Embed Size (px)
Citation preview

JPET #203356
1
1. Title page
Title: Metabolism and pathways for denitration of organic nitrates in the human liver
Authors: Mirco Govoni, Paola Tocchetti and Jon O Lundberg
Laboratories of origin: Department of Physiology and Pharmacology, Karolinska
Institute, Nanna Svartz väg 2, S-177 76 Stockholm, Sweden (M.G., J.O.L) and NicOx
S.A., Les Taissounières - 1681 Route des Dolines - Sophia Antipolis Cedex, France
(M.G., P.T.)
JPET Fast Forward. Published on April 17, 2013 as DOI:10.1124/jpet.113.203356
Copyright 2013 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
2
2. Running title page
a) Running title: Metabolism of organic nitrates in the human liver
b) Corresponding author: Department of Physiology and Pharmacology, Karolinska
Institute, Nanna Svartz väg 2, S-177 76 Stockholm, Sweden and NicOx S.A., Les
Taissounières - 1681 Route des Dolines - Sophia Antipolis Cedex, France; Telephone:
+39 3398657744, Fax: +46 8 332278, email: [email protected]
c) Number of text pages: 32; Number of tables: 0; Figures: 9; References: 36; Number of words in the Abstract: 250; Number of words in the Introduction: 747; Number of words in the Discussion: 1234; d) Nonstandard abbreviations:
CYP, cytochrome P-450; CYP1A2, cytochrome P-450 1A2; GST, glutathione S-
transferase; GTN, glyceryl trinitrate; HCT 1026, [1,1’-biphenyl]-4-acetic acid,2-fluoro-
a-methyl,4-(nitrooxy)butyl ester; HCT 1027, [1,1’-biphenyl]-4-acetic acid,2-fluoro-a-
methyl,4-hydroxybutyl ester; ISMN, isosorbide mononitrate; NCX 2057, 3-(4-hydroxy-
3-methoxyphenyl)-2-propenoic acid 4-(nitrooxy)butyl ester; NCX 2059, 3-(4-hydroxy-
3-methoxyphenyl)-2-propenoic acid 4-hydroxybutyl ester; NO, nitric oxide; NO2-,
nitrite; NO3-, nitrate; NOBA, (nitrooxy)butyl alcohol; NOC-5, 3-(aminopropyl)-1-
hydroxy-3-isopropyl-2-oxo-1-triazene; NOx, (NO3- + NO2
-); RSNO, S-nitrosothiols;
sGC, soluble guanylate cyclase;
e) Recommended section: Metabolism, Transport, and Pharmacogenomics
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
3
Abstract
Liver first-pass metabolism differs considerably among organic nitrates but little
information exists on the mechanism of denitration of these compounds in hepatic
tissue. The metabolism of nitrooxybutyl-esters of flurbiprofen and ferulic-acid, a class
of organic nitrates with potential therapeutic implication in variety of different
conditions, was investigated in comparison with glyceryl trinitrate (GTN) in human
liver by a multiple approach, using a spontaneous metabolism-independent NO donor
(NOC-5) as a reference tool. Nitrooxybutyl-esters were rapidly and quantitatively
metabolized to their respective parent compounds and the organic nitrate moiety
nitrooxybutyl-alcohol (NOBA). Differently from GTN which was rapidly and
completely metabolized to nitrite, NOBA was slowly metabolized to nitrate. In contrast
to the spontaneous NO donor NOC-5, NOBA and GTN did not generate detectable NO
and failed to suppress the activity of cytochromeP450; an enzyme known to be inhibited
by NO. The direct identification of NOBA following liver metabolism targets this
compound as the functional organic nitrate metabolite of nitrooxybutyl-esters.
Moreover the investigation of the pathways for denitration of NOBA and GTN suggests
that organic nitrates are not primarily metabolized to NO in the liver but to different
extents of nitrite or nitrate depending from their different chemical structure. It follows
that cytochromeP450-dependent metabolism of concomitant drugs is not likely to be
affected by oral co-administration of organic nitrates. However the first-pass may
differently affect the pharmacological profile of organic nitrates in connection with the
different extent of denitration and the distinct bioactive species generated and exported
from the liver (nitrate or nitrite).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
4
Introduction
Organic nitrates have been harnessed therapeutically since 1879 when glyceryl trinitrate
(GTN) was first described as a remedy for angina pectoris (Murrell, 1879). GTN, acting
through the liberation of nitric oxide (NO) in vascular tissue, is still widely used in the
treatment of coronary artery disease and congestive heart failure (Parker and Parker,
1998). This compound is rapidly and efficiently absorbed from the mouth leading to a
rapid onset of action that is suitable for the treatment of acute angina attacks. After
absorption from the mouth or the skin GTN is rapidly denitrated by hepatic metabolism
(systemic t1/2 ~ 1min) (Needleman et al., 1976) to its dinitrate form (GDN) which has a
higher systemic half-life (t1/2 ~ 2h) (Needleman et al., 1976) but a lower potency than
GTN (Münzel et al., 2005). When GTN is given by oral route it undergoes extensive
liver first-pass metabolism (Needleman et al., 1972) leaving its dinitrate form to
circulate systemically with little or no parent compound appearing in blood (Needleman
et al., 1976l; Yu et al., 1988). For this reason GTN is administered orally only for
prophylaxis purposes due to its slower onset of action and higher doses required to
reach therapeutic efficacy in comparison to sublingual or transdermal administration.
Efforts to increase its duration of action and prophylaxis have led to the synthesis of
other organic nitrates, of which the most important is isosorbide mononitrate (ISMN).
ISMN has similar pharmacological actions as GTN but it is longer acting and it
undergoes low hepatic metabolism after oral administration (Abshagen, 1992).
Furthermore, the growing realization that nitrates may represent new therapeutic agents
in different areas led to the development of organic nitrates containing adjunct
pharmacophores to introduce biological properties beyond those of the parent
compound (Hodosan et al., 1969; Keeble and Moore, 2002). In particular,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
5
nitrooxybutyl-ester derivatives is a novel class of hybrid organic nitrates consisting of a
(-ONO2) structural unit connected via a butyl linker to the parent compound by a
carboxyl-ester bond.
While the capacity of organic nitrates to liberate NO in vascular tissue is quite well
established (Münzel et al., 2005; Kleschyov et al., 2003; Mulsch et al.1995), the exact
mechanism of denitration is still a matter of debate. Moreover, whether organic nitrates
are at all capable of generating NO in extravascular tissues, as for example in the liver is
still controversial. Some studies have provided evidence for NO generation from GTN
incubated in subcellular liver fractions (Servent et al., 1989; Kozlov et al., 2003); but a
generalized concept (not supported by any data though) is that the liver cannot
metabolize GTN to NO. Indeed, it is well established that GTN undergoes extensive
hepatic metabolism causing loss of vascular bioactivity (Needleman et al., 1972) but
this does not necessarily imply that the liver is not capable of acute NO generation from
GTN. A better understanding of how organic nitrates are metabolized in the liver is
important when characterizing novel organic nitrates that are under development. As an
example, NO has been reported to be directly and indirectly involved in the inactivation
of cytochrome P450 (Khatsenko et al., 1993; Wink et al., 1993; Stadler et al., 1994)
suggesting that liver CYP-dependent drug metabolism may be drastically affected by
oral co-administration of organic nitrates undergoing first pass metabolism.
Nitrooxybutyl-ester derivatives of anti-inflammatory and/or anti-oxidant agents as
flurbiprofen and ferulic acid is a novel class of organic nitrates with potential
therapeutic implication in variety of different conditions (Keeble and Moore, 2002;
Scatena et al., 2004) as neurodegenerative diseases (Wenk et al., 2002; Wenk et al.,
2004, Gasparini et al., 2005, Prosperi et al., 2004), inflammation and nociception
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
6
(Ronchetti et al., 2009), learning and memory (Boultadakis et al., 2010), airway
inflammation and relaxation (Larsson et al., 2007; Larsson et al., 2009), osteoporosis
and bone resorption (Idris et al., 2004). Despite extensive pharmacological
characterization of these compounds, their metabolism and pathways of denitration
remain unknown. In particular, being targeted mainly for chronic diseases these
molecules are generally designed to be administered by oral route and an evaluation of
their liver metabolism is therefore critical.
