1987. Red Data Book of Indian Plants.1: BSI, Calcutta, India 1988.
Patil, V.M. 1998. Micropropagation studies in Ceropegia spp., Invitro Cell Dev. Biol-Plant, 34: 240-243.
Reddy, C.S., Reddy, K.N., Pattanaik, C. & Raju, V.S. 2006.Ethnobotanical observations on some endemic plants of EasternGhats, India. http//:www.siu.edu.beb/leaflets/ghats.htm
Reynolds, S. 2006. http://www.sagereynolds.com/cero/clist.comSharma, A., Kumar, V., Giridhar, P. & Ravishankar, G.A. 2008.
Induction of in vitro flowering in Capsicum frutescens underthe influence of silvernitrate and cobalt choride and pollentransformation. Electronic Journal of Biotechnology, 11: 84-89.
Singh, B., Sharma, S., Rani, G., Virk, G.S., Zaidi, A.A. & Nagpal,A. 2006. In vitro flowering in embryogenic cultures of Kinnowmandarin (Citrus nobilis Lour C. deliciosa Tenora). AfricanJournal of Biotechnology, 5: 1470-1474.
Walter, K.S. & Gillet, H.J. 1998. 1997 IUCN Red list of threatenedplants, compiled by the world conservation monitoring centre,IUCN, The World Conservation Union, Gland, Switzerland,Cambridge, pp 63-65.

A. BARUAH and J. SONOWAL – Comparative Vegetative Anatomy of Blue Vanda and Red Vanda 39Phytomorphology 60 (1 & 2) 2010, 39-42
IntroductionVanda coerulea Griff. Ex Lindl, is one of the mostcharming and sought-after orchids, popularly known as‘Blue Vanda’. In its native habitat, it is found growing inmixed pine forests on hills between 1300-2000 m altitudes,usually growing on species of Quercus. The plant is 30-60 cm high, having two opposite rows of symmetricalleaves which are strap-like. The erect scape bears 10-20large flowers, which are 7-10 cm in diameter. Floweringoccurs from October to November. In Northeast India, itsoccurrence is rarely recorded in sub-tropical zones in thestates like Assam, Arunachal Pradesh, Manipur,Meghalaya and Nagaland. This species is rather sun-loving. However, humid atmosphere in summer and drywinter helps healthy growth of the species. Vanda coeruleais extensively cultivated and several hybrids of commercehave been produced from it. The species is considered‘endangered’.
Renanthera imschootiana Rolfe, commonly knownas ‘Red Vanda’ is also an endangered species and growson large broad-leaved tree trunks preferably in sunnyareas with moderate rainfall. In Northeast India itsoccurrence is recorded in Manipur, Mizoram andNagaland (Hedge, 1984). Both the orchid species areenlisted in the ‘Red Data Book of Indian Plants’ (Nayar& Sastry, 1987, 1990).
Except a preliminary study on leaf anatomy(transection) of Renanthera imschootiana (Rao, 1998),no other works with regards to the anatomy of these two
Comparative Vegetative Anatomy of Blue Vanda (Vanda coeruleaGriff. Ex Lindl.) and Red Vanda (Renanthera imschootiana Rolfe.)
A. Baruah* and J. SonowalPhytodiversity Research Cell, Department of Botany, Darrang College, Tezpur-784001,India
ABSTRACT
A comparative anatomical study, on the leaf and roots of two endangered orchids namely – Blue Vanda (Vanda coeruleaGriff. Ex Lindl.) and Red Vanda (Renanthera imschootiana Rolfe.), was done. In both the species, the stomata areanomocytic, vascular bundles are arranged in a single series in leaves, while vascular strands are radial in roots. Thevegetative anatomical characters, such as distribution and colour of sclereids as well as number of vascular bundlesin leaves, presence or absence of striated velamen cells and the number of cortical cell layers in roots, are found helpfulin distinguishing the studied orchid taxa.
Key words: Vanda coerulea, Renanthera imschootiana, leaf, root, anomocytic stomata, sclereids, cortex, vascularbundles
Author for Correspondence: A. Baruah, e-mail: [email protected]
orchid species have so far been done. Anatomicalcharacters can be used to distinguish species in vegetativestage (Baruah 1998, Baruah & Nath 1997, Kaushik 1983,Khasim & Mahan Ram 1989, Stern & Judd 1999). Ourwork was restricted to the comparative vegetative anatomyof leaves and roots of Vanda coerulea and Renantheraimschootiana.
Materials and MethodsLeaves and roots were obtained from live plants,maintained in a home garden near Botanical Survey ofIndia (BSI), Shillong. The identity of the ‘Blue’ and‘Red’ Vanda were confirmed by Dr. R. Gogoi of BSI,Shillong. The transections of the leaf and root were madewith the help of a sharp razor blade by free hand sections.Epidermal peels from the fresh leaf fragments of size 3x5mm were obtained, after 30 min treatment at 400C using5% Potassium hydroxide as macerating agent for themesophyll. The peels were stained with aqueous safranin,mounted in 50% glycerine and sealed withDibutylphthalate xylol. Camera lucida sketches weredrawn from the prepared slides. The quantitative datawere calculated from the average of 10 readings of eachcharacter. The stomatal indices were also calculated(Baruah & Nath, 1997).
Observations and DiscussionBlue Vanda (Vanda coerulea)Leaf epidermis in surface view: Epidermis is single layeredand consists of straight to slightly curved, thin walled and

40 PHYTOMORPHOLOGY January-June 2010
tetragonal to polygonal cells (Fig. 1A & B). The numberof epidermal cells per mm2 on the upper surface is 5350,while on the lower epidermis, it is 5921. The size ofepidermal cells on upper surface is 40.79 x 14.48μm andon lower surface it is 27.05 x 19.94μm. Stomata are onlypresent in the lower epidermis of the leaf (hypostomatic).Stomata are anomocytic with 7-10 subsidiary cells(Fig. 1B). The number of stomata per mm2 is 192 andtheir size is 44.14 x 31.32 μm. The stomatal index is 3.14.Leaf transection: In outline leaf is dorsiventral and flat,V-shaped adaxially in the mid-vein region, taperingtowards the edges and slightly turning downwards (Fig.1C). Epidermis is uniseriate and consists of rectangular,thin walled cells. Cuticle present in both surfaces andthin. Epidermis is followed by fusiform striated sclereidswith a distinct lumen and wavy secondary wall which areintermingled with mesophyll cells. These sclereides arecolourless and scattered in the peripheral regions of theupper and lower mesophyll (Fig. 1E).
Mesophyll is undifferentiated and is 10-12 layered.Mesophyll cells of one or two upper and lower-mostlayers are comparatively smaller than those of middlelayers. Mesophyll cells are trigonal to polygonal, devoidof starch grains and rich in chloroplasts (Fig. 1E).
Vascular bundles, which are arranged in a singleseries in the mesophyll are comparatively smaller in size(106.80 x 95.40 μm) towards the margins than the midvein(225 x 132.43 μm). Vascular bundles 30-34 in number,collateral and their shapes are ranged from oval to globose(Fig. 1C). In each vascular bundle, xylem is lying towardsthe adaxial surface and the phloem towards the abaxialsurface.Root: It is almost circular in outline with smooth surface(Fig. 1D). The velamen consists of 4-5 layers of elongated,rectangular to polygonal cells with thin cutinized walls.Some velamen cells with striations are found scattered(Fig. 1F). The exodermis consists of a single layer ofpolygonal to rectangular, elongated cells with scatteredpassage cells (Fig. 1F). A few exodermal, passage andeven ground cortical cells also possess striations. Thecortex is of 27-32 cells thick and comprising rectangularto polygonal, thin walled, parenchymatous cells whoseouter one or two layers are comparatively smaller andslightly lignified but the innermost layers are unlignified.Cortical cells are packed with starch grains. Intercellularspaces are scattered. Vascular strands 15 in number andradial. The size of xylem vascular strand is 69.78 x 54.11μm. The xylem is exarch (Fig. 1D). Pith distinct,
comprising pentagonal to polygonal parenchymatouscells with small intercellular spaces.Red Vanda (Renanthera imschootiana):Leaf epidermis in surface view: Epidermis is singlelayered and consists of straight to slightly curved, thinwalled and tetragonal to polygonal cells (Fig. 1G & H).The number of epidermal cell per mm2 on the uppersurface is 3660, while on the lower epidermis, it is 4442.The size of epidermal cell on upper surface is 44.14 x21.26 μm and on lower surface it is 34.18 x 14.24 μm.Stomata are only present in lower epidermis of the leaf(hypostomatic). Stomata are anomocytic with 7-8subsidiary cells (Fig. 1H). The number of stomata permm2 is 189 and their size is 37.02 x 32.75 μm. Thestomatal index is 4.09.Leaf transection: In outline the leaf is dorsiventral andflat, V-shaped adaxially in the mid-vein region, taperingtowards the edges and slightly turning downwards(Fig. 1I).
Epidermis is uniseriate and consists of rectangular,thin walled cells. Cuticle present in both surfaces andthin. Epidermis is followed by fusiform striated sclereidswith a distinct lumen and wavy secondary wall which arepresent parallel to the leaf surface in sub-epidermal region.These sclereids are dull-red in colour (Fig. 1K).
Mesophyll is undifferentiated and is 13-14 layered.Mesophyll cells of one or two upper and lower-mostlayers are comparatively smaller than those of middlelayers. Mesophyll cells are tetragonal to polygonal, devoidof starch grains and rich in chloroplasts (Fig. 1K).
Vascular bundles which are arranged in a single seriesin the mesophyll are comparatively smaller in size (111.07x 79.74 μm) towards the margins than the midvein (160.91x 125.31 μm). Vascular bundles 12 in number, collateraland their shapes are ranged from oval to globose (Fig.1I). In each vascular bundle, xylem is lying towards theadaxial surface and the phloem towards the abaxialsurface.Root: It is almost circular in outline with smooth surface(Fig. 1J). The velamen consists of 5-6 layers of rectangularto polygonal cells with thin cutinized walls (Fig. 1L), Theexodermis consists of a single layer of rectangular topolygonal, elongated cells with some passage cellsintermingled with them (Fig. 1L). A few exodermal,passage and even ground cortical cells possess striations.The cortex is of 19-20 cells thick and comprisingrectangular to polygonal, thin walled, parenchymatous
A. BARUAH and J. SONOWAL – Comparative Vegetative Anatomy of Blue Vanda and Red Vanda 41
Fig. 1: A-F. Anatomical structures of Vanda coerulea. G-L. Anatomical structures of Renanthera imschootiana (cl, cuticle; ep, epidermalcell; sld, sclereid; vm, velamen; ed, exodermis; pc, passage cell). A & G. Upper epidermis of leaf. B & H. Lower epidermis of leaf. C &I. Outline drawing of transaction (T.S.) of leaf. D & J. Outline drawing of transaction (T.S.) of root. E & K. T.S. through leaf (upperside) showing cuticle, epidermis and sclereids. F & L. T.S. through root showing velamen, exodermis and passage cell.

