Embed Size (px)
Citation preview

Contents lists available at ScienceDirect
Journal of Proteomics
journal homepage: www.elsevier.com/locate/jprot
Serum metabolome profiling revealed potential biomarkers for milk proteinyield in dairy cows
Xuehui Wua, Huizeng Suna,b, Mingyuan Xuea, Diming Wanga, Le Luo Guanb,⁎, Jianxin Liua,⁎
a Institute of Dairy Science, MoE Key Laboratory of Molecular Animal Nutrition, College of Animal Sciences, Zhejiang University, Hangzhou 310058, ChinabDepartment of Agricultural, Food & Nutritional Science, University of Alberta, Edmonton T6G 2P5, Canada
A R T I C L E I N F O
Keywords:BiomarkersMilk protein yieldSerum metabolomicsHolstein dairy cow
A B S T R A C T
Milk yield (MY) and milk protein (MP) content are crucial milk performance traits of dairy cows that directlyaffect the dairy profits. This study first proposed milk protein yield (MPY) by considering MY and MP contenttogether. Forty multiparous cows were selected from the 348 Holstein dairy cows, which fed the same diet underthe same management condition, to investigate the serum metabolome profiles and to identify key metabolitesassociated with MPY. Among them, 20 cows with a higher MPY (MY > 34.5 kg/d and MP > 3.2%. i.e.,MPY > 1.11 kg/d) were defined as the HH group, and 20 cows with a lower MPY (MY < 31 kg/d andMP < 2.9%, i.e., MPY < 0.87 kg/d) as the LL group. The GC-TOF/MS and the ultra HPLC-MS/MS platformswere used to identify metabolites and quantify biomarkers, respectively. Orthogonal partial least squares dis-criminant analysis of serum metabolomes revealed a clear separation between the 2 groups. Thirty-six sig-nificantly different metabolites were identified, which mainly involved in valine, leucine and isoleucine bio-synthesis and glycine, serine and threonine metabolism. With biomarker analysis and validation, hippuric acid,nicotinamide and pelargonic acid may serve as key metabolites associated with MPY.Biological significance: This study reports the application of serum metabolomics to identify biomarkers related toMPY and to reveal the biological pathways affecting milk protein synthesis. Three novel serum biomarkers werediscovered to reflect the MPY variation of dairy cows, which may be useful in quality control in dairy cowproduction and for optimizing industrial production of dairy products. This study confirms that individualphysiological and metabolic differences contribute to the variations in MPY and provides directions for furtherimproving the MPY of dairy cows.
1. Introduction
Milk protein is a valuable nutritional component used to assess milkquality and is an important protein source for humans [1]. Milk proteincontent (MP, %) and milk yield (MY, kg/d) are two key economicaltraits for evaluating the lactation performance of dairy cows, but theyare negatively correlated [2]. To date, most studies have focused onenhancing MP or MY through nutritional strategies such as altering dietingredients [3], balancing amino acids (AAs) profiles in feed [4], andadding extra bio-available ingredients to feed [5]. Meanwhile, breedingand management strategies have also been applied to improve milkprotein production [6, 7]. However, the above researchers only focusedon either MP or MY, which could not meet the requirements for bothtraits. In this study, a new measure, milk protein yield (MPY, here de-fined as MP×MY) is introduced as a way to consider MY and MP at thesame time, which will assist in maximizing benefits for the dairy in-dustry.
It is known that even when animals are fed the same diet, they canhave divergent milk performance [8]. Many factors can affect suchvariation, including genetics, digestion and absorption processes, stressstatus, health, etc. [9]. We speculate that individual metabolic andphysiological variations can contribute to differences in MPY. Milkproduction and milk protein synthesis are complex biological processesthat involve many metabolic pathways. A comprehensive under-standing of their regulatory mechanisms is limited, which prevents itsfurther improvement through nutritional and management strategies.
Metabolomics, as a vital part of system biology, aims to investigaterelevant mechanisms by analyzing metabolic profiles of cells, biofluidsor tissues. Recent studies have revealed that blood metabolites used asphysiological biomarkers are able to reflect metabolic dysfunction,health and performance in vivo [10]. Our previous study showed thattwo key urine metabolites were associated with MY when cows con-sumed different quality forages [11], suggesting that metabolites couldserve as potential biomarkers for milk synthesis. Here, we further
https://doi.org/10.1016/j.jprot.2018.06.005Received 12 March 2018; Received in revised form 16 May 2018; Accepted 11 June 2018
⁎ Corresponding authors.E-mail addresses: [email protected] (L.L. Guan), [email protected] (J. Liu).
Journal of Proteomics 184 (2018) 54–61
Available online 18 June 20181874-3919/ © 2018 Published by Elsevier B.V.
T

hypothesized that serum metabolites of dairy cows with different MPYcould serve as potential biomarkers for milk production and milk pro-tein synthesis through their different metabolic functions.
In this study, to reflect the physiological mechanisms relating toMPY, we selected dairy cows with extremely high MPY and extremelylow MPY fed the same diet to characterize serum metabolome profiles.Moreover, biomarker analysis and quantitative validation were per-formed to identify key metabolites as potential biomarkers regardingMPY, which can provide high reliability and are feasible parameters forselection of high performance dairy cows.
2. Materials and methods
2.1. Animals and management
All the experimental protocols were approved by the Animal CareCommittee at the Zhejiang University (Hangzhou, P. R. China), and theanimal study was conducted according to the University's guidelines foranimal research. With high reliability of a power analysis(Supplementary Table S1), 40 multiparous Holstein dairy cows withdays in milk of 160.0 (± 28.7 SD) and parity of 3.3 (± 1.5 SD) wereselected from among 348 dairy cows housed at Hangjiang Dairy Farm(Hangzhou, China). They were kept in individually tethered stalls in abarn with good ventilation and were milked and fed 3 times daily at06:30, 14:00, and 20:00 h. The cows were fed an identical total mixedration with a concentrate-to-forage ratio of 57:43 (DM basis) and hadfree access to water.