In the present study we investigated the metabolism of nitrooxybutyl-ester compounds
in comparison with GTN and ISMN in the human liver by a multiple approach that
includes i) a chemiluminescent nitrite/nitrate/RSNO metabolic profile assessment ii) a
test of NO mediated inhibition of liver cytochrome P450 1A2 (CYP1A2) activity iii) a
direct electrochemical detection of NO and iv) the use of a spontaneous metabolism-
independent NO donor (NOC-5) as a reference tool.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
7
Materials and Methods
Biological material and chemicals
Human liver microsomes, cytosol and mitochondria were purchased from Tebu-bio,
Magenta, Italy, as pools of different mixed gender donors. HPLC-grade organic
solvents were purchased from Carlo Erba reagents (Milan, Italy). HPLC-grade water
was prepared with a Milli-Q water purification system. HCT 1026, [(1,1’-biphenyl)-4-
acetic acid,2-fluoro-α-methyl,4-(nitrooxy)butyl ester], HCT 1027, [1,1’-biphenyl]-4-
acetic acid,2-fluoro-α-methyl,4-hydroxybutyl ester, NCX 2057, 3-(4-hydroxy-3-
methoxyphenyl)-2-propenoic acid 4-(nitrooxy)butyl ester and NCX 2059, 3-(4-
hydroxy-3-methoxyphenyl)-2-propenoic acid 4-hydroxybutyl ester were provided by
NicOx SA (Sophia-Antipolis, France). GTN and NOBA were kindly provided by
Dipharma S.p.A. (Milan, Italy). All other chemicals were purchased from Sigma-
Aldrich (Milan, Italy).
Optimization of the conditions for the evaluation of the inhibitory potential of a
selected test drug on CYP1A2
Phenacetin was used as a probe substrate. This compound is selectively converted to
acetaminophen by the microsomal enzyme CYP1A2. The optimization of the conditions
for evaluation of the inhibitory potential of drugs on microsomal CYP1A2 was
conducted according to the FDA guidance for industries; drug interaction studies, study
design, data analysis and implications for dosing and labeling (2006). Briefly, the
kinetics of acetaminophen formation from phenacetin incubated in human liver
microsomes was tested at 3 different protein concentrations (chosen in order to avoid
enzymatic saturation) for 5 different substrate concentrations. The experimental
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
8
Michaelis-Menten constant values Km and Vmax were calculated by Lineweaver-Burk
equation for each protein concentration tested.
Inhibition experiments were further performed i) at the lowest microsomal protein
concentration tested (0.5 mg/ml) ii) incubating the substrate at the experimental
calculated Km value (50 µM) iii) incubating the substrate for a period of time in which
the formation of acetaminophen is linear and does not exceed the 30% of substrate
depletion (30 min).
Liver subcellular fractions incubations
In vitro incubations were performed in reconstituted human liver homogenate
(microsomes 0.5 mg·ml−1, mitochondria 0.65 mg·ml−1, cytosol 2.05 mg·ml−1) (Kozlov et
al., 2003) or in the single subcellular fraction at the same individual protein
concentration. The following cofactor mixtures were used for all incubations: NADPH
2 mM, NAD+ 0.5 mM, NADH 0.5 mM, reduced glutathione (GSH) 2.5 mM.
Reconstituted human liver homogenate (microsomes + mitochondria + cytosol) or
individual subcellular fractions and cofactors were incubated at the final working
concentration in Na-phosphate buffer pH 7.4 and kept under slight shaking at 37°C
during the experiments. Drugs (HCT 1026, NCX 2057, NOC-5, GTN, ISMN, NOBA)
were dissolved in acetonitrile or water and added to human liver fractions at a final
solvent concentration ≤ 0.5% v/v.
Since diazeniumdiolates like NOC-5 release 2 moles of NO per mole of parent
compound (Fitzhugh and Keefer, 2000), for comparison purposes the compounds were
incubated under equinormal conditions meaning a concentration of GTN, ISMN or
NOBA double that of NOC-5 ([GTN] = [ISMN] = [NOBA] = 2[NOC-5]).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
9
Incubation for HCT 1026 and NCX 2057 metabolic assessment: the experiment was
started adding the compound (HCT 1026 or NCX 2057 at a final concentration of 250
µM) to reconstituted homogenate. At fixed time points (0, 1, 5, 7, 10, 15, 20, 30, 45, 60,
90, 120, 180, 240 min) 50 µl of the incubation mixture was removed and deproteinized
with 100µl of methanol for HPLC-ENO20 NO2-, NO3
- and NOBA analysis. The
remaining 200 µl of incubation mixture was removed at the same time, deproteinized
with 400 µl of acetonitrile + phosphoric acid 1% (v/v) and processed for HPLC/UV
HCT 1026 or NCX 2057 metabolite profiling.
Incubations for nitrite (NO2-) and nitrate (NO3
-) assessment: the experiment was
started adding the compound (HCT 1026, NCX 2057, NOC-5 GTN, ISMN or NOBA at
a final concentration of 250-500µM) to reconstituted homogenate or single fractions. At
fixed time points, 50 µl of incubation mixture was removed, deproteinized with 100 µl
of methanol and submitted for HPLC-VIS NO2- and NO3
- analysis.
Incubations for CYP1A2 inhibition and nitrosothiol (RSNO) assessment: For each
incubation furafylline was used as standard control (reference CYP1A2 inhibitor). After
thirty minutes of incubation in reconstituted homogenate with the drug (NOC-5, GTN,
ISMN, NOBA; concentration range 1-1000 µM), phenacetin (dissolved in acetonitrile)
was added at the final concentration of 50 µM (solvent final concentration ≤ 0.5% v/v).
After one hour of incubation 200 µl of the incubation mixture was spiked with 20 µl of
internal standard (ketoprofen 5 mM) and deproteinized with 400 µl of acidified
acetonitrile (acetonitrile + formic acid 0,1% (v/v)) for HPLC-MS/MS analysis. At the
same time 600 µl of the incubation mixture was spiked with EDTA (final concentration
2 mM) and N-ethylmaleimide (final concentration 5 mM) and processed for RSNO
chemiluminescence analysis.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
10
Incubations for NO assessment: the electrochemical sensor was immersed in 4 ml of
reconstituted human liver homogenate under continuous stirring. The experiment was
started by adding the compound (NOC-5 at a final concentration of 250 µM and GTN,
ISMN or NOBA at a final concentration of 500 µM) and the NO signal was recorded
continuously over 1 hour.
Assay for HCT 1026 and NCX 2057 metabolite profiling
The metabolic profile of HCT 1026 and NCX 2057 and their respective metabolites
was carried out by liquid chromatography analysis on an Agilent 1100 system
equipped with a UV/Visible diode array programmable detector. Separations were
achieved by reverse phase elution with a Synergi Hydro-RP 80 Å column (150 x 4.6
mm i.d., particle size 4 µm) equipped with an AQ-C18 precolumn (4 x 3 mm i.d.)
maintained at 25°C. The mobile phase, acetonitrile (solvent A) and sodium
phosphate buffer 25 mM pH 2 (solvent B), was delivered at a flow rate of 1 ml·min-1.
HCT 1026 and its metabolites were detected at λ = 246 nm with the following
gradient composition: 0 min A 20%, 1-8 min A 80%, 9-11 min A 20%. Under these
conditions the retentions times for HCT 1026, its direct denitrated derivative HCT
1027, and flurbiprofen were 6.8; 4.7; 4.2 minutes, respectively.
NCX 2057 and its metabolites were detected at λ = 325 nm with the following
gradient composition: 0 min A 20%, 4-8 min A 80%, 10-12 min A 20%. Under these
conditions the retentions times for NCX 2057, its direct denitrated derivative NCX
2059 and ferulic acid were 6.1, 4.6 and 4.1 minutes, respectively.
NCX 2057, HCT 1026 and their metabolites were chromatographically separated and
characterized on the basis of the typical UV fingerprint of the pure compounds.
Chromatographic peaks were processed for spectral purity characterization by
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
11
statistical analysis for automated comparison of spectra (Agilent Chemstation
software). Only peaks showing spectral matching factors higher than 99% were
considered acceptable. Chromatograms were integrated with an Agilent Chemstation
software and area under the curve converted into concentration using reference
compounds,
Assays for NO2- and NO3
- and NOBA analysis
NO2- and NO3
- and NOBA concentrations in liver fractions were measured after
methanol precipitation of proteins (1:2 v/v) by using a dedicated HPLC system
(ENO-20; EiCom, Kyoto, Japan) coupled with and Agilent 1100 autosampler. The
method (Yamada and Nabeshima, 1997) is based on the separation of nitrite and
nitrate and NOBA by reverse-phase/ion exchange chromatography, followed by on-
line reduction of nitrate and NOBA to nitrite with cadmium and reduced copper.
Nitrite is then derivatized with the Griess reagent and the level of diazo compounds
is measured by a visible detector at 540 nm. The retentions times for nitrite, nitrate
and NOBA were 5.2, 8.6 and 29.5 minutes, respectively.
