Embed Size (px)
Citation preview

KAJIAN INHIBITOR α-GLUKOSIDASE AKTINOMISET ENDOFIT ASAL BROTOWALI
(Tinospora crispa)
SRI PUJIYANTO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012


PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi ”Kajian Inhibitor α-
Glukosidase Aktinomiset Endofit Asal Brotowali (Tinospora crispa)” adalah
karya saya dengan arahan dari komisi pembimbing dan belum diajukan
dalam bentuk apapun kepada perguruan tinggi manapun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan maupun
tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan
dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2012
Sri Pujiyanto
NRP: G361070031


ABSTRACT
SRI PUJIYANTO. The Study of α-Glucosidase Inhibitor of Endophytic Actinomycetes Isolated from Brotowali (Tinospora crispa). Under direction of YULIN LESTARI, ANTONIUS SUWANTO, SRI BUDIARTI and LATIFAH K. DARUSMAN.
An α-glucosidase inhibitor is one of the compounds for the treatment of diabetes. This inhibitor can retard the liberation of glucose from dietary complex carbohydrates and delay glucose absorption, resulting in reduced postprandial plasma glucose levels and suppress postprandial hyperglycaemia. Diabetic medicinal plants are potencial sources of α-glucosidase inhibitor-producing endophytic microorganisms. Actinomycetes has been known as a source of various bioactive compounds that have been used in human health. The purpose of this study were to isolate and to select α-glucosidase inhibitor-producing endophytic actinomycetes from Tinospora crispa and to characterize the selected
isolated and their α-glucosidase inhibitor. Endophytic actinomycetes were isolated from the roots, leaves and stems of T. crispa. Sterilized plant samples were inoculated on the HV Agar medium containing 50 ppm cycloheximide and 30 ppm nalidixic acid and were incubated for 2-3 weeks at room temperature. All actinomycetes isolates were tested for their ability to inhibit the α-glucosidase. Identification for the selected isolates was based on 16S rDNA sequences. The inhibitor activity to α-glucosidase was determined spectrophotometrically at 400 nm using p-nitrophenyl-α-D-glucopyranoside as a substrate, and acarbose as a positive control. Characterization and identification of inhibitor component was based on thin layer chromatography, column chromatography, phytochemicals, UV-Vis spectrophotometer and FTIR analyses.The results showed that endophytic actinomycetes isolated from T. crispa produced various inhibition activities. The highest inhibition activity to α-glucosidase was shown by BWA65 found from T. crispa. Production of α-glucosidase inhibitor compounds in this
plant largely related with the contribution of its actinomycetes endophytes. The identification based on 16S rDNA sequence revealed that the isolate has 98% similarity to Streptomyces diastaticus. Separation of bioactive components by
column chromatography obtained active fraction (F6), which has a low IC50 and high inhibitory activity. Based on phytochemicals, UV-Vis spectrophotometer and FTIR analyses, the active compound F6 is an auron group of flavonoid.
Key words: α-glucosidase inhibitor, endophytic actinomycetes, diabetes mellitus, Tinospora crispa.


RINGKASAN
SRI PUJIYANTO. Kajian Inhibitor α-Glukosidase Aktinomiset Endofit Asal Brotowali (Tinospora crispa). Dibimbing oleh YULIN LESTARI, ANTONIUS
SUWANTO, SRI BUDIARTI dan LATIFAH K. DARUSMAN
Pengobatan diabetes secara tradisional pada umumnya adalah dengan memanfaatkan berbagai jenis tanaman yang memiliki kandungan bahan aktif yang dapat menurunkan kadar gula dalam darah, diantaranya adalah senyawa yang dapat menghambat enzim α-glukosidase. Eksplorasi mikrob endofit diharapkan dapat menghasilkan metabolit sekunder penting yang memiliki khasiat sama dengan metabolit yang dihasilkan tanaman inangnya. Tanaman obat diabetes merupakan sumber mikrob potensial penghasil inhibitor α-glukosidase. Isolat potensial dari tanaman obat tersebut diharapkan dapat digunakan untuk memproduksi senyawa inhibitor α-glukosidase secara mikrobiologis.
Tujuan dari penelitian ini adalah untuk mendapatkan isolat aktinomiset endofit dari tanaman obat diabetes, khususnya Tinospora crispa (brotowali) yang berpotensi sebagai penghasil senyawa inhibitor α-glukosidase, mengkarakterisasi dan mengidentifikasi isolat aktinomiset endofit penghasil inhibitor α-glukosidase terpilih, serta mengkarakterisasi dan mengidentifikasi senyawa inhibitor α-glukosidase yang dihasilkan oleh isolat aktinomiset endofit terpilih.
Berdasar kerangka pemikiran bahwa: a) inhibitor α-glukosidase dapat dihasilkan oleh berbagai organisme, yaitu tanaman tingkat tinggi, alga, cendawan, bakteri (non aktinomiset) dan aktinomiset, b) terdapat beberapa temuan penelitian yang menyatakan bahwa beberapa mikrob endofit dapat menghasilkan senyawa yang serupa dengan tanaman inangnya, dan c) tanaman T. crispa secara empiris diketahui memiliki aktivitas antidiabetes serta menghasilkan senyawa inhibitor α-glukosidase, maka dapat dirumuskan suatu hipotesis: aktinomiset endofit penghasil senyawa inhibitor α-glukosidase dapat diisolasi dari tanaman T. crispa. Isolat aktinomiset yang diperoleh mampu menghasilkan senyawa inhibitor α-glukosidase secara in vitro.
Isolasi aktinomiset endofit dilakukan dari T. crispa menggunakan media Humic Acid Vitamin B (HV) agar dengan penambahan 50 ppm cycloheximide dan 30 ppm nalidixic acid. Sebelum dilakukan isolasi, sampel tanaman disterilisasi
permukaan menggunakan natrium biklorit dan alkhohol 70% secara bertahap. Aktivitas inhibitor α-glukosidase diuji berdasarkan pada penghambatan pemecahan substrat p-Nitrophenyl-α-D-glucopyranoside oleh enzim α-
glukosidase selama periode tertentu. Senyawa acarbose (Sigma) digunakan sebagai pembanding.
Untuk mengetahui peran aktinomiset endofit dalam menghasilkan inhibitor α-glukosidase, dilakukan pengujian aktivitas inhibitor α-glukosidase terhadap tanaman T. crispa bebas endofit yang diperoleh dari kultur jaringan tanaman,
tanaman asli yang diperoleh dari alam dan isolat aktinomiset endofit terpilih, kemudian hasilnya dibandingkan.
Identifikasi isolat terpilih dilakukan berdasarkan sekuen 16S rDNA. Pengamatan morfologi isolat dilakukan menggunakan mikroskop cahaya pada perbesaran 100 dan 400x dan SEM pada perbesaran 10.000x.
Deteksi keberadaan gen yang terlibat dalam biosintesis inhibitor α-glukosidase dilakukan dengan amplifikasi gen penyandi sedoheptulosa-7-fosfat siklase menggunakan primer VOG-F 5'GGSGGSGGSGTSCTSATGGACGT-

SGCSGG-3', dan primer VOG-R 5'GCCATGTCSACGCASACSGCSGCCTCS-CCGAG-3'. Hasil amplifikasi DNA dengan PCR selanjutnya disekuen dan dibandingkan dengan sekuen yang ada di database GenBank.
Produksi senyawa inhibitor menggunakan media produksi cair berisi 0.1% soluble starch, 0.5% pepton, dan 0.1% yeast extract . Ekstraksi senyawa inhibitor
dilakukan dengan menggunakan pelarut etil asetat. Fraksinasi senyawa aktif dilakukan dengan kromatografi kolom silika gel dengan eluen heksan:etil asetat 1:4. Analisis fraksi dilakukan dengan bantuan KLT analitik. Fraksi yang diperoleh diuji aktivitas inhibitor α-glukosidasenya. Fraksi terbaik dikarakterisasi lebih lanjut meliputi: nilai IC50, pengaruh konsentrasi substrat, pemeriksaan fitokimia, scanning serapan maksimum dengan spektrofotometer UV-Vis dan pemeriksaan
dengan spektrofotometer FTIR. Tiga puluh dua isolat aktinomiset endofit telah berhasil diisolasi dari
tanaman T. crispa. Isolat BWA65 dari tanaman T. crispa merupakan isolat paling berpotensi untuk dikaji lebih lanjut. Penemuan isolat aktinomiset endofit dari T. crispa yang menghasilkan inhibitor α-glukosidase dalam penelitian ini,
memperkuat pendapat bahwa tanaman dapat mengandung mikrob endofit yang dapat menghasilkan beberapa senyawa biologis atau metabolit sekunder yang diduga sebagai hasil transfer genetik (rekombinasi genetik) dari tanaman inang ke mikrob endofit.
Tanaman T. crispa bebas endofit yang diperoleh dari kultur jaringan
tanaman hanya memiliki kemampuan yang sangat rendah untuk menghasilkan senyawa inhibitor α-glukosidase. Tanaman T. crispa yang diperoleh dari alam
mampu memproduksi senyawa inhibitor jauh lebih besar dibandingkan tanaman bebas endofit yang diperoleh dari kultur jaringan. Namun demikian, kemampuan inhibitor α-glukosidase yang dihasilkan oleh aktinomiset endofit BWA65 lebih dari dua kali daripada aktivitas inhibitor α-glukosidase tanaman inang. Hasil ini mengindikasikan bahwa aktinomiset endofit dalam tanaman T. crispa tersebut
memberikan kontribusi besar terhadap produksi senyawa inhibitor α-glukosidase. Hasil identifikasi berdasarkan sekuen 16S rDNA sepanjang 1343 pasang
basa menunjukkan bahwa isolat BWA65 memiliki kesamaan 98% dengan Streptomyces diastaticus. Kajian tentang inhibitor α-glukosidase yang dihasilkan oleh S. diastaticus hingga saat ini belum pernah dilaporkan. Berdasarkan hal
tersebut, kemungkinan besar isolat BWA65 merupakan jenis baru penghasil inhibitor α-glukosidase.
Analisis menggunakan program BLASTX terhadap sekuen basa hasil amplifikasi gen sedoheptulosa-7-fosfat siklase dari isolat BWA65 menunjukkan kemiripan paling tinggi 92% dengan GAF sensor hybrid histidine kinase pada Streptomyces violaceusniger Tu4113 (no akses YP-004812094.1). Amplifikasi
ulang gen sedoheptulosa-7-fosfat siklase pada isolat aktinomiset BWA65 ini dengan primer Forward: 5’-CCTACGAGGTGCGCTTCCGGGACGACGT-3’ dan Reverse: 5’-GGCGGCCTGCAGCTCGGCGGCCGTCACGT-3’, berhasil meng-
amplifikasi gen tersebut secara spesifik sepanjang 300 bp. Analisis hasil sekuensing menggunakan program BLASTX menunjukkan kemiripan 100% dengan gen sedoheptulosa-7-fosfat siklase dari Actinoplanes sp. SE50/110 (no akses Y18523.4) (Velina 2012, unpublished).
Fraksinasi senyawa aktif dari ekstrak etil asetat kultur BWA65 diperoleh fraksi aktif (Fraksi F6) sebagai fraksi paling potensial. Fraksi ini memiliki aktivitas penghambatan sebesar 80.9% pada konsentrasi 200 ppm atau setara dengan 96.2% dari aktivitas acarbose (aktivitas penghambatan 84.1%). Pada pengujian nilai IC50, Fraksi F6 memiliki nilai IC50 10.9 ppm, lebih kecil dibanding nilai IC50
acarbose sebesar 36.65 ppm. Keunggulan senyawa pada fraksi F6 ini pada konsentrasi yang rendah (dibawah 100 ppm) aktivitasnya lebih tinggi dibanding

dengan acarbose. Pada konsentrasi 12.5; 25 dan 50 ppm fraksi F6 memiliki aktivitas penghambatan sebesar 143.1%; 152.4% dan 150.1% dibanding acarbose pada konsentrasi yang sama. Hasil pengujian aktivitas inhibitor dengan berbagai konsentrasi substrat p-Nitrophenyl-α-D-glucopyranoside menunjukkan
bahwa senyawa inhibitor α-glukosidase yang dihasilkan isolat BWA65 diduga memiliki tipe penghambatan kompetitif.
Berdasarkan hasil pemeriksaan dengan spektrofotometer FTIR dan penelusuran literatur mengindikasikan senyawa Fraksi F6 ini memiliki gugus fungsi: karbonil C=O, ikatan O-H, ikatan C-O, ikatan rangkap dan C=C cincin aromatik, serta memiliki serapan panjang gelombang maksimum 423 nm saat diuji dengan spektrofotometer UV-Vis. Berdasarkan hasil pemeriksaan menggunakan uji fitokimia, spektrofotometer UV-Vis maupun FTIR mengindikasikan bahwa senyawa aktif inhibitor α-glukosidase yang dihasilkan oleh isolat BWA65 merupakan senyawa flavonoid kelompok auron.
Kata kunci: aktinomiset endofit, inhibitor α-glukosidase, diabetes melitus,
tanaman obat


© Hak cipta milik IPB, tahun 2012
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apapun tanpa izin IPB


KAJIAN INHIBITOR α-GLUKOSIDASE AKTINOMISET
ENDOFIT ASAL BROTOWALI (Tinospora crispa)
SRI PUJIYANTO
Disertasi sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Mikrobiologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012

Penguji Luar pada Ujian Tertutup : 1. Prof. drh. Dondin Sajuthi, PhD
2. Dr. Yanti, MSi
Penguji Luar pada Ujian Terbuka : 1. Dr. Raymond R. Tjandrawinata
2. Dr. Aris Tri Wahyudi

HALAMAN PENGESAHAN
Judul Disertasi : Kajian Inhibitor α-Glukosidase Aktinomiset Endofit Asal
Brotowali (Tinospora crispa)
Nama : Sri Pujiyanto
NRP : G361070031
Disetujui
Komisi Pembimbing
Dr. Ir. Yulin Lestari Ketua
Prof. Dr. Ir. Antonius Suwanto, MSc. Anggota
Dr. dr. Sri Budiarti Anggota
Prof. Dr. Ir. Latifah K. Darusman, MS. Anggota
Diketahui
Ketua Program Studi Mikrobiologi
Dekan Sekolah Pascasarjana,
Dr. Ir. Gayuh Rahayu Dr. Ir. Dahrul Syah, MSc.Agr.
Tanggal Ujian:………………………. Tanggal Lulus:…………………


PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT, atas rahmat dan
karuniaNya sehingga penulis dapat menyelesaikan penulisan disertasi dengan
judul “Kajian Inhibitor α-Glukosidase Aktinomiset Endofit Asal Brotowali
(Tinospora crispa)”. Disertasi ini merupakan karya ilmiah penulis selama
mengikuti Program Doktor pada Program Studi Mikrobiologi, Sekolah
Pascasarjana Institut Pertanian Bogor.
Penulis menyadari bahwa karya disertasi ini tidak mungkin tercipta tanpa
bimbingan dari komisi pembimbing, untuk itu penulis menyampaikan
penghargaan yang setinggi-tingginya serta ucapan terimakasih kepada: Dr. Ir.
Yulin Lestari selaku ketua komisi pembimbing serta Prof. Dr. Ir. Antonius
Suwanto, MSc, Dr. dr. Sri Budiarti dan Prof. Dr. Ir. Latifah K. Darusman, MS
selaku anggota komisi pembimbing atas segala bimbingan, arahan, perhatian,
nasehat, motivasi serta keteladanan yang telah diberikan kepada penulis mulai
dari awal pemilihan tema penelitian, selama pelaksanaan penelitian hingga
penulisan disertasi.
Penelitian ini didanai oleh Hibah Penelitian Pascasarjana a.n. Dr. Ir. Yulin
Lestari, serta sebagian dari Beasiswa Pendidikan Pascasarjana (BPPS), untuk
itu penulis menyampaikan rasa terimakasih yang sebesar-besarnya.
Ucapan terimakasih penulis sampaikan kepada Prof. drh. Dondin Sajuthi,
PhD dari PS Satwa Primata, IPB dan Dr. Yanti, MSi dari Fakultas Teknobiologi
Universitas Atmajaya (selaku penguji luar pada sidang tertutup), Prof. Dr. Okky
Setyawati Dharmaputra (selaku wakil PS Mikrobiologi), drh. Sulistiyani, MSc,
PhD (selaku wakil Dekan FMIPA) serta Dr. Ir. Aris Tri Wahyudi, MSi dari PS
Mikrobiologi, IPB dan Dr. Raymond R. Tjandrawinata dari PT. DEXA MEDICA
(selaku penguji luar pada sidang terbuka), Dr. Ir. Ence Darmo Jaya Supena
(selaku wakil PS Mikrobiologi/Dept. Biologi) dan Dr. Ir. Sri Nurdiati (Dekan FMIPA
IPB) atas saran-saran yang diberikan untuk kesempurnaan penulisan disertasi ini.
Penulis menyampaikan rasa terimakasih kepada Dr. Ir. Gayuh Rahayu
selaku ketua PS Mikrobiologi serta seluruh staf pengajar PS Mikrobiologi atas
curahan ilmu selama menempuh studi di PS Mikrobiologi SPs IPB. Penulis juga
mengucapkan terimakasih kepada Kepala Bagian Mikrobiologi Departemen
Biologi dan Kepala Bagian Kimia Analitik Departemen Kimia, Fakultas MIPA IPB
yang telah memberi ijin penulis untuk melaksanakan penelitian ini. Kepada
seluruh tehnisi Lab. Mikrobiologi, Dept. Biologi dan Lab. Kimia Analitik, Dept.

Kimia dan PS Biofarmaka, penulis menyampaikan terimakasih atas segala
bantuannya dalam penyelesaian penelitian ini. Kepada seluruh mahasiswa S3,
S2 dan S1 sesama peneliti di Lab Mikrobiologi, penulis sampaikan terimakasih
atas kerjasama yang baik dan saling pengertiannya.
Kepada teman seperjuangan Dr. Ratih Dewi Hastuti dan Dr. Desniar serta
teman-teman dari Undip Semarang, Dr. Sunarno, Dr. Jumari, Dr. Sri Widodo
Agung Suedy dan Dr. Fuad Muhammad, penulis sampaikan terimakasih atas
diskusi-diskusi, kebersamaan dan bantuannya dalam penyelesaian studi ini.
Semoga kebersamaan ini tetap terjaga selamanya.
Kepada kedua orang tua penulis, Bapak Dwijosumarto dan Ibu Suresmi,
juga adik Sri Ristini dan keluarga, terimakasih atas doa yang tulus tiada henti
yang selalu mengiringi setiap langkah untuk penyelesaian studi S3 ini. Kepada
almarhum Bapak/Ibu mertua, penulis sampaikan terimakasih atas doa dan
curahan waktu untuk merawat anak-anak sebelum meninggalkan kami
selamanya. Kepada keluarga tercinta, istriku Nining Indrawati, SKom, MAB dan
anak-anakku Annisa Ismatul Jannah, Nadia Rizka Izzati dan Royan Rasyid
Habibie terima kasih atas kesabaran, pengorbanan, pengertian, dorongan dan
doa yang senantiasa menemani perjalanan S3 ini. Karya ini saya persembahkan
kepada keluarga tercinta.
Penulis menyadari bahwa tiada karya yang sempurna, untuk itu segala
masukan dan saran perbaikan senantiasa penulis harapkan. Semoga karya
ilmiah ini bermanfaat.
Bogor, Agustus 2012
Sri Pujiyanto

RIWAYAT HIDUP
Penulis dilahirkan di Klaten pada tanggal 13 Januari 1973, sebagai anak
pertama dari Bapak Dwijo Sumarto dan Ibu Suresmi. Pendidikan Sarjana Biologi
dalam bidang Mikrobiologi diselesaikan di Universitas Diponegoro pada tahun
1998. Pendidikan Magister Sains dalam bidang Mikrobiologi diselesaikan di
Institut Pertanian Bogor pada tahun 2001. Selanjutnya, penulis mendapatkan
kesempatan melanjutkan sekolah S3 di Program Studi Mikrobiologi, Sekolah
Pascasarjana, Institut Pertanian Bogor pada tahun 2007 melalui program
Beasiswa Pendidikan Pasca Sarjana (BPPS) Direktorat Jenderal Pendidikan
Tinggi. Penulis adalah staf pengajar di Jurusan Biologi, FMIPA Universitas
Diponegoro, Semarang. Hingga kini, penulis tercatat sebagai anggota himpunan
profesi Perhimpunan Mikrobiologi Indonesia (PERMI). Selama menjadi
mahasiswa S3, penulis telah mempresentasikan sebagian dari penelitian
disertasi pada International Seminar of Indonesian Society for Microbiology,
tahun 2010 di Bogor, dengan judul “The Activity of α-Glucosidase Inhibitor of
Endophytic Actinomycetes Isolated from Indonesian Diabetic Medicinal Plants”.
Pada tahun 2012, sebagian hasil penelitian ini telah dipublikasikan di
International Journal of Pharmacy and Pharmaceutical Science, Vol 4, tahun
2012, dengan judul “Αlpha Glucosidase Inhibitor Activity and Characterization of
Endophytic Actinomycetes Isolated from some Indonesian Diabetic Medicinal
Plants”.


DAFTAR ISI
Halaman
DAFTAR TABEL……………………………………………………………………. xiii
DAFTAR GAMBAR…………………………………………………………………. xv
DAFTAR LAMPIRAN……………………………………………………………….. xvi
PENDAHULUAN
Latar Belakang……………………………………………………………… 1
Perumusan Masalah……………………………………………………….. 6
Tujuan Penelitian…………………………………………………………… 6
Manfaat Penelitian…………………………………………………………. 6
Novelty………………………………………………………………………. 7
Hipotesis…………………………………………………………………….. 7
TINJAUAN PUSTAKA
Penyakit Diabetes Melitus………………………………………………… 9
Inhibitor α-Glukosidase dalam Pengobatan Diabetes…………………. 9
Tanaman Obat Diabetes…………………………………………………... 13
Tanaman Brotowali ………………………………………………….…….. 16
Mikrob Endofit………………………………………………………………. 18
Aktinomiset Endofit dan Potensinya……………………………………… 19
METODE PENELITIAN
Tempat dan Waktu Penelitian…………………………………………….. 23
Isolasi Aktinomiset Endofit………………………………………………… 23
Seleksi dan Uji Penghambatan Enzim α-Glukosidase…………………. 24
Peran Aktinomiset Endofit dalam Menghasilkan inhibitor α-Glukosidase ………………………………………………………………
24
Penentuan Waktu Produksi Optimum……………………………………. 25
Identifikasi Isolat Terpilih………………………………………………….. 25
Karakterisasi Morfologi Isolat……………………………………………... 26
Deteksi Gen Sedoheptulosa-7-Fosfat Siklase………………………….. 26

Isolasi dan Identifikasi Senyawa Inhibitor α-Glukosidase…………....... 27
HASIL DAN PEMBAHASAN
Isolasi Aktinomiset Endofit………………………………………………… 33
Penapisan Aktinomiset Endofit Penghasil Inhibitor α-Glukosidase....... 35
Karakterisasi Morfologi Isolat BWA65…………………………………… 40
Identifikasi Isolat Aktinomiset……………………………………………... 41
Deteksi Gen Sedoheptulosa-7-Fosfat Siklase………………………….. 43
Ekstraksi Senyawa Inhibitor ……………………………………………… 45
Uji Aktivitas Inhibitor α-Glukosidase Ekstrak……………………….. 46
Anaisis Fitokimia Ekstrak Etil Asetat………………………………… 48
Fraksinasi dengan Kromatografi Kolom…………………………………. 49
Pencarian Eluen Terbaik……………………………………………… 50
Uji Inhibisi α-Glukosidase Fraksi…………………………………….. 52
Karakterisasi dan Identifikasi Senyawa Inhibitor α-Glukosidase …….. 55
Pemeriksaan dengan KLT Analitik…………………………………… 55
Pengaruh Konsentrasi Substrat……………………………………… 57
Identifikasi dengan Uji Fitokimia……………………………………... 59
Analisis dengan Spektrofotometer UV-Vis………………………….. 60
Identifikasi Gugus Fungsi dengan FTIR …………………………… 64
Temuan Penting dan Implikasinya…………………………………… 66
KESIMPULAN DAN SARAN………………………………………………………. 69
DAFTAR PUSTAKA………………………………………………………………... 71
LAMPIRAN………………………………………………………………………...... 79

DAFTAR TABEL
Halaman
1 Beberapa organisme penghasil senyawa inhibitor α-glukosidase…... 13
2 Kajian tanaman obat diabetes di berbagai negara……………………..
14
3 Kajian aktinomiset endofit pada beberapa tanaman dan potensinya...
21
4 Komposisi reaksi PCR:amplifikasi gen 16S rRNA……………………...
25
5 Komposisi reaksi PCR amplifikasi gen Sedoheptulosa-7-fosfat siklase……………………………………………………………………….
27
6 Isolat aktinomiset endofit dari berbagai tanaman obat diabetes dan aktivitas inhibitor α-glukosidase yang dihasilkannya…………………..
35
7 Karakteristik kultur isolat BWA65 pada berbagai media……………….
40
8 Hasil analisis sekuen DNA hasil amplifikasi gen penyandi sedoheptulosa-7-fosfat siklase isolat BWA65 menggunakan program BLASTX……………………………………………………………………..
45
9 Hasil ekstraksi senyawa inhibitor dengan berbagai pelarut……………
46
10 Aktivitas inhibitor α-glukosidase berbagai ekstrak kultur aktinomiset BWA65 ……………………………………………………………………..
47
11 Kandungan fitokimia ekstrak etil asetat kultur BWA65…………………
49
12 Hasil percobaan KLT untuk menentukan pelarut terbaik……………....
50
13 Hasil fraksinasi senyawa menggunakan kromatografi kolom gel silika…………………………………………………………………………
52
14 Aktivitas inhibitor α-glukosidase hasil fraksinasi kolom………………...
52
15 Aktivitas inhibitor α-glukosidase beberapa senyawa dibanding acarbose ……………………………………………………………………
54
16 Hasil KLT analitik Fraksi F6 dengan beberapa eluen…………………..
57
17 Pengaruh konsentrasi substrat terhadap kemampuan inhibitor α-glukosidase hasil fraksinasi……………………………………………..
57
18 Rentangan serapan spektrum UV-Vis flavonoid ……………..………...
62
19 Ciri spektrum golongan flavonoid utama ………………………………..
62
20 Identifikasi gugus fungsi fraksi F6 dengan FTIR………………………. 65


DAFTAR GAMBAR Halaman
1 Diagram alur kerangka pemikiran penelitian …………………………......
5
2 Struktur senyawa acarbose …………………………………………………
10
3 Jumlah isolat aktinomiset endofit isolat dari tanaman obat………...........
33
4 Jumlah isolat aktinomiset endofit berdasarkan asal bagian tanaman.....
34
5 Aktivitas inhibitor α-glukosidase oleh aktinomiset endofit tanaman T. crispa………………………………………………………………………….
38
6 Aktivitas inhibitor α-glukosidase yang dihasilkan oleh tanaman hasil kultur jaringan tanaman, tanaman dari alam dan isolat aktinomiset endofit ………………………………………………………………………...
39
7 Morfologi isolat aktinomiset endofit BWA65 yang diamati pada media Oatmeal Agar, mikroskop cahaya dan SEM ………………………………
41
8 Pohon filogenetik isolat BWA65 berdasarkan sekuen 16S rDNA............
42
9 Hasil amplifikasi PCR menggunakan primer VOG-F dan VOG-R ….......
44
10 Nilai IC50 berbagai ekstrak kultur BWA65…………………………………. 48
11 Kromatografi lapis tipis dengan variasi campuran pelarut pengembang heksan: etil asetat 1:4………………………………………………………..
51
12 Nilai IC50 dari fraksi aktif hasil kromatografi kolom gel silika……………..
53
13 Aktivitas inhibitor α-glukosidase fraksi terpilih (Fraksi F3 dan Fraksi F6) dibandingkan acarbose………………………………………………….
55
14 Hasil kromatografi lapis tipis Fraksi F6 menggunakan berbagai jenis eluen …………………………………………………………………………..
56
15 Pengaruh konsentrasi substrat terhadap kemampuan inhibitor α-glukosidase hasil fraksinasi………………………………………………….
58
16 Kurva Lineweaver-Burke aktifitas α-glukosidase …………………………
58
17 Spektrum spektrofotometer UV-Vis Fraksi F6………………....................
61
18 Spektrum spektrofotometer UV-Vis Fraksi F3 …………………………….
61
19 Spektrum FTIR Fraksi F6……………………………………………………
54
20 Struktur dasar senyawa auron……………………………………………… 65 21
Tanaman brotowali…………………………………………………………...
87


DAFTAR LAMPIRAN
Halaman
1 Uji aktivitas inhibitor dan penentuan nilai IC50 ekstrak ……………………...
79
2 Hasil uji aktivitas hasil kromatografi kolom dan penentuan nilai IC50..........
81
3 Hasil sekuensing gen 16S rRNA isolat BWA65……………………………...
83
4 Hasil analisis BLAST sekuen gen 16S rRNA isolat BWA65……………….. 85
5 Tanaman brotowali……………………………………………………………... 87

