Embed Size (px)
Citation preview

Requirements for Pseudomonas aeruginosa Type I-F CRISPR-Cas Adaptation Determined
Using a Biofilm Enrichment Assay
1 Gary E. Heussler, Jon L. Miller, Courtney E. Price, Alan J. Collins, George A. O’Toole* 2
Running Title: Type I-F CRISPR-Cas Adaptation 3 4 5 *Address correspondence to George A. O’Toole, [email protected] 6 Department of Microbiology and Immunology 7 Geisel School of Medicine at Dartmouth 8 Rm 202 Remsen Building, Hanover, NH 03755 9 E-mail: [email protected] 10 Tel: (603) 650-1248 11 Fax: (603) 650-1728 12 13 Key words: Pseudomonas aeruginosa, CRISPR, Type I-F, adaptation, spacer 14
15 16
17 18
JB Accepted Manuscript Posted Online 29 August 2016J. Bacteriol. doi:10.1128/JB.00458-16Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

ABSTRACT 19 CRISPR/Cas systems are diverse and found in many archaea and bacteria. These systems have 20 been mainly characterized as an adaptive immune system; able to protect against invading 21 mobile genetic elements including viruses. The first step in this protection is acquisition of 22 “spacer” sequences from the invader DNA into the CRISPR array, termed CRISPR adaptation. 23 Progress in understanding the mechanisms and requirements of CRISPR adaptation has largely 24 been accomplished using overexpression of cas genes or plasmid loss assays; little work has 25 focused on endogenous CRISPR-acquired immunity from viral predation. Here, we have 26 developed a new biofilm-based assay system to enrich for P. aeruginosa strains with new spacer 27 acquisition. We use this assay to demonstrate P. aeruginosa rapidly acquires protective spacers 28 against DMS3vir, an engineered lytic variant of the Mu-like bacteriophage DMS3, through 29 primed CRISPR adaptation from spacers present in the native CRISPR2 array. We found that for 30 the P. aeruginosa Type I-F system, the cas1 gene is required for CRISPR adaptation, recG 31 contributes to (but is not required for) primed CRISPR adaptation, recD is dispensable for 32 primed CRISPR adaptation, and finally, the ability of a putative priming spacer to prime can 33 vary considerably depending on the specific sequences of the spacer. 34 35 IMPORTANCE 36 Our understanding of CRISPR adaptation has expanded largely through experiments in Type I 37 CRISPR systems using plasmid loss assays, mutants of E. coli, or cas1-cas2 overexpression 38 systems, but there has been little focus on studying adaptation of endogenous systems protecting 39 against a lytic bacteriophage. Here we describe a biofilm system that allows P. aeruginosa to 40 rapidly gain protective spacers against a lytic bacteriophage. This approach has allowed us to 41
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

probe requirements of CRISPR adaptation in the endogenous Type I-F system of P. aeruginosa. 42 Our data suggest that CRISPR-acquired immunity in a biofilm may be one reason many P. 43 aeruginosa strains maintain a CRISPR-Cas system. 44 45
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

INTRODUCTION 46 CRISPR-Cas (clustered regularly interspaced short palindromic repeats-CRISPR-47
associated proteins) systems are diverse and widespread, and currently classified into six 48 different types (1, 2) found in over 84% and 45% of sequenced archaeal and bacterial genomes, 49 respectively (3). The general function of a CRISPR-Cas system is to provide adaptive immunity 50 against mobile genetic elements (MGEs) including viruses and plasmids (4-6). This CRISPR- 51 mediated immunity is generated through three stages. In the first stage, termed CRISPR 52 adaptation, a small section of invading MGE nucleic acid (usually 30-40 base pairs, termed the 53 protospacer) is incorporated into the CRISPR array as a spacer along with an additional CRISPR 54 repeat. In the second step, CRISPR RNA (crRNA) generation, the CRISPR array is transcribed 55 into long, noncoding RNA and subsequently processed into individual spacers which associate 56 with Cas proteins forming the crRNA ribonucleoprotein complex. The final step is CRISPR 57 interference in which the spacer region of the crRNA binds invading MGEs at a complementary 58 site through Watson-Crick base pairing and recruits a Cas nuclease to degrade the bound target 59 (for a recent review see (7)). 60
Our understanding of CRISPR adaptation has greatly increased in the last several years 61 but still lags behind advances in understanding crRNA generation and CRISPR interference. 62 Most CRISPR adaption research in the Type I system focuses on the E. coli Type I-E system, but 63 since this CRISPR-Cas system is naturally H-NS silenced, CRISPR adaptation studies have 64 relied on either overexpression of Cas1 and Cas2 in the absence of CRISPR interference (8) or 65 using a Δhns strain of E. coli (9). E. coli has also been used to investigate adaptation in the Type 66 I-F system through expression of a heterologous Type I-F system from Pseudomonas aeruginosa 67 in E. coli (10). These studies have been instrumental in advancing our understanding of the 68
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

mechanisms and genetic requirements for adaptation, but the approaches used limit our 69 understanding of the role of the CRISPR-Cas system in its native context. The endogenous 70 CRISPR-Cas system of P. aeruginosa and its importance during challenge with DMS3vir has 71 been studied previously (11, 12) and we aim to expand upon these types of experiments by 72 studying the role of the P. aeruginosa CRISPR-Cas system in biofilm growth. 73
Pseudomonas aeruginosa strain UCBPP-PA14 contains a Type I-F CRISPR-Cas system 74 organized such that the canonical Type 1-F cas genes are flanked by two CRISPR arrays termed 75 CRISPR1 and CRISPR2 containing 13 and 21 spacers, respectively (Fig. 1)(13). Previously our 76 lab has shown this CRISPR-Cas system is active during normal laboratory growth conditions 77 (14) and capable of acquiring protective spacers during challenge with the bacteriophage 78 DMS3vir, a lytic-only variant of the bacteriophage DMS3 (15). However, even though the 79 native CRISPR2 locus contains 3 putative DMS3-targeting priming spacers (spacers that 80 partially match the phage target and increase the likelihood of incorporated new spacers against 81 the phage), after phage infection and isolation of resistant strains, less than one percent of these 82 resistant isolates had acquired new spacers while the vast majority (>99%) had gained resistance 83 through mutation of the type IV pilus (T4P), the known receptor of DMSvir (16) as well as many 84 other Pseudomonas phages (17). Thus, while co-inoculating P. aeruginosa with DMS3vir in a 85 culture tube results in spacer acquisition, this event is too inefficient to be used as a means of 86 investigating natural CRISPR adaptation in P. aeruginosa, necessitating a strategy for increasing 87 the frequency of native spacer acquisition. 88
Given that the T4P is known to play an important role in biofilm formation in P. 89 aeruginosa (18), we hypothesized that cells grown in a biofilm during phage challenge would be 90 less likely to gain resistance to DMS3vir through mutations of the T4P. We show here that a 91
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

biofilm-based enrichment system greatly increases the ability to detect CRISPR adaptation 92 events, allowing us to investigate the efficiency of different priming spacers, protospacer 93 sequences, and the genetic requirements of the P. aeruginosa Type I-F system in the context of 94 viral immunity. Furthermore, CRISPR-Cas systems are widespread in P. aeruginosa strains (19, 95 20), thus a better understanding of the role of the P. aeruginosa Type I-F CRISPR-Cas system in 96 the context of biofilm growth may help explain the prevalence of these systems. 97 98 MATERIALS AND METHODS 99 100 Strains and media. Strains, plasmids, and primers used in this study are listed in Supplementary 101 Table S1. The P. aeruginosa strain UCBPP-PA14 (abbreviated P. aeruginosa PA14) was used in 102 this study. P. aeruginosa and E. coli strains were routinely cultured in lysogeny broth (LB) at 103 37 degrees C. Growth media were supplemented with antibiotics at the following concentrations: 104 gentamicin (Gm), 10 μg mL-1 (E. coli) and 50 μg mL-1 (P. aeruginosa). The ΔrecD, ΔrecG, 105 ΔCRISPR1ΔCRISPR2sp1-19, ΔCRISPR1ΔCRISPR2sp1-19 + sp20-5MM, and CRISPR2-106 minimum strains and deletion plasmids were generated using allelic exchange and 107 Saccharomyces cerevisiae recombineering techniques described previously (21). The recG::TnM 108 strain used here is from the P. aeruginosa PA14 non-redundant library (22) and the transposon 109 insertion was confirmed via PCR. 110 111 Biofilm enrichment assay. The indicated P. aeruginosa strains were grown overnight and 112 standardized to an optical density of 3.0 at 600 nm, and then 100 μL of the culture (~2.5 x 108 113 CFU) was added to 5 ml of M63 minimal medium supplemented with 0.4% arginine and 1 mM 114
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

magnesium sulfate (a biofilm-inducing medium) per well in a 6 well tissue culture dish. The 115 indicated P. aeruginosa strains were co-inoculated with 2.5 x106 PFU of DMS3vir phage in each 116 well (an M.O.I. of 0.01) and grown overnight at 37 degrees C. After incubation, the medium was 117 aspirated and the wells were washed once with 5 ml of phosphate buffered saline (PBS) 118 removing any planktonic bacteria and leaving behind only the biofilm population. The biofilm 119 population was isolated by using a 25 cm Sarstedt™ cell scraper to remove any attached 120 bacterial cells at the air-liquid interface, and the cells subsequently collected in 1mL of PBS. 121 These cells were then washed with 1ml PBS, re-suspended in 50 μl of PBS and the entire 50 μl 122 inoculated into 5 mL LB and grown for either 10 hours (for the slow-growing ΔrecD mutant) or 123 6 hours (all other strains) at 37 degrees C. After the biofilm was allowed to form, the cells were 124 exposed to fresh DMS3vir at concentrations and in conditions exactly as described above. After 125 co-incubation of the biofilm cells and phage, the remaining biofilms cells were collected as 126 described above. Additionally, an aliquot of the planktonic population from the WT P. 127 aeruginosa + DMS3vir condition was collected prior to aspiration. Both planktonic and biofilm 128 cells were serially diluted, plated on LB agar, and incubated at 37 degrees C for 16 hours to yield 129 single colonies. For each condition, at least 200 single colonies were re-patched onto LB agar 130 using a sterile pipette tip and grown for 16 hours at 37 degrees C to assess their twitching 131 phenotype, as describe in the next section. 132 133 Twitch assay. The twitch assay was performed as described previously (23). Briefly, a sample 134 from re-patched isolates not displaying any bacteriophage-mediated lysis (as judged by lack of 135 obvious plaques) was collected using a sterile pipette tip and punctured through LB agar, and 136 deposited onto the hard plastic of the petri plate. The samples were then allowed to grow for 36 137
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