Gas phase chemiluminescence assay for RSNO
RSNOs were determined by gas-phase chemiluminescence with a NO analyzer
(Aerocrine AB, Sweden) after reductive cleavage and subsequent determination of the
NO released into the gas phase. The method has been described in detail elsewhere
(Lundberg and Govoni, 2004). NO signals were collected and reported as area under the
curve with Azur (v 3.0) chromatographic software (Datalys, Saint-Martin d'Héres,
France). RSNO were reduced to NO with a solution consisting of 45 mmol/l potassium
iodide (KI) and 10 mmol/l iodine (I2) in glacial acetic acid at 60°C and measurement
performed by direct sample injection (300 µl).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
12
Since the reducing solution is able to convert both nitrite and nitroso/nitrosyl
compounds to NO, RSNO were quantified by simple subtraction of the peak area of
sample aliquots pretreated with sulphanilamide + mercuric chloride at room temperature
for 30 min [10% (v/v) of a solution 5% sulfanilamide + 0.2% HgCl2 in 1 N HCl] from
that of sample aliquots treated only with sulfanilamide at room temperature for 15 min
[10% (v/v) of a solution 5% sulfanilamide in 1 N HCl]. Under these conditions, nitrite
reacts with sulfanilamide to form a stable diazonium ion that is not converted to any
appreciable extent to NO and HgCl2 selectively cleaves the S–NO bond (Feelisch et al.,
2002) without affecting peak shape or recovery of other detectable NO species. The
calibration curve was obtained with freshly prepared S-nitrosoglutathione.
HPLC-MS/MS analysis for CYP1A2 inhibition evaluation
The phenacetin O-deethylase activity of CYP1A2 was evaluated using liquid
chromatography. Analyses were carried out on an Agilent 1100 system coupled with a
triple quadrupole mass spectrometer (API 2000 LC-MS/MS system, Applied
Biosystems/MDS Sciex Toronto, Canada). Separations were achieved by reverse phase
elution with a Phenomenex Synergy Hydro-RP 80 Å (50 x 4 mm i.d., particle size 4
µm) equipped with a Phenomenex AQ-C18 precolumn (4 x 3 mm i.d.) maintained at
25°C. The mobile phase, water containing 0.1% formic acid (solvent A) and acetonitrile
(solvent B), was delivered at a flow rate of 0.35 ml·min-1 at the following gradient
composition: 0-1 min A 100%, 1.10-4.30 min A 0%, 4.40-15 min A 100%. Under these
conditions the retention times for acetaminophen, phenacetin and ketoprofen (internal
standard) were 3.87, 4.12 and 4.4 minutes, respectively.
The following mass spectrometer settings were used: turbo ionspray interface
maintained at 300°C; detection (positive ion mode) of the analytes using the multiple
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
13
reaction monitoring (MRM) scan mode was performed with collision activated
dissociation gas (CAD) at 5 psi; curtain gas (CUR) at 20 psi; turbo ion spray voltage at
+5500 V; channel electron multiplier (CEM) at 2400 V; source gases (GS1 and GS2)
pressures at 40 and 50 psi, respectively; declustering potential (DP), focusing potential
(FP) and entrance potential (EP) were set at 31, 370 and 10.05 V, respectively;
experiments were performed using a dwell time of 50 ms. The spectra were acquired at
unit resolution.
The mass transitions [MRM] of acetaminophen, phenacetin and ketoprofen were m/z
152→110, m/z 180→138 and m/z 255→105, respectively. Chromatograms were
integrated with Analyst software (version 1.2 Applied Biosystem, CA, USA) and area
under the curve converted into concentration using reference compounds.
Assay for electrochemical detection of NO
The measurement of NO release was performed with the inNO nitric oxide
measurement system using the amiNO-700 sensor electrode (Innovative Instruments
Inc., U.S.A.). The output current was monitored and recorded on a Windows 98
compatible system using analysis software provided with the inNO system.
The sensor was calibrated by adding known volumes of standard nitrite solution to an
acidified solution in the presence of iodide ion according to the manufacturer's
specifications and settled for temperature compensation before the experiment.
Data analysis
NOx (NO3- + NO2
-), nitrite and nitrate values in the incubations were subtracted from
the basal (time 0). One-way analysis of variance was used to evaluate differences
between groups and it was followed by a Tukey post hoc test for comparisons between
multiple groups. Differences were considered significant at p < 0.05. IC50 values for
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
14
enzyme inhibition were obtained by fitting to the sigmoid equation:Y= Y0 + [a /
1+(X/IC50)b] were Y = % inhibition and X = inhibitor concentration. Data were modeled
by nonweighted nonlinear regression analysis using SigmaPlot 8.02 (Systat Software,
Inc., Point Richmond, CA).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
15
Results
Liver metabolism of nitrooxybutyl-ester compounds.
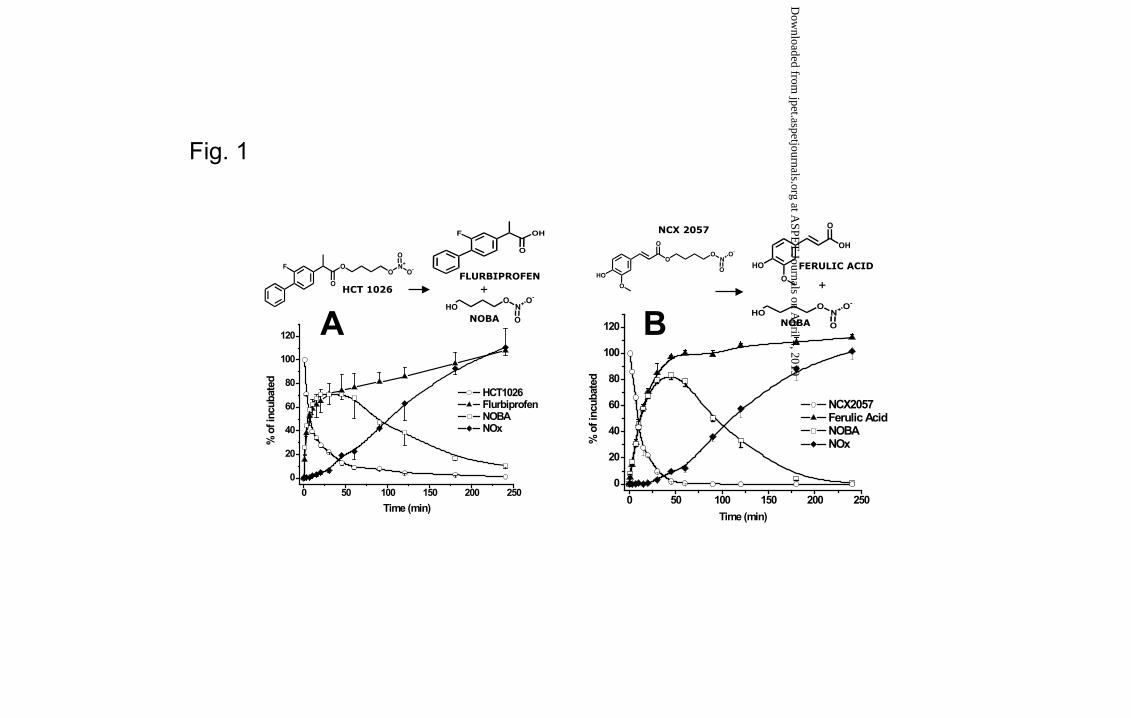
The chemical structures of HCT 1026, NCX 2057 and their metabolites arising from
biotransformation pathways involving hydrolytic cleavage of the ester functions in the
molecule are depicted in Fig. 1A and Fig. 1B, respectively.
HCT 1026 and NCX 2057 were rapidly and extensively metabolized at the carboxyl-
ester function. HCT 1026 and NCX 2057 rapidly disappeared (t1/2 = 7.03 ± 1.24, 9.15 ±
0.28 minutes, respectively) with a stoichiometric formation of flurbiprofen and ferulic
acid, respectively (Fig. 1A,B). At the same time the formation of NOBA and NOx
species coming from its metabolism, was consistent with the generation of flurbiprofen
and ferulic acid, respectively. In fact, the kinetics of NOBA generation from HCT 1026
and NCX 2057 overlapped the kinetics of flurbiprofen and ferulic acid, respectively,
over the first 20-30 minutes of incubation and thereafter declining consistent with the
slow metabolism of NOBA to NOx species (tmax = 30 and 45 minutes for HCT 1026 and
NCX 2057, respectively and Cmax = 70.78 ± 8.13 and 83.38 ± 3.06 % of incubated for
HCT 1026 and NCX 2057, respectively). The elimination of NOBA was consistent with
the generation of equimolar amount of NOx exclusively in the form of NO3- for both
compounds tested.
HCT 1026 was converted to flurbiprofen with V0 = - 19.32 ± 5.79 µM/min consistent
with V0 of flurbiprofen and NOBA formation of 18.63 ± 1.54 µM/min and 18.39 ± 3.05
µM/min , respectively, (- V0 HCT 1026 ≈ V0 Flurbiprofen ≈ V0 NOBA) (Fig. 1A). NCX 2057 was
converted to ferulic acid with V0 = - 13.87 ± 2.51 µM/min consistent with V0 of ferulic
acid and NOBA formation of 12.93 ± 0.15 µM/min and 11.30 ± 1.23 µM/min,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
16
respectively, (- V0 NCX 2057 ≈ V0 Ferulic Acid ≈ V0 NOBA) (Fig. 1B). HCT 1027 and NCX
2059 were not observed over the incubation time.
Incubations with the vehicle in liver homogenate or with the compounds alone in buffer
did not produce any increase in NOx concentrations.
