Upload
others
View
1
Download
0
Embed Size (px)
Citation preview
Jour
nal o
f Cel
l Sci
ence
RESEARCH ARTICLE
Transport along the dendritic endoplasmic reticulum mediates thetrafficking of GABAB receptors
José I. Valenzuela1,2, Matı́as Jaureguiberry-Bravo1,2, Daniela A. Salas1,2, Omar A. Ramı́rez1,2,3,Vı́ctor H. Cornejo1,2, Hsiangmin E. Lu4,5, Thomas A. Blanpied4,6 and Andrés Couve1,2,*
ABSTRACT
In neurons, secretory organelles within the cell body are
complemented by the dendritic endoplasmic reticulum (ER) and
Golgi outposts (GOPs), whose role in neurotransmitter receptor
trafficking is poorly understood. c-aminobutyric acid (GABA) type B
metabotropic receptors (GABABRs) regulate the efficacy of synaptic
transmission throughout the brain. Their plasmamembrane availability
is controlled by mechanisms involving an ER retention motif and
assembly-dependent ER export. Thus, they constitute an ideal
molecular model to study ER trafficking, but the extent to which the
dendritic ER participates in GABABR biosynthesis has not been
thoroughly explored. Here, we show that GABAB1 localizes
preferentially to the ER in dendrites and moves long distances within
this compartment. Not only diffusion but also microtubule and dynein-
dependent mechanisms control dendritic ER transport. GABABRs
insert throughout the somatodendritic plasma membrane but dendritic
post-ER carriers containing GABABRs do not fuse selectively with
GOPs. This study furthers our understanding of the spatial selectivity
of neurotransmitter receptors for dendritic organelles.
KEY WORDS: GABA, Dendrite, Endoplasmic reticulum, Golgi
outposts, Neuron, Trafficking
INTRODUCTIONThe control of the availability of neurotransmitter receptors at
appropriate subdomains of the plasma membrane is crucial for
neuronal function and synaptic plasticity. Availability is governed
by mechanisms of intracellular trafficking that deliver newly
synthesized receptors to the plasma membrane and remove
them for storage, recycling or degradation. These mechanisms
determine the activity-dependent strengthening or weakening of
synapses (Ehlers, 2000; Clem and Barth, 2006).
Two major secretory pathways operate in dendrites (Horton and
Ehlers, 2003). In a canonical pathway, proteins are synthesized and
post-translationally modified in the somatic ER, transported to a
perinuclear Golgi for additional modifications and targeted distally
in long-haul post-Golgi vesicles. The distribution of the continuous
ER throughout dendrites (Broadwell and Cataldo, 1983; Spacek
and Harris, 1997), and the localization of functional Golgi
outposts (GOPs), which are defined as discrete Golgi structures
discontinuous with the somatic organelle (Horton and Ehlers,
2003; Horton and Ehlers, 2004; Ye et al., 2007), suggest that local
secretory pathways also exist in neurons. Thus, proteins can be
synthesized and transported by diffusion along the somatodendritic
ER and exported through specialized ER exit sites (ERES) in
dendrites for local Golgi trafficking and delivery to the plasma
membrane (Aridor et al., 2004; Cui-Wang et al., 2012; Chen et al.,
2013). Importantly, glutamate receptors (AMPARs and NMDARs)
are differentially sorted to the perinuclear Golgi or GOPs (Jeyifous
et al., 2009). However, the role of dendritic organelles in the
trafficking of neurotransmitter receptors is not fully understood.
c-aminobutyric acid (GABA) is the major inhibitoryneurotransmitter in the central nervous system, and GABA type
B metabotropic receptors (GABABRs) mediate the slow and
prolonged phase of inhibitory postsynaptic potentials (Bettler
et al., 2004). The receptors are found in both somatodendritic and
axonal compartments, and are heteromers composed of two
subunits, GABAB1 and GABAB2, encoded by the Gabbr1 and
Gabbr2 genes, respectively. GABAB1 contains an arginine-X-
arginine (RXR)-type ER retention motif in the carboxyl-terminus
that is inactivated by GABAB2, resulting in the assembly of a
heterodimer and delivery to the plasma membrane (Margeta-
Mitrovic et al., 2000). In spite of their key function in regulating
synaptic transmission, the mechanisms that control their
availability remain for the most part unexplored, especially the
mechanisms that regulate their exocytic route in dendrites.
Conventional vesicular mechanisms regulate multiple stages of
GABABR trafficking (Pooler et al., 2009; Vargas et al., 2008;
Grampp et al., 2007; Biermann et al., 2010). However, segregated
GABAB1 and GABAB2 subunits are detected by light and electron
microscopy in hippocampal and visual cortex neurons (Gonchar
et al., 2001; Ramı́rez et al., 2009; Biermann et al., 2010). Because
GABAB1 is exported from the ER only upon binding to GABAB2 this
evidence suggests that they utilize an alternative ER transport
pathway into dendrites during the early stages of their biosynthetic
itinerary. However, the roles of dendritic organelles in the trafficking
of GABABRs are not well delineated. Here, we combine fixed and
live-cell confocal and super-resolution imaging of intracellular
GABAB subunits with the use of organelle reporters and trafficking
blockers to explore the contribution of the dendritic ER and GOPs to
the exocytic pathway of GABABRs in cultured hippocampal neurons.
RESULTSGABAB1 colocalizes with the ER in dendritesLocal dendritic trafficking requires newly synthesized membrane
proteins to dwell in, assemble in and exit from the
1Program of Physiology and Biophysics, ICBM, Faculty of Medicine, Universidad deChile, Santiago CP8380453, Chile. 2Biomedical Neuroscience Institute, BNI, Facultyof Medicine, Universidad de Chile, Santiago CP8380453, Chile. 3Program of Anatomyand Development, ICBM, Faculty of Medicine, Universidad de Chile, SantiagoCP8380453, Chile. 4Department of Physiology, University of Maryland School ofMedicine, Baltimore, MD 21201, USA. 5Program in Molecular Medicine, University ofMaryland School of Medicine, Baltimore, MD 21201, USA. 6Program in Neuroscience,University of Maryland School of Medicine, Baltimore, MD 21201, USA.
*Author for correspondence ([email protected])
Received 10 February 2014; Accepted 16 May 2014
� 2014. Published by The Company of Biologists Ltd | Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3382
mailto:[email protected]
Jour
nal o
f Cel
l Sci
ence
somatodendritic ER. Because GABABR assembly is requiredfor the export of the receptor from the ER, the existence of
segregated subunits in dendrites is a good reporter of residence inthe ER (Ramı́rez et al., 2009). To investigate this, we comparedthe colocalization of transfected GABAB1 and GABAB2, the twosubunits of the GABABR, to that of recombinant GABAAa4 andGABAAb3 (encoded by Gabra4 and Gabrb3, respectively),two subunits of the non-related ionotropic GABAA receptor(GABAAR). Colocalization was significantly lower for GABABthan for GABAA subunits (Fig. 1A,B). These observations areconsistent with previous reports (Gonchar et al., 2001; Ramı́rezet al., 2009) and indicate that segregated GABAB subunits are
abundant in dendrites, possibly revealing dendritic ER transportand assembly.
To determine whether segregated GABAB1 subunits are enriched
in the dendritic ER, we carried out quantitative colocalization withmarkers of secretory organelles. GABAB1, overexpressed eitheralone or in combination with GABAB2, colocalized preferentiallywith the ER and, to a lesser degree, with the ER-to-Golgi
intermediate compartment (ERGIC) and GOPs (Fig. 1C–E). Inagreement with the role of the GABAB2 subunit in controllingreceptor export from the ER, the colocalization of GABAB1 with
the ER decreased significantly in the presence of GABAB2(GABAB1, 5462%; GABAB1+GABAB2, 4563%; P50.025,Student’s t-test; 6s.e.m.) (Fig. 1D,E). The colocalization ofGABAB1 with the ER was comparable in regions of low andhigh network complexity, such as dendritic shafts and branchingpoints, respectively (Cui-Wang et al., 2012) (shaft, 5266%;branching points, 5064%; P50.758, Student’s t-test) (Fig. 1F).To rule out the possibility of artificial colocalization due tooverexpression, we analyzed neurons derived from GABAB1–
EGFP transgenic mice, which express GABAB1–EGFP atendogenous levels (Casanova et al., 2009). Colocalizationbetween GABAB1 and the ER in this model was comparable to
that observed with overexpressed subunits (Fig. 1G,H). Moreimportantly, blocking ER export in transgenic neurons with Sar1-H79G–HA, a dominant negative form of Sar1 that is locked in a
GTP-bound state and blocks export from the ER (Ward et al.,2001), increased the colocalization of GABAB1 with the ER(control, 3963%; Sar1-H79G–HA, 5064%; P50.04, Student’s t-test) (Fig. 1G,H), indicating that GABAB1 accumulates in the
dendritic ER when export is prevented. These results are consistentwith GABAB1 ER retention and export (Margeta-Mitrovic et al.,2000).
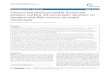
Fig. 1. GABAB1 is enriched in and traffics through the ER in dendrites. (A) Rat hippocampal neurons were co-transfected with GABAB1–Myc (GB1–MYC,green) and GABAB2–FLAG (GB2–FLAG, red) or with the GABAA subunit fusion proteins
pH/BBSb3 (GAb3, green) and RFP-BBSa4 (GAa4, red). Proximal dendritesare shown. Scale bar: 10 mm. (B) Left plot, percentage colocalization of GABAB2 with GABAB1 and of RFP-BBSa4 with pH/BBSb3. Right plot, percentagecolocalization of GABAB1 with GABAB2 or of
pH/BBSb3 with RFP-BBSa4. n54–8 neurons. (C) Neurons were transfected with GABAB1–Myc (green) and labeled withantibodies against Myc and KDEL (ER marker), ERGIC-53 (ERGIC marker) or GM130 (cis-Golgi marker) (red). The top panels are positive controls withGABAB1–Myc and two different secondary antibodies. (D) Same as in C for neurons co-transfected with GABAB1–Myc (red) and GABAB2 (purple).(E) Percentage of colocalization of GABAB1–Myc with KDEL, ERGIC-53 and GM130 (black bars) or of GABAB1–Myc with the same organelle markers in thepresence of GABAB2 (white bars). GABAB1–Myc was used as positive control (hatched bar). n55–7 neurons. (F) Percentage colocalization of GABAB1–Mycwith KDEL in dendritic shafts and branching points (BP). GABAB1–Myc was used as positive control (gray bar). n54–6 neurons. (G) Neurons from transgenicmice expressing GABAB1–GFP (green) were transfected with ER–RFP (red) in the presence or absence of Sar1-H79G–HA (purple). (H) Percentage ofcolocalization of GABAB1–GFP with ER–RFP. n56–7 neurons. All quantitative data show the mean6s.e.m.; *P,0.05; **P,0.01; n.s., non-significant;Student’s t-test.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3383
Jour
nal o
f Cel
l Sci
ence
GABAB1 transports within the ERWe then explored whether the ER functions as a transport
organelle for GABAB1. We first carried out live-cell imaging tocharacterize the dynamic relationship between intracellularGABAB1 and the ER in the soma and dendrites of neurons co-transfected with GABAB1–mRFP or GABAB1–EGFP and ER–
BFP, a vector that encodes a blue fluorescent protein fused to thehuman ER resident protein Sec61b – the encoded fusion proteinstrongly labels the ER (Zurek et al., 2011). Although the spatial
resolution was not sufficient to visualize the tubular networkbecause of its densely packed nature (Spacek and Harris, 1997),punctate and irregular structures containing fluorescent GABAB1were observed in the somatic and dendritic compartments, andtheir distribution patterns changed over time (Fig. 2A,B, toppanels, arrows). Not every puncta of GABAB1 colocalized with
the ER, in agreement with the results presented in Fig. 1, but theoverall structure often rearranged in synchrony. By contrast, thedynamics of the Golgi, labeled with either Golgi–EYFP (EYFPfused to the Golgi-targeted membrane-anchoring signal peptide
from human b-1,4-galactosyltransferase; labels Golgi in thesoma) or Galt2–mCherry (Galt2 is officially known asgalactosyltransferase 2; labels Golgi in the dendrites) and the
ERGIC, labeled with ERGIC-53–YFP (Ward et al., 2001), werenoticeably different from the dynamics of GABAB1 in bothcompartments (Fig. 2A–C).
