Embed Size (px)
Citation preview

JOURNAL OF THE AUSTRALIA NEW GUINEA FISHES ASSOCIATIONincorporated Registration No. ACO27788J
845
The new Laser Red Rainbowfish Melanoteania rubrivittata. G.L.
VOLUME TWENTY-NINENUMBER ONE
MARCH 2015
Main features in this issue:
MELANOTAENIA RUBRIVITTATA, A NEW SPECIES OF RAINBOWFISH
(MELANOTAENIIDAE) FROM NORTHWESTERN PAPUA PROVINCE, INDONESIA
Gerald R. Allen, Peter J. Unmack & Renny K. Hadziaty 846
WERE THOSE RED LASER “STRIPES” REALLY WORTH IT? –THE HUNT FOR THE WAPOGA RED LASER RAINBOWFISH
Gary Lange, Johannes Graf and Dan Dority 859
A COMMUNITY HERO – THE DWARF FLATHEAD GUDGEON
David Shoesmith 866
Habitat of Dwarf Flathead Gudgeon (page 866) D.S.

MELANOTAENIA RUBRIVITTATA, A NEW SPECIES OF RAINBOWFISH
(MELANOTAENIIDAE) FROM NORTHWESTERN PAPUA PROVINCE, INDONESIA
Gerald R. Allen1, Peter J. Unmack2 & Renny K. Hadiaty3
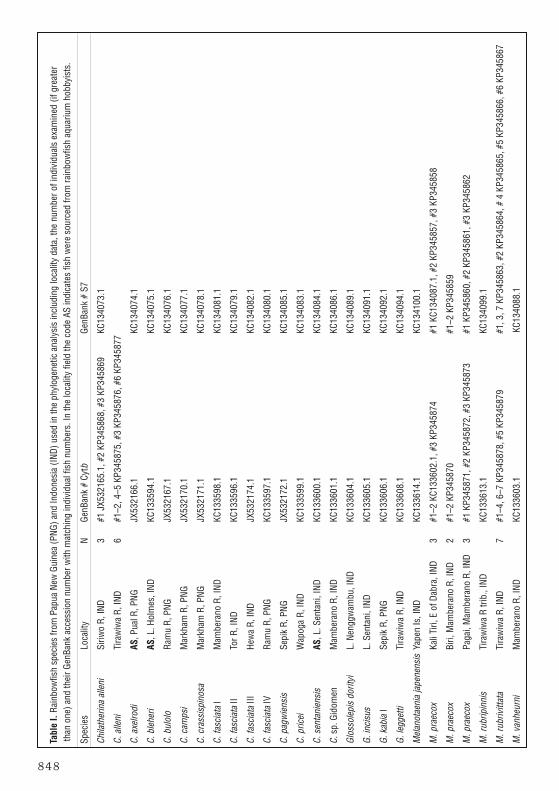
AbstractA new species of melanotaeniid rainbowfish, Melanotaenia rubrivittata, is described from theWapoga River system of northwestern Papua Province, Indonesia. The new taxon is describedon the basis of 26 specimens, 18.0–48.7 mm SL, collected from two sites near the TirawiwaRiver. It is similar to M. praecox in general appearance and morphology, but differs inpossessing red body stripes, a more slender body shape in adult males, a slightly longer snout,and fewer lateral scales. Genetic analysis provides additional evidence for the separation ofthese taxa.
IntroductionRainbowfishes of the family Melanotaeniidae are a spectacularly coloured group of freshwaterfishes endemic to the warmer portions of Australia and New Guinea. They live in a variety ofhabitats from clear streams and lakes through large turbid rivers, floodplain habitats, andisolated rocky pools or waterholes in arid regions. Most species form loose aggregations, whichswim either in midwater or just below the surface. The main dietary items include insects whichfall onto the surface and micro-crustaceans. Spawning occurs year round in most species, butreproductive activity often peaks at the onset of rainy periods. Rainbowfishes are popularaquarium fishes with dedicated hobbyist groups that specialise in them (Allen 1995; Tappin2011).
The families Melanotaeniidae, Eleotridae (gudgeons) and Gobiidae (gobies) collectively make upover 50% (~280 species) of the freshwater fish species richness of Australia and New Guinea
846
Fig. 1. Map of Papua Province, Indonesia showing type locality (star) of Melanotaenia rubrivittata nearSiewa, an abandoned mining exploration camp.

847
(Unmack et al. 2013). Despite the large number of rainbowfish species recognized, there aremany species that still require descriptions from Australia and New Guinea (Kadarusman et al.2012b; Unmack et al. 2013). Furthermore, almost any new regions explored in New Guinea(Allen et al. 2008, Allen & Hadiaty 2011, 2013; Kadarusman et al. 2012a), as well as othersre-explored (Allen & Unmack 2012; Allen et al. 2014; Kadarusman et al. 2010) continue toturn up new species at a high rate.
The broader biogeographic patterns within rainbowfishes are now well established from theearlier work of Zhu et al. (1994) and McGuigan et al. (2000) and more recent papers withexpanded taxonomic and molecular coverage (Kadarusman et al. 2012b; Unmack et al. 2013).Most species in the family are separated into three monophyletic groups representing westernNew Guinea (Birds Head/Birds Neck region), northern New Guinea and southern New Guineaand Australia. Western New Guinea can be further separated into three allopatrically distrib-uted lineages, while northern New Guinea contains three largely sympatric lineages andsouthern New Guinea/Australia has four broadly sympatric lineages. In addition, three earlybranching lineages represent Cairnsichthys Allen 1980, Rhadinocentrus Regan 1914 (bothrestricted to eastern Australia) and Iriatherina Meinken 1974 (found in northern Australia and
Fig. 2. Aquarium photograph of freshly collected holotype of Melanotaenia rubrivittata, male (upper),48.7 mm SL and female paratype, 40.8 mm SL (WAM P.31454), Siewa, Papua Province. G.R.A.
848
Tabl
e I.
Rain
bow
fish
spec
ies
from
Pap
ua N
ew G
uine
a (P
NG) a
nd In
done
sia
(IND)
use
d in
the
phyl
ogen
etic
ana
lysi
s in
clud
ing
loca
lity
data
, the
num
ber o
f ind
ivid
uals
exa
min
ed (i
f gre
ater
than
one
) and
thei
r Gen
Bank
acc
essi
on n
umbe
r with
mat
chin
g in
divi
dual
fish
num
bers
. In
the
loca
lity
field
the
code
AS
indi
cate
s fis
h w
ere
sour
ced
from
rain
bow
fish
aqua
rium
hob
byis
ts.
Spec
ies
Loca
lity
NGe
nBan
k #
Cytb
GenB
ank
# S7
Chila
ther
ina
alle
niSi
riwo
R, IN
D3
#1 J
X532
165.
1, #
2 KP
3458
68, #
3 KP
3458
69KC
1340
73.1
C. a
lleni
Tira
wiw
a R,
IND
6#1
–2, 4
–5 K
P345
875,
#3
KP34
5876
, #6
KP34
5877
C. a
xelro
diAS
, Pua
l R, P
NGJX
5321
66.1
KC13
4074
.1
C. b
lehe
riAS
, L. H
olm
es, I
NDKC
1335
94.1
KC13
4075
.1
C. b
ulol
oRa
mu
R, P
NGJX
5321
67.1
KC13
4076
.1
C. c
amps
iM
arkh
am R
, PNG
JX53
2170
.1KC
1340
77.1
C. c
rass
ispi
nosa
Mar
kham
R, P
NGJX
5321
71.1
KC13
4078
.1
C. fa
scia
taI
Mam
bera
no R
, IND
KC13
3598
.1KC
1340
81.1
C. fa
scia
taII
Tor R
, IND
KC13
3596
.1KC
1340
79.1
C. fa
scia
taIII
Hew
a R,
IND
JX53
2174
.1KC
1340
82.1
C. fa
scia
taIV
Ram
u R,
PNG
KC13
3597
.1KC
1340
80.1
C. p
agw
iens
isSe
pik
R, P
NGJX
5321
72.1
KC13
4085
.1
C. p
ricei
Wap
oga
R, IN
DKC
1335
99.1
KC13
4083
.1
C. s
enta
nien
sis
AS, L
. Sen
tani
, IND
KC13
3600
.1KC
1340
84.1
C.sp
. Gid
omen
Mam
bera
no R
, IND
KC13
3601
.1KC
1340
86.1
Glos
sole
pis
dorit
yiL.
Nen
ggw
ambu
, IND
KC13
3604
.1KC
1340
89.1
G. in
cisu
sL.
Sen
tani
, IND
KC13
3605
.1KC
1340
91.1
G. k
abia
ISe
pik
R, P
NGKC
1336
06.1
KC13
4092
.1
G. le
gget
tiTi
raw
iwa
R, IN
DKC
1336
08.1
KC13
4094
.1
Mel
anot
aeni
a ja
pene
nsis
Yape
n Is
, IND
KC13
3614
.1KC
1341
00.1
M. p
raec
oxKa
li Ti
ri, E
of D
abra
, IND
3#1
–2 K
C133
602.
1, #
3 KP
3458
74#1
KC1
3408
7.1,
#2
KP34
5857
, #3
KP34
5858
M. p
raec
oxBi
ri, M
ambe
rano
R, I
ND2
#1–2
KP3
4587
0#1
–2 K
P345
859
M. p
raec
oxPa
gai,
Mam
bera
no R
, IND
3#1
KP3
4587
1, #
2 KP
3458
72, #
3 KP
3458
73#1
KP3
4586
0, #
2 KP
3458
61, #
3 KP
3458
62
M. r
ubrip
inni
sTi
raw
iwa
R tri
b., I
NDKC
1336
13.1
KC13
4099
.1
M. r
ubriv
ittat
aTi
raw
iwa
R, IN
D7
#1–4
, 6–7
KP3
4587
8, #
5 KP
3458
79#1
, 3, 7
KP3
4586
3, #
2 KP
3458
64, #
4 K
P345
865,
#5
KP34
5866
, #6
KP34
5867
M. v
anhe
urni
Mam
bera
no R
, IND
KC13
3603
.1KC
1340
88.1