1
PENDAHULUAN
Latar Belakang
Diabetes melitus (DM) merupakan salah satu masalah utama kesehatan
dunia. Menurut perkiraan, penderita diabetes di seluruh dunia pada tahun 2025
mendatang akan mencapai 300 juta orang. Sebagian besar (lebih dari 95%)
penderita diabetes merupakan penderita diabetes tipe 2 atau sering disebut non-
insulin dependent diabetes (Bailey & Day 2003). DM atau kencing manis
merupakan penyebab kematian tertinggi di antara penyakit non infeksi lainnya.
Sekitar 1.08 juta kematian akibat penyakit kardiovaskular (pembuluh darah) yang
terjadi di seluruh dunia setiap tahunnya, sebanyak 851 ribu di antaranya
merupakan pasien DM.
Gejala penyakit diabetes adalah adanya keadaan hiperglikemia
(peningkatan kadar gula darah) yang terus-menerus dan bervariasi, terutama
setelah makan. Penyakit diabetes mellitus dapat menyebabkan adanya
komplikasi pada tingkat lanjut. Hiperglikemia dapat menyebabkan dehidrasi dan
ketoasidosis. Komplikasi jangka lama termasuk penyakit kardiovaskular,
kegagalan kronis ginjal, kerusakan retina yang dapat menyebabkan kebutaan,
serta kerusakan saraf yang dapat menyebabkan impotensi dan gangren dengan
risiko amputasi.
Penyakit DM merupakan penyakit yang tidak dapat disembuhkan, tetapi
dapat dikontrol. Pengobatan penyakit diabetes pada prinsipnya adalah suatu
usaha untuk menjaga agar kadar glukosa darah dapat dipertahankan pada
kondisi normal (80-120 mg/dl). Berbagai pilihan obat diabetes baik modern
maupun tradisional telah dikenal di masyarakat. Saat ini terdapat berbagai
pilihan obat diabetes oral yang tersedia untuk penderita diabetes tipe 2 (Bailey &
Day 2003).
Penggunaan inhibitor enzim dalam pengobatan penyakit telah lazim
digunakan dan mengalami perkembangan yang pesat. Nilai ekonomi obat yang
tergolong inhibitor enzim diperkirakan mencapai 95.57 milyar dolar pada tahun
2006 (Copeland 2005). Demikian juga dalam pengobatan diabetes, salah satu
target obat diabetes adalah menghambat enzim α-glukosidase. Enzim α-
glukosidase (EC 3.2.1.20) ini berperan dalam pencernaan karbohidrat komplek
(amilum) menjadi glukosa di dalam usus halus. Enzim ini menghidrolisis ikatan α-
1,4-D-glukosa dengan membebaskan molekul glukosa. Dengan dihambatnya

2
aktivitas enzim ini, maka asupan glukosa dari usus ke dalam darah dapat
dikurangi. Senyawa aktif yang memiliki aktivitas seperti ini adalah inhibitor α-
glukosidase. Beberapa senyawa kimia yang telah diketahui merupakan inhibitor
α-glukosidase antara lain: pradimicin (Yosuke et al. 1992), deoxynojirimycin
(Fischer et al. 1995), acarbose (Hemker et al. 2001), curcumin (Du et al. 2006)
dan ceptezole, suatu antibiotik beta laktam (Lee et al. 2007). Beberapa ekstrak
tanaman seperti: Pinus densiflora (Kim et al. 2003), Phalleria macrocarpa
(Sugiwati et al. 2006) dan Terminalia sp. (Anam et al. 2009) juga diketahui
memiliki aktivitas inhibitor α-glukosidase. Selain tanaman, organisme lain seperti
cendawan Ganoderma lucidum (Kim & Nho 2004) dan alga (Cannell et al. 1988)
serta aktinomiset juga diketahui memiliki aktivitas inhibitor α-glukosidase.
Beberapa aktinomiset yang diketahui menghasilkan inhibitor α-glukosidase
adalah: Actinomadura verrucosospora (Yosuke et al. 1992), Actinoplanes sp.
(Gown 2006), Streptomyces clavuligerus (Lee et al. 2007) dan Micromonospora
sp. (Suthindhiran et al. 2009).
Senyawa inhibitor α-glukosidase yang telah sukses dikomersialkan
adalah acarbose, suatu pseudooligosakarida yang memiliki struktur mirip glukosa.
Senyawa ini dihasilkan oleh Actinoplanes sp., suatu aktinomiset yang diisolasi
dari suatu daerah di Kenya dan dikomersialkan oleh perusahaan Bayer, Jerman
dengan nilai jual 379 juta US dolar pada tahun 2004 (Gown 2006).
Pengobatan diabetes secara tradisional pada umumnya dengan
memanfaatkan berbagai jenis tanaman yang memiliki kandungan bahan aktif
yang dapat menurunkan kadar gula dalam darah. Berbagai tanaman obat yang
secara empiris diketahui memiliki khasiat sebagai agen hipoglikemia antara lain
adalah: brotowali (Tinospora crispa), pinus (Pinus densiflora), lidah buaya (Alloe
vera), bawang putih (Allium sativum), bawang merah (Allium cepa), stevia
(Stevia rebaudiana), ubi jalar (Ipomoea batatas), sambiloto (Andrographis
paniculata), mengkudu (Morinda citrifolia), delima (Punica granatum), kelapa
(Cocos nucifera), jambelang (Eugenia jambolana), mahkota dewa (Phaleria
macrocarpha), ginseng (Panax sp), buah merah (Pandanus conoideus) dan pare
(Momordica charantia) (Subroto 2006; Klein et al. 2007; Bnouham et al. 2006).
Brotowali (Tinospora crispa) telah lama di kenal oleh masyarakat
tradisional Indonesia sebagai bahan pembuatan jamu yang di campur dengan
tanaman-tanaman herbal lainnya. Brotowali merupakan tanaman herba (perdu)
yang hampir semua bagian dari tubuhnya di manfaatkan sebagai obat serba

3
guna yang dapat menyembuhkan berbagai macam penyakit seperti: kencing
manis, hipertensi, dan demam. Kajian ilmiah terhadap tanaman brotowali sebagai
obat diabetes telah dilakukan para peneliti. Noor dan Ashcroft (1998) melaporkan
bahwa ekstrak T. crispa mampu menstimulasi peningkatan sekresi insulin,
sehingga dapat berperan sebagai antihiperglikemia. Noipa dan Ninlaaesong
(2011) melaporkan bahwa ekstrak T. crispa dapat meningkatkan laju transpor
glukosa ke dalam sel. Uji klinis oleh Sriyapai et al. (2009) menunjukkan bahwa
konsumsi serbuk T. crispa dapat menurunkan kadar gula darah setelah makan.
Chougale et al. (2009) melaporkan bahwa ekstrak tanaman T.cordifolia memiliki
aktivitas inhibitor α-glukosidase. Patela dan Mishrab (2012) menemukan tiga
senyawa dari tanaman Tinospora cordifolia yang memiliki kemampuan inhibitor
α-glukosidase, yaitu jatrorrhizine, palmatine dan magnoflorine. Senyawa-
senyawa tersebut memiliki nilai IC50 berturut-turut sebesar 22.05, 38.42 and 7.6
µg/mL untuk jatrorrhizine, palmatine dan magnoflorine. Hasil uji in vivo juga
menunjukkan adanya penekanan yang signifikan terhadap kenaikan kadar
glukosa plasma darah oleh ketiga senyawa tersebut pada konsentrasi 20 mg/kg
berat badan. Senyawa magnoflorine merupakan senyawa paling potensial
sebagai inhibitor α-glukosidase di antara ketiga senyawa yang ditemukan
tersebut.
Selama ribuan tahun, senyawa bioaktif yang berasal dari alam telah
memegang peran penting dalam pengobatan dan upaya mempertahankan
kesehatan manusia. Senyawa-senyawa tersebut dapat dihasilkan dari berbagai
sumber seperti tanaman, hewan maupun mikroorganisme (Chin et al. 2006).
Bahan bioaktif alami selain dapat digunakan sebagai obat secara langsung, juga
berperan sebagai bahan baku dalam pembuatan senyawa sintetik (Topliss et al.
2002). Tanaman, merupakan salah satu sumber yang sangat penting dalam
upaya pengobatan dan upaya mempertahankan kesehatan masyarakat. Sampai
saat ini menurut perkiraan badan kesehatan dunia (WHO), 80% penduduk dunia
masih menggantungkan dirinya pada pengobatan tradisional termasuk
penggunaan obat yang berasal dari tanaman. Sampai saat ini seperempat dari
obat-obat modern yang beredar di dunia berasal dari bahan aktif yang diisolasi
dan dikembangkan dari tanaman (Radji 2005).
Indonesia yang memiliki keanekaragaman hayati terbesar kedua di dunia
berpotensi besar untuk mengembangkan obat herbal yang berbasis pada
tanaman obat. Lebih dari 1000 spesies tumbuhan dapat dimanfaatkan sebagai

4
bahan baku obat. Tumbuhan tersebut menghasilkan metabolit sekunder dengan
struktur molekul dan aktivitas biologis yang beraneka ragam serta memiliki
potensi yang sangat baik untuk dikembangkan menjadi obat berbagai penyakit
(Radji 2005).
Permasalahan yang muncul dari pemakaian obat herbal adalah
bagaimana menjaga tingkat produksi obat herbal tersebut dengan bahan baku
yang terbatas karena sebagian besar bahan baku obat herbal diambil dari
tanaman induknya. Terdapat kekhawatiran bahwa sumberdaya hayati ini akan
musnah karena adanya kendala dalam budidayanya. Bahkan disinyalir bahwa
bahan obat herbal yang diproduksi dan diedarkan di Indonesia saat ini sebagian
besar bahan bakunya sudah mulai diimpor dari beberapa negara lain (Radji
2005).
Mikrob endofit merupakan mikrob yang hidup di dalam jaringan tanaman
pada periode tertentu dan mampu membentuk koloni dalam jaringan tanaman
tanpa membahayakan inangnya. Mikrob endofit ini dapat berupa cendawan dan
bakteri termasuk aktinomiset. Menurut Tan dan Zou (2001) setiap tanaman
tingkat tinggi dapat mengandung sejumlah mikrob endofit yang mampu
menghasilkan senyawa biologi atau metabolit sekunder serupa dengan inangnya
yang diduga sebagai akibat koevolusi atau transfer genetik (genetic
recombination). Beberapa penelitian telah menunjukkan hasil yang mendukung
pendapat tersebut. Strobell dan Daisy (2003) melaporkan bahwa Taxomyces
andreanae endofit pada tanaman Taxus menghasilkaan paclitaxel. Paclitaxel
adalah senyawa antikanker yang dihasilkan juga oleh tanaman Taxus brevivolia.
Taechowisan et al. (2007) melaporkan Streptomyces aureofaciens, endofit pada
tanaman jahe menghasilkan senyawa arylcoumarin yang memiliki aktivitas
antitumor, dimana tanaman jahe juga memiliki senyawa anti tumor seperti
dilaporkan oleh Katiyar et al. (1996). Penelitian Castillo et al. (2002)
menunjukkan bahwa Streptomyces NRRL 30562 endofit pada tanaman Kennedia
nigriscans mampu menghasilkan antibiotik spektrum luas. Tanaman ini secara
tradisional digunakan suku aborigin untuk mencegah infeksi mikrob pada luka.
Aktinomiset telah lama diketahui sebagai sumber berbagai senyawa
bioaktif yang telah dimanfaatkan dalam berbagai bidang seperti kesehatan,
pertanian, maupun industri. Sekitar 70% senyawa bioaktif baru dihasilkan oleh
kelompok aktinomiset (Takahashi 2004). Berdasarkan fenomena tersebut, mikrob
endofit khususnya aktinomiset pada tanaman obat diharapkan dapat menjawab

5
Diabetes mellitus (DM) merupakan masalah utama kesehatan
(WHO 1999)
permasalahan keterbatasan pemakaian tanaman obat sebagai sumber senyawa
bioaktif. Eksplorasi mikrob endofit diharapkan dapat menghasilkan metabolit
sekunder penting yang memiliki khasiat sama dengan metabolit yang dihasilkan
tanaman inangnya. Beberapa tanaman obat khas Indonesia seperti: brotowali,
sambiloto, mahkota dewa, ciplukan, dan lain-lain telah diketahui secara empiris
memiliki khasiat sebagai obat diabetes. Tanaman obat diabetes merupakan
sumber potensial penghasil inhibitor α-glukosidase (Benalla et al 2010).
Berdasarkan pendapat Tan dan Zou (2001) yang menyatakan bahwa endofit
dapat menghasilkan senyawa yang sama dengan senyawa yang dihasilkan
tanaman inangnya maka tanaman obat diabetes merupakan sumber potensial
isolat penghasil inhibitor α-glukosidase. Isolat potensial dari tanaman obat
tersebut, diharapkan dapat digunakan untuk memproduksi senyawa inhibitor α-
glukosidase secara mikrobiologis, dengan jumlah yang lebih banyak dan
kualitas yang lebih baik. Dengan demikian, penelitian tentang potensi mikrob
endofit terutama aktinomiset dari tanaman obat diabetes khususnya brotowali
untuk menghasilkan senyawa inhibitor α-glukosidase yang berguna dalam
pengobatan penyakit diabetes perlu dilakukan. Kerangka pemikiran dari
penelitian ini secara ringkas dapat dilihat pada Gambar 1.
Gambar 1 Diagram alur kerangka pemikiran penelitian.
Aktinomiset sumber bioaktif terbesar (70%)
(Takahashi 2004)
Endofit mampu menghasilkan bioaktif sama dengan inangnya (Tan & Zou 2001; Strobel & Daisy 2003; Castillo et al. 2002)
Potensi aktinomiset endofit tanaman T. crispa sebagai penghasil
senyawa inhibitor α-glukosidase
Pengobatan diabetes: menjaga glukosa darah tetap normal. Inhibitor alfa glukosidase menunda digesti karbohidrat (Bailey & Day 2003)
Brotowali secara empiris sebagai antidiabet, inhibitor α-glukosidase (Subroto 2006; Chougale et al.
2009; Patela & Mishrab 2012)
Beberapa Aktinomiset menghasilkan inhibitor α- glukosidase (Gown 2006; Lee et al. 2007; Suthindhiran et al. 2009)

6
Perumusan Masalah
Inhibitor α-glukosidase dapat dihasilkan oleh berbagai organisme, yaitu
tanaman tingkat tinggi, alga, cendawan, bakteri non aktinomiset dan aktinomiset.
Tanaman dapat mengandung mikrob endofit yang mampu menghasilkan
senyawa biologi atau metabolit sekunder serupa dengan tanaman inangnya.
Beberapa tanaman obat, khususnya T. crispa secara empiris diketahui memiliki
aktivitas antidiabetes dan menghasilkan α-glukosidase inhibitor. Dengan
demikian, dari tanaman obat yang telah diketahui memiliki aktivitas sebagai
antidiabetes serta diketahui menghasilkan senyawa inhibitor α-glukosidase akan
dapat diperoleh isolat endofit khususnya aktinomiset yang dapat menghasilkan
senyawa inhibitor α-glukosidase pula. Berdasarkan hal tersebut, dapat
dirumuskan masalah sebagai berikut:
1. Apakah dari tanaman T. crispa yang telah diketahui memiliki khasiat
sebagai obat diabetes dapat diperoleh isolat aktinomiset endofit yang
dapat menghasilkan senyawa inhibitor α-glukosidase?
2. Bagaimanakah karakteristik dan identitas isolat aktinomiset endofit
penghasil inhibitor α-glukosidase yang diperoleh dari tanaman T. crispa
tersebut?
3. Bagaimana karakteristik senyawa inhibitor α-glukosidase yang dihasilkan
oleh isolat aktinomiset endofit tersebut?
Tujuan Penelitian
Tujuan dari penelitian ini adalah:
1. Mendapatkan isolat aktinomiset endofit dari tanaman T. crispa yang telah
dikenal memiliki khasiat sebagai antidiabetes, yang berpotensi sebagai
penghasil senyawa inhibitor α-glukosidase.
2. Mengkarakterisasi dan mengidentifikasi isolat aktinomiset endofit
penghasil senyawa inhibitor α-glukosidase terpilih.
3. Mengkarakterisasi dan mengidentifikasi senyawa inhibitor α-glukosidase
yang dihasilkan oleh isolat aktinomiset endofit terpilih.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menjadi landasan ilmiah untuk
memproduksi senyawa inhibitor α-glukosidase secara mikrobiologis yang dapat
digunakan sebagai agen antihiperglikemia. Inhibitor α-glukosidase memiliki nilai

7
komersial tinggi karena kebutuhan obat diabetes semakin meningkat seiring
dengan meningkatnya jumlah penderita diabetes.
Produksi senyawa inhibitor α-glukosidase secara mikrobiologis ini
diharapkan lebih murah dan lebih efisien dibandingkan dengan cara mengekstrak
dari tanaman obat secara langsung karena dibutuhkan tanaman obat dalam
jumlah yang besar serta sulitnya penyeragaman kualitas bahan baku tanaman
obat. Dengan demikian, hasil penelitian ini diharapkan dapat memberi terobosan
teknologi dalam penyediaan bahan obat diabetes khususnya di Indonesia.
Novelty
Berdasarkan penelusuran literatur yang telah dilakukan, hingga saat ini
belum ada laporan kajian tentang inhibitor α-glukosidase dari aktinomiset endofit,
apalagi yang diisolasi dari tanaman T. crispa yang merupakan salah satu
tanaman obat tradisional di Indonesia. Beberapa informasi yang menyebutkan
adanya aktinomiset penghasil inhibitor α-glukosidase di atas semuanya adalah
bukan aktinomiset endofit.
Dengan demikian penelitian ini memiliki unsur kebaharuan yang akan
menambah khasanah ilmu pengetahuan tentang potensi aktinomiset sebagai
penghasil senyawa bioaktif, khususnya senyawa inhibitor α-glukosidase dari
aktinomiset endofit tanaman T. crispa.
Hipotesis Penelitian
Berdasar kerangka pemikiran dari uraian pada latar belakang bahwa:
a) inhibitor α-glukosidase dapat dihasilkan oleh berbagai organisme, yaitu
tanaman tingkat tinggi, alga, cendawan, bakteri non aktinomiset dan
beberapa aktinomiset,
b) terdapat beberapa temuan penelitian yang menyatakan bahwa beberapa
mikrob endofit dapat menghasilkan senyawa yang serupa dengan
tanaman inangnya, dan
c) tanaman T. crispa diketahui memiliki aktivitas inhibitor α-glukosidase,
maka dapat dirumuskan suatu hipotesis: aktinomiset endofit penghasil senyawa
inhibitor α-glukosidase dapat diisolasi dari tanaman brotowali. Isolat aktinomiset
yang diperoleh mampu menghasilkan inhibitor α-glukosidase secara in vitro.

8

9
TINJAUAN PUSTAKA
Penyakit Diabetes Melitus
Diabetes melitus (DM) atau dikenal sebagai kencing manis adalah
penyakit yang ditandai dengan hiperglikemia (peningkatan kadar gula darah)
yang terus-menerus dan bervariasi, terutama setelah makan. Sumber lain
menyebutkan bahwa yang dimaksud dengan diabetes melitus adalah keadaan
hiperglikemia kronik disertai berbagai kelainan metabolik akibat gangguan
hormonal, yang menimbulkan berbagai komplikasi kronik pada mata, ginjal, dan
pembuluh darah, disertai lesi pada membran basalis (Bailey & Day 2003).
Semua jenis diabetes melitus memiliki gejala yang mirip dan komplikasi
pada tingkat lanjut. Hiperglikemia sendiri dapat menyebabkan dehidrasi dan
ketoasidosis. Komplikasi jangka lama termasuk penyakit kardiovaskular,
kegagalan kronis ginjal (penyebab utama dialisis), kerusakan retina yang dapat
menyebabkan kebutaan, serta kerusakan saraf yang dapat menyebabkan
impotensi dan gangren dengan risiko amputasi. Komplikasi yang lebih serius
lebih umum terjadi bila kadar gula darah tidak dikendalikan dengan baik (WHO
1999).
Penyebab diabetes yang utama adalah karena kurangnya produksi insulin
(DM tipe 1, yang pertama dikenal), atau kurang sensitifnya jaringan tubuh
terhadap insulin (diabetes melitus tipe 2, bentuk yang lebih umum). Selain itu,
terdapat jenis diabetes melitus yang juga disebabkan oleh resistensi insulin yang
terjadi pada wanita hamil. Tipe 1 membutuhkan penyuntikan insulin, sedangkan
tipe 2 diatasi dengan pengobatan oral dan hanya membutuhkan insulin bila
obatnya tidak efektif. Diabetes melitus pada kehamilan umumnya sembuh
dengan sendirinya setelah persalinan (WHO 1999).
Inhibitor α-Glukosidase dalam Pengobatan Diabetes
Inhibitor α- glukosidase merupakan obat diabetes yang digunakan per
oral untuk pengobatan diabetes melitus tipe 2 yang bekerja dengan cara
mencegah digesti karbohidrat terutama amilum menjadi glukosa. Karbohidrat
secara normal akan dikonversi menjadi gula sederhana atau monosakarida yang
dapat diabsorbsi melalui instestinum. Dengan demikian inhibitor α- glukosidase
berperan dalam mengurangi pengaruh karbohidrat terhadap kandungan gula
darah.

10
Inhibitor α- glukosidase digunakan untuk mempertahankan kadar gula
agar tidak terjadi hiperglikemia di dalam diabetes melitus type 2. Obat ini dapat
digunakan sebagai monoterapi atau digunakan bersama obat diabetes lainnya.
Inhibitor α- glukosidase dapat berupa senyawa sakarida yang berperan sebagai
inhibitor kompetitif dari enzim yang diperlukan untuk mendigesti karbohidrat,
khususnya enzim α- glukosidase yang terdapat di dalam fili-fili usus kecil. Enzim
α-glukosidase yang terdapat pada membran sel-sel usus kecil menghidrolisis
oligosakarida, trisakarida dan disakarida menjadi glukosa dan monosakarida
lainnya yang terdapat di dalam usus kecil.
Salah satu contoh inhibitor α-glukosidase adalah acarbose. Acarbose
merupakan inhibitor α-glukosidase yang telah diterima di Amerika untuk
pengobatan diabetes tipe 2. Efek langsung dari antidiabetes ini adalah menunda
digesti karbohidrat komplek dan disakarida menjadi monosakarida yang mudah
diabsorbsi yaitu glukosa. Hal ini dapat tercapai oleh adanya penghambatan
reversibel terhadap enzim α-glukosidase (termasuk sukrase dan maltase) yang
terdapat di dalam duodenum. Pada pasien diabetes tipe 2, enzim ini
menghambat penundaan absorbsi glukosa sebagai kelanjutan proses
pencernaan karbohidrat kompleks. Acarbose tidak menimbulkan pengaruh
secara langsung terhadap resistensi insulin atau pengambilan glukosa yang
distimulasi insulin pada manusia (Clark 1998).
Acarbose merupakan pseudooligosakarida yang dihasilkan oleh genus
Actinoplanes dan telah digunakan untuk pengobatan pasien diabetes. Senyawa
ini sangat efektif untuk menghambat kerja enzim α-amilase, α-glukosidase, dan
sukrase. Struktur senyawa acarbose dapat dilihat pada Gambar 2 berikut.
Gambar 2 Struktur senyawa acarbose (Zhang et al. 2002).

11
Inhibitor enzim memiliki nilai potensi digunakan dalam pengendalian dan
pengobatan berbagai jenis penyakit. Pengendalian kinetika pencernaan
karbohidrat dan absorbsi glukosa dapat digunakan sebagai sarana mencegah
dan terapi terhadap penyakit diabetes, obesitas, hiperlipoproteinemia dan
hiperlipidemia. Kaitannya dengan hal ini, inhibitor α-glukosidase merupakan
enzim hidrolase amilolitik yang membebaskan glukosa dari ujung non reduksi
dari molekul polisakarida dan oligosakarida (Kim & Nho 2004).
Senyawa inhibitor α-glukosidase SKG-3 telah berhasil diisolasi dari
Ganoderma lucidum dengan fraksinasi menggunakan berbagai teknik
kromatografi. Senyawa yang telah dipurifikasi dikonfirmasi menggunakan
kromatografi lapis tipis (KLT) dan High Pressure Liquid Chromatography (HPLC).
Senyawa SKG-3 murni menunjukkan spot tunggal pada pelat TLC dan
menunjukkan satu puncak pada HPLC dengan waktu retensi 13 menit. Pengaruh
konsentrasi SKG-3 terhadap daya hambat terhadap berbagai glikosidase diuji
dan hasilnya secara jelas menunjukkan bahwa SKG-3 potensial dalam
menghambat α-glukosidase yang ditunjukkan nilai IC50 sebesar 4,6 µg/ml. SKG-3
tidak menunjukkan aktivitas penghambatan terhadap beta glukosidase, beta
galaktosidase atau α-mannosidase yang diujikan pada konsentrasi 100 µg/ml
(Kim & Nho 2004).
Daya hambat SKG-3 terhadap α-glukosidase akan meningkat dengan
melakukan preinkubasi senyawa SKG-3 dengan enzim mengindikasikan bahwa
senyawa ini bereaksi lambat dengan enzim α-glukosidase. Senyawa SKG-3
dapat dipisahkan dari α-glukosidase dengan dialisis. Ketika α-glukosidase
dicampur dengan sejumlah SKG-3 yang menghasilkan penghambatan 90%,
campuran reaksi ditempatkan pada kantong dialisis, hampir semua enzim dapat
diperoleh kembali dan memiliki aktivitas yang tinggi. Hasil ini menunjukkan
bahwa penghambatan SKG-3 terhadap α- glukosidase bersifat reversibel (Kim &
Nho 2004).
Penelitian Xiancui et al. (2005) menunjukkan bahwa beberapa spesies
makro alga juga dapat menghasilkan senyawa inhibitor α-glukosidase. Ekstrak
kasar makroalga seperti Rhodomela confervoides, Gracillaria textorii, Plocamium
telfairie, Ulva pertusa dan Enteromorpha intestinalis menunjukkan adanya
penghambatan yang kuat terhadap α-glukosidase pada konsentrasi 79,6 µg/ml.
Hasil penelitian Cannell et al. (1988) diperoleh senyawa pentagalloyl-glucose,
suatu inhibitor α-glukosidase dari alga air tawar Spirogyra varians. Aktivitas

12
inhibitor α- glukosidase juga ditunjukkan dari ekstrak air douchi (produk makanan
fermentasi kedelai dari Cina). Kultur murni Aspergillus oryzae yang digunakan
untuk fermentasi douchi di laboratorium ini, mampu menghasilkan inhibitor α-
glukosidase lebih tinggi dari pada isolat Actinomucor elegans dan Rhizopus
arrhizus (Chen et al. 2004).
Suatu antibiotik beta laktam ceftezole, dilaporkan memiliki aktivitas
menghambat α-glukosidase. Uji in vitro terhadap α-glukosidase, senyawa ini
menunjukkan adanya hambatan yang reversibel. Senyawa SKG-3 diketahui juga
merupakan hambatan non kompetitif. Percobaan pada mencit diabet
menunjukkan bahwa pemberian ceptezole (10 mg/kg/hari) dapat menurunkan
kadar glukosa darah sebesar 30% dalam waktu 20 menit setelah pemberian
obat tersebut (Lee at al. 2007).
Senyawa inhibitor α-glukosidase juga dapat dihasilkan oleh binatang spon
laut Penares sp. Senyawa yang dihasilkan hewan tersebut adalah penarolide
sulfat A1 dan A2, suatu makrolida yang merupakan kelompok senyawa poliketida.
Senyawa tersebut memiliki nilai IC50 berturut-turut 1.2 dan 1.5 μg/ml (Nakao et
al. 2000). Kajian inhibitor α-glukosidase pada beberapa organisme diringkaskan
pada Tabel 1.
Berdasarkan informasi pada tabel tersebut dapat dilihat bahwa inhibitor α-
glukosidase dapat dihasilkan oleh berbagai kelompok organisme seperti:
tanaman, alga, cendawan, aktinomiset maupun bakteri non aktinomiset. Dari
tabel tersebut terlihat bahwa kelompok aktinomiset telah cukup banyak
dilaporkan mampu menghasilkan senyawa inhibitor α-glukosidase. Namun
demikian, dari semua aktinomiset penghasil inhibitor α-glukosidase tersebut,
tidak satupun yang dilaporkan sebagai aktinomiset endofit. Berdasarkan fakta
tersebut terbuka peluang yang besar untuk mendapatkan isolat-isolat baru
aktinomiset penghasil inhibitor α-glukosidase, khususnya aktinomiset endofit
yang diisolasi dari tanaman obat.

13
Tabel 1 Beberapa organisme penghasil senyawa inhibitor α- glukosidase
No Organisme penghasil
Bahan aktif Pustaka
Tumbuhan 1 Pisonia alba ekstrak etanol Sunil et al. 2009 2 Terminalia sp. ekstrak etanol 96% Anam et al. 2009 3 Bergenia ciliata ekstrak kasar Bhandari et al. 2008 4 Piptadenia africana piptadienol Mbouangouere et al.
2008 5 Curcuma longa curcumin Du et al. 2006 6 Pinus densiflora ekstrak etanol 70% Kim et al. 2003
Cendawan 1 Ganoderma lucidum SKG-3, senyawa belum
diidentifikasi Kim et al. 2004
2 Penicillium sp Cyclo (dehydroala-L-Leu) Kwoon et al. 2000
Alga 1 Ecklonia stolonifera phlorotannin Moon et al. 2011 2 Spirogyra varians pentagalloyl glucosa Cannell et al. 1988
Aktinomiset 1 Micromonospora ekstrak etil asetat Suthindhiran et al.
2009
2 Streptomyces clavurigus
ceftezole, suatu antibiotik beta laktam
Lee et al. 2007
3 Actinoplanes sp. acarbose Hemker et al. 2001 4 Actinomadura
verrucospora pradimicin Q, C24H16O10 Yosuke et al. 1992
Bakteri non aktinomiset 1 Bacillus subtilis Ekstrak kasar Zhu et al. 2008
Tanaman Obat Diabetes
Bnouham et al. (2006) melaporkan bahwa setidaknya ada 176 spesies
tanaman yang berasal dari 84 famili yang telah dikaji di berbagai negara dan
menunjukkan potensi yang tinggi di dalam pengobatan penyakit diabetes.
Beberapa tanaman yang sangat potensial tersebut antara lain adalah famili:
Leguminoseae (11 spesies), Lamiaceae (7 spesies), Liliaceae (8 spesies),
Cucurbitaceae (7 spesies), Asteraceae (6 spesies), Moraceae (6 spesies),
Rosaceae (6 spesies), Euphorbiaceae (5 spesies) and Araliaceae (5 spesies).
Spesies yang paling banyak dikaji adalah: Citrullus colocynthis, Opuntia
streptacantha Lem. (Cactaceae), Trigonella foenum greacum L. (Leguminosae),
Momordica charantia L. (Cucurbitaceae), Ficus bengalensis L. (Moraceae),
Polygala senega L. (Polygalaceae) dan Gymnema sylvestre R. (Asclepiadaceae).