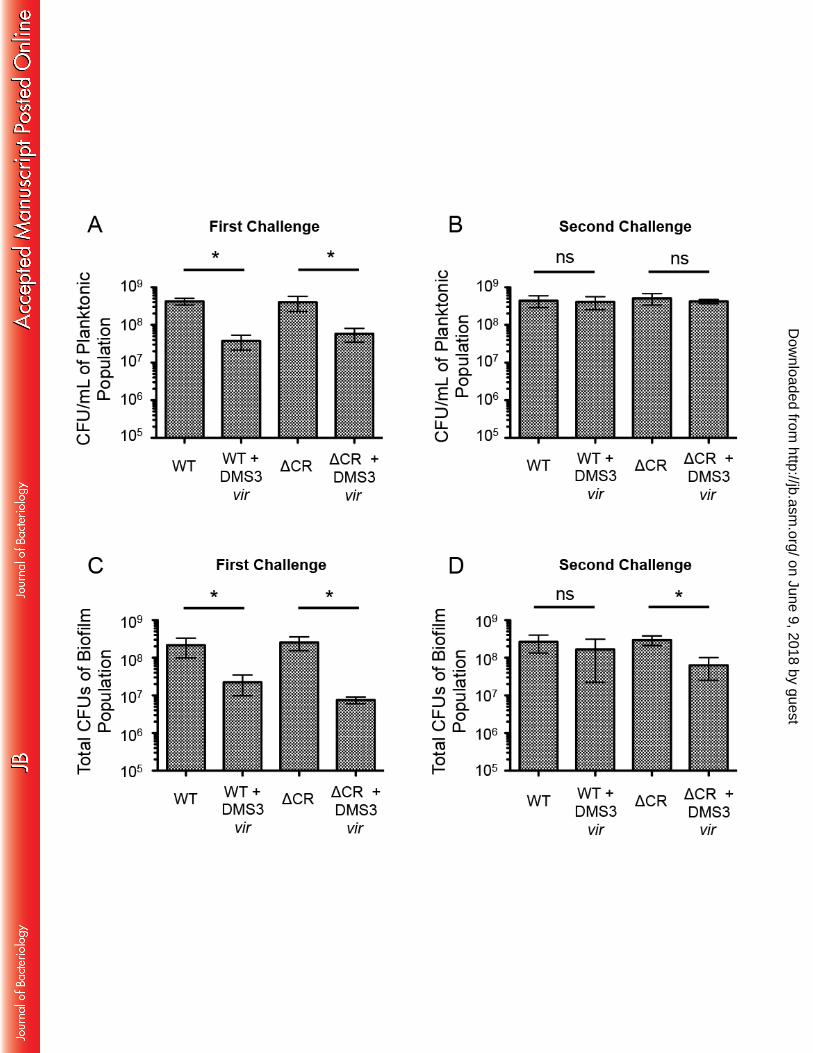
hours at 37 degrees C, at which point the LB agar is carefully peeled away. The bacteria were 138 visualized with the addition of 0.1% crystal violet. Twitch positive strains grow along the plastic 139 away from the inoculation point, while twitch negative cells remain at the inoculation point. 140 141 Detecting newly-acquired spacers. Any re-patched isolates from the biofilm-based, spacer 142 enrichment assay not showing signs of bacteriophage-mediated lysis and displaying a twitch-143 positive phenotype, was used as a template for PCR using primers amplifying the leader end of 144 either the CRISPR1 or CRISPR2 array. For the indicated isolates, one of these same primers was 145 used for Sanger sequencing to characterize the newly acquired spacers. 146 147 Statistical Analysis: The data presented in Figure 3 were analyzed using Graph Pad Prism 5. 148 The data represent the mean standard deviation of three independent experiments with multiple 149 replicates. All data were treated as normally distributed, and comparisons were tested with 150 Student's t-test. 151 152 RESULTS 153 154 P. aeruginosa rapidly gains resistance to the lytic bacteriophage DMS3vir in biofilm 155 inducing conditions 156
The biofilm enrichment system involves two consecutive incubations of P. aeruginosa 157 and DMS3vir in biofilm-inducing conditions. All the bacterial cells used to inoculate the medium 158 for the second biofilm incubation were isolated from the biofilm formed at the end of the first 159 incubation. Because the biofilm cells are exposed to phage DMS3vir, this approach strongly 160
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

enriches for efficient biofilm forming isolates that are resistant to DMS3vir infection. The 161 enrichment procedure is outlined in Figure 2. 162
To assess the impact of phage infection on the WT strain and a strain lacking the 163 CRISPR-Cas region, 2.5x108 CFU of either P. aeruginosa or P. aeruginosa in which the entire 164 CRISPR-Cas region was deleted (ΔCR) was added either alone or with 2.5x106 PFU of DMS3vir 165 to 5mL of biofilm inducing minimal medium in a 6 well plate. After 24 hours static growth at 37 166 degrees C, both the biofilm and planktonic populations were isolated and aliquots from each 167 population were serially diluted and plated to measure viable bacteria (CFU). As illustrated in 168 Figure 3, DMS3vir infection significantly reduced the level of both WT and ΔCR P. aeruginosa 169 in the planktonic population (Fig. 3A) as well as the biofilm population (Fig. 3C) after 24 hours 170 of incubation. The planktonic population of WT and ΔCR cells grown with DMS3vir had on 171 average 91% and 85% fewer cells, respectively, than the planktonic population of WT and ΔCR 172 grown without DMS3vir (Fig. 3A). Similarly, the biofilm population of WT and ΔCR cells 173 grown with DMS3vir had on average 90% and 97% fewer cells, respectively, than the biofilm 174 population grown without DMS3vir (Fig. 3C). 175 The biofilm cells isolated after the first incubation step were subsequently grown 176 planktonically for 6 hours in LB at 37 degrees C to generate enough cells to inoculate the second 177 biofilm assay plates required for our enrichment protocol. After the 6 hrs of planktonic growth, 178 the WT and ΔCR cells were added to 5mL of a biofilm-inducing minimal medium, either with or 179 without fresh DMS3vir, at the same counts used above. After 24 hours growth at 37 degrees C, 180 both the biofilm and planktonic populations were again isolated, and aliquots were serially 181 diluted and plated to measure CFU. In contrast to the results from the first round of incubation 182 (Fig. 3A,C), there was no significant decrease in either WT or ΔCR cells incubated with 183
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

DMS3vir compared to cells grown in the absence of the bacteriophage in the planktonic 184 population (Fig. 3B), indicating that both the WT and ΔCR planktonic cells had become resistant 185 to DMS3vir-mediated lysis. Interestingly while the biofilm population of WT cells grown with 186 DMS3vir was not significantly different than the biofilm population of the WT cells grown 187 alone, the biofilm population of ΔCR cells grown with DMS vir had on average 79% fewer cells 188 than the population of ΔCR cells grown alone, a significant (P<0.05) decrease (Fig. 3D). It 189 should be noted that while the difference between the WT biofilm and WT + DMS3vir biofilm 190 population was not significant, there were still on average 40% fewer cells in the WT + DMS3vir 191 population, and the variability of the assay may partially contribute to the lack of significance as 192 measured by Student’s t-test. Nevertheless, these data indicate that both WT and ΔCR P. 193 aeruginosa can rapidly gain resistance to DMS3vir during biofilm growth, and that the P. 194 aeruginosa cells with a functional CRISPR-Cas system had a slight but reproducible advantage 195 over CRISPR deficient P. aeruginosa cell when growing in a biofilm in the presence of a lytic 196 phage. 197 198 Pseudomonas aeruginosa incorporates protective spacers against the lytic bacteriophage 199 DMS3vir in biofilm inducing conditions 200 The biofilm-specific increase of WT versus ΔCR P. aeruginosa cells when grown in the 201 presence of DMS3vir suggested that the WT cells were gaining resistance to DMS3vir through 202 spacer acquisition in addition to, or perhaps in preference to, T4P mutations, especially given 203 that T4P mutations negatively impact the ability of P. aeruginosa to form a biofilm. To test this 204 idea, after isolation and serial dilution plating of the biofilm population from the second biofilm 205 incubation step (see Fig. 2), at least 200 colonies for each condition were re-patched onto LB 206
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