849
southern New Guinea). Lastly, a seventh lineage contains the recently discovered speciesMelanotaenia mairasi Allen & Hadiaty 2011, known from a single lake. This species wasshown by Kadarusman et al. (2012b) to represent an additional lineage restricted to theLengguru Arch, which was recovered as the sister group to the western lineage.
The current paper describes a new species of Melanotaenia from the Wapoga River systemof northern New Guinea (northwestern Papua Province, Indonesia, Fig. 1) collected by the firstauthor in 1998 while conducting an ichthyological survey in the vicinity of the Siewa explo-ration camp of Freeport Indonesia Mining Company (Allen & Renyaan 2000). It was initiallyidentified as M. praecox Weber & de Beaufort 1922, originally described from theMamberamo Drainage, which lies approximately 100 km to the east. However, geneticanalysis of recently collected tissue samples provided convincing evidence that the Wapogapopulation represents a distinct species.
Materials and methodsCounts and measurements in the description that appear in parentheses refer to the range forparatypes if different from the holotype. Type specimens are deposited at MuseumZoologicum Bogoriense, Cibinong, Java, Indonesia (MZB), National Museum of NaturalHistory, Washington, D.C. (USNM), and Western Australian Museum, Perth (WAM).Specimens of Melanotaenia praecox were also examined at the Naturalis Biodiversity Center,Leiden, Netherlands (formerly at Zoologisch Museum, Amsterdam and therefore bearing theacronym ZMA).
Comparative material examined: Melanotaenia praecox – WAM 26787, 40 mm SL, Pionier -bivak, approximately 2°18`S, 138°00`E, Mamberamo River system; WAM P.31027-001, 9specimens, 30–43 mm SL, near Dabra, 3°16`S, 138°46`E, Mamberamo River system; WAMP.31755-001, 4 specimens, 20–44 mm SL, Tiri Creek, approximately 3°18`S, 138°35`E,Mamberamo River system, about 5 km east of Dabra; ZMA 103.047, 40.0 mm SL,Pioniersbivak, Mamberamo River system; ZMA 103.142 (lectotype), 46.8 mm SL,Pioniersbivak; ZMA 110.165 (paralectotypes), 18 specimens, 28.4–46.5 mm SL.
The methods of counting and measuring are as follows: dorsal and anal rays – the last ray ofthe anal and second dorsal fins is divided at the base and counted as a single ray; pectoral finrays were counted on both sides; lateral scales – number of scales in horizontal row fromupper edge of pectoral-fin base to caudal-fin base, excluding the small scales posterior to thehypural junction; transverse scales – number of scales in vertical row (excluding small trun-cated scales along base of fins) between anal-fin origin and base of first dorsal fin; predorsalscales – number of scales along midline of nape in front of first dorsal fin; prepelvic scales –number of scales along ventral midline anterior to base of pelvic fins; circumpeduncular scales– number of scale rows completely encircling caudal peduncle at point of least depth; cheekscales – total number of scales covering suborbital and preoperculum; gill rakers – totalnumber of rakers (including rudiments) on first branchial arch, presented in two parts for therespective upper and lower limb of gill arch; standard length (SL) – measured from tip ofupper lip to caudal-fin base; head length (HL) – measured from tip of upper lip to upper rearedge of gill opening; body depth – measured vertically from anal-fin origin to base of firstdorsal fin; body width – measured at level of pectoral-fin base; snout length – measured fromtip of snout to anterior edge of eye; maxillary length – measured from tip of snout to poste-rior end of upper jaw; eye diameter – horizontal measurement across the middle of the eye;interorbital width – least width measured between the bony upper edges of the eye sockets;predorsal, preanal, and prepelvic distances – measured from tip of snout to origin of respec-tive dorsal, anal, and pelvic fins; length of second dorsal-fin base – measured from the origin
850
Tabl
e II.
Prop
ortio
nal m
easu
rem
ents
of s
elec
ted
type
spe
cim
ens
of M
elan
otae
nia
rubr
ivitt
ata
expr
esse
d as
per
cent
age
of th
e st
anda
rd le
ngth
.
Holo
type
Para
type
Para
type
Para
type
Para
type
Para
type
Para
type
Para
type
Para
type
MZB
WAM
WAM
USNM
USNM
MZB
WAM
MZB
WAM
2226
1P.
3145
4P.
3145
443
2453
4324
5322
262
P.31
454
2226
2P.
3145
4Se
xm
ale
mal
efe
mal
efe
mal
em
ale
fem
ale
mal
efe
mal
em
ale
Stan
dard
leng
th (m
m)
48.7
42.2
40.8
39.9
39.6
33.2
32.8
33.2
28.4
Body
dep
th37
.038
.334
.134
.637
.635
.239
.735
.235
.4
Body
wid
th12
.612
.113
.913
.912
.915
.012
.815
.010
.9
Head
leng
th25
.326
.326
.527
.427
.228
.228
.228
.229
.9
Snou
t len
gth
8.0
8.1
7.3
8.1
7.7
7.8
7.6
7.8
7.0
Max
illar
y le
ngth
8.4
8.3
8.4
8.4
8.2
9.1
7.8
9.1
8.7
Eye
diam
eter
7.7
8.9
8.4
8.5
9.1
9.6
9.5
9.6
10.7
Bony
inte
rorb
ital w
idth
10.4
11.2
10.1
11.9
10.8
10.9
10.9
10.9
11.3
Dept
h of
cau
dal p
edun
cle
11.0
11.7
10.3
10.3
11.6
10.7
12.5
10.7
10.9
Leng
th o
f cau
dal p
edun
cle
16.0
15.7
15.9
16.0
13.6
15.7
15.5
15.7
18.2
Pred
orsa
l dis
tanc
e46
.849
.549
.747
.347
.850
.646
.650
.650
.2
Prea
nal d
ista
nce
49.3
49.8
50.0
52.6
48.7
54.2
48.2
54.2
53.0
Prep
elvi
c di
stan
ce36
.936
.837
.038
.638
.640
.135
.940
.140
.2
2nd
dors
al-fi
n ba
se21
.423
.618
.922
.226
.221
.524
.621
.522
.7
Anal
-fin
base
38.0
40.3
34.2
33.5
39.1
32.1
39.8
32.1
31.5
Pect
oral
-fin
leng
th17
.818
.517
.717
.017
.620
.219
.520
.220
.6
Pelv
ic-fi
n le
ngth
13.3
15.2
13.8
13.2
13.3
15.6
13.4
15.6
16.4
Long
est r
ay 1
st d
orsa
l fin
13.9
16.1
12.6
12.9
15.9
12.7
16.7
12.7
16.9
Seco
nd d
orsa
l-fin
spi
ne11
.29.
811
.611
.511
.011
.812
.511
.812
.2
Long
est r
ay 2
nd d
orsa
l-fin
11.8
11.0
12.4
11.3
12.2
12.8
13.9
12.8
17.2
Anal
-fin
spin
e7.
16.
97.
97.
27.
57.
77.
97.
77.
0
Long
est a
nal r
ay10
.511
.512
.710
.812
.811
.511
.711
.515
.1
Caud
al-fi
n le
ngth
19.0
23.5
22.7
21.9
20.9
23.7
19.7
23.7
21.3
Caud
al c
onca
vity
3.8
9.9
6.8
7.5
9.0
6.5
4.4
6.5
6.8

