Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY ic 1984 by The American Society of Biological Chemists, Inc
Vol. 259, No. 1, Issue of January 10, pp. 475-480, 1984 Printed in U.S.A.
Detection of Pheromone Biosynthetic and Degradative Enzymes in Vitro*
(Received for publication, March 21, 1983)
David Morse$ and Edward Meighen From the Department of Biochemistry, McGill University, Montreal, Quebec, H3G 1 Y6 Canada
Highly sensitive, luminescent assays have been de- veloped to measure enzyme activities involved in the metabolism of a major class of insect pheromones which includes fatty aldehydes, alcohols, and their acetate esters. These assays have been applied to measure the in vitro biosynthesis and degradation of the sex pher- omone (transxis-1 1-tetradecenal, 96:4) of the eastern spruce budworm, Choristoneura fumiferana. Three activities were detected on analyses of extracts of the female moths: (a) an esterase that hydrolyzes both the cis and trans isomers of 1 1-tetradecenyl acetate, (b) an oxidase that converts fatty alcohols to aldehydes in the presence of Oar and (c) an NAD-dependent aldehyde dehydrogenase. The coupled luminescent response of bacterial luciferase to long chain aldehydes was used to measure rates of reaction as low as 0.1 pmol/min since only low amounts of material can be analyzed. Specific activities of these enzymes were higher in the pheromone producing gland than in other parts of the moth, implicating these enzymes, and the oxidase in particular, in the pathway of pheromone biosynthesis. The pathway was supported in vivo by demonstrating that topical application of 3H-labeled tetradecanyl ace- tate onto the insect gland resulted in the formation of [3H]tetradecanol and [3H]tetradecanoic acid, thus pro- viding evidence that all three enzymes were functional in the living insects.
Almost all insect species have highly effective communica- tion systems that involve the release of special compounds known as pheromones. A pheromone is a chemical, or more generally, a precise blend of chemicals, that affects the behav- ior of insects of the same species (Weaver, 1978; Silverstein, 1981; Mayer and McLaughlin, 1975). In view of the potential contamination of the environment by insecticides, phero- mones have practical importance since they can be used to monitor and/or control insect populations either as lures in traps, or by disrupting the normal communication process (Silverstein, 1981).
The identification of the chemical structures of pheromones has advanced very rapidly since 10,12-hexadecadien-1-01 was shown in 1959 to be the sex pheromone of the silkworm moth, Bombyx mori (Butenandt et al., 1959). The pheromone com- ponents of moths are typically long chain unsaturated ace- tates, alcohols, and aldehydes (Mayer and McLaughlin, 1975). However, biochemical studies on the biosynthesis of phero- mones are extremely limited (Bjostad and Roelofs, 1981;
* This research is supported by Medical Research Council Grant MT-4314. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a Medical Research Council Studentship.
Dillwith et al., 1981) and only in a few instances have in uitro enzyme activities been measured (Clearwater, 1975; Hedin, 1977; Inoue and Hamamura, 1972; Weatherston and Percy, 1976). The paucity of biochemical studies on insect phero- mones can be at least partially traced to the low amounts of material available and the absence of rapid and sensitive approaches for measuring the degradation and biosynthesis of pheromones. Biochemical investigations of pheromone me- tabolism are relevant to the effective application of phero- mones in controlling insect populations (Weaver, 1978; Sil- verstein, 1981) and could lead to alternative pest control methods.
The eastern spruce budworm, Choristoneura fumiferana, is one of the major insect pests in the United States and Canada, causing millions of dollars in damage annually to the forest industry. The sex pheromone consists of two long chain aldehydes, trans-11-tetradecenal (96%) and cis-ll-tetrade- cenal(4%) (Sanders and Weatherston, 1976; Silk et al., 1980). The pheromone, which attracts male moths of the same species, is secreted from a specialized gland located at the end of the abdomen of the female moth (Roelofs and Feng, 1968). The gland contains little of the aldehyde, but relatively large amounts of 11-tetradecenyl acetate (96:4, tram:cis) which has led to the suggestion that the acetate ester is a stored precursor of the pheromone (Silk et al., 1980).
Recently, we have developed an extremely sensitive and rapid bioluminescence assay for long chain aldehyde phero- mones found in insects (Meighen et al., 1981; Meighen et al., 1982; Grant et al., 1982). Since this analysis involves mea- surement of aqueous solutions of the compounds at the picom- ole level, it provides a readily applicable approach for studying the metabolism in uitro of aldehyde pheromones and their precursors.
