Embed Size (px)
Citation preview

T.C. SÜLEYMAN DEMİREL ÜNİVERSİTESİ
SAĞLIK BİLİMLERİ ENSTİTÜSÜ
KANTARON OTUNDAN (HYPERİCUM PERFORATUM L.) ELDE EDİLEN HYPERİSİN MADDESİNİN İNSAN
LENFOSİT KÜLTÜRLERİNDE KARDEŞ KROMATİD DEĞİŞİMİ (KKD) ÜZERİNE ETKİSİ
Nezahat KAŞTAN YÜCEL
TIBBİ BİYOLOJİ ANABİLİM DALI YÜKSEK LİSANS TEZİ
DANIŞMAN Doç. Dr. H. Ramazan YILMAZ
Bu Tez Süleyman Demirel Üniversitesi Araştırma Fonu tarafından 746 no’lu proje ile desteklenmiştir.
TEZ NO: 33
2006-ISPARTA

i
KABUL VE ONAY
Sağlık Bilimleri Enstitüsü Müdürlüğüne;
Süleyman Demirel Üniversitesi Sağlık Bilimleri Enstitüsü
Tıbbi Biyoloji Anabilim Dalı Yüksek Lisans Programı
Çerçevesinde yürütülmüş olan bu çalışma, aşağıdaki jüri üyeleri tarafından yüksek
lisans tezi olarak kabul edilmiştir.
Tez savunma tarihi: 25/01/2006
Tez Danışmanı: Doç Dr. H. Ramazan YILMAZ
Üye :Doç. Dr. H.Ramazan YILMAZ
Süleyman Demirel Üniversitesi Tıbbi Biyoloji Anabilim Dalı
Üye :Prof. Dr. Nurten ÖZÇELİK Süleyman Demirel Üniversitesi Tıbbi Biyoloji Anabilim Dalı
Üye :Yrd.Doç.Dr.Ekrem ÇİÇEK Süleymen Demirel üniversitesi Farmakoloji Anabilim Dalı
ONAY: Bu tez Enstitü Yönetim Kurulunca belirlenen yukarıdaki jüri üyeleri tarafından
uygun görülmüş ve enstitü yönetim kurulu kararıyla kabul edilmiştir.
Prof. Dr. Halis KÖYLÜ Enstitü Müdürü

ii
TEŞEKKÜR
Tıbbi Biyoloji Anabilim Dalında bana Yüksek lisans çalışmalarımı yapma
olanağı sağladığı için değerli hocam sayın Prof. Dr. Nurten ÖZÇELİK’e,
Tez çalışmalarım sırasında yardımları ve eleştirileri ile bana katkıda bulunan
tez danışman hocam sayın Doç. Dr. H. Ramazan YILMAZ’a,
Araştırmalarımda kullandığım yöntemleri öğrenmemde, laboratuar
çalışmalarımda ve tezimi yazmamda yardımlarını esirgemeyen Tıbbi Biyoloji A.D.
Öğretim Üyeleri Yrd. Doç.Dr. Efkan UZ’a, Yrd. Doç. Dr. Nilüfer CALAPOĞLU’na,
Araştırma Görevlileri: Arş. Gör. Pınar KOŞAR’a, Arş. Gör. Mustafa SOYÖZ’e,
Arş.Gör. Esin ÇETİN’e, Arş. Gör. Ayşe ALTUBAŞAK’a ve Arş. Gör. Barış
YAŞAR’a,
Her zaman destek olan iş arkadaşlarım Isparta Devlet Hastanesi Kan
Merkezi çalışanlarına,
Çalışmalarımda anlayışını ve desteğini esirgemeyen değerli eşim Bilal
Serdar YÜCEL’e, sevgili kızım R. Ahsen YÜCEL’e sonsuz teşekkür ediyorum.
Bu çalışmayı destekleyen Süleyman Demirel Üniversitesi Bilimsel
Araştırma Projeleri Yönetim Birimi (SDÜBAP)’ne teşekkür ederim.