2.2. Sample collection and measurement
Diets were sampled and collected at 05:30 and 19:00 h beforefeeding, and then stored at −80 °C in sealed plastic bags until lateranalysis of chemical composition (detailed methods are supplied in theSupplementary file). The ingredients and chemical composition arelisted in Supplementary Table S2.
Milk samples were collected at a proportion of 4:3:3 correspondingto the morning, noon and evening milking with a bronopol tablet (milkpreservative, D & F Control Systems, San Ramon, CA), and then storedat 4 °C before the infrared analysis of milk components and somaticcells using a spectrophotometer (Foss-4000, Foss, Hillerød, Denmark)within 2 days of collection [12]. Milk yields were recorded using milk-sampling devices (Waikato Milking Systems NZ Ltd., Waikato, Ha-milton, New Zealand) for 3 consecutive days.
Blood samples (10mL) were collected using two 5mL pro-coagu-lation tubes each from the jugular vein between 04:00 to 06:00 h. Then,the samples were centrifuged at 3000×g 4 °C for 15min to obtain thecorresponding serum within 30min of collection, which was thenstored at −80 °C for later analysis of biochemical parameters (detailsare specified in the Supplementary file) and metabolome profiles.
2.3. GC-TOF/MS detection and analysis
2.3.1. Identification and quantification of compounds detected by GC-TOF/MS
Serums were prepared and loaded into the GC-TOF/MS system asdepicted in the Supplementary file. The GC-TOF/MS analysis was per-formed using an Agilent 7890 GC system (Agilent 7890B, Agilent, USA)coupled with a Pegasus HT time-of-flight mass spectrometer (LECOChroma TOF PEGASUS HT, LECO, USA). The system was equipped witha DB-5MS capillary column coated with 5% diphenyl that was cross-linked with 95% dimethylpolysiloxane (30m×250 μM inner diameter,0.25-μM film thickness; J&W Scientific, Folsom, CA, USA). Proceduresof serum compounds separation, identification and relative quantifi-cation are described in the Supplementary file.
2.3.2. Identification of significantly different metabolites and pathwaysbetween HH and LL cows
The SIMCA 14.1 software package (V14.1, Sartorius Stedim DataAnalytics AB, Umea, Sweden) was applied to perform pattern recogni-tion multivariate analysis, which included principal component ana-lysis (PCA) and orthogonal partial least squares discriminant analysis(OPLS-DA), with the log transformation and unit variance scalingconversion mode.
The variable importance for the projection (VIP) > 1.0 and Pvalue < .05 was used to define the significantly different metabolites(SDMs) [13]. The SDMs were further identified and validated bysearching the online Kyoto Encyclopedia of Genes and Genomes(KEGG) and Bovine Metabolome Database (BMDB). Metaboanalyst 3.0(http://www.metaboanalyst.ca/) was employed to obtain relevantpathways. The Bos taurus (cow) pathway library was applied in thisprocedure.
2.3.3. Identification of biomarkers reflecting MPYEffective peaks data with similarity> 400 were imported into
Metaboanalyst 3.0 to manually select the minimum number of SDMs asbiomarkers to represent and explain the physiological difference be-tween HH and LL cows, based on the area under the curve (AUC),predicted class probabilities, and cross validation (CV) prediction(Specified procedures are depicted in the Supplementary file).
2.4. Biomarkers validation by ultra HPLC-MS/MS (UHPLC-MS/MS)
2.4.1. Standard solution and calibration curvesHippuric acid (HUA, 98%) from Sigma-Aldrich (St. Louis, MO), ni-
cotinamide (NAA, ≥99.5%) from Standards (Shanghai, China), andpelargonic acid (PEA, ≥90%) from Tokyo Chemical Industry (Tokyo,Japan) were separately dissolved or diluted at 10mmol/L as stock so-lutions. An aliquot of each stock solution was transferred to form amixed working standard solution. Then, a series of calibration standardsolutions were prepared by stepwise dilution, which were subjected toUHPLC-MS/MS analysis to form the calibration curves (SupplementaryTable S3).
2.4.2. UHPLC-MS/MS detection and analysisThe UHPLC separation was performed using an Agilent 1290
Infinity II series UHPLC System (Agilent Technologies) with a WatersACQUITY UPLC HSS T3 column (100×2.1mm, 1.8 μm). An Agilent6460 triple quadrupole mass spectrometer (Agilent Technologies),equipped with an AJS electrospray ionization interface, was applied forMS analysis. Details of metabolites extraction, quantitative parametersoptimization, UHPLC separation and MS identification processes arespecified in the Supplementary file.
2.4.3. LOD and LOQThe calibration standard solution was diluted stepwise then sub-
jected to UHPLC-MRM-MS analysis. The lower LOD (LLOD) and lowerLOQ (LLOQ) were defined as those peaks with S/N of 3 and 10, re-spectively [14].
2.4.4. Precision and accuracyThe quantitation precision was evaluated by RSD, determined by
injecting analytical replicates of a QC sample (quality control: themixed sample of HH and LL cows). The quantitation accuracy was de-termined by the analytical recovery of the QC sample. The percent re-covery was calculated as [(mean observed concentration)/(spikedconcentration)]× 100%.
2.5. Data analysis
The SAS MIXED model was used to analyze milk yield, milk com-position and blood biochemical parameters. P < .05 was defined as a
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
55

significant threshold, and P < .01 was regarded as an extremely sig-nificant difference.
3. Results
3.1. Lactation performance and blood biochemical parameters
The lactation performance of the 348 dairy cows was as follows: MYwas 31.9 (± 6.69 SD) kg/d, energy corrected milk yield (ECM) was30.9 (± 5.80) kg/d, and the content of milk protein, lactose and milkfat were 3.12 (± 0.29) %, 4.98 (± 0.18) %, and 3.16 (± 0.73) %,respectively. From this population, a total of 40 multiparous Holsteindairy cows were selected based on their MY and MP (Fig. 1, Supple-mentary Table S4). Twenty of them were defined as HH cows withhigher MY and higher MP (MY > 34.5 kg/d, MP > 3.2%), while 20other cows were defined as LL cows with lower MY and lower MP(MY < 31 kg/d, MP < 2.9%). The lactation performances of the HHand LL cows are listed in Table 1. MY, ECM, MPY and MP were sig-nificantly higher (P < .01) in HH cows than LL cows. No significantdifferences were found (P > .05) in the contents of milk fat, lactose, ormilk urea nitrogen between the two groups.