The present paper reports the discovery of three enzyme activities involved in pheromone metabolism in extracts of the gland of the eastern budworm: acetate esterase, alcohol oxidase, and aldehyde dehydrogenase activities. Lumines- cence assays were applied to measure the production and removal of long chain aldehyde as well as to measure the production of long chain alcohol using a coupled enzyme system to convert alcohol to aldehyde. Since analogous en- zyme activities may be present in other insects with phero- mones of similar structure, including such major pests as the corn earworm, the tobacco budworm, and the navel orange- worm (Grant et al., 1982), the approaches and techniques developed in this paper may be generally applicable for future biochemical investigations on enzymes involved in pheromone biosynthesis and degradation in a variety of different insects.
EXPERIMENTAL PROCEDURES’
’ Portions of this paper (including “Experimental Procedures,” Tables IS and IIS, and Figs. 15“) are presented in miniprint a t the
475
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from

476 Pheromone Metabolism in Vitro
RESULTS
Three enzyme activities involved in the metabolism of the aldehyde pheromone of the spruce budworm were detected in uitro in extracts of the gland of the female moth. The enzymes include an acetate esterase, a fatty alcohol oxidase, and a fatty aldehyde dehydrogenase catalyzing the reactions shown in the scheme below:
RCH20COCH3 - RCH20H 2 RCHO - RCOOH
Detection of these enzymes was achieved by the highly sen- sitive luminescent response of bacterial luciferase to long chain aldehydes enabling rates in the picomoles/min (nanom- olar/min) range to be measured directly (see Miniprint). Low amounts of the gland extracts were used in the enzyme assays as high protein concentrations resulted in a decrease in lu- minescent response to aldehydes, and only limited quantities of enzyme were available.
Acetate Esterase-Acetate esterase activity was determined using the luminescence assay after conversion of the alcohol product into the corresponding aldehyde with NAD and horse liver alcohol dehydrogenase. The rates obtained with trans- 11- and cis-11-tetradecenyl acetate as well as the saturated ester a t different substrate concentrations are shown in Fig. 4s (see Miniprint). The enzyme showed a slight preference for the trans-11 isomer, having a K,,, and Vmax of 0.3 p ~ , and 80 pmol/min/gland (4 pmol/min/pg), respectively, for this compound.
Alcohol Oxidase-The activity of alcohol oxidase in gland extracts was determined by the luminescence response of bacterial luciferase to the aldehyde product and also showed hyperbolic curves as a function of substrate concentration (Fig. 4s). This enzyme also appeared to have a preference for trans-1 1-tetradecenol over the cis-1 1 and saturated isomers. The K , values for the alcohols were between 0.1 and 0.2 p~ with the rate of oxidation of alcohol being 10 to 20 pmol/ min/gland at saturating substrate concentration. Although the maximum oxidase oxidase activity is about 5-fold lower than the esterase activity in the gland, these rates are more comparable at low, nonsaturating substrate concentrations ("0.1 p ~ ) since the K , for alcohol is about 3-fold lower than that of the ester.
The activity of the oxidase was clearly dependent on the presence of molecular oxygen as saturation of the reaction buffer with N, reduced the reaction velocity and rates were restored to normal when the mixture was exposed to the atmosphere (Fig. 1). This finding was in agreement with the inability of nicotinamide (NAD, NADP) or flavin (FMN, FAD) cofactors to stimulate the activity.
The metabolism of both tritium-labeled tetradecanyl ace- tate and tritium-labeled tetradecanol catalyzed by the gland extract is illustrated in Fig. 2. In the absence of NAD, alcohol is produced from the tetradecanyl acetate (lane 2), and alde- hyde is produced from the tetradecanol (lane 4 ) . In the pres- ence of NAD, the aldehyde is further oxidized to the acid (lane 5 ) , showing that an aldehyde dehydrogenase activity exists in the gland extract as well, and that the product of this reaction is a long chain carboxylic acid.