42 PHYTOMORPHOLOGY January-June 2010
Table 1. Comparative anatomical character distinctions of BlueVanda and Red Vanda orchids
Characters Blue Vanda Red VandaLeaf
Root
Epidermis: Epidermal cellmeasures 40.79 x 14.48μm in upper surface while27.05 X 19.94 μm in lowersurface. Number ofepidermal cells per mm2
in upper and lowersurfaces 5350 and 5921,respectively.Transection of Leaf:Epidermis is followed byfusiform striated sclereidswith a distinct lumen andwavy secondary wallwhich are intermingledwith mesophyll cells.Striated sclereids arescattered in peripheralregions of the upper andlower mesophyll. Thesesclereids are colourless.Mesophyll is 10-12layered. Mesophyll cellsare trigonal to polygonal.
Number of vascularbundles 30-34.
Transection: Somevelamen cells withstriations are present andscattered.Cortex 27-32 layered.Xylem vascular strandsmeasure 69.78x54.11 μm.
Epidermis: Epidermalcell measures 44.14 x21.26 μm in uppersurface while 34.18 x14.24 μm in lowersurface. Number ofepidermal cells per mm2
in upper and lowersurfaces 3660 and 4442,respectively.Transection: Epidermisis followed by fusiformstriated sclereids with adistinct lumen and wavysecondary wall which arepresent parallel to thesub-epidermal region ofupper and lower leafsurfaces. Sclereids aredull-red in colour.
Mesophyll is 13-14layered. Mesophyll cellsare tetragonal topolygonal.
Number of vascularbundles 12.
Transection: Striationsabsent in velamen cells.Cortex 19-20 layered.Xylem vascular strandsmeasure 49.84x37.02μm.
ReferencesBaruah, A. 1998. Vegetative anatomy of the endemic orchid Vanilla
pilifera Holt. Phytomorph, 48: 101-105.Baruah, A. & Nath. S.C. 1997. Studies on the foliar epidermal
characters of some members of the family Euphorbiaceae.Advances in Plant Sciences, 10: 115-121.
Hedge, S.N. 1984. Orchids of Arunachal Pradesh. Published by ForestDepartment, Arunachal Pradesh, Itanagar, India, 82pp.
Kaushik, P. 1983. Ecological and Anatomical marvels of the Himalayanorchids. Today & Tomorrow’s Printers & Publishers, New Delhi,India.
Khasim, S.M. & Mahan Ram, P.R. 1989. Anatomy of four speciesDendrobium. Journal of Swamy Botanical Club, 6: 99-104.
Nayar, M.P. & Sastry, A.R.K. 1987. Red Data Book of Indian Plants,(Vol. I) BSI Publication, Howrah, India, 278pp.
Nayar, M.P. & Sastry, A.R.K. 1990. Red Data Book of Indian Plants,(Vol.III) BSI Publication, Howrah, India, 202pp.
Rao, T.A. 1998. Conservation of wild orchids of Kodagu in the WesternGhats. Published by the Centre for Technology Development,The Agricultural Technology & Services Pvt. Ltd., KarnatakaAssociation for the Advancement of Science, Central CollegeBangalore & World Wide Fund for Nature, New Delhi, India,65pp.
Stern, W.L. & Judd, W.S. 1999. Comparative vegetative anatomy andsystematics of Vanilla (Orchidaceae). Botanical Journal of LinneanSociety, 131: 353-382.
cells whose outer one or two layers are comparativelysmaller and slightly lignified but the innermost layers areunlignified. Cortical cells are packed with starch grains.Intercellular spaces are scattered. Vascular strands 15 innumber and radial. The size of xylem vascular strand is49.84 x 37.02 μm. The xylem is exarch (Fig. 1j). Pithdistinct, comprising pentagonal to polygonalparenchymatous cells with small intercellular spaces.
While comparing the evaluated characters of leafand root, we found some significant anatomicalcharacter distinctions between the two studied orchidspecies (Table 1).

AMAN VERMA et al. – Effects of in vitro Nitric Oxide in Groundnut 43Phytomorphology 60 (1 & 2) 2010, 43-48
Effects of In Vitro Nitric Oxide on Photosynthesis, AntioxidantCapacity and Proline Accumulation in Groundnut Cultivar
RG-141 Subjected to Mercury StressAman Verma*, Varsha Sharma* and C.P. Malik**
*School of Life Sciences, Jaipur National University, Jaipur, India**Department of Biochemistry, Kurukshetra University, Kurukshetra
ABSTRACT
Nitric oxide is a bioactive molecule involved in mediation of various biotic and abiotic induced physiological responsesin plants. In the present study, using SNP (Sodium nitroprusside) as Nitric Oxide (NO) donor, phytotoxicity of mercuryon growing groundnut (Arachis hypogaea L.) cultivar. RG-141 was studied. Seeds were soaked for 24 h in varyingMercuric Chloride (HgCl2) concentrations (0.1, 0.5, 1.0 and 2.0 mM). Control set was devoid of any plant growth regulator(PGR). Embryonal axis with cotyledon was inoculated into two types of media, one containing HgCl2 concentrationsalone and other containing SNP as a NO donor along with it to see the effect of NO in counter acting the heavy metalstress. The extent of damage caused by HgCl2 and influence of NO in combination were observed through the variableresponses of selective bioindicators such as chlorophyll content, level of proline, peroxidase activity, etc.
Key words: Arachis hypogaea L., Sodium nitroprusside, Nitric Oxide, Embryonal axis with cotyledon, HgCl2,Heavy metal stress, antioxidant enzymes
* Author for Correspondence: C.P. Malik, e-mail: [email protected]
IntroductionMetal toxicities have gained a worldwide attention aslarge amounts are released into the environment limitingcrop productivity. They are known to cause irreversibledamage to a number of vital metabolic constituents andimportant biomolecules. They also cause irreversibleinjury to the plant cell wall and cell membrane. The toxicthreshold level of the metal in the tissue is defined bythe stress point’ for metal toxicity and beyond this level,the physiological state of the plant cell will be irreversiblydamaged (Van and Clijsters, 1990). During metal toxicity/ heavy metal stress plants may confer metal stress byrapidly accumulating certain compatible stress protect-ant compounds such as proline, glycine betain etc. (Bray,1993; Maggio et al., 2002).
Mercury (Hg) is a highly toxic, nonessential,persistent, immutable and nonbiodegradable metal thatundergoes many changes during transfer through differenttrophic levels of the food chain. Hg occurs naturally inthe environment in the various forms such as metallicmercury, inorganic mercury and organic mercury. Metallicmercury is the main form of the mercury released intothe air by natural processes. It can be absorbed fromthe soil and atmosphere, accumulate in the plant organsand show their phytotoxic effects.
Nitric oxide is a bioactive, simplest, lipophylic,lightest and only gaseous signal transmitting moleculeinvolved in mediation of various physiological andbiochemical functions in plants (Crawford & Guo, 2005;Del Rio et al., 2004; Lamattina et al., 2003; Neill et al.,2003; Wendehanne et al., 2004) induced during bioticand abiotic stress (Belgini & Lamattina, 2001). Thisenigmatic but unique diffusible multifunctional plantsignal molecule plays pivotal role in hormone modulation,programmed cell death and defence responses. Thecytoprotective role of NO are thought to be due to it’sreactivity with ROS (Del Rio et al., 2004; Lamattinaet al., 2003; Neill et al., 2003; Wendehanne et al., 2004).Exogenous application of NO confers resistance to salt(Uchida et al., 2002), heavy metals (Hsu & Kao, 2004),chilling (Neill et al., 2003) and ultra violet B- radiation(Shi et al., 2005) stresses. Effects of NO on plant growthare concentration dependent (Anderson & Mansfield,1979), lower concentrations (0-20 ppm) had proved tobe protective, whereas higher concentrations (40-80ppm) had proved to be toxic (Heeften et al., 1996;Leshem & Haramaty, 1996). NO acts as a chain breakerwherever damage is caused by ROS and limit damage(Lipton et al., 1993).

44 PHYTOMORPHOLOGY January–June 2010
This study was conducted to evaluate the in vitrorole of NO donor (SNP) in ameliorating the Hg toxicityin groundnut cultivar. The present study was also aimedto explore further the relation between NO and chlorophyllcontent, proline level and antioxidant enzyme systemunder normal and stressed conditions.
Materials and MethodsSeeds of groundnut cultivar RG-141 were obtained fromAgricultural Research station (ARS), Durgapura, Jaipur,India. Seeds were washed with running tap water andthen surface sterilized with 0.1% (w/v) aqueous mercuricchloride (HgCl2) solution for 7 min. Seeds weregerminated aseptically in 90×15 mm Petri dishes layeredwith wet/moistened 2-3 fold of blotting paper with: (1)Sterile water (control) and (2) Four differentconcentrations of HgCl2 i.e. 0.1, 0.5, 1.0, 2.0 mM for24 h. Embryonal axis with one cotyledon (Fig. 1A) wereseparated aseptically from the seeds used as an explant.Explants were then transferred aseptically to followingmedia: (1) MS basal medium (Murashige & Skoog,1962) with 3% (w/v) sucrose and 0.8% (w/v) agar(control; MS1); (2) MS1 + 4 mgL-1 6-Benzyl Adenine(BA) + 2 mgL-1 NAA (MS2); (3) MS2 + 0.1 mM SNP(MS3) (Fig. 1B). The pH of all media was adjusted to5.8 before autoclaving. Cultures were incubated at 25±2ºCunder white fluorescent light 32 μMm-2s-1 with 16 hoursphotoperiod. Rooting was induced in the developedshoots by using Indole Butyric Acid (IBA) as a PGR.Well rooted plantlets were hardened and transferred toearthen pots (Figs. 1C & D). Every after 20 days ofincubation, leaves were sampled to determine the relevantphysiological characteristics of treated and untreatedplants.
Chlorophyll quantificationThe pigments were extracted in 80% chilled acetone.The amount of total chlorophyll, chl.a and chl.b wasestimated spectrophotometrically according to Coombset al. (1985) against the solvent (80%) blank. Chlorophyllcontents were expressed as μM g-1 FW (fresh weight).
Measurement of proline concentrationProline was determined the method of Bates et al. (1973).Leaves were homogenized with 3% sulphosalicylic acidand the homogenate was centrifuged at 3000 g for 10min. After this acetic acid and acid ninhydrin wereadded, the supernatant was boiled for 1h and thenabsorbance of the supernatant at 520 nm was determined.Proline concentration was calculated with a standardcurve and expressed as mM g-1 FW.
Analysis of PeroxidasePeroxidase enzyme was estimated by using the methodof chance and Machlt (1955). Leaves were homogenizedin 0.1 mM phosphate buffer pH 6.8. Five ml of the assaymixture comprised 125 μM of 0.1 mM phosphate buffer(pH 6.8), 50 μM of pyrogallol, 50 μM of H2O2 and1 ml of the 2 times diluted enzyme extract. This wasincubated for 5 minutes at 25ºC after which the reactionwas stopped by adding 0.5 ml of 5% H2SO4. The amountof purpurogallin formed was determined by taking theabsorbance at 420 nm.
Analysis of PolyphenoloxidasePolyphenoloxidase was estimated using the method ofChance and Machlt (1955). Fresh leaves werehomogenized in 0.1 mM of phosphate buffer pH 6.8.Five ml of the assay mixture comprised 125 μM ofphosphate buffer (pH 6.8), 50 μM of pyrogallol and 1ml of the 2 times diluted enzyme extract. This wasincubated for 5 minutes at 25ºC after which the reactionwas stopped by adding 0.5 ml of 5% H2SO4. The amountof purpurogallin formed was determined by taking theabsorbence at 420 nm.
Statistical analysisIn all the experiments total seven levels of treatmentswas formed i.e. four levels of HgCl2 and two levels ofmedia along with one control. Their performance for theaforesaid parameters was judged by applying analysisof variance for completely randomized design using f-test. Since the treatments were formed with differentfactors, their marginal effects of HgCl2, media alongwith their interaction and control verses rest were tested.Correspondingly for the significant effects, CDs wereestimated at 5% and 1% level of significance.
ResultsNitric oxide prevented groundnut leaves from oxidativedamage caused by heavy metal stress. Mercury treatedleaves lost chlorophyll in gradually dose- and time-dependent manners, whereas NO could apparentlycounteract the decay of chlorophyll in the leaves. Asdepicted in Table 5, chlorophyll b in groundnut leavesdecreased markedly with increasing concentrations ofheavy metal. Addition of NO donor (MS3) restoredchl. b content when compared with MS2 even at lowconcentration. Total chlorophyll content decreased withincreasing concentration of heavy metal in MS2 medium.With NO donor chlorophyll content was restoredsubstantially. Clearly, HM treated leaves lost chlorophyll