Next, we explored the diffusion of GABAB1 within thedendritic ER using fluorescence recovery after photobleaching(FRAP, Fig. 2D) on an 8-mm dendritic segment of neuronstransfected with GABAB1–EGFP. ER-retained GABAB1–EGFPwas mobile in dendrites, and lateral mobility in ER tubules wasobserved by high-rate imaging in thick dendrites (Fig. 2E).Prolonged NMDA receptor activation with glutamate produces
reversible ER fission and disrupts the transport of luminal ERproteins (Kucharz et al., 2009; Ramı́rez et al., 2011). Thus, weused this treatment in neurons that were co-transfected with
GABAB1–EGFP and ER–RFP to confirm the ER mobility ofGABAB1–EGFP. As expected, glutamate receptor activationsignificantly altered the structure of the ER and resulted in the
co-rearrangement of ER–RFP and GABAB1–EGFP (Fig. 2F).More importantly, the mobility of GABAB1–EGFP was severelyimpaired (Fig. 2G).
These results demonstrate that GABAB1 localizes to the ER,
which allows the transport of unassembled GABAB1 in dendrites.Because the subunit is not an ER-resident protein but a traffickingcargo we will refer to it as ‘ER-associated GABAB1’ throughout
the rest of the text.
Characterization of GABAB1 transport within the dendritic ERTo gain a better understanding of ER-associated GABAB1 subunittransport we first used single-particle tracking photoactivatedlocalization microscopy (sptPALM) (Manley et al., 2008; Frost
et al., 2010). This technique is well suited to examine thedynamic properties of single intracellular molecules within smallregions, and allows a spatial resolution of ,10–50 nm (Senguptaet al., 2012). To track receptor subunits, we generated mEos2-
tagged GABAB1 (GABAB1–mEos2). mEos2 is a bright andphotostable photoconvertible fluorescent protein that is switchedfrom green to red fluorescence by UV irradiation (McKinney
et al., 2009). This construct was retained intracellularly andmaintained the characteristic trafficking properties of GABAB1(data not shown). Individual molecular locations within
transfected neurons were recorded in a 20,000-frame series, and
the summation of these positions revealed that GABAB1–mEos2was distributed throughout a complex heterogeneous structure
(Fig. 3A, right panel), similar to that reported for the dendritic ERusing structured illumination or electron microscopy (Cui-Wanget al., 2012; Broadwell and Cataldo, 1983; Spacek and Harris,1997). Single photoswitched GABAB1–mEos2 molecules were
tracked over multiple image frames and were found to be highlymobile, and their motility was not oriented in relation to the axisof the dendritic shaft (Fig. 3A, left panel; Fig. 3B), consistent
with the expected organization of the ER. The molecular velocityspanned a range from nearly immobile to 1 mm/s, with a fractionof the population moving at rates .2 mm/s. Notably, thedistribution of velocities was not different in proximal anddistal segments of a dendrite (Fig. 3B, right panel). Because mostmolecules were tracked for only brief periods over which the
influence of directed transport or confinement is minimized (,4frames or 200 ms), the molecular velocity can be used to estimatean effective diffusion coefficient D, given that the mean squaredmolecular displacement increases as 4DDt, where Dt is theobservation interval. Thus, the modal velocity of 200 nm/sobserved here indicates that a substantial proportion of GABAB1moves within the dendritic ER with a relatively slow effective D,
,0.01 mm2/s, whereas the subpopulation of molecules undergoingrapid motion does so with D surpassing 1 mm2/s. These values areconsistent with the general expectation for motion of
transmembrane proteins in the lipid bilayer (Saxton and Jacobson,1997) and are comparable to those of ER-resident proteins (Bannaiet al., 2004). It should be noted that because each molecule is
tracked only briefly, it is not possible to determine with thisapproach how frequently molecules transition from one rate ofmobility to another, and thus for how long they remain immobile.These observations suggest that two or more subpopulations of
different mobility contribute to the ER transport of GABAB1 indendrites.
We used FRAP to further examine the transport of ER-
associated GABAB1 fused to EGFP or mRFP. We used thepercentage recovery as an indication of the mobile fraction, andrecovery rate (t1/2) was used as an indication of transport velocity
within the ER. The mobile fraction of GABAB1–EGFP was5062% (6s.e.m.) and t1/2 was 7562 s, with minimal interferencefrom acquisition bleaching (Fig. 3C). The percentage recoveryand t1/2 were not affected by the fluorescent tag because the same
results were obtained with GABAB1–mRFP (Fig. 3C). Thetransport of GABAB1 was bidirectional and significantly slowerthan that of ER–RFP (Fig. 3D). The percentage recovery and t1/2were identical in primary, secondary and tertiary branches(percentage recovery of primary branches versus secondary,P50.919; versus tertiary, P50.833; t1/2 of primary branchesversus secondary, P50.242; versus tertiary, P50.275, Student’st-test) (Fig. 3E,F). Significant recovery was observed afterbleaching a large 27-mm stretch of dendritic volume, indicatingthat the ER allows long-distance protein transport (Fig. 3G).
Dendritic transport of ER-associated GABAB1 is controlled bymultiple componentsPrevious studies have suggested that the transport ofneurotransmitter receptors within the dendritic ER is controlledby lateral diffusion (Cui-Wang et al., 2012). Long-distance
transport of ER-associated GABAB1 raises the possibility thatsimple diffusion is not sufficient to explain its mobility. First, toconfirm that our FRAP-based transport assay was not confounded
by the appearance of newly synthesized GABAB1, we blocked
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3384
Jour
nal o
f Cel
l Sci
ence
Fig. 2. GABAB1 is mobile within the neuronal ER. (A) Neurons were co-transfected with GABAB1–mRFP (GB1–RFP, red), ER–BFP (green) and Golgi–EYFP(purple) and were imaged live. A somatic region is shown. Arrows show synchronous rearrangement of GABAB1–mRFP and ER–BFP signals. (B) Neuronswere co-transfected with GABAB1–EGFP (green), ER–BFP (purple) and Galt2–mCherry (red) and were imaged live. A dendrite is shown. Arrows showsynchronous rearrangement of GABAB1–EGFP and ER–BFP signals. (C) Neurons were co-transfected with GABAB1–mRFP (red), ERGIC–YFP (green) andER–BFP (purple), and imaged live. A dendrite is shown. Arrows show the asynchronous rearrangement of GABAB1–mRFP and ERGIC–YFP signals.(D) Diagram illustrating the FRAP experiments carried out for ER proteins in dendrites. (E) The ER mobility of GABAB1–EGFP was evaluated by FRAP in athick dendrite. The image was saturated to allow visualization of the tubular ER structure. (F) Dendrites of neurons transfected with GABAB1–EGFP (green) andER–RFP (red) and imaged live in the absence or presence of 100 mM glutamate (20 min). (G) The mobility of GABAB1–EGFP was evaluated by FRAP (bleachedregion 8 mm) before (left) or after (right) the glutamate treatment. Scale bars: 10 mm.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3385
Jour
nal o
f Cel
l Sci
ence
protein synthesis by treating neurons with cycloheximide (CHX,0.1 mg/ml, 90 min). Importantly, the percentage recovery and t1/2were unaffected by cycloheximide (Fig. 4A,B, GABAB1–EGFP6CHX percentage recovery, P50.166; t1/2, P50.642,Student’s t-test). Next, taking into account the fact that multiple
cytoskeletal mechanisms drive protein transport in neurons, weexamined their contribution to the ER-mediated transport ofGABAB1 in proximal dendrites. The mobile fraction of GABAB1decreased significantly after high doses of nocodazole (NocoHigh, 100 mM, 4 h), but was unaffected by low doses ofnocodazole (Noco Low, 100 nM, 5–70 min), trichostatin A
(TSA, 125 nM, 30 min) or cytochalasin D (cytoD, 5 mM, 2 h)(Noco High, 3763%, P50.002; Noco Low, 5362%, P50.214;TSA, 4762%, P50.397; cytoD, 5065%, P50.901, Student’s t-test; 6s.e.m.) (Fig. 4C). These results indicate that, in additionto diffusion, a proportion of subunit transport depends onpolymerized microtubules (which are affected by high doses of
nocodazole), but not on dynamic microtubules (which areaffected by low doses of nocodazole), acetylated microtubules
(the levels of which are increased by TSA) or the actincytoskeleton (which is disrupted by cytochalasin D).Interestingly, luminal ER–RFP and the ER membrane protein
cytochrome-b5–EGFP (Cytb5–GFP) were unaffected by highdoses of nocodazole (percentage recovery of ER–RFP6NocoHigh, P50.966; percentage recovery of Cytb5–GFP6NocoHigh, P50.827. t1/2 for ER–RFP6Noco High, P50.361; forCytb5–GFP6Noco High, P50.239, Student’s t-test) (Fig. 4E–H) arguing against a global effect of nocodazole in the transport
of proteins through the ER. To confirm that nocodazole affectedER-retained GABAB1, we preincubated neurons with brefeldinA (BFA), a treatment that results in the dispersal of the Golgiand accumulation of cargo in the ER. Under these conditions,
high doses of nocodazole still decreased the transport ofGABAB1 (control BFA, 6162%; BFA+Noco High, 3266%,
Fig. 3. Characterization of the ER transport of GABAB1. (A) The mobility of GABAB1–mEos2 was evaluated by using sptPALM in dendrites. Left panel, quiverplots show GABAB1–mEos2 (GB12mEos2) molecules mobilizing in multiple directions (inset is 26magnification). Scale bar: 1 mm. Right panel, GABAB1–mEos2 was imaged in a live neuron for 10 seconds. Scale bar: 2 mm. (B) Net velocity of GABAB1–mEos2 single molecules at initial and distal segments of thedendrite (black and red curves, respectively). (C) FRAP of GABAB1–EGFP (green symbols) and GABAB1–mRFP (red symbols) in dendrites. The uppercurves (empty symbols) correspond to the experimental bleaching without FRAP. n514–28 neurons. (D) FRAP of GABAB1–EGFP (green symbols) and ER–RFP (red symbols) in dendrites. n58 neurons. (E) Percentage recovery of GABAB1–EGFP in primary (1 )̊, secondary (2 )̊ and tertiary (3 )̊ dendriticbranches. n53–5 neurons. (F) t1/2 of recovery of GABAB1–EGFP in primary, secondary and tertiary dendritic branches. n53–5 neurons. All quantitative datashow the mean6s.e.m.; n.s., non-significant. (G) The long-range mobility of GABAB1–EGFP was evaluated by bleaching a 27-mm dendritic region in live (left) orfixed (right) cells. Scale bar: 10 mm.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3386
Jour
nal o
f Cel
l Sci
ence
P50.016, Student’s t-test) (Fig. 4I,J). Although nocodazolemight be affecting a minor BFA-independent component ofpost-ER COPII-coated carriers, the robust effect of BFA
confirms that microtubule dependence occurs mostly on ER-associated GABAB1. Control experiments demonstrated theeffectiveness of all pharmacological treatments (data notshown).