iii
İÇİNDEKİLER
KABUL VE ONAY ...................................................................................................... i TEŞEKKÜR................................................................................................................. ii İÇİNDEKİLER ...........................................................................................................iii SİMGELER VE KISALTMALAR............................................................................. iv ŞEKİL, TABLO, GRAFİK ve RESİM LİSTESİ......................................................... v 1. GİRİŞ ....................................................................................................................... 1 2. GENEL BİLGİLER ................................................................................................. 4
2.1. Hypericum Perforatum L. Hakkında Genel Bilgiler ......................................... 4 2.1.1. Hypericum perforatum L.’nin Sistematiği ................................................. 4 2.1.2. Hypericum L.’nin Genel Özellikleri .......................................................... 4 2.1.3. Hypericum L. Genusunun Taşıdığı Maddeler............................................ 6 2.1.3.1. Hyperisin ................................................................................................ 7 2.1.3.2. Hyperforin ............................................................................................ 19
2.2. Kardeş Kromatid Değişimi (KKD) ................................................................. 20 2.2.1. KKD’nin Tarihçesi................................................................................... 21 2.2.2. KKD’nin Kullanım Alanları .................................................................... 21 2.2.3. KKD Mekanizmaları ................................................................................ 22 2.2.4. KKD Oluş Mekanizması ile İlgili İleri Sürülen Modeller ....................... 24 2.2.5. KKD Yöntemini Etkileyen Faktörler ....................................................... 29
3. MATERYAL VE METOD.................................................................................... 32 3.1. Materyal .......................................................................................................... 32
3.1.1. Kimyasallar .............................................................................................. 32 3.1.2. Diğer Malzemeler.................................................................................... 32 3.1.3. Deney Gruplarının Oluşturulması ............................................................ 32
3.2. Metod .............................................................................................................. 33 3.2.1. Hücre Kültürlerinin Kurulması ................................................................ 33 3.2.2. Kromozom Eldesi ....................................................................................34 3.2.3. KKD Boyama Tekniği .............................................................................35 3.2.4. KKD’nin Değerlendirilmesi..................................................................... 36 3.2.5. İstatistiksel İşlemler ................................................................................. 37 3.2.6. Fotografik İşlemler................................................................................... 37
4. BULGULAR.......................................................................................................... 38 5. TARTIŞMA VE SONUÇ ...................................................................................... 44 ÖZET ......................................................................................................................... 48 SUMMARY ............................................................................................................... 49 KAYNAKLAR .......................................................................................................... 50

iv
SİMGELER VE KISALTMALAR
KKD : Kardeş Kromatid Değişimi
KA : Kromozom Aberasyonu
PDT : Fotodinamik Tedavi
MAP kinaz : Mitoz Aktive Edici Protein Kinaz
PUFA : Poliansatüre Yağ Asidi
PKC : Protein Kinaz C
CDK : Siklin Depended Kinaz
ROS : Reaktif Oksijen Radikalleri
RT : Revers Transkriptaz
ATP : Adenozin Trifosfat
MHO : Monoamin Oksidaz
COMT : Katekolamin Metil Tranferaz
GABA : Gama Amino Bütirik Asit
FPG : Floresans ve Giemsa Yöntemi
3 HdTh : 3H-deoksi-timidin, Radyoaktif Timidin
BrdU : 5-Bromo-2 deoxyüridin
TNFR : Tümör Nekroz Faktör Reseptörü
TRAIL : TNF-releated apopitozis indused ligand

v
ŞEKİL, TABLO, GRAFİK VE RESİM LİSTESİ
Şekil 1: Hypericum perforatum L.’nin genel görünümü …………………….. 4
Şekil 2: Hyperisin’in aromatik formülü …………………………… ………… 9
Şekil 3: Hyperforini’n aromatik görünümü …………………………… ……... 19
Şekil 4: Kromozomlarda oluşabilecek KKD’lerin şematik gösterimi ………… 23
Şekil 5: KKD oluşum mekanizması …………….……………………………… 24
Şekil 6: Timinin ve BrdU’in halkasal yapısı ……………….…………………. 25
Şekil 7: BrdU varlığına bağlı spirilizasyon gecikmesi ………………….……... 26
Şekil 8: Evens isimli araştırıcının ileri sürdüğü KKD oluş mekanizması ...…… 27
Şekil 9: KKD oluş mekanizması …………….………………………………..... 29
Tablo 1: Hypericum perforatum metanol ekstraktının taşıdığı bileşikler ……..... 7
Tablo 2: Hypericum perforatum’um antidepresan etkisi kanıtlanmış extraktları ... 17
Tablo 3: Gruplar arasındaki KKD değerleri …….…………………………….… 38
Tablo 4: Kadın ve erkek KKD değerleri ……………………………………..... 41
Grafik 1: Grup I ve II’nin KKD değerlerinin karşılaştırılması …………….......... 39
Grafik 2: Grup I ve III’ün KKD değerlerinin karşılaştırılması ………………….. 40
Grafik 3: Grup II ve III’ün KKD değerlerinin karşılaştırması …………………. 40
Resim 1: Grup I’e ait bir metafaz örneği ………………………………………... 42
Resim 2: Grup II’e ait metafaz örneği ……………………………………….….. 42
Resim 3: Grup III’e ait bir metafaz örneği …………………………………….... 43