851
of second dorsal fin to the base of last ray of second dorsal fin; length of anal-fin base –measured from the origin of anal fin to base of last ray of anal fin; pectoral, pelvic, andcaudal-fin lengths – measured from base of fin to distal tip of longest ray; caudal peduncledepth is least depth and caudal peduncle length is measured between two vertical lines, onepassing through base of last anal ray and the other through caudal-fin base; caudalconcavity –horizontal distance between verticals at tips of shortest and longest rays.
All of the rainbowfish DNA sequences from the monophyletic “Chilatherina” group and severalfrom the “Glossolepis” group sequenced by Unmack et al. (2013) were phylogeneticallyanalysed, plus new additional sequences obtained from a larger number of individuals from C. alleni, M. praecox, and the new species described here from the Wapoga River system(Table I). We sequenced the complete mitochondrial cytochrome b (cytb) gene and a portionof the S7 nuclear gene (part of exons 1 and 3 and complete exon 2 and introns 1 and 2) andused GARLI 2.0 (Zwickl 2006) to obtain the best maximum likelihood trees and 1000 boot-strap replicates. Methods for obtaining DNA sequence data and their analyses follows Allen &Unmack (2012), Unmack et al. (2013) and Allen et al. (2014) except where noted as follows:S7 sequences were manually phased for only M. praecox and the new species, of those, sevenindividuals had a single heterozygous position, while one (M. praecox Pagai 2) had six (phasedindividuals have the letters a and b to designate each allele); S7 sequences were then alignedusing the online version of MAFFT 7.205 (Katoh and Standley, 2013) using the slow iterativerefinement G-INS-i algorithm with the scoring matrix for nucleotide sequences set to 1PAM/K=2, a gap opening penalty of 1.53 and an offset value of 0.1; the model of sequence evolu-tion GTR+I+G and GTR+G were the best ones identified by ModelTest 3.7 (Posada &Crandall, 1998) for cytb and S7 respectively; we used attachmentspertaxon = 66 and treeswere rooted with species from the “Glossolepis” group. GenBank accession numbers areprovided in Table I for all sequences examined in this study.
Melanotaenia rubrivittata n. sp.Laser Red RainbowfishHolotype. MZB 22261, male, 48.7 mm SL, small pond beside Tirawiwa River, 3° 01.970’S,136° 22.231’ E, Wapoga River system, Papua Province, Indonesia, 0.3 m, seine net, G. Allen& S. Renyaan, 5 April 1998.
Paratypes (collected with holotype unless stated otherwise): MZB 22262, 6 speci-mens, 23.2–43.5 mm SL; USNM 427072, 5 specimens, 26.0–39.3 mm SL; WAM P.31449-001, 6 specimens: 18.0–42.2 mm SL; WAM P.31454-001, 8 specimens, 28.4–41.9mm SL, small pond beside Tirawiwa River, 3° 01.399’S, 136° 21.892’ E, Wapoga Riversystem, Papua Province, Indonesia, 0.3 m, seine net, G. Allen & D. Polhemus, 9 April 1998.
Table III. Summary of dorsal, anal, and pectoral fin-ray, predorsal scale, and cheek scale counts for Melanotaenia rubrivittata.Pectoral-ray counts were taken on both sides of each individual.
First Dorsal-fin spines Soft Dorsal-fin rays Soft Anal-fin rays
IV V VI 9 10 11 12 17 18 19 20 21
4 15 4 1 7 11 4 2 5 9 6 1
Pectoral-fin rays Predorsal scales Cheek scales
10 11 12 13 13 14 15 16 6 7 8 9 10 11
1 27 17 1 3 10 9 1 1 3 1 11 5 2

Diagnosis: A species of melanotaeniid rainbowfish distinguished by the following combina-tion of characters: dorsal rays IV–VI-I,9–12 (usually V-I,10–11); anal rays I,17–21 (usuallyI,18–20); pectoral rays 10–13 (usually 11–12); lateral scales 32–33, predorsal scales 13–16(usually 14–15); circumpeduncular scales 12; total gill rakers on first arch 12–14; total scalescovering cheek (preopercle) 6-11 (average 8.9); snout length 3.2 (3.0–4.3) in HL; averagebody depth of adult males 37.7 % SL; colour in life brilliant neon blue on upper two-thirds ofbody with five red stripes (one between each scale row); ventral portion of head and bodysilvery white to blue with slight pinkish hue; first dorsal fin blue; second dorsal fin blue with redbasal stripe and broad red zone encompassing outer margin and posterior portion of fin; anal
852
Fig. 3. Male specimens of Melanotaenia rubrivittata raised in captivity, approximately 45 mm SL. The nuptial display is shown in the upper photograph. G.L.