Interestingly, only TSA decreased t1/2 significantly (t1/2,6862 s; P50.026, Student’s t-test) (Fig. 4D). Although thisexperiment does not directly identify a specific transport
mechanism, tubulin acetylation could decrease GABAB1 confinementby increasing the recruitment of the molecular motors tomicrotubules (Reed et al., 2006; Dompierre et al., 2007; Cai
et al., 2009) or by decreasing ER complexity (Friedman et al.,2010; Cui-Wang et al., 2012). Because the former interpretationinvolves molecular motors and we have demonstrated that a
component of transport is microtubule dependent, we evaluated therole of kinesin-1 and dynein in the transport of dendritic GABAB1.Kinesin-1 co-immunoprecipitates with GABAB1 in brain extracts(Vidal et al., 2007), participates in the axonal localization of
GABAB1 (Valdés et al., 2012) and has been implicated inER dynamics (Woźniak et al., 2009). However, the expressionof a dominant-negative version of kinesin-1 comprising the
cargo-binding domain of Kif5C that interferes with endogenouskinesin–cargo interactions (KIF5CDN–RFP, data not shown)(Falley et al., 2009) had no effect on the GABAB1 mobile
fraction (control RFP, 4463%; KIF5CDN–RFP, 4368%;P50.872, Student’s t-test) or recovery kinetics in dendrites(control RFP, 7164 s; KIF5CDN–RFP, 6963 s; P50.290,Student’s t-test) (Fig. 5A,B).
Because the mixed microtubule orientation in proximaldendrites allows minus-end-directed dynein to drive transportbidirectionally (Kapitein et al., 2010), we used overexpression of
GFP-tagged p50 (also known as dynamitin or DCTN2) (p50–EGFP) to inhibit dynein function (Echeverri et al., 1996;Burkhardt et al., 1997; Presley et al., 1997). Importantly, p50–EGFP overexpression significantly decreased the percentage
recovery of GABAB1 (control EGFP, 5764%; p50–EGFP,4663%; P50.049, Student’s t-test), whereas it had no effect ont1/2 (Fig. 5C,D). GFP-tagged N5-bIII-spectrin (N5-bIII–EGFP),an established blocker of the bIII-spectrin–dynein complex(Devarajan et al., 1997), produced a mild but non-significantdecrease in the percentage recovery (4763%; P50.077,Student’s t-test) and did not alter t1/2 (Fig. 5C,D).
A prediction of our FRAP experiments is that dynein inhibitioncould limit the ER distribution of GABAB1 in dendrites. Thus, we
tested the intracellular distribution of transfected GABAB1–mRFP in the presence of p50–EGFP. As previously reported,dendritic morphology was altered by p50–EGFP (Kapitein et al.,2010) (Fig. 5H). More importantly, the distribution of GABAB1in p50–EGFP-expressing neurons was dramatically different fromthat of control neurons. Specifically, the distribution of ER-associated GABAB1 was restricted to the proximal regions in
dendrites, whereas it was unaltered in axons [GABAB1–mRFPmean intensity (arbitrary units) in dendrites for control EGFP,69.964.8; for p50–EGFP, 39.863.8, P50.00003. In axons forcontrol EGFP, 10.161.3; for p50–EGFP, 14.362.0, P50.095,Student’s t-test] (Fig. 5E,G,H). By contrast, dynein inhibition didnot affect EGFP or MAP2 distribution in dendrites or axons
(MAP2 mean intensity of control EGFP versus p50–EGFP for
Fig. 4. The ER transport of dendritic GABAB1 is controlled by multiple components. (A) Percentage recovery of GABAB1–EGFP for control samples orafter treatment with 0.1 mg/ml cycloheximide (CHX) for 90 min. n54 and 5 neurons, respectively. (B) Same as above for the t1/2. (C,D) Same as in A,B forneurons under control conditions or treated with 100 mM nocodazole for 4 h (Noco High), 100 nM nocodazole for 5–70 min (Noco Low), 125 nM TSA for30 min or 5 mM cytochalasin D for 2 h (Cyto D). n528, 9, 8, 20 and 4 neurons, respectively. At least three independent experiments were performed percondition. (E,F) Same as in A,B for neurons transfected with ER–RFP under control conditions or following treatment with 100 mM nocodazole for 4 h (ER–RFP+Noco). n54 and 7 neurons, respectively. (G,H) Same as in A,B for neurons transfected with Cytb5–GFP under control conditions or followingtreatment with 100 mM nocodazole for 4 h (Cytb5–GFP+Noco). n55 and 6 neurons, respectively. (I,J) Same as in A,B for neurons treated with 0.5 mg/ml BFA for20 h or 0.5 mg/ml BFA for 20 h+100 mM nocodazole for 4 h (BFA+Noco). n53 and 4 neurons, respectively. All data show the mean6s.e.m.; *P,0.05; **P,0.01;n.s., non-significant; Student’s t-test.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3387
Jour
nal o
f Cel
l Sci
ence
dendrites, P50.309; for axons, P50.144. EGFP mean intensity ofcontrol EGFP versus p50–EGFP for dendrites, P50.334; foraxons, P50.101, Student’s t-test). To further analyze the effect ofp50–EGFP on the intracellular distribution of GABAB1–mRFP,we calculated a polarity index, where an index of .0 or ,0indicates polarization towards dendrites or axons, respectively(Kapitein et al., 2010). The polarity index of ER-associatedGABAB1 in control neurons was 0.7560.03 (Fig. 5F), consistentwith the preferential somatodendritic distribution of GABAB1subunits (Biermann et al., 2010; Valdés et al., 2012). By contrast,upon expression of p502EGFP, the polarity index of GABAB1decreased significantly (0.4760.06; P50.0005, Student’s t-test)(Fig. 5F). As expected, the polarity index of MAP2 and EGFPwere unaffected by dynein inhibition (MAP2, P50.111; EGFP,P50.386, Student’s t-test). Taken together, these datademonstrate that dynein is essential for one component ofdendritic ER transport and for the targeting of GABAB1 subunits.
GABABRs traffic through canonical and local secretoryroutesTransport through a subcompartment of the dendritic ER
determines the specific sorting of NMDARs to GOPs (Jeyifouset al., 2009). Likewise, the localization and transport of nascentGABAB1 subunits within the dendritic ER might result in local
trafficking through GOPs. To examine whether GABABRs trafficthrough GOPs, we co-transfected GABAB1–Myc and GABAB2–
FLAG, which assemble and traffic to the plasma membrane,together with ARF1-Q71I–HA, a constitutively active mutant ofthe small GTPase ARF1 – involved in COPI recruitment – that
prevents export from the Golgi and produces the accumulation ofcargo in the somatic Golgi and GOPs in neurons (Zhang et al.,1994; Jeyifous et al., 2009). Under steady-state conditions,
GABAB1 was mostly excluded from the Golgi in the perinuclearregion and in dendrites (Fig. 6A,B). Upon expression of ARF1-Q71I–HA, GABAB1 accumulated significantly in both somaticand dendritic Golgi structures (P50.0061 and P50.0001,respectively, Student’s t-test) (Fig. 6A,B,E). These resultsindicate that newly synthesized receptors traffic through theperinuclear Golgi and GOPs.
It has been suggested that dwell times and diffusion within theER define the exploration volumes of nascent proteins and,therefore, spatially determine ER export domains (Herpers and
Rabouille, 2004; Cui-Wang et al., 2012). Thus, we reasoned thatGABAB1 subunits with long ER residency would preferably feedGOPs, whereas rapidly exported subunits might preferentially use
the perinuclear Golgi. To test this hypothesis, we used GABAB1-DC–Myc, a mutant subunit that contains a deletion of 104 aminoacids in its carboxyl terminus. This mutant protein lacks the ER
Fig. 5. Dendrite-specific ERtransport and targeting of GABAB1depends on dynein. (A) Percentagerecovery of GABAB1–EGFPfluorescence for neurons transfectedwith RFP or KIF5DN–RFP. n517 and8 neurons, respectively. Threeindependent experiments wereperformed per condition. (B) Same asin A for the t1/2. (C,D) Same as in A,Bfor neurons co-transfected withGABAB1–mRFP and EGFP,dynamitin (p50–EGFP) or N5-bIII-spectrin (N5-bIII-EGFP). n513–26neurons. Four independentexperiments were performed percondition. (E) Quantification ofGABAB1–mRFP fluorescenceintensities in axons and dendrites ofneurons transfected with controlEGFP or p50–EGFP. n516 neuronsper condition. (F) Polarity index ofGABAB1–mRFP for neuronstransfected with EGFP or p50–EGFP.n516 neurons per condition. Allquantitative data show themean6s.e.m.; *P,0.05; ***P,0.001;n.s., non-significant; Student’s t-test.(G,H) Representative images ofneurons co-transfected withGABAB1–mRFP (GB1–mRFP, red)and a double dose of control EGFPplasmid (EGFP/EGFP, green) (G) orp50–EGFP plus EGFP (p50–EGFP/EGFP, green) (H) and labeled with thedendritic marker MAP2 (purple).Dendritic segments outlined in whiteare enlarged below the main image.Arrowheads mark the axon. Scalebars: 10 mm.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3388
Jour
nal o
f Cel
l Sci
ence
retention domain, does not need GABAB2 for ER export and isefficiently transported to the plasma membrane (Restituitoet al., 2005). Unexpectedly, even under basal conditions, the
colocalization of this mutant subunit with GOPs was 70% higherthan that of full-length GABAB1 (percentage colocalizationwith Golgi–EYFP for GABAB1–Myc+GABAB2, 2468%; forGABAB1-DC–Myc, 6668%; P50.0002, Student’s t-test)(Fig. 6C–E). These results indicate that dendritic ER transportdoes not restrict the sorting of GABABRs to GOPs.
To further explore the ER-to-Golgi trafficking of GABABRs,we used a recently described pulse-chase system based on thefusion of four repeats of mutant FKBP12 (each repeat is referredto as an FM domain) to a transmembrane protein of interest. This
fusion results in ER retention due to aggregation of the FMdomains (Al-Bassam et al., 2012). Addition of the rapamycinanalog D/D solubilizer allows disaggregation of the FM domains,
release from the ER and visualization of a synchronous pool oftrafficking cargo. We engineered the FM domains at the N-terminus of GABAB1-DC (FM–GABAB1-DC–EGFP), which isexposed to the ER lumen during trafficking (Fig. 7A). Afteraddition of D/D solubilizer, FM–GABAB1-DC–EGFP wasefficiently released from the ER and reached the plasma
membrane (data not shown). Importantly, in neurons, FM–GABAB1-DC–EGFP was released from the ER after the additionof D/D solubilizer and subsequently condensed in the somaticGolgi and in GOPs (Fig. 7B). Individual carriers of ER-released
GABAB1-DC–EGFP traveled bidirectionally in dendrites with amild retrograde bias and merged with stationary GOPs that wereat variable distances from their site of appearance (the percentage
of carriers that merged with GOPs that traveled ,2.5 mm or.2.5 mm was 60% and 40%, respectively. Retrograde transport,70%; anterograde transport, 30%; six neurons, three independent
experiments) (Fig. 7C). Instantaneous velocities of post-ERcarriers were in the range of those reported for carriers of theERGIC (Appenzeller-Herzog and Hauri, 2006) (Fig. 7D). Taken
together, these results indicate that GABAB1 traffics through boththe perinuclear Golgi and GOPs, and that their use is notdetermined by ER residency times.