Aldehyde Dehydrogenase-Aldehyde dehydrogenase activ- ity in gland extracts could be rapidly measured from the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Rethesda, MD 20814. Request Document No. 83 "0764, cite the authors, and include a check or money order for 54.40 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
Hz0 0 NAD
0 1 0 10 20 30
TIME ( m i n )
FIG. 1. Dependence of alcohol oxidase activity on the pres- ence of oxygen. Enzyme from a gland extract was added in a small volume (10 pl) to 5 ml of 0.05 M phosphate buffer, pH 7.0, that had been purged of oxygen by boiling and then cooling under nitrogen. The reaction was initiated by the addition of 1.0 p~ truns-ll-tetra- decenol in 5 pl of isopropanol and the solution either was opened immediately (0) or maintained under nitrogen for 15 min (0) before being exposed to the atmosphere. The luminescence response was measured a t various times over 15 min after exposure to the atmo- sphere.
1 2 3 4 5 -FRONT
-TETRADECANYL -TETRA DECA NAL
-TETRADECANOIC
ACETATE
ACiD
-1ETRADECANOL
-ORIGIN
FIG. 2. Metabolism of [3H]tetradecanyl acetate and [3H]te- tradecanol by extracts of the spruce budworm. ["HjTetrade- canyl acetate (2.3 nmol) was added to 1.0 ml of 0.05 M phosphate (pH 8.0) and left to react for 15 min in either the absence (lune I ) or the presence (lane 2 ) of 100 pl of gland extract. ['HJTetradecanol (1.7 nmol) was added to 1.0 ml of 0.05 M phosphate buffer (pH 7) and left to react for 20 min in the absence of gland extract (lune 3 ) , the presence of 100 pl gland extract (lune 4 ) , or 100 pl of gland extract plus 1 mM NAD (lune .5). All reactions were stopped by addition of tetradecanol (100 pg) , tetradecanoic acid (120 pg), tetradecanyl ace- tate (40 pg), and tetradecanal (20 pg). These compounds also func- tioned as carriers when the aqueous solution was extracted with 1.0 ml of hexane. The hexane phase was concentrated to approximately 15 pl, spotted onto silica gel plates and chromatographed in hex- ane:ether:acetic acid (90:10:2). Unlabeled standards were visualized with I, vapor before the plate was enhanced and fluorographed.
NAD-dependent rate of disappearance of aldehyde using a luminescence coupled assay. Since changes of <25% in the initial aldehyde concentration could not be accurately mea- sured, an integrated form of the Michaelis-Menten equation and not the initial velocity was used to determine the enzyme
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from

Pheromone Metabolism in Vitro 477
TABLE I Morphological distribution of alcohol oxidase, acetate esterase, and
aldehyde dehydrogenase activities in the spruce budworm Enzyme assays and protein determinations were performed as
described under “Experimental Procedures” using centrifuged ex- tracts. Experimental data represent the average of five separate experiments. Units are given in picomoles/min.
Oxidase Esterase Dehydro. genase
1 2 3
e- units fpg
Head 0.14 f 0.06 1.5 f 2.0 Thorax
21 f 8 0.14 f 0.12 0.4 f 0.5 22 f 22
Legs 0.07 rf: 0.08 1.0 f 1.0 16 f 21 Abdomen 0.15 f 0.02 1.7 f 1.3 Gland
18 f 18 1.1 f 0.7 4.2 f 4.8 38 f 17
activity (see Miniprint). The K , for trans-11-tetradecenal was found to be approximately 100 nM and the maximum velocity between 1 and 2 nmol/min/gland. The NAD was present at saturating concentrations (0.5 mM) since changing its concentration (0.01-1.0 mM) only resulted in a decrease in the activity at concentrations below 0.2 mM. Although activity was not observed if NADP replaced NAD, the aldehyde de- hydrogenase was found to function equally well with both the 1 I-unsaturated and saturated 14-carbon aldehydes.
Morphological Distribution of Enzymes-Since the lumines- cence coupled assays gave activities proportional to the amount of extract for not only aldehyde dehydrogenase but also for the oxidase and esterase (Figs. 2s and 3S, Miniprint), it was thus possible to examine the morphological distribution of the enzymes in the female moth of the eastern spruce budworm. All three enzymes had their highest specificity (unitslpg of protein) in extracts of the gland with lower levels in the head, thorax, legs, and abdomen (Table I). Since the percentage of the wet weight as protein was essentially iden- tical (7.2-8.696) for all tissues investigated, the specific activ- ity based on units/pg of wet weight gave similar conclusions. The alcohol oxidase appeared to have the greatest specific activity in the gland in terms of its morphological distribution, consistent with it being the enzyme directly responsible for the biosynthesis of the pheromone released from the gland. Mixing of the gland extract with extracts of other tissues showed that the lower levels of enzyme in other parts were not due to inhibitory factors present in these extracts (see Miniprint, Table IS). The polypeptide distributions by SDS2 gel electrophoresis of the extracts from the different body parts were very similar, indicating that the oxidase is not a major protein component in the gland.