1
1. GİRİŞ
Bitkilerin tedavide kullanımları insanlık tarihi kadar eskiye dayanmaktadır.
İnsanoğlu ilk çağlarda, hastalıkları iyileştirebilmek için tabiata ve hayvanlara
özellikle de bitkilere yönelmiştir. Sınama yanılma yöntemi ile bazen etkili ilaçlar
bulup onlardan istifade etmiştir. Afyon bitkisinden saflaştırılmış olan morfin, kodein
gibi ilaçlarla, digoksin ve digitoksin gibi kalp glikozitleri de halkın ilaç olarak
kullandığı bitkilerden elde edilmiştir (1,2).
Tez konusu olarak seçtiğimiz halk arasında “sarı kantoran, binbirdelik otu”
olarak bilinen Hypericum perforatum, Türkiye’de ve Avrupa’da yaygın yetişen
yabani bir bitkidir. H. perforatum Hypericaceae (Guttiferae) familyasından olup bu
familyanın Türkiye’de 89 türü yetişmektedir. Bunların 43’ü ise endemiktir (3). H.
perforatum geleneksel folklorik ilaç olarak dahilen ağrı giderici, yatıştırıcı, parazit
düşürücü, ülser tedavi edici ve haricen de yara iyileştirici olarak ülkemizde
kullanılmaktadır (4).
Yapılan araştırmalarla bu tıbbi bitkinin antitümör (5), antiviral (6),
antidepresan (7), antibakteriyal (8), antiinflamatuar (9), analjezik (10) ve
hepatoprotektif (11) etkilerinin olduğu belirlenmiştir. Avrupada St. John Wort olarak
tanınan bu bitkinin en zengin hyperisin (Hy) ve türevleri (pseudohyperisin) kaynağı
olduğu bulunmuştur (12). Hy bilinen en güçlü doğal fotosensitizerdir (13). Son
yıllarda bu bileşikler tümöral ve viral hastalık tedavisinde, ayrıca ılımlı depresyon
yatıştırmada önem kazanmıştır.
1988’den beri antiviral etkisi araştırılmakta olup etki mekanizması henüz
açıklanamamıştır (14). Işığa duyarlaştırıcı olan ve lipitte çözünen Hy’nin tümör
hücreleri ve zarflı virüsler üzerine güçlü fotodinamik etkisi olduğu rapor edilmiş
fakat Hy’in tümör hücresi ölüm mekanizmasını nasıl teşvik ettiği çeşitli teoriler ileri
sürülmesine rağmen henüz tam olarak açığa kavuşturulamamıştır. Hy’nin bu
antitümör ve antiviral etkisinin fotosensitizer özelliğinden kaynaklandığı
bilinmektedir. Hy’nin düşük dozlarda antiproliferatif etki gösterdiği, yüksek dozlarda
ise hem apopitozis hem de nekroz yoluyla tümör hücrelerini yok ettiği
düşünülmektedir (15).