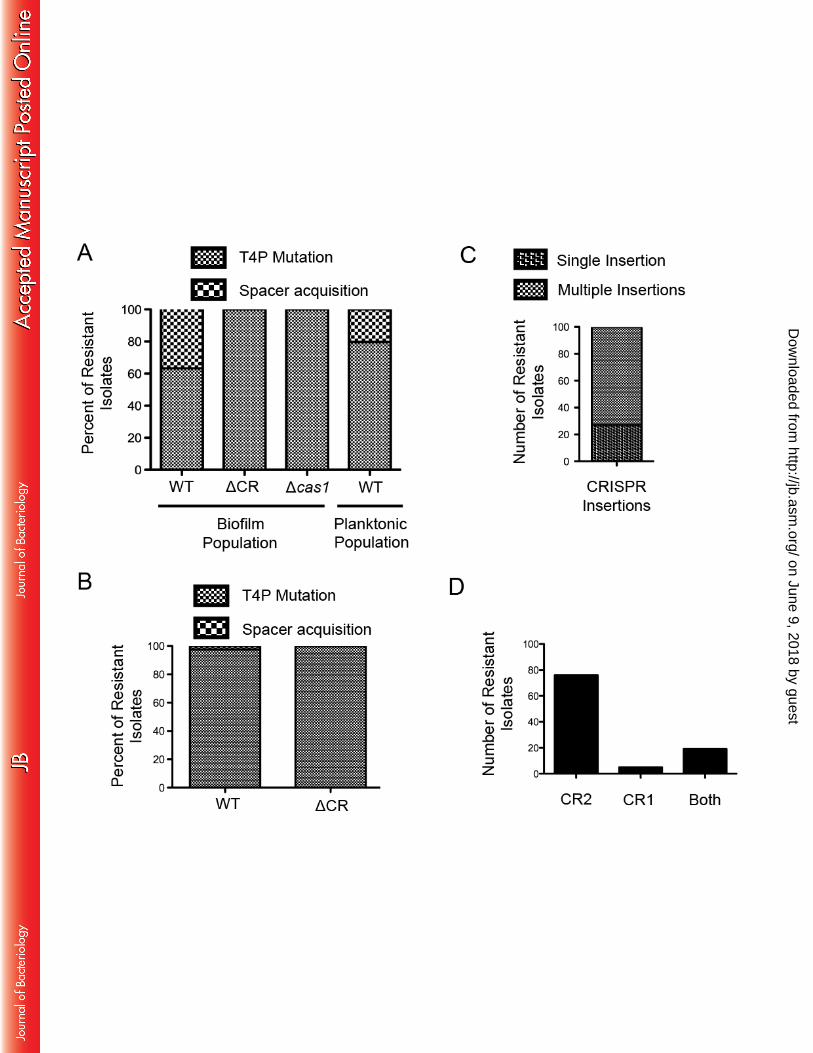
agar in three separate assays. Not all cells in the biofilm gained resistance to DMS3vir, and to 207 eliminate non-resistant cells any isolate that showed signs of lysis from DMS3vir particles that 208 had been retained from the infection step were eliminated from further analysis (Fig. 2, compare 209 NR versus R patches). All the remaining isolates were then assayed for twitching motility as 210 described previously (24) and shown in Figure 2. Isolates that had gained DMS3vir resistance 211 through a T4P mutation were easily distinguishable due to their lack of twitching, and any isolate 212 that was twitch-positive was presumed to have acquired a new spacer that blocked infection and 213 thus was screened for spacer acquisition using primers that amplify the leader-proximal end of 214 both CRISPR1 and CRISPR2. 215 Unlike the low rate (>1%) of spacer acquisition previously observed during P. 216 aeruginosa and DMS3vir co-incubation (15), on average 37.7% of the DMS3 resistant isolates of 217 the WT strain had acquired new spacers while the remaining 62.3% had gained resistance 218 through T4P mutation (Fig. 4A). The ΔCR biofilm resistant population was 100% twitch-219 negative in these experiments suggesting no other means of resistance was acquired besides a 220 mutation in the T4P apparatus. 221 222 Cas1 is required for new spacer acquisition 223
Cas1 is universally conserved across CRISPR-Cas systems and has been shown to be 224 required for CRISPR adaptation (8, 25), including for a heterologous P. aeruginosa Type I-F 225 system over-expressed in E. coli (10) and for the endogenous Type I-F system of Pectobacterium 226 atrosepticum during CRISPR adaptation against plasmids (26). To test the requirement of 227 endogenous Cas1 in CRISPR adaptation by P. aeruginosa versus a lytic phage, a Δcas1 mutant 228 was assayed for spacer acquisition using the biofilm enrichment system. Similar to the CRISPR-229
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

deficient ΔCR mutant, all of the resistant isolates after the second biofilm incubation had lost 230 T4P functionality (Fig. 4A), confirming that the cas1 gene is required for CRISPR adaptation by 231 P. aeruginosa challenged with lytic bacteriophage. 232 233 Biofilm growth enhances spacer acquisition 234
The underlying premise of our assay is that growth in a biofilm should enhance the 235 frequency of spacer acquisition by selection for the sub-population of cells with functional T4P, 236 a cell appendage required for robust biofilm formation (18). It has been shown previously that P. 237 aeruginosa will more likely gain resistance to DMS3vir through CRISPR adaptation versus 238 mutations in genes encoding for T4P during co-incubation in a minimal medium (11). Therefore 239 to determine if the CRISPR adaptation frequency of ~38% observed in WT P. aeruginosa is 240 specifically the result of biofilm growth and not simply growth in a minimal medium (previous 241 assays were performed in rich LB medium), the planktonic population of WT P. aeruginosa 242 incubated with DMS3vir isolated after the second incubation step was assayed for CRISPR 243 adaptation in the same manner as the biofilm population in three separate experiments. Of the 244 resistant planktonic cells, on average 20.3% had gained resistance through CRISPR adaptation 245 with the remaining 79.7% gaining resistance through T4P mutations (Fig. 4A), significantly less 246 (P=0.01) than the CRISPR adaptation frequency (37.7%) observed in the resistant biofilm 247 population (see Supplementary Table 2 for raw data used in the statistical analysis). 248
Since the level of spacer acquisition in the planktonic population was higher than what 249 was previously observed after co-inoculating P. aeruginosa with DMS3vir in LB medium 250 overnight at 37 degrees with shaking (15), the culture tube-based P. aeruginosa and DMS3vir 251 co-incubation was repeated using WT and ΔCR P. aeruginosa in LB medium to confirm the low 252
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

adaptation frequency observed previously. Of the >300 resistant isolates, 2.3% of WT P. 253 aeruginosa and 0% of ΔCR had gained resistance through CRISPR adaptation with the 254 remaining isolates gaining resistance through T4P mutation (Fig. 4B), confirming the low 255 frequency of spacer acquisition in rich medium for planktonically growing bacteria. 256
Taken together, these data suggest that P. aeruginosa grown planktonically and 257 challenged with DMS3vir acquire resistance to DMS3vir via CRISPR adaptation at a much 258 higher rate (20% versus 2%) when grown in a minimal medium with static growth compared to a 259 rich medium with shaking. Furthermore, the highest rate of CRISPR adaptation to DMS3vir 260 (38%) was observed in the biofilm population of cells grown in a minimal medium, supporting 261 our strategy for increasing the rate of spacer acquisition in P. aeruginosa by enriching for cells 262 with a functional T4P during challenge with DMS3vir. 263 264 Multiple spacers are preferentially incorporated with a biased incorporation into the 265 CRISPR2 array 266
Of the P. aeruginosa isolates that incorporated new spacers providing resistance to 267 DMS3vir, 100 were selected randomly to assay the specific number of spacers acquired and to 268 determine into which CRISPR array the spacers were inserted. A strong bias is observed for 269 multiple CRISPR insertions in our system, with only 27 out of 100 isolates inserting a single 270 spacer (Fig. 4C). These data are similar to observations in the Type I-F system of P. 271 atrosepticum; during CRISPR adaptation against a transformed plasmid only 24% (9/37) of 272 plasmid-insensitive strains inserted a single spacer (27) suggesting similarities in CRISPR 273 adaptation against a plasmid and versus a lytic bacteriophage in the Type I-F system. Multiple 274
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

incorporated spacers are likely important since it has been shown in P. aeruginosa that spacer 275 diversity against DMS3vir is beneficial to the survival of the bacteria (12). 276
Additionally, of the 100 resistant isolates, 76 inserted new spacers into only CRISPR2, 5 277 inserted new spacers into only CRISPR1, and 19 inserted new spacers into both CRISPR1 and 278 CRISPR2 (Fig. 4D), demonstrating that CRISPR2 and CRISPR1 are both functional in CRISPR 279 interference, although there appears to be a bias for inserting new spacers into CRISPR2 in these 280 experiments similar to what had been observed previously in P. aeruginosa (11). These data are 281 also similar to observations in the Type I-F system of P. atrosepticum; of 105 new spacers 282 inserted into three CRISPR arrays in P. atrosepticum, 65%, 32%, and <3% were found in 283 CRISPR1, CRISPR2, and CRISPR3 respectively (27) further suggesting common themes of the 284 Type I-F system regardless of target (plasmid versus lytic bacteriophage) or host (P. 285 atrosepticum versus P. aeruginosa). 286 287 Spacers incorporated using the biofilm enrichment system are the result of primed 288 CRISPR adaptation 289
Recent studies of the Type I-E CRISPR-Cas system of E. coli separate Type I CRISPR 290 adaptation into two distinct types. In the first type, naïve adaptation, new spacers are 291 incorporated into a CRISPR array in the absence of CRISPR interference components. This 292 naïve adaptation was demonstrated via the overexpression of the cas1 and cas2 genes (8, 28). In 293 the second type of adaptation, termed primed adaptation, a nucleic acid target is bound by a 294 crRNA ribonucleoprotein complex that can not engage in CRISPR interference due to 295 mismatches between the protospacer and spacer or a non consensus protospacer adjacent motif 296
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