fin bluish with red basal stripe and red outer margin; caudal fin red, grading to bluish or translu-cent along posterior margin.
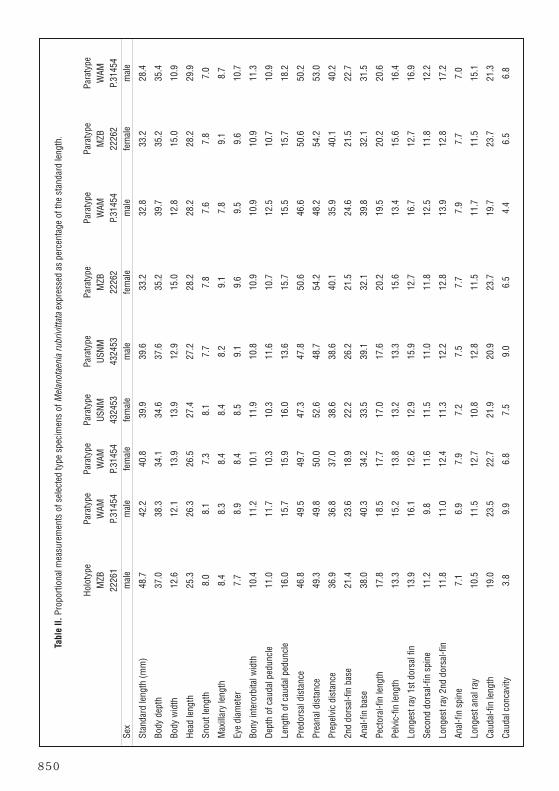
Description (morphometrics based on 17 specimens, 28.4–48.7 mm SL and counts on 23specimens, 26.0-48.7 mm SL; see also Tables II and III): Dorsal rays V-I,10 (IV–VI-I,9–12);anal rays I,19 (I,17–21); pectoral rays 11 (10–13); pelvic rays I,5; branched caudal rays 13(15); principal caudal rays 15 (17); upper and lower procurrent caudal rays 5 (5–6); lateralscales 33 (32–33); transverse scales 10; predorsal scales 15 (13–16); prepelvic scales 16(13–17); cheek scales 9 (6–11); circumpeduncular scales 12; gill rakers on first branchial arch1 + 12 (1–2 + 11–13), total gill rakers on first arch 13 (12–14).
Body depth 2.7 (2.5–3.3) in SL, head length 3.9 (3.3–3.9) in SL; greatest width of body 2.9(2.2–3.2) in greatest body depth; snout length 3.2 (3.0–4.3) in HL; eye diameter 3.3 (2.8–3.2)in HL; interorbital width 2.4 (2.4–3.0) in HL; depth of caudal peduncle 2.3 (2.2–2.7) in HL;length of caudal peduncle 1.6 (1.6–2.0) in HL.
Jaws about equal, oblique, premaxilla with an abrupt, ventrally-directed bend between theanterior horizontal portion and lateral part; maxilla ends below about anterior edge of eye;maxillary length 3.0 (2.9–3.7) in HL; lips thin; teeth conical with slightly curved tips, extendingon to outer surface of lips; teeth of upper jaw in about five irregular rows anteriorly with largestin outer row, reduced to single row posteriorly, where exposed when mouth closed; teeth inlower jaw in about 4–6 irregular rows anteriorly, reduced to 1 or 2 rows posteriorly; narrowrow containing small, conical teeth on vomer and palatines (usually embedded in congealedmucous).
Cephalic sensory pores well developed, consisting of six large pores along margin of preop-ercle, five pores on maxilla, four pores on preorbital, five pores on interorbital/supraocular,and five pores along infraorbital (postocular) canal.
Scales of body cycloid, relatively large, and arranged in regular horizontal rows; scale marginsweakly crenulate; predorsal scales extending forward to about middle of interorbital space;preopercle with 1-2 scale rows between its posterior angle and eye.
Predorsal length 2.1 (2.0–2.1) in SL; preanal length 2.0 (1.8–2.1) in SL; prepelvic length 2.7(2.5–2.9) in SL; length of second-dorsal fin base 4.7 (3.8–5.3) in SL; length of anal-fin base2.6 (2.5–3.2).
First dorsal fin-origin about level with anal fin-origin; longest spine (usually second or third) offirst dorsal fin 1.8 (1.5–2.5) in HL, its depressed tip reaching base of spine of second dorsal
853
Fig. 4. Melanotaenia rubrivittata, preserved male holotype, 48.7 mm SL.. G.R.A.

fin in females and reaching to about base of first soft ray in mature males; second dorsal-finspine 2.3 (1.7–2.7) in HL; longest (generally anteriormost in females and penultimate in adultmales) rays of second dorsal fin 2.1 (1.8–2.4) in HL, depressed posterior rays extending lessthan one-half length of caudal peduncle in females and about one-half to two-thirds length ofcaudal peduncle in mature males; anal-fin spine 3.6 (2.9–4.3) in HL; longest (more or lesssubequal in adult males and females) anal rays 2.4 (1.9–2.5) in HL; pelvic-fin tips whendepressed reaching to base of anal spine or first soft anal ray in mature adults; length of pelvicfins 1.9 (1.7–2.1); length of pectoral fins 1.4 (1.3–1.8) in HL; length of caudal fin 1.3(1.2–1.4) in HL; caudal fin moderately forked, caudal concavity 6.7 (2.7–6.4) in head length.
Colour of freshly captured male in life (Fig. 2): head greyish dorsally, silvery blue on cheek andoperculum; brilliant neon blue on upper two-thirds of body with five red stripes; ventral portionof head and body silvery white to blue with slight pinkish hue; first dorsal fin blue; second dorsalfin blue with red basal stripe and broad red zone encompassing outer margin and posteriorportion of fin; anal fin bluish with red basal stripe and red outer margin; caudal fin red, gradingto bluish or translucent along posterior margin; pelvic fins bluish to translucent with redanterior margin and reddish posterior tip; pectoral fins translucent. Illustrations of captiveaquarium specimens are presented in Fig. 3. Nuptial males are capable of “switching on” agolden-orange mid-dorsal stripe, extending from the dorsal-fin origin to the snout tip (Fig. 3,upper fish).
Colour pattern of freshly captured female in life (Fig. 2, lower): generally similar to that of maleexcept overall pattern is much duller and lacks brilliant red markings on fins; second dorsal andanal fins with blue outer margin.
Colour of holotype in alcohol (Fig. 4): snout and dorsal surface of head bluish-grey, grading totan on propercle and opercle; upper half of body brown with red stripes (see live descriptionabove) replaced by tan markings; longitudinal scale row at eye level slightly darker brown;lower half of body with large brown area below pectoral fin, gradually fading to tan posteri-orly; fins mainly translucent. Female paratypes greyish on snout and dorsal surface of head,tan on preopercle and opercle; upper half of body brown with darker brown scale margins;lower half of body mainly tan; fins translucent.
Sexual dimorphism: Typical of most Melanotaenia, males have a deeper body and longerdorsal and anal fin-rays. The longest soft dorsal-fin rays of males are located in the posteriorportion of the fin, in contrast to that of females, which are situated anteriorly. In addition, thedepressed first dorsal fin of adult males extends to the base of the first soft ray of the seconddorsal fin, compared with the origin of the fin in females.
Body depth generally increases, especially in males, with increased growth and is thereforemost useful as a distinguishing character when assessed on the basis of sex and size range asfollows: males – 40–50 mm SL, 37.0–40.2 % SL (average 38.4, n = 4); males – under 30mm SL, 35.2–39.7% SL (average 37.0, n = 4); all males combined 28.4–48.7 mm SL,35.2–40.2% SL (average 37.7, n = 8); females 40–45 mm SL – 30.0–34.1% SL (average32.1, n = 2); females – under 40 mm SL, 27.5–35.4% SL (average 30.5, n = 12); all femalescombined – 30.2–42.6 mm SL, 27.5–35.4% SL (average 30.7, n = 14) This species reachessexual maturity at a relatively small size as indicated by the presence of ripe gonads, approxi-mately 28.5 mm SL in males and 30.0 mm SL in females.
Remarks: The new species is most similar in appearance to Melanotaenia praecox (Fig. 5)from the Mamberamo River system, lying approximately 100 km to the east. The two speciesshare a number of similarities including a neon-blue ground colour, brilliant red markings onthe median fins, relatively small maximum size (less than about 50 mm SL), overlapping countsfor dorsal rays, anal rays, pectoral rays, predorsal scales, and cheek scales, and most morpho-
854

855
metric proportions. Genetic results (Fig. 6) reveal the two species are clearly separable. Inaddition, M. praecox lacks the characteristic red body stripes of male M. rubrivittata. Themales of M. praecox also tend to be deeper bodied (average depth 41.0% SL, n = 14) thanmales of M. rubrivittata (average depth 37.7% SL, n = 8). There is also a slight difference insnout length with M. praecox generally having a shorter snout (average length 7.2% SL, n =15 versus 7.7% SL, n = 15 for M. rubrivittata). Lastly, M. praecox has fewer lateral scales(usually 29–30 versus 32–33) and frequently has 11 transverse scales (always 10 in M.rubrivittata).
Genetic results and discussion: A total of 44 individuals were sequenced for cytb and 36for S7 which was reduced to 33 sequences in both datasets once identical samples wereremoved. Analysis of the 33 sequences yielded 863/895 invariant characters, 83/40 variablebut parsimony uninformative characters, and 195/67 parsimony informative characters forcytb and S7 respectively. Maximum likelihood analysis recovered one tree for each dataset withlikelihood scores of -4253.549833 and -2260.786885 respectively (Fig. 6). The relationshipsrecovered from cytb were broadly congruent with the larger sequence dataset in Unmack et al.
Fig. 5. Aquarium photographs of Melanotaenia praecox, males, approximately 45 mm SL, vicinity ofDabra, Mamberamo River system, Papua Province. G.R.A.