2
Hücre ve organellerin yıkımını ışık aracılığı ile katalize eden fotosensitizer
ajanların kullanılmasına fotodinamik terapi (FDT) denilmektedir. FDT kanser
tedavisi için kullanılan yeni ve umut verici yöntemlerden biridir. Antitümoral PDT
bir fotosensitizerin sistemik uygulamasını ve tümöral lezyona oksijen varlığında
görülebilir ışığın dağıtılmasını hedefler (16). Bu sağlandığı zaman tip I veya tip II
reaksiyonu vasıtasıyla reaktif oksijen radikallerinin lokal üretimi lipit membranlarda
zincirleme reaksiyonları gerçekleştirerek tümör harabiyetine sebep olur (17). FDT
virüslere uygulanırsa zarf yapısının bozulmasını sağlayarak virüsün infekte etme
özelliğinin kaybına neden olur (18). Son yapılan araştırmalarda ışıkla aktive edilen
Hy’nin intrinsik veya ekstrinsik apopitozis yollarından her ikisini de aktifleyebileceği
ileri sürülmüş kesin bir açıklama getirilememiştir (19).
Tarih boyunca insanlar H. perforatum’u ruhsal sıkıntı, hafif depresyon,
yorgunluk, bitkinlik tedavisi için kullanmışlardır. Günümüzde ise H. perforatum
eksraktları, depresyon yatıştırmada doğal teropatik ajanların en faydalı
olanlarındandır (20). Ayrıca diğer antidepresanların etki mekanizmalarından farklı
bir etki mekanizmasına sahip olduğu düşünülmektedir. Nörokimyasal etkisinin
santral sinir sisteminde H. perforatum’a ya bir çok düzenleyicinin cevap vermesi ya
da enzimatik aktivite mekanizmasının inhibisyonu şeklinde olduğu ileri sürülmüştür.
Antidepresan etkinin, H. perforatum’un içerdiği hyperisinler, hyperforinler,
pikosiyanidinler ve flavanoidlerin farklı etki şekilleriyle olduğu varsayılmaktadır
(21).
Çeşitli Hypericum türlerinin yapraklarından hazırlanan metanol ekstrakları
çeşitli tiplerdeki yaralara uygulanmış, standart yara iyileştirici merhemlerle
karşılaştırılabilir seviyede iyi sonuçlar elde edilmiştir (22). Türkiye’de geleneksel
olarak halk arasında zeytinyağı içerisine bu bitkinin çiçeklerinin katılarak güneşte
bekletilip elde edilen karışımın yara iyileştirici olarak haricen uygulandığı
bilinmektedir (23).
Kardeş Kromatid Değişimi (KKD) analizi, genotoksik ajanların
deoksiribonükleik asitte (DNA) oluşturduğu hasarın kromozom düzeyinde tespit
edilmesini sağlayan doğrudan metodlardan birisidir (24). KKD, DNA replikasyonu

3
sırasında kardeş kromatidler arasında özdeş segmentlerin karşılıklı simetrik yer
değiştirmesidir. Bu olay kromozomun her iki kromatidinin birden kırılması ile ortaya
çıkmaktadır. KKD’de yeni duplike olmuş kromatid ve eski kendi kardeş kromatidi
arasında, kromozom morfolojisi değişmeden simetrik olarak özdeş segmentlerin
değişimi söz konusudur (25). KKD, çoğalmakta olan hücrelerde spontan olarak
meydana gelir. Özellikle kromozom hasarı, instabilitesi ve DNA tamir bozukluğu
sendromlarında duyarlı bir parametre olarak kullanılmaktadır. KKD, DNA’daki çok
küçük harabiyetlerin bile hassas göstergesi olarak kabul edilir (25, 26).
Bu çalışmada, tümör hücrelerinde apopitozisi indüklediği bilimsel
çevrelerce kabul edilmiş fakat genotoksit etkisinden bahsedilmemiş olan Hy’nin
insan lenfosit hücrelerinde genotoksik etkisinin olup olmadığının araştırılması
amaçlanmaktadır.

4
2. GENEL BİLGİLER
2.1. Hypericum Perforatum L. Hakkında Genel Bilgiler
2.1.1. Hypericum perforatum L.’ nin Sistematiği
Divisio : Spermatophyta
Subdivisio: Angiospermae
Clasis : Dicotyledoneae
Subclasis : Magnolipsida
Ordo : Theales
Familya : Cluciaceae ( Hypericaceae, Guttiferae)
Genus : Hypericum perforatum (Kantaron otu, Binbirdelik otu) (3)
Şekil 1: Hypericum perforatum L.’nin genel görünümü (20)
Türkçe: Binbirdelik otu, Sarı kantaron, kan otu, yara otu, koyun kıran, kılıç
otu, püren (4,20,27,28).
İngilizce : St. John’s wort (4,20)
Almanca : Johanniskraut (4,20)
Fransızca : Millepertius (4,20)
2.1.2. Hypericum L.’nin Genel Özellikleri
Hypericum L. (Hypericaceae): Çok yıllık otsu çalımsıdır. Yapraklar
perfoliat (sarıcı) veya değil, nadiren aurikulat (kulaklı) olup sepaller ve petallar 5,