AMAN VERMA et al. – Effects of in vitro Nitric Oxide in Groundnut 45
in gradually dose-dependent manner, whereas NO couldcounteract the decay of chlorophyll caused by heavymetal. Comparatively chl.a was less affected (Table 4).
The data given in Table 1 show that HM stressinduced accumulation of proline in peanut leaves. Undernormal growth conditions (MS1) proline as an osmoticmediator and antioxidative substrate showed nearly steadystates (Table 1). The accumulation of proline in HM-treated peanut leaves was enhanced with the dose-dependent manner. The addition of NO further inducedthe accumulation of proline in HM-treated leaves.Thus, 0.1 and 0.5 mmol heavy metal had 1.5 and2.48 mM ml-1 g-1 FW. This is in comparison to1.16 mM ml-1 g -1 FW. SNP further accentuated the levelof free proline.
Further studies indicated that the activity ofperoxidase increased in the MS2 growth conditions. 0.1mM of HM enhanced the activity and with the additionof NO (MS3), the activity significantly increased at this
level of stress. With the HM stress level enhanced theactivity of the enzyme declined and addition of NO didnot produce any effect. At 2 mM concentration of HM,the peroxidase activity decreased over the control. BrieflyNO had no apparent effect on Peroxides activity (Table2). Further, the changes of this H2O2 scavenging enzymebehaved differently under HM stress. Polyphenol oxidaseactivity enhanced with HM stress, but as the concentrationwas enhanced the activity of this enzyme decreasedirrespective of the two media used (MS2 and MS3). NOincreased the activity of polyphenol oxidase at lowconcentrations, but at high concentrations of HM it wasunaffected (Table 3).
DiscussionMercury causes many morphological, physiological andbiochemical changes in growing plants. Chlorophyllcontent acts as a primary bioindicator of Hg toxicity.In the present study with increase in the Hg concentration,decrease in the chlorophyll a and b was observed (Jiang
Table 1. Effect of NO on the proline content (mM g-1 FW) in leaves under different concentrations of mercury after 24 h of incubation
HgCl2 (μM) MS2 MS3 Marginal Mean0.1 1.548 1.600 1.5740.5 2.144 2.322 2.2331.0 1.987 2.268 2.1282.0 1.581 2.232 1.907Marginal Mean 1.815 2.106 1.960MS1 Mean 1.118
For comparison of SE (d) CD 5% CD 1%Media means 0.001 0.002 0.002HgCl2 means 0.001 0.002 0.003Int. means 0.002 0.003 0.005Control v/s Treatments 0.001 0.003 0.004
Table 2. Effect of NO on the peroxidase activity (A420) in leaves after treatment with different concentrations of mercury after 24h ofincubation
HgCl2 (μM) MS2 MS3 Marginal Mean0.1 0.349 0.322 0.3360.5 0.285 0.274 0.2801.0 0.241 0.232 0.2372.0 0.267 0.234 0.251Marginal Mean 0.286 0.266 0.276MS1 Mean = 0.291
For comparison of SE (d) CD 5% CD 1%Media means 0.011 0.022 0.031HgCl2 means 0.015 0.032 0.043Int.means 0.021 0.045 0.061Control v/s Treatments 0.016 0.033 0.046
Table 3. Effect of NO on the polyphenol oxidase activity (A420) in leaves after treatment with different concentrations of mercury after24h of incubation
HgCl2 (μM) MS2 MS3 Marginal Mean0.1 0.362 0.389 0.3760.5 0.288 0.349 0.3191.0 0.214 0.231 0.2232.0 0.227 0.244 0.236Marginal Mean 0.273 0.303 0.288MS1 Mean = 0.282
For comparision of SE(d) CD 5% CD 1%Media means 0.010 0.021 0.028HgCl2 means 0.014 0.029 0.040Int. means 0.020 0.041 0.056Control v/s Treatments 0.015 0.031 0.042

46 PHYTOMORPHOLOGY January–June 2010
Table 4. Effect of NO on the chlorophyll-a content (μM g-1 FW) in leaves under different levels of mercury stress after 24 h of incubation
HgCl2 (μM) MS2 MS3 Marginal Mean0.1 53.450 68.800 61.1250.5 51.350 68.700 60.0251.0 47.550 58.400 52.9752.0 32.250 55.150 43.700Marginal Mean 46.150 62.763 54.456MS1 Mean = 77.95
For comparison of SE(d) CD5 % CD 1%Media means 0.086 0.180 0.247HgCl2 means 0.121 0.255 0.349Interaction means 0.171 0.360 0.493Control v/s Treatments 0.129 0.270 0.370
Table 5. Effect of NO on the chlorophyll-b content (μM ml-1g-1FW) leaves under different levels of mercury stress after 24 h ofincubation
HgCl2 (μM) MS2 MS3 Marginal Mean0.1 60.250 88.550 74.4000.5 53.700 70.300 62.0001.0 50.250 73.950 62.1002.0 40.800 65.550 53.175Marginal Mean 51.250 74.588 62.919MS1 Mean = 85.65
For comparison of SE (d) CD 5% CD 1%Media means 0.071 0.149 0.204HgCl2 means 0.100 0.210 0.288Interaction means 0.142 0.297 0.407Control v/s Treatments 0.106 0.223 0.306
Fig. 1: Response of groundnut cultivar RG-141 under the influence of HM stress and SNP as NO donor. A – Embryonal axis as an explant,B–Response of MS1, 3 and 2 respectively, C–Rooted plantlet in a thermocol pot during hardening, D–Hardened plants in the earthenpot.
A B
DC
MS1MS3
MS2

AMAN VERMA et al. – Effects of in vitro Nitric Oxide in Groundnut 47
et al., 1994). Sodium nitroprusside (SNP) is reportedas an important NO donor (Malik et al., 2007; Vermaet al., 2009). Low concentrations of SNP generated asteady-state NO concentration.
Our data for chlorophyll showed that exogenous NOcould significantly mitigate the oxidative stress causedby HM stress in peanut leaves. Significant decrease inthe chlorophyll a and b in the presence of HM can beeither by inhibiting protochlorophyllide reductase or byinhibiting the water spliting enzymes located at theoxidising site of photosystem II (Van Assche & Ciljsters,1990). Application of NO donor in in vitro conditionswas proved to be evident in alleviating the HM stressinduced damage. Among the overproduction of ROSunder HM stress has been reported by various workers(Jiang et al., 1999). It would be interesting to quantifyMDA, the product of lipid peroxidation. One pointwhich deserves special mention is the role of NO whichpresumably eliminates ROS (O2 and H2). The role ofperoxidase in scavenging the oxygen ions is also deduced.
Peroxidases are associated with physiologicalprocesses including disease and pathogen responses,lignin and suberin biosynthesis and more so with thebiotic and abiotic stresses (Fry, 1986).They constitutea large family having substrate specificity and functionin different peroxidation reactions in the cell. They areconsidered as antioxidant enzymes protecting cells fromthe destructive influence of derived oxygen species(Chen & Schopfer, 1999). In our NO is shown to affectperoxidase activities with substantial knock-on effectson reactive oxygen species (Clark et al., 2000).
The present results on the effect of SNP on peroxidaseactivities were HM concentration dependent. At lowconcentrations it enhanced the peroxidase activity buthigher concentrations decreased the activity. In view ofthe observations of some workers it would be interestingto estimate catalase activity as well. These workers havedemonstrated decreased catalase activity with NO.
ReferencesAnderson, L. & Mansfield, T.A. 1979. The effects of nitric oxide
pollution on the growth of tomato. Environmental Pollution, 20:113-121.
Bates, L.S., Waldren, R.P. & Teare, I.D. 1973. Rapid determinationof free proline for water stress studies. Plant and Soil, 109:372-381.
Belgini, M.V. & Lamattina, L. (2001(b)). Nitric oxide: A non-traditional regulator of plant growth. Trends Plant Science, 6:508-509.
Bray, E. 1993. Molecular responses of water deficit. Plant physiology,103: 1035-1040.
Chance, B. & Maehlt, A.C. 1955. Assays of catalase and peroxidases.Methods in Enzymology, 2: 764-775.
Chen, S. & Schopfer, P. 1999. Hydroxyl radicals productions inphysiological reactions. A novel function of peroxides. EuropeanJournal of Biochemistry, 260: 726-735.
Clark, D., Durner, J., Navrre, D.A. & Klessing, D.F. 2000. Nitricoxide inhibition of tobacco catalase and ascorbate peroxidase.Molecular Plant-Microbe Interactions, 13: 1380-1384.
Coombs, J.D., Hall, O., Long, S.P. & Scunlacki, J.M.O. (Eds.) 1985.Techniques in Bio-productivity and Photosynthesis. PergamonPress, Oxford, New York. 298pp.
Crawford, N.M. & Guo, F.Q. 2005. New insights into nitric oxidemetabolism and regulatory functions. Trends in Plant Science,10: 195-200.
Del Rio, L.A., Corpas, F.J. & Barroso, J.B. 2004. Nitric oxide andnitric oxide synthase activity in plants. Phytochemistry, 65: 783-792.
Fry, S.C. 1986. Cross-linking of matrix polymers in the growing cellwalls of angiosperms. Annual Review of Plant Physiology, 37:165-186.
Heeften, C.A., Besford, R.T. & Wellburn, A.R. 1996. Effects of NO(+ NO2) pollution on growth, nitrate reductase activities andassociated protein contents in glasshouse lettuce grownhydroponically in winter with CO2 enrichment. New Phytologist,133: 495-501.
Hsu, Y.T. & Kao, C.H. 2004. Cadmium toxicity is reduced by nitricoxide in rice leaves. Plant Growth Regulator, 42: 227-238.
Jiang, M.Y., Yang, W.Y. & Xu, J. 1994. Active oxygen damage effectof chlorophyll degradation in rice seedling under osmotic stress.Acta Botanica Sinica, 41(3): 229.
Lamattina, L., Garcia-Mata, C., Graziano, M. & Pagnussat, G. 2003.Nitric oxide: the versatility of an extensive signal molecule. AnnualReview Plant Biology, 54: 109-136.
Leshem, Y.Y. & Haramaty, E. 1996. Plant aging: the emission of NOand ethylene and the effect of NO-releasing compounds on growthof pea (Pisum sativum) foliage. Journal of Plant Physiology, 148:258-263.
Lipton, S.A., Choi, Y-B., Pan, Z.H., Le, S.Z., Chen, H.S-V., Sucher,N.J., Singel, D.J., Loscalzo, J. & Stamler, J.S. 1993. A redoxbased mechanism for the neuroprotective and neurodestructiveeffects of nitric oxide and related nitroso-compounds. Nature,364: 626-632.
Maggio, A., Miyasaki, S., Veronese, P., Fujita, T., Ibeas, J.I., Damasz,B., Narasimhan, M.L. Hasegawa, P.M., Joly, R.J. & Bressan,R.A. 2002. Does proline accumulation play an active role in stressinduced growth reduction? The Plant Journal, 31: 699-712.
Malik, C.P., Kaur, B., Verma, A., & Wadhwani, C. 2007. The role ofNitric Oxide and Brassinolide in the amelioration of lowtemperature effect on fiber elongation in Gossypium arboreum L.Journal of Plant Science Research, 23(1): 47-50.
Malik, C.P. & Verma, A. (Eds.) 2009. The mechanisms of cotton fiberdevelopment. In: Biotechnology: Cracking new Pastures. M. D.Publications Pvt. Ltd., Delhi
Murashige. T. & Skoog, F. 1962. A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol. Plant 15 473-497
Neill, S.J., Desikan, R. & Hancock, J.T. 2003. Nitric oxide signallingin plants. New Phytologist, 159: 11-35.

48 PHYTOMORPHOLOGY January–June 2010
Shi, S., Wang, Y., Zhang, L. & Zhang, L. 2005. Protective effect ofnitric oxide against oxidative stress under ultraviolet-B radiation.Nitric Oxide, 13(1): 1-9.
Uchida, A., Jagendorf, A.T., Hibino, T., Takabe, T. & Takabe, T. 2002.Effects of hydrogen peroxide and nitric oxide on both salt andheat stress tolerance in rice. Plant Science, 163: 515-523.
Van Assche, F. & Clijsters, H. 1990. Effect of heavy metals on enzymeactivity in plants. Plant Cell Environment, 13: 195-206.
Wendehanne, D., Durner, J. & Klessig, D.F. 2004. Nitric oxide: a newplayer in plant signalling and defense responses. Current Opinionin Plant Biology, 7: 449-455.