(2013), albeit with reduced support for some nodes and slightly different relationships due tofewer characters versus the 6827 combined base pairs in Unmack et al. (2013). The S7 resultsare overall quite similar to cytb (except in the cases of two introgressed species, see below), butwith reduced resolution between some closely related species (which is typical for nuclearsequence data with this gene). The S7 results differ slightly from those in Unmack et al. (2013)in that Melanotaenia praecox is nested more deeply within the “Chilatherina” group and theposition of the lineage containing C. fasciata is slightly different. This difference is likely dueto the larger number and type of genes included in Unmack et al. (2013).
We found evidence for mitochondrial introgression between Melanotaenia rubrivittata andChilatherina alleni, based on their sister group relationship for cytb (Fig 6). Introgression is aresult of an initial hybridisation event followed by an extensive backcrossing of the hybrids onlyinto one of the parental species (usually), such that over time genetic material from one of thespecies becomes incorporated into the other (essentially via horizontal transfer rather than viacommon descent). This can arise under various scenarios, such as where one species issubstantially more abundant than the other and thus limited in their choice of conspecificmates. Mitochondrial DNA can provide very strong evidence for this as it is only maternallyinherited and is passed along clonally (meaning there is no recombination, which is quitedifferent to nuclear DNA). Introgression has been recorded between a number of rainbowfishspecies, usually between those with sympatric distributions (Unmack et al. 2013). Clearly thisintrogression is not recent given there is a pairwise p-distance of between 1.0–1.6% for cytbbetween individuals within both species. As with the morphological evidence presented above,nuclear sequence data clearly place M. praecox and M. rubrivittata as sister species well
856
Fig. 6. Maximum likelihood trees for selected rainbowfish species based on analysis of mitochondrialcytochrome b sequences (left) and nuclear S7 sequences (right). Bootstrap values were obtained from1,000 replicates. Numbers after species names represent the individual within that population, Romannumerals match those used in Unmack et al. (2013). For S7, the letter a or b indicates different phasedalleles for heterozygous individuals.

857
separated from each other by a greater sequence divergence than most sister species shownin Fig. 6. Note that C. alleni is placed in the same location for both cytb and S7 relative toother Chilatherina species, but quite distant from M. rubrivittata for the nuclear gene (Fig.6). The same patterns of relatedness can be see with Melanotaenia vanheurni which hasexperienced introgression with an ancestor to Chilatherina fasciata and its closely relatedsister species (which they would have been sympatric with), but is clearly placed very differentlywith nuclear sequence data in the “Glossolepis” group (Fig. 6).
The phylogenetic position of M. praecox and M. rubrivittata within the “Chilatherina” groupis one of the more interesting relationships within rainbowfishes. Neither species shares muchresemblance to Chilatherina species (or any other melanotaeniid for that matter) based ongeneral appearance. Molecular clock estimates place the divergence of the ancestor to M. praecox and M. rubrivittata and its sister group at a mean age of 9.9 million years(Unmack et al. 2013). Even if only approximately accurate, this leaves a long time for diver-gence especially given how quickly rainbowfishes can evolve differences.
Distribution, habitat and zoogeography: The new species is known only from theWapoga River system in the vicinity of Siewa, former site of a Freeport Mining Companyexploration camp. This location (Fig. 1) is approximately 100 km northeast of Nabire, a rela-tively large town lying on the edge of Cenderawasih Bay. The type locality (Fig. 7) is situatedabout 130 km upstream in the Wapoga system at an elevation of approximately 70 m on theedge of a broad floodplain adjacent to nearby foothills. At the time of collection (April 1998)it consisted of a small pond, resulting from previous flooding of the nearby Tirawiwa (some-times spelt Tiawiwa) River. The pond was fully shaded and about 12 m in diameter with anaverage depth of only 30 cm. The substratum was composed of soft mud covered with a layerof organic debris, mainly dead leaves. Additional paratypes were collected in a small (2–5 mwidth), closed-canopy rainforest stream 1.3 km northwest from the type locality. The newspecies and Mogurnda wapoga Allen et al. 1999 (Eleotridae) were the only fishes collected atboth sites, although a few juvenile Glossolepis leggetti Allen & Renyaan 1998 (Melano -taeniidae) were obtained from the pond habitat. Temperature and pH values of 27.0°C/6.6and 28.7°C/8.0 were recorded for the respective pond and stream habitats. Specimensobtained for genetic analysis in 2012 from the same general area were found in a small (about
Fig. 7. Floodplain habitat of Tirawiwa River new Siewa, West Papua Province and type locality (right) ofMelanotaenia rubrivittata. G.R.A.

1.5–2 m wide) rainforest stream that was also inhabited by Melanotaenia rubripinnis Allen &Renyaan 1998 and Chilatherina alleni Price 1997. The Wapoga system is an importantlocation for freshwater fish endemism with all the previously mentioned melanotaeniids knownthus far only from this region, except C. alleni, which ranges to the Siriwo River drainage,approximately 70 km to the southwest. In addition, Mogurnda wapoga and Sicyopteruserythropterus Keith et al. 2012 (Gobiidae) are known only from this area.
Etymology: The new species is named rubrivittata (Latin: red-stripes) with reference to thedistinctive markings that distinguish it from the similar appearing M. praecox.
Acknowledgements:We are grateful for the generous support of the National Geographic Society for funding (NGS grant 5215-94) thatenabled the first author to survey fishes in western New Guinea from 1995–1999. Collection and specimen exportpermits during this period were provided by Lembaga Ilmu Pengetahuan Indonesia (LIPI). We also thank ConservationInternational and the Freeport Indonesia Company for providing funds and logistical support for the 1998 Wapogasurvey. Thanks are also due to Samuel Renyaan, who accompanied the first author on this expedition. The Wapogacollections were assisted by Burke Burnett, Mike Moore, Hendrite Ohee, Dan Polhemus, and Sylvester Tenege. Criticaltissue samples of the new species were generously donated by Dan Dority, Johannes Graf, and Gary Lange. We are alsograteful to Gary Lange for his superb live photographs of the new species. Tarmo Raadik and Martin Gomon bothprovided excellent feedback on our earlier drafts.
References:Allen, G. R. 1980. A genetic classification of the rainbowfishes (family Melanotaeniidae). Records of the Western
Australian Museum 8: 449-490.Allen, G. R. 1995. Rainbowfishes in Nature and in the Aquarium. Tetra Verlag, Melle, Germany,180 pp.Allen, G. R. & Hadiaty, R. K. 2011. A new species of rainbowfish (Melanotaeniidae), from western New Guinea (West
Papua Province, Indonesia). Fishes of Sahul 25 (1): 602-607.Allen, G. R. & Hadiaty, R. K. 2013. Melanotaenia sneideri, a new species of rainbowfish (Melanotaeniidae), from West
Papua Province, Indonesia. aqua, Journal of Ichthyology and Aquatic Biology 19 (3): 137-146.Allen, G. R., Jenkins, A. P. & Renyaan, S. J. 1999. Mogurnda wapoga, a new species of freshwater gudgeon (Eleotridae)
from Irian Jaya. Revue française d’Aquariologie Herpétologie 26 (1-2): 63-66.Allen, G. R. & Renyaan, S. J. 1998. Three new species of rainbowfishes (Melanotaeniidae) from Irian Jaya, Indonesia.
aqua, Journal of Ichthyology and Aquatic Biology 3 (2): 69-80.Allen, G. R. & Renyaan, S. J. 2000. Chapter 6. Fishes of the Wapoga River system, northwestern Irian Jaya, Indonesia.
In: A biological assessment of the Wapoga River area of northwestern Irian Jaya, Indonesia (Eds. A. L. Mack & L. E.Alonso). RAP Bulletin of Biological Assessment 14, Conservation International, Washington, DC.
Allen, G. R. & Unmack, P. J. 2012. A new species of rainbowfish (Chilatherina: Melanotaeniidae), from the Sepik RiverSystem of Papua New Guinea. aqua, International Journal of Ichthyology 18 (4): 227-237.
Allen, G. R., Unmack, P. J. & Hadiaty, R. K. 2008. Two new species of rainbowfishes (Melanotaenia: Melanotaeniidae),from western New Guinea (Papua, Barat Province, Indonesia). aqua, International Journal of Ichthyology 14 (4):209-224.
Allen, G. R., Hadiaty, R. K. & Unmack, P.J. 2014. Three new species of rainbowfishes (Melanotaeniidae) from the BirdsHead Peninsula, West Papua Province, Indonesia. aqua, Journal of Ichthyology and Aquatic Biology 20 (3): 139-158.
Kadarusman, Hadiaty, R. K., Segura, G., Setiawibawa, G., Caruso, D. & Pouyaud, L. 2012a. Four new species of rain-bowfishes (Melanotaeniidae) from Arguni Bay, West Papua, Indonesia. Cybium 36: 369-382.
Kadarusman, Hubert, N., Hadiaty, R. K., Paradis, E. & Pouyaud, L. 2012b. Cryptic diversity in Indo-Australian rainbow-fishes revealed by DNA barcoding: implications for conservation in a biodiversity hotspot candidate. PloS one 7(7):e40627.
Kadarusman, Sudarto, Paradis, E. & Pouyaud, L. 2010. Description of Melanotaenia fasinensis, a new species of rain-bowfish (Melanotaeniidae) from West Papua, Indonesia with comments on the rediscovery of M. ajamaruensis and theendangered status of M. parva. Cybium 34: 207-215.
Katoh, K. & Standley, D.M. 2013. MAFFT multiple sequence alignment software version 7: improvements in perform-ance and usability. Molecular Biology and Evolution 30: 772-780.
Keith, P., Allen, G. R., Lord, C. & Hadiaty, R. 2012. Five new species of Sicyopterus (Gobioidei: Sicydiinae) from PapuaNew Guinea and Papua. Cybium 35 (for 2011): 299-318.
McGuigan, K., Zhu, D., Allen, G. R., & Moritz, C. (2000). Phylogenetic relationships and historical biogeography ofmelanotaeniid fishes in Australia and New Guinea. Marine and Freshwater Research 51: 713-723.
Meinken, H. 1974. Mitteilung der Fischbestimmungsstelle des VDA, Nr. 77. Aus Neu-Guinea kommt eine neue Gattungund Art der Ährenfische (Pisces, Atherinidae). Aquarium Aqua Terra 8 (55): 9-11.
858