5
serbest, petallar genellikle sarı, stamenler 5 grup halinde ve petallerin önündedir.
Ovaryum 3-5 veya tek lokuluslu, 2-çok ovüllü. Meyvesi pestisid kapsula veya
nadiren bakka. Kapsulanın üzerinde boyuna çizgiler (vittae) veya enine kabartılar
(vesicle) bulunur. Avrupa ve Anadolu’da yaygın bir türdür. Dünyada sıcak ve ılıman
bölgelerde yayılış gösteren 400’ün üzerinde türü olup Avrupa’da 10, Türkiye’de 89
türü bulunur. Bunlardan 43’ü endemiktir. Hypericum L.’nin türkiyede en yaygın
temsil edilen türleri, Hypericum perforatum L., H. Trigqetrifolium, Hypericum
calycinum (Büyük çiçekli binbirdelik otu), H. empetrifolium Willd. (püren, sarı
püren), H. scabrum L (mayasıl otu, kepirotu), H. tedrapetum Fries’dir (3,4).
Hypericum perforatum L: Tarla yol orman kıyılarında, tepelerde ve
çayırlarda temmuzdan eylül ayına kadar çiçeklenen ve ülkemizde , sarı kantaron,
kanotu, kılıç otu mayasıl otu, kuzukıran, koyunkıran ve yara otu gibi yöresel adlara
da sahip olan şifalı bir bitkidir. Bitki 30-100 cm boyunda, çok senelik ve otsudur.
Yapraklar 10-35 mm uzunlukta, elips biçiminde ve hemen hemen sapsızdır.
Yapraklar ışığa karşı tutulduğunda yağ guddeleri, çok miktarda parlak noktacıklar
halinde kolaylıkla görülür. Bu özelliğinden dolayı binbirdelik otu denmiştir. Çiçek
beş parçalı petaller altın sarısı renkli, kenarları siyah benekli gudde tüyleri ile
çevrilidir. Stamenler çok adette ve 3 demet halinde toplanmış. Bütün Türkiye ve
Avrupada yabani olarak yetişir. Ülkemizde yetişen üç varyetesi vardır: H. perforatum
var. perforatum, H. Perforatum var. Angustifolium, H. perforatum var. microfhyllum
(3).
Hypericum türleri farklı salgısal yapı tiplerinin varlığı ile karakterize edilir:
Saydam bezler ya da oyuklar, siyah nodüller ve salgı kanalları (29).
Hıristiyan mistiziminde bu bitkinin önemli bir yeri vardır. Yaklaşık 1300
yıldan beri Hristiyan geleneğinin sembolik bir bitkisi olarak kabul görmektedir. Bitki
Avrupa Ülkelerinde çoğunlukla St. John Wort olarak bilinmektedir. Rivayetlere göre
bitkinin ismi Yahya (Johannas) peygamberden gelmektedir. İncil’de geçen bir
olaydan ötürü Yahya Peygamber, Hıristiyanlar için mucizevi bir bitki getirmiştir. Bu
bitki H. Perforatum’dur. Diğer bir rivayet ise haçlı seferleri sırasında yaralanan St.
John şövalyelerinin yaralarının tedavisi bu bitki ile yapıldığından bu ismi aldığı
şeklindedir (20).

6
Hazırlanan tentürleri orta şiddetteki depresif durumlarda, özellikle menepoz
sıkıntılarını giderici olarak kullanılır. Haricen bitkisel yağlar içinde hazırlanıp yara
ve yanık tedavi edici olarak kullanılır (9).
H. trigqetrifolium: Akdeniz Doğu Avrupa’da doğal olarak yetişir.
Türkiyenin hemen hemen bütün bölgelerinde yaygındır. Antalya; Akseki, Manavgat,
Muğla, Mersin, Tarsus’da bulunmaktadır. Geleneksel sedatif, antihelmintik
antiinflematuar ve antiseptik etkileri için kullanılır. Gastrointestinal hastalıklar ve
yanık tedavisinde kullanılır (3).
Hypericum calycinum (Büyük çiçekli binbirdelik otu): 20-60cm boyunda
büyük çiçekli bir türdür. Bilhassa Kuzey Anadolu’da yaygındır (22).
H. empetrifolium Willd. ( püren, sarı püren): 20-60 cm boyunda, dik, dallı
ve küçük çiçekli bir türdür. Bilhassa Batı Anadolu’da kumaşları sarıya boyamak için
kullanılır (22).
H. scabrum L (mayasıl otu, kepirotu): 15-45 cm yükseklikte gövdesi
genellikle pürtüklü olan bir türdür. Çiçekler genellikle gövdenin ucunda, birçoğu bir
arada bulunur. Anadoluda yaygın bir türdür. Kayseri-Yozgat yörelerinde çiçekli
dallarından hazırlanan infüzyon (%1), dahilen basura ve kabıza karşı
kullanılmaktadır (22).
H. tedrapetum Fries: 10-130 cm yükseklikte, gövdesi dört kanatlı ve küçük
çiçekli bir türdür. Kuzey ve Güney Anadolu’da yaygındır (22).
2.1.3. Hypericum L. Genusunun Taşıdığı Maddeler
H. perforatum bitkisinde naftodiantoronlar (hyperisin, pseudohypersin,
izohyperisin), flovonlar (hyperosit rutin, quarsetol, quersitin, Biflovanoit), uçucu
yağlar (carophyllene, α- pinene, seskiterpenler), protein, tanen, karoten, vitamin C,
resin, hyperforin bileşikler bulunmaktadır (Tablo 1).