14
Tanaman Lagerstroemia speciosa (Lythraceae), yang di kawasan Asia
Tenggara dikenal dengan nama banaba, secara tradisional telah dikonsumsi
masyarakat dalam berbagai bentuk untuk pengobatan diabetes dan penyakit
ginjal. Pada tahun 1990-an popularitas tanaman ini telah menarik perhatian para
ilmuwan di berbagai negara. Sejak saat itu, konsistensi aktivitas anti diabetes
dari tanaman ini diuji secara in vitro maupun in vivo (Klein et al. 2007).
DM merupakan penyakit yang umum terjadi di berbagai negara, sehingga
upaya pengobatan penyakit DM mulai tradisional hingga modern banyak
dilakukan. Sejumlah penelitian tanaman obat yang memiliki khasiat sebagai
antidiabetes juga telah dilakukan oleh ilmuwan di berbagai negara (Tabel 2).
Tabel 2 Kajian tanaman obat diabetes di berbagai negara
No Tanaman Negara Informasi bahan aktif Referensi
1 Ceiba pentandra Afrika Selatan
ekstrak C. pentandra pada dosis rendah 40 mg/kg bb menurunkan glukosa darah 40.0% dan 48.9%, pada tikus normal dan tikus diabet
Djoemeni et al. 2006
2 Helichrysum odoratissimum, Helichrysum nudifolium, Artemisia afra, Vernonia oligocephala
Afrika Selatan
secara tradisional dikonsumsi sebagai sayur atau lalap
Mahop & Mayet 2007
3 Coccinia cordifolia, Catharanthus roseus
Bangla-desh
ekstrak metanol daun tanaman C. cordifolia dan C. roseus memiliki efek antihiperglikemik pada tikus diabet yang diinduksi dengan aloksan
Akhtar et al. 2007
4 Cissus sicyoides
Brazilia senyawa aktif yang terkandung pada tanaman ini juga memiliki aktivitas antimikrob
Beltrame et al. 2002
5 Panax ginseng
Cina senyawa ginsenosida hasil ekstraksi secara signifikan sebagai antihiperglikemik pada mencit yang kegemukan
Attele et al. 2002
6 Andrographis paniculata
Malaysia pemberian secara oral ekstrak etanol Andrographis paniculata berperan dalam meningkatkan sensitivitas insulin dan penundaan perkembangan resistensi insulin
Subramanian et al. 2008
7 Murraya konigii, Ocimum tenuiflorum
India ekstrak kloroform Ocimum tenuiflorum dan Murraya konigii menghambat α- glukosidase
Bhat et al. 2008
8 Syzygium cumini
India diperoleh senyawa mycaminose yang memiliki efek antidiabet pada tikus
Kumar et al. 2008
9 Acacia catechu, India berbagai tanaman tersebut Jeyacand

15
Alpinia calcarata, Budea monosperma
secara tradisional digunakan dalam ayurveda sebagai obat antidiabet
ran & Mahes 2007
10 Melia dubia India ekstrak alkhohol M. dubia efektif sebagai agen hipoglikemik
Susheela et al. 2008
11 Phyllanthus fraternus
India ekstrak etanol P. fraternus memiliki efek antidiabet dan antioksidan dibanding obat standar tolbutamide.
Garg et al. 2008
12 Tribullus terrestris India ekstrak T. terrestris dapat menurunkan gula darah mencit hiperglikemia
Lamda et al. 2011
13 Terminalia sp.
Indonesia ektrak metanol daun terminalia mampu menghambat α-glukosidase, nilai IC50 5µg/ml
Anam et al. 2009
14 Shorea balanocarpoides
Indonesia ekstrak kayu raru dari jenis S. balanocarpoides memiliki inhibisi terhadap α- glukosidase berkisar 88-97%.
Pasaribu, 2009
15 Andrographis paniculata
Indonesia ekstrak etanol herba sambiloto mempunyai efek menurunkan glukosa darah pada uji toleransi glukosa
Yulinah et al. 2001
16 Phalleria macrocarpa
Indonesia ekstrak buah mahkota dewa memiliki aktivitas inhibitor α- glukosidase yang memiliki efek antihiperglikemia pada tikus putih
Sugiwati et al. 2006
17 Tanaman famili Apocynaceae,Clusiaceae, Euphorbiaceae, Rubiaceae
Indonesia berbagai ekstrak memiliki aktivitas lebih tinggi dari acarbose
Elya et al. 2012
18 Verbascum cermanensis, Rosa damascene, Rosmarinus officinnalis
Iran ekstrak metanol dan air beberapa tanaman tersebut menghambat di atas 50% terhadap α- glukosidase
Gholamhoseinian et al. 2008
19 Viscum album Inggris terdapat senyawa aktif dalam tanaman Viscum album yang berperan meningkatkan sekresi insulin, sebagai senyawa antidiabetes
Gray & Flat 1999
20 Chrysanthemum coronarium, Dioscorea batatas, Morus alba, Citrus unshiu
Korea pemberian ekstrak C. coronarium dan M. alba memiliki efek hipoglikemik pada tikus diabet setara dengan glibenclamide. Pemberian ekstrak C.unshiu menunjukkan efek antihiperlipidemik dibanding antidiabetik.
Kim et al. 2006
21 Guaiacum coulteri, Psacalium peltatum dan Psidium guajava
Mexico P. peltatum dan G. coulteri memiliki efek antihiperglikemik pada kelinci diabet dan kelinci sehat.
Aguilar et al. 2003
22 Azadirachta indica Nigeria ekstrak tanaman tersebut baik Ebong et

16
dan Vernonia amygdalina
secara kombinasi atau secara sendiri dapat mereduksi kadar gula darah yang setara dengan clorpropamid.
al. 2008
23 Lagerstroemia speciosa
Philipina digunakan sebagai obat diabetes di Philipina, mengandung tannic acid dan penta galoil glucose (PGG)
Klein et al. 2007
24 Flemingia sp Taiwan ekstrak air Flemingia sp memiliki IC50 253 µg/ml terhadap α- glukosidase
Hsieh et al. 2010
Di Indonesia, juga dikenal berbagai tanaman yang dikenal memiliki kasiat
sebagai obat diabetes seperti sambiloto, brotowali, ciplukan, daun dewa, pare,
dan mahkota dewa. Tanaman tersebut secara empiris telah digunakan oleh
masyarakat Indonesia untuk pengobatan diabetes. Beberapa kajian ilmiah juga
telah dilakukan untuk mengetahui kandungan bahan aktif tanaman ataupun
mekanisme kerja bahan aktif tanaman dalam menurunkan kadar gula darah.
Yulinah et al. (2001) melaporkan bahwa ekstrak etanol tanaman sambiloto
mampu menurunkan kadar gula darah tikus putih yang menderita hiperglikemia.
Ekstrak buah mahkota dewa juga dilaporkan memiliki kemampuan sebagai anti
hiperglikemia melalui aktivitas inhibitor α-glukosidase (Sugiwati et al. 2006).
Pasaribu (2009) melaporkan bahwa kayu raru (Shorea sp) memiliki aktivitas
inhibitor α- glukosidase. Di samping, itu masih banyak jenis-jenis tanaman dii
Indonesia dari famili Apocynaceae, Clusiaceae, Euphorbiaceae, dan Rubiaceae
yang memiliki aktvitas inhibitor α-glukosidase (Elya et al. 2011).
Tanaman brotowali (T. crispa)
Klasifikasi tanaman brotowali adalah sebagai berikut:
Kingdom: Plantae
Subkingdom: Tracheobionta
Super Divisi: Spermatophyta
Divisi: Magnoliophyta
Kelas: Magnoliopsida
Sub Kelas: Magnoliidae
Ordo: Ranunculales
Famili: Menispermaceae
Genus: Tinospora
Spesies: Tinospora crispa

17
Tanaman brotowali merupakan salah satu tanaman obat yang terkenal
dari Asia Tenggara. Tanaman ini sudah berabad-abad dikenal sebagai salah satu
tanaman obat yang sangat manjur. Penyebaran brotowali di Asia meliputi:
wilayah Cina, Semenanjung Melayu, Filipina, India dan Indonesia. Brotowali
mulai tersebar ke seluruh penjuru benua setelah terjadi hubungan dagang antar
benua. Hal tersebut di karenakan manfaatnya yang sangat besar sebagai bahan
baku obat-obatan. Brotowali merupakan tanaman obat tradisional Indonesia yang
biasa ditanam di pekarangan atau tumbuh liar di hutan. Rebusan batangnya yang
terasa sangat pahit biasa dijadikan obat rematik, menurunkan kadar gula darah
dan menurunkan panas. Di Indonesia, selain dikenal dengan nama bratawali,
tanaman ini juga dikenal dengan nama daerah andawali, antawali, putrawali atau
daun gadel.
Tinospora crispa, termasuk suku Menispermaceae, klas Magnoliopsida
(Dicotyledoneae), divisi Magnoliophyta (Spermatophyta). Ciri-ciri penting dari
tumbuhan ini: liana, membelit dengan batang dan ranting, batang sukulen dan
berbenjol-benjol; daun tunggal, tanpa stipula, tulang daun menjari, fitotaksis
tersebar; bunga uniseksual, trimeros, aksiler atau cauliflorous; buah batu; tipe
daun dorsiventral, stomata anomositik; berkas pembuluh kolateral terbuka; pada
bagian korteks batang terdapat lengkungan sklerenkim; kandungan kimianya
terdiri atas: amilum, pikroretin, pikroretosida, alkaloida, saponin, tanin. (Santa et
al. 1998)
Brotowali atau Tinospora crispa (L.) Miers ex Hoff. memiliki sinonim T.
cordifolia (Thunb.) Miers, T. rumphii Boerl, T. tuberculata, Cocculus crispum,
Menispermum crispum, M. tuberculatum, M. verrucosum. Di setiap daerah dan
negara juga memiliki berbagai macam nama yang berbeda-beda. Walaupun
persebarannya sudah ke seluruh benua tetapi belum di temui keragaman tingkat
subgenus yang mencolok. Hal tersebut disebabkan karena tanaman ini termasuk
tanaman yang mudah bertahan hidup dalam kondisi ekstrim.
Hasil penelitian menunjukkan bahwa brotowali mengandung berbagai
macam zat kimia yang sangat membantu sekali dalam proses penyembuhan.
Brotowali telah lama di kenal oleh masyarakat tradisional Indonesia sebagai
bahan pembuatan jamu yang di campur dengan tanaman-tanaman herbal
lainnya. Brotowali merupakan tanaman herba (perdu) yang hampir semua
bagian dari tubuhnya di manfaatkan sebagai obat serba guna yang dapat

18
menyembuhkan berbagai macam penyakit seperti: diabetes melitus, hipertensi,
dan demam. Kajian ilmiah terhadap tanaman brotowali sebagai obat diabetes
telah dilakukan para peneliti. Noor dan Ashcroft (1997) melaporkan bahwa
ekstrak T. crispa mampu menstimulasi peningkatan sekresi insulin, sehingga
dapat berperan sebagai antihiperglikemia. Noipa dan Ninlaaesong (2011)
melaporkan bahwa ekstrak T. crispa dapat meningkatkan laju transpor glukosa
ke dalam sel. Chougale et al. (2009) melaporkan bahwa ekstrak tanaman T.
cordifolia memiliki aktivitas inhibitor α-glukosidase. Patela dan Mishrab (2012)
menemukan tiga senyawa dari tanaman Tinospora cordifolia yang memiliki
kemampuan inhibitor α-glukosidase, yaitu jatrorrhizine, palmatine dan
magnoflorine. Senyawa-senyawa tersebut memiliki nilai IC50 berturut-turut
sebesar 22.05, 38.42 and 7.6 µg/mL untuk jatrorrhizine, palmatine dan
magnoflorine. Hasil uji in vivo juga menunjukkan adanya penekanan yang
signifikan terhadap kenaikan kadar glukosa plasma darah oleh ketiga senyawa
tersebut pada konsentrasi 20 mg/kg berat badan. Senyawa magnoflorine
merupakan senyawa paling potensial sebagai inhibitor α-glukosidase diantara
ketiga senyawa yang ditemukan tersebut. Uji klinis oleh Sriyapai et al. (2009)
menunjukkan bahwa konsumsi serbuk T. crispa dapat menurunkan kadar gula
darah setelah makan.
Mikrob Endofit
Mikrob endofit adalah mikrob yang hidup di dalam jaringan tanaman pada
periode tertentu dan mampu hidup dengan membentuk koloni dalam jaringan
tanaman tanpa membahayakan inangnya. Setiap tanaman tingkat tinggi dapat
mengandung beberapa mikrob endofit yang mampu menghasilkan senyawa
biologi atau metabolit sekunder yang diduga sebagai akibat koevolusi atau
transfer genetik (genetic recombination) dari tanaman inangnya ke dalam mikrob
endofit (Tan & Zou 2001).
Kemampuan mikrob endofit memproduksi senyawa metabolit sekunder
sesuai dengan tanaman inangnya merupakan peluang yang sangat besar dan
dapat diandalkan untuk memproduksi metabolit sekunder dari mikrob endofit
yang diisolasi dari tanaman inangnya tersebut. Sekitar 300000 jenis tanaman
yang tersebar di muka bumi ini, masing-masing tanaman mengandung satu atau
lebih mikrob endofit yang terdiri dari bakteri dan cendawan (Strobel & Daisy
2003). Apabila endofit yang diisolasi dari suatu tanaman obat dapat

19
menghasilkan metabolit sekunder sama dengan tanaman inangnya atau bahkan
dalam jumlah yang lebih tinggi, maka tidak perlu mengambil tanaman aslinya
untuk diambil sebagai simplisia.
Berbagai jenis endofit telah berhasil diisolasi dari tanaman inangnya, dan
telah berhasil dibiakkan dalam media sintetik yang sesuai. Demikian pula
metabolit sekunder yang diproduksi oleh mikrob endofit tersebut telah berhasil
diisolasi dan dimurnikan serta telah dielusidasi struktur molekulnya. Berbagai
produk mikrob endofit yang telah dilaporkan adalah sebagai antibiotik, antivirus,
antikanker, antioksidan, antidiabetes, imunosupresif dan lain-lain (Strobel &
Daisy 2003).
Endofit yang telah dilaporkan memiliki aktivitas antidiabet adalah
Pseudomassaria sp. Endofit yang diisolasi dari hutan dekat Kinshasa, Republik
Congo ini mampu menghasilkan senyawa non peptida (L-783,281). Senyawa ini
memiliki aktivitas yang menyerupai insulin, tetapi tidak seperti insulin karena tidak
terdegradasi di dalam sistem pencernaan, sehingga dapat digunakan secara oral.
Pemberian secara oral senyawa L-783,281 terhadap hewan model mencit
diabetes menunjukkan adanya penurunan kadar gula darah secara signifikan.
Hasil ini memberi kemungkinan terapi baru terhadap diabetes (Strobel & Daisy
2003).
.
Aktinomiset Endofit dan Potensinya
Aktinomiset telah lama diketahui sebagai sumber berbagai senyawa
bioaktif yang telah dimanfaatkan dalam berbagai bidang seperti kesehatan,
pertanian, maupun industri. Beberapa hasil penelitian di bawah ini merupakan
contoh potensi aktinomiset endofit yang mampu menghasilkan senyawa bioaktif
yang berguna dalam bidang medis.
Streptomyces sp. strain NRRL 30562 merupakan isolat endofit dari
tanaman obat snakevine (Kennedia nigriscans) yang mampu menghasilkan
antibiotik berspektrum luas yang disebut munumbicin. Senyawa ini dapat
menghambat pertumbuhan Bacillus anthracis, dan Mycobacterium tuberculosis
yang multi resisten terhadap berbagai obat anti tuberkulosis, sehingga senyawa
ini berpotensi digunakan untuk pengobatan terhadap penyakit tuberkulosis
(Castillo et al. 2002).
Jenis endofit lainnya yang juga menghasilkan antibiotik berspektrum luas
adalah aktinomiset endofit yang diisolasi dari tanaman Grevillea pteridifolia.

20
Endofit ini menghasilkan metabolit kakadumycin. Aktivitas anti bakterinya sama
seperti munumbicin (Castillo et al. 2003). Streptomyces sp. (MSU-2110) yang
merupakan isolat endofit pada tanaman Monstrea sp. mampu menghasilkan
antibiotik coronamycin yang dapat menghambat cendawan Cryptococcus
neoformans yang bersifat patogen pada manusia (Ezra et al. 2004).
Paclitaxel dan derivatnya merupakan zat yang berkhasiat sebagai anti
kanker yang pertama kali ditemukan yang diproduksi oleh mikrob endofit.
Paclitaxel merupakan senyawa diterpenoid yang terdapat dalam tanaman Taxus.
Senyawa yang dapat mempengaruhi molekul tubulin dalam proses pembelahan
sel-sel kanker ini, dapat diproduksi oleh endofit Pestalotiopsis microspora yang
terdapat pada tanaman Taxus andreanae, T. brevifolia, dan T. wallichiana
(Strobel 2001). Beberapa aktinomiset endofit lainnya seperti Kitasatospora dan
Micromonospora telah dapat diisolasi dari tanaman Taxus yang mampu
menghasilkan berbagai senyawa taxane yang memiliki aktivitas sebagai
antitumor (Caruso et al. 2000).
Streptomyces aureofaciens CMUAc130 yang diisolasi dari jaringan akar
tanaman jahe (Zingiber officinale) dapat menghasilkan senyawa 4-arylcoumarin
yang memiliki aktivitas anti tumor. Pemberian senyawa ini secara intra peritonial
dapat menghambat perkembangan sel Lewis Lung Carcinoma (LLC). Senyawa
4-arylcoumarin menunjukkan aktivitas anti tumor dengan nilai T/C 80.8 dan
50,0% pada dosis 1 dan 10 mg/kg berat badan (dalam bentuk 5,7-dimetoxy-4-
pherylcoumarin) dan 81.5 dan 44.9% pada dosis 1 dan 10 mg/kg (dalam bentuk
5,7-dimetoxy-4-phenylcoumarin). Kontrol positif yang berupa senyawa
adriamycin menunjukkan nilai T/C 55.9% pada dosis 2 mg/kg berat badan
(Taechowisan et al. 2007). Senyawa pterocidin yang dihasilkan oleh
Streptomyces hygroscopicus merupakan senyawa sitotoksik terhadap beberapa
human cancer cell lines sehingga berpotensi digunakan untuk pengobatan
kanker (Igarashi et al. 2006).
Munumbicin C dan D yang dihasilkan dari Streptomyces endofit sangat
berpotensi digunakan sebagai obat anti malaria. Hasil pengujian terhadap
Plasmodium falciparum menunjukkan bahwa nilai IC50 dari munumbicin tersebut
masing-masing 6.5 dan 4.5 ng/ml, lebih rendah dari IC50 cloroquin yang
merupakan gold standar obat anti malaria yaitu 7.0 ng/ml (Castillo et al. 2002).
Streptomyces sp. (MSU-2110) yang merupakan isolat endofit pada tanaman
Monstrea sp. mampu menghasilkan antibiotik peptida yang disebut coronamicin.

21
Senyawa ini juga memiliki kemampuan dalam menghambat Plasmodium
falciparum dengan IC50 sebesar 9 ng/ml sehingga berpotensi juga digunakan
sebagai obat antimalaria (Ezra et al. 2004). Ringkasan kajian aktinomiset endofit
pada beberapa tanaman dan potensinya dapat dilihat pada Tabel 3.
Tabel 3 Kajian aktinomiset endofit pada beberapa tanaman dan potensinya
No Aktinomiset endofit
Tanaman inang Potensi Pustaka
1 Microbispora Streptomyces Micromonospora
Brassica campestris
biokontrol terhadap Plasmodiospora brassicae
Sun et al. 2008
2 Pseudonocardia oroxyli sp.
Oroxylum indicum
belum dilaporkan Gu et al. 2006
3 Streptomyces Microbispora Micromonospora Nocardiades
Gandum biokontrol patogen tanaman
Coombs & Franco 2003
4 Streptomyces sp Zingiber officinale Alpinia galanga
biokontrol terhadap Candida albicans dan Fusarium oxysporum
Taecowisan & Lumyong 2003
5 Streptomyces sp Aegiceras comiculatum
menghasilkan cyclopentenone
Lin et al. 2005
6 Streptomyces hygroscopicus
- senyawa sitotoksik (pterocidin)
Igarashi et al. 2006
7 Streptomyces sp Monstrea menghasilkan coronamicin (antifungi, antimalaria)
Ezra et al. 2004
8 Streptomyces aureofaciens
Zingiber officinale
menghasilkan coumarin (antitumor)
Taecowisan et al. 2007
9 Kitasatospora Micromonospora Streptomyces
Taxus sp menghasilkan taxane (antikanker)
Caruso et al. 2000
10 Streptomyces sp Kennedia nigriscans
menghasilkan munumbicin (antifungi, antimalaria, anti TBC
Castillo et al. 2002
Studi awal telah dilakukan Irawan (2008) dengan melakukan isolasi
aktinomiset endofit dari tanaman ciplukan, temulawak dan brotowali. Lima belas
isolat aktinomiset yang telah diperoleh, dua diantaranya mampu menghasilkan
inhibitor α-glukosidase, yaitu isolat Tc-2.1 (dari brotowali) dan Cx-10.1 (dari temu
lawak). Hasil pengujian awal menunjukkan ekstrak kasar isolat Cx-10.1 memiliki
daya hambat lebih besar dibandingkan dengan Glucobay 0.1% terhadap enzim
α-glukosidase.
Tabel 3 di atas menunjukkan bahwa sejumlah aktinomiset endofit dapat
diisolasi dari berbagai jenis tanaman. Isolat-isolat aktinomiset endofit tersebut

22
juga dilaporkan mampu menghasilkan senyawa bioaktif dalam medium sintetik di
laboratorium. Kenyataan ini mengindikasikan adanya peluang yang besar untuk
mendapatkan isolat-isolat baru aktinomiset endofit dalam rangka pencarian
senyawa bioaktif baru, khususnya senyawa inhibitor α-glukosidase.

23
METODE PENELITIAN
Tempat dan Waktu Penelitian
Penelitian ini dilakukan di Laboratorium Mikrobiologi Departemen Biologi
dan di Laboratorium Kimia Analitik Departemen Kimia, FMIPA IPB dari bulan
April 2009 sampai dengan Maret 2012.
Isolasi Aktinomiset Endofit
Sampel tanaman brotowali (Tinospora crispa) diperoleh dari Kebun
Koleksi Tanaman Obat, Pusat Studi Biofarmaka, Institut Pertanian Bogor, Bogor.
Sampel tanaman berupa bagian akar, batang dan daun tanaman brotowali.
Disamping brotowali, beberapa tanaman obat yang telah diketahui memiliki
kasiat sebagai obat diabetes juga dilakukan isolasi aktinomiset endofitnya, yaitu:
lidah buaya (Alloe vera), mahkota dewa (Phaleria macrocarpa), temu ireng
(Curcuma aeruginosa), pegagan (Centela asiatica), tempuyung (Xoncus
arvensis), sambiloto (Andrographis paniculata), secang (Caesalpinia sappan),
temu lawak (Curcuma xanthorriza), pete (Parcia speciosa) sambung nyawa
(Gynura procumbens), ciplukan (Physalis peruviana) dan rosela (Hibiscus
sabdariffa).
Bagian tanaman yang diisolasi dicuci dengan air lalu dilakukan sterilisasi
permukaan mengikuti metode seperti yang diterangkan Coombs dan Franco
(2003). Sampel tanaman direndam dalam alkhohol 70% selama 1 menit, larutan
hipoklorit 1% selama 5 menit, lalu alkhohol 70% selama satu menit, dan terakhir
dibilas dengan akuades steril. Sampel tanaman yang telah steril selanjutnya
dihaluskan secara aseptik dengan mortar dan ditambahkan 4 ml 12.5 mM buffer
fosfat. Sebanyak 100 µl suspensi sampel diinokulasikan pada medium Humic
Acid Vitamin B (HV) agar dengan penambahan 50 ppm cycloheximide dan 30
ppm nalidixic acid. Inkubasi dilakukan selama 14-21 hari pada suhu ruang.
Untuk konfirmasi keberhasilan proses sterilisasi permukaan, bilasan terakhir
akuades juga dilakukan pencawanan pada medium HV agar. Koloni-koloni
aktinomiset yang tumbuh diisolasi dan dimurnikan lebih lanjut menggunakan
medium Yeast Malt Extract Agar (YMA). Koleksi isolat disimpan dalam medium
YMA miring pada suhu 4 0C dan larutan gliserol 15% pada suhu -20 0C.