859
Posada, D. & Crandall, K. A. 1998. ModelTest: testing the model of DNA substitution. Bioinformatics 14: 817-818.Price, D. S. 1997. Chilatherina alleni, a new species of rainbowfish (Melanotaeniidae) from Irian Jaya. Revue française
d’Aquariologie Herpétologie 24 (3-4): 79-82.Regan, C. T. 1914. Report on the freshwater fishes collected by the British Ornithologists’ Union expedition and the
Wollaston expedition in Dutch New Guinea. Transactions of the Zoological Society of London 20 (6:1): 275-286.Tappin, A. 2011. Rainbowfishes, their Care and Keeping in Captivity. 2nd Edition, published online at http://rain-
bowfish.angfaqld.org.au/Book.htm Electronic version accessed 9 November 2014.Unmack, P. J., Allen, G. R. & Johnson, J. B. 2013. Phylogeny and biogeography of rainbowfishes (Melanotaeniidae)
from Australia and New Guinea. Molecular Phylogenetics and Evolution 67: 15-27.Weber, M. & de Beaufort, L. F. 1922. The fishes of the Indo-Australian Archipelago. IV. Heteromi, Solenichthyes,
Synentognathi, Percesoces, Labyrinthici, Microcyprini. E. J. Brill, Leiden. 410 pp.Zhu, D., Jamieson, B. G., Hugall, A. & Moritz, C. 1994. Sequence evolution and phylogenetic signal in control-region
and cytochrome b sequences of rainbow fishes (Melanotaeniidae). Molecular Biology and Evolution 11, 672-683.Zwickl, D. J. 2006. Genetic algorithm approaches for the phylogenetic analysis of large biological sequence datasets
under the maximum likelihood criterion. Ph.D. dissertation, The University of Texas at Austin.
1Western Australian Museum, Locked Bag 49, Welshpool DC, Perth, Western Australia 6986. E-mail:[email protected] for Applied Ecology and Collaborative Research Network for Murray-Darling Basin Futures, University ofCanberra, ACT 2601, Australia.3Museum Zoologicum Bogoriense (MZB), Division of Zoology, Research Centre for Biology, Indonesian Institute ofSciences (LIPI), Jalan Raya Bogor, Km 46, Cibinong 16911, Indonesia.
WERE THOSE RED LASER “STRIPES” REALLY WORTH IT? – THE HUNT FOR THE
WAPOGA RED LASER RAINBOWFISHGary Lange, Johannes Graf and Dan Dority
Can you remember when you had a close call in your car and almost crashed, cheating injurymaybe by inches? How about as a youngster jumping off of a tall cliff into the lake then secondguessing yourself after it was too late to turn back, hoping things didn’t end badly. “I shouldn’thave been following that other car so closely” and “How did I let my friend Randy talk me intosuch a stupid decision as jumping off this cliff” were the thoughts racing through my mind inthose moments. And here I was in a pathless jungle, up the creek without a paddle. Actuallywithout the boat, because they had abandoned us and now we were several hours into a verylong hike. Those thoughts were again running through my mind that I had indeed made a questionable decision.
But let’s back up a few months to January of 2012. It’s time to plan another trip withJohannes Graf and Dan Dority. Where should we go for new fish and adventure? We didn’thave to look any further than the cover of Aqualog’s “All Rainbowfish”. Let’s go to the Siewaairfield (Figure 1), home of Glossolepis leggetti and Melanotaenia rubripinnis. This was alsohome to the Aqualog cover fish Chilatherina alleni and of course that super electric blue “M.praecox” with the striking red laser stripes thru its body. These were fish that Dr. Gerry Allenhad photographed from a 1998 expedition but none were brought back alive. It was easy tosay you want to go there but now much harder to actually get there. The Siewa airstrip builtfor mining exploration was no longer in use and overgrown, the jungle had taken it back. Theonly way would be to take a small boat from Nabire travelling on the ocean for five hours andthen up Sungai Wapoga and its tributary, Sungai Tirawiwa, for another eight hours to get asclose to the Siewa airstrip as possible. We badly wanted to collect the wonderfully yellowChilatherina alleni and especially that electric blue “striped praecox”. We have had more thana few conversations with aquarists wondering out loud whether that cover picture was“doctored” because the blue and the red stripes were so intense. Hopefully we would find outfor ourselves.

Our first stroke of luck was that Dan made connections with someone in Nabire that might beable to help us, a friend of a friend as it were. Anderson, or Andy was able to also find us aboat captain, who said he could take us where we wanted to go. That always sounds good inthe emails but you wonder if things will change once you get there. We made our flight fromJayapura to Nabire and met up with Andy and eventually our captain for the trip, Guntur. Soonafterwards we would learn of our first “problem”. This was our third trip together so we nowhave as our motto “It’s not a problem, it’s an adventure”. This long journey would need a lotof gas. The captain couldn’t just take his collection of 20 litre gas cans to the gas station andhave them filled; that would be too easy. That would be hoarding and that wasn’t allowed, youcould only get about 35 litres at a time and it could only be pumped into an automobile. Thoseare the rules, so the captain had to go fill up his car, drive to the harbor, siphon the gas intothe containers and repeat over and over. This probably would have only taken him a full dayexcept for another problem, electricity. Electricity doesn’t always run 24/7 in Nabire and itwas acting up again. That evening the captain came back and told us it might take another dayor two before we could get enough gas for the trip. Instead of moping around we decided tohead off on another adventure for a few days (see FOS Vol27#4 762–772).
Two days later we started on our long delayed journey. Our vessel was a large dugout outriggercanoe with two 40-horsepower motors to speed us on our way. It had a very small cabin thattwo people could barely get into to avoid sun or rain but a great place to store our gear. It’swonderful speeding along the beautiful blue ocean heading up the coast toward our destina-tion for the first hour. By the time we got to the stop some five hours later at Wireri we werequite tired of our boat ride. The captain had family here so it made it a convenient place torest. We were the entertainment for much of the village that evening as it wasn’t often thatwhite people came this way. One of the older villagers who was familiar with the collectingarea would also continue with us in the morning. We had a nice dry bed on the chief’s patio
Fig 1. GPS trace from Nabire to Wireri to Wapoga collection site and back (Google Earth Map)
860