7
Tablo 1: Hypericum perforatum metanol ekstraktının taşıdığı bileşikler (4,11).
Bileşik Türü Miktar Bulunduğu
yer
Naftodiantron
(% 0,1-0,3)
hypericin
pseudohypericine
izohypericine
95-4660 ppm
kök,gövde,
yaprak,çicek
tohum
Flovanlar
(%4-5)
hyperosit
rutin
quarsetol
biflovanoit
quercitin
117.100 ppm
çiçekte
Uçucu yağlar
(%0,1)
carophyllene
α- pinene
seskiterpenler
yaprak
çiçek
kök
Tanen (%10) çiçek
Karoten 165 ppm tohum
Vitamin C 395 ppm kök, tohum
Hyperforin
Protein 181.000-207.000 ppm
kök, tohum
Resin
2.1.3.1. Hyperisin
Hypericum perforatum’da hyperisin adı verilen bir diantron bulunmaktadır.
Hy, hegzahidroksi dimetil naftodiantron’dur (2). Hy hypericum genusunun en önemli
metabolitlerinden ikincisidir (30). Hy ve pseudohyperisin bitkiyi böcek ve zararlı
hayvanlara karşı koruyan allokimyasal fotosensitizer bileşiklerdir. Hyperisin ve
onun protoformu pseudhyperisin, bitkinin kökü, yaprakları, sepal, petal, stamen ve
taze ovüllerinde ışığa tutulunca görülen küçük koyu bezlerde lokalize granüller
olarak bulunur (31). Bitkide pseudohyperisin (protonaftodiontoron) / hyperisin
(naftodiantoron) oranının yüksek tutulması veya içinde bulundukları veziküllerin
geçirgen olmaması sayesinde bitki ışığa bağlı fotoharabiyetten korunmaktadır (15).

8
Hy fotosensibilizasyon meydana getiren bir maddedir. Bu bitkiyi yiyen
hayvanlardan, yalnız beyaz tüylü olanlarda, güneş ışığı tesiri ile, deri ve mukozada
yaralar ve genel metabolizma bozuklukları ile beliren özel bir hastalık
(hypericismus) meydana gelir. Hy küçük dozlarda mental depresyona karşı tonik ve
stimülan olarak kullanılmaktadır (2,4) Hy’nin tümor hücreleri ve viruslar üzerine
güçlü bir fotodinamik etkiye sahip olduğu tespit edilmiş fakat bu bileşiğin tümör
ölüm mekanizmasını nasıl teşvik ettiği henüz açığa kavuşturulamamıştır. Hy’nin
zarflı virüsleri inaktif hale getirdiği fakat zarfsız virüslere bir etkisinin olmadığı rapor
edilmiştir. Bu kabiliyetinin ışığa bağlı arttığı da belirtilmiştir (6). Hy’nin apopitozisi
uyardığı birkaç tümör: Glioma, hipofiz adenomu, retinal pigment epitel hücreleri
maling T lenfositler. Hyperisinin protein kinaz C’nin potansiyel inhibitörü olduğu
gösterilmiştir. Hy MAP kinaz ve EGF reseptör trozin kinaz inhibitörü olarak ve
usülüne uygun ışık ve uygun konsantrasyonda uygulanınca HeLa hücreleri içinde
mitokondriyal hekzokinaz inhibisyonuna da etkili olduğu belirtilmiştir (32, 33).
A) Hyperisin Molekülünün Genel Özellikleri
Hyperisin molekülünün 54 atom ve 260 elektronu olan polisiklik
naftodiontorondur. 7 ve 14 pozisyonundaki karbonil grupları ile çok sağlam bir yapı
oluşturur (34).
Kimyasal formülü: C30H16O8
Çözünürlüğü: DMSO, %100 etenol, metanol, aseton, etilmetilketon, pridin
ve diğer organik çözücüler.
Molekül ağırlığı: 504,4
Aromatik formülü (Şekil 2).