24
Seleksi dan Uji Aktinomiset Endofit Penghasil Inhibitor α-glukosidase
Semua isolat yang diperoleh ditumbuhkan ke dalam medium cair berisi
0.1% soluble starch, 0.5% pepton, dan 0.1% yeast extract (pH 7) selama 14 hari
dengan kecepatan agitasi 120 rpm pada temperatur ruang. Biomassa sel
dipisahkan dengan sentrifugasi dengan kecepatan 1432 x g selama 20 menit dan
supernatan yang diperoleh diuji aktivitas inhibitor α-glukosidasenya. Aktivitas
inhibitor α-glukosidase diuji menurut Anam et al. (2009). Uji penghambatan
enzim dilakukan berdasarkan pada pemecahan substrat untuk menghasilkan
produk berwarna, yang diukur absorbansinya selama periode waktu tetentu.
Enzim α-glukosidase (Sigma, St. Louis) dilarutkan dalam 0.1 M buffer fosfat pH 7
dengan konsentrasi 0.25 unit/ml. Substrat yang digunakan adalah p-Nitrophenyl
α-D-glucopyranoside (Sigma, St. Louis) 20 mM yang dilarutkan dalam 0.1 M
buffer fosfat pH 7. Campuran reaksi terdiri dari 125 l substrat, 240 l 0.1 M
buffer fosfat pH 7 dan 10 l sampel. campuran reaksi diinkubasi pada suhu 370C
selama 5 menit, ditambahkan 125 l larutan enzim dan diinkubasi selama 15
menit pada suhu 37 0C. Reaksi dihentikan dengan penambahan 500 l larutan
Na2CO3 200 mM, dan p-nitrophenol yang dihasilkan diukur absorbansinya pada
panjang gelombang 400 nm. Sebagai pembanding digunakan larutan acarbose 1
mg/ml. Penghambatan aktivitas enzim α-glukosidase ditentukan dengan rumus:
Penghambatan (%) = (Ak-(As1-As0))/Ak x 100%
(Ak: absorbansi kontrol, AS0: absorbansi sampel tanpa enzim, As1: absorbansi sampel)
Peran Aktinomiset Endofit dalam Menghasilkan Inhibitor α-glukosidase
Untuk mengetahui peran isolat aktinomiset endofit dalam menghasilkan
inhibitor α-glukosidase, digunakan tanaman T. crispa bebas endofit yang hasil
kultur jaringan tanaman yang diperoleh dari Lab. Konservasi Sumberdaya Hutan,
Fakultas Kehutanan IPB. Sebanyak 0.5 gram sampel tanaman hasil kultur
jaringan digerus secara aseptik dan ditambah 0.5 ml buffer fosfat, kemudian
dipisahkan antara biomassa dan supernatan dengan sentrifugasi pada kecepatan
1432 x g. Supernatan yang diperoleh diuji aktivitas inhibitor α-glukosidasenya.
Dengan cara yang sama dilakukan pula uji aktivitas inhibitor α-glukosidase
terhadap tanaman T. crispa yang diperoleh dari alam. Kemampuan aktivitas

25
inhibitor α-glukosidase kemudian dibandingkan dengan aktivitas inhibitor dari
kultur aktinomiset endofit terpilih.
Penentuan Waktu Produksi Optimum
Kurva produksi digunakan untuk mengetahui waktu optimum yang
digunakan untuk memproduksi inhibitor α-glukosidase dalam jumlah besar. Isolat
terpilih ditumbuhkan dalam medium produksi menurut Chen et al. (2004) yang
berisi 0.1% soluble starch, 0.5% pepton dan 0.15 yeast extract (pH 7) steril.
Sebanyak 1% starter diinokulasikan ke dalam media produksi dan diinkubasi
pada suhu ruang dengan kecepatan agitasi 150 putaran per menit dengan rotary
shaker. Setiap 5 hari dilakukan panen kultur dengan cara sentrifugasi pada
kecepatan 1432 x g selama 10 menit pada suhu 40C. Supernatan yang diperoleh
diuji aktivitas penghambatannya terhadap enzim α-glukosidase. Pengukuran
berat kering pelet dilakukan untuk mengetahui pertumbuhan isolat. Percobaan
diakhiri jika produksi inhibitor α-glukosidase telah mengalami penurunan.
Identifikasi Isolat Terpilih
Identifikasi isolat terpilih dilakukan berdasarkan sekuen 16S rDNA.
Ekstraksi DNA isolat aktinomiset BWA65 dilakukan menggunakan Genomic DNA
Mini Kit (Geneaid) dilanjutkan dengan amplifikasi gen 16S rRNA menggunakan
Primer 20F (5'GATTTTGATCCTGGCTCAG3') dan 1500R (5'GTTACCTT-
GTTACGACTT3'). Komposisi reaksi PCR seperti tertera pada Tabel 4.
Tabel 4 Komposisi reaksi PCR untuk amplifikasi gen 16S rRNA
Komponen Volume (µl)
dH2O 19.5 Buffer PCR 4 MgCl2 5 dNTP 1.5 Primer 20-F (10 pM) 1 Primer 1500-R (10 pM) 1 DMSO 0.5 DNA template 7 Taq DNA polimerase 0.5
Reaksi amplifikasi dilakukan dengan menggunakan thermal cycler (model
480, Perkin-Elmer, USA), dengan siklus sebagai berikut: 5 menit denaturasi awal
pada 94 0C, 45 detik denaturasi pada 94 oC, dan 45 detik annealing primer pada

26
57 oC, 1 menit ekstensi pada 72 oC dan 7 menit ekstensi final pada 72 oC,
sebanyak 30 siklus. Hasil amplifikasi kemudian dilarikan menggunakan gel
agarosa 1% pada 70 Volt selama 45 menit. Hasil elektroforesis selanjutnya
diwarnai menggunakan etidium bromida selama 15 menit dan divisualisasi
menggunakan lampu UV transluminator. Pita DNA yang muncul didokumentasi
menggunakan Gel Doc.
Hasil PCR disekuen menggunakan jasa sekuensing MacroGen, Korea.
Analisis sekuen DNA dilakukan menggunakan program BioEdit dan dilakukan
analisis BLAST pada Bank Gen NCBI dataLibrary. Analisis filogenetik
menggunakan program multiple aligment Clustal X dan konstruksi pohon
filogenetik menggunakan program NJ plot.
Karakterisasi Morfologi Isolat
Karakterisasi morfologi isolat dilakukan dengan mengamati pertumbuhan
isolat pada media Yeast Malt Extract Agar (YMA), Yeast Extract Agar (YSA) dan
Oatmeal Agar (OA). Pengamatan makroskopis meliputi pertumbuhan koloni,
warna miselium aerial, warna miselium substrat dan keberadaan pigmen terlarut.
Pengamatan mikroskopis isolat dilakukan menggunakan mikroskop cahaya pada
perbesaran 100 dan 400x, serta Scanning Electron Microscope (SEM)(JSM-
5310LV) pada perbesaran 10000x.
Deteksi Gen Sedoheptulosa-7-Fosfat Siklase
Isolasi DNA Genom. DNA genom aktinomiset diisolasi menggunakan
Genomic DNA Mini Kit (Geneaid). Tahapan kerjanya adalah sebagai berikut.
Kultur sel umur 3 hari sebanyak 1.5 µl ditransfer ke dalam tabung mikro, lalu
disentrifus 3354 xg selama 3 menit. Supernantan dibuang dan pelet sel yang
diperoleh diresuspensi dengan 200 µl buffer lisozim. Suspensi tersebut
diinkubasi 10 menit pada suhu ruang, tiap 2-3 menit dibolak-balik. Ke dalam
suspensi kemudian ditambahkan 200 µl buffer GB dan diinkubasi 70 0C selama
10 menit, dan dibolak-balik tiap 3 menit hingga suspensi terlihat jernih. Tahap
berikutnya adalah penambahan 200 µl etanol 96% ke dalam lisat dan diaduk
dengan pipet perlahan-lahan.
Hasil pelisisan sel selanjutnya ditransfer ke dalam kolom GD yang telah
ditempatkan ke dalam tabung koleksi. Tabung disentrifus 8586 xg selama 2
menit, dan tabung koleksi dipisahkan. Kolom GD ditempatkan pada tabung

27
koleksi yang baru lalu ditambahkan 400 µl buffer W1, lalu disentrifus 8586 xg
selama 30 detik. Kolom GD diambil dan ditempatkan pada tabung koleksi yang
baru. Ke dalam kolom GD ditambah 600 µl wash buffer dan disentrifus 8586 x g
selama 30 detik. Kolom GD ditempatkan kembali pada tabung koleksi dan
disentrifus lagi selama 3 menit untuk mengeringkan matriks pada kolom GD.
Kolom GD selanjutnya dipindahkan ke dalam tabung mikro dan ditambah 100 µl
buffer elusi yang telah dipanaskan 70 0C. Tabung dibiarkan 3-5 menit hingga
buffer elusi meresap ke dalam matriks, lalu disentrifugasi 8586 x g dan larutan
DNA yang diperoleh disimpan di suhu -20 0C.
Amplifikasi Gen Penyandi Sedoheptulosa-7-Fosfat Siklase.
Amplifikasi gen penyandi Sedoheptulosa-7-fosfat siklase menggunakan primer
yang didesain oleh Hyun et al. (2005) sebagai berikut: primer VOG-F
5'GGSGGSGG-SGTSCTSATGGACGTSGCSGG-3' (GGGVLMDVAG), dan
primer VOG-R 5'GCCATGTCSACGCASACSGCSGCCTCSCCGAG-3'
(HGEAVCVDMA). Amplifikasi dengan PCR dilakukan menurut Hyun et al. (2005)
dengan modifikasi. Sebagai kontrol positip dilakukan pula amplifikasi fragmen
DNA dari Actinoplanes sp. SE50/100. Komposisi reaksi campuran PCR yang
digunakan dalam percobaan ini seperti tertera pada Tabel 5.
Tabel 5 Komposisi PCR untuk amplifikasi gen sedoheptulosa-7-fosfat siklase
Komponen Volume (µl)
dH2O 12.9 Buffer PCR 5 MgCl2 1 dNTP 0.5 Primer VOG-F (10 pM) 2 Primer VOG-R (10 pM) 2 DMSO 0.5 DNA template 2 Taq DNA polimerase 0.1
Reaksi amplifikasi dilakukan dengan menggunakan thermal cycler (model
480, Perkin-Elmer, USA), dengan siklus sebagai berikut: 5 menit denaturasi awal
pada 95 0C, 20 detik denaturasi pada 98 oC, dan 1 menit annealing primer pada
67 oC, 45 detik ektensi pada 72 oC dan 7 menit ekstensi final pada 72 oC,
sebanyak 30 siklus. Hasil amplifikasi kemudian dilarikan menggunakan gel
agarosa 1% pada 70 V selama 45 menit. Hasil elektroforesis selanjutnya
diwarnai menggunakan etidium bromida selama 15 menit dan divisualisasi

28
menggunakan lampu UV transluminator. Pita DNA yang muncul didokumentasi
menggunakan Gel Doc.
Sekuensing dan Analisis Molekuler. Hasil PCR disekuen menggunakan
jasa sekuensing 1ST BASE Singapura. Hasil sekuensing kemudian dibandingkan
dengan sekuen yang diambil dari database di GenBank, menggunakan program
BLASTX pada situs NCBI.
Isolasi dan Identifikasi Senyawa Inhibitor α-Glukosidase
Persiapan Kultur dan Media Fermentasi. Isolat BWA65 ditumbuhkan
pada media YSA selama 7 hari selanjutnya diinokulasikan ke dalam medium cair
berisi 0.1% soluble starch, 0.5% pepton, dan 0.1% yeast extract (pH 7). Kultur
diinkubasi selama 7 hari dengan kecepatan agitasi 120 putaran per menit pada
temperatur ruang. Kultur ini selanjutnya digunakan sebagai starter.
Produksi Senyawa Inhibitor α-Glukosidase. Untuk keperluan
karakterisasi senyawa inhibitor α-glukosidase diperlukan kultur produksi dalam
jumlah besar (5 liter). Pada penelitian ini produksi skala 5 liter dilakukan dengan
menggunakan galon steril bekas kemasan air minum ukuran 19 liter, dengan
aerasi yang dilewatkan mikro filter 0.2 µm. Produksi inhibitor α-glukosidase
dilakukan dengan menginokulasikan sebanyak 150 ml (berat kering biomassa sel
0.1 gram) kultur aktinomiset umur 7 hari ke dalam 5 liter media produksi, dan
dilakukan inkubasi selama 15 hari dengan aerasi. Pemanenan kultur dilakukan
dengan menyaring kultur untuk memisahkan supernatan dan biomassa.
Ekstraksi Senyawa Inhibitor α-Glukosidase. Supernatan yang
diperoleh dari kultur isolat BWA65 diekstraksi dengan menggunakan berbagai
pelarut untuk mendapatkan senyawa aktif. Pelarut yang dicoba digunakan adalah:
kloroform, etanol, metanol, butanol, dan etil asetat. Ekstraksi dilakukan dengan
cara menambahkan pelarut ke dalam supernatan dengan perbandingan 1:1,
selanjutnya dihomogenkan dengan menggunakan magnetic stirer selama 2 jam
dan dibiarkan selama 2 jam hingga membentuk fraksi air dan fraksi pelarut.
Fraksi pelarut kemudian dipisahkan dan dilakukan pemekatan dengan rotary
evaporator hingga diperoleh fraksi pekat. Fraksi pekat yang diperoleh kemudian
dikeringkan, dihitung bobotnya dan siap digunakan untuk tahap selanjutnya.

29
Fraksinasi Senyawa Inhibitor α-Glukosidase dengan Kromatografi
Kolom. Sebelum kromatografi kolom dilakukan, terlebih dahulu dilakukan
pencarian eluen terbaik dengan bantuan Kromatografi Lapis Tipis (KLT).
Tahapan ini dilakukan untuk mencari pelarut terbaik yang dapat memisahkan
komponen aktif dari ekstrak etil asetat. Tahapan ini dilakukan dengan cara coba-
coba dengan menggunakan pelarut tunggal atau gabungan dari beberapa pelarut.
Pelarut yang dicobakan antara lain: etanol, metanol, etil asetat, heksan, aseton
dan klorofom. Hasil dari tahapan ini digunakan untuk fraksinasi senyawa aktif
menggunakan kromatografi kolom.
Fraksinasi dengan kromatografi kolom dilakukan dengan menggunakan
kolom silika gel dengan ukuran panjang 38 cm dan diameter 1.5 cm. Preparasi
kolom diawali dengan memasukkan 20 gram silika gel (Merck) ditambah dengan
eluen yang akan digunakan yaitu campuran heksan : etil asetat 1:4, sambil
diketuk-ketuk berulang kali hingga terbentuk kolom yang stabil. Setelah kolom
siap digunakan, sebanyak 5 ml ekstrak etil asetat diinjeksikan ke dalam kolom
dan dielusi dengan heksan : etil asetat 1:4 secara isokratik dengan kecepatan
elusi 1 ml/menit. Fraksi-fraksi sebanyak ± 5 ml ditampung pada tabung reaksi.
Tahap selanjutnya adalah menggabungkan fraksi-fraksi yang memiliki
kromatogram yang sama dengan bantuan KLT. Fraksi-fraksi yang diperoleh
selanjutnya diuji aktivitasnya dalam menghambat α-glukosidase.
Pengujian Aktivitas Inhibitor α-Glukosidase Fraksi. Aktivitas inhibitor
α-glukosidase diuji menurut Moon et al. (2011) dengan modifikasi. Uji
penghambatan enzim dilakukan berdasarkan pada pemecahan substrat untuk
menghasilkan produk berwarna, yang diukur absorbansinya selama periode
waktu tetentu. Enzim α-glukosidase (Sigma) dilarutkan dalam 0.1 M buffer fosfat
pH 7 dengan konsentrasi 0.2 unit/ml. Sebagai substrat digunakan p-Nitrophenyl
α-D-glucopyranoside (Sigma) 2.5 mM yang dilarutkan dalam 0.1 M buffer fosfat
pH 7. Campuran reaksi terdiri dari 50 l substrat, 50 l 0.1 M buffer fosfat pH 7
dan 50 l sampel. Setelah campuran reaksi diinkubasi pada suhu 37 0C selama 5
menit, sebanyak 50 l larutan enzim ditambahkan dan selanjutnya diinkubasi
selama 15 menit pada suhu 37 0C. Reaksi dihentikan dengan penambahan 800
l larutan Na2CO3 200 mM. Senyawa p-nitrofenol yang dihasilkan dari reaksi ini
diukur absorbansinya pada panjang gelombang 405 nm. Sebagai pembanding
digunakan larutan acarbose.

30
Penentuan Nilai IC50. Nilai IC50 merupakan nilai yang menunjukkan
konsentrasi ekstrak yang menyebabkan penghambatan sebesar 50% terhadap
aktivitas enzim α-glukosidase. Penentuan nilai ini dilakukan dengan cara menguji
aktifitas penghambatan ekstrak pada berbagai macam konsentrasi. Setelah
didapat nilai penghambatan dari masing-masing konsentrasi ekstrak, selanjutnya
dibuat persamaan garis yang merupakan fungsi dari konsentrasi ekstrak dan
besaran penghambatan yang dihasilkan. Nilai-nilai konsentrasi ekstrak sebagai
variabel X dan besarnya penghambatan sebagai variabel Y.
Pengaruh Konsentrasi Substrat. Pengaruh konsentrasi substrat
digunakan untuk menduga tipe inhibitor dari senyawa Fraksi F6. Penentuan tipe
inhibitor dilakukan dengan cara menguji aktivitas inhibitor pada berbagai macam
konsentrasi substrat yaitu: 20, 10, 5, 2.5, dan 1.25 mM. Data yang diperoleh
kemudian dikonversi dan diinterpretasikan ke dalam persamaan Lineweaver-Burk
dalam bentuk grafik. Konsentrasi substrat diubah menjadi 1/[S] pada sumbu X,
dan kecepatan reaksi pembentukan produk hasil hidrólisis enzim diubah menjadi
1/V pada sumbu Y. Tahap selanjutnya menentukan persamaan garis yang
terbentuk dan tipe hambatannya berdasarkan perpotongan garis dengan sumbu
X dan sumbu Y antara kinetika enzim tanpa inhibitor dan kinetika enzim setelah
mendapat perlakuan inhibitor. Nilai Vmak dan KM ditentukan dengan melihat
perpotongan garis terhadap sumbu X dan sumbu Y dari persamaan Lineweaver-
Burke yang diperoleh. Perpotongan dengan sumbu X merupakan nilai -1/KM dan
perpotongan dengan sumbu Y merupakan nilai 1/Vmak (Copeland 2005; Wu et al
2012).
Identifikasi dengan Spektroskopi UV-Vis. Identifikasi senyawa
menggunakan spektrofotometer UV dilakukan dengan cara mengukur pola
spektrum serapan senyawa dalam larutan yang sangat encer dengan
pembanding blanko pelarut, menggunakan spektrofotometer UV-Vis
(Pharmaspec UV-1700 Shimadzu) yang dapat merekam secara otomatis. Pelarut
yang digunakan dalam pengukuran adalah etanol. Senyawa dalam sampel diukur
pada panjang gelombang 200-900 nm. Identifikasi senyawa dilakukan dengan
membandingkan pola spektrum yang dihasilkan dengan pola spektrum referensi.
Identifikasi dengan FTIR. Contoh dalam bentuk serbuk sebanyak ± 2 mg
dihaluskan bersamaan dengan 0.198 gram KBr dalam mortal agate. Contoh yang
telah dihaluskan dan bercampur dengan KBr dimasukkan ke dalam alat pencetak

31
pelat KBr kemudian ditekan sehingga diperoleh lempeng serbuk yang transparan.
Contoh lempeng tadi kemudian dimasukkan ke dalam spektrofotometer FTIR.
Spektrum yang diperoleh berupa kurva transmitan dengan bilangan gelombang.
Identifikasi gugus fungsi berdasarkan pada nilai bilangan gelombang gugus yang
sudah diketahui (Pavia 2001)

32

33
HASIL DAN PEMBAHASAN
Isolasi Aktinomiset Endofit
Pada penelitian ini sebanyak 32 isolat aktinomiset endofit telah berhasil
diisolasi dari tanaman T. crispa dan 33 isolat dari tanaman lainnya menggunakan
media agar HV. Semua isolat aktinomiset endofit yang diperoleh dalam penelitian
ini merupakan isolat yang dapat dikulturkan dan tidak mewakili semua populasi
mikrob endofit yang hidup di tanaman-tanaman obat tersebut. Hal ini disebabkan
sebagian besar mikrob (lebih dari 99%) adalah unculturable atau tidak dapat
ditumbuhkan pada medium sintetik (Sharma et al. 2005).
Sekitar 69,2% dari tanaman yang diuji mengandung aktinomiset endofit
Tananam T. crispa merupakan tanaman obat dengan jumlah isolat aktinomiset
tertinggi (32 isolat). Sejumlah isolat diperoleh dari tanaman lainnya, yaitu:
Curcuma aeruginosa (9 isolat), Gynura procumbens (6 isolat), Curcuma
xanthorryza (5 isolat) dan tanaman lainnya (1-4 isolat) (Gambar 3).
Gambar 3 Jumlah isolat aktinomiset endofit yang berhasil diisolasi dari masing-masing tanaman obat.
Hasil isolasi dari keseluruhan tanaman menunjukkan bahwa aktinomiset
endofit paling banyak diperoleh dari bagian akar tanaman (45 isolat), diikuti
bagian rimpang (14 isolat), lalu batang dan daun (masing-masing 3 isolat).
Hampir 70% aktinomiset endofit diperoleh dari bagian akar (Gambar 4). Hal ini
dapat dipahami karena secara umum aktinomiset merupakan bakteri tanah,
6 5
1
32
13 3 4
9
00
5
10
15
20
25
30
35
Jum
lah
iso
lat
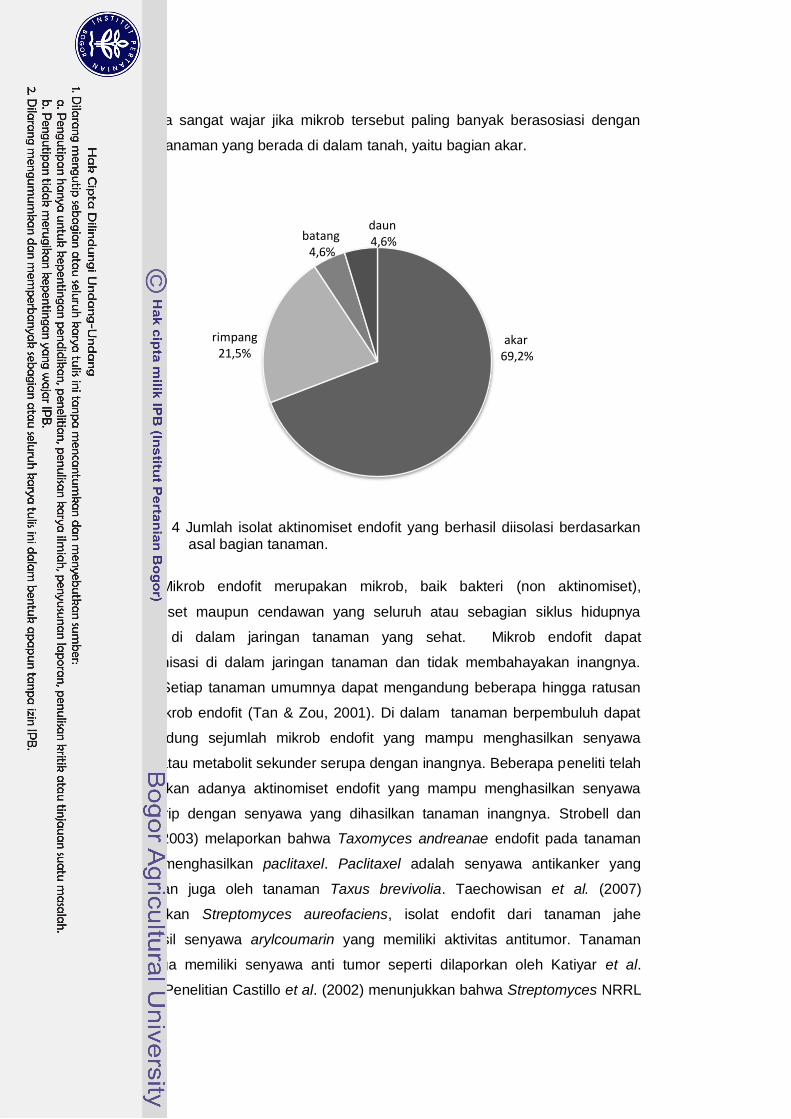
34
sehingga sangat wajar jika mikrob tersebut paling banyak berasosiasi dengan
bagian tanaman yang berada di dalam tanah, yaitu bagian akar.
Gambar 4 Jumlah isolat aktinomiset endofit yang berhasil diisolasi berdasarkan asal bagian tanaman.
Mikrob endofit merupakan mikrob, baik bakteri (non aktinomiset),
aktinomiset maupun cendawan yang seluruh atau sebagian siklus hidupnya
berada di dalam jaringan tanaman yang sehat. Mikrob endofit dapat
berkolonisasi di dalam jaringan tanaman dan tidak membahayakan inangnya.
Setiap tanaman umumnya dapat mengandung beberapa hingga ratusan
jenis mikrob endofit (Tan & Zou, 2001). Di dalam tanaman berpembuluh dapat
mengandung sejumlah mikrob endofit yang mampu menghasilkan senyawa
biologi atau metabolit sekunder serupa dengan inangnya. Beberapa peneliti telah
melaporkan adanya aktinomiset endofit yang mampu menghasilkan senyawa
aktif mirip dengan senyawa yang dihasilkan tanaman inangnya. Strobell dan
Daisy (2003) melaporkan bahwa Taxomyces andreanae endofit pada tanaman
Taxus menghasilkan paclitaxel. Paclitaxel adalah senyawa antikanker yang
dihasilkan juga oleh tanaman Taxus brevivolia. Taechowisan et al. (2007)
melaporkan Streptomyces aureofaciens, isolat endofit dari tanaman jahe
penghasil senyawa arylcoumarin yang memiliki aktivitas antitumor. Tanaman
jahe juga memiliki senyawa anti tumor seperti dilaporkan oleh Katiyar et al.
(1996). Penelitian Castillo et al. (2002) menunjukkan bahwa Streptomyces NRRL
akar69,2%
rimpang 21,5%
batang4,6%
daun4,6%
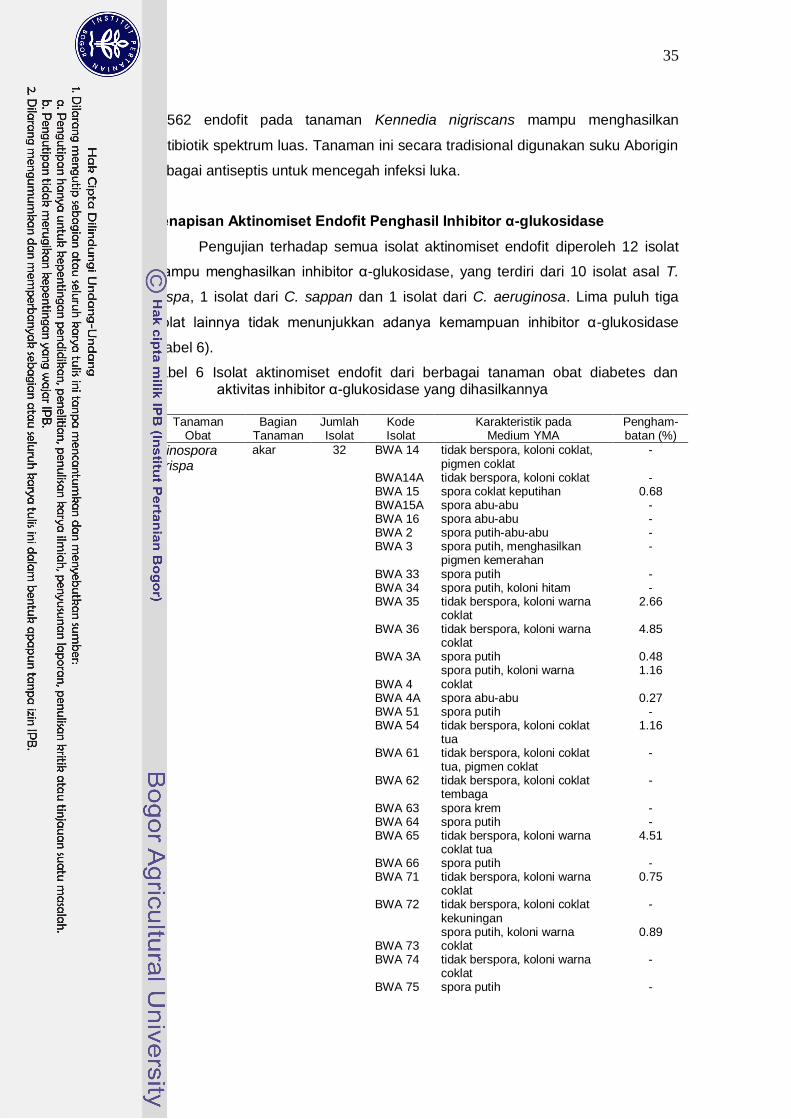
35
30562 endofit pada tanaman Kennedia nigriscans mampu menghasilkan
antibiotik spektrum luas. Tanaman ini secara tradisional digunakan suku Aborigin
sebagai antiseptis untuk mencegah infeksi luka.
Penapisan Aktinomiset Endofit Penghasil Inhibitor α-glukosidase
Pengujian terhadap semua isolat aktinomiset endofit diperoleh 12 isolat
mampu menghasilkan inhibitor α-glukosidase, yang terdiri dari 10 isolat asal T.
crispa, 1 isolat dari C. sappan dan 1 isolat dari C. aeruginosa. Lima puluh tiga
isolat lainnya tidak menunjukkan adanya kemampuan inhibitor α-glukosidase
(Tabel 6).
Tabel 6 Isolat aktinomiset endofit dari berbagai tanaman obat diabetes dan aktivitas inhibitor α-glukosidase yang dihasilkannya
Tanaman
Obat Bagian
Tanaman Jumlah Isolat
Kode Isolat
Karakteristik pada Medium YMA
Pengham- batan (%)
Tinospora crispa
akar 32 BWA 14 tidak berspora, koloni coklat, pigmen coklat
-
BWA14A tidak berspora, koloni coklat - BWA 15 spora coklat keputihan 0.68 BWA15A spora abu-abu - BWA 16 spora abu-abu - BWA 2 spora putih-abu-abu - BWA 3 spora putih, menghasilkan
pigmen kemerahan -
BWA 33 spora putih - BWA 34 spora putih, koloni hitam - BWA 35 tidak berspora, koloni warna
coklat 2.66
BWA 36 tidak berspora, koloni warna coklat
4.85
BWA 3A spora putih 0.48
BWA 4 spora putih, koloni warna coklat
1.16
BWA 4A spora abu-abu 0.27 BWA 51 spora putih - BWA 54 tidak berspora, koloni coklat
tua 1.16
BWA 61 tidak berspora, koloni coklat tua, pigmen coklat
-
BWA 62 tidak berspora, koloni coklat tembaga
-
BWA 63 spora krem - BWA 64 spora putih - BWA 65 tidak berspora, koloni warna
coklat tua 4.51
BWA 66 spora putih - BWA 71 tidak berspora, koloni warna
coklat 0.75
BWA 72 tidak berspora, koloni coklat kekuningan
-
BWA 73 spora putih, koloni warna coklat
0.89
BWA 74 tidak berspora, koloni warna coklat
-
BWA 75 spora putih -
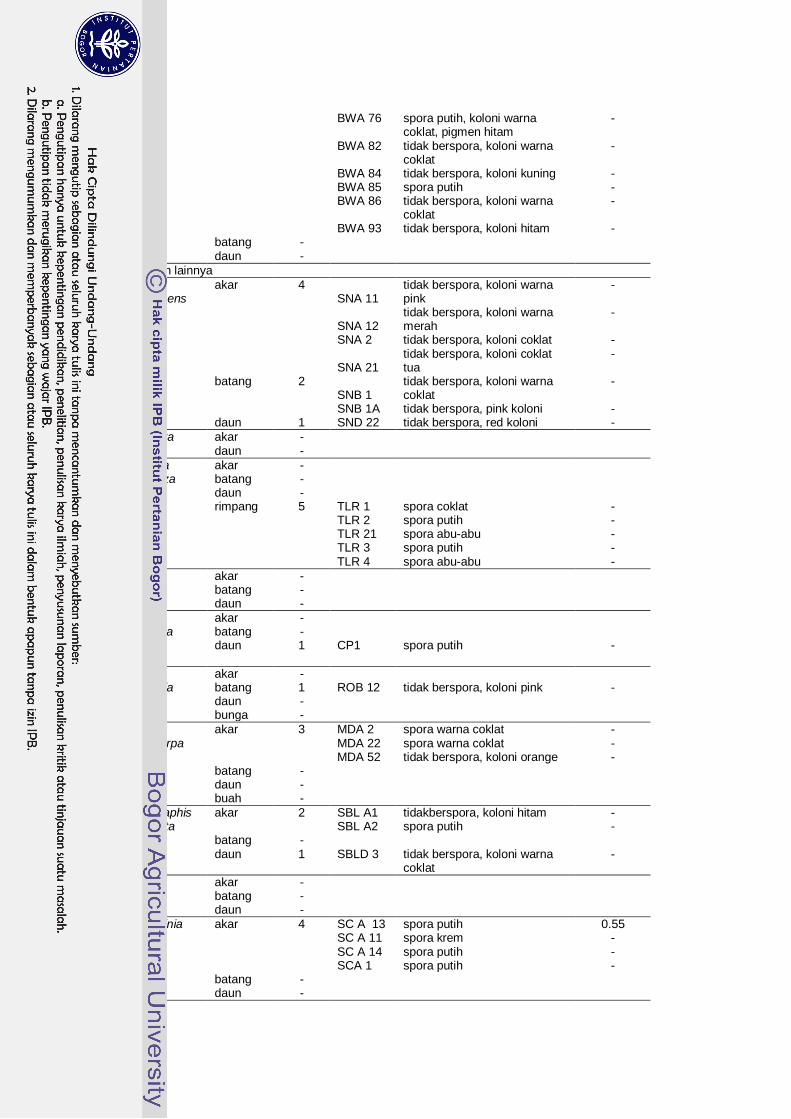
36
BWA 76 spora putih, koloni warna coklat, pigmen hitam
-
BWA 82 tidak berspora, koloni warna coklat
-
BWA 84 tidak berspora, koloni kuning - BWA 85 spora putih - BWA 86 tidak berspora, koloni warna
coklat -
BWA 93 tidak berspora, koloni hitam - batang - daun -
Tanaman lainnya
Gynura procumbens
akar 4 SNA 11
tidak berspora, koloni warna pink
-
SNA 12 tidak berspora, koloni warna merah
-
SNA 2 tidak berspora, koloni coklat -
SNA 21 tidak berspora, koloni coklat tua
-
batang 2 SNB 1
tidak berspora, koloni warna coklat
-
SNB 1A tidak berspora, pink koloni - daun 1 SND 22 tidak berspora, red koloni -
Alloe vera
akar - daun -
Curcuma xanthoriza
akar - batang - daun - rimpang 5 TLR 1 spora coklat -
TLR 2 spora putih - TLR 21 spora abu-abu - TLR 3 spora putih - TLR 4 spora abu-abu -
Centela asiatica
akar - batang - daun -
Physalis peruviana
akar - batang - daun 1 CP1 spora putih -
Hibiscus sabdariffa
akar - batang 1 ROB 12 tidak berspora, koloni pink - daun - bunga -
Phaleria macrocarpa
akar 3 MDA 2 spora warna coklat - MDA 22 spora warna coklat - MDA 52 tidak berspora, koloni orange -
batang - daun - buah -
Andrographis paniculata
akar 2 SBL A1 tidakberspora, koloni hitam - SBL A2 spora putih -
batang - daun 1 SBLD 3 tidak berspora, koloni warna
coklat -
Xoncus arvensis
akar - batang - daun -
Caesalpinia sappan
akar 4 SC A 13 spora putih 0.55 SC A 11 spora krem - SC A 14 spora putih - SCA 1 spora putih -
batang - daun -