Enzyme Actiuities in Vivo-To demonstrate the activity of all three enzymes in uiuo, radiolabeled tetradecanyl acetate was applied to the glands, and the products formed were analyzed by thin layer chromatography. As shown in Fig. 3, the tetradecanyl acetate was readily metabolized by the gland into both tetradecanol and tetradecanoic acid, indicating that the esterase, oxidase, and dehydrogenase could function in series in the intact gland. The absence of tetradecanal, the intermediate between the alcohol and acid, is presumably a reflection of the presence of NAD in the tissue. As shown in Fig. 2, addition of NAD to in uitro assays converts the fatty aldehyde to fatty acid. When tetradecanyl acetate was applied to either the head or the abdomen in uiuo, tetradecanol was observed with the production of tetradecanol acid being sig- nificantly lower than that obtained for the gland (Fig. 3). This in uiuo specificity is in agreement with the morphological
* The abbreviations used are: SDS, sodium dodecyl sulfate; HLAD, horse liver alcohol dehydrogenase; BSA, bovine serum albumin; DMSO, dimethyl sulfoxide.
-FRONT
(. - TETRADECANYL ACETATE -TETRADECANAL
-TETRADECANOIC ACID
- TETRADECANOL
-ORIGIN
FIG. 3. Metabolism of tetradecanyl acetate in vivo. Tritium- labeled tetradecanyl acetate was applied topically to living insects as described (Bjostad and Roelofs, 1981). The distribution of radiolabel was determined by fluorography after thin layer chromatography of the hexane extracts obtained from the tissues after a 15-min incu- bation in vivo with the glands (lane 2). the heads (lane 3). and the abdomens (lane 4 ) . The dimethyl sulfoxide solution of tetradecanyl acetate before application is shown in lane 1.
distribution of the enzyme activities measured in the in uitro assays (Table I) .
DISCUSSION
Studies on identifying and resolving the enzymes involved in pheromone metabolism have been primarily limited by the low amounts of material and the lack of suitable assays. Thus, the development of the bioluminescence assay for aldehyde pheromones, and its application in the present study to mea- sure long chain acetate esterases, fatty alcohol oxidases, and fatty aldehyde dehydrogenases, provided a significant increase in sensitivity so that enzyme activities involved in sex pher- omone metabolism could be detected in vitro. The assays are sensitive enough to measure all three activities in a homoge- nate from a single gland.
Two of the enzyme activities, an acetate esterase and an oxidase, which catalyze the formation of the aldehyde phero- mone (tranxcls-11-tetradecenal; 96:4) from trans:cis-11-tetra- decenyl acetate, are soluble enzymes. Partial resolution of the enzymes by anion exchange chromatography had no effect on their kinetic properties even though multiple molecular forms of the esterase were observed (see Miniprint). Both enzymes had higher specific activities in extracts from the pheromone- releasing gland than in extracts from the other parts of the insect. Tetradecenyl acetate has been shown by Silk et al. (1980) to be present in the same isomeric ratio and in signif- icantly higher amounts (10-40-fold) in the spruce budworm gland compared to the aldehyde pheromone. Furthermore, an increase in the acetate ester levels (-2-fold) in the gland during the calling period of the insect has led to the proposal that the acetate ester may be the precursor of the aldehyde pheromone (Silk et al., 1980). Although a daily rhythm in aldehyde levels was not observed by these workers, recent experiments have demonstrated that aldehyde levels in the gland are also significantly higher in the calling period (Morse et al., 1982). The detection of the esterase and oxidase activ-
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from

478 Pheromone Metabolism in Vitro
ities in the extracts of the budworm and the morphological specificity for the gland both in vivo and in vitro provides direct support to the proposal that the biosynthesis of the aldehyde pheromone occurs via a fatty alcohol acetate ester. Some caution must be exercised in this interpretation, how- ever, as it is possible these enzymes, and the esterase in particular, may have a more genera: role in the insect in addition to or instead of pheromone metabolism. It is of interest that the reactions catalyzed by the esterase and oxidase require no substrates other than H20 and 02, and are highly favored thermodynamically ( i e . irreversible), thus making the acetate ester an ideal precursor and storage form for the aldehyde pheromone. Conversely, the irreversibility of these reactions clearly indicates that the acetate ester is synthesized in a different set of reactions.