The liver metabolism of other compounds belonging to the class of nitrooxybutyl-ester
derivatives was investigated and showed very similar metabolic properties to HCT 1026
and NCX 2057 (data not shown).
NOC-5, GTN, ISMN and NOBA liver metabolism
NOC-5 and GTN were rapidly and extensively metabolized to NOx species. The
kinetics of NOx generation from the two compounds overlapped over 90 min of
incubation with an almost identical V0 (13.0 ± 0.9 µM/min and 12.9 ± 0.4 µM/min for
NOC-5 and GTN, respectively) (Fig. 2). However, the generation of nitrite and nitrate
as individual species was different. While GTN generated nitrite as the main metabolite
(V0 NO2- = 13.1 ± 0.1 µM/min ≈ V0 NOx >> V0 NO3
- = 0.4±0.1 µM/min) (Fig. 3A),
NOC-5 generated both nitrite and nitrate with similar V0 (V0 NO2- = 7.3 ± 0.2 µM/min,
V0 NO3- = 5.7 ± 0.2 µM/min) (Fig. 3B).
NOBA incubated under the same experimental conditions displayed a slower rate of
metabolism to form NOx than GTN while NOx formation from ISMN was very low
(Fig. 4). An analysis of the individual species generated from NOBA also showed
differences in terms of metabolic products. In fact, NOBA led exclusively to the
generation of NO3- with a V0 NO3
- ≈ V0 NOx = 1.2±0.1 µM/min and a generation of NO3
-
70 µM and 460 µM after 60 and 240 minutes of incubation, respectively (Fig. 3C).
Incubations with the vehicle in liver homogenate or with the compounds alone in buffer
did not produce any increase in NOx concentrations.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
17
RSNO formation
Fig. 5A shows the generation of RSNO measured 1 hour after incubation of NOC-5 and
GTN at different concentrations. Although the amount of RSNO formed was much
lower in comparison to the initial concentration, NOC-5 showed a higher nitrosating
capacity than GTN,in particular at the highest concentrations tested, where RSNO levels
reached 9.8 ± 2.1µM for NOC-5 and 3.5 ± 0.3µM for GTN (Fig. 5A). Incubations with
NOBA or ISMN under the same experimental conditions did not produce any RSNOs,
nor did incubations with vehicle in liver homogenate or with the compounds alone in
buffer.
Inhibition of CYP1A2
The effects of GTN, ISMN, NOBA and NOC-5 on CYP1A2 activity are shown in Fig.
5B. The estimated IC50 values obtained showed that organic nitrates as ISMN, NOBA
and GTN did not significantly inhibit the phenacetin O-deethylase activity of CYP1A2.
In contrast, NOC-5 completely inhibited CYP1A2 activity at a concentration of 500µM
with an IC50 of 114.2 µM. Incubations were performed in homogenate (microsomes +
mitochondria + cytosol) to guarantee the presence of a complete pool of liver enzymes
and complete liver metabolic activity.
Direct measurement of NO formation
Generation of NO was assessed by an electrochemical sensor and large differences were
noted between NOC-5 and the organic nitrates GTN, ISMN and NOBA. The
spontaneous metabolism-independent NO donor NOC-5, showed a rapid generation of
NO peaking (tmax) at 6 min and reaching a maximum concentration (Cmax) of 4.15 µM
(Fig. 5C). In contrast, GTN, NOBA and ISMN did not show any NO increase over the
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
18
study period (Fig. 5C). Lower concentrations of organic nitrates were also tested but
again, no formation of NO was detected (data not shown).
Incubations with the vehicle in reconstituted homogenate or with compounds alone in
buffer did not produce any NO.
Metabolism of GTN in different liver compartments
Conversion of GTN to NOx. differed between the various liver compartments studied.
Among the fractions, the cytosol was able to metabolize GTN with the highest rate V0
cytosol = 7.8 ± 0.5 µM/min being more than 4 times higher than the metabolism in the
other fractions (V0 mitochondria = 1.0 ± 0.1 µM/min, V0 microsomes = 1.7 ± 0.1 µM/min) (Fig.
6A). No significant differences were observed when GTN was incubated in
homogenate (microsomes + mitochondria + cytosol) compared to incubations in the
cytosolic fraction (V0 Cytosol ≈ V0 microsomes+mitochondria+cytosol = 7.9 ± 0.6 µM/min) (Fig.
6A). Moreover the generation of nitrite in the first part of the pharmacokinetic profile
(0-10 min) in homogenate and cytosol were similar (Fig. 6B)
Although GTN was found to be stable in phosphate buffer (data not shown), it was
slightly unstable in the presence of cofactors. Incubations performed in presence of each
cofactor alone (NAD+, NADH, NADPH or GSH) showed that this instability is due to
GSH slowly reacting with GTN and generating NOx species (data not shown).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
19
Discussion
In a previous study (Govoni et al., 2006) we showed that nitrooxybutyl-ester derivatives
undergoes ubiquitous and fast esterase-mediated carboxyl-ester hydrolysis in rat liver
and plasma, yielding to the formation of the parent compound and an organic nitrate
moiety speculatively identified as the nitrooxybutyl-alcohol species (NOBA). The direct
and unequivocal identification of NOBA following metabolism of nitrooxybutyl-ester
(Fig. 1) has been made possible only in the current study with the development of a
novel chromatographic method based on the on line conversion of organic nitrates to
nitrite and subsequent derivatization with the griess reagent. It follows that NOBA is in
fact the active organic nitrate metabolite of nitrooxybutyl-esters and for these reasons it
has been used on its own to investigate the mechanism of denitration of this class of
compounds in human liver in comparison to GTN, ISMN and the spontaneous NO
donor NOC-5. The various organic nitrates displayed considerably different rates of
liver metabolism to NOx, with GTN being rapidly metabolized NOBA showing an
intermediate rate and ISMN being negligibly metabolized (Fig. 4). Notably, NOC-5 and
GTN generated identical quantities of NOx (Fig. 2). Assuming that the first metabolic
step of organic nitrates in the liver is associated with the release of NO which is
consequently oxidized to nitrite and nitrate, these data might have been taken as
evidence for GTN and NOC-5 having the same NO releasing kinetics in the liver.
However, when looking at the generation of NO2- and NO3
- separately (Fig. 3A,B) it
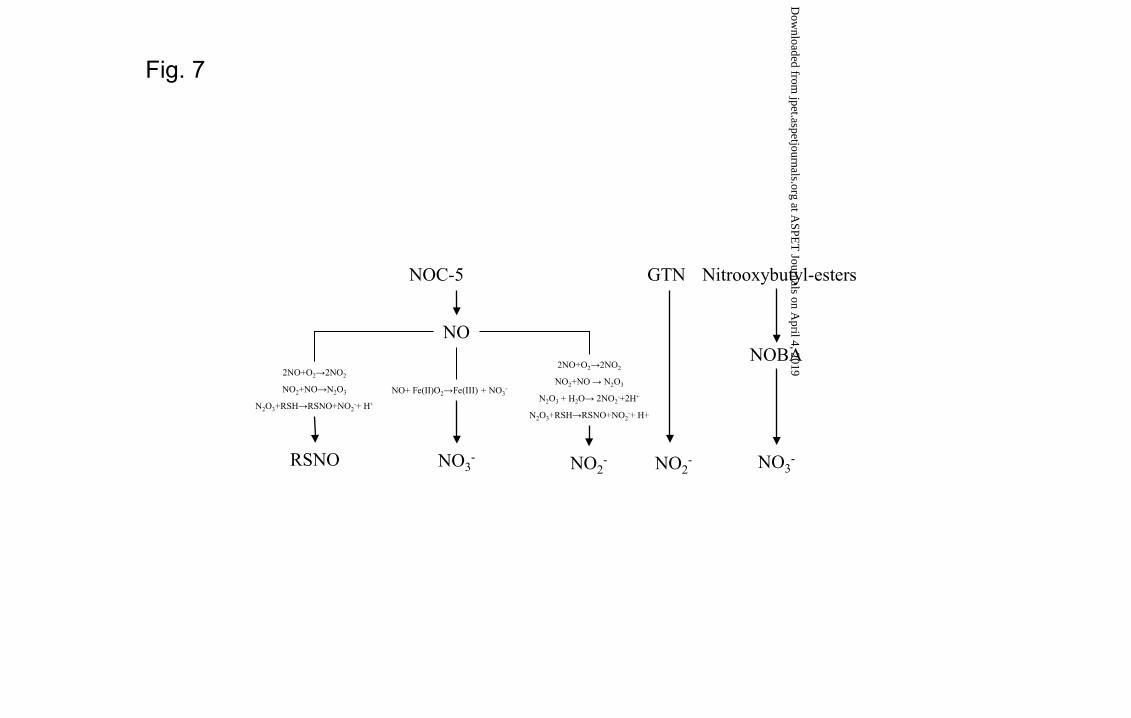
became evident that the mechanism of denitration is different from these two molecules.