37
Parcia speciosa
akar - batang - daun -
Curcuma aeruginosa
akar - batang - daun - rimpang 9
TIR 11 spora putih, koloni warna coklat
-
TIR 12 spora krem - TIR 13 spora keemasan, koloni warna
coklat 3.62
TIR 14 tidak berspora -
TIR 1A spora putih, koloni warna coklat
-
TIR 1B spora putih -
TIR 1B2 spora putih, koloni warna coklat
-
TIR 2 spora abu-abu, koloni coklat tua
-
TIR 3 spora putih abu-abu - Jumlah isolat 65
Pada pengujian lebih lanjut secara kuantitatif menggunakan p-nitrophenyl
α-D-glucopyranoside sebagai substrat menunjukkan bahwa ekstrak kasar
(supernatan) dari empat isolat aktinomiset endofit (BWA36, BWA65, BWA35,
BWA54) dari T. crispa memiliki aktivitas α-glukosidase tertinggi. Ekstrak kasar
BWA65 isolat dari T. crispa menghasilkan penghambatan tertinggi (11.01%)
terhadap α-glukosidase, yaitu sebesar 80% jika dibandingkan dengan
penghambatan 1mg/ml acarbose (13.61%) yang digunakan sebagai kontrol
(Gambar 5).
Penemuan aktinomiset endofit pada tanaman brotowali belum pernah
dilaporkan oleh para peneliti sebelumnya, sehingga dapat dikatakan adanya
aktinomiset endofit pada tanaman brotowali pada penelitian ini merupakan hal
yang baru. Laporan ini juga yang pertama kali menyebutkan adanya aktinomiset
endofit penghasil senyawa inhibitor α-glukosidase. Laporan-laporan sebelumnya
yang menyebutkan sejumlah aktinomiset penghasil senyawa inhibitor α-
glukosidase adalah bukan merupakan isolat endofit yang berasal dari jaringan
tanaman.

38
Gambar 5 Aktivitas inhibitor α-glukosidase oleh supernatan dari kultur aktinomiset endofit asal T. crispa.
Penemuan isolat aktinomiset endofit dari tanaman T. crispa yang
menghasilkan inhibitor α-glukosidase dalam penelitian ini memiliki arti penting
dalam memperkuat pendapat Tan dan Zou (2001) yang menyatakan bahwa
tanaman dapat mengandung mikrob endofit yang dapat menghasilkan senyawa
biologis atau metabolit sekunder yang sama dengan inangnya. Informasi tentang
keberadaan agen hipoglikemik pada tanaman Tinospora telah dilaporkan oleh
beberapa peneliti sebelumnya ( Rajalakshmi et al. 2009; Chougale et al. 2009).
Mereka melaporkan bahwa ekstrak tanaman T. cordifolia memiliki aktivitas
inhibitor α-glukosidase. Hasil penelitiannya menunjukkan bahwa pemberian
ekstrak etanol dan air tanaman T. cordifolia pada dosis 400 mg/kg dapat
menurunkan gula darah pada tikus dan kelinci hiperglikemia yang diinduksi
dengan aloksan.
Pada penelitian ini juga dibandingkan aktivitas inhibitor α-glukosidase
yang dihasilkan oleh tanaman inang T. crispa, tanaman T. crispa bebas endofit
hasil kultur jaringan tanaman dan isolat aktinomiset endofit. Hasil pengujian ini
diharapkan dapat menggambarkan peran aktinomiset endofit dalam memberikan
kontribusi terhadap produksi senyawa inhibitor. Hasil penelitian menunjukkan
bahwa tanaman bebas endofit yang diperoleh dari kultur jaringan tanaman umur
3 bulan hanya memiliki kemampuan yang sangat rendah untuk menghasilkan
senyawa inhibitor. Tanaman T. crispa yang diperoleh dari alam mampu
memproduksi senyawa inhibitor jauh lebih besar (Gambar 6). Namun,
6,35
11,01
5,646,34
13,61
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
16,00
BWA 36 BWA65 BWA 35 BWA 54 Ac
Inh
ibis
i (%
)
Isolat aktinomiset

39
kemampuan inhibitor α-glukosidase yang dihasilkan oleh aktinomiset endofit
BWA65 lebih dari dua kali lipat dibanding aktivitas inhibitor α-glukosidase yang
dihasilkan tanaman inang. Data tersebut mengindikasikan bahwa aktinomiset
endofit dalam tanaman T. crispa tersebut memberikan kontribusi yang besar
terhadap produksi senyawa inhibitor α-glukosidase.
Gambar 6 Aktivitas inhibitor α-glukosidase yang dihasilkan oleh tanaman hasil
kultur jaringan tanaman (KJT, kalus), tanaman dari alam (batang, akar, daun) dan isolat aktinomiset endofit BWA65.
Selain tanaman hasil kultur jaringan, diuji pula aktivitas inhibitor α-
glukosidase dari kalus. Kalus merupakan sekumpulan hasil proliferasi sel
tanaman yang membentuk biomassa tetapi tidak dapat berdiferensiasi
membentuk organ tanaman seperti akar, batang dan daun. Kultur kalus biasanya
dilakukan dengan tujuan untuk menghasilkan senyawa bioaktif dari suatu
tanaman. Pada gambar di atas terlihat bahwa kalus tanaman brotowali memiliki
aktivitas inhibitor α-glukosidase yang relatif besar jika dibanding dengan tanaman
hasil kultur jaringan. Hal ini dapat disebabkan karena pada kultur kalus,
biosintesis senyawa metabolit sekunder lebih besar jika dibanding dengan
metabolisme sekunder pada tanaman hasil kultur jaringan. Dengan demikian
untuk keperluan produksi metabolit sekunder, seringkali digunakan kultur kalus.
Beberapa peneliti memilih menggunakan kultur kalus untuk memproduksi
senyawa bioaktif. Purwianingsih dan Hamdiyati (2009). menggunakan kultur
kalus Morinda citrifolia L. (Mengkudu) yang dielisitasi menggunakan
Sacharomyces cerevisiae untuk memproduksi senyawa bioaktif kuinon. Sutini et
0,06
3,05
1,64
3,39
4,52
10,98
0,00
2,00
4,00
6,00
8,00
10,00
12,00
KJT Kalus Batang Akar Daun BWA65
Pe
ngh
amb
atan
(%)
Sampel

40
al. (2008) juga menggunakan kultur kalus Camellia sinensis untuk memproduksi
senyawa bioaktif flavonoid, flavan-3-ol suatu metabolit sekunder yang terdapat
dalam daun teh muda yang yang memiliki khasiat sebagai antiobesitas dan
antioksidan. Adanya aktivitas inhibitor α-glukosidase pada kultur kalus tanaman
Tinospora belum pernah dilaporkan sebelumnya. Kultur kalus tanaman Tinospora
cordifolia dilaporkan memiliki aktifitas antimikrob terhadap Eschericia coli dan
Staphylococcus aureus (Khalilsaraie et al. 2011).
Karakterisasi Morfologi dan Identifikasi Isolat BWA65
Dalam penelitian ini digunakan tiga jenis media untuk melihat karakteristik
kultur isolat BWA65 yaitu: Yeast Extract Malt Extract Agar (YMA), Yeast Extract
Soluble Starch Agar (YSA) dan Oatmeal Agar (OA). Hasil penelitian
menunjukkan bahwa isolat BWA65 memiliki pertumbuhan yang baik pada media
YSA dan OA, dan pertumbuhan moderat pada media YMA. Pada semua media,
isolat BWA65 menghasilkan miselium aerial berwarna putih. Miselium substrat
isolat BWA65 berwarna coklat apabila ditumbuhkan pada media OA dan coklat
tua apabila ditumbuhkan pada media YSA maupun YMA. Isolat BWA65 ini juga
memproduksi pigmen terlarut (soluble pigment) berwarna coklat tua pada media
YSA dan merah muda pada media OA (Tabel 7).
Tabel 7 Karakteristik kultur isolat BWA65 pada berbagai media (7 hari inkubasi
pada suhu ruang)
Karakteristik Kultur Media
YMA YSA OA
pertumbuhan moderat baik baik
miselium aerial putih putih putih
miselium substrat coklat tua coklat tua coklat
pigmen terlarut tidak ada coklat merah muda
Pengamatan morfologi isolat BWA65 di bawah mikroskop cahaya
dengan perbesaran 400x menunjukkan adanya rantai spora berbentuk spiral.
Susunan rantai spora berbentuk spiral ini merupakan karakter yang khas untuk
Streptomyces. Pengujian lebih lanjut dengan menggunakan mikroskop elektron
SEM menunjukkan bahwa isolat BWA65 memiliki hifa udara yang tidak

41
bercabang (unbranched aerial hyphae) serta rantai spora spiral dengan spora
berbentuk silinder dan permukaannya halus (Gambar 7).
Gambar 7 Morfologi isolat aktinomiset endofit BWA65 yang ditumbuhkan pada media Oatmeal Agar umur 7 hari (A), dilihat dengan mikroskop cahaya (400x) (B) dan SEM (10,000x) (C), tanda panah menunjukkan rantai spora.
Identifikasi Isolat Aktinomiset
Hasil identifikasi molekuler menggunakan sekuen parsial 16S rDNA
menunjukkan bahwa BWA65 memiliki kesamaan paling tinggi dengan
Streptomyces olivochromogenes dengan tingkat kemiripan 92%. Berdasarkan
studi sebelumnya, S. olivochromogenes dikenal untuk menghasilkan senyawa
aktif seperti: glukosa isomerase, xilosa isomerase dan fosfolipase (Azin et
al.1997; Simkada et al. 2009). Sampai sekarang, belum ada laporan tentang
inhibitor α-glukosidase yang dihasilkan oleh S. olivochromogenes. Identifikasi
ulang isolat BWA65 dengan sekuen gen 16S rRNA yang lebih lengkap dilakukan
untuk meyakinkan hasil identifikasi ini. Hasil identifikasi menggunakan sekuen
16S rDNA yang lebih lengkap sepanjang sekitar 1343 pasang basa menunjukkan
bahwa BWA65 memiliki kesamaan 98% dengan Streptomyces diastaticus.
Berdasarkan studi sebelumnya, S. diastaticus dikenal dapat menghasilkan
senyawa aktif seperti: L-arabinofuranosidase (Tajana et al. 1992) dan antibiotik
makrolida poliene (Seco et al. 2005). Hingga saat ini juga belum ada laporan
tentang inhibitor α-glukosidase yang dihasilkan oleh S. diastaticus. Berdasarkan
hal tersebut, kemungkinan besar isolat BWA65 merupakan jenis baru penghasil
inhibitor α-glukosidase. Pohon filogenetik isolat aktinomiset endofit BWA65
berdasarkan sekuen 16S rDNA dapat dilihat pada Gambar 8.
C A B

42
Pohon filogenetik tersebut terlihat jelas bahwa isolat BWA65 memiliki
kekerabatan paling dekat dengan S. diastaticus. Isolat BWA65 berada satu grup
dengan beberapa Streptomyces lainnya seperti S. coeliflavus, S.
olivochromogenes dan S. abikoensis. S. coeliflavus dikenal mampu menghaslkan
senyawa inhibitor alfa amilase dan S. abikoensis dilaporkan dapat menghasilkan
senyawa inhibitor α-glukosidase. Pada gambar pohon filogenetik tersebut juga
dapat dilihat dua jenis aktinomiset non Streptomyces yaitu Actinoplanes dan
Saccarothrix espanaensis. Kedua jenis anggota aktinomiset non Streptomyces ini
dikenal juga dapat menghasilkan senyawa inhibitor α-glukosidase (Hyun et al.
2005). Sebagai outer group pada gambar pohon filogenetik tersebut terdapat
spesies Bacillus subtilis yang merupakan wakil dari bakteri Gram positif non
aktinomiset.
Gambar 8 Pohon filogenetik isolat BWA65 berdasarkan sekuen 16S rDNA.

43
Deteksi Gen Sedoheptulosa- 7- Fosfat Siklase
Ketersediaan gen penyandi biosintesis senyawa inhibitor α-glukosidase
akan memudahkan untuk mengembangkan primer dan probe yang dapat
digunakan untuk mendeteksi adanya organisme penghasil senyawa inhibitor α-
glukosidase. Hyun et al. (2005) telah berhasil membuat primer untuk gen
penyandi inhibitor α-glukosidase pada aktinomiset berdasarkan sekuen gen yang
telah diketahui. Oleh karena itu, deteksi gen penyandi senyawa inhibitor α-
glukosidase pada aktinomiset akan menjadi lebih mudah. Primer yang spesifik
dapat membantu mempercepat pencarian galur-galur aktinomiset penghasil
senyawa inhibitor α-glukosidase. Pendekatan yang dapat dilakukan untuk
mendeteksi adanya gen penyandi inhibitor α-glukosidase pada aktinomiset
BWA65 adalah dengan menggunakan primer spesifik yang telah dikenal tersebut.
Dengan cara ini akan dapat diketahui apakah isolat aktinomiset BWA65
mempunyai gen penyandi biosintesis senyawa inhibitor α-glukosidase yang mirip
dengan acarbose, validamisin dan lainnya yang jalur biosintesisnya melibatkan
senyawa sedoheptulosa-7-fosfat. Dengan cara ini juga dapat ditemukan dengan
cepat aktinomiset yang mempunyai gen penyandi senyawa inhibitor α-
glukosidase tersebut atau tidak. Jika deteksi gen tidak menunjukkan hasil positif
maka senyawa inhibitor α-glukosidase tersebut kemungkinan merupakan
senyawa baru dan belum diketahui jalur biosintesisnya.
Hasil amplifikasi gen penyandi sedoheptulosa-7-fosfat siklase
menggunakan primer VOG-R dan VOG-R setelah divisualisasi menggunakan gel
agarosa 1% seperti tertera pada Gambar 9. Fragmen DNA target berukuran
sekitar 540 pasang basa tampak terlihat jelas pada hasil amplifikasi
menggunakan PCR yang divisualisasi pada gel agarosa. Fragmen DNA
berukuran 540 pasang basa ini kemudian dipurifikasi untuk kemudian disekuen
dan hasilnya dianalisis menggunakan BLASTX pada situs NCBI. Hasil analisis
BLASTX dari hasil sekuen terhadap urutan asam amino yang terdapat pada bank
data NCBI menunjukkan kemiripan paling besar terhadap protein histidin kinase
pada Streptomyces violaceusniger (No akses YP-004812094.1) (Tabel 8) dan
bukan sebagai protein sedoheptulosa-7-fosfat siklase.

44
Gambar 9 Hasil amplifikasi PCR menggunakan primer VOG-F dan VOG-R (lajur 1,2 kontrol positif; 3,4,5 isolat BWA 65; M marker; 6,7 kontrol negatif.
Histidin kinase sendiri merupakan protein kelas transferase yang
biasanya berupa protein transmembran yang berperan dalam transduksi sinyal
melintasi membran selular. Sebagian besar histidin kinase merupakan
homodimer yang memiliki kemampuan autokinase, fosfotransfer dan aktivitas
fosfatase. Molekul histidin kinase biasanya memiliki bagian reseptor di bagian
luar sel (domain ekstraseluler), bagian yang melintasi membran sel (domain
transmembran), dan bagian dalam sel (domain intraseluler) yang mengandung
aktivitas enzimatik. Selain aktivitas kinase, domain-domain intraseluler biasanya
memiliki wilayah yang mengikat ke molekul efektor sekunder atau kompleks
molekul yang lebih lanjut menyebarkan transduksi sinyal dalam sel. Histidin
kinase memainkan peran utama dalam transduksi sinyal di prokariot untuk
adaptasi selular terhadap kondisi lingkungan dan cekaman (Dutta et al. 1999)
Analisis gen penyandi sedoheptulosa-7-fosfat siklase pada penelitian ini
belum sesuai dengan hasil yang diharapkan karena fragmen DNA yang didapat
menggunakan primer VOG-F dan VOG-R memiliki kemiripan yang lebih besar
terhadap gen-gen bukan penyandi sedoheptulosa-7-fosfat siklase. Primer VOG-F
dan VOG-R pada awalnya dipilih karena primer ini telah digunakan oleh peneliti
sebelumnya untuk mendeteksi adanya gen penyandi sedoheptulosa-7-fosfat
siklase pada aktinomiset (Hyun et al. 2005). Ketidaksesuaian hasil ini

45
kemungkinan besar disebabkan karena primer tersebut tidak menempel secara
spesifik hanya pada gen sedoheptulosa-7-fosfat siklase, tetapi juga dapat
menempel pada gen lain yang ujungnya memiliki urutan basa yang komplemen
dengan sebagian urutan basa pada primer VOG-F dan VOG-R, misalnya gen
histidin kinase tersebut. Ketidak spesifikan ini berhasil dibuktikan oleh Velina
(2012, unpublished) yang berhasil mendesain ulang primer untuk
mengamplifikasi gen sedoheptulosa-7-fosfat siklase pada isolat BWA65 ini.
Primer dengan urutan basa Forward: 5’-ACCTACGAGGTGCGCTTCCGG-
GACGACGT-3’ dan Reverse : 5’-GGCGGCCTGCAGCTCGGCGGCCGTCA-
CGT-3’. Hasil amplifikasi menggunakan primer tersebut diperoleh amplikon
berukuran sekitar 300 pasang basa. Fragmen DNA yang diperoleh kemudian
dikloning ke plasmid pMD20 dan kemudian disekuen. Hasil analis sekuen
fragmen DNA hasil amplifikasi tersebut menunjukkan kemiripan 100% dengan
sedoheptulosa-7-fosfat siklase pada Actinoplanes SE50/100. Terdeteksinya gen
penyandi sedoheptulosa-7-fosfat siklase pada isolat BWA65 ini memberikan
informasi terdapatnya jalur biosintesis senyawa inhibitor α-glukosidase yang
mirip dengan jalur biosintesis acarbose pada isolat BWA65.
Tabel 8 Hasil analisis sekuen DNA hasil amplifikasi gen penyandi sedoheptulosa-
7-fosfat siklase isolat BWA65 menggunakan program BLASTX
No akses Homologi
Tingkat kemiripan
YP-004812094.1 GAF sensor hybrid histidine kinase pada Streptomyces violaceusniger Tu 4113
92%
ZP_07294440.1 Sensor histidine kinase/respone regulator pada Streptomyces himastatinicus ATCC 53653)
89%
AD106478.1 Two-component system sensor kinase pada Streptomyces bingchenggensis BCW-1
83%
Ekstraksi Senyawa Inhibitor α-Glukosidase
Pelarut terbaik yang digunakan untuk ekstraksi senyawa inhibitor dari
kultur BWA65 diperoleh dengan cara melakukan percobaan pendahuluan
dengan menggunakan berbagai macam pelarut yaitu; kloroform, etil asetat,
metanol, etanol dan butanol. Hasil ekstraksi dengan berbagai macam pelarut
tersebut disajikan pada Tabel 9. Hasil percobaan menunjukkan penggunaan etil

46
asetat memberikan hasil terbaik diantara pelarut lainnya dengan rendemen
ekstrak 0.03%, diikuti dengan pelarut etanol, metanol, butanol dan kloroform . Etil
asetat selanjutnya digunakan untuk keperluan ekstraksi senyawa inhibitor α-
glukosidase dari kultur aktinomiset endofit BWA65.
Tabel 9 Hasil ekstraksi senyawa inhibitor dengan berbagai pelarut
Pelarut Rendemen (%)
kloroform 0.0100
etil asetat 0.0300
metanol 0.0260
etanol 0.0299
butanol 0.0243
Uji Aktivitas Inhibitor α-glukosidase Hasil Ekstraksi dengan Berbagai
Pelarut. Ekstrak kultur BWA 65 selanjutnya diuji aktivitas penghambatannya
terhadap enzim α-glukosidase. Pada pengujian ini enzim α-glukosidase
menghidrolisis substrat p-nitrophenyl α-D-glucopyranoside menjadi p-nitrophenol
yang berwarna kuning dan glukosa. Aktivitas enzim diukur berdasarkan
absorbansi p-nitrophenol yang berwarna kuning. Dengan adanya ekstrak kultur
BWA65 yang berperan sebagai inhibitor α-glukosidase maka p-nitrophenol yang
dihasilkan akan berkurang yang ditandai oleh berkurangnya intensitas warna
kuning. Aktivitas penghambatan α-glukosidase berbagai ekstrak kultur BWA65
dapat dilihat pada Tabel 10.
Hasil pengujian tersebut dapat dilihat aktivitas inhibitor kultur BWA65
yang diekstrak dengan berbagai macam pelarut. Ekstrak etil asetat memberikan
aktivitas penghambatan paling tinggi dibandingkan dengan ekstrak lainnya. Pada
konsentrasi 1000 μg/ml ekstrak etil asetat memberikan penghambatan sebesar
77.77%, diikuti ekstrak butanol, metanol, kloroform dan etanol dengan nilai
penghambatan sebesar 75.98%, 69.94%, 69.51% dan 68.05%. Semakin rendah
konsentrasi ekstrak menghasilkan penghambatan yang semakin rendah pula
terhadap aktivitas enzim α-glukosidase. Berdasarkan hasil uji pendahuluan
bahwa penggunaan etil asetat menghasilkan rendemen ekstrak tertinggi dan
aktivitas inhibitor α-glukosidase tertinggi dibanding pelarut lainnya, serta
senyawa etil asetat diketahui tidak bersifat toksik, maka untuk keperluan
ekstraksi selanjutnya dipilih senyawa etil asetat.

47
Tabel 10 Aktivitas inhibitor α-glukosidase berbagai ekstrak kultur aktinomiset
BWA65
Konsentrasi
μg/ml
Penghambatan ekstrak (%)
etil asetat kloroform butanol metanol etanol
1000 77.77 69.51 75.98 69.94 68.05
500 71.80 64.80 66.99 66.03 59.92
250 65.59 58.59 60.15 64.23 59.92
125 61.75 54.25 58.20 59.06 52.22
62.5 56.20 52.46 55.74 41.31 44.92
Penggunaan etil asetat untuk mengekstrak senyawa bioaktif khususnya
senyawa inhibitor α-glukosidase telah dilakukan oleh beberapa peneliti
sebelumnya. Xu et al. (2005) melakukan ekstraksi tanaman Glycyrrhiza uralensis
dengan beberapa jenis pelarut. Hasil pengujian ekstrak menunjukkan bahwa
ekstrak etil asetat dari tanaman tersebut menghasilkan hambatan terhadap α-
glukosidase tertinggi sebesar 83.2% dibanding dengan ekstrak dari pelarut yang
lain. Wu et al. (2009) juga melaporkan telah mengekstrak tanaman
Crossostephium chinense dengan etil asetat untuk mendapatkan senyawa
inhibitor α-glukosidase. Hasil fraksinasi dari ekstrak etil asetat tersebut akhirnya
diperoleh senyawa scopoletin, tanacetin, hispidulin, quercetagetin, celagin yang
menunjukkan aktivitas penghambatan yang kuat terhadap enzim α-glukosidase
secara in vitro.
Pada penelitian ini ditentukan juga nilai IC50 masing-masing ekstrak yaitu
nilai yang menunjukkan konsentrasi ekstrak yang menyebabkan penghambatan
sebesar 50% terhadap aktivitas enzim α-glukosidase yang diuji. Semakin kecil
nilai IC50 berarti semakin kuat daya hambat senyawa inhibitor terhadap enzim α-
glukosidase tersebut. Nilai ini didapat dengan cara menguji aktifitas
penghambatan suatu ekstrak pada berbagai macam konsentrasi. Setelah didapat
nilai penghambatan dari masing-masing konsentrasi ekstrak, selanjutnya dibuat
persamaan garis yang merupakan fungsi dari konsentrasi ekstrak dan besaran
penghambatan yang dihasilkan. Nilai-nilai konsentrasi ekstrak sebagai absis
(variabel X) dan besarnya penghambatan sebagai ordinat (variabel Y). Hasil
pengujian menunjukkan bahwa nilai IC50 terkecil diperoleh dari ekstrak etil asetat
yaitu sebesar 28.6 μg/ml, dan diikuti berturut-turut oleh ekstrak butanol, kloroform,

48
metanol dan etanol dengan nilai IC50 berturut-turut sebesar 38, 54, 84 dan 102
μg/ml (Gambar 10).
Gambar 10 Nilai IC50 dari berbagai ekstrak kultur BWA65 yang diekstraksi menggunakan beberapa pelarut.
Analisis Fitokimia Ekstrak Etil Asetat. Analisis fitokimia dilakukan untuk
mengetahui kandungan kualitatif senyawa metabolit sekunder dari suatu bahan
alam. Dengan analisis ini dapat diketahui golongan utama senyawa aktif yang
terkandung di dalam suatu ekstrak. Hasil pengujian fitokimia terhadap ekstrak etil
asetat kultur BWA65 dapat dilihat pada Tabel 11. Hasil uji fitokimia secara umum
menunjukkan bahwa ekstrak etil asetat kultur BWA65 mengandung senyawa
golongan flavonoid, phenol hidroquinon, steroid dan triterpenoid. Daya
penghambatan ekstrak etil asetat dari kultur aktinomiset endofit BWA65 diduga
karena kandungan senyawa fitokimia yang terdapat di dalam ekstrak tersebut.
Beberapa peneliti telah melaporkan adanya senyawa yang termasuk golongan
senyawa yang tedapat pada ekstrak tersebut yang diketahui mampu
menghambat kerja enzim α-glukosidase.
Senyawa flavonoid merupakan kelompok senyawa yang telah banyak
dilaporkan memiliki aktivitas inhibitor α-glukosidase maupun sebagai anti
hiperglikemia oleh para peneliti sebelumnya. Kim et al. (2000) telah menemukan
senyawa kelompok flavonoid, yaitu luteolin yang memiliki aktivitas penghambatan
terhadap enzim α-glukosidase maupun α-amilase. Lee et al. (2008) telah
28,6
54
38
84
102
0
20
40
60
80
100
120
etil asetat kloroform butanol metanol etanol
Nila
i IC
50
(μg/m
l)
Ekstrak

49
mengisolasi senyawa flavonoid dari kelompok flavonol dari daun tanaman
Machilus philippinensis yang memiliki aktivitas inhibitor terhadap enzim α-
glukosidase dari Bacillus stearothermophilus. Ghosh dan Konishi (2007)
menyatakan bahwa ekstrak flavonoid kelompok antosianin diketahui memiliki
potensi aktivitas inhibitor α-glukosidase dan dapat menekan peningkatan kadar
glukosa setelah makan. Tadera et al. (2006) menguji enam kelompok flavonoid
terhadap aktivitas enzim alfa amilase dan α-glukosidase. Hasil pengujian
menunjukkan bahwa flavonoid dari kelompok antosianidin, isoflavon dan flavonol
dapat menghambat enzim α-glukosidase dengan nilai IC50 kurang dari 15 µM.
Wu et al. (2012) menemukan beberapa senyawa flavonoid dari kelompok
isoflavon dari rimpang Belamcanda chinensis yaitu: rhamnosylswertisin, genistein,
genistin, mangiferin dan daidzin yang memiliki penghambatan kuat terhadap
enzim α-glukosidase secara in vitro.
Tabel 11 Kandungan fitokimia ekstrak etil asetat kultur BWA65
No Kelompok senyawa Keberadaan
1 alkaloid -
2 flavonoid ++++
3 phenol hidroquinon +++
4 steroid +
5 triterpenoid +++
6 tanin -
7. saponin -
Fraksinasi dengan Kromatografi Kolom
Pencarian Eluen Terbaik. Ekstrak etil asetat yang diperoleh pada
percobaan ini selanjutnya difraksinasi lebih lanjut menggunakan kromatografi
kolom gel silika. Sebelum dilakukan fraksinasi dengan kromatografi kolom,
terlebih dahulu perlu ditentukan eluen yang tepat sebagai fasa geraknya, yang
dapat memisahkan komponen aktif yang terdapat pada ekstrak etil asetat. Untuk
mencari eluen terbaik (fasa gerak) yang dapat memisahkan senyawa yang paling
baik, dilakukan pengujian kromatografi lapis tipis (KLT) terlebih dahulu. Pengujian
ini meliputi pencarian pelarut tunggal maupun campuran yang dapat memisahkan
senyawa-senyawa aktif yang terkandung di dalam ekstrak etil asetat tersebut.