KISHORE S. RAJPUT et al. – Stem Anatomy of Strychnos Bicolor Prog. (Loganiaceae) from Brazilian Cerrado 49Phytomorphology 60 (1 & 2) 2010, 49-57
IntroductionIt is well known that lianas are important constituentsof forest ecosystems contributing to many vital aspectsof forest dynamics, structure, and composition (Isnard& Silk, 2009). Occurrence of climbing plants has beenreported in more than 133 families of angiosperms(Gentry, 1991) and they may represent 10–45% of woodystems in some tropical forests (Schnitzer et al., 2005).Climbers are also said to comprise as much as 40% ofthe diversity of woody species (Schnitzer & Bongers,2002). Due to their species diversity and abundance inforests, it is now widely admitted that lianas have to betaken into account in silviculture before or after logging(Putz et al., 2008) as well as in forest vegetation models(Phillips et al., 2002).
According to Phillips et al. (2002) and Wright etal., (2004) lianas abundance in forest ecosystems isapparently increasing day by day as compared to treespecies. They proposed that lianas respond strongly toincreasing CO2 concentration and/or benefit from
Stem Anatomy of Strychnos bicolor Prog. (Loganiaceae) fromBrazilian Cerrado
Kishore S. Rajput1,2, Marina B. Fiamengui1 and Carmen R. Marcati11 Depto. Recursos Naturais - Ciências Florestais, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista,
Campus de Botucatu, SP, CP237, CEP18603-970, Brasil2 Department of Botany, The M.S. University of Baroda, Vadodara-390002, India
ABSTRACT
Stem anatomy was investigated in a woody climber, Strychnos bicolor Prog., of the Loganiaceae. Inter- and intraxylarysecondary phloem was present. Secondary xylem was diffuse porous with indistinct growth rings and was composedof tracheids, vessels (both wider and narrow vessels), fibres with bordered pits, axial and ray parenchyma cells. Severalirregularly arranged phloem islands of various shapes and dimensions (interxylary phloem) were embedded in thickwalled xylem cells. Interxylary phloem development begins as small segments of the cambium inward developmentof secondary xylem was retarded and gradually stopped while outward development of secondary phloem continued.The vascular cambium formed a depression in the areas where formation of xylem was stopped. Development of thesedepressions resulted in detachment of such segments from rest of the cambial cylinder. New cambial cells were originatedon outer margin from the phloem parenchyma cells located outside to these segments. These newly formed cambialcells joined with the detached ends and completed the cambial cylinder. The cambial cylinder continues dividingbidirectionally producing secondary xylem internally and phloem externally. Formation of each new segment followedsimilar pattern thus leading to complete embedding of the sunken segments of the cambium that ceased temporarilyto divide internally. In the thick stems, peripheral pith parenchyma cells located between protoxylem and interxylaryphloem dedifferentiated into an internal cambium. This internal cambium produced secondary phloem centripetally, theintraxylary phloem.
Key words: Included phloem, Interxylary phloem, intraxylary phloem, Loganiaceae, Strychnos
Author for Correspondence: Kishore S. Rajput, e-mail: [email protected]
anthropic perturbation and increasing forestfragmentation (Londré & Schnitzer, 2006; Mohan et al.,2006; Zotz et al., 2006). These publications have openeda large debate on the potential societal and economicimpacts of increasing vigour and abundance of lianas,particularly in the tropics (Isnard & Silk, 2009). Therefore,it is an important issue to understand that differentspecies may respond differently to environmental changessuch as atmospheric CO2 concentration, forestfragmentation, and changes in tree communities. Thus,there is a great need to study development, physiology,ecology, and functional ecology of climbing plants inresponse to environmental changes (Isnard & Silk, 2009).
Anatomical/structural transformation is one of theimportant aspects of the climbing species. In the earlystage of life they are self supporting, therefore, the stemstiffness is higher but as they start climbing theirarchitecture changes and develop wide vessels, widerays, and abundant parenchyma as they rely on thesupporting object. In climbers such as Condylocarpon

50 PHYTOMORPHOLOGY January–June 2010
(Apocynaceae), Strychnos (Loganiaceae) and Bauhinia(Caesalpiniaceae) with a woody, self-supporting growthphase, there is an abrupt change from latewood withthick-walled fibers and high frequency of narrow vesselsto early compliant wood, with a higher frequency oflarge diameter vessels which causes a drastic reductionof stem stiffness (Rowe & Speck, 2005; Rowe et al.,2004; Isnard & Silk, 2009). Origin of different typesof cambial variants are said to be result of the climbinghabit, which is achieved by different patterns of secondarygrowth (Isnard & Silk, 2009), being the formation ofinterxylary phloem one of it.
Interxylary or included phloem are the terms appliedto the phloem produced centripetally from the cambiumor externally from isolated cambial segments embeddedwithin the secondary xylem as in Strychnos and inseveral species of the Bignoniaceae (Mikesell & Popham,1976; Carlquist, 1988). Outer and Veenendaal (1995)are of the opinion that scattered, isolated strands ofsecondary phloem embedded in secondary xylem shouldbe referred as diffuse interxylary phloem instead ofinterxylary or interxylary phloem of the foraminate typeas stated by IAWA Committee (1989). The term“included” is a misnomer in the case of dicotyledonswith successive cambia because conjunctive tissue inthose species is either formed as a background tissueor as bands between one vascular band and another, andby definition the phloem is thus not “included” withinsecondary xylem in species with successive cambia(Carlquist, 1988). According to him the term includedphloem is not to be confused with the phloem producedby successive cambia. In the latter, phloem is producedoutwardly from each of the successive cambia andtherefore lies between conjunctive tissue (which is notsecondary xylem) and the secondary xylem producedby each of the cambia. Thus, the phloem is neither“interxylary” nor “included” in dicotyledons withsuccessive cambia. Interxylary phloem is producedcentripetally or centrifugally by a single cambium andthus occurs as strands intercalated in a secondary xylembackground.
Occurrence of interxylary phloem in the species ofStrychnos is a constant feature and the opinion on itsorigin seems to be almost unanimous. Development ofinterxylary phloem in different species of Strychnos hasbeen studied time to time by different workers in differentspecies with varying habit such as trees, shrubs,sarmentose lianas etc., (Scott & Brebner, 1889; Solereder,1908; Chalk & Chattaway, 1937; Cockrell, 1941;
Duvigneaud et al., 1952; Obaton, 1960; Leeuwenberg,1969; Metcalfe, 1983; Veenendaal & Outer, 1993) butthere is no information on Strychnos bicolor. Moreover,most of these studies are based on the development ofinterxylary phloem in different species of Strychnos.Therefore, present investigation is aimed to provide theinformation on general histological events of inter- andintraxylary phloem developed in Strychnos bicolor, itscorrelation with the habit and comparison with theavailable literature on different species of Strychnos.
Materials and MethodsStem samples from two plants of Strychnos bicolor Prog.(Loganiaceae) growing at Rio Bonito, a cerradão area(one of the physiognomies of the cerrado sensu lato)at Botucatu municipality, São Paulo state, Brazil, wascollected. The climate is Cwb (mesothermal with drywinters), according to Köeppen’s (1931) classification,with July being the coldest and driest month of the year.Brazilian Cerrado, a savanna-like ecosystem, ischaracterized by dystrophic, acidic soils, which areextremely low in available nutrients and have high levelsof soluble few form Aluminium. These conditions resultin the typical xeromorphic appearance of its vegetation(Ribeiro & Walter, 1998; Coutinho, 2002; Franco, 2002).
Eight to ten segments of about 8–30 mm thick and40–60 mm in length from the base (30 cm above theground), middle (5-6 meter above ground) and top portion(about 8-10 mm thick and 10-12 meter above ground)were collected once in two different years (i.e. September2008, June 2009). The samples were fixed immediatelyin FAA70 (Berlyn & Miksche, 1976) and passed by avacuum pump for half an hour. After 12 hrs of fixationin FAA, suitable trimmed samples were transferred to70% alcohol. Samples were embedded in PEG (30%,50%, 70%, 90% followed by 2X100% PEG) andprocessed by routine method as described by Richter(1981), modified by Antonio Carlos Franco Barbosafrom Laboratório de Madeiras e Produtos Derivados(LMPD), CT-Floresta, IPT, São Paulo, Brazil. Transverse,radial and tangential longitudinal sections of 12–15 μmthickness were obtained with sliding microtome andstained with 1% aqueous solution of safranin and astrablue (Roeser, 1972). After staining the slides weremounted in Entelan® synthetic medium.
To obtain the length and width of cells, small piecesof xylem adjacent to the outermost cambium ring weremacerated with Franklin’s solution (Berlyn & Miksche,1976) at 55 to 60oC for 12-24 hrs, and stained with

KISHORE S. RAJPUT et al. – Stem Anatomy of Strychnos Bicolor Prog. (Loganiaceae) from Brazilian Cerrado 51
ethanolic safranin to study general morphology anddimensional details. Length and width of the sieve tubeelements were measured directly from the tangentiallongitudinal sections. Thirty measurements were chosenrandomly to obtain mean and standard deviation for eachcell type. Important results were micro-photographedwith Carl Zeiss trinocular research microscope.
ResultsStructure and development of the young stem: a singlevascular cambium is responsible for the secondary growthin the main stem. After forming 2-3 mm of secondaryxylem, small segments of the cambium ceased to divideinwardly but continue to produce secondary phloemoutside. Such behaviour of cambium led to the formationof depressions. A new set of cambial cells originatedoutside to these depressions from the parenchyma cellsand retained the circular outline of the cambium. Thisresulted into a formation of several interxylary phloemislands embedded in the secondary xylem (Fig. 1A). Inthe young stem, 4-6 layered cork cells formed outermostlayer but as the secondary growth progress further,number of cells in the periderm region increased (2–3 mm in thickness) in the 20 mm thick stems (Fig. 1B).These cells were characterised by relatively wide lumenwith thick, sclerosed walls on peripheral side of the stem.Oval to polygonal parenchyma cells beneath peridermcomposed the cortex with a continuous ring of sclereidsand can be found in the middle of it. In the thicker stems,the cortical parenchyma became tangentially elongateddue to increase in the stem diameter that exerted apressure on the cortical parenchyma (Fig. 1B). Therefore,a continuous ring of stone cells was also fragmented butits continuity was retained by the re-differentiation ofadjacent cortical cells into new stone cells. Pericycleconsisted of rod shaped elongated cells which were notmuch distinct but can be identified by the isolated strandsof pericyclic fibres.
The central portion of the stem was composed ofthin walled parenchymatous pith. In the centre, pith cellswere often intermingled with group of more or lessisodiametric stone cells in the centre of pith. Peripheralportion of the pith was also characterised by the presenceof isolated bundles of intraxylary phloem. Abundanceof prismatic/rod shaped crystals were also observed inthe parenchyma cells of the pith as well as in the corticalregion of all the samples studied.Structure of secondary xylem: Secondary xylem wasdiffuse porous with indistinct growth rings (Fig. 1A).Several irregularly arranged phloem islands of various
shapes and dimensions were embedded in thick walledxylem cells (Fig. 1A). Xylem was composed of tracheids,vessels (both wider and narrow vessels), fibres withbordered pits, axial and ray parenchyma cells whilefibriform vessels were not seen in any of the samplesinvestigated.
Wider vessels were mostly solitary and possessedsimple perforation plate on its oblique end walls. Theywere 273–571 (417) μm in length and 79–176 (124) μmin tangential diameter. On the other hand, narrow vesselswere relatively larger than the fibriform vessels reportedfor other scandent species. They were relatively longerthan the wider vessel element with 293–588 (378) μmin length and 28–63 (44) μm in tangential diameter.
The xylem fibres were characterised by the presenceof bordered pits (Fig. 1E, F), thick lignified walls, narrowlumen and tapering ends. The bordered pits were ovalto circular in outline measuring about 4.3–6.2 (4.6) μmin tangential diameter with very distinct margins. Thepit apertures formed a narrow angle with the fibre axis.Fibres were 739-1200 (960) μm in length and 7–16 (12)μm in width.
Occurrence of pith flecks was a common feature inall the samples investigated (Fig. 1C, D). In transverseview they appear as conspicuous irregular islands oflarger parenchyma cells with different dimensions andshape (Fig. 1C, D) ranging from 500 μm to 3000 μmin tangential and 95 μm to 2785 μm in radial plane.
Rays were mostly uniseriate but 2–3 seriate rayswere also observed frequently. They were heterocellular,in which terminal ray cells were procumbent and verticallyelongated while rest of them were more or lessisodiametric. They were thick and lignified in the xylemregion while unlignified and thin walled when traversingthrough the interxylary phloem island with relativelylarge, oval to circular simple pits.Development of interxylary phloem: Initially thecambium formed secondary xylem internally andsecondary phloem externally. In small segments of thecambium inward development of secondary xylem wasretarded and gradually stopped (Fig. 2A) while outwarddevelopment of secondary phloem continued. Thus, thevascular cambium formed a depression in the areaswhere formation of xylem was stopped (Fig. 2A).Development of these depressions resulted in detachmentof such segments from rest of the cambial cylinder (Figs.1B, 2B). New cambial cells were originated on outermargin from the phloem parenchyma cells located outsideto these segments (Fig. 2C–F). These newly formed