While NOC-5 denitration is consistent with a direct generation of NO followed by
oxidation to both NO2- and NO3
-, GTN denitration led almost exclusively to the
generation of NO2- (Fig. 7). Notably, also the metabolic profile of NOBA was not
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
20
consistent with a direct generation of NO leading exclusively to the generation of NO3-
species (Fig. 3C). In addition to the different NO2-/NO3
- metabolic profile, GTN, NOBA
and NOC-5 also showed different nitrosating capacity measured as S-nitrosothiol
generation. In fact, consistent with the direct generation of NO in turn forming the
potent nitrosating agent N2O3 (Fig. 7), S-nitrosothiols formation was higher for NOC-5
(Fig. 5A). A further indirect assessment of GTN and NOBA metabolism comes from
the experiments looking at the activity of CYP1A2, an enzyme potently inhibited by
NO (Stadler et al., 1994). NOC-5 was found to extensively inhibit CYP1A2 activity
while the different organic nitrates did not (Fig. 5B).
These results clearly suggest that organic nitrates share a common pathway of
denitration in the liver not primarily being metabolized to NO. Instead, depending on
their chemical structure, they are metabolized to nitrate or nitrite. This was further
demonstrated by measuring NO formation as shown in Fig. 5C. While NOC-5 directly
released NO, GTN and NOBA did not show any detectable NO formation.
Importantly, the denitration of GTN in liver homogenate is exclusively carried out by
the cytosolic fraction with a negligible contribution from the mitochondrial as well as
the microsomal compartments. This is clearly evidenced by the fact that the denitration
of GTN in the presence of a complete pool of liver enzymes (microsomes +
mitochondria + cytosol) was the same as when using only cytosolic enzymes (Fig. 6).
Interestingly it has been demonstrated previously (Kozlov et al., 2003) that the cytosolic
fraction of the liver is not capable of significative GTN bioactivation to NO. Taken
together these findings suggest that the absence of intrahepatic NO generation is
explained by the much higher rate of denitration carried out by the cytosolic enzymes
thereby consuming GTN before it reaches the other compartments.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
21
Recently it has been reported that the enzymatic metabolism of organic nitrates might
be concentration-dependent (Daiber et al., 2009) and therefore a careful evaluation of
the test concentrations used is needed when investigating the metabolism of these
compounds. GTN, ISMN and nitrooxybutylester-compounds can be taken by the oral
route in doses of 46 µmoles, 628 µmoles and 2,2 mmoles, respectively (Parker and
Parker, 1998; Fagerholm et al., 2005). These molecules are lipophilic and very rapidly
absorbed to reach the liver. GTN is almost completely metabolized during first passage
leaving the dinitrate metabolite GDN to circulate systemically (Needleman et al., 1976,
Yu et al., 1988). The tmax of intact detectable GTN in vivo in plasma after oral intake is
5 minute after administration (Yu et al., 1988). This suggests that the entire dose taken
is rapidly concentrated in the liver during first pass. Nitrooxybutylester-derivatives are
rapidly absorbed from the GI tract and their parent compound appears in the circulation
with almost complete bioavailability (Fagerholm et al., 2005). This suggests formation
of NOBA at relevant concentration in the liver. Following these considerations the mid
micromolar concentrations tested in the present work seem appropriate for analyzing the
metabolic pathway during first passage. These concentrations might however not be
suitable to assess the functional metabolism of organic nitrates after the first passage
since the concentrations of intact drug reaching the target tissues is orders of magnitude
lower.
NO donor compounds are known to inhibit CYP by forming nitrosyl-heme CYP
reversibly in competition with oxygen (Ignarro et al., 2002). Such decreased CYP
mediated activity might have implications for drug interactions when NO donors are
given in association with other drugs metabolized by CYPs. In this in vitro study
organic nitrates were not found to inhibit CYP1A2 (Fig. 5B) most likely due to lack of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
22
NO release (Fig. 5C). Translated in vivo these findings suggest that CYP-dependent
drug metabolism of concomitant drugs would not be affected by co-administration of
organic nitrates.
Another implication of the results of current study is that while the systemic
bioavailablity of organic nitrates might be decreased by its liver metabolism to nitrate or
nitrite this specific metabolic route may support another recently described NO
pathway. Nitrate can be converted in vivo to nitrite that is emerging as a stable
circulating NO pool and a variety of pathways have been described for the reduction of
nitrite to NO in blood and tissues (Lundberg et al., 2008; Lundberg et al., 2009). Such
NO formation is expected to be slower and more prolonged than direct release of NO
from most organic nitrates and can significantly contribute to the final therapeutic effect
of these molecules in connection to the extent and identity of species generated (nitrate
or nitrite) and exported from the liver.
In conclusion, our data suggests that organic nitrates are not primarily metabolized to
NO in the liver and their different chemical structures not only dramatically change the
extent of denitration in this organ but also the entity of the metabolic species generated.
ISMN undergoes negligible liver metabolism while GTN is rapidly and extensively
metabolized primarily to nitrite. Nitrooxybutyl-ester compounds on the other hand are
first rapidly metabolized to the organic nitrate moiety NOBA which is eventually
slowly metabolized to inorganic nitrate. If the lower extent of denitration of NOBA in
comparison to GTN was also reflected in vivo during first passage, NOBA might have
the potential to overcome the liver and reach (at least in part) the systemic circulation
allowing for bioactivation to NO in the vascular compartment or elsewhere where a
different enzymatic pool than in the liver exists. In summary the first-pass may
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
23
differently affect the pharmacological profile of organic nitrates in connection with the
different extent of denitration and the distinct species generated and exported from the
liver (nitrate or nitrite).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
24
Authorship Contributions.
Participated in research design: Govoni, Tocchetti, Lundberg Conducted experiments: Govoni Contributed new reagents or analytic tools: Govoni, Lundberg Performed data analysis: Govoni Wrote or contributed to the writing of the manuscript: Govoni, Lundberg
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
25
References
Abshagen UW (1992) Pharmacokinetics of isosorbide mononitrate. Am J Cardiol 70: 61G-66G.
Boultadakis A, Liakos P and Pitsikas N (2010) The nitric oxide-releasing derivative of ferulic acid NCX 2057 antagonized delay-dependent and scopolamine-induced performance deficits in a recognition memory task in the rat. Prog Neuropsychopharmacol Biol Psychiatry 34: 5-9.
Daiber A, Wenzel P, Oelze M, Schuhmacher S, Jansen T and Münzel T (2009) Mitochondrial aldehyde dehydrogenase (ALDH-2)--maker of and marker for nitrate tolerance in response to nitroglycerin treatment. Chem Biol Interact 178: 40-47.
Fagerholm U and Björnsson MA (2005) Clinical pharmacokinetics of the cyclooxygenase inhibiting nitric oxide donator (CINOD) AZD3582. J Pharm Pharmacol 57: 1539-1554.
Feelisch M, Rassaf T, Mnaimneh S, Singh N, Bryan NS, Jourd'Heuil D and Kelm M (2002) Concomitant S-, N-, and heme-nitros(yl)ation in biological tissues and fluids: implications for the fate of NO in vivo. FASEB J 16: 1775-1785.
Fitzhugh AL and Keefer LK (2000) Diazeniumdiolates: pro- and antioxidant applications of the "NONOates". Free Radic Biol Med 28: 1463-1469.
Gasparini L, Ongini E, Wilcock D and Morgan D (2005) Activity of flurbiprofen and chemically related anti-inflammatory drugs in models of Alzheimer's disease. Brain Res Brain Res Rev 48: 400-408.
Govoni M, Casagrande S, Maucci R, Chiroli V and Tocchetti P (2006) In vitro metabolism of (nitrooxy)butyl ester nitric oxide-releasing compounds: comparison with glyceryl trinitrate. J Pharmacol Exp Ther 317: 752-761.
Hodoşan F, Mantsch O, Jude I, Breazu D, Cuparencu B and Sterescu N (1969) Nitrate esters of steroid hormones and related compounds. Preparation and biological properties. Arzneimittelforschung 19: 684-685.
Idris AI, Del Soldato P, Ralston SH and van't Hof RJ (2004) The flurbiprofen derivatives HCT1026 and HCT1027 inhibit bone resorption by a mechanism independent of COX inhibition and nitric oxide production. Bone 35: 636-643.
Ignarro LJ, Napoli C and Loscalzo J (2002) Nitric oxide donors and cardiovascular agents modulating the bioactivity of nitric oxide: an overview. Circ Res 90: 21-28.
Keeble JE and Moore PK (2002) Pharmacology and potential therapeutic applications of nitric oxide-releasing non-steroidal anti-inflammatory and related nitric oxide-donating drugs. Br J Pharmacol 137: 295-310.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
26
Khatsenko OG, Gross SS, Rifkind AB and Vane JR (1993) Nitric oxide is a mediator of the decrease in cytochrome P450-dependent metabolism caused by immunostimulants. Proc Natl Acad Sci U S A 90: 11147-11151.