(PAM), but the presence of this mismatched spacer results in rapid and efficient spacer 297 acquisition from regions proximal to the priming protospacer (25, 29). 298
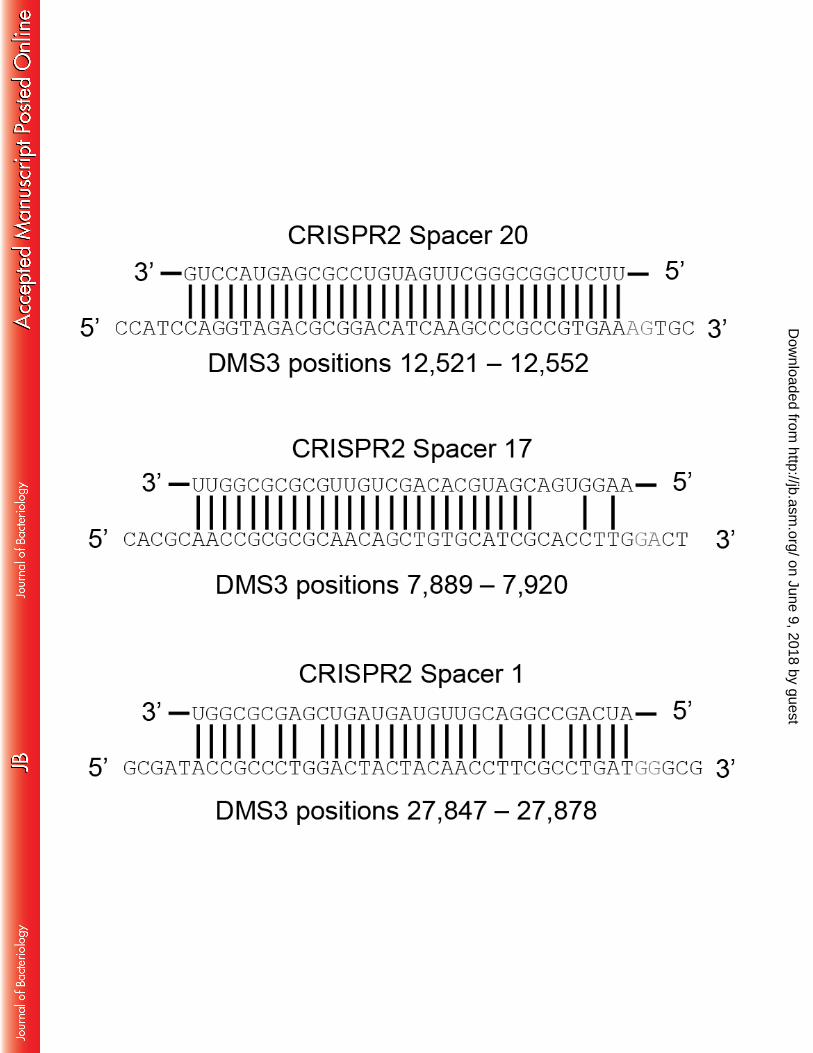
P. aeruginosa contains three spacers in the native CRISPR2 array that can potentially 299 participate in primed adaptation against the bacteriophage DMS3, as illustrated in Figure 5A. 300 The first, CRISPR2 Spacer 20 (CR2_sp20), is 100% complementary to DMS3 but cannot 301 interfere due to a non-consensus PAM (AG instead of GG). The second, CR2_sp17, is partially 302 complementary to DMS3 with 5 mismatches including 4 in the seed region, in addition to 303 targeting a protospacer with a non-consensus PAM (GA instead of GG). It should be noted the 304 mismatches between CR2_sp17 and DMS3 likely exist because of a single nucleotide deletion 305 on the phage genome; if a thymine is added where the identity between the spacer and target end, 306 CR2_sp17 would be a 100% match with the correct PAM. The third, CR2_sp1, is partially 307 complementary to DMS3 with 5 mismatches but the target has a consensus PAM. Notably, 308 binding of CR2_sp1 to the DMS3 protospacer inserted on the P. aeruginosa chromosome is 309 responsible for the CRISPR-dependent modification of biofilm formation reported previously by 310 our group (10). 311
To assay if the CRISPR adaptation observed in the biofilm enrichment system is the 312 result of primed adaptation or naïve adaptation, a mutant was generated in which the CRISPR1 313 array was deleted and all spacers in the CRISPR2 array were deleted, leaving only a single repeat 314 and the leader sequence (Fig. S1) since a single repeat and leader sequence has been shown to be 315 sufficient for naïve adaptation in the E. coli Type I-E system (8). Therefore any newly acquired 316 spacer in this mutant background would be the result of naïve adaptation. This construct, termed 317 CRISPR-minimum, was then analyzed via the biofilm enrichment assay. To increase the 318 likelihood of detecting an acquired spacer, the assay was modified slightly in that rather than re-319
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

patch the colonies (step 6 in Fig. 2) the bacteria were plated on freshly poured LB agar (therefore 320 with a higher moisture level than typical plates) so that any twitching-positive colony develops a 321 distinct ‘rough’ appearance, as previously reported (15), and can therefore be easily identified. 322 While this modification of the assay prevents the determination of rates of T4P mutations in 323 resistant isolates since DMS3vir-resistant isolates are indistinguishable from non-DMS3vir-324 resistant isolates and not every twitch positive colony will display the ‘rough’ appearance, these 325 modifications allow rapid screening of a large number of isolates for putative spacer acquisition 326 events. Over 10,000 colonies were screened and 768 ‘rough’ colonies were assayed for spacer 327 acquisition by PCR with no new spacers detected. Therefore since no naïve adaptation was 328 detected, our data suggest primed adaptation is the predominant mode of spacer acquisition in 329 the P. aeruginosa Type I-F CRISPR-Cas system. 330
To determine which of the three putative priming spacers illustrated in Figure 5 331 contribute to primed adaptation, the CRISPR arrays of 35 of the CRISPR-positive resistant 332 bacteria isolated from the biofilm enrichment system were sequenced resulting in the 333 characterization of 87 new spacers (Table S3). Of the 87 spacers examined, the vast majority 334 (84/87) are likely primed by CR2_sp1 based on their distribution around the CR2_sp1 DMS3 335 target (Table S3). This finding is similar to previous work in which the vast majority of newly 336 acquired spacers in resistant P. aeruginosa after co-incubation with DMS3vir were likely primed 337 by CR2_sp1 (11). Previously, when the P. aeruginosa Type I-F CRISPR-Cas system was 338 overexpressed in E. coli, newly acquired primed spacers targeting regions up to 5000 bp from the 339 location of the priming protospacer was detected (10). In our system, putative primed spacers 340 targeting regions up to 6000 bp away from the CR2_sp1 target were detected, suggesting the 341 long range of Type I-F CRISPR priming observed previously in E. coli is not due an artifact of 342
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