861
Melanotaenia sp Wapoga Red Laser constantly moving and chasing G.L.
and with a few tarps lashed to the side poles it kept us dry from the relentless rain that poundedus through the night.
We were up quite early the next morning and on our way by 7 am. There is only so muchdaylight in the tropics and when the sun sets at six pm it gets very dark very quickly. Soon wewere at the mouth of the mighty Sungai Wapoga. It’s quite wide, some 2 km where it entersthe ocean. Soon afterwards it narrows to about 500 meters and stayed that width for hours aswe twisted through turn after turn on the river. The torrential rains from the night before hadwashed a lot of debris into the river; it was a virtual traffic jam of logs that our captain had toexpertly navigate. His pilot was in the back of the boat manning the outboard motors. Thecaptain perched high on the roof of the tiny cabin could see most of the dangers in the muddywater, the moving ones and the sunken logs that were waiting to rip into our canoe or tear upour motor or, even worse, to crash the outriggers. Logs that were much larger than our canoeshot by in the swift current. The captain motioned his pilot with hand signals and the two ofthem did an excellent job steering clear of most of the debris.
Finally we turned into Sungai Tirawiwa and the massive log assault ceased. We made goodtime for a while but the river was getting narrower with every turn. We could see that this riverwas not so muddy or silty but rather dark with tannins, a good sign for finding bright colouredfishes. Soon the outriggers were slicing through the grasses on the sharper turns. We were still a long way from the original Siewa airstrip, a distance too far to hike. The captain startedto pull his boat into a spot where he had camped before. Dan talked him into going furtherand so Guntur continued onward. He had never gone that far upstream before. To us it seemedlike maybe only fifteen or twenty minutes further but it was that much closer to the airstriplocation. Finally we could go no further the river was only as wide as the outrigger itself.
It was time to set up our tarp “tent” for the night. Soon it was raining again. Even though wewere several feet above the river level we wondered if we would end up in the river later thatnight. If the rains came down as hard as they did the night before it was a distinct possibility.We had a feast of rice, ramen noodles and canned mackerel as well as a hot cup of instantcoffee. So off to crawl under the mosquito netting to try to get some sleep. It wasn’t thatcomfortable but we were close to realizing our dreams and capturing some remarkable fishes.

Top: “The Outrigger” – our transportation for this adventure. J.G.Above (left) Dan and Johannes on the way back to Nabire (G.L.), (right): Gary after rainstorm. J.G.Below: Anderson (Andy) waiting on the long boat ride to Wireri. G.L.
862

863
Melanotaenia praecox – Pagai; beautiful but the colours aren’t quite as intense as the fish from Wapogaand it also gets much broader as it matures. G.L.
What would tomorrow morning bring? That made it harder to sleep than the wet ground, theheat or the buzzing mosquitoes. The fruit bats fluttered above in the canopy during the nightand in the morning we also discovered a large pile of fresh cassowary dung not too far fromour campground.
In the morning we set off right away to catch our fishes, it was still raining just enough to getyou good and wet. The captain and his pilot would break camp and then wait for us to return.He told us that we had to be back by noon. The older Papuan came with us because he hadbeen here many years before. The plan was to find the old logging road which was visible onGoogle Earth maps. Well in the jungle it’s “use it or lose it” and the road when we finally foundit was worse that walking through the uncut forest. We quickly realized that we were not goingto make it all of the way to the airstrip and started checking the streams around us. They wereonly one half meter wide and maybe 6 cm deep and yielded no fish.
A little further on we found a larger 2 m wide creek. The creek was nothing but slippery mudand a few feet of tannin soaked water. It was an adventure just to get into the water withoutfalling. No rocks on the bottom just more mud to sink into and the occasional thorny stick totear at your legs or get caught in the seine. Our fishing method was to have two people holdthe seine across the width of the creek. The other two would go upstream a few feet and thenattempt to run towards the seine splashing and jumping hoping to herd some fish into thewaiting net. It was not an easy task and the “runners” had to spread their toes wide in theirshoes to avoid losing a sneaker to the mud.
Our first fish were Melanotaenia rubripinnis followed shortly by Chilatherina alleni. Still noelectric blue and red striped fish in our nets though. Around the next bend was a large pile ofdead wood in the water that looked like a good place for snakes or fishes. Naturally we beatinto the woodpile chasing imaginary fishes downstream into our waiting seine. But this timeour efforts were rewarded with a bright blue flash in the net. We had our “striped praecox”.The colours were as intense as the cover of the Aqualog. The fish in these small muddy creekswere quite sparse so we had to work hard for what we caught. The fish gods were not goingto give them up easily, we would have to earn them. We also realized that we had again proventhe myth to be false, that you can only have one Melanotaenia species per creek or river. After

Large Melanotaenia rubripinnis; too large to transport so we set him free. G.L.
hours we finally had enough fishes and placed the C. alleni, the “striped praecox” and M.rubripinnis into breather bags for the short walk back to the boat. Photos would have to wait,it was still raining off and on and we had a boat to catch. We headed back to the main riverabove our camp spot. We decided to have a few pulls of the seine here in this now somewhatclear fast running water with an easy to fish gravel bottom. And with that one haul we hadfound our missing fish, Glossolepis leggetti. They were all pretty small so we quickly stashedthem into bags and continued our walk back to the boat.
When we got back to the camp there was no boat! The old Papuan then told Dan that thismight be a possibility if the river dropped and it indeed had dropped quite a bit. No worries,the captain will be meeting us at the first place he wanted to camp which seemed to us about15 or 20 minutes downriver. There are no trails in this area so walking was not easy to saythe least. We were in swimming trunks and water shoes not hiking boots and long pants. Wepaid the price with a great many scratches from thorns.
That 15-20 minute hike to the first camp ground ended up being over a five hour walk. Wegot back to the boat just before dark. Instead of making camp for the night the captain decidedto push on to Wireri. We were exhausted so it seemed like it might be a good idea. When youtravel at night you get to see wondrous things like trees that attract blue fireflies. The captain
Chilatherina alleni from Wapoga. Just as yellow and bright as the cover of the Aqualog. G.L.
864

865
Melanotaenia rubripinnis Wapoga – The lone male that survived the ordeal. G.L.
used his torch sitting in the front of the boat sweeping back and forth to look for logs beatingon the left or right side of the bow to signal the pilot which way to turn. It was then that werealized that there indeed were a lot of “red eyes” on the shore especially when we got to thelarger section of the river. Away from civilization there still were a lot of crocodiles in thesewaters. Exhausted we decide to just leave the fish in the boat’s cabin out of the rain. Thisalmost proved to be a fatal mistake. Sometime during the march some bags leaked and thebreather bags were no longer breathing. The G. leggetti and M. rubripinnis were not goingto make it and we weren’t sure we would have enough C. alleni or “striped praecox” to makea breeding group either.
In the end we did successfully bring back enough of the C. alleni and striped praecox to breedthem and get them out to other hobbyists in North America and Europe.
So were those extra red Laser stripes in our “Wapoga praecox” worth all of that effort? Firstof all after getting samples back in Dr. Unmack’s hands he told us that it was indeed a uniquespecies, now formally named M. rubrivittata. From the hobbyist’s point of view we could alsotell that they behaved very differently than Melanotaenia praecox. We had recollectedMelanotaenia praecox from the Mamberamo system back in 2008 and the two fish lookedand behaved a lot differently. First of all the M. praecox from Pagai and elsewhere becomevery broad (from dorsal to anal fin) as they mature. As one of the smaller rainbowfishes theyhave subdued behaviour in our hands, meaning they are very peaceful and rather slow movingcompared to some other rainbowfishes.
The “Striped praecox from Wapoga” or as Gary calls them “The Wapoga Red LaserRainbowfish” are a much more active fish. The dominant male is constantly chasing the othermales and always flashing at the females. They move so quickly as to be quite a challenge tophotograph. The blue on the body is indeed brighter and deeper in colour than the truepraecox. As they mature they do not get anywhere near as broad as true Melanotaeniapraecox do. Even though we collected both species in sparse tannin laden or even muddycreeks both really appreciate a planted tank and show their best colours when kept that way.Like their cousin though the “Wapoga Red Lasers” are quite prolific with fry that are easy torear. Feed them well, change the water and soon you will have eggs and fry. In conclusion thiswas the adventure of a lifetime and yes “those red laser stripes” were really worth it.