52 PHYTOMORPHOLOGY January–June 2010
cambial cells joined with the detached ends and completedthe cambial cylinder (Fig. 3A). The cambial cylindercontinues dividing bidirectionally producing secondaryxylem internally and phloem externally. Formation ofeach new segment followed similar pattern thus leadingto complete embedding of the sunken segments of thecambium that ceased temporarily to divide internally.
The embedded cambial segments of each phloem islandremained active forming secondary phloem externallywhile differentiation of xylem cells was very slow.
Sieve tubes were rarely formed in the externalsecondary phloem therefore; most of the phloem elementswere developed in the form of interxylary islands (Fig.2B, D). Phloem formed by the cambium cylinder wasmostly composed of parenchyma cells that contain
Fig. 1: Transverse (A–D) and tangential longitudinal (E, F) viewof secondary xylem of Strychnos bicolorA. Mature stem showing islands of interxylary phloem
(arrowheads). Arrow indicates predominance of uniseriaterays while multi-seriate rays are rare.
B. Gross structure of stem showing, periderm, cortex(arrowhead) and part of the cambial segment that ceased toform xylem (lower arrow). Arrow on upper right cornerindicates part of the functionally normal cambium.
C. Pith flecks in the mature stem showing deformed xylemderivatives as compared to other part (arrowhead).
D. Enlarged view of pith fleck showing cell morphology(arrowhead).
E. Xylem fibres with bordered pits (arrowheads).F. Xylem fibres with bordered pits (arrowheads).Figure 1(A–D): Scale bar = 100 μm; E, F: Scale bar = 75 μm
Fig. 2(A–F): Transverse view of stem showing development ofinterxylary phloem in Strychnos bicolorA. Cessation of secondary xylem formation in small segment of
cambium (arrow). Note the sunken part of the cambium ascompared to other ends of the cambium.
B. Disconnected cambial segment. Arrowhead indicatesfunctionally normal cambium on upper right corner.Arrowhead on lower side showing functionally retardedsegment of cambium that ceased to produce secondary xylem.
C. Cessation of secondary xylem formation in small segment ofcambium (arrow). Note the other ends of the cambium.Arrowhead indicates newly developing cambial segment outside to the portion that ceased to divide.
D. Part of the cambial segment that stopped production ofsecondary xylem. Arrowheads indicate phloem parenchymathat dedifferentiates into new segment of the cambium.
E. Development of new cambial segment (arrowhead) outsideto the cambium that stopped producing secondary xylem.
F. Development of new cambial segment (arrowhead) outsideto the cambium that stopped producing secondary xylem.
(A–D): Scale bar = 100 μm; E, F: Scale bar = 75 μm

KISHORE S. RAJPUT et al. – Stem Anatomy of Strychnos Bicolor Prog. (Loganiaceae) from Brazilian Cerrado 53
rhomboidal to rod shaped crystals. As compared to thisphloem, the interxylary phloem was characterized byhaving parenchyma cells larger in diameter (Fig. 3B,C) and higher in length. The interxylary phloem wascomposed of sieve tube elements, companion cells, axialand ray parenchyma cells. Length and width of interxylarysieve tube elements were measured and was about 372–385 (378) μm and 26–32 (29) μm respectively, whileit was 364 –371 (366) μm and 26–30 (27) μm, respectivelyfor external phloem. Rays were mostly uniseriate butoccasionally bi- to tri-seriate rays passing through thephloem islands that may be seen in transverse view.These rays were thin walled in the phloem islands andthick walled outside the phloem islands, in the secondaryxylem (Fig. 3B). Accumulation of more and moresecondary phloem in the older phloem islands exerteda pressure on the earlier formed phloem islands thusleading to crush and obliterate the non-functional phloem(Fig. 3B, C).Structure of the interxylary phloem islands: Interxylaryphloem islands were arranged in an irregular way (Fig.1A) and they were smaller and fewer centripetally whileconsiderably larger and more numerous towards theperiphery of the stems. Occasionally larger islands maybe seen in the inner part of the stem (Fig. 1A). Theseislands were usually tangentially elongated and moreor less oval to oblong in shape. With the developmentof secondary growth, the islands became older andembedded deep inside the thick walled xylem derivatives.Therefore, sieve tube elements became non-functionaland showed heavy accumulation of callose. Consequently,they became crushed and underwent obliteration (Fig.3B, C). The cambial arcs embedded to the inside marginof each phloem islands continued to divide periclinallyfor a fairly long period producing more phloem cells.Since islands were surrounded by thick walled lignifiedxylem elements, the new phloem cells had no chanceto expand outwards. Thus, newly formed secondaryphloem always crushed the older one, especially thesieve tubes and companion cells. Accumulation of crushedand obliterated phloem cells as a cap on outer borderof each island can be seen very well in the older phloemislands of thicker stems (Fig. 3C).Development of intraxylary phloem: Young stems ofStrychnos showed presence of intraxylary phloem fromthe beginning of primary growth. In mature stems, oldernonfunctional internal phloem strands are replaced bythe addition of new elements from adjacent pith cells(Fig. 3E, F). As the internal phloem became nonfunctional,
the strands begin to collapse; thus the space formed byobliteration of sieve tube elements is filled by theenlargement of adjacent parenchyma cells. Over timethese parenchyma cells divide and differentiate into newsieve tube elements (Fig. 3F). In some samples, smallarcs/segments of internal cambium were formed on theouter side of the internal phloem strands (Fig. 3F). Thisinternal cambium was originated by the dedifferentiationof fully matured pith parenchyma cells. Parenchymacells that gave rise to the internal cambium were locatedalong the outer periphery of internal phloem strands. Theinternal cambium was functional unidirectional and cutsoff only phloem elements centripetally whiledifferentiation of xylem derivatives were not observedin any of the samples studied. Structurally this phloemwas more or less similar to that of interxylary phloem.Like normal cambium, the internal cambium wasstructurally composed of vertically arranged fusiformcambial cells and radially elongated uni- to multi-seriaterays (Fig. 3E).DiscussionInterxylary or included phloem is referred to theoccurrence of strands of secondary phloem that areenclosed by the secondary xylem (Mikesell & Popham,1976). Various modes of interxylary phloem are reportedby the earlier workers. It may be concentric or diffusetype also referred as foraminate or island types (IAWACommittee). The diffuse or foraminate type of phloemdevelopment may be categorised into four subtypes: (i)Combretum subtype, according to Eames & MacDaniels(1947) in genera like Combretum small segments ofcambium produce phloem cells towards the inside(centripetally) for a brief period, in place of the xylemcells which are normally produced. After a brief periodof such activity, these cambium segments restore theirnormal function and bury the phloem formed inwardlywithin the xylem. A similar type of interxylary phloemdevelopment was described in Salvadora and othermembers of the Salvadoraceae (Carlquist, 2002), andin Leptadenia (Singh, 1943; Patil & Rajput, 2008). (ii)In Strychnos subtype xylem production retards oversmall arcs of the cambium and the original circular outline of the cambium was restored by complementarysegment formed outside to it (Eames & MacDaniels,1947; Philipson, 1990, Veenendaal & Outer, 1993; Outer& Veenendaal, 1995). (iii) In Azima subtype certaingroups of parenchyma cells of the secondary xylemdivide (dedifferentiate) and redifferentiate into phloemelements (Philipson 1990, Outer & Veenendaal, 1981)

54 PHYTOMORPHOLOGY January–June 2010
Fig. 3(A–E): Transverse view of stem showing development of cambium and structure of interxylary phloem and secondary xylem ofStrychnos bicolorA. Development of new cambial segment (arrowheads) outside to the cambium that stopped producing secondary xylem (arrows).
Arrow on the upper right corner shows disconnected part of the functionally normal cambium.B. Recently formed interxylary phloem island showing functional phloem (arrowhead).C. Interxylary phloem island in the inner part of the stem showing obliterated phloem (arrow) that form a cap above functional phloem
(arrowhead).D. Portion of the pith in the mature showing intraxylary phloem. Note the non-functional obliterated phloem cap (arrow). Arrowhead
indicates functional intraxylary phloem.E. Development of internal cambium on the periphery of the pith (arrowhead). Arrow indicates functional sieve tube elements. Note
the uni-multi-seriate rays developed from the internal cambium (small arrowheads).(A–C, E): Scale bar = 75 μm; D: Scale bar = 100 μm

KISHORE S. RAJPUT et al. – Stem Anatomy of Strychnos Bicolor Prog. (Loganiaceae) from Brazilian Cerrado 55
and (iv) In Calycopteris subtype, in small segments ofthe cambium, cells in the middle of the cambial zonedifferentiate into their derivatives, thus splitting thecambial zone into outer and inner segments. The outersegment restore cambial cylinder by joining with existingcambial ring while the inner one became encircled bydifferentiating xylem (Rajput et al., 2009). In the presentstudy, small segments cambium cease to producesecondary xylem like other Strychnos species and circularoutline of the cambial cylinder is restored by formingnew segment.
Occurrence of interxylary phloem is a frequent featureof the Loganiaceae and said to be widely distributed ingenera belonging to families with internal phloem(Solereder, 1908). Interxylary phloem occurs in sixgenera including Strychnos (Metcalfe, 1983). All thespecies of Strychnos are said to possess interxylaryphloem except for three species belonging to the taxonSpinosae, being S. henningsii Gilg (Leeuwenberg , 1969),S. afzelii Gilg and S. lecomtii A. Chev. (Obaton, 1960).Veenendaal and Outer (1993) concluded that absenceof interxylary phloem in S. afzelii Gilg and S. lecomtiiA. Chev. may be associated with their weedy habit asthey are sarmentose lianas.
Phloem anastomoses of axial interxylary phloemstrands in tangential and radial directions occur commonlyin most of the plant species with interxylary phloem andare regularly observed in Combretum nigricans andStrychnos (Veenedaal & Outer ,1993; Outer &Veenendaal, 1995). These strands establish a threedimensional network of secondary interxylary phloemand are considered to be the main assimilate transportingsystem of the stem. In the present investigation, we didnot notice phloem anastomoses in any of the samplesinvestigated. Absence of phloem anastomoses in S. bicolormay be associated with its habit. Other species ofStrychnos have a thick stem and are either trees or woodylianas. However, the stem diameter S. bicolor is muchsmaller. Less stem diameter could be the reason for theabsence of phloem anastomoses.
It is considered that during the course of evolutionplants have adapted various modes of secondarythickening and development of cambial variants (Gibson,1973, 1978; Carlquist, 1962, 1970, 2001). The functionof interxylary phloem is related to sudden outbursts ofgrowth and flowering events (Carlquist, 2001). Itsfunction also seems to be related with the protection ofconducting elements from the external damage due tofluctuations in climate, and other mechanical damage
including insect attack. Formation of sieve tube elementswith very small and narrow diameter on the normalposition by the external cambium while formation ofsieve tube elements with more and larger diameter byembedded cambial segments may be a kind of protectionand regeneration mechanism from the mechanical injuriesin addition to functions endorsed by earlier workers(Gibson, 1973, 1978; Carlquist, 1970, 2001; Rajput,2003). Frequent occurrence of pith flecks in all thesamples investigated is an additional evidence for theirrole in wound healing and mechanical injuries.
Various functions have been ascribed to the presenceof inter and intraxylary phloem. Schenck (1893) andmost latter workers hypothesised that different anomalousarrangements of secondary tissue including internalphloem greatly increase the mechanical flexibility ofliana stems, and prevent their breakage when stems aretwisted, coiled or bent. Dobbins and Fisher (1986),however, hypothesised that segments of vascularcambium, living phloem and parenchyma tissueembedded deeply within the secondary xylem providea reservoir for the regeneration of tissue when the stemis injured. Experimentally it has been proved that internaland included phloem helps lianas to maintain the apicaldominance in the girdle stems. It also reduces the chancesof disruption of translocated material to the root systemand gives an advantage of rapid and vigorous regenerationin wounded stems. Since lianas have narrow stems andmore leaf area ratio as compared to other woody speciesin which the same leaf area is supplied by wider stem(Dobbins & Fisher, 1986)
Occurrence of both wide and narrow vessels in asingle stem of lianas has already been documented inseveral lianoid genera of different families of angiosperms(Ewers & Fisher, 1989, Ewers et al., 1991; Carlquist &Hanson, 1991; Rajput et al., 2008). Strychnos possessboth wide as well as narrow vessels but the narrowvessels differ from the fibriform vessels of other dicot.Wide vessels are considered to be more vulnerable toair embolism (Carlquist, 1991), whereas narrow vessels(particularly fibriform vessels) are probably as effectiveas tracheids in preventing the formation of air embolism.This is because air embolisms occur far less commonlyin narrow vessels than wide ones (Ellmore & Ewers,1985, Carlquist, 1991). The combination of differentvessel diameters is considered to be an adaptive featuretowards greater potential for conduction (efficiency)while preventing air embolism (Carlquist, 1988, 1991).Wider as well as narrow vessels are reported in several