heterologous expression. Additionally, of these 84 CR2_sp1-primed spacers, only 4 targeted a 343 region of DMS3 with a non-consensus PAM (GC, AG, TG and GC in spacer number 11, 27, 33, 344 and 69 respectively) with three of these spacers (spacer 11, 27, and 33) targeting a region of 345 DMS3 with a consensus PAM shifted one nucleotide away (i.e. GG in the +1/-1 or -2/-3 position 346 instead of the canonical -1/-2 position) an occurrence observed in other Type-I CRISPR systems 347 and termed “PAM slippage” (27, 30). 348
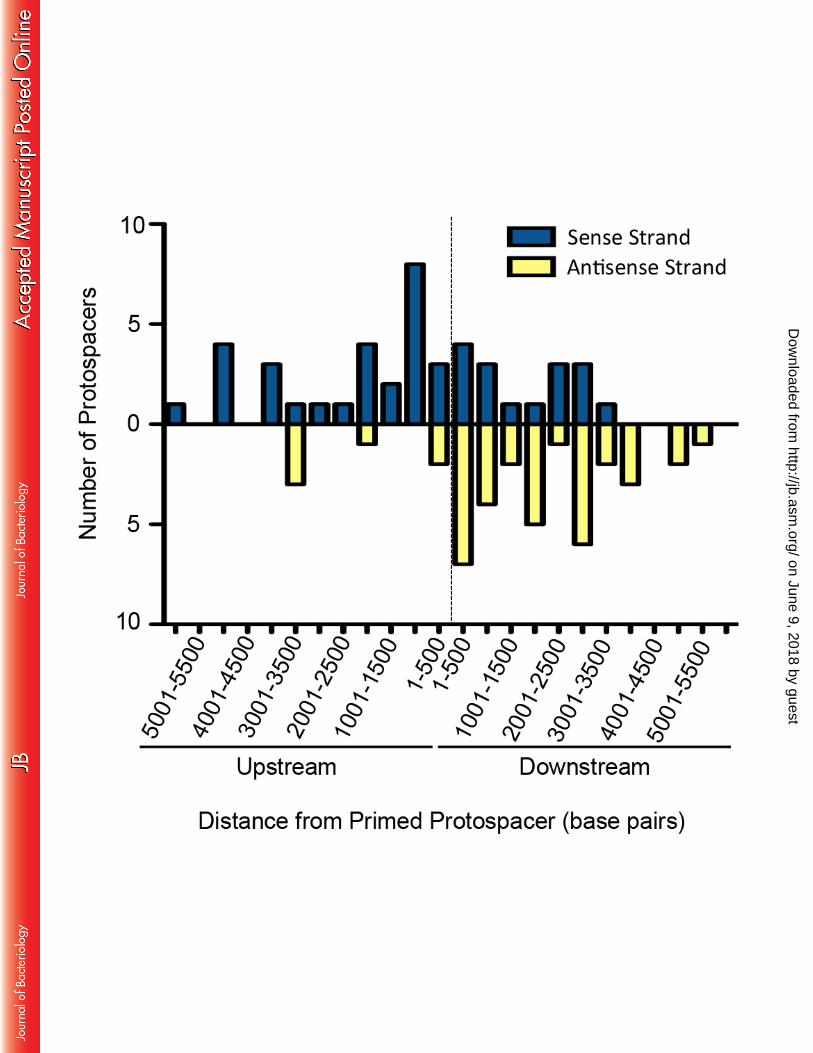
The remaining 3 spacers we identified from this collection of 87 spacers are potentially 349 primed by spacer CR2_sp20. The first two of these spacers target DMS3 within 250 base pairs of 350 the CR2_sp20 target and the third target DMS3 within 3000 base pairs (Table S3). All three of 351 these putative CR2_sp20-primed spacers were found in the same isolate, strongly suggesting that 352 spacer incorporation likely occurred only once in these resistant isolates, with the number of 353 insertions dependent on how many new spacers were acquired during a single priming event, in 354 line with the model proposed in the Type I-F system of P. atrosepticum (27). 355 356 Spacer acquisition in the biofilm enrichment assay occurs in a biased manner. A key 357 difference between Type I-E and Type I-F primed spacer acquisition is the observed biases 358 during protospacer selection. It was shown in the Type I-E CRISPR-Cas system of E. coli that 359 during CRISPR priming, new spacers are incorporated that target the same strand as the priming 360 spacer (9, 25). Conversely, in Type I-F CRISPR priming it was demonstrated that acquired 361 spacers show no bias in respect to the targeted versus non targeted strand but instead new spacers 362 preferentially target sites 5’ of the priming spacer target, either on the targeted or the non-363 targeted strand (10, 11, 27); this same bias has also been reported in the Type I-B system (31). 364
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

Of the 83 CR2_sp1 putatively primed spacers that target DMS3 within 6000 base pairs of 365 the CR2_sp1 target, protospacer selection was similar to other Type I-F systems. There was 366 minimal bias between the target and non-target strand with 53% of new spacers targeting the 367 same strand as CR2_sp1 versus 46% targeting the opposite strand (Figure 6). However, there is 368 an observed bias between protospacers selected in the 3’ or 5’ direction relative to the CR2-sp1 369 target with 73% of new spacers targeting a protospacer present in the 5’ direction relative to the 370 CR2_sp1 target and only 27% targeting a protospacer in the 3’ direction of the CR2_sp1 target 371 (Figure 6). These data are similar to what has been previously shown with endogenous Type I-F 372 CRISPR adaptation against a lytic bacteriophage (11), and confirms that the protospacer 373 selection biases previously observed reflect a general property of the Type I-F system. 374 375 RecD is dispensable for primed acquisition in the P aeruginosa Type I-F CRISPR-Cas 376 system while RecG contributes to efficient priming 377
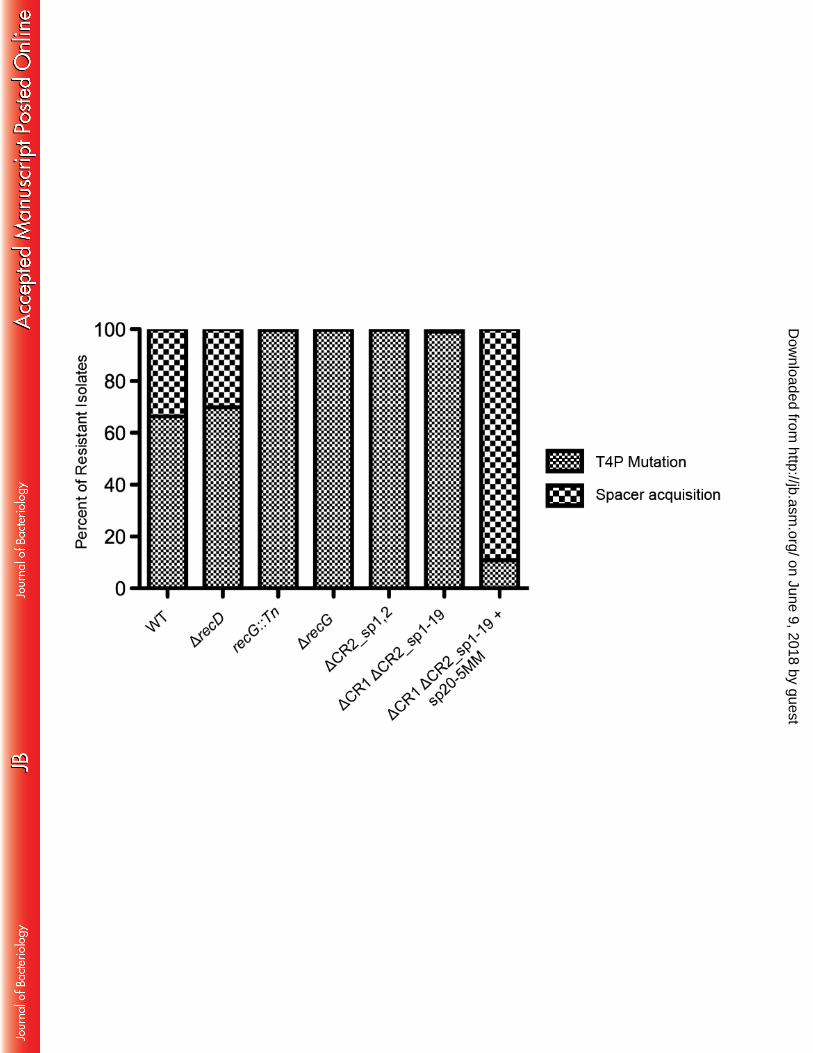
Recent work has demonstrated involvement of both the RecBCD complex and RecG 378 protein in naïve and primed CRISPR adaptation, respectively (28, 32). The RecBCD complex is 379 required for naïve adaptation in the Type I-E system of E. coli, and likely contributes to 380 adaptation through single stranded DNA generation at double stranded breaks, such as those 381 encountered during replication, thereby providing a substrate for Cas1 (28). Regarding primed 382 adaptation, in the E. coli Type I-E system RecG and PriA were shown to be required for primed 383 adaptation presumably through R-loop removal after crRNA binding, allowing the Cas1-Cas2 384 complex access to the single stranded DNA generated by Cas3 (32). We tested if these proteins 385 were playing a similar role in the Type I-F system of P. aeruginosa using the biofilm enrichment 386 system and hypothesized that since all spacer acquisition in our system is likely primed, deletion 387
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

of the recD gene would not impact spacer acquisition while deletion of the recG gene would 388 impact new spacer insertion. 389 A deletion of the recD gene was generated in P. aeruginosa and this mutant strain was 390 incubated with DMS3vir in the biofilm enrichment scheme outlined in Figure 2. Of note, during 391 the LB growth step between the two biofilm incubations, the cells carrying the recD mutation 392 were incubated for 10 hours instead of 6 due to a slight growth defect caused by the deletion of 393 the recD gene, otherwise the biofilm enrichment proceeded was performed as described above. 394 At least 200 colonies were re-patched after isolation of the biofilm population from the second 395 incubation, and of the isolates that had gained resistance to DMS3vir, 30.1% had gained 396 resistance through spacer acquisition while the remaining 69.9% were T4P mutants (Fig. 7A), 397 similar to the spacer acquisition frequency observed for the WT population described above 398 (Figure 6). To test if the acquired spacers were primed, the CRISPR arrays of 7 ΔrecD isolates 399 with newly acquired spacers were sequenced, resulting in the characterization of 14 newly 400 acquired spacers. As listed in Table S4, CR2_sp1 likely primed all 14 newly acquired spacers, 401 supporting the hypothesis that RecD, and presumably the RecBCD complex, is dispensable for 402 primed adaption in the Type I-F CRISPR-Cas system of P. aeruginosa. 403 To test the requirement of RecG in primed CRISPR adaptation, a P. aeruginosa strain 404 with a transposon stably inserted into the recG gene was assayed in the biofilm enrichment assay 405 at the same counts of bacteria and bacteriophages as described above (see Figure 2). Of the 406 resistant mutants isolated from the biofilm population after the second incubation, 99.6% were 407 T4P mutants while 0.4% (two isolates) had gained resistance through CRISPR adaptation (Fig. 408 7). The CRISPR arrays of these two isolates were sequenced, and the 4 newly acquired spacers 409 were likely primed by CR2_sp1 (Table S4). To confirm that these data were not due to any polar 410
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