Finally, we speculated that the use of dendritic organelles
might influence the site of delivery at the neuronal plasmamembrane. To spatially characterize the insertion of newlysynthesized GABABRs, the kinetics of de novo insertion were
evaluated by quantifying the appearance of plasma membranebgtx-Myc–GABAB1a, a modified version of the GABAB1a subunit(one of the two alternative isoforms of GABAB1) containing an a-bungarotoxin-binding site engineered at the extracellular N-terminus. De novo exocytosed bgtx-Myc–GABAB1a subunitswere specifically and irreversibly labeled with fluorescent a-bungarotoxin (Bgtx-594) after saturating the recycling pool(Bgtx-646) as reported previously (Wilkins et al., 2008;Terunuma et al., 2010). We measured the mean intensity ofBgtx-594 at the outline of the soma or dendrite to estimate the
Fig. 6. GABABRs traffic throughsatellite GOPs. (A) Neurons were co-transfected with GABAB1-Myc (GB1–MYC, red) and Golgi–EYFP (green), orwith GABAB1–Myc (red), Golgi–EYFP(green) and ARF1-Q71I–HA (purple) toblock exit from the Golgi. GABAB2–FLAGis not shown. (B) Same as in A forproximal dendrites and GOPs. (C) Somasof neurons co-transfected with GABAB1-DC–Myc (GB1-DC–MYC, red) and Golgi–EYFP (green). (D) Same as in C forproximal dendrites and GOPs.(E) Percentage of colocalization ofGABAB1–Myc with Golgi–EYFP in theabsence (white bars) or presence ofARF1-Q71I–HA (black bars) or GABAB1-DC–Myc (gray bars) in the soma (left) anddendrites (right). Data show themean6s.e.m.; n54–8 neurons; **P,0.01;***P,0.001; Student’s t-test. Scale bars:10 mm.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3389
Jour
nal o
f Cel
l Sci
ence
number of newly inserted receptors. In neurons transfected with
bgtx-Myc–GABAB1a and GABAB2–EGFP, de novo insertion wasevident after 15 min of Bgtx-594 labeling, and graduallyincreased over 60 min (Fig. 8A). The timecourses of de novo
insertion of bgtx-Myc–GABAB1a in the soma and in dendriteswere remarkably similar (Fig. 8B). Indeed, the ratio ofsomatic:dendritic insertion revealed only a mild positive slope
(Fig. 8C). Overall these data do not support an early exocytosisevent in the cell body followed by subsequent diffusion andredistribution to dendrites (Rosenberg et al., 2001), but rather the
simultaneous delivery of newly synthesized receptors to the entiresomatodendritic compartment. These data are in agreement withthe use of the entire ensemble of exocytic organelles.
DISCUSSIONThe large size and morphological complexity of neuronsintroduce significant challenges to the delivery of membrane
cargoes, which might be difficult to convey exclusively throughlong-haul post-Golgi vesicles. Thus, local trafficking involvingthe dendritic ER and GOPs provides an alternative means to
regulate the supply of membrane components, but its role in thedelivery of neuronal proteins is still poorly understood (Hortonand Ehlers, 2003). Here, we demonstrate that GABAB1 transport
in dendrites is independent of COPI vesicle formation and takes
place within the dendritic ER. The transport of ER-associatedGABAB1 is not controlled by diffusion alone, but also by anactive mechanism involving microtubules and dynein. GABABRs
traffic through both the perinuclear Golgi and GOPs, and insertat the plasma membrane throughout the somatodendriticcompartment. We conclude that GABABRs utilize a long-
distance ER transport mechanism and both perinuclear andlocal secretory routes prior to reaching the plasma membrane.Our observations argue against the view that dendritic ER
transport restricts the trafficking of neurotransmitter receptors toGOPs, and they are consistent with the hypothesis that ERassembly of GABABRs is the rate-limiting step for receptortrafficking.
ER localization of GABAB1 in dendritesWe demonstrate that the ER retention of GABAB1 produces a
broad intracellular ER-enriched distribution in dendrites, with asmall proportion localizing at the ERGIC and Golgi, possiblyrevealing an ER–ERGIC–Golgi retrieval mechanism (Ramı́rez
et al., 2009; Biermann et al., 2010; Brock et al., 2005). Otherneurotransmitter receptors and ion channels are also enriched inthe ER and display a variety of ER retention signals (Ramı́rez and
Fig. 7. GABAB1 carriers merge with GOPs after ER export. (A) Diagram of the FM-based pulse-chase system implemented for a GABAB1 construct withEGFP fused to the C-terminus and four FM domains fused to the N-terminus (FM4x4). (B) Neurons were transfected with FM4-GB1-DC–EGFP (green) and fixedbefore or after a 60–120-min treatment with D/D solubilizer (DD). The Golgi was stained with GM130 (red). Arrows point to Golgi structures that show littlecolocalization before the addition of D/D solubilizer (0 min), but precise colocalization after incubation with D/D solubilizer (60–120 min). Scale bar: 10 mm.(C) Neurons were co-transfected with FM4-GB1-DC–EGFP (green) and Galt2–mCherry (red) to visualize GOPs in dendrites (upper panels). After 12 h ofexpression, protein synthesis was inhibited with cycloheximide and neurons were incubated with D/D solubilizer for 30 min at 37˚C and imaged live. Lowerpanels show a time series of the boxed dendritic segment. Arrows indicate moving carriers that merge with GOPs. Arrowheads show a static GABAB1 structure.Scale bar: 5 mm. (D) Diagram of the trajectory of the moving carrier visualized in C. The graph shows velocities in each interval.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3390
Jour
nal o
f Cel
l Sci
ence
Couve, 2011) but it is still unclear to what extent ER retention is
related to local dendritic checkpoints and how these regulate latertrafficking stages.
ER cargoes in neurons might be confined by regions of high
network complexity, especially those at dendritic branching points(Cui-Wang et al., 2012). However, the continuity of the ER allowsproteins to disperse throughout the organelle. In our experiments,the time of expression was sufficient to distribute GABAB1 through
regions of low and high complexity and to localize the subunitintracellularly within the entire somatodendritic compartment. Thisphenomenon was not an artifact of overexpression, because the
colocalization with the ER in dendrites was similar in transgenicneurons that expressed GABAB1–EGFP at endogenous levels andincreased after blocking ER export. By contrast, NMDARs
accumulate exclusively in the soma after blockade of ER export,providing evidence that multiple ER subcompartments control thetrafficking of different neurotransmitter receptors (Jeyifous et al.,2009).
ER transport in dendritesThe intracellular velocities of GABAB1 are comparable to those
reported for ER resident proteins (GFP–KDEL, RFP–KDEL, GFP–SERCA2a and GFP–IP3R1) (Bannai et al., 2004). Although diffusionhas been elegantly characterized for other neurotransmitter receptors
– namely, ER-retained AMPA subunits (Cui-Wang et al., 2012) –lateral diffusion alone is not sufficient to explain the ER transport of
GABAB1 in neurons. Three pieces of evidence strongly support this
conclusion: the microtubule-dependent transport of GABAB1, themicrotubule dependence even after BFA-induced ER retention, andthe dynein-dependent transport and localization of ER-associated
GABAB1.Regarding the underlying transport mechanism, our results show
that a component of ER-associated GABAB1 transport is impairedafter depolymerization of microtubules. However, our data do not
support the idea that microtubule-dependent ER remodelingcontrols the mobility of GABAB1, because the transport of ER–RFP and Cytb5–GFP were unchanged by nocodazole. Others have
proposed that vesicular-like carriers mediate a motor-assistedcomponent of ER transport in dendrites, potentially identifying anactive mechanism (Bannai et al., 2004; Jeyifous et al., 2009). With
our available data we cannot conclude whether the mobility ofGABAB1 is mediated by a vesicular-like component or by transportalong the continuous ER network. Although it is unusual toenvisage a motor dragging cargo within the plane of the bilayer, we
favor this view, and we speculate that the diffusion ofneurotransmitter receptors within the plane of the ER membraneprovides a basal dynamics that might be suitable for modification
by molecular motors. Supporting this notion, microtubule andmotor-driven motility of ERES occurs independently of themovement of ER membranes (Gupta et al., 2008).
We also speculate that the active component of ER-retainedGABAB1 introduces an additional mechanism for the confinement
Fig. 8. GABAB1 insertion is somatodendritic. (A) Neurons were co-transfected with bgtx-Myc–GABAB1a (GB1-Bgtx) and GABAB2–EGFP (GB2–EGFP,green). The population of recycling receptors was saturated with Bgtx-646 (blue). De-novo-inserted receptors were visualized by binding to Bgtx-594 (red)between 0 and 60 min. Scale bar: 20 mm. (B) The intensity of red fluorescence (de novo insertion) was measured at the somas (black) and proximal dendrites(white) and plotted against time. A.U., arbitrary units. n53 independent experiments; data show the mean6s.e.m.; 17–26 neurons; *P,0.05; ***P,0.001;Student’s t-test. (C) The ratio of somatic versus dendritic intensity was calculated from the data in B.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3391
Jour
nal o
f Cel
l Sci
ence
of ER cargoes that might counteract the dispersing tendency ofdiffusion. As a matter of fact, our data suggest that microtubule
acetylation increases the recruitment of a higher proportion offaster molecular motors that decrease the confinement ofGABABRs.
Are canonical and local trafficking in dendritesmutually exclusive?Our data show that GABAB1 localizes to the dendritic ER evenafter co-transfection with GABAB2, that GABABRs accumulate
in GOPs and that they are delivered to the plasma membranethroughout the somatodendritic compartment. Thus, the evidencesuggests that a local secretory route operates for GABABRs.
Other receptors also employ local routes, but differences in howthese routes are utilized are already apparent. For instance,endogenous and overexpressed NMDARs accumulate exclusively
on GOPs upon expression of ARF1-Q71I–HA (Jeyifous et al.,2009), whereas GABABRs accumulate in the somatic Golgi andin GOPs. Although the intrinsic limitations of overexpression
should be considered, the differences between GABABRs andNMDARs suggest that distinct neurotransmitter receptors displaydifferent degrees of spatial selectivity for organelles.