Sixteen blood biochemical parameters of the HH and LL cows, in-cluding protein metabolism, energy supply, liver and kidney function,oxidation and stress, are listed in Table 2. Malonaldehyde (P < .01)and total cholesterol were significantly higher (P < .05) in HH than inLL cows. No significant differences were found (P > .05) in otherblood biochemical parameters between the two groups. Detailed in-formation from the blood test results are listed in Supplementary TableS4.
3.2. Metabolites profiling using GC-TOF/MS
A total of 308 effective peaks were obtained after data pre-proces-sing, among which 178 compounds were relatively quantified and 130effective peaks were labeled “analyte” or “unknown” in light of theLECO-Fiehn Rtx5 database. The detailed information of all the effectivepeaks are listed in Supplementary Table S5.
3.3. Statistical comparison of metabolites in serum between the HH and LLgroups
The multivariate analysis of the metabolic profile differences be-tween the HH and LL cows are shown in Fig. 2. The PCA score plotdemonstrated no clear distinction between the two groups, and twooutliers went beyond the 95% Hotelling's T-squared ellipse (Fig. 2A). Inthe PCA model, the R2X value was 0.524, and Q2 was −0.175. TheOPLS-DA score plot showed significantly separated clusters between theHH and LL samples, whereas 2 samples from the LL group were not inthe 95% Hotelling T-squared ellipse (Fig. 2B). Two evaluating para-meters for the classification from the OPLS-DA model wereR2Y=0.905 and Q2= 0.688. Fig. 2C shows the 200 permutation testsresult, with intercept R2= (0, 0.754), Q2= (0, −0.216), indicatingthat the robustness of the OPLS-DA model was appropriate for down-stream analysis.
Fig. 1. Scatter plot of milk protein content and milk yield for the 347 mid-lactation dairy cows. The dotted red lines represent the grouping standard. Thecows with milk protein> 3.2% and milk yield exceeding 34.5 kg/d were re-garded as the HH group, and those with milk protein< 2.9% and milk yieldno> 31 kg/d were included in the LL group. (For interpretation of the refer-ences to colour in this figure legend, the reader is referred to the web version ofthis article.)
Table 1Milk yield and milk composition of experimental cows.
Item Group1 SEM P value
HH LL
Cow quantity, n 20 20 – –Yield, kg/dMilk 37.1a 26.9b 0.807 < 0.01ECM2 37.0a 25.0b 1.26 < 0.01Milk protein 1.23a 0.74b 0.0271 <0.01
Milk composition, %Protein 3.31a 2.77b 0.0284 <0.01Fat 3.15 3.03 0.199 0.66Lactose 5.04 4.96 0.0387 0.13Total solids 12.4 11.8 0.249 0.10
Milk urea nitrogen, mg/dL 14.0 14.6 0.481 0.45Somatic cells (×105) 3.90 3.32 1.72 0.81
1 HH means cows with higher milk yield (MY > 34.5 kg/d) and higher milkprotein content (MP > 3.2%); and LL means cows with lower milk yield(MY < 31 kg/d) and lower milk protein content (MP < 2.9%).
2 Energy-corrected milk, ECM=0.3246×milk yield (kg)+ 13.86×milkfat yield (kg)+ 7.04×milk protein yield (kg) [45].
a Means within a row with different superscripts differ (P < .05).b Means within a row with different superscripts differ (P < .05).
Table 2Blood parameters of experimental cows.
Item Group1 SEM P value
HH LL
Urea nitrogen, mmol/L 4.31 4.21 0.223 0.77Total protein, g/L 64.3 66.0 1.64 0.47Albumin, g/L 39.1 38.3 1.09 0.63Glucose, mmol/L 4.34 4.41 0.180 0.79Non-esterified fatty acid, mmol/L 0.217 0.247 0.0249 0.40Total cholesterol, mmol/L 6.30a 5.21b 0.300 0.01β-hydroxybutyrate, mmol/L 0.519 0.575 0.0391 0.32Triglyceride, mmol/L 0.297 0.300 0.0211 0.92Glutamic-pyruvic transaminase, U/L 5.74 5.15 0.487 0.40Glutamic-oxalacetic transaminease, U/L 9.79 9.03 0.631 0.40Creatinine, mmol/L 64.2 63.3 3.01 0.84Total bilirubin, mmol/L 5.05 4.46 0.334 0.22Malonaldehyde, nmol/mL 4.53a 2.86b 0.289 < 0.01Superoxide dismutase, U/mL 140 135 3.06 0.25Glutathione peroxidase, U/mL 133 127 4.11 0.34Total antioxidant capability, U/mL 3.72 3.43 0.189 0.27
1 HH means cows with higher milk yield (MY >34.5 kg/d) and higher milkprotein content (MP > 3.2%), and LL means cows with lower milk yield(MY < 31 kg/d) and lower milk protein content (MP < 2.9%).
a Means within a row with different superscripts differ (P < .05).b Means within a row with different superscripts differ (P < .05).