9
Şekil 2: Hyperisin’in aromatik formülü (35).
Hy molekülünün son derece dinamik bir yapısı vardır. Hy hücre membranı
lipid bileyerleri arasına girebilir. Çoğunlukla lipofilik özelliktedir. Sulu çözeltilerinde
nötral, anyonik ve katyonik halde bulunabilir (5).
Yanlarda bulunan iki hidroksi ve iki metil grup Hy’nin sekizli halkasal
yapısının her iki yanında kanat gibi durur ve Hy molekülü aynı düzlemde uzanmaz.
Hidroksi ve metil gruplar birbirini iter. Bunlar hyperisin molekülünün sabit durmayıp
burkulma hareketi yapmasını sağlar (36).
Hy bir yanı hidrofobik bir yanı hidrofilik olan eşsiz bir moleküler yapıya
sahiptir. Hidrofobik metil grupları içeren hyperisin molekülünün tepe alt ve yan
tarafı nonpolardır. Tahminen hidrofobik kısım yağ içine gömülür. Triplet oksijenden
daha az reaktif olan tekil oksijen iki elektronu çiftli bağ içeren hedeflere bağlar
(PUFA’da olduğu gibi). Hidrofilik kısım sulu ortamda hidrojen bağlayabilir (37).
Hy’nin antiviral ve antitumoral özellikleri tamamıyla olmasa da büyük
oranda onun hyperisin’nin fotodinamik özelliklerine bağlıdır. Hy’nin fotodinamik
reaksiyonlarının altında yatan mekanizma tam olarak anlaşılamamıştır. Onun hücre
membranına bağlanan sensitize bir pigment olarak ışık ve oksijen arasında
interaksiyonlar sağladığı bilinmektedir. Hy ile yapılan fotodinamik reaksiyonlarda
ana oksidant olarak tekil oksijen hizmet verir (37 ).

10
Hy absorbsiyon spectromu 600-700 nm arasındadır (36). Bu da kırmızı ve
yeşil ışığa tekabül eder. Teorik olarak ışık quantasını absorbe eder ve oksijen
bulunan ortamda tekil oksijen formuna çevirir. 650-700 nm ışığa maruz bırakılırsa
tekil oksijenin üretildiği tip II fotosensitizasyonuna uğrar (38). Hy eritrositlerin
fotohemolizi gibi hücresel komponentlerin fotooksidasyonunu tetikler (39 ).
Tekil oksijenin polisiklik dionlarda bulunan pi-elektron sistemine karşı
güçlü bir affinitesi vardır. Pi elektronlar hyperisin’nin fotoaktif özelliklerinden
sorumludur (37).
Hipotez olarak bu yapı pi elektronların enerji seviyesini düşürür ve tekil
oksijenin geçici olarak bağlanmasına neden olur. Daha sonraki bir noktada viral
replikasyon mekanizmasını bozmak için tekil oksijenleri salıverebilir (37).
B) Hyperisinin Fotodinamik Özellikleri
Son 10 yıl içersinde yapılan bir çok çalışma ile Hy fotodinamik aktiviteleri
açıklanmıştır. Bunlar:
1) Işıkla uyarılan Hy’nin protein kinaz C (PKC) ve diğer büyüme
faktörleri ile uyarılan protein kinazları inhibe ettiği (40),
2) Membran lipid peroksidasyonuna neden olduğu (41),
3) Hücresel glutatyon seviyelerinin düşürülmesine ve süperoksit dismutaz
yükselmesi (42,43),
4) Mitokondriyal fonksiyonun bozulmasına (43),
5) Asetil kolinesterazın erimiş globül formunda çapraz bağlanmaların
oluşmasına (44),
6) α-kristalin merceğin fotooksidasyonuna sebep olduğu gösterilmiştir
(45).
C) Hyperisinin Hücre İçinde Bulunduğu Yerler ve Hücresel Hedefleri
Fizyolojik şartlarda monobazik bir tuz olarak bulunan Hy’in hidroksil (OH)
gruplarını taşıdığı kısımlardan kaynaklanan asiditesi de vardır. Hy tuzları (F, Br, Cr)