ER export kinetics and lateral diffusion determine the spatial
range over which export occurs (Herpers and Rabouille, 2004;Cui-Wang et al., 2012). Although most of our findings areconsistent with this proposal, our results with GABAB1-DCindicate to the contrary that an improved ability to exit the ERincreases trafficking through GOPs. One interpretation of theseobservations is that post-ER carriers fuse indiscriminately with
the perinuclear Golgi and GOPs, which act in concert to increasethe trafficking capacity of the neuron. Interestingly, upon ERrelease, the temperature-sensitive VSVG mutant or BDNF use
both the perinuclear Golgi and GOPs (Horton and Ehlers, 2003).Our experiments using GABAB1 are consistent with this view ofnon-selective processing and suggest that, for a subset of neuronalcargoes, ER transport does not restrict sorting to GOPs and that
selected proteins use the complete ensemble of Golgi structures.In this context, the absence of dendritic Golgi in ,30% ofneurons (Horton and Ehlers, 2003; Horton et al., 2005) and the
presence of GOPs in only ,18% of mature dendrites (Hortonet al., 2005), in a scenario where all dendrites containneurotransmitter receptors, suggest that local trafficking cannot
be exclusive. However, metabotropic glutamate receptor(mGluR) activity confines ER transport (Cui-Wang et al., 2012)and enhances the formation of ERES containing NMDARs(Aridor et al., 2004). Therefore, it is still unknown how, or if, the
balance of canonical and local trafficking is modified by activity,and whether preferential use of local routes influences the localavailability of receptors at synapses in response to activity.
Local supply of GOPsThe dendritic distribution of the ER and GOPs suggests thatintermediate transport carriers might operate locally. Indeed,VSVG carriers that merge with GOPs have been reported (Horton
and Ehlers, 2003). Additionally, packets containing VSVGbecome concentrated at branching points, presumably GOPs,after local ER export (Chen et al., 2013). However, a relevant
issue that remains to be addressed is to what extent the local ERcontributes to supply nearby GOPs. We have observed post-ERcarriers merging with GOPs after both long and short trajectories,
suggesting that dendritic ER export is not necessarily coupled tolocal trafficking. Therefore, although our results indicate that the
dendritic ER is a crucial component for the transport of GABABsubunits, they do not support the notion that GOPs feed
exclusively on the local ER, presenting a more complexscenario of local trafficking in dendrites.
MATERIALS AND METHODSAnimalsAdult pregnant female Sprague-Dawley rats were purchased from the
Central Animal Facility at Universidad Católica de Chile and were
euthanized by asphyxia in a CO2 chamber according to the Guide for
Care and Use of Laboratory Animals of The National Research Council
(1996). GABAB1–EGFP mice were kindly provided by Bernhard Bettler
(University of Basel, Switzerland). They carry a bacterial artificial
chromosome containing GABAB1-EGFP in a homozygous knockout
background for GABAB1. The construct is expressed under control of
GABAB1 promoter elements (Schuler et al., 2001; Casanova et al., 2009).
CellsCOS-7 cells were transfected by electroporation (Couve et al., 1998).
Primary hippocampal neurons were cultured from embryonic day (E)18
rats or E18 GABAB1–EGFP transgenic mice (Goslin and Banker, 1991)
and transfected by using Ca2+ phosphate at 7–15 days in vitro (Valdés
et al., 2012; Jiang and Chen, 2006). Endogenous subunits do not
assemble with recombinant subunits until 4–5 days post-transfection
(Valdés et al., 2012); therefore, all experiments were analyzed between
12 and 48 h post-transfection.
Reagents and DNA plasmidsBFA, cycloheximide, cytochalasin D, TSA and nocodazole were
purchased from Sigma (St Louis, MO). D/D Solubilizer was purchased
from Clontech (Mountain View, CA). Myc–GABAB1, FLAG–GABAB2,
HA–GABAB2, Myc–GABAB1-DC, GABAB1a–EGFP, GABAB2–EGFP,GABAB1a–mRFP and Bgtx-Myc–GABAB1 were as described previously
(Couve et al., 1998; Calver et al., 2001; Couve et al., 2002; Restituito
et al., 2005; Ramı́rez et al., 2009; Terunuma et al., 2010). To generate
GABAB1a–mEos2, rat GABAB1a was subcloned into pmEos2-N1
(between the SpeI and BamHI sites). mEos2, kindly provided by Sean
McKinney (Howard Hughes Medical Institute, Janelia Farm Research
Campus, MD), is fused to the intracellular carboxyl terminus of
GABAB1a. To generate FM–GABAB1-DC–EGFP, four FM domainswere fused to the N-terminus of GABAB1-DC by amplifying GABAB1-DC using the primers 59-ATCTCGAGGAGCAGAAGCTGATCTCAG-39 and 39-CTTTTGTCCTAGTAGGTGGTCTTAAGCG-59, andsubcloning the resulting amplicon into the Type I FM4-EGFP vector
(Thuenauer et al., 2004) between the XhoI and EcoRI sites.
The GABAA subunitsRFP-BBSa4 and pH/BBSb3 have been described
previously (Abramian et al., 2010; Bogdanov et al., 2006). pDsRed-C1
(RFP), pEYFP-Golgi, pEYFP-ER and pDsRed2-ER (ER-RFP) were
obtained from Clontech (Mountain View, CA). Galt2–mCherry was
kindly provided by Alfredo Cáceres (Instituto de Investigación Médica
Mercedes y Martı́n Ferreyra, Argentina). Cytb5–EGFP was kindly
provided by Claudio Hetz (Universidad de Chile, Chile). BFP–Sec61b(ER–BFP), ARF1-Q71I–HA and Sar1-H79G–HA have been described
previously (Ward et al., 2001; Jeyifous et al., 2009; Zurek et al., 2011).
Kif5CDN–RFP corresponds to amino acids 678–955 of KIF5C
(accession number NM_001107730). This construct is also referred to
as DN2 (Falley et al., 2009). Dynamitin subunit p50–EGFP and N5-bIII-spectrin were kindly provided by Anna Akhmanova (Erasmus Medical
Center, The Netherlands) and Michael Stankewich (Yale University,
New Haven, CT), respectively (Devarajan et al., 1997; Splinter et al.,
2012). All manipulations and the fidelity of DNA constructs were
verified by sequencing.
AntibodiesAntibodies against Myc and ERGIC-53 were purchased from Sigma.
Antibodies against KDEL and GM130 were purchased from StressGen
(Ann Arbor, MI). Antibodies against HA were purchased from Roche
(Indianapolis, IN). Anti-GFP antibodies (ab6556) were purchased from
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3392
Jour
nal o
f Cel
l Sci
ence
Abcam (Cambridge, UK). Fluorescently labeled secondary antibodies
were purchased from Jackson Immuno Research Laboratories (West
Grove, PA).
Immunofluorescence, time-lapse microscopy and FRAPImmunofluorescence was performed in permeabilized neurons (Ramı́rez
et al., 2009). Fixed and live-cell imaging was performed on a spectral
confocal microscope Olympus FluoView FV1000 (Center Valley, PA)
with a UPLSAPO 606/1.35 objective and 36 digital zoom, in Tyrodemedium (124 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2,
30 mM D-glucose and 25 mM HEPES pH 7.4) in an equilibrated
microscopy suite at 23 C̊. For FRAP experiments, dendritic regions of
interest (ROIs) of 60660 pixels (small ROI) or 2006200 pixels (largeROI), ,15 mm away from the soma, were bleached for 2 s using thetornado mode with the argon 473-nm laser at 100% power for EGFP-
tagged proteins or with the argon 405-nm and 559-nm laser at 100%
power for mRFP-tagged proteins. Fluorescence recovery was measured
every 10 s at 20% laser power using KALMAN2 by imaging the entire
field. Boxes of 8 mm2 were selected within the photo-bleached ROI toquantify recovery using ImageJ. Individual FRAPs were fitted to a first-
order exponential decay curve using established criteria, except for those
for Cytb5–EGFP, which were best fitted to an asymptotic type I curve
(Snapp et al., 2003), and selected by fitting (R2§0.95).The average intensity of the fluorescent signals in dendrites and axons
was determined using ImageJ (method modified from Kapitein et al.,
2010). Briefly, confocal images were acquired with a UPLSAPO 606/1.35 objective using sequential acquisition settings and KALMAN2 at
204862048 pixel resolution. Each image was a z-series of 5–10 imagesof which the maximum projection was used. Lines of width 1 pixel were
traced along axons and the major dendrite. Dendrites and axons were
identified based on their morphology and by immunostaining for MAP2.
Average pixel intensities were determined along the traced lines and the
background was subtracted. The average dendrite (Id) and axonal (Ia)
intensities were used to calculate the polarity index (PI) using
PI5(Id2Ia)/(Id+Ia).
ColocalizationColocalization analysis was performed as described previously (Ramı́rez
et al., 2010). All parameters were kept constant for a particular type of
experiment. Two-channel fluorescent image stacks [intensity I(x,y,z),
(0,255), voxel size Dx/Dy/Dz5137/137/250 nm] were recorded in themulti-track mode and KALMAN4. Four optical slices per neuron were
selected and deconvolved by Huygens Scripting software (Scientific
Volume Imaging, Hilversum, The Netherlands) using an algorithm based
on the Classic Maximum Likelihood Estimator. Image processing routines
for segmentation, visualization, calculation of colocalization coefficients
and statistical validation were based on Interactive Data Language (ITT,
Boulder, CO). Percentage colocalization for Manders coefficients (M1 and
M2) was calculated using the CDA (confined displacement algorithm)
plugin for ImageJ, followed by interpolation of the Manders coefficients
using the formula: Percentage colocalization5[M(0)2M(random)/M(100)2M(random)]6100, where M(0) is Manders at the original position,M(100) is the theoretical maximum (1), and M(random) is the
corresponding value for the random. Statistical validation was performed
using CDA.
sptPALMsptPALM was carried out by modifying a previously reported approach
(Frost et al., 2010). Briefly, imaging was conducted on an Olympus IX81
inverted microscope with a 1006/1.45 total internal reflectionfluorescence (TIRF) oil-immersion objective. Output from a set of
diode lasers (Coherent, Santa Clara, CA) was directed to the rear of the
microscope using a custom optical path. Molecules were simultaneously
photoconverted and excited using 405-nm (,100 mW) and 561-nm(20 mW) light through oblique illumination (near total internal
reflection) to reduce background fluorescence. A total of 10,000–
20,000 frames were collected at 5 Hz with 10-ms laser pulses to
minimize receptor motion during the frame (Frost et al., 2012).
Fluorescence was detected by an iXon+ 897 EM-CCD camera (Andor
Technology, South Windsor, CT) placed after a 1.66magnifying optic,resulting in a pixel size of 100 nm. Following acquisition, molecules
were localized by fitting a two-dimensional elliptical Gaussian function
to a 969 pixel array surrounding the peak, and locations were assembledinto tracks using available algorithms (Manley et al., 2008). Molecules
were segregated into those that were tracked over multiple frames and
those that appeared in a single frame. The distribution of tracked
molecules was calculated based on the density of molecules within a
search radius of 500 nm. Local net velocities were calculated using the
first and last points of tracks originating within this radius. Velocities and
vectors were mapped only in pixels with at least three tracks originating
within the search radius.
a-bungarotoxin labelingNeurons were co-transfected with bgtx-Myc–GABAB1a and GABAB2–
EGFP. At 48 h post-transfection, live neurons were incubated for
30 min with 13 mM (+)-tubocurarine chloride hydrate (Sigma). Theywere then incubated with a-bungarotoxin (Invitrogen, Eugene, OR)conjugated to Alexa FluorH 646 for 120 min at 37 C̊, washed with PBS
and incubated with a-bungarotoxin conjugated to Alexa FluorH 594 fordifferent periods of time at 37 C̊. Neurons were fixed and imaged on an
Olympus BX61WI upright microscope, an Olympus disk-scanning unit
and an UPlanFLN 606/1.25. Mean intensity was quantified usingImageJ in a 261-mm ROI at the outline of the somatic or dendritic(20 mm from the soma) regions to estimate the fluorescence associatedwith the plasma membrane.