Kleschyov AL, Oelze M, Daiber A, Huang Y, Mollnau H, Schulz E, Sydow K, Fichtlscherer B, Mülsch A and Münzel T (2003) Does nitric oxide mediate the vasodilator activity of nitroglycerin? Circ Res 93: e104-112.
Kozlov AV, Dietrich B and Nohl H (2003). Various intracellular compartments cooperate in the release of nitric oxide from glycerol trinitrate in liver. Br J Pharmacol 139: 989-997.
Larsson AK, Bäck M, Lundberg JO and Dahlén SE (2009) Specific mediator inhibition by the NO donors SNP and NCX 2057 in the peripheral lung: implications for allergen-induced bronchoconstriction. Respir Res 10: 46.
Larsson AK, Fumagalli F, DiGennaro A, Andersson M, Lundberg J, Edenius C, Govoni M, Monopoli A, Sala A, Dahlén SE and Folco GC (2007) A new class of nitric oxide-releasing derivatives of cetirizine; pharmacological profile in vascular and airway smooth muscle preparations. Br J Pharmacol 151: 35-44.
Lundberg JO, Gladwin MT, Ahluwalia A, Benjamin N, Bryan NS, Butler A, Cabrales P, Fago A, Feelisch M, Ford PC, Freeman BA, Frenneaux M, Friedman J, Kelm M, Kevil CG, Kim-Shapiro DB, Kozlov AV, Lancaster JR Jr, Lefer DJ, McColl K, McCurry K, Patel RP, Petersson J, Rassaf T, Reutov VP, Richter-Addo GB, Schechter A, Shiva S, Tsuchiya K, van Faassen EE, Webb AJ, Zuckerbraun BS, Zweier JL and Weitzberg E (2009) Nitrate and nitrite in biology, nutrition and therapeutics. Nat Chem Biol 5: 865-869.
Lundberg JO and Govoni M (2004) Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic Biol Med 37: 395-400.
Lundberg JO, Weitzberg E and Gladwin MT (2008) The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov 7: 156-167.
Mülsch A, Bara A, Mordvintcev P, Vanin A and Busse R (1995) Specificity of different organic nitrates to elicit NO formation in rabbit vascular tissues and organs in vivo. Br J Pharmacol 116: 2743-2749.
Münzel T, Daiber A and Mülsch A (2005) Explaining the phenomenon of nitrate tolerance. Circ Res 97: 618-628.
Murrell W (1879) Nitro-glycerine as a remedy for angina pectoris. Lancet 16: 80-81.
Needleman P (1976) Organic nitrate metabolism. Annu Rev Pharmacol Toxicol 16: 81-93.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
27
Needleman P, Lang S and Johnson EM (1972) Organic nitrates: relationship between biotransformation and rational angina pectoris therapy. J Pharmacol Exp Ther 181: 489-497.
Parker JD and Parker JO (1998) Nitrate therapy for stable angina pectoris. N Engl J Med 338: 520-531.
Prosperi C, Scali C, Barba M, Bellucci A, Giovannini MG, Pepeu G and Casamenti F (2004) Comparison between flurbiprofen and its nitric oxide-releasing derivatives HCT-1026 and NCX-2216 on Abeta(1-42)-induced brain inflammation and neuronal damage in the rat. Int J Immunopathol Pharmacol 17: 317-330.
Ronchetti D, Borghi V, Gaitan G, Herrero JF and Impagnatiello F (2009) NCX 2057, a novel NO-releasing derivative of ferulic acid, suppresses inflammatory and nociceptive responses in in vitro and in vivo models. Br J Pharmacol 158: 569-579.
Scatena R (2004) Nitroflurbiprofen (NicOx). Curr Opin Investig Drugs 5: 551-556.
Servent D, Delaforge M, Ducrocq C, Mansuy D and Lenfant M (1989) Nitric oxide formation during microsomal hepatic denitration of glyceryl trinitrate: involvement of cytochrome P-450. Biochem Biophys Res Commun 163: 1210-1216.
Stadler J, Trockfeld J, Schmalix WA, Brill T, Siewert JR, Greim H and Doehmer J (1994) Inhibition of cytochromes P4501A by nitric oxide. Proc Natl Acad Sci U S A 91: 3559-3563.
Wenk GL, McGann-Gramling K, Hauss-Wegrzyniak B, Ronchetti D, Maucci R, Rosi S, Gasparini L and Ongini E (2004) Attenuation of chronic neuroinflammation by a nitric oxide-releasing derivative of the antioxidant ferulic acid. J Neurochem 89: 484-493.
Wenk GL, Rosi S, McGann K and Hauss-Wegrzyniak B (2002) A nitric oxide-donating flurbiprofen derivative reduces neuroinflammation without interacting with galantamine in the rat. Eur J Pharmacol 453: 319-324.
Wink DA, Osawa Y, Darbyshire JF, Jones CR, Eshenaur SC and Nims RW (1993) Inhibition of cytochromes P450 by nitric oxide and a nitric oxide-releasing agent. Arch Biochem Biophys 300: 115-123.
Yamada K and Nabeshima T (1997) Simultaneous measurement of nitrite and nitrate levels as indices of nitric oxide release in the cerebellum of conscious rats. J Neurochem 68: 1234-1243.
Yu DK, Williams RL, Benet LZ, Lin ET and Giesing DH (1988) Pharmacokinetics of nitroglycerin and metabolites in humans following oral dosing. Biopharm Drug Dispos 9: 557-565.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
28
Footnotes.
a) This work was supported by NicOx S.A., Sophia Antipolis Cedex, France
c) Person to receive reprint requests:
Name: Mirco Govoni
Affiliation: Department of Physiology and Pharmacology, Karolinska Institute
Street address: Nanna Svartz väg 2
Postal code: S-177 76
City: Stockholm
State:Sweden
Telephone: +39 339 8657744
Fax: +46 8 332278
email: [email protected]
d) Authors name:
Mirco Govoni1,2, Paola Tocchetti2 and Jon O Lundberg1
1Department of Physiology and Pharmacology, Karolinska Institute, Nanna Svartz väg
2, S-177 76 Stockholm, Sweden
2NicOx S.A., Les Taissounières - 1681 Route des Dolines - Sophia Antipolis Cedex,
France
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
29
Legends for figures.
Fig. 1. Chemical structures and metabolic profile of HCT 1026 (A), NCX 2057 (B) and
their metabolites arising from biotransformation pathways involving hydrolytic
cleavage of the carboxylic and nitric ester functions in reconstituted human liver
homogenate (cytosol + microsomes + mitochondria) with cofactors. HCT 1026 (A) and
NCX 2057 (B) were incubated at a final concentration of 250µM and their metabolic
profile followed over the incubation time. Filled up triangles (�) show the kinetic of
flurbiprofen and ferulic acid formation from HCT 1026 and NCX 2057 (�),
respectively, while the direct denitrated metabolites HCT 1029 or NCX 2059 were not
observed over the incubation time. Open squares represent the kinetics of NOBA
generation from HCT 1026 (A) and NCX 2057 (B) (�) and filled up diamonds the time
course of NOx species generated over the incubation time (�). NOx values are
subtracted from the basal (time 0). Data are presented as mean ± S.D., n ≥ 3.
Fig. 2. Extent of denitration of GTN and NOC-5 in reconstituted human liver
homogenate (microsomes + mitochondria + cytosol) with cofactors. GTN (�) and
NOC-5 (�) were incubated at equinormal conditions (500µM and 250µM for GTN and
NOC-5, respectively) and the profile of NOx species generated followed over the
incubation time. Values are subtracted from the basal (time 0). Data are presented as
mean ± S.D., n ≥ 3.
Fig. 3. Metabolism of GTN (A), NOC-5 (B) and NOBA (C) in reconstituted human
liver homogenate (microsomes + mitochondria + cytosol) with cofactors. GTN, ISMN
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
30
and NOC-5 were incubated at equinormal conditions (500µM for GTN and ISMN and
250µM for NOC-5) and the profile of NO2- and NO3
- species generated followed over
the incubation time. Values are subtracted from the basal (time 0). Data are presented as
mean ± S.D., n ≥ 3.
Fig. 4. Extent of denitration of organic nitrates in reconstituted human liver homogenate
(microsomes + mitochondria + cytosol) with cofactors. GTN (�), ISMN (�) and
NOBA (�) were incubated at a final concentration of 500 µM and the profile of NOx
species generated followed over the incubation time. Values are subtracted from the
basal (time 0). Data are presented as mean ± S.D., n ≥ 3.
Fig. 5. (A) RSNO generation from GTN and NOC-5 in reconstituted human liver
homogenate (microsomes + mitochondria + cytosol) with cofactors. GTN and NOC-5
were incubated at different concentrations ranging from 5µM to 1000µM and the
amount of RSNO species generated was measured after 1 hour of incubation.