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
56

3.4. Identification of significantly different metabolites and pathwaysbetween HH and LL cows
Thirty-six SDMs and 19 significantly different unidentified peaksbetween the HH and LL groups are shown in the volcano plot (Fig. 3).Among these, 24 SDMs were more abundant in the HH group, with 8metabolites belonging to AAs and AA conjugates, including allantoicacid (Log2 fold change (FC)= 0.850), creatine (Log2 (FC)= 0.560),kyotorphin (Log2 (FC)= 0.509), asparagine (Log2 (FC)= 0.462), iso-leucine (Ile, Log2 (FC)= 0.336), 3-aminoisobutyric acid (Log2(FC)= 0.315), serine (Ser, Log2 (FC)= 0.285) and HUA (Log2(FC)= 0.253); four SDMs belonging to carbohydrates (CHOs) and CHOconjugates, including D-glucoheptose (Log2 (FC)= 0.796), salicin (Log2(FC)= 0.755), sedoheptulose (Log2 (FC)= 0.349) and threitol (Log2(FC)= 0.244); and three organic acids. In addition, 12 SDMs wereenriched in the LL group, with six organic acids including 2 fatty acids,PEA (Log2 (FC)=−0.558) and caprylic acid (Log2 (FC)=−0.556);and three alcohols including resveratrol (Log2 (FC)=−10.4), picea-tannol (Log2 (FC)=−9.95) and 3,4-dihydroxymandelic acid (Log2(FC)=−6.92). Detailed information on the SDMs are included inSupplementary Table S6.
Thirty-six SDMs enriched metabolic pathways were identified andare illustrated in a metabolome view map (Fig. 4). These SDMs weremainly involved in valine (Val), leucine (Leu) and Ile biosynthesis(P= .021, impact value= 0.333), glycine (Gly), Ser and threonine(Thr) metabolism (P= .027, impact value= 0.243), the tricarboxylicacid cycle (TCA cycle, P= .063, impact value= 0.125), nicotinate andNAA metabolism (P= .241, impact value= 0.159), glyoxylate and
Fig. 2. PCA score map, OPLS-DA score plot and corresponding validation plot of OPLS-DA derived from the metabolite profiles of experimental cows.The red box represents HH, while the blue circle represents LL. PCA=principal component analysis. OPLS-DA=orthogonal partial least squares discriminantanalysis. HH= cows with higher milk yield (MY > 34.5 kg/d) and higher milk protein content (MP > 3.2%). LL= cows with lower milk yield (MY < 31 kg/d)and lower milk protein content (MP < 2.9%). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of thisarticle.)
Fig. 3. Volcano plot of serum metabolites for HH and LL.The x-axis represents 1og2 (FC) value, and the y-axis means −log10 (P value).The red dot indicates a significantly different metabolite (SDM) that is moreabundant in the HH group, while the blue dot represents a higher concentrationof SDM in the LL cows. The dot size represents the variable importance in theprojection (VIP) value. FC= fold change, mean value of peak area obtainedfrom the HH group/mean value of peak area obtained from the LL. HH=cowswith higher milk yield (MY > 34.5 kg/d) and higher milk protein content(MP > 3.2%). LL= cows with lower milk yield (MY < 31 kg/d) and lowermilk protein content (MP < 2.9%). (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of thisarticle.)
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
57

dicarboxylate metabolism (P= .287, impact value=0.296), pyruvatemetabolism (P= .373, impact value= 0.188) and glutathione meta-bolism (P= .425, impact value=0.361) with impact value>0.1(Supplementary Table S7).
3.5. Identification of biomarkers reflecting MPY
The HUA, NAA and PEA were selected as potential biomarkers todistinguish between higher and lower MPY cows, with the AUCvalue=0.943 (Fig. 5A) and an average accuracy= 0.899 based on 100cross validations (Fig. 5C). However, there were still three outliers inthe probability view image (Fig. 5B).
3.6. Validation of biomarkers reflecting MPY using UHPLC-MS/MS
Fig. S1 shows the extracted ion chromatographs (EICs) from thestandard solution and a representative sample (A13 in HH group) ofHUA and NAA. Both analytes showed symmetrical peak shapes and thebaseline separations were appropriate for the following quantification.The LLOD and LLOQ of HUA were 3.1 nmol/L and 6.1 nmol/L, re-spectively. The LLOD and LLOQ of NAA were 6.1 nmol/L and12.2 nmol/L, respectively. The correlation coefficients (R2) of regres-sion fitting were 0.999 and 0.997 for HUA and NAA, respectively,which showed a good quantitative relationship between the MS re-sponses and the analyte concentrations (Supplementary Table S3).
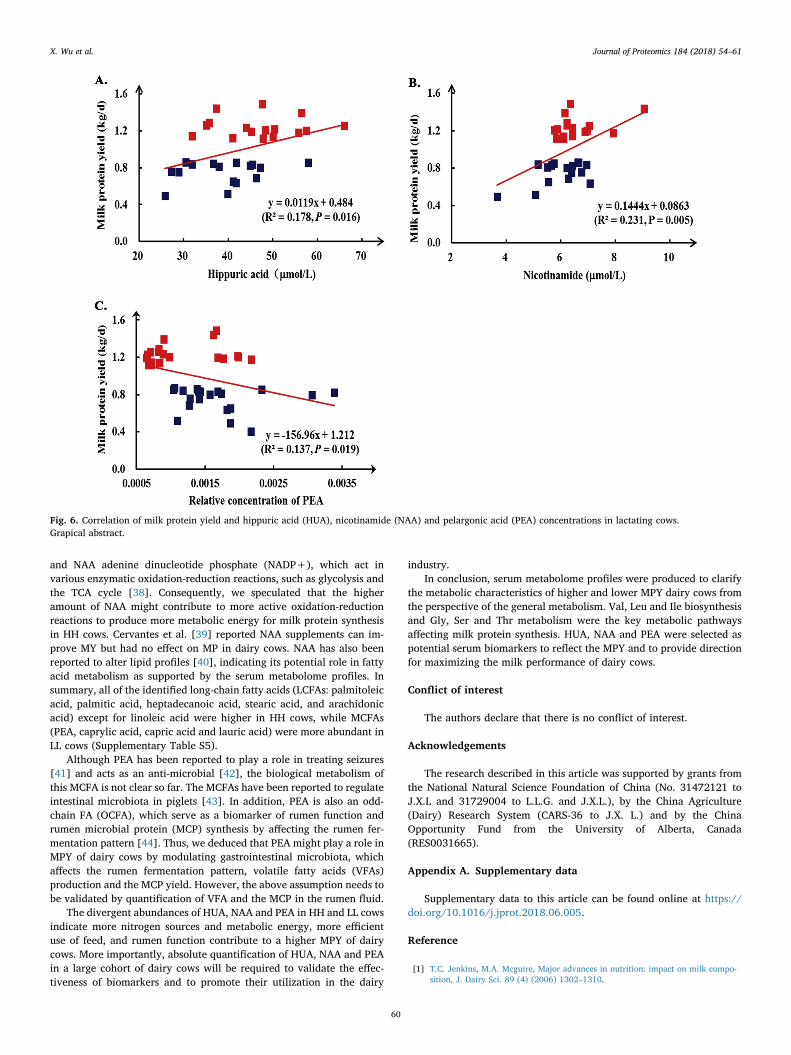
The quantification results are shown in Supplementary Table S8.The absolute concentration of HUA (P= .019) and NAA were sig-nificantly higher or tended to be higher in the HH group than in the LLgroup (P= .057). In addition, MPY was significantly positive correlatedwith HUA (R2= 0.178, P= .016) and NAA (R2=0.231, P= .005),respectively. A significant negative correlation was obtained betweenMPY and the relative concentration of PEA (R2=0.137, P= .019)(Fig. 6).