AcknowledgementsWe thank Richard Saliba (Tufts University, Boston, USA), Viviana Valdés(Universidad Mayor, Santiago, Chile), José Cánovas and Marı́a A. Juricic (bothUniversidad de Chile, Santiago, Chile) for technical assistance and for criticalreading of the manuscript.
Competing interestsThe authors declare no competing interests.
Author contributionsA.C, J.I.V. and T.A.B. conceived and designed the experiments. J.I.V. and M.J-Bperformed the experiments and analyzed the data. J.I.V., A.C. and T.A.B.interpreted the data. H.L. and O.R. contributed image analysis routines. V.H.C.and D.S. generated reagents. J.I.V. and A.C. wrote the manuscript.
FundingThis work was supported by Iniciativa Cientı́fica Milenio [grant number P09-015-Fto A.C. and J.I.V.]; National Institutes of Health [grant number MH80046 to H.L.and T.A.B.]; Fondo Nacional de Desarrollo Cientı́fico y Tecnológico [grant number3110157 to O.R., 1100137 and 1140617 to A.C. and M.J.-B.]. D.S., J.I.V. andV.H.C. were supported by Comisión Nacional de Investigación Cientı́fica yTecnológica. Deposited in PMC for release after 12 months.
ReferencesAbramian, A. M., Comenencia-Ortiz, E., Vithlani, M., Tretter, E. V., Sieghart,W., Davies, P. A. and Moss, S. J. (2010). Protein kinase C phosphorylationregulates membrane insertion of GABAA receptor subtypes that mediate tonicinhibition. J. Biol. Chem. 285, 41795-41805.
Al-Bassam, S., Xu, M., Wandless, T. J. and Arnold, D. B. (2012). Differentialtrafficking of transport vesicles contributes to the localization of dendriticproteins. Cell Rep. 2, 89-100.
Appenzeller-Herzog, C. and Hauri, H. P. (2006). The ER-Golgi intermediatecompartment (ERGIC): in search of its identity and function. J. Cell Sci. 119,2173-2183.
Aridor, M., Guzik, A. K., Bielli, A. and Fish, K. N. (2004). Endoplasmic reticulumexport site formation and function in dendrites. J. Neurosci. 24, 3770-3776.
Bannai, H., Inoue, T., Nakayama, T., Hattori, M. and Mikoshiba, K. (2004).Kinesin dependent, rapid, bi-directional transport of ER sub-compartment indendrites of hippocampal neurons. J. Cell Sci. 117, 163-175.
Bettler, B., Kaupmann, K., Mosbacher, J. and Gassmann, M. (2004). Molecularstructure and physiological functions of GABA(B) receptors. Physiol. Rev. 84,835-867.
Biermann, B., Ivankova-Susankova, K., Bradaia, A., Abdel Aziz, S.,Besseyrias, V., Kapfhammer, J. P., Missler, M., Gassmann, M. and Bettler,B. (2010). The Sushi domains of GABAB receptors function as axonal targetingsignals. J. Neurosci. 30, 1385-1394.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3393
http://dx.doi.org/10.1074/jbc.M110.149229http://dx.doi.org/10.1074/jbc.M110.149229http://dx.doi.org/10.1074/jbc.M110.149229http://dx.doi.org/10.1074/jbc.M110.149229http://dx.doi.org/10.1016/j.celrep.2012.05.018http://dx.doi.org/10.1016/j.celrep.2012.05.018http://dx.doi.org/10.1016/j.celrep.2012.05.018http://dx.doi.org/10.1242/jcs.03019http://dx.doi.org/10.1242/jcs.03019http://dx.doi.org/10.1242/jcs.03019http://dx.doi.org/10.1523/JNEUROSCI.4775-03.2004http://dx.doi.org/10.1523/JNEUROSCI.4775-03.2004http://dx.doi.org/10.1242/jcs.00854http://dx.doi.org/10.1242/jcs.00854http://dx.doi.org/10.1242/jcs.00854http://dx.doi.org/10.1152/physrev.00036.2003http://dx.doi.org/10.1152/physrev.00036.2003http://dx.doi.org/10.1152/physrev.00036.2003http://dx.doi.org/10.1523/JNEUROSCI.3172-09.2010http://dx.doi.org/10.1523/JNEUROSCI.3172-09.2010http://dx.doi.org/10.1523/JNEUROSCI.3172-09.2010http://dx.doi.org/10.1523/JNEUROSCI.3172-09.2010
Jour
nal o
f Cel
l Sci
ence
Bogdanov, Y., Michels, G., Armstrong-Gold, C., Haydon, P. G., Lindstrom, J.,Pangalos, M. and Moss, S. J. (2006). Synaptic GABAA receptors are directlyrecruited from their extrasynaptic counterparts. EMBO J. 25, 4381-4389.
Broadwell, R. D. and Cataldo, A. M. (1983). The neuronal endoplasmicreticulum: its cytochemistry and contribution to the endomembrane system. I.Cell bodies and dendrites. J. Histochem. Cytochem. 31, 1077-1088.
Brock, C., Boudier, L., Maurel, D., Blahos, J. and Pin, J. P. (2005). Assembly-dependent surface targeting of the heterodimeric GABAB Receptor is controlledby COPI but not 14-3-3. Mol. Biol. Cell 16, 5572-5578.
Burkhardt, J. K., Echeverri, C. J., Nilsson, T. and Vallee, R. B. (1997).Overexpression of the dynamitin (p50) subunit of the dynactin complex disruptsdynein-dependent maintenance of membrane organelle distribution. J. Cell Biol.139, 469-484.
Cai, D., McEwen, D. P., Martens, J. R., Meyhofer, E. and Verhey, K. J. (2009).Single molecule imaging reveals differences in microtubule track selectionbetween Kinesin motors. PLoS Biol. 7, e1000216.
Calver, A. R., Robbins, M. J., Cosio, C., Rice, S. Q., Babbs, A. J., Hirst, W. D.,Boyfield, I., Wood, M. D., Russell, R. B., Price, G. W. et al. (2001). The C-terminal domains of the GABA(b) receptor subunits mediate intracellulartrafficking but are not required for receptor signaling. J. Neurosci. 21, 1203-1210.
Casanova, E., Guetg, N., Vigot, R., Seddik, R., Julio-Pieper, M., Hyland, N. P.,Cryan, J. F., Gassmann, M. and Bettler, B. (2009). A mouse model forvisualization of GABA(B) receptors. Genesis 47, 595-602.
Chen, D., Gibson, E. S. and Kennedy, M. J. (2013). A light-triggered proteinsecretion system. J. Cell Biol. 201, 631-640.
Clem, R. L. and Barth, A. (2006). Pathway-specific trafficking of native AMPARsby in vivo experience. Neuron 49, 663-670.
Couve, A., Filippov, A. K., Connolly, C. N., Bettler, B., Brown, D. A. and Moss,S. J. (1998). Intracellular retention of recombinant GABAB receptors. J. Biol.Chem. 273, 26361-26367.
Couve, A., Thomas, P., Calver, A. R., Hirst, W. D., Pangalos, M. N., Walsh, F. S.,Smart, T. G. and Moss, S. J. (2002). Cyclic AMP-dependent protein kinasephosphorylation facilitates GABA(B) receptor-effector coupling. Nat. Neurosci. 5,415-424.
Cui-Wang, T., Hanus, C., Cui, T., Helton, T., Bourne, J., Watson, D., Harris, K. M.and Ehlers, M. D. (2012). Local zones of endoplasmic reticulum complexityconfine cargo in neuronal dendrites. Cell 148, 309-321.
Devarajan, P., Stabach, P. R., De Matteis, M. A. and Morrow, J. S. (1997). Na,K-ATPase transport from endoplasmic reticulum to Golgi requires the Golgispectrin-ankyrin G119 skeleton in Madin Darby canine kidney cells. Proc. Natl.Acad. Sci. USA 94, 10711-10716.
Dompierre, J. P., Godin, J. D., Charrin, B. C., Cordelières, F. P., King, S. J.,Humbert, S. and Saudou, F. (2007). Histone deacetylase 6 inhibitioncompensates for the transport deficit in Huntington’s disease by increasingtubulin acetylation. J. Neurosci. 27, 3571-3583.
Echeverri, C. J., Paschal, B. M., Vaughan, K. T. and Vallee, R. B. (1996).Molecular characterization of the 50-kD subunit of dynactin reveals function forthe complex in chromosome alignment and spindle organization during mitosis.J. Cell Biol. 132, 617-633.
Ehlers, M. D. (2000). Reinsertion or degradation of AMPA receptors determinedby activity-dependent endocytic sorting. Neuron 28, 511-525.
Falley, K., Schütt, J., Iglauer, P., Menke, K., Maas, C., Kneussel, M., Kindler, S.,Wouters, F. S., Richter, D. and Kreienkamp, H. J. (2009). Shank1 mRNA:dendritic transport by kinesin and translational control by the 59untranslatedregion. Traffic 10, 844-857.
Friedman, J. R., Webster, B. M., Mastronarde, D. N., Verhey, K. J. and Voeltz,G. K. (2010). ER sliding dynamics and ER-mitochondrial contacts occur onacetylated microtubules. J. Cell Biol. 190, 363-375.
Frost, N. A., Shroff, H., Kong, H., Betzig, E. and Blanpied, T. A. (2010). Single-molecule discrimination of discrete perisynaptic and distributed sites of actinfilament assembly within dendritic spines. Neuron 67, 86-99.
Frost, N. A., Lu, H. E. and Blanpied, T. A. (2012). Optimization of cell morphologymeasurement via single-molecule tracking PALM. PLoS ONE 7, e36751.
Gonchar, Y., Pang, L., Malitschek, B., Bettler, B. and Burkhalter, A. (2001).Subcellular localization of GABA(B) receptor subunits in rat visual cortex.J. Comp. Neurol. 431, 182-197.
Goslin, K. and Banker, G. (1991). Rat hippocampal neurons in low-densitycultures. In Culturing Nerve Cells (ed. G. Banker and K. Goslin), pp. 251-281.Cambridge: MA, MIT.
Grampp, T., Sauter, K., Markovic, B. and Benke, D. (2007). Gamma-aminobutyric acid type B receptors are constitutively internalized via theclathrin-dependent pathway and targeted to lysosomes for degradation. J. Biol.Chem. 282, 24157-24165.
Gupta, V., Palmer, K. J., Spence, P., Hudson, A. and Stephens, D. J. (2008).Kinesin-1 (uKHC/KIF5B) is required for bidirectional motility of ER exit sites andefficient ER-to-Golgi transport. Traffic 9, 1850-1866.
Herpers, B. and Rabouille, C. (2004). mRNA localization and ER-based proteinsorting mechanisms dictate the use of transitional endoplasmic reticulum-golgiunits involved in gurken transport in Drosophila oocytes. Mol. Biol. Cell 15,5306-5317.