56 PHYTOMORPHOLOGY January–June 2010
lianas and it is a common feature of weak stem plants(Dobbins & Fisher, 1986; Ewers & Fisher, 1989; Fisher& Ewers, 1989).
Bordered pits mainly occur in water-conducting andmechanical cells of the secondary xylem such as invessel elements, tracheids and various fibres (Esau,1965). They may also be found in fibres and sclereidsoutside the secondary xylem (Esau, 1965). In the presentinvestigation we report presence of bordered pits inxylem fibres of Strychnos bicolor. Their presence inStrychnos may be associated with its habit and appearsto be an adaptive feature to serve as mechanical as wellas conductive tissue. As there are very few narrow vesselelements in S. bicolor, therefore along with mechanicalsupport these fibres may be playing important role likeimperforated tracheary elements in water conduction.
Intraxylary phloem, also called as medullary,perimedullary or internal phloem is known to occur inseveral families of dicotyledons. Metcalfe and Chalk(1950) listed 27 dicotyledonous families with internalphloem differentiation, but comparatively very few ofthem develop an internal cambium in addition to internalphloem (Philipson & Ward, 1965; Fukuda, 1967; Mikesell& Schroeder, 1984; Patil & Rajput, 2008; Rajput et al.2008). In S. bicolor, the intervening tissue between theprotoxylem and internal phloem underwent division(dedifferentiate) into an internal cambium. Initially thiscambium originates between the primary xylem andprimary internal phloem at two opposing areas, and laterarises in association with the other areas of internalphloem to form a discontinuous cylinder. At first thiscambium may be unidirectional producing only secondaryphloem inwardly but later it may become bidirectionalproducing both secondary phloem and secondary xylem(Rajput et al. 2008). In the present study we have notnoticed any xylem production, and only phloem wasobserved from the inner side of the cambium.AcknowledgementOne of the authors (KSR) is grateful to CAPES-PVEfor the financial assistant under Visiting ProfessorFellowship and to CNPq for grants to Fiamengui MB(Proc. 830858/1999-7-Master Grants) and Marcati CR(Proc. 301352/2008-9 - Researcher Grants).ReferencesBerlyn, G.P. & Miksche, J.P. 1976. Botanical Microtechnique and
Cytochemistry. Ames, Iowa: The Iowa State University Press,326 pp.
Carlquist, S. 1962. A theory of paedomorphosis in dicotyledonouswood. Phytomorphology, 12: 30-45.
Carlquist, S. 1970. Wood anatomy of insular species of Plantagoand problem of raylessness, Bulletin of Torrey Botanical Club,
97: 353-361.Carlquist, S. 1988. Comparative wood anatomy: Systematic,
Ecological, and Evolutionary Aspects of Dicotyledonous Wood.Springer Verlag, Berlin and Heidelberg, Germany.
Carlquist, S. 1991. Anatomy of vines and lianas stems: a reviewand synthesis. In: Putz, F.E. & Mooney H.A. (eds.) Biologyof vines. Cambridge University Press, Cambridge, pp 53-71.
Carlquist, S. 2001. Comparative wood anatomy: Systematic,ecological and evolutionary aspect of dicotyledon wood.Springer, Berlin, Heidelberg, New York.
Carlquist, S. 2002. Wood and bark anatomy of Salvadoraceae:ecology, relationships, histology of interxylary phloem. Journalof Torrey Botanical Society, 129: 10-20.
Carlquist, S. & Hanson, M.A. 1991. Wood and stem anatomy ofConvolvulaceae: A survey. Aliso, 13: 51-94.
Chalk, L. & Chattaway, M.M. 1937. Identification of woods withincluded phloem. Tropical Woods, 50: 1-31.
Cockrell, R.A. 1941. A comparative study of the wood structureof several South American species of Strychnos. AmericanJournal of Botany, 28: 32-41.
Coutinho, M.M. 2002. O bioma cerrado. In: H.L. Klein (ed.). EugenWarming e o cerrado brasileiro: um século depois. Editora Unesp,São Paulo, p.77-92.
Dobbins, D.R. & Fisher, J.B. 1986. Wound responses in girdledstems of lianas. Botanical Gazette, 147: 278-289.
Duvigneaud, P., Staquet, J. & Dewit, J. 1952. Contribution a l’ etudeanatomique des rameaux chez les sections africaines du genreStrychnos. Bull. Soc. Roy. Bot., Belgique, 85: 39-67.
Eames, A. & MacDaniels, L.H. 1947. An introduction to plantanatomy. Tata McGraw-Hill Publication Co. Ltd. pp. 311.
Ellmore, G.S. & Ewers, F.W. 1985. Hydraulic conductivity in trunkxylem of elm, Ulmus Americana. IAWA J, 6: 303-307.
Esau, K. 1965. Vascular differentiation of plants. Holt Rinehart andWinston Inc, New York.
Ewers, F.W. & Fisher, J.B. 1989. Variation in vessel length anddiameter in stems of six tropical and subtropical lianas. AmericanJournal of Botany, 76: 1452-1459.
Ewers, F.W., Fisher, J.B. & Fitchner, K. 1991. Water flux and xylemstructure in vines. In: Putz, F.E. & Mooney, H.A. (eds.), Biologyof vines. Cambridge University Press, Cambridge.
Fischer, J.B. & Ewers, F.W. 1989. Wound healing in stems of lianasafter twisting and girdling injury. Botanical Gazette, 150: 251-265.
Franco, A.C. 2002. Ecophysiology of woody plants. In: Oliveira,P.S. & Marquis, R.J. (eds.) The cerrados of Brazil. ColumbiaUniversity Press, New York, p. 178-197.
Fukuda, Y. 1967. Anatomical study of the internal phloem in thestems of dicotyledons, with special reference to its histogenesis.Jour. Fac. Sci., Univ. Tokyo III, 9: 313-375.
Gentry, A.G. 1991. The distribution and evolution of climbing plants.In: Putz, F.E. & Mooney, H.A. (eds.), Biology of vines,Cambridge University Press, Cambridge.
Gibson, A.C. 1973. Comparative anatomy of secondary xylem inCactoidae (Cactaceae). Biotropica, 5: 29-65.

KISHORE S. RAJPUT et al. – Stem Anatomy of Strychnos Bicolor Prog. (Loganiaceae) from Brazilian Cerrado 57
Gibson, A.C. 1978. Rayless secondary xylem of Halophytum.Bulletin of Torrey Botanical Club, 105: 39-44.
IAWA Committee, 1989. List of microscopic features for hardwoodidentification. IAWA Bull ns, 10: 221-332.
Isnard, S. & Silk, W.K. 2009. Moving with climbing plants fromCharles Darwin’s time into the 21st century. American Journalof Botany, 96: 1205-1221.
Köeppen, W. 1931. Climatologia. Fondo de Cultura Econômica,Buenos Aires.
Leeuwenberg, A.J.M. 1969. The Loganiaceae of Africa. VIII.Strychnos III. Meded. Landbouwhogeschool Wageningen, 69:1-36.
Londre, R.A. & Schnitzer, S.A. 2006. The distribution of lianasand their change in abundance in temperate forest over the past45 years. Ecology, 87: 2973-2978.
Metcalfe, C.R. 1983. Anomalous structure. In Metcalfe C. R. &Chalk, L. (eds.) Anatomy of dicotyledons II. Clarendon Press.pp. 52-63, 210-211.
Metcalfe, C.R. & Chalk, L. 1950. Anatomy of the dicotyledons.Oxford: Clarendon Press.
Mikesell, J.E. & Popham, R.A. 1976. Ontogeny and correlativerelationships of the primary thickening meristem in four-o’ clockplants (Nyctaginaceae) maintained under long and shortphotoperiods. American Journal of Botany, 63: 427-437.
Mikesell, J. & Schroeder, A.C. 1984. Internal phloem developmentin Pharbitis nil Chois. (Convolvulaceae). Botanical Gazette,145: 196-203.
Mohan, J.E., Ziska, L.H., Schlesinger, W.H., Thomas, R.B., Sicher,R.C., Jeorge, K. & Clark, J.S. 2006. Biomass and toxicityresponses of poison ivy (Toxicodendron radicans) to elevatedatmospheric CO2. Proceedings National Academy of Sciences,USA, 103: 9086-9089.
Obaton, M. 1960. Les lianes ligneuses a structure anomales desforets denses d’Afrique occidentale. Ann. Sc. Nat. Bot., 12: 1-220.
Outer, R.W. den & Veenendaal, W.L.H. van 1981. Wood and barkanatomy of Azima tetracantha Lam. (Salvadoraceae) withdescription of its included phloem. Acta Botanica Neerlandica,30: 199-207.
Outer, R.W. den & Veenendaal, W.L.H. van 1995. Developmentof included phloem in the stem of Combretum nigricans(Combretaceae). IAWA Jour., 16: 151-158.
Patil, V.S. & Rajput, K.S. 2008. Structure and development of inter-and intraxylary phloem in Leptadenia reticulata (Retz.) W. &A. (Asclepiadaceae). Polish Bot. Jour., 53: 5-13.
Phillips, O.L., Vasquéz Martínez, R., Arroyo, L., Baker, T.R.,Killeen, T., Lewis, S.L. & Malhi, Y. 2002. Increasing dominanceof large lianas in Amazonian forests. Nature 418: 770-774.
Philipson, W.R. & Ward, J.M. 1965. The ontogeny of vascularcambium in the stem of seed plants. Biological Reviews, 40:534-579.
Philipson, W.R. 1990. Anomalous cambia. In: Iqbal, M. (Ed.) Thevascular cambium. John Wiley and Sons, New York. pp. 201-212.
Putz, F.E., Sist, P., Fredericksen, T. & Dykstra, D. 2008. Reducedimpact logging: Challenges and opportunities. Forest Ecologyand Management, 256: 1427-1433.
Rajput, K.S. 2003. Structure of cambium and its derivatives in thecompressed stem of Canavalia ensiformis (L) DC (Fabaceae).Phyton, 41: 135-146.
Rajput, K.S., Raole, V.M. & Gandhi, D. 2008. Radial secondarygrowth, formation of successive cambia and their products inIpomoea hederifolia L (Convolvulaceae). Botanical Journal ofLinnaeus Society, 158: 30-40.
Rajput, K.S., Patil, V.S. & Rao, K.S. 2009. Development of includedphloem in Calycopteris floribunda Lamk. (Combretaceae).Journal of Torrey Botanical Society, 136: 302-312.
Ribeiro, J.F. & Walter, B.M.T. 1998. Fitofisionomias do biomacerrado. In: Sano, S.M. & Almeida, S.P. (eds.). Cerrado:ambiente e flora, São Paulo, p.89-166.
Richter, H.G. 1981. Anatomie des sekundären Xylems und der Rindeder Lauraceae. Sonderbände des Naturwiss, Vereins Hamburg5, Verlag Paul Parey: Hamburg & Berlin, p.148.
Roeser, K.R. 1972. Die nadel der schwarzkiefermassenprodukt undkunstwerk der nature. Mikrokosmos, 61: 33-36.
Rowe, N.P. & Speck, T. 2005. Plant growth forms: An ecologicaland evolutionary perspective, New Phytologist, 166: 61-72.
Rowe, N.P., Isnard, S. & Speck, T. 2004. Diversity of mechanicalarchitecture in climbing plants: An evolutionary perspective.Journal of Plant Growth Regulation, 23: 108-128.
Schenck, H. 1893. Beitrage zur Anatomie der Liane. In: Schimper,A.F.W. (ed.) Botanisches Mittheilungen. Tropica. Heft. 5, Teil2. Verlag von Gustav Fischer, Jena. pp. 1-271.
Schnitzer, S.A. 2005. A mechanistic explanation for global patternsof liana abundance and distribution. American Naturalist, 166,262–276.
Schnitzer, S.A. & Bongers, F. 2002. The ecology of lianas and theirrole in forests, Trends in Ecology and Evolution, 17: 223-230.
Scott, D.H. and Brebner, G. 1889. On the anatomy and histogenyof Strychnos. Ann. Bot., 3: 275-204.
Solereder, H. 1908. Systematische Anatomie der Dicotyledonen,405-407. Enke, Stuttgart.
Veenendaal, W.L.H. van and Outer, R.W. den. 1993. Developmentof included phloem and organisation of the phloem networkin the stem of Strychnos millepunctata (Loganiaceae). IAWAJour., 14: 253-365.
Wright, S.J., Calderón, O., Hernández, A. & Paton, S. 2004. Arelianas increasing in importance in tropical forests? A 17-yearrecord from Panama. Ecology, 85: 484-489.
Zotz, G., Cueni, N. & Körner, C. 2006. In situ growth stimulationof a temperate zone liana Hedera helix in elevated CO2,Functional Ecology, 20, 763-769.