A NAD-dependent fatty aldehyde dehydrogenase was also found in extracts of the budworm. This enzyme, in contrast to the esterase and oxidase, appears to be membrane-bound which suggests that it may be located in a different subcellular location than the esterase and oxidase. The dehydrogenase could function to remove high concentrations of aldehydes that would be toxic to the cell. This enzyme has also been found in the male budworm including the antennae and could serve in a pheromone-clearing role which may be essential for the continuous response to pheromone molecules by the male moth. The dehydrogenase activity is high in gland homoge- nates and could explain why aldehyde was not observed in the in vivo labeling studies when tetradecanyl acetate was applied topically to the gland in dimethyl sulfoxide. The i n vivo labeling technique could not be used to show incorpora- tion of radiolabel into the volatile pheromone, as the aldehyde release rates, in the calling period (Morse et al., 1982), dropped to very low levels after application of dimethyl sulfoxide (data not shown). Nevertheless, the labeling studies did indicate that all three enzymes could function sequentially in vivo.
A wide variety of insects have been shown to have long chain aldehydes as their pheromones (Mayer and Mc- Laughlin, 1975; Grant et al., 1982). Although the luminescent assay is not advantageous for studying the specificities of enzymes for different aldehydes since luciferase only gives a high response to low concentrations of 14- to 16-carbon aldehydes (Meighen et al., 1982), fortunately, the major insect pests such as the corn earworm, the tobacco earworm, the tobacco budworm, and the navel orangeworm all have unsat- urated aldehyde pheromones of this chain length. Similarly, an even greater number of insects have been shown to have acetate esters of 14- and 16-carbon alcohols as their major pheromone component (Mayer and McLaughlin, 1975). Con- sequently, the luminescent assays should be generally appli-
cable for studying pheromone biosynthesis and degradation in a wide variety of insects. Recent experiments have shown that these esterase, oxidase, and dehydrogenase activities can be readily detected in gland extracts of the corn earworm Heliothis zea. These investigations have the potential for identification of a common pathway for pheromone metabo- lism in insects using long chain alcohols, acetate esters, or aldehydes as pheromones.
Acknowledgments-We thank Dr. Gary Grant, of the Forest Pest Management Institute, Sault Ste. Marie, for his generous supply of spruce budworm pupae, and Rozsa Szittner for her technical assist- ance.
REFERENCES Bjostad, L. B., and Roelofs, W . L. (1981) J . Biol. Chem. 256, 7936-
Butenandt, A., Beckmann, R., Stamm, D., and Hecker, E. (1959) Z.
Clearwater, J. R. (1975) Insect Biochem. 5 , 737-746 Dillwith. J. W., Blomauist, G. J., and Nelson, D. R. (1981) Insect
7940
Naturforsch. 14b, 283-284
Biochem. 11,' 247-253 Grant. G. G.. Slessor. K. N.. Szittner. R. B.. Morse. D.. and Meiahen.
E. A. (1982) J . Chem. Ecol. 8, 923-933 '
H., and Hastings, J. W. (1972) J. Biol. Chem. 247,398-404
. , - .
Gunsalus-Miguel, A., Meighen, E. A., Zeigler Nicoli, M., Nealson, K.
Hedin, P. A. (1977) J. Chem. Ecol. 3, 279-289 Inoue, S., and Hamamura, Y. (1972) Proc. Jpn. Acad. 48, 323-326 Laemmli, U. K. (1970) Nature (Lond.) 227, 680-685 Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951)
Mayer, M. S., and McLaughlin, J. R. (1975) Fl. Agric. Exp. Stn.
Meighen. E. A., Slessor, K. N., and Grant, G. G . (1981) Experientia
J . Biol. Chem. 193, 265-275
Monogr. 6
37,555-557 Meiehen. E. A.. Slessor. K. N.. and Grant. G. G. (19821 J. Chem.
Epol. 8 , 911-921 Moreau, R. A., and Huang, A. H. C. (1979) Arch. Biochem. Biophys.