Horton, A. C. and Ehlers, M. D. (2003). Dual modes of endoplasmic reticulum-to-Golgi transport in dendrites revealed by live-cell imaging. J. Neurosci. 23, 6188-6199.
Horton, A. C. and Ehlers, M. D. (2004). Secretory trafficking in neuronaldendrites. Nat. Cell. Biol. 6, 585-591.
Horton, A. C., Rácz, B., Monson, E. E., Lin, A. L., Weinberg, R. J. and Ehlers,M. D. (2005). Polarized secretory trafficking directs cargo for asymmetricdendrite growth and morphogenesis. Neuron 48, 757-771.
Jeyifous, O., Waites, C. L., Specht, C. G., Fujisawa, S., Schubert, M., Lin, E. I.,Marshall, J., Aoki, C., de Silva, T., Montgomery, J. M. et al. (2009). SAP97and CASK mediate sorting of NMDA receptors through a previously unknownsecretory pathway. Nat. Neurosci. 12, 1011-1019.
Jiang, M. and Chen, G. (2006). High Ca2+-phosphate transfection efficiency inlow-density neuronal cultures. Nat. Protoc. 1, 695-700.
Kapitein, L. C., Schlager, M. A., Kuijpers, M., Wulf, P. S., van Spronsen, M.,MacKintosh, F. C. and Hoogenraad, C. C. (2010). Mixed microtubules steerdynein-driven cargo transport into dendrites. Curr. Biol. 20, 290-299.
Kucharz, K., Krogh, M., Ng, A. N. and Toresson, H. (2009). NMDA receptorstimulation induces reversible fission of the neuronal endoplasmic reticulum.PLoS ONE 4, e5250.
Manley, S., Gillette, J. M., Patterson, G. H., Shroff, H., Hess, H. F., Betzig, E. andLippincott-Schwartz, J. (2008). High-density mapping of single-moleculetrajectories with photoactivated localization microscopy. Nat. Methods 5, 155-157.
Margeta-Mitrovic, M., Jan, Y. N. and Jan, L. Y. (2000). A trafficking checkpointcontrols GABA(B) receptor heterodimerization. Neuron 27, 97-106.
McKinney, S. A., Murphy, C. S., Hazelwood, K. L., Davidson, M. W. andLooger, L. L. (2009). A bright and photostable photoconvertible fluorescentprotein. Nat. Methods 6, 131-133.
Pooler, A. M., Gray, A. G. and McIlhinney, R. A. (2009). Identification of a novelregion of the GABA(B2) C-terminus that regulates surface expression andneuronal targeting of the GABA(B) receptor. Eur. J. Neurosci. 29, 869-878.
Presley, J. F., Cole, N. B., Schroer, T. A., Hirschberg, K., Zaal, K. J. andLippincott-Schwartz, J. (1997). ER-to-Golgi transport visualized in living cells.Nature 389, 81-85.
Ramı́rez, O. A. and Couve, A. (2011). The endoplasmic reticulum and proteintrafficking in dendrites and axons. Trends Cell Biol. 21, 219-227.
Ramı́rez, O. A., Vidal, R. L., Tello, J. A., Vargas, K. J., Kindler, S., Härtel, S. andCouve, A. (2009). Dendritic assembly of heteromeric gamma-aminobutyric acidtype B receptor subunits in hippocampal neurons. J. Biol. Chem. 284, 13077-13085.
Ramı́rez, O., Garcı́a, A., Rojas, R., Couve, A. and Härtel, S. (2010). Confineddisplacement algorithm determines true and random colocalization influorescence microscopy. J. Microsc. 239, 173-183.
Ramı́rez, O. A., Härtel, S. and Couve, A. (2011). Location matters: theendoplasmic reticulum and protein trafficking in dendrites. Biol. Res. 44, 17-23.
Reed, N. A., Cai, D., Blasius, T. L., Jih, G. T., Meyhofer, E., Gaertig, J. andVerhey, K. J. (2006). Microtubule acetylation promotes kinesin-1 binding andtransport. Curr. Biol. 16, 2166-2172.
Restituito, S., Couve, A., Bawagan, H., Jourdain, S., Pangalos, M. N., Calver,A. R., Freeman, K. B. and Moss, S. J. (2005). Multiple motifs regulate thetrafficking of GABA(B) receptors at distinct checkpoints within the secretorypathway. Mol. Cell. Neurosci. 28, 747-756.
Rosenberg, M., Meier, J., Triller, A. and Vannier, C. (2001). Dynamics of glycinereceptor insertion in the neuronal plasma membrane. J. Neurosci. 21, 5036-5044.
Saxton, M. J. and Jacobson, K. (1997). Single-particle tracking: applications tomembrane dynamics. Annu. Rev. Biophys. Biomol. Struct. 26, 373-399.
Schuler, V., Lüscher, C., Blanchet, C., Klix, N., Sansig, G., Klebs, K., Schmutz,M., Heid, J., Gentry, C., Urban, L. et al. (2001). Epilepsy, hyperalgesia,impaired memory, and loss of pre- and postsynaptic GABA(B) responses inmice lacking GABA(B(1)). Neuron 31, 47-58.
Sengupta, P., Van Engelenburg, S. and Lippincott-Schwartz, J. (2012).Visualizing cell structure and function with point-localization superresolutionimaging. Dev. Cell 23, 1092-1102.
Snapp, E. L., Altan, N. and Lippincott-Schwartz, J. (2003). Measuring proteinmobility by photobleaching GFP chimeras in living cells. Curr. Protoc. Cell Biol.19, 21.1.1-21.1.24.
Spacek, J. and Harris, K. M. (1997). Three-dimensional organization of smoothendoplasmic reticulum in hippocampal CA1 dendrites and dendritic spines of theimmature and mature rat. J. Neurosci. 17, 190-203.
Splinter, D., Razafsky, D. S., Schlager, M. A., Serra-Marques, A., Grigoriev, I.,Demmers, J., Keijzer, N., Jiang, K., Poser, I., Hyman, A. A. et al. (2012).BICD2, dynactin, and LIS1 cooperate in regulating dynein recruitment to cellularstructures. Mol. Biol. Cell 23, 4226-4241.
Terunuma, M., Vargas, K. J., Wilkins, M. E., Ramı́rez, O. A., Jaureguiberry-Bravo, M., Pangalos, M. N., Smart, T. G., Moss, S. J. and Couve, A. (2010).Prolonged activation of NMDA receptors promotes dephosphorylation and alterspostendocytic sorting of GABAB receptors. Proc. Natl. Acad. Sci. USA 107,13918-13923.
Thuenauer, R., Hsu, Y. C., Carvajal-Gonzalez, J. M., Deborde, S., Chuang,J. Z., Römer, W., Sonnleitner, A., Rodriguez-Boulan, E. and Sung, C. H.(2014). Four-dimensional live imaging of apical biosynthetic trafficking reveals apost-Golgi sorting role of apical endosomal intermediates. Proc. Natl. Acad. Sci.USA 111, 4127-4132.
Valdés, V., Valenzuela, J. I., Salas, D. A., Jaureguiberry-Bravo, M., Otero, C.,Thiede, C., Schmidt, C. F. and Couve, A. (2012). Endoplasmic reticulumsorting and kinesin-1 command the targeting of axonal GABAB receptors. PLoSONE 7, e44168.
Vargas, K. J., Terunuma, M., Tello, J. A., Pangalos, M. N., Moss, S. J. andCouve, A. (2008). The availability of surface GABA B receptors is independent
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3394
http://dx.doi.org/10.1038/sj.emboj.7601309http://dx.doi.org/10.1038/sj.emboj.7601309http://dx.doi.org/10.1038/sj.emboj.7601309http://dx.doi.org/10.1177/31.9.6309951http://dx.doi.org/10.1177/31.9.6309951http://dx.doi.org/10.1177/31.9.6309951http://dx.doi.org/10.1091/mbc.E05-05-0400http://dx.doi.org/10.1091/mbc.E05-05-0400http://dx.doi.org/10.1091/mbc.E05-05-0400http://dx.doi.org/10.1083/jcb.139.2.469http://dx.doi.org/10.1083/jcb.139.2.469http://dx.doi.org/10.1083/jcb.139.2.469http://dx.doi.org/10.1083/jcb.139.2.469http://dx.doi.org/10.1371/journal.pbio.1000216http://dx.doi.org/10.1371/journal.pbio.1000216http://dx.doi.org/10.1371/journal.pbio.1000216http://dx.doi.org/10.1002/dvg.20535http://dx.doi.org/10.1002/dvg.20535http://dx.doi.org/10.1002/dvg.20535http://dx.doi.org/10.1083/jcb.201210119http://dx.doi.org/10.1083/jcb.201210119http://dx.doi.org/10.1016/j.neuron.2006.01.019http://dx.doi.org/10.1016/j.neuron.2006.01.019http://dx.doi.org/10.1074/jbc.273.41.26361http://dx.doi.org/10.1074/jbc.273.41.26361http://dx.doi.org/10.1074/jbc.273.41.26361http://dx.doi.org/10.1016/j.cell.2011.11.056http://dx.doi.org/10.1016/j.cell.2011.11.056http://dx.doi.org/10.1016/j.cell.2011.11.056http://dx.doi.org/10.1073/pnas.94.20.10711http://dx.doi.org/10.1073/pnas.94.20.10711http://dx.doi.org/10.1073/pnas.94.20.10711http://dx.doi.org/10.1073/pnas.94.20.10711http://dx.doi.org/10.1523/JNEUROSCI.0037-07.2007http://dx.doi.org/10.1523/JNEUROSCI.0037-07.2007http://dx.doi.org/10.1523/JNEUROSCI.0037-07.2007http://dx.doi.org/10.1523/JNEUROSCI.0037-07.2007http://dx.doi.org/10.1083/jcb.132.4.617http://dx.doi.org/10.1083/jcb.132.4.617http://dx.doi.org/10.1083/jcb.132.4.617http://dx.doi.org/10.1083/jcb.132.4.617http://dx.doi.org/10.1016/S0896-6273(00)00129-Xhttp://dx.doi.org/10.1016/S0896-6273(00)00129-Xhttp://dx.doi.org/10.1111/j.1600-0854.2009.00912.xhttp://dx.doi.org/10.1111/j.1600-0854.2009.00912.xhttp://dx.doi.org/10.1111/j.1600-0854.2009.00912.xhttp://dx.doi.org/10.1111/j.1600-0854.2009.00912.xhttp://dx.doi.org/10.1083/jcb.200911024http://dx.doi.org/10.1083/jcb.200911024http://dx.doi.org/10.1083/jcb.200911024http://dx.doi.org/10.1016/j.neuron.2010.05.026http://dx.doi.org/10.1016/j.neuron.2010.05.026http://dx.doi.org/10.1016/j.neuron.2010.05.026http://