4. Discussion
To our knowledge, the current study is the first to propose theconcept of MPY by combining the MY and MP traits together and tostudy how serum metabolomes could be related to MPY, which willprovide novel implications for achieving maximized MPY of dairy cowsand maximized benefits for the dairy industry. We also investigatedmetabolic pathways and biomarkers associated with MPY that couldprovide directions for enhancing MPY beyond nutritional and man-agement strategies.
4.1. Significantly different metabolites
The SDMs analysis identified the majority (8 out of 9) of AAs and AAconjugates were more abundant in HH cows, suggesting more AAsavailable for milk protein synthesis, consistent with a previous study[15]. Further, these enriched AAs could stimulate milk protein synth-esis by increasing the phosphorylation of the mammalian target of ra-pamycin, ribosomal protein S6 kinase 1 and ribosomal protein S6 aspreviously reported [16]. All of the SDMs classified into the CHOs andCHO conjugates were more enriched in HH cows. In detail, sedo-heptulose serves as a constituent of sedoheptulose-7-phosphate and isan intermediate metabolite of the pentose phosphate pathway (PPP)[17]. Threitol is an end product of xylose metabolism that is also relatedto PPP through the glucuronate pathway [18]. PPP is most active in themammary gland and adipose tissue in rats [19]. Therefore, the higherrelative concentration of sedoheptulose and threitol may indicate moreenergy supply to improve MPY by glucose oxidation in the mammarygland [20]. On the other hand, two medium chain fatty acids (MCFAs),caprylic acid and PEA, were enriched in LL cows. It has been reportedthat MCFAs regulate the leptin expression of bovine mammary epi-thelial cells or bovine adipocytes, which further regulate feed intakeand energy expenditure in vitro [21, 22]. In vivo, supplementing withMCFAs can decrease the dry matter intake, MP and lactose levels oflactating dairy cows [23]. Thus, the relatively higher concentration ofAAs and CHOs and the lower concentration of fatty acids reveals thatmore AAs and more metabolic energy were provided for HH cows toimprove MPY.
4.2. Metabolic pathways
This study not only identified the SDMs between HH and LL cowsbut also pinpointed the most influential pathways these metabolites areinvolved in: Val, Leu and Ile biosynthesis and Gly, Ser and Thr meta-bolism.
Val, Leu and Ile biosynthesis are different as indicated by the Ile(Log2 (FC)= 0.336) and pyruvic acid (Log2 (FC)=−0.399)(Supplementary Table S7). Ile infusion has been reported to enhanceMY [24] and MP [25]. As a branched-chain AA, Ile not only serves as anAA component in MP but also regulates MP synthesis by the mTORcomplex 1 signaling pathway [16], indicating that the higher Ile con-tributes to higher MPY of dairy cows. Pyruvic acid, as a substrate of Ile[26], is significantly lower in HH cows, probably attributable to moreIle synthetized by HH cows. Pyruvic acid can be converted into acetyl-CoA by oxidative decarboxylation, and the acetyl-CoA then participatesin the TCA cycle to produce adenosine triphosphate (ATP) to meet thehigher requirement of energy for milk protein production in HH cows.The higher relative concentrations of citric acid (Log2 (FC)= 0.466)and succinic acid (Log2 (FC)= 1.18) also support the concept of a moreactive TCA cycle in HH cows.
The Gly, Ser and Thr metabolism pathways are characterized by thedivergence of Ser (Log2 (FC)= 0.285), creatine (Log2 (FC)= 0.560)and pyruvic acid in this study (Supplementary Table S7). Although Glywas not significantly different between HH and LL cows, its down-stream products creatine and L-allothreonine were significantly higherin the HH group (P < .05). Creatine synthesis is closely related to Gly
Fig. 4. Metabolome view map of the significantly different metabolites iden-tified in HH and LL.The x-axis represents the pathway impact, and the y-axis represents thepathway enrichment. Larger sizes and darker colors represent higher pathwayimpact values and higher pathway enrichment, respectively. HH=cows withhigher milk yield (MY > 34.5 kg/d) and higher milk protein content(MP > 3.2%). LL= cows with lower milk yield (MY < 31 kg/d) and lowermilk protein content (MP < 2.9%).
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
58

metabolism, and mainly occurs in the liver and kidney, then it istransported via the blood to tissues with high energy demands, such asthe brain and muscle [27]. Given that its main role is recycling ade-nosine diphosphate (ADP) to ATP, the higher relative concentration ofcreatine in the serum might indicate the availability of more ATP forphysiological metabolism in HH cows. L-allothreonine can be directlygenerated through Gly by L-threonine aldolase and L-allo-threoninealdolase. L-Allothreonine is also the substrate of Ser hydroxymethyltransferase, a coenzyme that catalyzes the invertible interaction of Glyand Ser [28]. Ser, as a nonessential AA, can also be synthesized from 3-phosphoglycerate by phosphoglycerate dehydrogenase, phosphoserinetransaminase and phosphoserine phosphatase in the body [29]. Thenewly synthesized Ser could be transported in the blood to the mam-mary gland to synthesize MP [30]. It has been previously reported thatGly, Ser and Thr metabolism was more active in dairy cows fed high-quality forage diets and they might serve as biomarkers for higher MYand MP quality [13]. Thus, a more active Gly, Ser and Thr metabolismmight partly contribute to more MY and MP production in HH cows.