effect of the transposon in recG, an in-frame deletion of recG was generated and assayed in the 411 biofilm enrichment assay at the same counts of bacteria and bacteriophages as described above. 412 Of 479 resistant isolates, 99.6% were T4P mutants with only two isolates (0.4%) gaining 413 resistance through spacer acquisition, confirming that while RecG is necessary for efficient 414 CRISPR adaptation, priming can still occur in a recG mutant strain, albeit at a greatly reduced 415 rate. 416 417 Spacers incorporated using the biofilm enrichment are biased towards CR2_sp1 priming 418 over CR2_sp20 priming due to the specific location of mismatches between the spacer and 419 the target. One observation made using the biofilm enrichment assay is that almost all of the 420 newly incorporated sequenced spacers were apparently primed by CR2_sp1. The CRISPR2 array 421 of isolate #33 listed in Table S3 contains three newly acquired spacers which were likely primed 422 by CR2_sp20 due to the proximity of the spacer targets on DMS3 to the target of CR2_sp20. 423 This is the only example using the biofilm enrichment assay of a spacer other than CR2_sp1 424 likely mediating priming, and we hypothesized that deleting CR2_sp1 would enrich for 425 alternatively primed spacer acquisitions. A strain in which the first two spacers of CRISPR2 426 were deleted (ΔCR2_sp1,2) was co-inoculated with DMS3vir in the biofilm enrichment assay 427 under the same counts of bacteria and bacteriophage described above. Interestingly, CRISPR 428 adaptation was greatly reduced without CR2_sp1, with only a single isolate (0.3%) of the 429 resistant biofilm population gaining resistance through spacer acquisition (Fig. 7). This isolate 430 was sequenced and the resulting single acquired spacer appeared to be primed by CR2_sp20 431 since target of the new spacer was only 1.2 kb upstream of the CR2_sp20 DMS3 target. These 432
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

data suggest that a large difference in priming efficiency can exist between two priming spacers 433 in an endogenous Type I-F CRISPR-Cas system. 434
One possibility for the reduced rate of spacer acquisition observed with the ΔCR2_sp1,2 435 strain is the distance between CR2_sp20 and the leader sequence of CRISPR2 since it has been 436 observed in P. furiosus that crRNA from spacers closest to the leader accumulate to a higher 437 level that crRNA from spacers farther from the leader sequence (33). To test if the lower ability 438 of CR2_sp20 to prime is related to the distance of CR2_sp20 to the CRISPR2 leader sequence, a 439 strain was generated in which the entire CRISPR1 array and CRISPR2 spacers 1-19 were deleted 440 (ΔCR1 ΔCR2_sp1-19, Fig. S1) resulting in CR2_sp20 being the closest spacer to the CRISPR2 441 leader sequence. To confirm CR2_sp20 is functional in this strain, a mutant of DMS3vir was 442 generated in which the CR2_sp20 target contains the correct PAM (GG) instead of the non-443 functional PAM (AG) normally present in wild type DMS3. This A to G mutation is 444 synonymous and therefore unlikely to affect the ability of the phage to infect P. aeruginosa. This 445 mutant phage, referred to as DMS3vir-PC (in reference to PAM corrected), should therefore not 446 be able to infect the ΔCR1 ΔCR2_sp1-19 strain (as measured by plaque assay) if CR2_sp20 is 447 functional due to CRISPR interference. As illustrated in Figure S2, DMS3 vir-PC can infect 448 CRISPR deficient P. aeruginosa (ΔCR) but not the ΔCR1 ΔCR2_sp1-19 strain, confirming 449 CR2_sp20 is still functional. 450
The ΔCR1 ΔCR2_sp1-19 strain was assayed in the biofilm enrichment assay at the same 451 counts of bacteria and bacteriophages as described above. Interestingly, even with CR2_sp20 452 proximal to the leader, the rate of spacer acquisition was still low with 99% (513 out of 518 453 resistant isolates) gaining resistance to DMS3vir through T4P mutations and only 1% (5 out of 454
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

518) isolates gaining resistance to DMS3vir through spacer acquisition (Fig. 7, bar second from 455 the right). 456
The CRISPR array of isolates of the ΔCR1 ΔCR2_sp1-19 mutant that gained resistance 457 through spacer acquisition was sequenced to determine the location on the DMS3 genome from 458 which the newly acquired spacers were derived. As expected, three of the isolates had spacers 459 derived from target regions close to the target of CR2_sp20 (Table S4), however all of the 460 spacers in isolate #4 target regions of DMS3 were at least 14.5 kilobases from the target region 461 of CR2_sp20. Interestingly, the region of DMS3 targeted by these new spacers, while far from 462 the target region of CR2_sp20, is very close (about 1000 bps) to the target region of CR2_sp1 463 even though CR2_sp1 was deleted in this strain, suggesting the region near the CR2_sp1 target is 464 potentially more amenable to spacer acquisition. 465
An alternate explanation as to why CR2_sp1 is more efficient at priming than CR2_sp20 466 is the difference in sequences that prevent CRISPR interference. While CR2_sp1 has 5 467 mismatches between the spacer and the protospacer, CR2_sp20 instead has a only single 468 mutation in the PAM. Therefore to test the hypothesis that CR2_sp20 would be more efficient at 469 priming if it had sequence changes in position similar to CR2_sp1, a version of the ΔCR1 470 ΔCR2_sp1-19 strain was constructed in which CR2_sp20 had mismatches at identical positions 471 as CR2_sp1, referred to as ΔCR1 ΔCR2_sp1-20 + sp20-5MM (for 5 mismatches) and illustrated 472 in Figure S3. This strain was assayed in the biofilm enrichment assay at the same counts of 473 bacteria and bacteriophages as described above with a key difference being DMS3vir-PC was 474 used instead of DMS3vir. We used DMS3vir-PC here because to be similar to CR2_sp1, 475 CR2_sp20-5MM would need to target a region of DMS3 with the correct PAM. For the biofilm 476 enrichment assay, three biological replicates were performed with at least 200 isolates each. 477
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

Strikingly, unlike the low rate of CRISPR adaptation observed with the ΔCR1 ΔCR2_sp1-19 478 strain (5 out of 518 resistant isolates), with the ΔCR1 ΔCR2_sp1-20 + sp20-5MM strain, 424 out 479 of 476 resistant isolates gained resistance through spacer acquisition (89%, Figure 7). This was 480 the highest rate of CRISPR adaptation in resistant isolates observed among any strain, including 481 wild-type P. aeruginosa. 482
Overall these data suggest that certain spacers are much better at priming than others, and 483 one reason for the variability is the specific sequences causing mismatches between the spacer 484 and the protospacer, rather than the location of the target or overall sequence of the spacer. 485 Furthermore, primed adaptation is the predominant (and perhaps only) mode of spacer 486 acquisition in the P. aeruginosa Type I-F CRISPR-Cas system as we were unable to detect naïve 487 adaptation using the biofilm enrichment assay. 488
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

DISCUSSION 489 By developing a strategy of enriching for biofilm-grown cells during challenge with the 490
lytic bacteriophage DMS3vir, we have reduced the frequency of phage receptor mutations 491 dominating the resistant population. We also developed an assay system that allows robust 492 spacer acquisition through endogenous expression of the P. aeruginosa Type I-F CRISPR 493 system. This assay system allowed us to characterize the genetic requirements of primed 494 CRISPR adaptation against a lytic bacteriophage, and finding that the cas1 gene is required for 495 CRISPR adaptation, the recG gene contributes to efficient primed CRISPR adaptation, and gene 496 recD gene is dispensable for primed CRISPR adaptation. Furthermore, due to the presence of 3 497 putative priming spacers against DMS3vir in the native CRISPR2 array in P. aeruginosa, the 498 contribution of specific spacers to CRISPR acquired immunity against DMS3vir was 499 investigated, finding a strong bias wherein CR2_sp1 is efficient in conferring CRISPR immunity 500 through primed adaptation. This high efficiency of priming by CR2_sp1 is not simply due to the 501 proximity of CR2_sp1 to the CRISPR2 leader sequence and is likely the result of the specific 502 sequences that prevent functional CRISPR interference. 503
Preventing bacteriophage adsorption is a common strategy employed by bacteria to gain 504 resistance to lytic bacteriophages, analogous to plasmid-based adsorption interference (34), but 505 mutants that directly affect the function of surface receptors can come with a fitness cost (35). 506 Accordingly, the goal of the biofilm enrichment system was to not simply investigate 507 mechanisms of CRISPR adaption in P. aeruginosa but to also develop an assay that may provide 508 insight on why the Type I-F CRISPR-Cas system is typically found in a broad range of P. 509 aeruginosa strains. The advantage provided by CRISPR adaptation when grown in a biofilm in 510
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