Morse, D., Szittner, R. B., Grant, G. G., and Meighen, E. A. (1982)
Roelofs, W. L., and Feng, K. (1968) Ann. Entomol. SOC. Am. 61,312-
Sanders, C. J., and Weatherston, J. (1976) Can. Entomol. 108,1285-
Silk, P. J., Tan, S. H., Wiesner, C. J., Ross, R. J., and Lonergan, G.
Silverstein, R. M. (1981) Science (Wash. D. C.) 213, 1326-1332 Walker, A. C., and Schmidt, C. L. A. (1944) Arch. Biochem. Biophys.
Weatherston, J., and Percy, J. E. (1976) Insect Biochem. 6,413-417 Weaver, W. N. (1978) in Biochemistry of Insects (Rockstein, M., ed)
Wray, W., Boulikas, T., Wray, V. P., and Hancock, R. (1981) Anal.
194,422-430
J . Insect Physiol. 28, 863-866
316
1290
C. (1980) Enuiron. Entomol. 9, 640-644
5,445-467
pp. 359-389, Academic Press, New York
Biochem. 118, 197-203
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from

Pheromone Metabolism in Vitro 479
SIIPPLEWENTARY IUTERlAL
t o
DETECTION OF PHEROMONE lDSYNTHET!C AND DEGRllOATlVE ENZYMES I N VlTRO
by
David Morse and Edward Meighen
EXPERlWENTAL PROCEDURES
Mater ia ls - Luciferase was p u r i f i e d frm Benerkea h a w e y i ( m a r V i b r i o h w v e y i l by a procPduie $ ? m i l a ? t o t h a t of %unsalur-Mig-n¶7r. T h e m i F Z T E j % e # d l I tOred 31 described (Herghen e t 11.. 19821. HLPE , NAO. FW and BSA e r e
O f >95% upon gar chromatographic analysis, were Supplied by Chemramppco. (Albany supplied by Slgma. Long chain aldehydes. . lcoholr. and aceta te es te rs w i th d purity
I n t e r n i t l O n a l l Colunbur. Ohio. Tr l t ium labe l led te t radecano l was prepared by L iA lH+ reduc t lo " O f [9,10-'n]-tetradecanoic acid (New En land Nuclear1 i n dry e ther and p u r i f i e d by t h l n layer chromatography (TLC1. C'H?-Tetradecanyl acetate, prepared by reac t ion Of [ )HI tetradecanol wi th an e x c e l l Of dCetlC anhydride * I S 2150 TLC p u r l f i e d p r i o r t o use. A l l nc was c a r r i e d cut an naacheray-Nagel SLL N-I(R/UV2 I p la te r . heat ac t i va ted fo r l h a t 1ZD'C and then developed I n herane:ether:ace&c x l d 190:10:21. Thin layer chrOmt0gram *em Prepared for fluOrOgrdphy by spraying l l g h t l y w i t h Enhance (HENI; the procedure I d s a5 given by the manufacturer. DEE Sepharore CL 6B fov anion exchange chrmatography war c4tained from Pharmacia.
100 pupae from the Forest Pest M n a g m n t InPtltUtP, SaUlt Ste Marie. Ontario and a l l owed to emrge under natura l l ight ing. Ext racts 4 r e pwpared by exc is ion o f the glands from 2- t o 3-day o l d female nothr and hmogen iza t lon of the glands for 2 min a t 5W rm w i t h a mtw d r i v e n p e s t l e i n 0.05 M phosphate buffer pH 7.0110 glands per m l l . The so lu t ions were C l a r i f l e d by a 10 m r n cen t r i f uga t io ; a t 13000 19 . Prote in con ten t was I p p m x i m t e l y 0.2 mg per a l . I n t h e 1SSayys desc r ibed be ln , on l y
e x t r a c t f r a L 0.1 gland. Phosphate bu f fe rs were prepared by d i l u t l n g a p p r o p r i a t e 5 to LO v l o f t h e e x t r a c t per nil of assay m i r t u r e was analyzed corresponding t o
dnountl O f 1.0 M NaHIPOh and 1.0 M K 2 H P O I .