50
Pemilihan pelarut campuran didasarkan pada studi literatur yang telah
dilakukan peneliti sebelumnya (Kim & Nho 2004). Pengujian dengan KLT
menggunakan fasa diam berupa pelat KLT gel silika GF254 dan fasa geraknya
ditentukan dengan cara coba-coba (trial and error). Dari beberapa percobaan
yang dilakukan, akhirnya diperoleh pelarut yang dapat memisahkan komponen
aktif dalam ekstrak yang ditandai dengan adanya hasil kromatografi yang berupa
spot-spot yang terpisah dengan baik. Pelarut terbaik yang dapat memisahkan
komponen aktif dari ekstrak BWA65 adalah pelarut campuran heksan dan etil
asetat dengan perbandingan 1:4 (Tabel 12).
Tabel 12 Hasil percobaan KLT untuk menentukan pelarut terbaik
Pelarut Jumlah
spot Nilai Rf
aceton 1 0.142
etanol 1 0.071
kloroform 1 0.014
metanol 1 0.014
etil asetat: aseton (1:1) 2 0.428; 0.500
etil asetat:aseton (3:1) 3 0.071; 0.142; 0.214
kloroform:metanol (4:1) 1 0.428
kloroform:metanol (4:11) 1 0.857
kloroform:metanol (5:10) 1 0.857
kloroform:metanol (3:2) 1 0.428
heksan:etil asetat (1:1) 3 0.142; 0.285; 0.714
heksan:etil asetat (3:2) 2 0.214; 0.285
heksan:etil asetat (1.5:3.5) 5 0.142; 0.285; 0.428; 0.500; 0.786
heksan:etil asetat (1:4) 7 0.240; 0.300; 0.360; 0.430; 0.460; 0.610; 0.780
heksan:etil asetat (9:1) 1 0.285
heksan:etil asetat (1:1) 3 0.214; 0.285; 0.714

51
Gambar 11 Kromatografi lapis tipis dengan variasi campuran pelarut pengem-
bang heksan: etil asetat 1:4.
Hasil pengujian diperoleh pelarut campuran heksan : etil asetat (1:4)
merupakan eluen terbaik untuk pemisahan senyawa aktif inhibitor α-glukosidase
yang terdapat pada ekstrak. Eluen ini dapat memisahkan komponen yang
terdapat dalam ekstrak menjadi 7 spot dengan nilai Retention factor (Rf) sebesar
0.24; 0.30; 0.36; 0.43; 0.46; 0.61; dan 0.78 (Gambar 11)
Setelah eluen sebagai fase gerak terbaik ditemukan, kemudian dilakukan
fraksinasi dengan kromatografi kolom kilas (flash chromatography). Pada tahap
ini, fasa diam yang digunakan adalah silika gel yang bersifat polar dan fase gerak
yang digunakan adalah heksan : etil asetat dengan perbandingan 1:4. Hasil
fraksinasi diperoleh 201 fraksi. Fraksi-fraksi ini kemudian dianalisis dengan
menggunakan kromatografi lapis tipis. Fraksi-fraksi dengan pola kromatogram
yang sama kemudian digabung menjadi satu. Hasil fraksinasi kolom dan
pemeriksaan dengan KLT diperoleh 7 fraksi gabungan (Tabel 13).
Fraksi F1 merupakan gabungan dari fraksi nomor 1-2 dengan spot
tunggal dengan nilai Rf 0.88. Fraksi F2 merupakan gabungan dari fraksi nomor 3-
7 dengan spot tunggal dengan nilai Rf 0.86. Fraksi F3 merupakan gabungan dari
fraksi nomor 8-20 dengan spot dua dengan nilai Rf 0.62 dan 0.50. Fraksi F4
merupakan gabungan dari fraksi nomor 21-55 dengan spot dua dengan nilai Rf
0.40 dan 0.35. Fraksi F5 merupakan gabungan dari fraksi nomor 58-85 dengan
spot dua dengan nilai Rf 0.31 dan 0.21. Fraksi F6 merupakan gabungan dari
fraksi nomor 86-155 dengan spot tunggal dengan nilai Rf 0.19. Fraksi F7
Spot ke Nilai Rf
1 0.78
2 0.61
3 0.46
4 0.43
5 0.36
6 0.30
7 0.24

52
merupakan gabungan dari fraksi nomor 156-201 dengan spot tunggal dengan
nilai Rf 0.15.
Tabel 13 Hasil fraksinasi senyawa menggunakan kromatografi kolom gel silika.
Fraksi Nomor fraksi Jumlah
spot RF
Berat (gram)
Rendemen (%)
F1 1-2 1 0.88 0.0580 7.25
F2 3-7 1 0.86 0.1039 12.99
F3 8-20 2 0.62; 0.50 0.0273 3.41
F4 21-55 2 0.40; 0.30 0.0362 4.56
F5 56-85 2 0.31; 0.23 0.0331 4.14
F6 86-155 1 0.19 0.1452 18.15
F7 156-201 1 0.10 0.0186 2.33
Masing-masing fraksi gabungan tersebut kemudian dipekatkan
menggunakan vacum rotary evaporator. Hasil perolehan fraksi bervariasi dengan
rendemen 2.33-18.15%. Perolehan tertinggi diperoleh pada Fraksi F6 dengan
berat ekstrak 0.1452 gram, diikuti oleh fraksi F2, F1, F4, F5, F3 dan F7 dengan
berat berturut-turut: 0.1039; 0.0580; 0.0362; 0.0331; 0.0273; dan 0.0186 gram.
Uji Penghambatan α-glukosidase Fraksi. Hasil pengujian aktivitas
inhibitor α-glukosidase senyawa aktif hasil fraksinasi menggunakan kromatografi
kolom menunjukkan hasil yang bervariasi (Tabel 14).
Tabel 14 Aktivitas inhibitor α-glukosidase hasil fraksinasi kolom
Konsentrasi
μg/ml
Penghambatan Fraksi (%)
F1 F2 F3 F4 F5 F6 F7 Acb
200 59.80 72.71 83.98 66.32 30.32 80.90 57.01 84.10
100 56.42 63.13 75.32 62.49 25.25 72.09 46.69 76.57
50 53.08 56.00 62.81 58.53 24.43 67.24 39.55 44.63
25 45.62 46.77 55.45 51.24 23.01 62.21 39.03 40.82
12.5 44.05 39.15 45.62 45.77 17.91 49.00 38.28 34.25
Aktivitas Fraksi F1-F7 dalam menghambat enzim α-glukosidase pada
konsentrasi 200 ppm menunjukkan adanya kemampuan penghambatan yang
bervariasi antara 30.32-80.93%. Aktivitas tertinggi dihasilkan oleh fraksi F3
dengan aktivitas peghambatan sebesar 83.98% pada konsentrasi 200 ppm atau
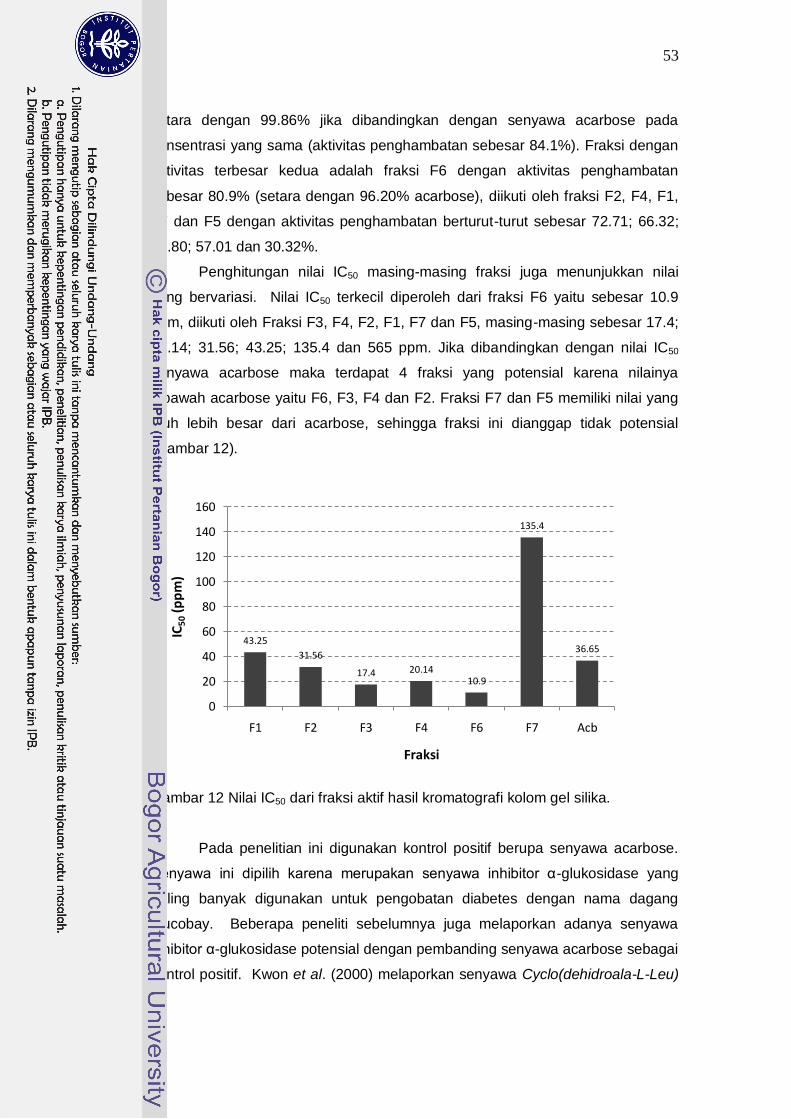
53
setara dengan 99.86% jika dibandingkan dengan senyawa acarbose pada
konsentrasi yang sama (aktivitas penghambatan sebesar 84.1%). Fraksi dengan
aktivitas terbesar kedua adalah fraksi F6 dengan aktivitas penghambatan
sebesar 80.9% (setara dengan 96.20% acarbose), diikuti oleh fraksi F2, F4, F1,
F7 dan F5 dengan aktivitas penghambatan berturut-turut sebesar 72.71; 66.32;
59.80; 57.01 dan 30.32%.
Penghitungan nilai IC50 masing-masing fraksi juga menunjukkan nilai
yang bervariasi. Nilai IC50 terkecil diperoleh dari fraksi F6 yaitu sebesar 10.9
ppm, diikuti oleh Fraksi F3, F4, F2, F1, F7 dan F5, masing-masing sebesar 17.4;
20.14; 31.56; 43.25; 135.4 dan 565 ppm. Jika dibandingkan dengan nilai IC50
senyawa acarbose maka terdapat 4 fraksi yang potensial karena nilainya
dibawah acarbose yaitu F6, F3, F4 dan F2. Fraksi F7 dan F5 memiliki nilai yang
jauh lebih besar dari acarbose, sehingga fraksi ini dianggap tidak potensial
(Gambar 12).
Gambar 12 Nilai IC50 dari fraksi aktif hasil kromatografi kolom gel silika.
Pada penelitian ini digunakan kontrol positif berupa senyawa acarbose.
Senyawa ini dipilih karena merupakan senyawa inhibitor α-glukosidase yang
paling banyak digunakan untuk pengobatan diabetes dengan nama dagang
Gucobay. Beberapa peneliti sebelumnya juga melaporkan adanya senyawa
inhibitor α-glukosidase potensial dengan pembanding senyawa acarbose sebagai
kontrol positif. Kwon et al. (2000) melaporkan senyawa Cyclo(dehidroala-L-Leu)
43.25
31.56
17.4 20.1410.9
135.4
36.65
0
20
40
60
80
100
120
140
160
F1 F2 F3 F4 F6 F7 Acb
IC5
0(p
pm
)
Fraksi

54
yang memiliki aktivitas inhibitor α-glukosidase dengan nilai IC50 sebesar 35 µg/ml
(nilai IC50 acarbose sebesar 61 µg/ml). Lee et al. (2008) menemukan senyawa
flavonol monorhamniside yang memiliki aktivitas inhibitor α-glukosidase dengan
nilai IC50 6.10 µM dan 1.00 µM, sementara nilai IC50 acarbose sebesar 0.046 μM.
Senyawa phlorotannin yang dilaporkan oleh Moon et al. (2011) memiliki aktivitas
inhibitor α-glukosidase dengan nilai IC50 sebesar 1.37-6.13 dan nilai IC50
acarbose sebesar 187 µg/ml. Elya et al. (2011) mendapatkan ekstrak berbagai
tanaman dengan nilai IC50 2.33-112.02 µg/ml dibanding nilai IC50 acarbose
sebesar 117.2 µg/ml. Peneliti lain (Nie et al. 2011) melaporkan senyawa oleanic
acid memiliki aktivitas inhibitor α-glukosidase dengan nilai IC50 sebesar 98.5 μM
dan nilai IC50 acarbose sebesar 388 μM (Tabel 15). Hasil pengujian oleh para
peneliti tersebut menunjukkan adanya nilai IC50 yang bervariasi dari senyawa
acarbose yang digunakan. Hal ini sangat tergantung pada komposisi reaksi yang
digunakan untuk menguji aktivitas inhibitor α-glukosidase senyawa bioaktif
tersebut.
Tabel 15 Aktivitas inhibitor α-glukosidase beberapa senyawa dibanding acarbose
Bahan aktif Nilai IC50 senyawa
Nilai IC50
acarbose Author
oleanolic acid 98.5 µM 388 µM Nie et al. 2011
cyclo(dehidroala-L-Leu)
35 µg/ml 61 µg/ml Kwon et al. 2000
flavonol monorhamnosida
1.0-6.10 µM
0.046 µM Lee et al. 2008
phlorotannin 1.37-6.13 µg/ml 187 µg/ml Moon et al. 2011
ekstrak berbagai tanaman
2.33-112.02 µg/ml
117.2 µg/ml Elya et al. 2011
Fraksi F6 10.9 µg/ml 36.7 µg/ml Penelitian ini
Fraksi F3 dan Fraksi F6 merupakan fraksi yang paling potensial karena
memiliki aktivitas yang setara dengan acarbose (Gambar 13). Bahkan pada
konsentrasi yang rendah (dibawah 100 ppm) aktivitas kedua fraksi ini lebih tinggi
dibanding dengan acarbose. Pada konsentrasi 12.5; 25 dan 50 ppm fraksi F6
memiliki aktivitas inhibitor sebesar 49.00; 62.21 dan 67.24% atau setara dengan
143.1%; 152.4% dan 150.1% dibanding acarbose pada konsentrasi yang sama.
Fraksi F3 pada konsentrasi 12.5; 25 dan 50 ppm memiliki aktivitas inhibitor
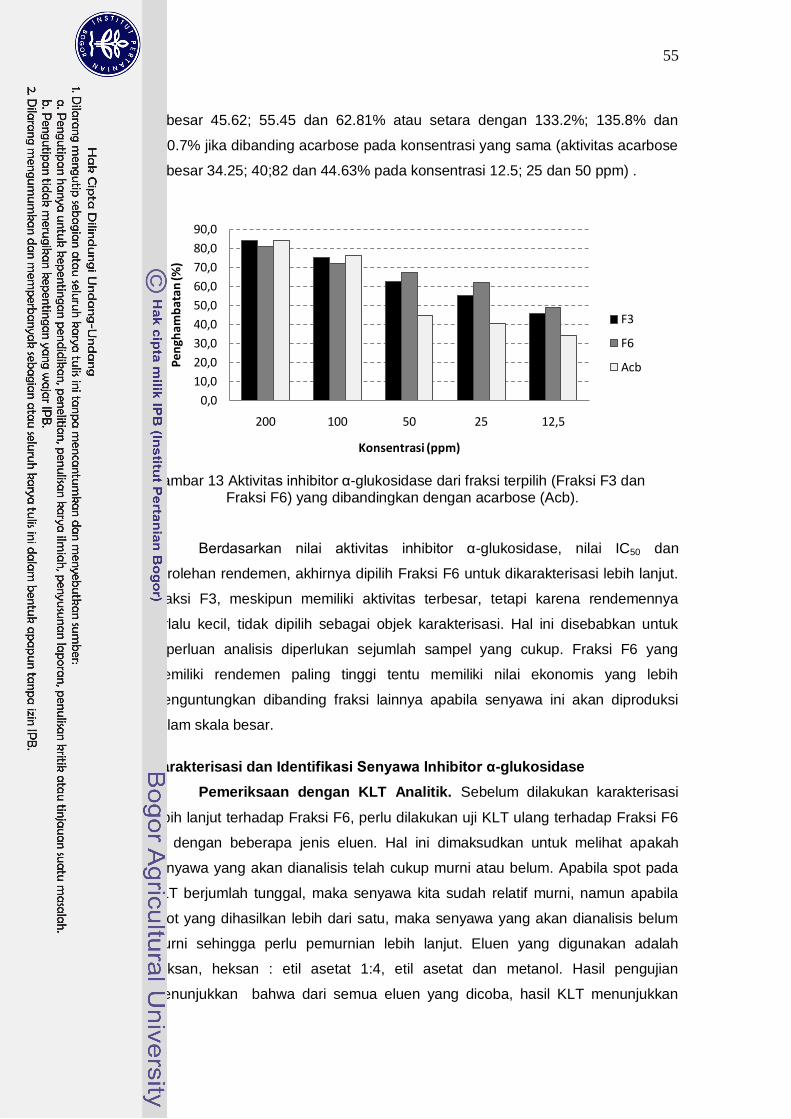
55
sebesar 45.62; 55.45 dan 62.81% atau setara dengan 133.2%; 135.8% dan
140.7% jika dibanding acarbose pada konsentrasi yang sama (aktivitas acarbose
sebesar 34.25; 40;82 dan 44.63% pada konsentrasi 12.5; 25 dan 50 ppm) .
Gambar 13 Aktivitas inhibitor α-glukosidase dari fraksi terpilih (Fraksi F3 dan
Fraksi F6) yang dibandingkan dengan acarbose (Acb).
Berdasarkan nilai aktivitas inhibitor α-glukosidase, nilai IC50 dan
perolehan rendemen, akhirnya dipilih Fraksi F6 untuk dikarakterisasi lebih lanjut.
Fraksi F3, meskipun memiliki aktivitas terbesar, tetapi karena rendemennya
terlalu kecil, tidak dipilih sebagai objek karakterisasi. Hal ini disebabkan untuk
keperluan analisis diperlukan sejumlah sampel yang cukup. Fraksi F6 yang
memiliki rendemen paling tinggi tentu memiliki nilai ekonomis yang lebih
menguntungkan dibanding fraksi lainnya apabila senyawa ini akan diproduksi
dalam skala besar.
Karakterisasi dan Identifikasi Senyawa Inhibitor α-glukosidase
Pemeriksaan dengan KLT Analitik. Sebelum dilakukan karakterisasi
lebih lanjut terhadap Fraksi F6, perlu dilakukan uji KLT ulang terhadap Fraksi F6
ini dengan beberapa jenis eluen. Hal ini dimaksudkan untuk melihat apakah
senyawa yang akan dianalisis telah cukup murni atau belum. Apabila spot pada
KLT berjumlah tunggal, maka senyawa kita sudah relatif murni, namun apabila
spot yang dihasilkan lebih dari satu, maka senyawa yang akan dianalisis belum
murni sehingga perlu pemurnian lebih lanjut. Eluen yang digunakan adalah
heksan, heksan : etil asetat 1:4, etil asetat dan metanol. Hasil pengujian
menunjukkan bahwa dari semua eluen yang dicoba, hasil KLT menunjukkan
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
200 100 50 25 12,5
Pe
ngh
amb
atan
(%)
Konsentrasi (ppm)
F3
F6
Acb

56
adanya 1 spot saja (Gambar 14). Hal ini menunjukkan bahwa senyawa yang
terkandung dalam Fraksi F6 sudah cukup murni. Meskipun demikian, akan lebih
baik apabila dilakukan pemurnian lebih lanjut terhadap Fraksi F6 ini dan
dilakukan konfirmasi menggunakan HPLC untuk memastikan kemurnian
senyawa pada Fraksi F6 tersebut.
Gambar 14 Hasil kromatografi lapis tipis Fraksi F6 menggunakan berbagai jenis eluen (H: heksan, HEA: heksan etil asetat 1:4, EA: etil asetat dan M: metanol)
Laju migrasi senyawa pada Fraksi F6 untuk masing-masing eluen yang
digunakan pada KLT bervariasi tergantung kepolaran eluen yang digunakan
(Tabel 16). Semakin polar eluen yang digunakan semakin cepat laju migrasi
senyawa pada pelat KLT sehingga semakin tinggi nilai Rf yang dihasilkan.
Penggunaan metanol sebagai eluen memberikan nilai Rf terbesar (Rf 0.81)
dibandingkan dengan penggunaan eluen etil asetat (Rf 0.44) , heksan-etil asetat
(Rf 0.19) maupun heksan saja (Rf 0.00). Kepolaran metanol lebih besar dari etil
asetat, dan kepolaran etil asetat lebih besar dari heksan. Berdasarkan hal
tersebut dapat diketahui bahwa senyawa pada Fraksi F6 merupakan senyawa
yang polar. Hal ini karena senyawa yang polar akan lebih mudah larut dalam
pelarut polar. Markham (1988) menegaskan bahwa flavonoid merupakan
senyawa yang polar karena memiliki sejumlah gugus hidroksil atau gugus gula
(bagi glikosida) sehingga akan mudah larut dalam pelarut polar seperti metanol
tersebut.
H HE
A
E
A
M

57
Tabel16 Hasil KLT analitik Fraksi F6 dengan beberapa eluen
Eluen Jumlah spot Rf
heksan 1 0.00
heksan: etil asetat 1:4 1 0.19
etil asetat 1 0.44
metanol 1 0.81
Pengaruh Konsentrasi Substrat terhadap Aktivitas Inhibitor α-
glukosidase. Percobaan ini digunakan untuk menduga tipe penghambatan yang
dimiliki oleh senyawa Fraksi F6. Pengujian aktivitas inhibitor dilakukan dengan
menggunakan substrat p-nitrophenyl α-D-glucopyranoside dengan berbagai
konsentrasi mulai dari 20 mM, 10 mM, 5 mM, 2,5 mM dan 1.25 mM. Hasil
pengujian pengaruh substrat terhadap kemampuan inhibitor Fraksi F6 dapat
dilihat pada Tabel 17 dan Gambar 15. Berdasarkan hasil tersebut terlihat bahwa
semakin tinggi konsentrasi substrat semakin kecil hambatan yang dihasilkan oleh
senyawa pada Fraksi F6. Hal ini mengindikasikan bahwa Fraksi F6 merupakan
inhibitor kompetitif. Tipe penghambatan yang dimiliki Fraksi F6 ini serupa dengan
tipe hambatan yang dihasilkan oleh senyawa acarbose. Sebagaimana diketahui
bahwa acarbose merupakan senyawa yang memiliki jenis hambatan kompetitif.
Berdasarkan analisis kinetika enzim menurut persamaan Lineweaver-Burke
diperoleh grafik seperti tertera pada Gambar 16.
Tabel 17 Pengaruh konsentrasi substrat terhadap kemampuan inhibitor α-
glukosidase hasil fraksinasi
Konsentrasi substrat (mM) Penghambatan (%)
Fraksi F6 Acarbose
20 55.88 30.34
10 76.90 63.71
5 79.66 81.35
2.5 87.12 88.16
1.25 89.30 87.73
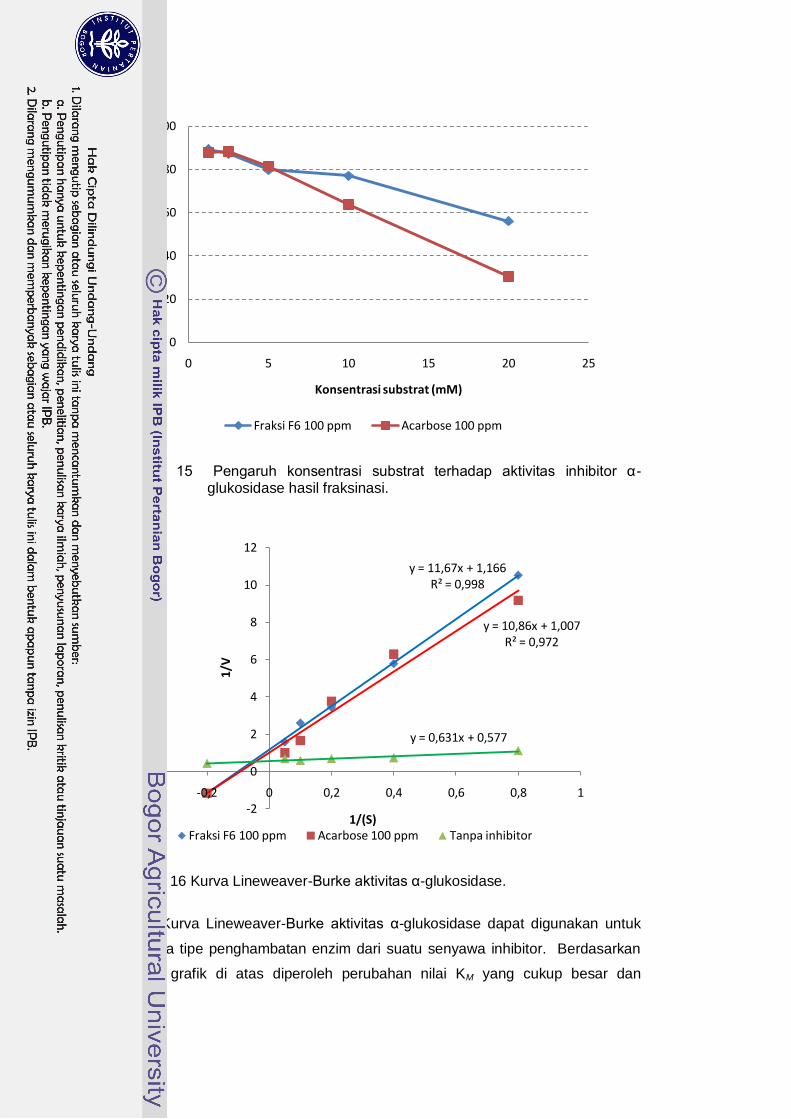
58
Gambar 15 Pengaruh konsentrasi substrat terhadap aktivitas inhibitor α-
glukosidase hasil fraksinasi.
Gambar 16 Kurva Lineweaver-Burke aktivitas α-glukosidase.
Kurva Lineweaver-Burke aktivitas α-glukosidase dapat digunakan untuk
menduga tipe penghambatan enzim dari suatu senyawa inhibitor. Berdasarkan
analisis grafik di atas diperoleh perubahan nilai KM yang cukup besar dan
0
20
40
60
80
100
0 5 10 15 20 25
Inh
ibis
i (%
)
Konsentrasi substrat (mM)
Fraksi F6 100 ppm Acarbose 100 ppm
y = 11,67x + 1,166R² = 0,998
y = 10,86x + 1,007R² = 0,972
y = 0,631x + 0,577
-2
0
2
4
6
8
10
12
-0,4 -0,2 0 0,2 0,4 0,6 0,8 1
1/V
1/(S)Fraksi F6 100 ppm Acarbose 100 ppm Tanpa inhibitor