Val, Leu and Ile biosynthesis, and Gly, Ser and Thr metabolism havebeen reported to be involved in some pathological changes, such as type2 diabetes [31], ischemic stroke [32], and hepatitis C virus [33] inhumans based on plasma or urine metabolome profiles, which mightreveal a close relationship between milk performance and animalhealth. Thus, the above metabolic pathways suggest that AAs, energylevel and health status played a vital role in milk protein synthesis.
4.3. Biomarkers
HUA is derived from dietary protein degradation and can be re-synthesized from aromatic compounds or cyclic sugar type-compoundsby the gastrointestinal microbiota through the shikimate pathway [34].Urinary HUA can reflect the dietary composition, hepatic function,disease state and even metabolism alterations [34]. As Sun et al. [11]reported, a higher urinary HUA level leads to more nitrogen loss andlower MPY in cows fed low-quality forages, which supports the hy-pothesis that HUA as excretory products are mainly produced fromdietary protein degradation. Thus, we assumed that the higher serumHUA level in HH cows might be derived from higher protein degrada-tion, which is transported by the blood to provide a nitrogen source formilk protein synthesis. On the other hand, Vanessa et al. [35] reportedfasting serum HUA concentrations were elevated when humans con-sumed a high-anthocyanins diet, which is also related to better glucoseand insulin metabolism. Consequently, we speculated that the higherserum HUA levels in the HH group might also indicate more energysupplied by glucose metabolism and hormone regulation. Karisa et al.[36] reported that HUA may serve as a potential biomarker of residualfeed intake in beef steers, which might reveal a divergent feed effi-ciency in HH and LL cows. In addition, it has been reported that HUA,as an aromatic compound, could be converted into aromatic AAs suchas tryptophan, tyrosine and phenylalanine, then be transformed intoNAA [37], which was another potential biomarker found in this study.
NAA can be incorporated into NAA adenine dinucleotide (NAD+)
Fig. 5. Biomarker analysis results (ROC view) of serum metabolites for milk protein yield in experimental cows.Statistical method to evaluate treatment effectiveness using hippuric acid, nicotinamide and pelargonic acid (A). Probability view image (B) and cross validation plot(C). HH= cows with higher milk yield (MY > 34.5 kg/d) and higher milk protein content (MP > 3.2%). LL= cows with lower milk yield (MY < 31 kg/d) andlower milk protein content (MP < 2.9%).
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
59
and NAA adenine dinucleotide phosphate (NADP+), which act invarious enzymatic oxidation-reduction reactions, such as glycolysis andthe TCA cycle [38]. Consequently, we speculated that the higheramount of NAA might contribute to more active oxidation-reductionreactions to produce more metabolic energy for milk protein synthesisin HH cows. Cervantes et al. [39] reported NAA supplements can im-prove MY but had no effect on MP in dairy cows. NAA has also beenreported to alter lipid profiles [40], indicating its potential role in fattyacid metabolism as supported by the serum metabolome profiles. Insummary, all of the identified long-chain fatty acids (LCFAs: palmitoleicacid, palmitic acid, heptadecanoic acid, stearic acid, and arachidonicacid) except for linoleic acid were higher in HH cows, while MCFAs(PEA, caprylic acid, capric acid and lauric acid) were more abundant inLL cows (Supplementary Table S5).
Although PEA has been reported to play a role in treating seizures[41] and acts as an anti-microbial [42], the biological metabolism ofthis MCFA is not clear so far. The MCFAs have been reported to regulateintestinal microbiota in piglets [43]. In addition, PEA is also an odd-chain FA (OCFA), which serve as a biomarker of rumen function andrumen microbial protein (MCP) synthesis by affecting the rumen fer-mentation pattern [44]. Thus, we deduced that PEA might play a role inMPY of dairy cows by modulating gastrointestinal microbiota, whichaffects the rumen fermentation pattern, volatile fatty acids (VFAs)production and the MCP yield. However, the above assumption needs tobe validated by quantification of VFA and the MCP in the rumen fluid.
The divergent abundances of HUA, NAA and PEA in HH and LL cowsindicate more nitrogen sources and metabolic energy, more efficientuse of feed, and rumen function contribute to a higher MPY of dairycows. More importantly, absolute quantification of HUA, NAA and PEAin a large cohort of dairy cows will be required to validate the effec-tiveness of biomarkers and to promote their utilization in the dairy
industry.In conclusion, serum metabolome profiles were produced to clarify
the metabolic characteristics of higher and lower MPY dairy cows fromthe perspective of the general metabolism. Val, Leu and Ile biosynthesisand Gly, Ser and Thr metabolism were the key metabolic pathwaysaffecting milk protein synthesis. HUA, NAA and PEA were selected aspotential serum biomarkers to reflect the MPY and to provide directionfor maximizing the milk performance of dairy cows.
Conflict of interest
The authors declare that there is no conflict of interest.
Acknowledgements
The research described in this article was supported by grants fromthe National Natural Science Foundation of China (No. 31472121 toJ.X.L and 31729004 to L.L.G. and J.X.L.), by the China Agriculture(Dairy) Research System (CARS-36 to J.X. L.) and by the ChinaOpportunity Fund from the University of Alberta, Canada(RES0031665).
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jprot.2018.06.005.
Reference
[1] T.C. Jenkins, M.A. Mcguire, Major advances in nutrition: impact on milk compo-sition, J. Dairy Sci. 89 (4) (2006) 1302–1310.
Fig. 6. Correlation of milk protein yield and hippuric acid (HUA), nicotinamide (NAA) and pelargonic acid (PEA) concentrations in lactating cows.Grapical abstract.
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
60

[2] S. Bedö, E. Nikodémusz, K. Percsich, L. Bárdos, Variations in the milk yield and milkcomposition of dairy cows during lactation, Acta Vet. Hung. 43 (1) (1995) 163.