FelMle e ls te rn budworm (Chor i l toneura fua i fe rand l 4111 received as batche l O f
prepared I n hexane frm the neat l i qu id . The long chaln Compaundr (genera l l y 5 m o l ) Stock w l u t i o n s o f aldehydes, alcohols and long chain acetate esters RW
I n I D VI Of hexane were t r a n s f e r r e d i n t o aqueous So lu t ion by lenaval a f the hexane under a c o n t r o l l e d f l e w o f N f o r 2 3 n before incubat ion 15-10 min) in bu f fe r
cmpoundr are t r d n l f e r r e d d i r e c t l y i n t o aqueous m d l d by using d solvent misc ib le (genera l l y 3.5 m11. Although2 s imi la r Tesu l ts can be obtained i f the long chain
r l t h water 1e.g. 1 r a p r o p a n o l . d i r e t h y 1 f o m n i d e I , care r u s t be taken i n these carer t h a t the amunt Of s o l v e n t w e d 7 1 low enough so 1s not to i n t e r f e r e i n the d i f f e r e n t enzymatic assays 1e.g. 0.1% i s o p r o p a n o l i n h i b i t s a l c o h o l o r i d a r e a c t r r i t y l .
regression ana1ysir
T I M E immn I TIME i m m )
TABLE IS
Tissue A c t i v i t y " ( p m l / m n l
Minus Gland Plus Gland
Head 0.1 f 0.2 3.2 * 0.4
Abdonpn 0.2 * 0.3 3.8 + 0.6
Leg 0.0 t 0.2 3 .6 * 0.6
Thorax 0.3 f 0.2 3.2 t 0.8
Gland 2.6 f 0.8 6.0 * 0.6
EXTRACT l,~g/mll
I I
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from
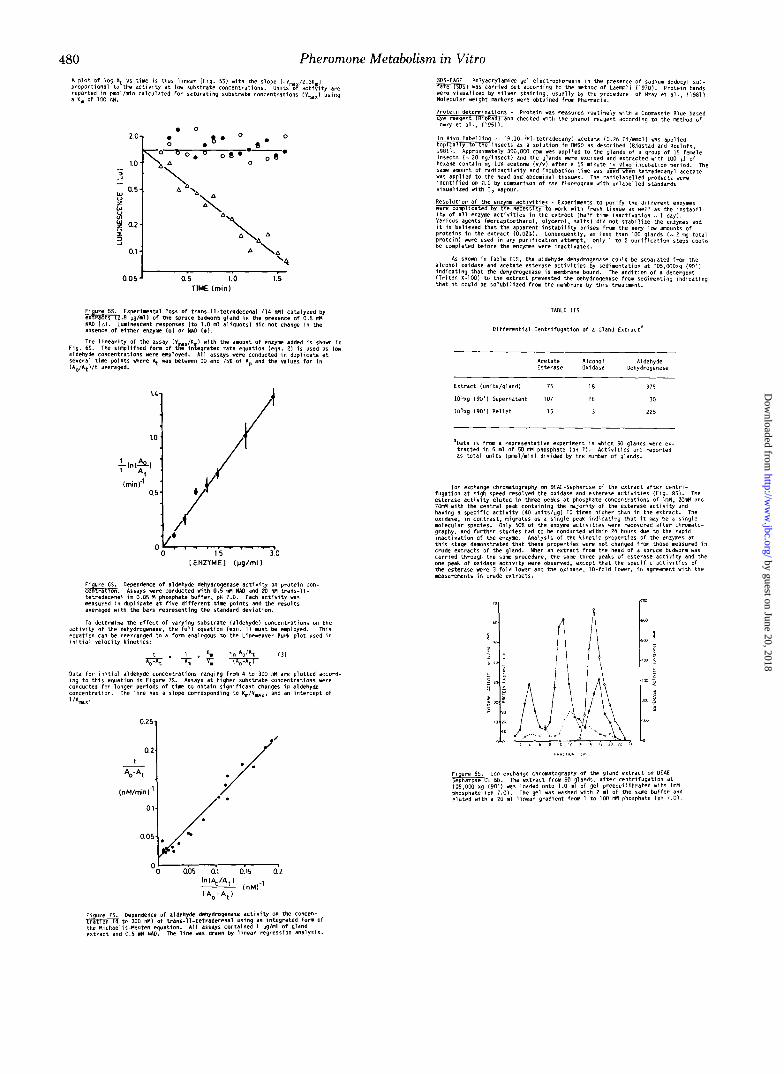
480 Pheromone Metabolism in Vitro R p l o t O f log & V I t i n e i s thus l i n e a r I f l g . 581 with the s lope 1 - V m 12.31: 1 proportional t o t h e a c t i v i t y a t low s u b s t m e c o n c e n t r a t l o n r . U n i t s of a c t i 7 i t y w e reported i n pmol/min ca l cu la ted f o r I a tu rd t l ng I ubs tmte concen t ra t i ons iYmil " r i n g I X. of 1 0 0 nM.