Comparisons were made between groups incubated at equinormal concentrations
([GTN] = 2[NOC-5]). Data are presented as mean ± SEM, n ≥ 3. *, p < 0.05. Notably
NOBA and ISMN incubated in the same experimental conditions did not show any
RSNO increase and were therefore omitted from the present figure. (B) CYP1A2
inhibition in reconstituted human liver homogenate (microsomes + mitochondria +
cytosol) with cofactors. NOC-5 (�), GTN (�), ISMN (�) and NOBA (�) were
preincubated for 30 minutes at different concentrations (ranging from 1 to 500µM) and
the phenacetin O-deethylase activity of CYP1A2 measured 30 minutes after addition of
phenacetin. The CYP1A2 activity was considered 100% in presence of the vehicle. Data
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #203356
31
are presented as mean ± S.D., n ≥ 3. *, p < 0.05. (C) Direct measurement of NO by
electrochemical sensor in reconstituted human liver homogenate (microsomes +
mitochondria + cytosol) with cofactors. ISMN, NOBA, GTN and NOC-5 were
incubated at equinormal conditions (500µM for ISMN, NOBA, GTN and 250µM for
NOC-5) and the generation of NO followed over 1 hour of incubation.
Fig. 6. (A) Extent of denitration of GTN in the different liver compartments. GTN was
incubated at a final concentration of 250 µM and the profile of NOx species generated
followed over the incubation time in reconstituted human liver homogenate
(microsomes + mitochondria + cytosol) with cofactors (�), human liver cytosol with
cofactors (�), human liver microsomes with cofactors (�), human liver mitochondria
with cofactors (�), buffer + cofactors (�). (B) Nitrite formation in liver homogenate
(microsomes + mitochondria + cytosol) and cytosol in the first 10 min of incubation.
Values are subtracted from the basal (time 0). Data are presented as mean ± S.D., n ≥ 3.
Fig. 7. Metabolism of NOC-5, GTN and NOBA in human liver. While NOC-5 is
spontaneously converted to NO which in turn is metabolized in the liver to nitrite,
nitrate and nitrosothiols, the organic nitrates GTN and NOBA are directly metabolized
to nitrite and nitrate, respectively.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