[3] G.A. Broderick, A.P. Faciola, L.E. Armentano, Replacing dietary soybean meal withcanola meal improves production and efficiency of lactating dairy cows, J. DairySci. 98 (8) (2015) 5672–5687.
[4] Z. Zhou, M. Vailatiriboni, E. Trevisi, J.K. Drackley, D.N. Luchini, J.J. Loor, Betterpostpartal performance in dairy cows supplemented with rumen-protected me-thionine compared with choline during the peripartal period, J. Dairy Sci. 99 (11)(2016) 8716–8732.
[5] N. Anantasook, M. Wanapat, A. Cherdthong, P. Gunun, Effect of tannins and sa-ponins in Samanea saman on rumen environment, milk yield and milk compositionin lactating dairy cows, J. Anim. Physiol. Anim. Nutr. 99 (2) (2015) 335–344.
[6] S.D. Cochran, J.B. Cole, D.J. Null, P.J. Hansen, Discovery of single nucleotidepolymorphisms in candidate genes associated with fertility and production traits inHolstein cattle, BMC Genet. 14 (1) (2013) 49.
[7] C.V. Phyn, J.K. Kay, A.G. Rius, S.R. Morgan, C.G. Roach, T.M. Grala, J.R. Roche,Temporary alterations to postpartum milking frequency affect whole-lactation milkproduction and the energy status of pasture-grazed dairy cows, J. Dairy Sci. 97 (11)(2014) 6850–6868.
[8] T.C. Linden, R.C. Bicalho, D.V. Nydam, Calf birth weight and its association withcalf and cow survivability, disease incidence, reproductive performance, and milkproduction, J. Dairy Sci. 92 (6) (2009) 2580–2588.
[9] R.L. Laben, R. Shanks, P.J. Berger, A.E. Freeman, Factors affecting milk yield andreproductive performance, J. Dairy Sci. 65 (6) (1982) 1004–1015.
[10] T. Soga, R. Baran, M. Suematsu, Y. Ueno, S. Ikeda, T. Sakurakawa, Y. Kakazu,T. Ishikawa, M. Robert, T. Nishioka, Differential metabolomics reveals ophthalmicacid as an oxidative stress biomarker indicating hepatic glutathione consumption, J.Biol. Chem. 281 (24) (2006) 16768–16776.
[11] H.Z. Sun, B. Wang, J.K. Wang, H.Y. Liu, J.X. Liu, Biomarker and pathway analysesof urine metabolomics in dairy cows when corn Stover replaces alfalfa hay, J. Anim.Sci. Biotechnol. 7 (1) (2016) 49.
[12] M.F. Laporte, P. Paquin, Near-infrared analysis of fat, protein, and casein in cow'smilk, J. Agric. Food Chem. 47 (7) (1999) 2600–2605.
[13] H.Z. Sun, D.M. Wang, B. Wang, J.K. Wang, H.Y. Liu, L.L. Guan, J.X. Liu,Metabolomics of four biofluids from dairy cows: potential biomarkers for milkproduction and quality, J. Proteome Res. 14 (2) (2015) 1287–1298.
[14] B. Booth, M.E. Arnold, B. Desilva, L. Amaravadi, S. Dudal, E. Fluhler, B. Gorovits,S.H. Haidar, J. Kadavil, S. Lowes, Workshop report: crystal city v - quantitativebioanalytical method validation and implementation: the 2013 revised FDA gui-dance, AAPS J. 17 (2) (2015) 277–288.
[15] L. Doepel, D. Pacheco, J.J. Kennelly, M.D. Hanigan, I.F. López, H. Lapierre, Milkprotein synthesis as a function of amino acid supply, J. Dairy Sci. 87 (5) (2004)1279–1297.
[16] J.A. Appuhamy, N.A. Knoebel, W.A. Nayananjalie, J. Escobar, M.D. Hanigan,Isoleucine and leucine independently regulate mTOR signaling and protein synth-esis in MAC-T cells and bovine mammary tissue slices, J. Nutr. 142 (3) (2012)484–491.
[17] P.A. Marks, A newer pathway of carbohydrate metabolism; the pentose phosphatepathway, Diabetes 5 (4) (1956) 276–283.
[18] E. Pitkänen, The conversion of d -xylose into d -threitol in patients without liverdisease and in patients with portal liver cirrhosis, Clin. Chim. Acta 80 (1) (1977)49–54.
[19] H. Cabezas, R.R. Raposo, E. Meléndez-Hevia, Activity and metabolic roles of thepentose phosphate cycle in several rat tissues, Mol. Cell. Biochem. 201 (1–2)(1999) 57.
[20] M.S. Allen, P. Piantoni, Carbohydrate nutrition: managing energy intake and par-titioning through lactation, Vet. Clin. N. Am. Food Anim. Pract. 30 (3) (2014)577–597.
[21] M. Soliman, K. Kimura, M. Ahmed, D. Yamaji, Y. Matsushita, Y. Okamatsu-Ogura,K. Makondo, M. Saito, Inverse regulation of leptin mRNA expression by short- andlong-chain fatty acids in cultured bovine adipocytes, Domest. Anim. Endocrinol. 33(4) (2007) 400–409.
[22] T. Yonezawa, S. Yonekura, M. Sanosaka, A. Hagino, K. Katoh, Y. Obara, Octanoatestimulates cytosolic triacylglycerol accumulation and CD36 mRNA expression butinhibits acetyl coenzyme a carboxylase activity in primary cultured bovine mam-mary epithelial cells, J. Dairy Res. 71 (71) (2004) 398–404.
[23] R. Fukumori, T. Sugino, H. Shingu, N. Moriya, H. Kobayashi, Y. Hasegawa,M. Kojima, K. Kangawa, T. Obitsu, S. Kushibiki, Ingestion of medium chain fattyacids by lactating dairy cows increases concentrations of plasma ghrelin, Domest.