dx.doi.org/10.1371/journal.pone.0036751http://dx.doi.org/10.1371/journal.pone.0036751http://dx.doi.org/10.1002/1096-9861(20010305)431:23.0.CO;2-Khttp://dx.doi.org/10.1002/1096-9861(20010305)431:23.0.CO;2-Khttp://dx.doi.org/10.1002/1096-9861(20010305)431:23.0.CO;2-Khttp://dx.doi.org/10.1074/jbc.M702626200http://dx.doi.org/10.1074/jbc.M702626200http://dx.doi.org/10.1074/jbc.M702626200http://dx.doi.org/10.1074/jbc.M702626200http://dx.doi.org/10.1111/j.1600-0854.2008.00811.xhttp://dx.doi.org/10.1111/j.1600-0854.2008.00811.xhttp://dx.doi.org/10.1111/j.1600-0854.2008.00811.xhttp://dx.doi.org/10.1091/mbc.E04-05-0398http://dx.doi.org/10.1091/mbc.E04-05-0398http://dx.doi.org/10.1091/mbc.E04-05-0398http://dx.doi.org/10.1091/mbc.E04-05-0398http://dx.doi.org/10.1016/j.neuron.2005.11.005http://dx.doi.org/10.1016/j.neuron.2005.11.005http://dx.doi.org/10.1016/j.neuron.2005.11.005http://dx.doi.org/10.1038/nn.2362http://dx.doi.org/10.1038/nn.2362http://dx.doi.org/10.1038/nn.2362http://dx.doi.org/10.1038/nn.2362http://dx.doi.org/10.1038/nprot.2006.86http://dx.doi.org/10.1038/nprot.2006.86http://dx.doi.org/10.1016/j.cub.2009.12.052http://dx.doi.org/10.1016/j.cub.2009.12.052http://dx.doi.org/10.1016/j.cub.2009.12.052http://dx.doi.org/10.1371/journal.pone.0005250http://dx.doi.org/10.1371/journal.pone.0005250http://dx.doi.org/10.1371/journal.pone.0005250http://dx.doi.org/10.1038/nmeth.1176http://dx.doi.org/10.1038/nmeth.1176http://dx.doi.org/10.1038/nmeth.1176http://dx.doi.org/10.1016/S0896-6273(00)00012-Xhttp://dx.doi.org/10.1016/S0896-6273(00)00012-Xhttp://dx.doi.org/10.1038/nmeth.1296http://dx.doi.org/10.1038/nmeth.1296http://dx.doi.org/10.1038/nmeth.1296http://dx.doi.org/10.1111/j.1460-9568.2009.06636.xhttp://dx.doi.org/10.1111/j.1460-9568.2009.06636.xhttp://dx.doi.org/10.1111/j.1460-9568.2009.06636.xhttp://dx.doi.org/10.1038/38891http://dx.doi.org/10.1038/38891http://dx.doi.org/10.1038/38891http://dx.doi.org/10.1016/j.tcb.2010.12.003http://dx.doi.org/10.1016/j.tcb.2010.12.003http://dx.doi.org/10.1074/jbc.M900575200http://dx.doi.org/10.1074/jbc.M900575200http://dx.doi.org/10.1074/jbc.M900575200http://dx.doi.org/10.1074/jbc.M900575200http://dx.doi.org/10.1111/j.1365-2818.2010.03369.xhttp://dx.doi.org/10.1111/j.1365-2818.2010.03369.xhttp://dx.doi.org/10.1111/j.1365-2818.2010.03369.xhttp://dx.doi.org/10.4067/S0716-97602011000100003http://dx.doi.org/10.4067/S0716-97602011000100003http://dx.doi.org/10.1016/j.cub.2006.09.014http://dx.doi.org/10.1016/j.cub.2006.09.014http://dx.doi.org/10.1016/j.cub.2006.09.014http://dx.doi.org/10.1016/j.mcn.2004.12.006http://dx.doi.org/10.1016/j.mcn.2004.12.006http://dx.doi.org/10.1016/j.mcn.2004.12.006http://dx.doi.org/10.1016/j.mcn.2004.12.006http://dx.doi.org/10.1146/annurev.biophys.26.1.373http://dx.doi.org/10.1146/annurev.biophys.26.1.373http://dx.doi.org/10.1016/S0896-6273(01)00345-2http://dx.doi.org/10.1016/S0896-6273(01)00345-2http://dx.doi.org/10.1016/S0896-6273(01)00345-2http://dx.doi.org/10.1016/S0896-6273(01)00345-2http://dx.doi.org/10.1016/j.devcel.2012.09.022http://dx.doi.org/10.1016/j.devcel.2012.09.022http://dx.doi.org/10.1016/j.devcel.2012.09.022http://dx.doi.org/10.1091/mbc.E12-03-0210http://dx.doi.org/10.1091/mbc.E12-03-0210http://dx.doi.org/10.1091/mbc.E12-03-0210http://dx.doi.org/10.1091/mbc.E12-03-0210http://dx.doi.org/10.1073/pnas.1000853107http://dx.doi.org/10.1073/pnas.1000853107http://dx.doi.org/10.1073/pnas.1000853107http://dx.doi.org/10.1073/pnas.1000853107http://dx.doi.org/10.1073/pnas.1000853107http://dx.doi.org/10.1073/pnas.1304168111http://dx.doi.org/10.1073/pnas.1304168111http://dx.doi.org/10.1073/pnas.1304168111http://dx.doi.org/10.1073/pnas.1304168111http://dx.doi.org/10.1073/pnas.1304168111http://dx.doi.org/10.1371/journal.pone.0044168http://dx.doi.org/10.1371/journal.pone.0044168http://dx.doi.org/10.1371/journal.pone.0044168http://dx.doi.org/10.1371/journal.pone.0044168http://dx.doi.org/10.1074/jbc.M802419200http://dx.doi.org/10.1074/jbc.M802419200
Jour
nal o
f Cel
l Sci
ence
of gamma-aminobutyric acid but controlled by glutamate in central neurons.J. Biol. Chem. 283, 24641-24648.
Vidal, R. L., Ramı́rez, O. A., Sandoval, L., Koenig-Robert, R., Härtel, S. andCouve, A. (2007). Marlin-1 and conventional kinesin link GABAB receptors to thecytoskeleton and regulate receptor transport. Mol. Cell. Neurosci. 35, 501-512.
Ward, T. H., Polishchuk, R. S., Caplan, S., Hirschberg, K. and Lippincott-Schwartz, J. (2001). Maintenance of Golgi structure and function depends onthe integrity of ER export. J. Cell Biol. 155, 557-570.
Wilkins, M. E., Li, X. and Smart, T. G. (2008). Tracking cell surface GABAB receptorsusing an alpha-bungarotoxin tag. J. Biol. Chem. 283, 34745-34752.
Woźniak, M. J., Bola, B., Brownhill, K., Yang, Y. C., Levakova, V. and Allan,V. J. (2009). Role of kinesin-1 and cytoplasmic dynein in endoplasmic reticulummovement in VERO cells. J. Cell Sci. 122, 1979-1989.
Ye, B., Zhang, Y., Song, W., Younger, S. H., Jan, L. Y. and Jan, Y. N. (2007). Growingdendritesandaxonsdiffer in their relianceon thesecretory pathway.Cell130, 717-729.
Zhang, C. J., Rosenwald, A. G., Willingham, M. C., Skuntz, S., Clark, J. andKahn, R. A. (1994). Expression of a dominant allele of human ARF1 inhibitsmembrane traffic in vivo. J. Cell Biol. 124, 289-300.
Zurek, N., Sparks, L. and Voeltz, G. (2011). Reticulon short hairpintransmembrane domains are used to shape ER tubules. Traffic 12, 28-41.
RESEARCH ARTICLE Journal of Cell Science (2014) 127, 3382–3395 doi:10.1242/jcs.151092
3395
http://dx.doi.org/10.1074/jbc.M802419200http://dx.doi.org/10.1074/jbc.M802419200http://dx.doi.org/10.1016/j.mcn.2007.04.008http://dx.doi.org/10.1016/j.mcn.2007.04.008http://dx.doi.org/10.1016/j.mcn.2007.04.008http://dx.doi.org/10.1083/jcb.200107045http://dx.doi.org/10.1083/jcb.200107045http://dx.doi.org/10.1083/jcb.200107045http://dx.doi.org/10.1074/jbc.M803197200http://dx.doi.org/10.1074/jbc.M803197200http://dx.doi.org/10.1242/jcs.041962http://dx.doi.org/10.1242/jcs.041962http://dx.doi.org/10.1242/jcs.041962http://dx.doi.org/10.1016/j.cell.2007.06.032http://dx.doi.org/10.1016/j.cell.2007.06.032http://dx.doi.org/10.1083/jcb.124.3.289http://dx.doi.org/10.1083/jcb.124.3.289http://dx.doi.org/10.1083/jcb.124.3.289http://dx.doi.org/10.1111/j.1600-0854.2010.01134.xhttp://dx.doi.org/10.1111/j.1600-0854.2010.01134.x
Fig 1Fig 2Fig 3Fig 4Fig 5Fig 6Fig 7Fig 8Ref 1Ref 2Ref 3Ref 4Ref 5Ref 6Ref 7Ref 8Ref 9Ref 10Ref 11Ref 12Ref 13Ref 14Ref 15Ref 16Ref 17Ref 18Ref 19Ref 20Ref 21Ref 22Ref 23Ref 24Ref 25Ref 26Ref 27Ref 28Ref 29Ref 30Ref 31Ref 32Ref 33Ref 34Ref 35Ref 36Ref 37Ref 38Ref 39Ref 40Ref 41Ref 42Ref 43Ref 44Ref 45Ref 46Ref 47Ref 48Ref 49Ref 50Ref 51Ref 52Ref 53Ref 54Ref 55Ref 56Ref 57Ref 58Ref 59Ref 60Ref 61Ref 62Ref 63Ref 64Ref 65Ref 66Ref 67Ref 68
/ColorImageDict > /JPEG2000ColorACSImageDict > /JPEG2000ColorImageDict > /AntiAliasGrayImages false /CropGrayImages true /GrayImageMinResolution 150 /GrayImageMinResolutionPolicy /OK /DownsampleGrayImages true /GrayImageDownsampleType /Bicubic /GrayImageResolution 200 /GrayImageDepth 8 /GrayImageMinDownsampleDepth 2 /GrayImageDownsampleThreshold 1.50000 /EncodeGrayImages true /GrayImageFilter /FlateEncode /AutoFilterGrayImages false /GrayImageAutoFilterStrategy /JPEG /GrayACSImageDict > /GrayImageDict > /JPEG2000GrayACSImageDict > /JPEG2000GrayImageDict > /AntiAliasMonoImages false /CropMonoImages true /MonoImageMinResolution 1200 /MonoImageMinResolutionPolicy /OK /DownsampleMonoImages true /MonoImageDownsampleType /Bicubic /MonoImageResolution 600 /MonoImageDepth -1 /MonoImageDownsampleThreshold 1.50000 /EncodeMonoImages true /MonoImageFilter /CCITTFaxEncode /MonoImageDict > /AllowPSXObjects false /CheckCompliance [ /None ] /PDFX1aCheck false /PDFX3Check false /PDFXCompliantPDFOnly true /PDFXNoTrimBoxError false /PDFXTrimBoxToMediaBoxOffset [ 0.00000 0.00000 0.00000 0.00000 ] /PDFXSetBleedBoxToMediaBox false /PDFXBleedBoxToTrimBoxOffset [ 0.00000 0.00000 0.00000 0.00000 ] /PDFXOutputIntentProfile (Euroscale Coated v2) /PDFXOutputConditionIdentifier (FOGRA1) /PDFXOutputCondition () /PDFXRegistryName (http://www.color.org) /PDFXTrapped /False
/CreateJDFFile false /SyntheticBoldness 1.000000 /Description >>> setdistillerparams> setpagedevice