D O
'4 0.05 0.5 1.0 1.5
TIME lmin)
iAo/Rtl / t averaged. "
Dependence O f aldehyde dehydrogenase a c t i v i t y on p r o t e i n con- cen ra Assays *ere Conducted w i t h 0.5 M NAD and Xl M t w n l - 1 1 -
measured ~n d u p l i c a t e a t f i v e d i f f e r e n t t i m e p o i n t s and the resu l t s te t iadecena l i n 0.05 M phosphate buffer, pH 7.0. Each a c t i v i t y was
averaged w i t h the bars represent ing the standard deviat ion.
TO determine the effect of vary ing subst rate la ldehydel concentrat ions on the a c t i v i t y o f t h e dehydrogenase. the f u l l equa t ion leqn. 11 Mlt be employed. This equation can be rear ranged to a form analogous t o t h e l i n e e a v e r - B u r 4 p l o t used i n I n i t i a l v e l o c 7 t y k i n e t i c s :
t 1 h I n 'oint 131 T x T + K T=w
0.251
0.2. t -
%-At ,
(nM/minl"
alcohol oxidase and aceta te es te rase ac t i v r t ie r by red lmentat ion a t 105 WOxg ( 9 0 ' 1 i n d i c a t i n g t h a t t h e dehydrogenase i s membrane bound. The add i t l on o f a ' de te rgen t ( T r i t o n X-100) fa the extract prevented the dehydrogenase from s e d i m n t i n g i n d i c a t i n g t h a t It could be IOIUb l l i2ed frm the membrane by th ls t rea tment .
AS shorn i n Table 118, the aldehyde dehydrogenase could be separated f m m the
TABLE I18
D i f f e r e n t i a l Centrifugation Of d Gland E x t r a c t a
Acetate Alcohol Aldehyde Esterase Oxidare Dehydrogenase
375
30
225
Ion exchange Chromatography on DEAL-Sephdfole o f t h e e x t r a c t a f t e r C e n t r l - fugat ion a t h igh speed resolved the oxldare and e s t e r a s e d c t i w t l e r (Fig. 881. The esterale a c t i v i t y e l u t e d i n t h r e e peak$ a t phosphate COnEentTdtlanL Of lM, 20nH and 7 6 M with the Centra l peak con ta in ing t he ma jo r i t y o f t he es te rase ac t i v i t y and having a s p e c i f i c a c t i v i t y 140 UnitS/ygl 10 t i m s h i g h e r than i n t h e e x t r a c t . The oxidase, I n Contrast. migrates as a 11ngle peak I n d i c a t i n g t h a t It m y be a I l n g l e m l e c u l a r species. Only 50% of the enzyme a c t l v i t i e r were recovered after chromato- graphy. and further studies had TO be Conducted w i t h l n 20 hours due t O t h e m p i d l n a c t i r a t i a n Of the enzyme. Analysis Of t h e k l n e t i c p r o p e r t i e s of t h e e n r y m r a t
Cwde ex t rac ts O f the gland. When an e x t r a c t from the head O f a spruce budworm was t h i s stage denanstrated that these Proper t ies *ere not changed from those malured l n
car r ied th rough the sane procedure, the sdm three peaks of esterare a c t i n t y and the one peak o f Oxidase a c t i v r t y e r e OQleTved, except tha t the rpecr f i c activities of the esterase were 3- fo ld lower and the Oxidase. IO- fo ld l o re r , I " agreemnt wi th the maru renen t r i n c rude ex t rac ts .
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from

D Morse and E MeighenDetection of pheromone biosynthetic and degradative enzymes in vitro.
1984, 259:475-480.J. Biol. Chem.
http://www.jbc.org/content/259/1/475Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/259/1/475.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 20, 2018http://w
ww
.jbc.org/D
ownloaded from













![[ pheromone ] 01](https://img.pdfslide.net/doc/110x75/568caab71a28ab186da2ad9b/-pheromone-01.jpg)