Anim. Endocrinol. 45 (4) (2013) 216–223.[24] P.H. Robinson, W. Chalupa, C.J. Sniffen, W.E. Julien, H. Sato, T. Fujieda,
K. Watanabe, H. Suzuki, Influence of postruminal supplementation of methionineand lysine, isoleucine, or all three amino acids on intake and chewing behavior,ruminal fermentation, and milk and milk component production, J. Anim. Sci. 77(10) (1999) 2781–2792.
[25] C.G. Schwab, L.D. Satter, A.B. Clay, Response of lactating dairy cows to abomasalinfusion of amino acids, J. Dairy Sci. 59 (7) (1976) 1254–1270.
[26] M. Kisumi, S. Komatsubara, I. Chibata, Pathway for isoleucine formation formpyruvate by leucine biosynthetic enzymes in leucine-accumulating isoleucine re-vertants of Serratia marcescens, J. Biochem. 82 (1) (1977) 95–103.
[27] E.E. Edison, M.E. Brosnan, C. Meyer, J.T. Brosnan, Creatine synthesis: production ofguanidinoacetate by the rat and human kidney in vivo, Am. J. Physiol. RenalPhysiol. 293 (6) (2007) F1799–F1804.
[28] J.C. Heo, S.H. Lee, Alleviation of asthma-related symptoms by a derivative of L-Allothreonine, Int. J. Mol. Med. 31 (4) (2013) 881–887.
[29] L.I. Pizer, Enzymology and regulation of serine biosynthesis in cultured humancells, J. Biol. Chem. 239 (1964) 4219–4226.
[30] T.R. Mackle, D.A. Dwyer, K.L. Ingvartsen, P.Y. Chouinard, D.A. Ross, D.E. Bauman,Evaluation of whole blood and plasma in the interorgan supply of free amino acidsfor the mammary gland of lactating dairy cows, J. Dairy Sci. 83 (6) (2000)1300–1309.
[31] A.H. Zhang, H. Sun, G.L. Yan, Y. Yuan, Y. Han, X.J. Wang, Metabolomics study oftype 2 diabetes using ultra-performance LC-ESI/quadrupole-TOF high-definition MScoupled with pattern recognition methods, J. Physiol. Biochem. 70 (1) (2014)117–128.
[32] Y. Huang, C.Y. Gu, X.S. Xia, X. Li, Metabolomics analysis on patients with ischemicstroke based on ultra-performance liquid chromatography with quadrupole-time-of-flight mass spectrometry, Int. J. Clin. Exp. Med. 10 (7) (2017) 10277–10286.
[33] A. Zhang, H. Sun, Y. Han, G. Yan, Y. Yuan, G. Song, X. Yuan, N. Xie, X. Wang,Ultraperformance liquid chromatography–mass spectrometry based comprehensivemetabolomics combined with pattern recognition and network analysis methods forcharacterization of metabolites and metabolic pathways from biological data sets,Anal. Chem. 85 (15) (2013) 7606–7612.
[34] H.J. Lees, J.R. Swann, I.D. Wilson, J.K. Nicholson, E. Holmes, Hippurate: the naturalhistory of a mammalian–microbial cometabolite, J. Proteome Res. 12 (4) (2013)1527–1546.
[35] D.D.M. Vanessa, M.A. Lankinen, J. Lindström, R. Puupponen-Pimiä,D.E. Laaksonen, J. Pihlajamäki, M. Lehtonen, M. Uusitupa, J. Tuomilehto,M. Kolehmainen, Fasting serum hippuric acid is elevated after bilberry (Vacciniummyrtillus) consumption and associates with improvement of fasting glucose levelsand insulin secretion in persons at high risk of developing type 2 diabetes, Mol.Nutr. Food Res. 61 (9) (2017) 1700019.
[36] B.K. Karisa, J. Thomson, Z. Wang, C. Li, Y.R. Montanholi, S.P. Miller, S.S. Moore,G.S. Plastow, Plasma metabolites associated with residual feed intake and otherproductivity performance traits in beef cattle, Livest. Sci. 165 (1) (2014) 200–211.
[37] R.W. Pero, Health consequences of catabolic synthesis of hippuric acid in humans,Curr. Clin. Pharmacol. 5 (1) (2010) 67.
[38] P. Belenky, K.L. Bogan, C. Brenner, NAD+ metabolism in health and disease,Trends Biochem. Sci. 32 (1) (2007) 12–19.
[39] A. Cervantes, T.R. Smith, J.W. Young, Effects of nicotinamide on milk compositionand production in diary cows fed supplemental fat, J. Dairy Sci. 79 (1) (1996)105–113.
[40] A. Zák, M. Zeman, M. Vecka, E. Tvrzická, Nicotinic acid: an unjustly neglectedremedy, Cas. Lek. Cesk. 145 (11) (2006) 825–831.
[41] P. Chang, N. Terbach, N. Plant, P.E. Chen, M.C. Walker, R.S.B. Williams, Seizurecontrol by ketogenic diet-associated medium chain fatty acids, Neuropharmacology69 (2013) 105–114.
[42] J. Pimentel, K. Richardson, Antimicrobial Formulations with Pelargonic Acid, WO,(2013).
[43] J. Zentek, F. Ferrara, R. Pieper, L. Tedin, W. Meyer, W. Vahjen, Effects of dietarycombinations of organic acids and medium chain fatty acids on the gastrointestinalmicrobial ecology and bacterial metabolites in the digestive tract of weaning pig-lets, J. Anim. Sci. 91 (7) (2013) 3200–3210.
[44] V. Fievez, E. Colman, J.M. Castromontoya, I. Stefanov, B. Vlaeminck, Milk odd- andbranched-chain fatty acids as biomarkers of rumen function - an update, Anim. FeedSci. Tech. 172 (1–2) (2012) 51–65.
[45] R. Orth, Sample Day and Lactation Report, DHIA 200, Fact Sheet A-2. Mid-StatesDairy Records Processing Center (DRPC), Ames, IA, 1992.
X. Wu et al. Journal of Proteomics 184 (2018) 54–61
61





