JUDY ROUSE-MILLER et al. – Anatomy of Abscission Zones in Cajanus cajan (Millp.) 59Phytomorphology 60 (1 & 2) 2010, 59-65
IntroductionIn the Caribbean Cajanus cajan (Millp.) (pigeonpea) isused fresh as a pulse vegetable and is an importantprotein source. Reduced pod yield due to prematureabscission of flowers and young pods has been observedin many pigeonpea varieties with reported abscissionpercentages as high as ninety (Ariyanayagam 1975;Sheldrake & Saxena, 1979).
The shedding of plant organs such as leaves, fruits,flowers and floral parts at specific locations and times inthe life cycle of the plant is referred to as abscission(Roberts et al., 2002). Abscission functions primarily asa strategy to remove diseased or senescing organs andsecondarily for propagation by shedding of ripe fruitwhich contain seeds (Osborne, 1989). The study of themechanisms responsible for abscission is of scientificinterest but also has relevance for manipulation ofharvesting and yield (Osborne, 1989; Roberts et al., 2002).
Abscission zones (AZs) are usually located at thebase of petioles and pedicels where the organ is attachedto the axis, and are frequently discernable externally asa constricted area (Addicott, 1982). Anatomically thezone usually develops at the same time as the organ andis characterised by small brick-shaped cells which may
Anatomy of Abscission Zones in Cajanus cajan (Millp.)Judy Rouse-Miller2, E. Julian Duncan, Brian Cockburn and Grace Sirju-Charran
Department of Life Sciences, Faculty of Science and Agriculture, The University of the West Indies, St Augustine,Trinidad and Tobago, West Indies
ABSTRACT
Reduced pod yield due to premature abscission of flowers and young pods has been observed in Cajanas cajan (pigeonpea)with reported abscission rates as high as ninety percent. Based on histological investigations, the anatomy of the abscissionprocess in pigeonpea leaves and flowers is described and compared. Typically, in both leaf (rachis/stem and lateral rachis/rachillae) and flower (pedicle/peduncle) junctions an anatomically distinct layer is not present prior to induction ofabscission, however, at the terminal (apical) rachis/rachilla junction one is observed. At all junctions, separation occursas a result of dissolution of the middle lamellae leaving intact cells on each separation face, except at the rachis/stemsite where cell fracture occurs. Protection occurs as a result of suberisation of the exposed faces at all vegetative siteswhereas at pedicel/peduncle junctions, periderm formation was observed. The three vegetative abscission junctions differin two respects; (1) time of development and manifestation of the abscission zone and (2) separation occurs either bycell fracture or by dissolution of the middle lamellae. The vegetative and reproductive abscission junctions differ withrespect to the method of protection.
Key words: Flower junction, leaf junction, pigeonpea
Abbreviations: AZ(s) – abscission zone(s)
Author for Correspondence: e-mail: [email protected]
be two to many layers thick, however in some speciesabscission takes place in the absence of this band of cells.In others secondary cell division results in thedifferentiation of an AZ which was not previously formed(Gawali & Avery, 1950). There are two steps to theprocess; firstly a separation layer develops within theAZ and two cell layers including the separation layermove apart, and secondly protection of exposed surfacesoccurs either as a result of periderm formation orsuberisation (Von Mohl, 1960). AZ cells can be identifiedphysiologically before abscission initiation by theirresponse to ethylene (Osborne et al., 1985). This andother features suggest that abscission zone cells arebiochemically as well as morphologically distinct beforethe separation process (Roberts et al., 2000).
An understanding of the anatomy of abscissionzones in pigeonpea and of the enzymes involved in theprocess would facilitate application of non-traditionalapproaches to regulating the process. The anatomicaldifferentiation at pigeonpea leaf and flower abscissionjunctions has not been previously described. Theobjectives of this work were to identify and comparethe time of development and the anatomical changesleading to the formation AZs, separation and protectionat pigeonpea leaf and flower junctions.

60 PHYTOMORPHOLOGY January–June 2010
Materials and Methods
Plant MaterialPigeonpea seeds (cultivar ICPL 88027) were germinatedin peatmoss in seedling traysin a greenhouse under a12 hour photoperiod and 30°C-33°C daytime and28°C-30°C nighttime temperatures. One tray ofseedlings was watered daily (unstressed) until leafjunctions were harvested from non-senescent leaves ofeight-weeks old seedlings. Another tray of seedlings waswatered daily for four weeks then weekly until leafjunctions of senescent leaves were harvested fromstressed, eight-week old seedlings. Ten unstressed eight-week old seedlings were transferred to soil in aluzinctins (15 x 10 x 10 cm) and maintained under greenhouseconditions until flowering. Floral junctions wereharvested from buds and flowers at anthesis and postanthesis stages. ICPL 88027 is photoperiod insensitiveand comes into flowering approximately 50-75 daysafter sowing.
Excision of leaf and flower junctionsFor ease of description of the trifoliolate leaf junctionsthe terms rachis and rachilla will be used throughoutwith reference to petiole and leaflet stalk respectively.Both rachis and rachilla possess pulvini.
Rachis/stemJunctions were excised by making cuts along the stem0.5 cm above and below the node and mid way alongthe rachis between the point of attachment to the stemand the lateral leaflets.
Rachis/rachillaJunctions were prepared by removing the lamina of theleaflets and cutting the rachis just above and below thelateral leaflets separating the lateral from the terminal(apical) junctions.
Pedicel/peduncleJunctions were harvested from flowers at stages asdescribed by Ganpat (1991) as follows: stage 5 – anthesis,stage 6 – early post-anthesis. Cuts were made in thepeduncle above and below the point of insertion of thepedicel and midway along the latter.
All junctions at nodes 1-7 (acropetally) of non-senescent, trifoliolate leaves along the length of eachof five, unstressed, eight-week old seedlings weresampled. Five junctions of each flower stage weresampled.
Histology and MicroscopyLeaf and flower junctions were fixed in formalin-aceticacid (FAA), dehydrated in ethyl alcohol series andinfiltrated with paraffin wax (Johansen, 1940). Permanentslide of longitudinal sections of 10 μm thickness, stainedwith safranin and fast green were viewed using anOlympus BX50 microscope, and digital images weretaken using a Pixera 5.8 megapixel 48-bit CCD camerahosted on a Dell 8300graphics-intensive computer.Electron microscopy: longitudinal sections cut at 10 μmthickness on a Leitz 1512 rotary microtome and mountedon glass slides, were dried in a Tousimis critical pointdryer for 2 h before mounting on metal stubs. Themounted slides were placed in a Denton sputter coaterand coated with a thin layer of gold. The sections wereviewed and photographed with a Philips SEM 515scanning electron microscope.
Observations
Anatomical description of abscission zones, vasculatureat the junction and modes of separation and protectionThe photomicrographs in Figures 1-5 are representativeof all junctions of presenescent or senescent trifoliolateleaves at nodal positions (nodes 1-7) and of pedicel/peduncle junctions of pre or post anthesis, non-inducedor induced flowers (stages 5 and 6).
Abscission zones (presenescent leaf and non-inducedflower junctions)The points where separation of the leaflets, rachis andflower take place are indicated externally by a constrictionat the proximal end of the rachilla, rachis and pedicelrespectively. Internally at the rachis/stem junction thereare no anatomically distinct layers at the interface betweenthe cortical tissues of the stem and rachis to indicate theposition of an abscission zone (Fig. 1A).
However, at the terminal (apical) rachis/rachillajunction approximately four to five rows of small andsomewhat brick-shaped cells at the interface of therachis/rachilla are anatomically distinct from the adjacenttissues and comprise the abscission zone (Fig. 1B).Again at the lateral rachis/rachillae junctions as was thecase with the rachis/stem junction, there are no distinctrow(s) of cells at the interface where the rachis andrachilla tissue meet which can be described as theabscission zone (Fig. 1C). The photomicrograph of theflower junction also shows no evidence of an abscissionzone at the interface between pedicel and peduncle tissue(Fig. 1D).
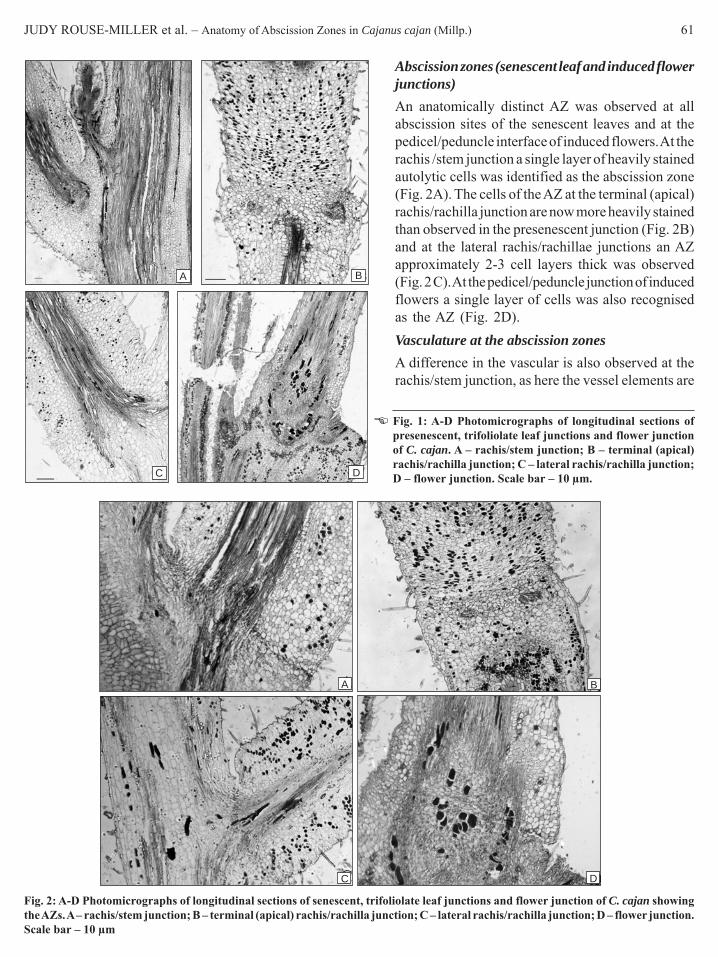
JUDY ROUSE-MILLER et al. – Anatomy of Abscission Zones in Cajanus cajan (Millp.) 61
A B
DC
Abscission zones (senescent leaf and induced flowerjunctions)An anatomically distinct AZ was observed at allabscission sites of the senescent leaves and at thepedicel/peduncle interface of induced flowers. At therachis /stem junction a single layer of heavily stainedautolytic cells was identified as the abscission zone(Fig. 2A). The cells of the AZ at the terminal (apical)rachis/rachilla junction are now more heavily stainedthan observed in the presenescent junction (Fig. 2B)and at the lateral rachis/rachillae junctions an AZapproximately 2-3 cell layers thick was observed(Fig. 2 C). At the pedicel/peduncle junction of inducedflowers a single layer of cells was also recognisedas the AZ (Fig. 2D).Vasculature at the abscission zonesA difference in the vascular is also observed at therachis/stem junction, as here the vessel elements are
Fig. 1: A-D Photomicrographs of longitudinal sections ofpresenescent, trifoliolate leaf junctions and flower junctionof C. cajan. A – rachis/stem junction; B – terminal (apical)rachis/rachilla junction; C – lateral rachis/rachilla junction;D – flower junction. Scale bar – 10 μm.
Fig. 2: A-D Photomicrographs of longitudinal sections of senescent, trifoliolate leaf junctions and flower junction of C. cajan showingthe AZs. A – rachis/stem junction; B – terminal (apical) rachis/rachilla junction; C – lateral rachis/rachilla junction; D – flower junction.Scale bar – 10 μm
A B
C D