59
perubahan Vmax yang kecil. Pola kinetika yang terbentuk setelah penambahan
inhibitor Fraksi F6 mengakibatkan peningkatan KM sebesar 8.92 mM dari 1.09
mM menjadi 10.01 mM dan penurunan Vmax sebesar 0.05 A/menit dari 0,11
A/menit menjadi 0.06 A/menit. Menurut Copeland (2005) jenis penghambatan
kompetitif adalah penghambatan yang terjadi dengan peningkatan nilai KM
dengan nilai Vmax yang tetap. Pada penghambatan kompetitif, inhibitor
berkompetisi dengan subtrat untuk memperebutkan sisi aktif enzim dan peristiwa
ini menyebabkan nilai KM menjadi lebih besar dari keadaan normalnya dengan
nilai Vmax relatif tetap. Secara teoritis, kurva ideal dari tipe penghambatan
kompetitif adalah terjadi perpotongan garis kurva pada titik yang sama pada
sumbu Y antara kurva reaksi enzim dengan penambahan inhibitor dan kurva
tanpa penambahan inhibitor. Berdasarkan hal tersebut, dapat diduga bahwa tipe
penghambatan Fraksi F6 adalah tipe penghambatan kompetitif. Ketidaktepatan
perpotongan kurva pada sumbu Y antara kurva tanpa inhibitor dan kurva dengan
penambahan inhibitor kemungkinan dapat disebabkan oleh beberapa hal antara
lain kemungkinan senyawa Fraksi F6 belum murni. Kemungkinan lainnya adalah
konsentrasi senyawa yang diuji belum tepat, sehingga perlu dilakukan pengujian
menggunakan berbagai konsentrasi inhibitor, seperti dilakukan oleh Kimura et al.
(2004). Konsekuensi dari tipe inhibitor kompetitif ini apabila nanti akan
diaplikasikan pada manusia adalah harus cermat di dalam memperhitungkan
dosis dengan pola makan pasien.
Beberapa senyawa lain juga telah diketahui mekanisme inhibitornya.
Diantara flavonoid yang dikenal memiliki aktivitas inhibitor α-glukosidase,
senyawa genistein (kelompok isoflavon) memiliki mekanisme penghambatan non
kompetitif (Lee & Lee 2001). Wu et al. (2012) juga melaporkan senyawa
swertisin yang merupakan flavonoid kelompok isoflavon juga memiliki aktivitas
penghambatan non kompetitif.
Identifikasi Inhibitor α-glukosidase dengan Uji Fitokimia. Uji fitokimia
terhadap Fraksi F6 dilakukan untuk memperkuat dugaan kelompok metabolit
utama dari senyawa kimia yang terdapat pada Fraksi F6. Hasil uji fitokimia
menunjukkan bahwa Fraksi F6 positif sebagai flavonoid. Hasil pengujian ini juga
semakin memperkuat adanya keterkaitan antara metabolit yang dihasilkan oleh
mikrob endofit dengan tanaman inangnya. Beberapa peneliti sebelumnya telah
melaporkan adanya senyawa kelompok flavonoid pada tanaman T. crispa. Hasil

60
pengujian oleh Amom et al (2009) menunjukkan bahwa ekstrak tanaman T.
crispa mengandung flavonoid yang memiliki aktivitas antioksidan yang tinggi
setara dengan antioksidan yang telah umum dikenal seperti BHT (butil hidroksi
toluen) dan vitamin C. Flavonoid yang terdeteksi pada tanaman T. crispa ini
antara lain catechin (1.58 μg/μl), luteolin (0.85 μg/μl), morin (1.44 μg/μl), dan rutin
(1.38 μg/μl). Adanya beberapa senyawa flavonoid ini diyakini secara kolektif
berfungsi sebagai antioksidan.
Patela dan Mishrab (2012) menemukan tiga senyawa dari tanaman T.
cordifolia yang memiliki kemampuan inhibitor α-glukosidase, yaitu jatrorrhizine,
palmatine dan magnoflorine. Senyawa-senyawa tersebut memiliki nilai IC50
berturut-turut sebesar 22.05; 38.42 and 7.6 µg/ml untuk jatrorrhizine, palmatine
dan magnoflorine. Hasil uji in vivo juga menunjukkan adanya penekanan yang
signifikan terhadap kenaikan kadar glukosa plasma darah oleh ketiga senyawa
tersebut pada konsentrasi 20 mg/kg berat badan. Senyawa magnoflorine
merupakan senyawa paling potensial sebagai inhibitor α-glukosidase diantara
ketiga senyawa yang ditemukan tersebut. Jika dibandingkan dengan aktivitas
senyawa yang dilaporkan oleh Patela dan Mishrab (2012) di atas maka senyawa
Fraksi F6 memiliki aktivitas yang lebih baik dibanding jatrorrhizine, palmatine
(nilai IC50 22.05 dan 38.42) tetapi lebih rendah dari pada magnoflorine (nilai IC50
7.6 µg/ml). Namun demikian untuk aktivitas in vivo dalam menurunkan kadar
gula darah pada mencit diabetes belum dapat dibandingkan, karena uji coba
senyawa inhibitor dari isolat BWA65 ini masih dalam pelaksanaan.
Disamping diketahui mengandung senyawa yang memiliki kemampuan
inhibitor α-glukosidase, tanaman T. crispa juga diketahui memiliki kandungan
senyawa bioaktif yang dapat menurunkan kadar gula darah melalui aktivitas lain.
Ruana et al (2012, in press) melaporkan adanya senyawa borapetoside dari
tanaman T. crispa yang mampu meningkatkan sensitivitas insulin sehingga
berperan sebagai anti hiperglikemia pada tikus diabetes.
Identifikasi dengan Spektrofotometer UV-Vis. Spektrum flavonoid
biasanya ditentukan dalam larutan dengan pelarut metanol atau etanol. Spektrum
khas terdiri dari atas dua puncak pada rentang 240-285 nm (pita II) dan 300-550
nm (pita I). Hasil pengujian Fraksi F6 dan Fraksi F3 menggunakan
spektrofotometer UV-Vis dapat dilihat pada Gambar 17 dan 18.

61
Gambar 17 Spektrum serapan Fraksi F6 menggunakan spektrofotometer UV-Vis.
Gambar 18 Spektrum serapan Fraksi F3 menggunakan spektrofotometer UV-Vis.
Hasil pengujian menggunakan spektrofotometer UV-Vis yang berupa
kurva yang menggambarkan nilai serapan senyawa kemudian diidentifikasi
menggunakan tabel referensi. Petunjuk mengenai rentang puncak utama yang
diperkirakan untuk setiap jenis flavonoid disajikan pada Tabel 18 dan Tabel 19.

62
Tabel 18 Rentangan serapan spektrum UV-Vis flavonoid (Markham 1988)
Pita II (nm) Pita I (nm) Jenis flavonoid
250-280 310-350 flavon 250-280 330-360 flavonol (3-OH tersubtitusi) 250-280 350-385 flavonol (3-OH bebas) 245-275 310-330 bahu
kira-kira 320 puncak isoflavon
275-295 300-330 bahu flavonon dan dihidroflavonol
230-270 340-390 khalkon 230-270 380-430 auron
270-280 465-560 antosianidin dan antosianin
Berdasarkan spektrum serapan dengan spektrofotometer UV-Vis
(Gambar 17), Fraksi F6 memiliki serapan maksimum pada panjang gelombang
423 nm dan tiga puncak pada daerah UV yaitu pada 255, 248 dan 242 nm maka
diduga Fraksi F6 ini merupakan flavonoid yang termasuk dalam kelompok auron
(Markham, 1988; Harborne, 2006). Fraksi F3 memiliki serapan maksimum pada
panjang gelombang 431 nm dan 319 nm. Berdasarkan penelusuran pada tabel
spektrum flavonoid (Tabel 18 dan 19) menurut Markham (1988) dan (Harborne
2006), nilai serapan maksimum Fraksi F3 tidak ada yang memenuhi kriteria
dalam tabel tersebut, sehingga diduga senyawa aktif pada Fraksi F3 tersebut
bukan merupakan kelompok flavonoid. Merujuk pada hasil uji fitokimia,
kemungkinan senyawa Fraksi F3 ini dapat berasal dari kelompok terpenoid. Hal
ini diperkuat adanya laporan yang menyatakan adanya senyawa kelompok
terpenoid dari tanaman T. crispa yang mempunyai aktivitas inhibitor α-
glukosidase.
Table 19 Ciri spektrum golongan flavonoid utama (Harborne 2006)
Panjang gelombang maks (nm)
Panjang gelombang tambahan (nm)
Petunjuk
475-560 ± 275 antosianin 390-430 240-270 auron
365-390 240-260 khalkon 350-390 ± 300 flavonol 250-270 330-350 Tidak ada flavon dan biflavonil 250-270 275-290 310-330 flavonon dan flavonol ± 255 255-265 310-330 isoflavon

63
Menurut Markham (1988) auron bersama dengan khalkon, flavonon,
dihidrokhalkon dan isoflavon merupakan flavonoid minor karena penyebaran
masing-masing kelas ini terbatas. Auron dan khalkon merupakan pigmen kuning
yang dapat dijumpai pada tanaman Compositae dan beberapa famili tanaman
lainnya. Salah satu contoh auron adalah aureusidin. Senyawa ini terdapat di
alam dalam bentuk glikosida.
Glikosida sendiri merupakan bentuk flavonoid yang terikat dengan gula,
sedangkan yang tidak terikat dengan gula disebut aglikon. Adanya gula yang
terikat pada flavonoid (bentuk yang umum ditemukan) cenderung menyebabkan
flavonoid lebih mudah larut dalam air dan dengan demikian campuran pelarut di
atas dengan air merupakan pelarut yang lebih baik untuk glikosida. Sebaliknya
aglikon yang kurang polar seperti isoflavon, flavonon, flavon serta flavonol yang
termetoksilasi cenderung lebih mudah larut dalam pelarut seperti eter dan
kloroform. Aglikon flavonoid adalah polifenol dan karena itu mempunyai sifat
kimia senyawa fenol, yaitu bersifat agak asam sehingga larut dalam basa
(Markham 1988).
Flavonoid mempunyai sejumlah gugus hidroksil atau suatu gula sehingga
flavonoid merupakan senyawa polar, maka umumnya flavonoid larut dalam
pelarut polar seperti etanol, metanol, butanol, aseton, dimetilsulfoksida,
dimetilformamida, air dan lain-lain. Adanya gugus berupa gula pada flavonoid
diduga kuat sebagai bagian aktif yang memiliki aktivitas inhibitor α-glukosidase.
Seperti halnya acarbose, adanya struktur gula menyebabkan senyawa ini mampu
bersaing dengan substrat untuk menempati sisi aktif enzim. Percobaan pengaruh
konsentrasi substrat mendukung adanya dugaan ini, dimana semakin tinggi
konsentrasi sustrat, maka hambatan yang dihasilkan oleh Fraksi F6 semakin
kecil. Fraksi F6 memiliki ciri-ciri seperti yang didiskripsikan di atas sehingga
senyawa Fraksi F6 ini diduga kuat merupakan flavonoid kelompok auron.
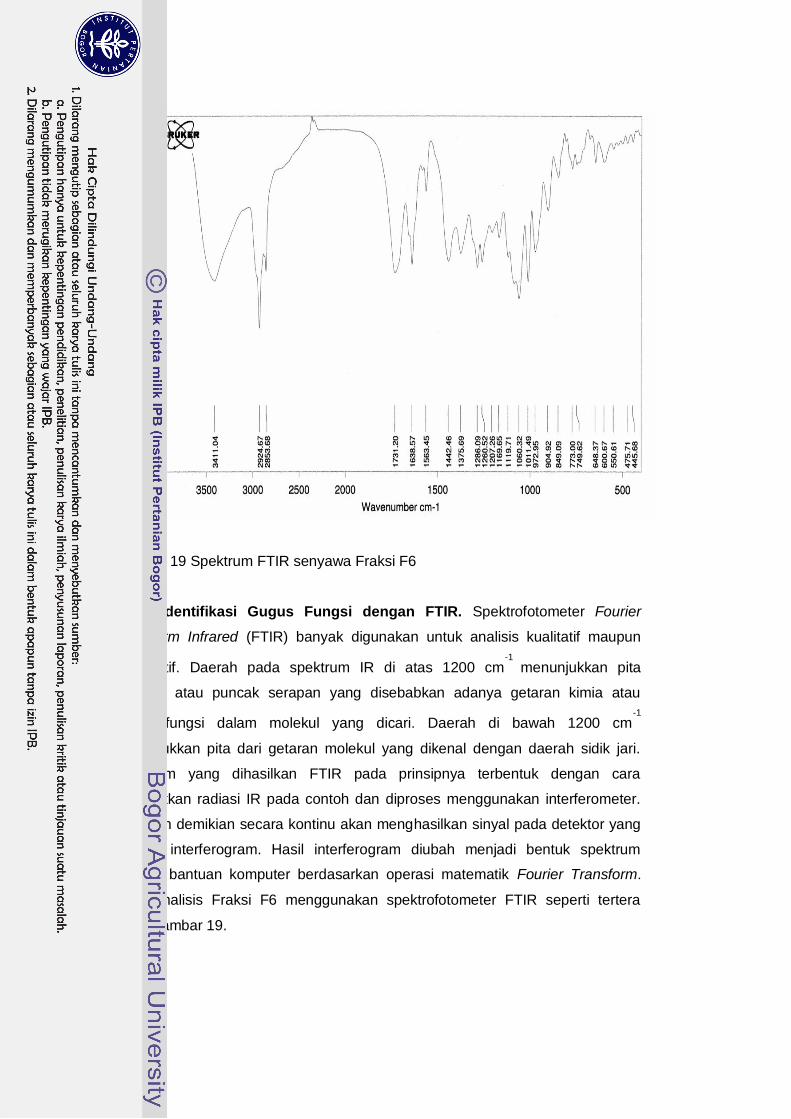
64
Gambar 19 Spektrum FTIR senyawa Fraksi F6
Identifikasi Gugus Fungsi dengan FTIR. Spektrofotometer Fourier
Transform Infrared (FTIR) banyak digunakan untuk analisis kualitatif maupun
kuantitatif. Daerah pada spektrum IR di atas 1200 cm-1
menunjukkan pita
serapan atau puncak serapan yang disebabkan adanya getaran kimia atau
gugus fungsi dalam molekul yang dicari. Daerah di bawah 1200 cm-1
menunjukkan pita dari getaran molekul yang dikenal dengan daerah sidik jari.
Spektrum yang dihasilkan FTIR pada prinsipnya terbentuk dengan cara
melewatkan radiasi IR pada contoh dan diproses menggunakan interferometer.
Keadaan demikian secara kontinu akan menghasilkan sinyal pada detektor yang
disebut interferogram. Hasil interferogram diubah menjadi bentuk spektrum
dengan bantuan komputer berdasarkan operasi matematik Fourier Transform.
Hasil analisis Fraksi F6 menggunakan spektrofotometer FTIR seperti tertera
pada Gambar 19.

65
Tabel 20 Identifikasi gugus fungsi fraksi F6 dengan FTIR (Pavia et al. 2001)
Bilangan gelombang Fraksi F6 cm
-1 Bilangan gelombang
referensi cm-1
Tipe vibrasi
3411.04 dekat 3400-2400 O-H Ikatan H
2924.67 3000-2850 C-H alkana
2853.68 2900-2800
C-H aldehid
1731.20 1740-1720 1750-1730
C=O aldehid atau ester
1638.57 1680-1630
C=O amida
1563.45 1600-1475
1640-1550
C=C N-H
aromatik atau amina dan amida primer dan sekunder
1442.46 1465
-CH2- alkana
Daerah sidik jari 900-1400
849.09 900-690
1000-650
C-H aromatik atau alkena
773.00 900-690
1000-650
C-H aromatik atau alkena
749.62 900-690
1000-650
C-H aromatik atau alkena
Berdasarkan hasil pemeriksaan dengan spektrofotometer FTIR (Gambar
17) dan penelusuran literatur (Tabel 20) diduga senyawa Fraksi F6 ini memiliki
gugus karbonil C=O pada bilangan gelombang 1731 cm-1, terdapat ikatan O-H
pada bilangan gelombang 3411.04 cm-1, terdapat ikatan C-O pada bilangan
gelombang 1300-1000 cm-1, terdapat ikatan rangkap C=C cincin aromatik pada
bilangan gelombang 1563 cm-1. Hasil analisis ini memiliki kesesuaian dengan
struktur dasar senyawa auron (Gambar 20).
Gambar 20 Struktur dasar senyawa auron.

66
Temuan Penting dan Implikasinya
Penemuan isolat aktinomiset endofit dari tanaman T. crispa yang dapat
menghasilkan inhibitor α-glukosidase semakin menambah data atau bukti ilmiah
yang dapat memperkuat teori yang menyatakan bahwa suatu tanaman dapat
mengandung mikrob endofit yang dapat menghasilkan metabolit sekunder sama
dengan yang dihasilkan oleh tanaman inangnya. Fenomena ini tentu akan
menjadi peluang besar penelitian dalam bidang mikrobiologi untuk melakukan
eksplorasi mikrob dalam rangka pencarian senyawa bioaktif baru melalui
pendekatan empiris-ilmiah. Resep-resep pengobatan tradisional yang
memanfaatkan tanaman khas dari berbagai wilayah nusantara yang diwariskan
secara kultural oleh nenek moyang merupakan modal informasi yang sangat
berharga dalam rangka pemilihan tanaman inang sebagai sumber mikrob endofit
baru penghasil senyawa bioaktif yang bermanfaat. Penelitian-penelitian tentang
senyawa bioaktif yang berasal dari mikrob endofit khususnya dari tanaman-
tanaman obat tradisional masih sangat langka, sehingga peluang untuk
mendapatkan mikrob maupun senyawa jenis baru sangat besar. Temuan-temuan
ini diharapkan dapat berkontribusi terhadap perkembangan ilmu pengetahuan.
Tanaman T. crispa bebas endofit yang diperoleh dari hasil kultur jaringan
memiliki kemampuan yang lebih rendah dalam menghasilkan senyawa inhibitor
α-glukosidase jika dibandingkan dengan tanaman T. crispa yang diperoleh dari
alam. Isolat aktinomiset endofit BWA65 mampu menghasilkan aktivitas inhibitor
α-glukosidase yang lebih besar dari tanaman T. crispa yang diperoleh dari alam.
Fenomena tersebut mengindikasikan bahwa mikrob endofit, dalam hal ini
aktinomiset endofit pada tanaman T. crispa tersebut memberikan kontribusi
besar terhadap produksi senyawa inhibitor α-glukosidase. Indikasi adanya peran
penting mikrob endofit dalam menghasilkan senyawa bioaktif ini merupakan
fenomena menarik untuk dikaji lebih lanjut, khususnya dalam budidaya tanaman
obat (penghasil senyawa bioaktif). Dalam budidaya tanaman khususnya untuk
keperluan produksi senyawa bioaktif, faktor genetis tanaman dan faktor
lingkungan (ekologi) sangat menentukan kualitas dan kuantitas senyawa bioaktif
yang dihasilkan. Faktor lingkungan di sini meliputi antara lain adalah kandungan
unsur hara tanah dan mikro klimat lokasi budidaya tanaman. Salah satu faktor
yang belum banyak dikaji adalah adanya peran mikrob endofit dalam
meningkatkan produksi senyawa bioaktif. Berdasarkan indikasi adanya peran
penting mikrob endofit dalam menghasilkan senyawa bioaktif, maka dalam

67
budidaya tanaman obat perhatian terhadap eksistensi dan kebugaran mikrob
endofit perlu dilakukan dalam rangka memaksimalkan peran mikrob endofit
dalam memproduksi bahan aktif. Penelitian peningkatan senyawa bioaktif
tanaman melalui pendekatan peran mikrob endofit juga masih sangat langka.
Adanya fenomena peran endofit dalam menghasilkan senyawa bioaktif
pada tanaman juga memiliki potensi besar dalam bidang kultur jaringan tanaman.
Dalam bidang kultur jaringan, permasalahan utama yang sering dijumpai adalah
adanya mikrob kontaminan yang tumbuh dalam eksplan yang ditanam. Selama
ini setiap mikrob yang tumbuh dari eksplan tanaman dianggap kontaminan yang
harus dihilangkan. Untuk keperluan propagasi tanaman, maka adanya mikrob
yang tumbuh pada eksplan mutlak harus dicegah, tetapi untuk keperluan
produksi seyawa bioaktif, adanya mikrob yang tumbuh tidak selalu harus dicegah.
Berdasarkan fenomena bahwa mikrob endofit dapat memiliki kontribusi yang
besar dalam menghasilkan senyawa bioaktif pada tanaman, maka adanya
mikrob yang selalu hadir pada eksplan dapat berasal dari endofit sehingga tidak
harus selalu dicegah, melainkan perlu dikaji peluangnya di dalam meningkatkan
produksi senyawa bioaktif dari kultur jaringan tersebut.
Isolat aktinomiset BWA65 memiliki kesamaan 98% dengan Streptomyces
diastaticus. Adanya inhibitor α-glukosidase yang dihasilkan oleh S. diastaticus
hingga saat ini belum pernah dilaporkan. Berdasarkan hal ini, kemungkinan
besar isolat BWA65 merupakan Streptomyces penghasil senyawa inhibitor α-
glukosidase baru, sehingga senyawa yang dihasilkan kemungkinan besar
merupakan senyawa inhibitor α-glukosidase jenis baru.
Hasil fraksinasi senyawa aktif inhibitor α-glukosidase yang dilakukan
dalam penelitian ini diperoleh Fraksi F6 yang memiliki aktivitas setara dengan
acarbose. Acarbose merupakan senyawa inhibitor α-glukosidase yang sudah
dikomersialkan dan digunakan secara luas untuk pengobatan diabetes. Pada
konsentrasi yang rendah (di bawah 100 μg/ml) aktivitas Fraksi F6 ini lebih tinggi
dibanding dengan acarbose. Berdasarkan hasil pemeriksaan menggunakan uji
fitokimia, spektrofotometer UV-Vis maupun FTIR mengindikasikan bahwa
senyawa aktif inhibitor α-glukosidase yang dihasilkan oleh isolat BWA65
merupakan senyawa flavonoid kelompok auron. Berdasarkan kajian literatur,
hingga saat ini juga belum ada laporan tentang senyawa kelompok auron yang
memiliki aktivitas inhibitor α-glukosidase. Hal ini juga membuka peluang kajian
pengembangan lebih lanjut dari senyawa aktif Fraksi F6 ini.

68

69
KESIMPULAN DAN SARAN
KESIMPULAN
Pada penelitian ini diperoleh isolat aktinomiset endofit BWA65 dari
tanaman T. crispa yang mampu menghasilkan senyawa inhibitor α-glukosidase.
Berdasarkan hasil pengujian aktivitas inhibitor α-glukosidase antara tanaman T.
crispa bebas endofit, tanaman dari alam dan isolat endofit BWA65,
mengindikasikan aktinomiset endofit tersebut memiliki kontribusi penting di dalam
menghasilkan senyawa inhibitor α-glukosidase pada tanaman T. crispa. Hasil
identifikasi berdasarkan sekuen 16S rDNA menunjukkan isolat BWA65 memiliki
kesamaan 98% dengan Streptomyces diastaticus, yang belum pernah dilaporkan
memiliki aktivitas inhibitor α-glukosidase. Isolat aktinomiset endofit BWA65
merupakan isolat potensial untuk dikembangkan sebagai penghasil inhibitor α-
glukosidase karena selain memiliki unsur kebaharuan organisme penghasilnya,
juga memiliki kemampuan inhibisi yang tinggi terhadap enzim α-glukosidase.
Senyawa inhibitor α-glukosidase yang dihasilkan isolat ini dapat diproduksi
menggunakan medium cair dan dapat diekstrak menggunakan pelarut etil asetat.
Hasil fraksinasi komponen bioaktif menggunakan kromatografi kolom diperoleh
Fraksi aktif F6 yang memiliki aktivitas inhibitor α-glukosidase setara dengan
acarbose. Fraksi F6 memiliki aktivitas penghambatan sebesar 80.9% pada
konsentrasi 200 ppm atau setara dengan 96.2% dari aktivitas acarbose (aktivitas
inhibisi 84.1%). Pada pengujian nilai IC50, Fraksi F6 memiliki nilai IC50 10.9 ppm
lebih kecil dibanding nilai IC50 acarbose sebesar 36.65 ppm.
Berdasarkan hasil pemeriksaan dengan spektrofotometer FTIR dan
penelusuran literatur mengindikasikan senyawa Fraksi F6 ini memiliki gugus
fungsi: karbonil C=O, ikatan O-H, ikatan C-O, ikatan rangkap C=C dan cincin
aromatic. Pada pengukuran dengan spektrofotometer UV-Vis senyawa ini
memiliki serapan panjang gelombang maksimum 423 nm. Berdasarkan hasil
pemeriksaan menggunakan uji fitokimia, spektrofotometer UV-Vis maupun FTIR
mengindikasikan bahwa senyawa aktif inhibitor α-glukosidase yang dihasilkan
oleh isolat BWA65 merupakan senyawa flavonoid kelompok auron.
SARAN
Pembuktian secara langsung melalui uji kemampuan kolonisasi isolat
BWA65 pada tanaman T. crispa bebas endofit perlu dilakukan untuk lebih

70
menegaskan bahwa isolat BWA65 benar-benar merupakan endofit pada
tanaman T. crispa.
Perlu dilakukan uji lanjut untuk menentukan struktur kimia senyawa aktif
Fraksi F6. Penentuan struktur ini berguna untuk keperluan verifikasi kebaharuan
senyawa yang ditemukan dalam penelitian ini.
Pengembangan lebih lanjut isolat BWA65 sebagai penghasil bahan aktif
untuk obat diabetes perlu dilanjutkan melalui tahapan uji in vivo dan uji klinis agar
dapat sampai ke komersialisasi produk.

71
DAFTAR PUSTAKA
Aguilar FJ, Dorantes TB, Leon AG, Carrillo LV, Saenz JL, Ramos RR. 2003.
Study of the antihyperglycemic effect of antidiabetic plants in rabbits with impaired glucose tolerance. Proc West Pharmacol Soc 46:148-152.
Akhtar MA, Wahed II, Islam MR, Shaheen SM, Islam MA, Amran MS, Ahmed M.
2007. Comparison of long-term antihyperglycemic and hypolipidemic effects between Coccinia cordifolia (Linn.) and Catharanthus roseus (Linn.) in alloxan-induced diabetic rats. J Med Med Sci 2:29-34.
Amom Z, Bahari H, Isemaail S, Ismail NA, Shah MD, Arsyad MS. 2009.
Nutritional composition, antioxidant ability and flavonoid content of Tinospora crispa stem. Adv Nat Appl Sci 3:88-94.
Anam K, Widharma RM, Kusrini D. 2009. Alpha glucosidase inhibitor of
Terminalia species. Int J Pharmacol 5:277-280.
Attele AS, Zhou YP, Xie JT, Wu JA, Zhang L, Dey L, Pugh P, Rue PA, Polonsky
KS, Yuan C. 2002. Antidiabetic effects of Panax ginseng berry extract and the identification of an effective component. Diabetes 51:1851-1859.
Azin M, Maozami N, Bogan MN. 1997. Production, purification and immobilization
of glucose isomerase from Streptomyces olivochro-mogenes PTCC 1547. World J Microbiol Biotechnol 13:597-598.
Bailey CJ, Day C. 2003. Antidiabetic drugs. Br J Cardiol 10:128-136.
Benalla W, Bellahcen S, Bnouham M. 2010. Antidiabetic medicinal plants as a
source of α-glucosidase inhibitors. Curr Diabetes Rev 6:247-54.
Beltrame FM, Pessini GL, Doro DL, Dias BP, Bazotte RB, Garcia DA. 2002.
Evaluation of the antidiabetic and antibacterial activity of Cissus sicyoides. Braz Archiv Biol Technol 45:21-25.
Bhandari MR, Anurakkum NJ, Hong G, Kawabata J. 2008. Alpha glucosidase
and alpha amilase inhibitory activities of Nepalese medicinal herb Pakhanbhed (Berginia ciliata). Food Chem 106:247-252.
Bhat M, Zinjarde SS, Bhargava SY, Kumar AR, Joshi BN. 2008. Antidiabetic
Indian plants: a good source of potent amilase inhibitor. Evid Compl Altern Med 11:1-6.
Bnouham M, Ziyyat A, Mekhfi H, Tahri A. 2006. Medicinal plants with potential
antidiabetic activity - a review of ten years of herbal medicine research (1990-2000). Int J Diabet Metabol 14:1-25.
Cannell RJP, Farmer P, Walker JM. 1988. Purification and characterization of
pentagalloylglucose, an α-glucosidase inhibitor / antibiotic from freshwater green algae Spirogyra varians. J Biochem 255:937-941.

72
Caruso MA, Colombo L, Fedeli L, Pavesi A, Quaroni S, Saracchi M, Ventrella G. 2000. Isolation of endophytic fungi and actinomycetes taxane producers. Ann Microbiol 50:3-13.
Castillo UF, Strobel GA, Ford EJ, Hess WM, Porter H, Jensen JB, Albert H,
Robison R, Condron MA, Teplow DB, Stevens D, Yaver D. 2002. Munumbicins, wide-spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscans. Microbiology 148:2675-2685.
Chen H, Yan X, Lin W, Zheng L, Zhang W. 2004. A new method for screening α-
glucosidase inhibitors and application to marine microorganisms. Pharm Biol 42:416-421.
Chin YW, Balunas WJ, Chai HB, Kinghorn AD. 2006. Drug discovery from
natural sources. Aaps J 8:239-252.
Chougale AD, Ghadyale VA , Panaskar SN, Arvindekar AU. 2009. Alpha
glucosidase inhibition by stem extract of Tinospora cordifolia. J Enz Inhib Med Chem 24:998-1001.
Clark CM. 1998. Oral therapy in type 2 diabetes: pharmacological properties and
clinical use of currently available agents. Diabet Spectrum 11:211-221.
Coombs JT, Franco CM. 2003. Isolation and identification of actinobacteria from
surface –sterilized wheat roots. Appl Environ Microbiol 69:5603-5608.
Copeland RA. 2005. Evaluation of Enzyme Inhibitors in Drug Discovery: a Guide
for Medical Chemists and Pharmacologists. New Jersey: Wiley-
Interscience. Djomeni PD, Tedong L, Asongalem EA, Dimo T, Sokeng SC, Kamtchouing P.
2006.
Hypoglycaemic and antidiabetic effect of root extracts of Ceiba pentandra in normal and diabetic rats . Afr J Trad Compl Altern Med
3:129-136. Du ZY, Liu RR, Shao WY, Mao X, Ma L, Gu L, Huang Z, Chan ASC. 2006. Α-
glucosidase inhibition of natural curcuminoids and curcumin analogs. Eur J Med Chem 41:213-218.
Dutta R, Qin L, Inouye M. 1999. Histidine kinases: diversity of domain
organization. Mol Microbiol 34:633-640. Ebong PE, Atangwho IJ, Eyong EU, Egbung GE. 2008. The antidiabetic efficacy
of combined extracts from two continental plants: Azadirachta indica (A. Juss) (Neem) and Vernonia amygdalina (Del.) (African Bitter Leaf). American J Biochem Biotech 4:239-244.
Elya B, Basah K, Munim A, Yuliastiti. 2012. Screening of α-glucosidase inhibitory
activity from some plants of Apocynaceae, Clusiaceae, Euphorbiaceae, and Rubiaceae. J Biomed Biotechnol 12:1-6.
Ezra D, Castillo UF, Strobel GA, Hess WM, Porter H, Jensen JB, Condron MAM,
Teplow DB, Sears J, Maranta M, Hunter M, Weber B, Yaver D. 2004.