A COMMUNITY HERO – THE DWARF FLATHEAD GUDGEONDavid Shoesmith
The amazing variety of habitats that Australian native freshwater fishes occupy, and the subsequent variety of fishes that occur across those habitats, often means that a communitytank of even relatively peaceful natives is limited in what can be mixed. This is especially sowhen dealing with fishes from the cooler parts of the country, where temperature and salinitycan be critical for survival of individual species, e.g. do not expect fish like Spotted Galaxias(Galaxias truttaceus) to survive in a typical rainbowfish tank. The main reason why speciestanks are so popular and useful is to prevent hybridisation, but another reason is that some fishjust don’t mix!
I have kept a large number of different species and want to draw attention to one of them thatI think is a real community tank champion, the Dwarf Flathead Gudgeon, (Philypnodonmacrostomus). These guys are widespread in south eastern Australia, everywhere fromWilsons Promontory (Victoria) up the east coast to just north of Bundaberg (Queensland) andin parts of the Murray-Darling Basin, primarily along the Murray River proper, especially in thelower sections in South Australia.
They are easy to catch, as their benthic habits lead to them often being stationary amongstplants or debris with the survival instinct to stay put or to move quickly to the nearest plant ordebris and stay put! They will swim into bait traps readily, when the trap is placed on thebottom of the waterway. However I have caught them mostly by dip netting, by scooping alongthe bottom in the spots where water flow is slowest, i.e. along the banks in flowing streams oranywhere in still pools. When disturbed they will usually flee to the nearest vegetation or otherunderwater obstacle – but if they consider themselves already hidden then they will stay still,
866
A typical Dwarf Flathead Gudgeon habitat – The Murray River in South Australia. P.J.U.

867
i.e. if they’re already in some vegetation just pass your dip net through it and they’ll end up inyour net.
They don’t mind some variety in water quality, especially salinity, as they can be found in lovelyclear streams or in muddy salty estuarine waters. I found some on the recent ANGFA combinedclubs field trip in the lovely clear flowing freshwater Friday Creek in the Orara valley inlandfrom Coffs Harbour. Over the years I have collected them on the south coast of NSW in shortcreeks flowing into closed estuaries, and in large numbers on the muddy or sandy bottoms inclosed estuaries that have a decent inflow of fresh water. I have kept them in freshwater andbrackish tanks with no noticeable change in their health or behaviour. Temperature is not anissue for them, in that I have collected them in all seasons in Victoria, New South Wales andQueensland, and kept them in ambient and heated tanks in Melbourne. That has happenedsporadically over fifteen years and any differences that may exist in the various geographicalpopulations has gone unnoticed by myself. That should not preclude another enthusiast takingnotes over time and looking for differences. Who knows what that may lead to …?
They are universally small and peaceful, don’t be put off by the reputation of the much larger-growing Flathead Gudgeon (Philypnodon grandiceps) – the latter is a nasty piece of workwhen it comes to any other small fishes, including rivals amongst their species. I have keptDwarf Flathead Gudgeon with Empire Gudgeon (Hypseleotris compressa), Smelt (Retropinnasemoni) and various Rainbowfishes, Galaxiids, Blue-eyes, Glassfishes, southern Gobies andGlass Shrimps. I have not seen them interact with each other in my aquariums, so no schoolingor breeding behaviour has been observed. However there are plenty of places for fish that sizeto carry on some private business, and perhaps I have just been too casual to notice. I havecertainly seen the males looking a bit spiffed up, hoping to impress I expect, always polite,never aggressive – but can’t reveal anything more. One reason is probably that I have alwayskept them with other species! In all cases their presence made the tank more interesting.
Philypnodon macrostomus – the Dwarf Flathead Gudgeon. Photographer unknown

They are typically 1 to 3 cm when caught, and grow to about 3 cm maximum size. They seemto acquire that size relatively quickly and in my aquariums they survived for up to three years.They are generally brown to grey in body colour, with transparent fins with pale striping. I havenot bred them, so cannot be sure but the males and females are quite different in appearance.The male has two main distinguishing features; firstly they end up with a wide and long mouththat splays out from his head and extends to behind their eyes, secondly they have distinctstriping in their dorsal fin with an orange edge which varies in intensity. The female has a verysmall mouth and is generally paler and greyer than the male.
Scavenging is not beneath them, they will clean up any uneaten food that makes it to thebottom of the tank. In fact you need to ensure that some food does make it the bottom so theycan feed. Having some reliable and efficient scavengers is very handy for maintaining tankhealth and makes the addition of a few Dwarf Flathead Gudgeon in every tank without largepredators a great idea in my experience! Sometimes they will hover in mid-water and feed onpassing food, they will also come to the surface to feed if there are not a lot of other fishflashing around feeding at the surface. Of course they love live foods, but do not make a fussabout accepting dried or frozen prepared foods. I’ve tried all of my commercial dried foodoptions on them and as long as they can fit it into their mouth they will eat it, i.e. flake to verysmall sinking pellets. They’ll take flake off the surface, in mid water, or the next day off thegravel. In this sense I like to think of them as an Australian Corydoras in the way that theyfascinate, scavenge and hardily take a variety of water conditions.
They’re so available (if you live near some to collect, but otherwise hard to come by), hardy,peaceful, fun and native that I think most people would enjoy keeping them. You don’t haveto take my word for it – Doc Bruce Hansen recommends them as fun to collect with the kidsby rock-rolling in a shallow creek, and he has also seen plenty by spotlighting in the shallowsat night.
They are hardier (certainly easier to feed) than Carp Gudgeons, more peaceful than FlatheadGudgeon, and do not grow up to devour your other fishes. They are fascinating to watch inan aquarium, complement other native fishes in their appearance and behaviour, and arepretty unique as not many people seem to keep them. “Can’t be bothered” hey? Well you’remissing out. In fact, the whole experience from locating and capturing them through tokeeping them healthy and happy at home is enjoyable. You should try some of these little fish– go for a community tank and they’ll play their part to a tee.
868
ANGFA (Australia New Guinea Fishes Association) Incorporated.Postal Address: PO Box 1040, Richmond North, Victoria, 3121, Australia. http://www.angfa.org.auFISHES OF SAHUL is printed and published by ANGFA and is distributed free to all financial members.HON EDITOR/DESIGN: Ken Smales. [email protected] COORDINATOR: Neil Armstrong. [email protected] TREASURER: John Lenagan. [email protected] MEMBERSHIP OFFICER: Derek Tustin. [email protected] direct reprint request to authors via membership officer.ANGFA invites submission of articles by its members as well as non-members who would like to contribute. Please post to aboveaddress or contact the editor or coordinator for instructions or assistance.Proof reading for this issue of the journal by: Ron Bowman and Glenn Briggs.Photos in this issue of the journal by: Gerald R. Allen (G.R.A.), Johannes Graf (J.G.), Gary Lange (G.L.), David Shoesmith (D.S.)and Peter J. Unmack) (P.J.U.).© photographs: actual photographer. © text: the author(s). Printed by Galloping Press, Kirrawee, NSW





