62 PHYTOMORPHOLOGY January–June 2010
shorter at the junction than elsewhere (Fig. 3A). At theterminal rachis /rachilla junction the ring of vascularstrands of the rachis anastomose to form a plexus (Fig.3B) and at the lateral rachis/rachilla junction the vascularsystem is at its narrowest at the AZ (Fig. 3C). Againat the pedicel/peduncle AZ the vasculature is at itsnarrowest point; two strands enter the pedicel oneacropetally and the other basipetally and these strandsmerge at the junction forming an inverted v. The pointof the v is at the AZ with the pith developing just beyondthis point (Fig. 3D).
Mode of separation at the junctionsAt the rachis/stem junction cell fracture is observed(Fig. 4A). However, at the terminal (apical) rachis/rachilla and lateral rachis/rachilla leaf junctions andpedicel/peduncle flower junctions separation occurs asa result of dissolution of the middle lamella, leavingintact cells on each fracture face (Fig. 4B, C, D).
Fig. 3: A-D Photomicrographs of longitudinal sections of senescent,trifoliolate leaf junctions and flower junction of C. Cajan showingvasculature at the AZs. A – rachis/stem junction; B – terminal(apical) rachis/rachilla junction; C – lateral rachis/rachillajunction; D – flower junction. Scale bar-10 μm
A
B
C C
Fig. 4: A – D. Longitudinal sections of senescent, trifoliolate leaf junctions and flower junction of C. Cajan showing separation at theAZs. A – Electron micrograph of rachis/stem junction; photomicrographs; B – terminal (apical) rachis/rachilla junction; C – lateralrachis/rachilla junction; D – flower junction. Scale bar-10 μm
A
B
C D

JUDY ROUSE-MILLER et al. – Anatomy of Abscission Zones in Cajanus cajan (Millp.) 63
Mode of protection at the junctionsAt leaflet and rachis abscission sites the exposed cellsof the subtending axis is protected as a result ofsuberisation (Fig. 5A, B, C). However at the pedicel/peduncle junction, protection occurs as a result of thedevelopment of a periderm (Fig. 5D). Periderm is alsoseen on the exposed surface of the bract which subtendsthe flower (Fig. 5D).
The three vegetative abscission junctions differ intwo respects; (1) time of development and manifestationof the abscission zone and (2) separation occurs eitherby cell fracture or by dissolution of the middle lamellae.
Fig. 5: A-D Photomicrographs of longitudinal sections of senescent, trifoliolate leaf junctions and flower junction of C. Cajan showingprotective layer(s) at the AZs. A – rachis/stem junction; B – terminal (apical) rachis/rachilla junction; C – lateral rachis/rachillajunction; D – flower junction. Scale bar-10 μm
The vegetative and reproductive abscission junctionsdiffer with respect to the method of protection(Table 1).
DiscussionThe constriction seen at the base of the rachis, rachilla,and pedicel in pigeonpea indicated the position at whichseparation occurred. This has been observed in otherspecies such as in P. vulgaris where the region ofseparation is visible externally as a narrow, constrictedband of hyaline cells (Webster, 1968). Constricted areashave also been observed in tomato floral junctions

64 PHYTOMORPHOLOGY January–June 2010
(Roberts et al., 1984) and in cotyledon abscission ofL. angustifolius I (Clements & Atkins, 2001).
A typical abscission zone which develops at the timeof organ formation or as a result of secondary celldivisions just prior to separation does not occur inpigeonpea rachis, lateral rachillae and pedicel junctions.Gawali and Avery (1950) and van Doorn and Stead(1997) noted that abscission occurs in many species inwhich neither the typical abscission zone nor secondarycell development prior to separation occurs and concludedthat it was not a prerequisite for abscission, this appearsto be the case for pigeonpea. The zone in pigeonpea ishowever functionally distinct from the cell layers aboveand below it, as a polygalacuronase gene fragment wasamplified from the junction but not from cells aboveand below it (Rouse-Miller et al., unpublished).
Different manifestations of the AZ are observedwithin the same organ as a distinct zone is seen at theterminal (apical) rachis/rachilla junction of non senescentleaves and is not observed at the other junctions. At allother sites the AZ is anatomically distinct just prior toabscission of the organ and the number of cell layersdistinguished vary depending on the site.
In pigeonpea, separation and protection is achievedin two ways. Different ways of separation within thesame species has also been observed in peach where leafabscission zones show cell fracture and cell wall lysiswhereas at fruit abscission junctions separation occurleaving intact cells (Rascio et al., 1985; Ramina et al.,1993).
Changes in vasculature seen at the pigeonpeaabscission junctions are typical and facilitate easiershedding as one fracture point is presented. Addicott(1982) and Webster (1968) reported anatomical changesat the AZs such as shorter, less secondarily thickenedvessel elements; and protosteles formed as a result ofvascular anastomosis (Addicott, 1982). This reductionin vasculature at the junction allows easier shedding ofappendages as less mechanical force would be required
Table 1. Summary of anatomical observations of the abscission process at vegetative and reproductive abscission sites in C. cajan(pigeonpea)
Feature / junction type Rachis/ stem Lateral rachis/ Terminal (apical) Pedicel/rachillae rachis/rachilla peduncle
Distinct abscission zone present No No Yes NoSeparation-cell fracture Yes No No NoSeparation -dissolution of the middle lamella No Yes Yes YesProtection-periderm formation No No No YesProtection-suberisation of the fracture face Yes Yes Yes No
to fracture the xylem vessels following separation inother tissues (Addicott, 1982). This decreased amountof vascular tissue at abscission junctions has beenobserved for P. vulgaris leaf AZs (Webster, 1968) andoil palm fruit abscission zones (Henderson et al., 2001).Additionally, the number of points from which watercould be lost prior to formation a protective layer is alsoreduced significantly.
The anatomy of the junctions before, during andafter abscission has been described for the vegetativeand reproductive abscission junctions of pigeonpea.This is the first reference to different manifestations ofthe abscission zone in legumes. The result indicates thatfew cell layers at the junctions are involved in abscissionand as a result further investigation of the abscissionprocess should proceed to the molecular level asbiochemical studies involving enzyme activity in thecells of the zone may prove difficult.
ReferencesAbeles, F.B. 1969. Abscission: Role of Cellulase. Plant Physiology,
44: 447-452.Abeles, F.B., Morgan, P.W. & Saltveit, M.E. 1992. Ethylene in Plant
Biology. Academic Press Inc., San Diego.Abernathy, R. H., Palmer, R.G., Shibles R. & Anderson, I.C. 1977.
Histological observations on abscising and retaining soybeanflowers. Canadian Journal of Plant Sciences, 57: 713-716.
Addicott, F.T. 1968. Enviromental factors in the physiology ofabscission, Plant Physiol. 43: 1471-1479.
Addicott, F.T. 1982. Abscission. University of California Press,Berkley.
Ariyanayagam, R.P. 1975. Status of research on pigeonpea inTrinidad. In: Proceedings of the International Workshop onpigeonpeas, ICRISAT/ICAR Vol 1 ICRISAT, Pantancheru, India.
Clements, J. & Atkins, C. 2001. Characterization of a non-abscissionmutant in Lupinus angustifolius I.: Genetic and structuralaspects. American Journal of Botany, 88: 31-42.
Dolly, D.I. 1993. Pigeonpea variety acceptance, an analysis ofresearch and adoption issues in Trinidad and Tobago. PhD. Diss.The University of the West Indies: St. Augustine.
Ganpat, M.M. 1991. Physiological and Biochemical studies onabscission of reproductive structures of pigeonpea (Cajanus

JUDY ROUSE-MILLER et al. – Anatomy of Abscission Zones in Cajanus cajan (Millp.) 65
cajan (L.) Millsp. M.Phil. Thesis. The University of the WestIndies, St. Augustine.
Gawali, A.G. & Avery Jr, G.S. 1950. Leaf abscission and the so-called abscission layer. American Journal of Botany, 37: 172-180.
Henderson, J., Davies, H.A., Heyes, S.J. & Osborne, D.J. 2001.The study of a monocotyledon abscission zone using microscopic,chemical, enzymatic and solid state 13 C CP/MAS NMRanalyses. Phytochemistry, 56: 131-139.
Johansen, D.A. 1940. Plant Microtechnique, (Mc Graw Hill: NewYork).
Osborne, D.J. 1989. Abscission. Crit. Critical Reviews in PlantSciences, 8: 103-129.
Osborne, D.J., Mc Manus, M.T. & Webb, J. 1985. Target cells forethylene action, In: Roberts, J.A. & Tucker, G.A. (eds.) Ethyleneand Plant Development. Butterworths, London. pp. 197-212.
Osborne, D.J. & Sargent, J.A. 1976. The positional differentiationof abscission zones during the development of leaves ofSambucus nigra and the response of the cells to auxin andethylene. Planta, 132: 197-204.
Ramina, A. Casadoro, G. & Rascio, N. 1993. Structural biochemicaland molecular aspects of abscission in peach. Acta Hort., 329:211-217.
Rascio, N., Casadoro, G., Ramina, A, & Masia, A. 1985. Structuraland biochemical aspects of peach fruit abscission (Prunus,persica L. Batsch). Planta, 164: 1-11.
Roberts, J.A., Elliott, K.A. & Gonzaìlez-Carranza, Z.H. 2002.Abscission, dehiscence, and other cell separation processes.Annual Review of Plant Biology, 53: 131-58.
Roberts, J.A., Schindler, B. & Tucker, G.A. 1984. Ethylene promotedtomato flower abscission and the possible involvement of aninhibitor. Planta, 160: 159-163.
Roberts, J.A., Whitelaw, C.A., Gonzaìlez-Carranza, Z.H. & McManus, M.T. 2000. Cell separation processes in plants – models,mechanisms and manipulation. Annals of Botany, 86: 223-225.
Sexton, R. & Hall, J.L. 1974. Fine structure and cytochemistry ofthe abscission zone cells of Phaseolus leaves. I. Ultrastructuralchanges occurring during abscission. Annals of Botany, 38: 849-854.
Sheldrake, A.R. & Saxena, N.P. 1979. Comparisons of earlier andlater formed pods of chickpeas (Cicer arietinum L.). Annalsof Botany, 43: 467-473.
van Doorn, W.G. & Stead, A.D. 1997. Abscission of flowers andfloral parts. Journal of Experimental Botany, 48: 821-837.
Ververidis, P. & John, P. 1991. Complete recovery in vitro ofethylene-forming enzyme activity. Phytochemistry, 30: 725-727.
von Mohl, H. 1960. Ueber die anatomischen Veranderungen desBlattgelenkes. Welche das Adfallen de Blatter herbeifuhren, Bot.Zeitg., 18: 1-7 Addicott FT 1982 Abscission (University ofCalifornia Press: Berkley).
Webster, B.D. 1968. Anatomical studies of abscission. PlantPhysiology, 43: 1512-1544.
Webster, B.D. & Chui, H.W. 1975. Ultrastructural studies ofabscission in Phaseolus: characteristics of the floral abscissionzone. Journal of American Society of Horticultural Science, 100:613-618.
Wiersema, J. & Leon, B. 1999. World Economic Plants. A StandardReference, CRC Press, London.





