73
Coronamycins, peptide antibiotics produced by a verticillate Streptomyces Sp. (Msu-2110) endophytic on Monstera sp. Microbiology 150:785-793.
Fischer PB, Chollin M, Karlsson GB, James W, Butter TD. 1995. The α-
glucosidase inhibitor N-butyldeoxynojirimycin inhibit human immunodeficiency virus entry at the level of post CD4 binding. J Virol
69:5791-5797. Garg M, Dhar VJ, Kalia AN. 2008. Antidiabetic and antioxidant potential of
Phyllanthus fraternus in alloxan induced diabetic animals. Pharmacog Mag 4:138-143
Gholamhoseinian A, Fallah H, Sharifi F, Mirtajaddini M. 2008. The inhibitory
effect of some Iranian plants extract on the α-glucosidase. Iran J Basic Sci
11:1-9. Ghosh D, Konishi T. 2007. Anthocyanins and anthocyanin-rich extracts: role in
diabetes and eye function. Asia Pac J Clin Nutr 16:200-208. Gray AM, Flat PR. 1999. Insulin-secreting activity of the traditional antidiabetic
plant Viscum album (mistletoe). J Endocrinol 160:409-414. Gown J. 2006. Diabetes drug produced by a microbe. Di dalam: Beth Burrows,
editor. Out of Africa: Mysteries of Access and Benefit Sharing.
Washington: The Edmonds Institute. hlm 1-4. Gu Q, Luo H, Zheng W, Liu Z, Huang Y. 2006. Pseudonocardia oroxyli sp. Nov.,
a novel actinomycete isolated from surface–sterilized Oroxylum indicum root. Int J Syst Evol Microbiol 56:2193-2197.
Harborne JB. 2006. Metode Fitokimia: Penuntun Cara Modern Menganalisis
Tumbuhan. Penerbit ITB Bandung.
Hemker M, Stratmann A, Goeke K, Schro¨Der W, Lenz J, Piepersberg W, Pape
H. 2001. Identification, cloning, expression, and characterization of the extracellular acarbose-modifying glycosyltransferase, AcbD, from Actinoplanes sp. strain SE50. J Bacteriol 183:4484-4492.
Hsieh PC, Huang GJ, Ho YL, Lin YH, Huang SS, Chiang YC, Tseng MC, Chang
YS. 2010. Activities of antioxidants, α-glucosidase inhibitors and aldose reductase inhibitors of the aqueous extracts of four Flemingia species in Taiwan. Bot Std 51:293-302.
Hyun CG, Kim SY, Hur JH, Seo MJ, Suh JW, Kim SO. 2005. Molecular detection
of α-glucosidase inhibitor-producing actinomycetes. J Microbiol 43:313-318.
Igarashi Y, Miura S, Fujita T, Furumai T. 2006. Cpterocidin, a cytotoxic
compound from the endophytic Streptomyces hygroscopicus. J Antibiot
59:193-195. Irawan D. 2008. Isolasi aktinomiset endofit tanaman obat yang berpotensi
sebagai antidiabetes melalui kajian aktivitas α-glukosidase [skripsi].

74
Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Jeyacandran R, Mahesh A. 2007. Enumeration of antidiabetic herbal flora of
Tamil Nadu. Resc J Med Plant 1:144-148.
Katiyar SK, Arwal R, Muhtar, H. 1996. Inhibition of tumor promotion in SENCAR
mouse skin by etanol extract of Zingiber officinale rhizome. Cancer Resc
56:1023-1030. Khalilsaraie MF, Saima N, Meti NT, Bhadekar NK, Nerkar DP. 2011.Cytological
study and antimicrobial activity of embryogenic callus induced from leaf cultures of Tinospora cordifolia (Willd.) Miers. J Med Plants Resc 5:3002-
3006. Kim JS, Kwon CS, Son KH. 2000. Inhibition of α-glucosidase and amylase by
luteolin, a flavonoid. Biosci Biotechnol Biochem 11:2458-2461. Kim JS, Ju JB, Choi CW, Kim SC. 2006. Hypoglycemic and antihyperlipidemic
effect of four Korean medicinal plants in alloxan induced diabetic rats. Am J Biochem Biotechnol 2:154-160.
Kim SD, Nho HJ. 2004. Isolation and characterization of α-glucosidase inhibitor
from fungus Ganoderma lucidum. J Microbiol 42:223-227. Kim YM, Wang MH, Rhee HI. 2003. A novel α-glucosidase inhibitor from pine
bark. Carbo Resc 339:715-717. Kimura A, Lee JH, Lee IS, Lee HS, Park KH, Chiba S, Kim D. 2004. Two potent
competitive inhibitors discriminating α-glucosidase family I from family II. Carbo resch 339:1035-1040.
Klein G, Kim J, Himmeldirk K, Cao Y, Chen X. 2007. Antidiabetes and antiobesity
activity of Lagerstroemia speciosa. Evid Compl Altern Med 4:401-407. Kumar A, Ilavarasan R, Jayachandran T, Deecaraman M, Aravindan P,
Padmanabhan N, Krishan MRV. 2008. Antidiabetic activity of Syzygium cumini and its isolated compound against streptozotocin-induced diabetic rats. J Med Plant Resc 29:246-249.
Kwon OS, Park SH, Yun BS, Pyun YR, Kim CJ. 2000. Cyclo(Dehydroala-L-Leu),
an α-glucosidase inhibitor from Penicillium sp. F70614. J Antibiot 53:954-
958. Lamda HS, Bhargava CS, Thakur M, Bhargava S. 2011. α-glucosidase and
aldose reductase inhibitory activity in vitro and antidiabetic activity in vivo of tribulus terrestris (Dunal). Int J Pharm Pharm Sci 3:270-272.
Lee DS, Lee JM, Kim SU, Chang KT, Lee SH. 2007. Ceptezole, a cephem
antibiotic is an α-glucosidase inhibitor with in vivo antidiabetic activity. Int J Molec Med 20:279-283.

75
Lee DS, Lee SH. 2001. Genistein, a soy isoflavone, is a potent α-glucosidase inhibitor. FEBS Letters 501:84-86.
Lee SS, Lin HC, Che CH. 2008. Acylated flavonol monorhamnosides, α-
glucosidase inhibitors. Phytochemistry 69:2347-2353.
Lin WL, Fu LH, Sattler I, Huang X, Grabley S. 2005. New cyclopentenone
derivatives from an endophytic Streptomyces sp. isolated from the mangrove plant. Aegiceras comiculatum. J Antibiot 58:594-598.
Mahop MP, Mayet M. 2007. En route to biopiracy: ethnobotanical research on
anti medicinal plants in the Eastern Cape Province, South Africa. Afr J Biotechnol 6:2945-2952.
Markham KR. 1988. Cara Mengidentifikasi Flavonoid. Bandung: Penerbit ITB.
Mbouangouere RN, Tane P, Choudary MI. 2008. Piptadenol A-C and α-
glucosidase inhibitor from Piptadenia africana. Resc J Phytochem 2:27-34.
Moon HE, Islam MN, Ahn BR, Chowdhury SS, Sohn HS, Jung HA, Choi JS. 2011.
Protein tyrosine 1B and α-glucosidase inhibitory phlorotannins from edible brown algae, Ecklonia stolonifera and Eisena bicyclis. Biosci Biotechnol Biochem 75:1472-1480.
Nakao Y, Maki T, Matsunaga S, Soest RWM, Fusetani N. 2000. Penarolide
sulfates A1 and A2, new α-glucosidase inhibitors from a marine sponge Penares sp. Tetrahedron 56: 8977-8987.
Nie W, Luo JG, Wang XB, Yin H, sun HB, Yao HQ, Kong LY. 2011. Synthesis of
new α-glucosidase inhibitors based on oleanolic acid incorporating cinnamic amides. Chem Pharm Bull 59:1051-056.
Noipha K, Ninlaaesong P. 2011. The activation of GLUT1, AMPKα and PPARγ
by Tinospora crispa in L6 myotubes. Spatula 1:245-249. Noor H, Ashcroft. 1998. Pharmacological characterization of the anti-
hyperglycaemic properties of Tinospora crispa extract. J Ethnopharmacol 62:7-13.
Pasaribu GT. 2009. Zat Ekstraktif kayu raru dan pengaruhnya terhadap penurun
kadar gula darah secara in vitro [tesis]. Bogor. Sekolah Pascasarjana, Institut Pertanian Bogor.
Patela MB, Mishrab SM. 2012. Magnoflorine from Tinospora cordifolia stem
inhibits α-glucosidase and is antiglycemic in rats. J Func Food 4:78-86.
Pavia DL, Lampman GM, Kriz GS. 2001. Introduction to Spectroscopy: a Guide
for Students of Organic Chemistry. USA: Thomson Learning.
Purwianingsih W, Hamdiyati Y. 2009. Metode elisitasi menggunakan ragi
Sacharomyces cerevisiae untuk meningkatkan kandungan bioaktif kuinon kalus Morinda citrifolia L. (Mengkudu). Biosainstifika 1:167-178.

76
Radji M. 2005. Peranan bioteknologi dan mikrob endofit dalam pengembangan obat herbal. Majalah Ilmu Kefarmasian 2:113-126.
Rajalakshmi M, Eliza J, Priya CE, Nirmala A, Daisy P. 2009. Antidiabetic
properties of Tinospora cordifolia stem extracts on streptozotocin-induced diabetic rats. Afr J Pharm Pharm 3:171-180.
Ruana T, Lamb SH, Chic TC, Leeb SS, Sua MJ. 2012. Borapetoside C from
Tinospora crispa improves insulin sensitivity in diabetic mice. Phytomedicine (in press)
Seco EM, Cuesta T, Fotso S, Laatsch H, Malpartida F. 2005. Two polyene
amides produced by genetically modified Streptomyces diastaticus var 108. Chem Biol 12:535-543.
Santa IG, Prajogo E, Bambang W. 1998. Studi taksonomi brotowali (Tinospora
crispa (L.) Miers ex Hook F. and Thoms. Jakarta: Kelompok Kerja Nasional Tumbuhan Obat Indonesia. http://isjd.pdii.lipi.go.id/index.php-/Search.html [6 Juli 2012].
Sharma R, Ranjan R, Kapardar RK, Grover A. 2005. Unculturable bacterial
diversity: an untapped resource. Current Sci 89:71-77.
Simkada JR, Lee HJ, Jang SY, Cho SS, Park EJ, Sohng JK, Yoo JC. 2009. A
novel alkalo and thermostable phospholipase D from Streptomyces olivochromogenes. Biotechnol Lett 31:429-435.
Sriyapai C, Dumma R, Kongkathip N, Kritiyanunt S. 2009. Hypoglycemic effect of
Tinospora crispa dry powder in out patients with metabolic syndrome. J Healt Resc 23:125-133.
Strobel GA, Daisy B. 2003. Bioprospecting for microbial endophytes and their
natural products. Microbiol Molec Biol Rev 67:491-502.
Strobel GA. 2001. Microbial gifts from rain forests. Can J Plant Pathol 24:14-20.
Subramanian RM, Asmawi Z, Sadikun A. 2008. Effect of ethanolic extract of
Andrographis paniculata nees on a combination of fat-fed diet and low dose streptozotocin induced chronic insulin resistance in rats. Diabet Croatica 37:13-22.
Subroto A. 2006. Ramuan Herbal untuk Diabetes Mellitus. Jakarta: Penebar
Swadaya. Sugiwati S, Kardono LBS, Bintang M. 2006. Alpha glucosidase inhibitory activity
and hipoglycemic effect of Phalleria macrocarpa fruit pericarp extracts by oral administration to rats. J Appl Sci 6:2312-2316.
Sun OL, Choi GJ, Choi YH, Kim CJ, Kim JC. 2008. Isolation and characterization of endophytic actinomycetes from Chinese cabbage roots and antagonists to Plasmodiophora brassicae. J Microbiol Biotechnol 18:1741-1746.

77
Sunil C, Latha G, Mohanraj P, Kalichevan K, Agastian P. 2009. Alpha glucosidase inhibitor and antidiabetic activities of ethanolic extract of Pisonia alba leaves. Int J Integrat Biol 6:41-45.
Susheela T, Balaravi P, Theophilus J, Reddy JN, Redd PUM. 2008. Evaluation of
hypoglycaemic an antidiabetic effect of Melia dubia CAV fruits in mice. Curr Sci 94:1191-1195.
Sutini B, Tatik W, Wahyu W,Sumitro SB. 2008. Meningkatkan produksi flavan-3-
ol melalui kalus Camellia sinensis L. dengan elisator CU2+. Berk. Penel. Hayati: 14:39-44.
Suthindhiran K, Jayasri MA, Kannabiran K. 2009. Alpha glucosidase inhibitor and
α-amylase inhibitor of Micromonospora sp. VITSDK3. Int J Integrat Biol 6:115-120.
Tadera K, Minami Y, Takamatsu K, Matsuoka T. 2006. Inhibition of α-glucosidase
and α-amylase by flavonoid. J Nutr Sci Vitaminol 52:149-153.
Taechowisan T, Lu C, Shen Y, Lumyong S. 2007. Antitumor activity of 4-
arylcoumarins from endophytic Streptomyces aureofaciens CMU Ac130. J Cancer Resc Ther 3:86-91.
Taechowisan T, Lumyong S. 2003. Activity of endophytic actinomycetes from
roots of Zingiber officinale and Alpinia galanga against phytopathogenic fungi. Ann Microbiol 53:291-298.
Tajana E, Fiechter A, Zimmermann W. 1992. Purification and characterization of
two alpha-L-arabinofuranosidases from Streptomyces diastaticus. Appl Environ Microbiol 58:1447-1450.
Takahashi Y. 2004. Exploitation of new microbial resources for bioactive
compounds and discovery of new actinomycetes. Actinomycetologica
18:54-61. Tan RX, Zou WX. 2001. Endophytes: a rich source of functional metabolites. Nat
Prod Rep 18:448-459. Topliss JG, Clark AM, Ernst E, Hufford CD, Johnston GAR, Rimoldi JM,
Weimann BJ. 2002. Natural and synthetic substances related to human health. Pure Appl Chem 74:1957-1985.
[WHO] World Health Organization.1999. Definition, diagnosis and classification of
diabetes mellitus and its complification. Dept. of noncomunicable and disease surveillance. Geneva.
Wu C, Shen J, He P,Chen Y, Li L, Zhang L, Li Y, Fu, Y, Dai R, Meng W, Deng Y.
2012. The α-glucosidase inhibiting isoflavones isolated from Belamcanda chinensis leaf extract. Rec Nat Prod 6:110-120.
Wu Q, Yang X, Zou L, Fu D. 2009. Bioactivity guided isolation of alpha-
glucosidase inhibitor from whole herbs of Crossostephium chinense. Chin J Chin Mat Med. 34:2206-2211.

78
Xiancui L, Rongli N, Xiao F, Lijun H, Lixin Z. 2005. Macroalgae as a source of
alpha-glucosidase inhibitors. Chin J Oceanol Limnol 23:354-356. Xu Y, Ni J, Meng Q, Gao Y, Fan B. 2005. Preliminary study on the alpha-
glucosidase inhibitor from Glycyrrhiza uralensis. J Chin Med Mat 28:890-
891. Yosuke SU, Tokomazu, Takashi T, Toshikazu O. 1992. Novel α-glucosidase
inhibitor, pradimicin Q. US Patent 5091418. Yulinah E, Sukrasno, Fitri MA. 2001. Aktivitas antidiabetika ekstrak etanol herba
sambiloto (Andrographis paniculata Nees, Acanthaceae). J Mat Sains
6:13-20. Zhang CS, Stratmann S, Block O, Bruckner R, Podeschwa M, Altenbach HJ,
Wehmeier, UF, Piepersberg W. 2002. Biosynthesis of the C7-cyclitol moiety of acarbose in Actinoplanes Species SE50/110. J Biol Chem
277:22853-22862. Zhu YP, Yin LJ, Cheng YQ, Yamaki K, Mori Y, Su YC, Li LT. 2008. Efffect of
source of carbon and nitrogen on production of α-glucosidase inhibitor by newly isolated strain of Bacillus subtilis B2. Food Chem 109:737-742.
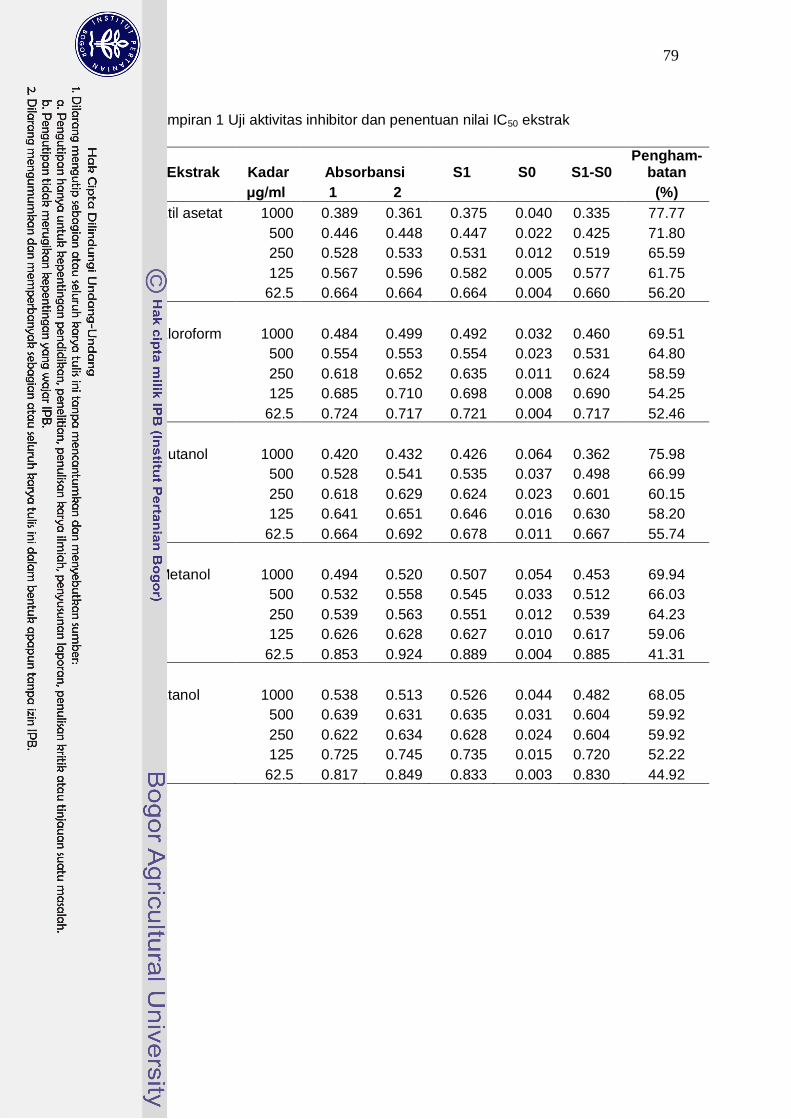
79
Lampiran 1 Uji aktivitas inhibitor dan penentuan nilai IC50 ekstrak
Ekstrak Kadar Absorbansi S1 S0 S1-S0 Pengham-
batan
μg/ml 1 2
(%)
Etil asetat 1000 0.389 0.361 0.375 0.040 0.335 77.77
500 0.446 0.448 0.447 0.022 0.425 71.80
250 0.528 0.533 0.531 0.012 0.519 65.59
125 0.567 0.596 0.582 0.005 0.577 61.75
62.5 0.664 0.664 0.664 0.004 0.660 56.20
Kloroform 1000 0.484 0.499 0.492 0.032 0.460 69.51
500 0.554 0.553 0.554 0.023 0.531 64.80
250 0.618 0.652 0.635 0.011 0.624 58.59
125 0.685 0.710 0.698 0.008 0.690 54.25
62.5 0.724 0.717 0.721 0.004 0.717 52.46
Butanol 1000 0.420 0.432 0.426 0.064 0.362 75.98
500 0.528 0.541 0.535 0.037 0.498 66.99
250 0.618 0.629 0.624 0.023 0.601 60.15
125 0.641 0.651 0.646 0.016 0.630 58.20
62.5 0.664 0.692 0.678 0.011 0.667 55.74
Metanol 1000 0.494 0.520 0.507 0.054 0.453 69.94
500 0.532 0.558 0.545 0.033 0.512 66.03
250 0.539 0.563 0.551 0.012 0.539 64.23
125 0.626 0.628 0.627 0.010 0.617 59.06
62.5 0.853 0.924 0.889 0.004 0.885 41.31
Etanol 1000 0.538 0.513 0.526 0.044 0.482 68.05
500 0.639 0.631 0.635 0.031 0.604 59.92
250 0.622 0.634 0.628 0.024 0.604 59.92
125 0.725 0.745 0.735 0.015 0.720 52.22
62.5 0.817 0.849 0.833 0.003 0.830 44.92

80
Contoh penentuan nilai IC50
Nilai IC50 ekstrak etil asetat:
Dari data konsentrasi ekstrak dan aktivitas inhibitor diperoleh persamaan garis
yang menunjukkan hubungan antara konsentrasi inhibitor (x) dengan aktivitas
inhibitor (y) sebagai berikut:
Y = 7.673 ln(x) + 24.25
50 = 7.673 ln(x) + 24.25
ln(x) = (50-24.25):7.673
ln(x) = 3.356
x = 28.6
y = 7,673ln(x) + 24,25R² = 0,994
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
0 200 400 600 800 1000 1200
Inh
ibis
i (%
)
Konsentrasi (μg/ml)
Etil asetat

81
Lampiran 2 Hasil uji aktivitas hasil kromatografi kolom dan penentuan nilai IC50
Fraksi Kadar Absorbansi S1 S0 S1-S0 Pengham-
batan
μg/ml 1 2 3 (%)
F1 200 0.541 0.581 0.578 0.567 0.028 0.539 59.80
100 0.603 0.616 0.608 0.609 0.025 0.584 56.42
50 0.649 0.647 0.644 0.647 0.018 0.629 53.08
25 0.712 0.764 0.755 0.744 0.015 0.729 45.62
12.5 0.776 0.748 0.74 0.755 0.005 0.750 44.05
F2 200 0.551 0.436 0.422 0.470 0.104 0.366 72.71
100 0.547 0.568 0.571 0.562 0.068 0.494 63.13
50 0.668 0.627 0.618 0.638 0.048 0.590 56.00
25 0.792 0.739 0.744 0.758 0.045 0.713 46.77
12.5 0.843 0.841 0.84 0.841 0.026 0.815 39.15
F3 200 0.325 0.194 0.203 0.241 0.026 0.215 83.98
100 0.31 0.372 0.364 0.349 0.018 0.331 75.32
50 0.521 0.547 0.475 0.514 0.016 0.498 62.81
25 0.61 0.605 0.621 0.612 0.015 0.597 55.45
12.5 0.705 0.751 0.739 0.732 0.003 0.729 45.62
F4 200 0.41 0.519 0.503 0.477 0.026 0.451 66.32
100 0.511 0.53 0.527 0.523 0.02 0.503 62.49
50 0.54 0.636 0.545 0.574 0.018 0.556 58.53
25 0.638 0.691 0.676 0.668 0.015 0.653 51.24
12.5 0.747 0.713 0.747 0.736 0.009 0.727 45.77
F5 200 0.914 1.125 0.96 1.000 0.066 0.934 30.32
100 0.925 1.16 0.983 1.023 0.021 1.002 25.25
50 0.941 1.168 0.989 1.033 0.02 1.013 24.43
25 0.945 1.171 1.033 1.050 0.018 1.032 23.01
12.5 0.997 1.208 1.128 1.111 0.011 1.100 17.91
F6 200 0.286 0.278 0.279 0.281 0.025 0.256 80.90
100 0.367 0.41 0.411 0.396 0.022 0.374 72.09
50 0.5 0.435 0.433 0.456 0.017 0.439 67.24
25 0.56 0.508 0.502 0.523 0.017 0.506 62.21
12.5 0.785 0.655 0.634 0.691 0.008 0.683 49.00
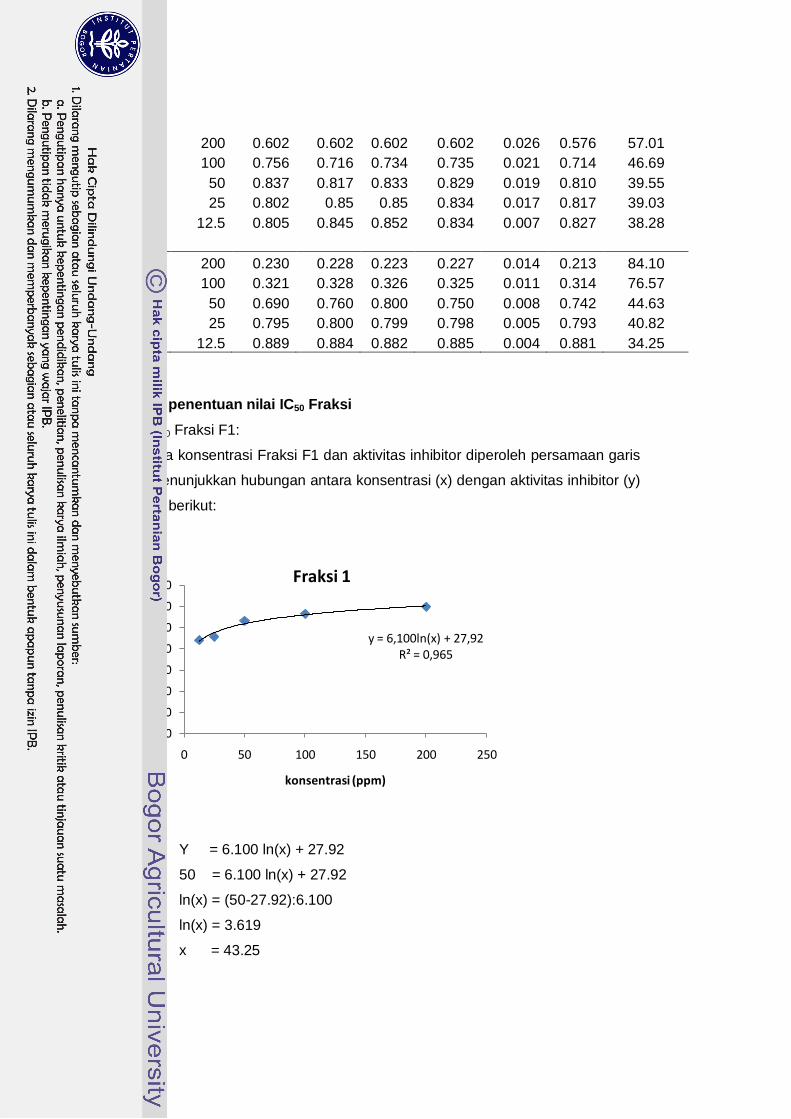
82
F7 200 0.602 0.602 0.602 0.602 0.026 0.576 57.01
100 0.756 0.716 0.734 0.735 0.021 0.714 46.69
50 0.837 0.817 0.833 0.829 0.019 0.810 39.55
25 0.802 0.85 0.85 0.834 0.017 0.817 39.03
12.5 0.805 0.845 0.852 0.834 0.007 0.827 38.28
Acb 200 0.230 0.228 0.223 0.227 0.014 0.213 84.10
100 0.321 0.328 0.326 0.325 0.011 0.314 76.57
50 0.690 0.760 0.800 0.750 0.008 0.742 44.63
25 0.795 0.800 0.799 0.798 0.005 0.793 40.82
12.5 0.889 0.884 0.882 0.885 0.004 0.881 34.25
Contoh penentuan nilai IC50 Fraksi
Nilai IC50 Fraksi F1:
Dari data konsentrasi Fraksi F1 dan aktivitas inhibitor diperoleh persamaan garis
yang menunjukkan hubungan antara konsentrasi (x) dengan aktivitas inhibitor (y)
sebagai berikut:
Y = 6.100 ln(x) + 27.92
50 = 6.100 ln(x) + 27.92
ln(x) = (50-27.92):6.100
ln(x) = 3.619
x = 43.25
y = 6,100ln(x) + 27,92R² = 0,965
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
0 50 100 150 200 250
Inh
ibis
i (%
)
konsentrasi (ppm)
Fraksi 1

83
Lampiran 3 Hasil sekuensing gen 16S rRNA isolat BWA65

84
85
Lampiran 4 Hasil analisis BLAST sekuen gen 16S rRNA isolat BWA65

86

87
Lampiran 5. Tanaman brotowali
Gambar 21 Tanaman brotowali (Koleksi Kebun Tanaman Obat ,PS Biofarmaka
IPB, Cikabayan, Bogor).
akar
daun
batang





