0 50 100 150 200 2500
20
40
60
80
100
120
% o
f inc
ubat
ed
Time (min)
NCX2057 Ferulic Acid NOBA NOx
Fig. 1
A B HCT 1026
O
O
FO
N+O
O-FLURBIPROFEN
OH
O
F
+ HO
ON+
O
O-
NOBA
NCX 2057
HO
O
O
OO
N+
O
O-
FERULIC ACID HO
O
O
OH
+
HOO
N+
O
O-
NOBA
0 50 100 150 200 2500
20
40
60
80
100
120
% o
f inc
ubat
ed
Time (min)
HCT1026 Flurbiprofen NOBA NOx
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
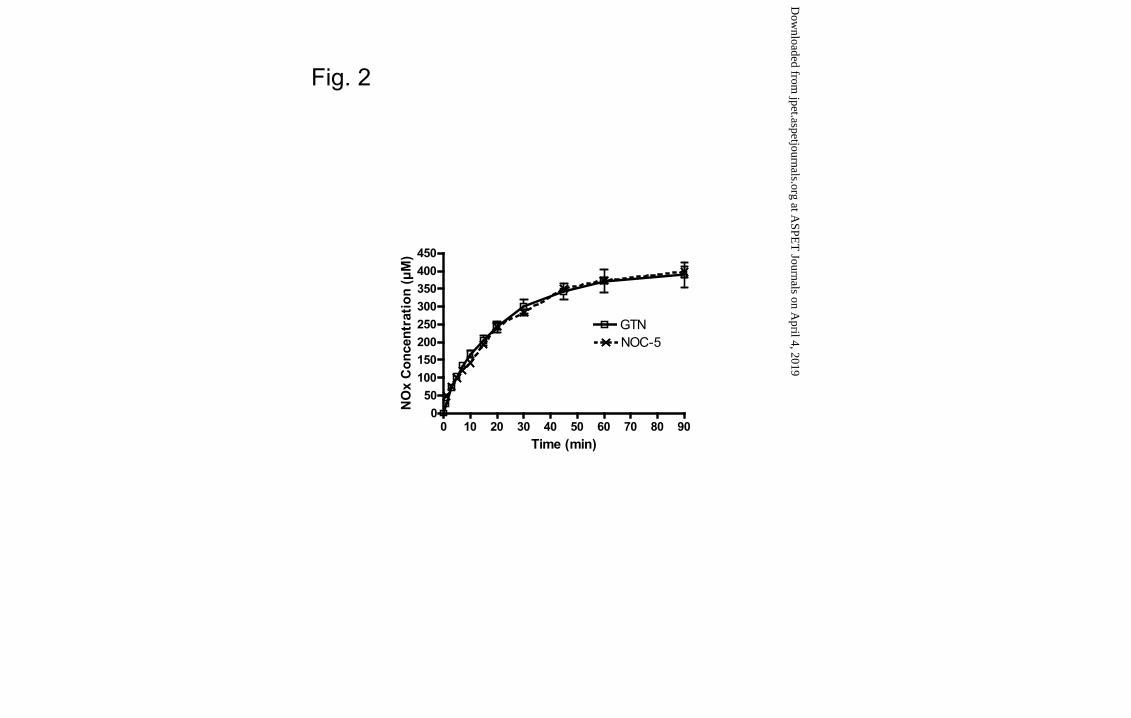
Fig. 2
0 10 20 30 40 50 60 70 80 900
50100150200250300350400450
GTNNOC-5
Time (min)
NO
x C
once
ntra
tion
(µM
)
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
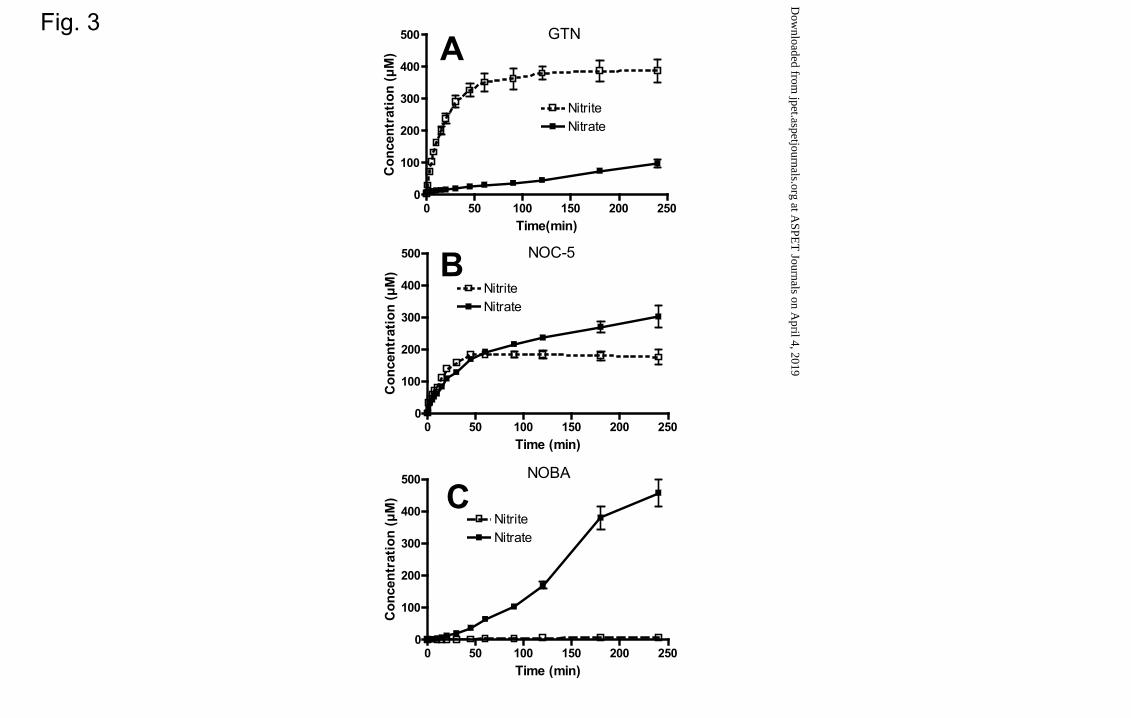
Fig. 3 A
B
0 50 100 150 200 2500
100
200
300
400
500
NitriteNitrate
GTN
Time(min)
Con
cent
ratio
n (µ
M)
0 50 100 150 200 2500
100
200
300
400
500
NitriteNitrate
NOC-5
Time (min)
Con
cent
ratio
n (µ
M)
C
0 50 100 150 200 2500
100
200
300
400
500
NitriteNitrate
NOBA
Time (min)
Con
cent
ratio
n (µ
M)
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
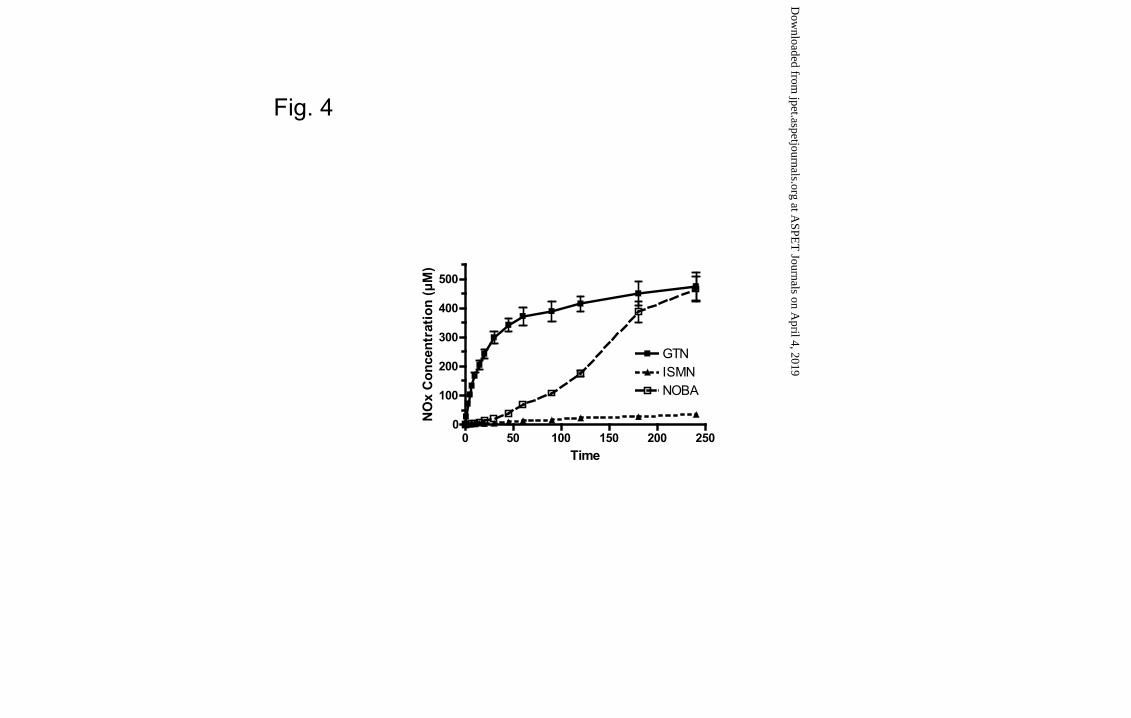
Fig. 4
0 50 100 150 200 2500
100
200
300
400
500
GTNISMNNOBA
Time
NO
x C
once
ntra
tion
(µM
)
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
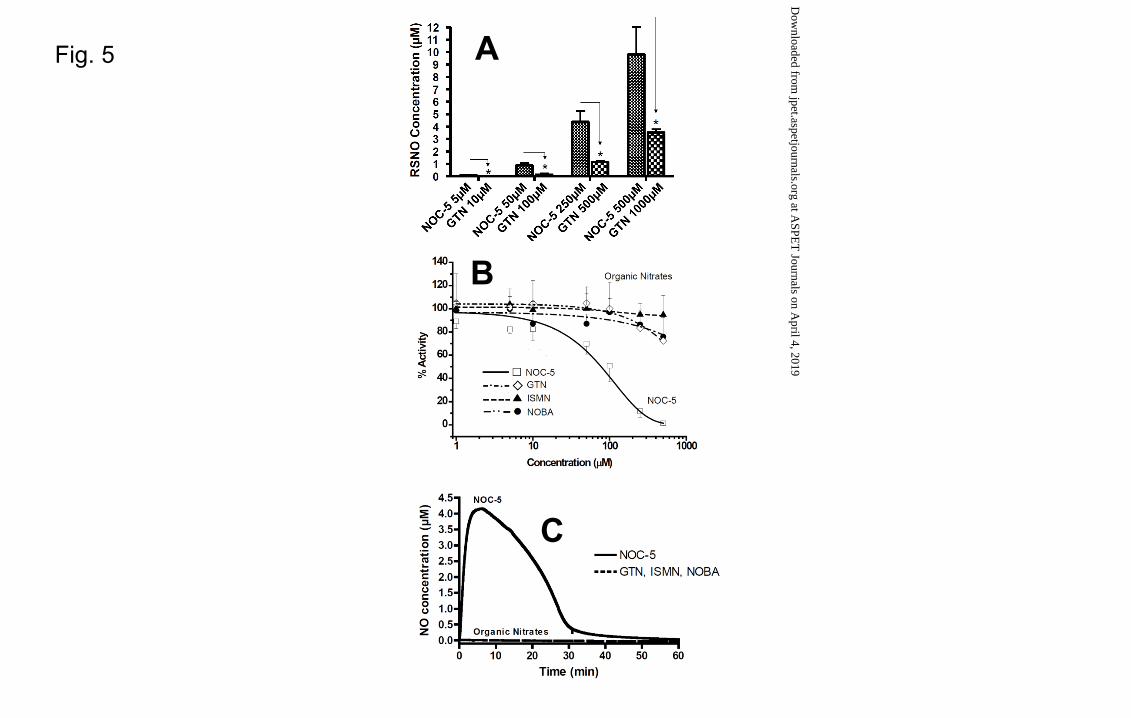
Fig. 5
B
A
C
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
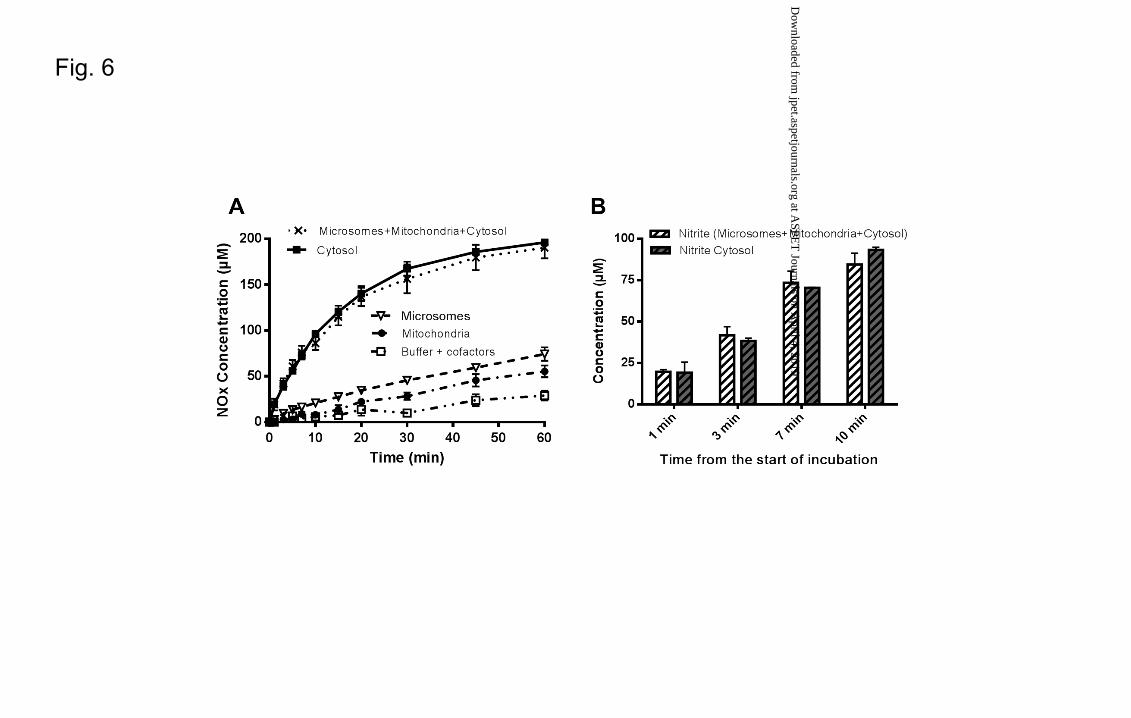
Fig. 6
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from
GTN
2NO+O2→2NO2
NO2+NO → N2O3
N2O3 + H2O→ 2NO2-+2H+
N2O3+RSH→RSNO+NO2-+ H+
NO2-
NO
NO2- RSNO
NO+ Fe(II)O2→Fe(III) + NO3-
NO3-
2NO+O2→2NO2
NO2+NO→N2O3
N2O3+RSH→RSNO+NO2-+ H+
NOC-5
Fig. 7
NOBA
NO3-
Nitrooxybutyl-esters
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on April 17, 2013 as DOI: 10.1124/jpet.113.203356
at ASPE
T Journals on A
pril 4, 2019jpet.aspetjournals.org
Dow
nloaded from













![Hsin-Yueh Chang Jason T.C. Tzen, Su-Jen Lin, Yi-Ting …jpet.aspetjournals.org/content/jpet/early/2010/12/28/jpet.110...JPET #174839 2 Running Title page [Running title] Prednisolone](https://img.pdfslide.net/doc/110x75/5ac22fbe7f8b9ae45b8e3f75/hsin-yueh-chang-jason-tc-tzen-su-jen-lin-yi-ting-jpet-174839-2-running.jpg)