the presence of bacteriophage compared to CRISPR-deficient P. aeruginosa was significant and 511 reproducible, perhaps suggesting one reason why this system is conserved in microbes. 512
Similar to what was observed in other studies involving the native Type I-F system of P. 513 aeruginosa, there were many similarities in the spacers incorporated using the biofilm 514 enrichment assay described here and what has been published using a plasmid loss-based system 515 with the Type I-F system of P. atrosepticum, despite the differences between the two assay 516 systems. In both systems during a primed spacer acquisition event, multiple spacers are 517 incorporated at a higher rate than single spacers (~75% versus 25%), spacer incorporation is not 518 equal among CRISPR arrays, and a nearly identical strand bias exists regarding selected 519 protospacers. Therefore these observations likely reflect general characteristics of the Type I-F 520 system and are not associated with the type of target (i.e. plasmid versus lytic bacteriophage) or 521 the bacterium that encodes the CRISPR system. 522
The observation that multiple spacers were typically inserted during an acquisition event 523 suggests the ability of the P. aeruginosa to rapidly modify its CRISPR loci. This finding is at 524 odds with our previous work (19) showing surprisingly conserved spacer arrays across isolates of 525 this microbe from India and two sites in the United States (Hanover, NH and Pittsburgh, PA). It 526 is possible that the rate of acquisition observed in the lab does not reflect the rates actually 527 occurring in natural settings, or alternatively, perhaps such newly acquired spacers are easily 528 lost. 529
The biofilm selection assay was used to test the requirements of genome stability proteins 530 previously implicated in CRISPR adaptation. We found that RecD was dispensable for CRISPR 531 adaptation in P. aeruginosa, which was not surprising since the hypothesized role of the 532 RecBCD complex in P. aeruginosa adaptation is to generate single stranded DNA for Cas1 533
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

during naïve adaptation, and only primed adaptation was observed in the biofilm selection assay. 534 Intriguingly, we found that priming could still occur in the absence of RecG, albeit at a greatly 535 reduced rate. The putative role of RecG in CRISPR adaptation is to relieve R-loops formed at 536 sites where the crRNA ribonucleoprotein complex has partially bound its target, thereby 537 allowing Cas1 to access the single stranded DNA generated by Cas3. Our data support this 538 model, but demonstrate that RecG is not completely required for Cas1 to access the single 539 stranded target and incorporate new spacers. 540
One of the most striking results obtained with the biofilm enrichment assay is the 541 difference in priming efficiency between CR2_sp20 and CR2_sp1 spacers. It has been shown in 542 the E. coli Type I-E CRISPR-Cas system that spacers that match their protospacer with 100% 543 complementarity but lacks a consensus PAM (such as CR2_sp20) are efficient priming spacers 544 (29). Yet when either CR2_sp1 or all spacers except CR2_sp20 and 21 were deleted, spacer 545 acquisition frequency was greatly reduced to levels similar to those observed in the recG mutant, 546 indicating that CR2_sp20 is a very poor priming spacer despite 100% complementarity with a 547 non-consensus PAM. The inefficiency of CR2_sp20 at priming is not due to location of 548 CR2_sp20 relative to the CRISPR2 leader since both a leader-distant and a leader-proximal 549 CR2_sp20 had a poor priming efficiency. Instead, the inefficiency of CR2_sp20 at priming can 550 be resolved by engineering CR2_sp20 to have the same mismatch sites as CR2_sp1 between the 551 spacer and the bacteriophage target, which results in rates of priming more efficient than 552 CR2_sp1. Further research is needed to determine which of the 5 mismatches are the most 553 important for this phenomenon, and why. 554
Interestingly, none of the sequenced newly acquired spacers were likely primed by 555 CR2_sp17, suggesting that when “escaping” CRISPR interference, a single nucleotide deletion 556
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

can be more beneficial to the bacteriophage than a single nucleotide substitution since our data 557 indicate the substitution can be more easily over come through CRISPR priming. Avoidance of 558 priming for a bacteriophage infecting P. aeruginosa is especially important since no naïve 559 adaptation was detected using the biofilm enrichment assay, suggesting that primed adaptation is 560 the dominant form of CRISPR adaptation in the P. aeruginosa Type I-F. 561
Overall the biofilm enrichment assay described here provides a system to study 562 endogenous CRISPR adaptation in the P. aeruginosa Type I-F system against a lytic 563 bacteriophage and provides a complementary dataset generated by CRISPR studies in which 564 artificial expression constructs or mutants are used to investigate the mechanisms of CRISPR 565 adaptation. 566 567 References 568 1. Makarova KS, Wolf YI, Alkhnbashi OS, Costa F, Shah SA, Saunders SJ, 569
Barrangou R, Brouns SJ, Charpentier E, Haft DH, Horvath P, Moineau S, Mojica 570 FJ, Terns RM, Terns MP, White MF, Yakunin AF, Garrett RA, van der Oost J, 571 Backofen R, Koonin EV. 2015. An updated evolutionary classification of CRISPR-Cas 572 systems. Nature reviews. Microbiology 13:722-736. 573
2. Shmakov S, Abudayyeh OO, Makarova KS, Wolf YI, Gootenberg JS, Semenova E, 574 Minakhin L, Joung J, Konermann S, Severinov K, Zhang F, Koonin EV. 2015. 575 Discovery and functional characterization of diverse class 2 CRISPR-Cas systems. 576 Molecular cell 60:385-397. 577
3. Grissa I, Vergnaud G, Pourcel C. 2007. The CRISPRdb database and tools to display 578 CRISPRs and to generate dictionaries of spacers and repeats. BMC bioinformatics 8:172. 579
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4. Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero 580 DA, Horvath P. 2007. CRISPR provides acquired resistance against viruses in 581 prokaryotes. Science 315:1709-1712. 582
5. Brouns SJ, Jore MM, Lundgren M, Westra ER, Slijkhuis RJ, Snijders AP, Dickman 583 MJ, Makarova KS, Koonin EV, van der Oost J. 2008. Small CRISPR RNAs guide 584 antiviral defense in prokaryotes. Science 321:960-964. 585
6. Marraffini LA, Sontheimer EJ. 2008. CRISPR interference limits horizontal gene 586 transfer in staphylococci by targeting DNA. Science 322:1843-1845. 587
7. Barrangou R, Marraffini LA. 2014. CRISPR-Cas systems: Prokaryotes upgrade to 588 adaptive immunity. Molecular cell 54:234-244. 589
8. Yosef I, Goren MG, Qimron U. 2012. Proteins and DNA elements essential for the 590 CRISPR adaptation process in Escherichia coli. Nucleic acids research 40:5569-5576. 591
9. Swarts DC, Mosterd C, van Passel MW, Brouns SJ. 2012. CRISPR interference 592 directs strand specific spacer acquisition. PloS one 7:e35888. 593
10. Vorontsova D, Datsenko KA, Medvedeva S, Bondy-Denomy J, Savitskaya EE, 594 Pougach K, Logacheva M, Wiedenheft B, Davidson AR, Severinov K, Semenova E. 595 2015. Foreign DNA acquisition by the I-F CRISPR-Cas system requires all components 596 of the interference machinery. Nucleic acids research 43:10848-10860. 597
11. Westra ER, van Houte S, Oyesiku-Blakemore S, Makin B, Broniewski JM, Best A, 598 Bondy-Denomy J, Davidson A, Boots M, Buckling A. 2015. Parasite exposure drives 599 selective evolution of constitutive versus inducible defense. Current biology 25:1043-600 1049. 601
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

12. van Houte S, Ekroth AK, Broniewski JM, Chabas H, Ashby B, Bondy-Denomy J, 602 Gandon S, Boots M, Paterson S, Buckling A, Westra ER. 2016. The diversity-603 generating benefits of a prokaryotic adaptive immune system. Nature 532:385-388. 604
13. Zegans ME, Wagner JC, Cady KC, Murphy DM, Hammond JH, O'Toole GA. 2009. 605 Interaction between bacteriophage DMS3 and host CRISPR region inhibits group 606 behaviors of Pseudomonas aeruginosa. Journal of bacteriology 191:210-219. 607
14. Cady KC, O'Toole GA. 2011. Non-identity-mediated CRISPR-bacteriophage interaction 608 mediated via the Csy and Cas3 proteins. Journal of bacteriology 193:3433-3445. 609
15. Cady KC, Bondy-Denomy J, Heussler GE, Davidson AR, O'Toole GA. 2012. The 610 CRISPR/Cas adaptive immune system of Pseudomonas aeruginosa mediates resistance 611 to naturally occurring and engineered phages. Journal of bacteriology 194:5728-5738. 612
16. Budzik JM, Rosche WA, Rietsch A, O'Toole GA. 2004. Isolation and characterization 613 of a generalized transducing phage for Pseudomonas aeruginosa strains PAO1 and PA14. 614 Journal of bacteriology 186:3270-3273. 615
17. Ceyssens PJ, Lavigne R. 2010. Bacteriophages of Pseudomonas. Future microbiology 616 5:1041-1055. 617
18. O'Toole GA, Kolter R. 1998. Flagellar and twitching motility are necessary for 618 Pseudomonas aeruginosa biofilm development. Molecular microbiology 30:295-304. 619
19. Cady KC, White AS, Hammond JH, Abendroth MD, Karthikeyan RS, Lalitha P, 620 Zegans ME, O'Toole GA. 2011. Prevalence, conservation and functional analysis of 621 Yersinia and Escherichia CRISPR regions in clinical Pseudomonas aeruginosa isolates. 622 Microbiology 157:430-437. 623
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

20. van Belkum A, Soriaga LB, LaFave MC, Akella S, Veyrieras JB, Barbu EM, 624 Shortridge D, Blanc B, Hannum G, Zambardi G, Miller K, Enright MC, Mugnier 625 N, Brami D, Schicklin S, Felderman M, Schwartz AS, Richardson TH, Peterson TC, 626 Hubby B, Cady KC. 2015. Phylogenetic distribution of CRISPR-Cas systems in 627 antibiotic-resistant Pseudomonas aeruginosa. mBio 6. 628
21. Shanks RM, Caiazza NC, Hinsa SM, Toutain CM, O'Toole GA. 2006. 629 Saccharomyces cerevisiae-based molecular tool kit for manipulation of genes from gram-630 negative bacteria. Applied and environmental microbiology 72:5027-5036. 631
22. Liberati NT, Urbach JM, Miyata S, Lee DG, Drenkard E, Wu G, Villanueva J, Wei 632 T, Ausubel FM. 2006. An ordered, nonredundant library of Pseudomonas aeruginosa 633 strain PA14 transposon insertion mutants. Proceedings of the National Academy of 634 Sciences of the United States of America 103:2833-2838. 635
23. Ha DG, Richman ME, O'Toole GA. 2014. Deletion mutant library for investigation of 636 functional outputs of cyclic diguanylate metabolism in Pseudomonas aeruginosa PA14. 637 Applied and environmental microbiology 80:3384-3393. 638
24. Kuchma SL, Ballok AE, Merritt JH, Hammond JH, Lu W, Rabinowitz JD, O'Toole 639 GA. 2010. Cyclic-di-GMP-mediated repression of swarming motility by Pseudomonas 640 aeruginosa: the pilY1 gene and its impact on surface-associated behaviors. Journal of 641 bacteriology 192:2950-2964. 642
25. Datsenko KA, Pougach K, Tikhonov A, Wanner BL, Severinov K, Semenova E. 643 2012. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial 644 immunity system. Nature communications 3:945. 645
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

26. Wilkinson ME, Nakatani Y, Staals RH, Kieper SN, Opel-Reading HK, McKenzie 646 RE, Fineran PC, Krause KL. 2016. Structural plasticity and in vivo activity of Cas1 647 from the type I-F CRISPR-Cas system. The Biochemical journal 8:1063-1072. 648
27. Richter C, Dy RL, McKenzie RE, Watson BN, Taylor C, Chang JT, McNeil MB, 649 Staals RH, Fineran PC. 2014. Priming in the Type I-F CRISPR-Cas system triggers 650 strand-independent spacer acquisition, bi-directionally from the primed protospacer. 651 Nucleic acids research 42:8516-8526. 652
28. Levy A, Goren MG, Yosef I, Auster O, Manor M, Amitai G, Edgar R, Qimron U, 653 Sorek R. 2015. CRISPR adaptation biases explain preference for acquisition of foreign 654 DNA. Nature 520:505-510. 655
29. Fineran PC, Gerritzen MJ, Suarez-Diez M, Kunne T, Boekhorst J, van Hijum SA, 656 Staals RH, Brouns SJ. 2014. Degenerate target sites mediate rapid primed CRISPR 657 adaptation. Proceedings of the National Academy of Sciences of the United States of 658 America 111:E1629-1638. 659
30. Shmakov S, Savitskaya E, Semenova E, Logacheva MD, Datsenko KA, Severinov K. 660 2014. Pervasive generation of oppositely oriented spacers during CRISPR adaptation. 661 Nucleic acids research 42:5907-5916. 662
31. Li M, Wang R, Zhao D, Xiang H. 2014. Adaptation of the Haloarcula hispanica 663 CRISPR-Cas system to a purified virus strictly requires a priming process. Nucleic acids 664 research 42:2483-2492. 665
32. Ivancic-Bace I, Cass SD, Wearne SJ, Bolt EL. 2015. Different genome stability 666 proteins underpin primed and naive adaptation in E. coli CRISPR-Cas immunity. Nucleic 667 acids research 43:10821-10830. 668
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

33. Hale C, Kleppe K, Terns RM, Terns MP. 2008. Prokaryotic silencing (psi)RNAs in 669 Pyrococcus furiosus. Rna 14:2572-2579. 670
34. Forde A, Daly C, Fitzgerald GF. 1999. Identification of four phage resistance plasmids 671 from Lactococcus lactis subsp. cremoris HO2. Applied and environmental microbiology 672 65:1540-1547. 673
35. Avrani S, Wurtzel O, Sharon I, Sorek R, Lindell D. 2011. Genomic island variability 674 facilitates Prochlorococcus-virus coexistence. Nature 474:604-608. 675
676 677 678
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

Figures Legends 679 680 Figure 1. Cartoon of the Type 1-F CRISPR-Cas system in Pseudomonas aeruginosa strain 681 UCBPP-PA14. Gene names and spacers numbers are indicated, and described in the text. 682 683 Figure 2. Schematic of biofilm enrichment assay. Pseudomonas aeruginosa and DMS3vir are 684 co-inoculated at a multiplicity of infection of 0.01 using 2.5x108 CFU and 2.5x106 PFU of P. 685 aeruginosa and DMS3 vir, respectively, during each of the two biofilm incubations. 686 687 Figure 3. Number of viable P. aeruginosa cells in the planktonic and biofilm population after 688 the biofilm enrichment assay. After both the first (A) and second (B) 24 hour challenge in 689 biofilm-inducing medium, 1 mL of the planktonic culture was collected, serially dilutions 690 prepared, and plated to measure CFUs. Additionally, after both the first (C) and second (D) 24 691 hour challenge in biofilm-inducing medium the total biofilm population at the air-liquid interface 692 in each well was isolated using a cell scraper, washed, re-suspended in PBS, serially diluted, and 693 plated to measure CFUs. Error bars represent standard deviation from three replicates and (*) 694 indicates significant difference (P<0.05, Student’s t-test) of the specified condition compared to 695 the equivalent condition without DMS3vir. 696 697 Figure 4. Resistance to DMS3 vir in the biofilm enrichment assay is gained either through Type-698 IV pilus loss-of-function or Cas1-dependent spacer acquisition. After the second biofilm 699 incubation in the biofilm enrichment assay both the biofilm population and planktonic population 700 of P. aeruginosa were plated for single colonies. At least 200 colonies in each specified 701
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

condition from at least two replicates were re-patched on LB-agar. (A) Isolates that were 702 resistant to DMS3vir and twitch-negative were scored as a Type IV pilus (T4P) mutant, while the 703 isolates that were resistant to DMS3vir, twitch-positive, and demonstrated spacer acquisition as 704 determined by PCR of the CRISPR arrays were scored as a spacer acquisition-positive isolate. 705 Type IV pilus mutant and spacer acquisition-positive isolates are displayed in each condition as a 706 percentage of the total DMS3vir resistant population in that condition. (B) After 24 hours co-707 incubation of either WT or the CRISPR deficient (∆CR) P. aeruginosa with DMS3vir at a 708 multiplicity of infection of 0.01 at 37 degrees C in 5mL of LB, 200 single colonies from two 709 replicates in each condition were isolated, scored and displayed as described in panel A. (C and 710 D) 100 randomly selected spacer acquisition positive isolates from the WT and DMS3vir co-711 incubation condition in the biofilm enrichment assay were scored for (C) inserting either a single 712 spacer or multiple spacers and (D) inserting a new spacer into the CRISPR1, CRISPR2, or both 713 CRISPR1 and CRISPR2 arrays as determined by PCR. 714 715 Figure 5. The three putative priming spacers located in the native CRISPR2 locus of P. 716 aeruginosa strain UCBPP-PA14 (top) along with their cognate DMS3 target (bottom). The PAM 717 position is underlined and the DMS3 location listed indicates the position of the 32 base pair 718 protospacer in the DMS3 genome. 719 720 Figure 6. DMS3 targets of newly acquired spacers demonstrate a bias. The 83 sequenced spacers 721 acquired by WT P. aeruginosa when incubated with DMS3vir in the biofilm enrichment assay 722 within 6000 base pairs of the CRISPR2 spacer 1 (CR2_sp1) target are displayed as a function of 723 both the distance from the CR2_sp1 target and which DMS3 strand (sense or antisense) each 724
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

targets. The dashed line represents the CR2_sp1 target location with the x-axis representing the 725 distance from the CR2_sp1 target divided into segments of 500 base pairs either upstream or 726 downstream of the CR2_sp1 target. Each bars represent number of spacers targeting within that 727 particular 500 base pair segment of DMS3, with blue bars plotted above the X axis representing 728 targets on the positive sense DMS3 strand with the yellow bars plotted below the X axis 729 representing targets on the negative sense DMS3 strand. 730 731 Figure 7. Contributions to new spacer acquisition. RecD is dispensable for primed spacer 732 acquisition, RecG and CRISPR2 spacer 1 are necessary for efficient primed spacer acquisition, 733 and CRISPR2 spacer 20 is sufficient for primed spacer acquisition when the mismatches are in 734 similar positions as CRISPR2 spacer 1 in the biofilm enrichment assay. After the second 735 challenge in the biofilm enrichment assay, the biofilm population of P. aeruginosa (either WT or 736 the specified mutant) co-incubated with either DMS3vir (first six columns) or DMS3vir-PC (last 737 column on the right) was plated for single colonies. At least 200 colonies in each specified 738 condition from at least two replicates were re-patched on LB-agar. Isolates that were resistant to 739 DMS3vir and twitch-negative were scored as a Type IV pilus (T4P) mutant, while isolates that 740 were resistant to DMS3vir, twitch-positive, and demonstrated spacer acquisition as determined 741 by PCR of the CRISPR arrays were scored as a spacer acquisition-positive isolate. Type IV pilus 742 mutant and spacer acquisition positive isolates from each condition are displayed as a percentage 743 of the total DMS3 vir resistant population in that condition. 744 745
on June 9, 2018 by guesthttp://jb.asm
.org/D
ownloaded from