11
hücresel lipit membran yapıları tarafından tutulur ve bu yüzden lipofilik iyon çifti
gibi davranır (29). Bu hidrofobik karakter sayesinde hyperisin sitoplazma içinde 2-4
saat kalabilir. Yapılan çalışmalar hyperisinin hücre içinde özellikle endoplazmik
retikülüm ve golgi kompleksinde biriktiğini mitokondrinin ise dışında kaldığı ortaya
çıkmıştır. Hyperisinin hücre içi dağılımının hücre tipinden de etkilendiği kolon Caco-
2 hücrelerinde nükleer membranda toplanmış halde bulunmasıyla anlaşılmıştır (15).
Hyperisin’in hücre içine girişinin enerji gerektirmemesi ve reseptör
bağlanmalarının olmamasından difüzyon veya çözünülürlükle olduğu
düşünülmektedir. Hücre içi lokalizasyonundan dolayı da esas hedeflerinin
membranlar olduğu kabul edilmektedir. Membranlar ışık bağlantılı reaktif oksijen
türleri (ROS) üretilen yerler veya hücresel hedeflerin yakın çevresidir (42,37,46).
Nöronlarla yapılan çalışmada, Hy’in çok yüksek konsantrasyonlarda (20
μM) uygulanması halinde nükleus içinde birkaç bölümde gözlenmiştir (47). Nükleer
lümen dışında izole DNA’da ve oligonükleotidlerle interaksiyona girmiş olduğu,
membran hasarına neden olan ROS üretimi ile DNA hasarına da neden olabileceği
belirtilmiştir (48-50).
D) Hyperisinin bilinen etkileri:
1- Antikanser etkisi
2-Antivirütik etkisi
3- Antidepresant etkisi
1- Antikanser etkisi
Hyperisin bilinen en güçlü doğal fotosensitizerdir. Hyperisinin antikanser
etkisi de bu fotosensitizer özelliğinden kaynaklanmaktadır. Fotodinamik terapi
(PDT) kanser tedavisi için kullanılan yeni ve umut verici yöntemlerden biridir. Hücre
ve organellerin yıkımını ışık aracılığı ile katalizleyen fotosensitizer ajanların
kullanılmasına fotodinamik terapi (PDT) denilmektedir. PDT, tümör hücresi
tarafından tutulan bir fotosensitizer ajan ve bu ajanın absorbsiyon spektrumuna uyan
görülebilir bir ışık dalga boyunu gerektirmektedir. Genellikle fotosensitizerler
oksijenin hazır bulunması ve ışık stimülasyonunun olmasıyla peroksit ve hidroksil
radikallerine dönüşen süperoksit radikallerini üreten (tip I reaksiyonu) veya non-

12
radikal tekil oksijen moleküllerini üreten (tip II reaksiyonu) bileşikler olarak
tanımlanırlar (13, 51).
Antitümöral fotodinamik tedavi bir fotosensitizerin sistemik uygulanmasını
ve tümör lezyonuna görülebilir ışığın dağıtılmasını hedefler. Bunlar sağlandığı
zaman tip I veya tip II reaksiyonları vasıtasıyla reaktif oksijen radikallerinin lokal
üretimi lipid membranlarda zincirleme oksidasyon reaksiyonlarını gerçekleştirerek
tümör harabiyetine sebep olur (52).
Reaktif oksijen radikalleri, hücre metabolizması esnasında meydana gelen
biyokimyasal reaksiyonlarla ortaya çıkan, dış yörüngelerinde bir süre için bile olsa
ortaklanmamış elektron taşıyan atom veya moleküller olarak tanımlanırlar (53,54).
Başlıca oksijen radikelleri:
-Süperoksit anyon radikali (O2˙- ): Oksijen molekülüne bir elektron transfer
edilirse süperoksit anyon radikali (52).
-Hidrojen peroksit radikali (H2O2): İki elektron transfer edilirse hidrojen
peroksit radikali oluşur (52).
-Hidroksil radikali (OH.): Hidrojen peroksit radikali demir (Fe) gibi bir
geçiş metal iyonu ile reaksiyona girerse parçalanıp canlı dokular için daha zararlı
olan hidroksil radikaline dönüşür (52).
- Tekil oksijen (O21): Tekil oksijen gerçekte bir serbest radikal değildir,
ancak serbest radikal reaksiyonları sırasında üretilen bir reaktif oksijen türüdür (52).
Serbest oksjen radikallerinin başlıca endojen kaynakları:
1- NADPH oksidaz, lipooksijenaz, prostoglandin sentetaz gibi plazma
membranı enzimleri ve lipid peroksidasyonu,
2- İntoksikasyon, iskemi, travma gibi durumlara bağlı oksidatif stres,
3- Ksantin oksidaz, triptofan oksijenaz gibi enzimler ve hemoglobin,
4- Mitokondrideki ETS,
5- Nükleus ve ER’de bulunan ETS (sit p-450),
6- Peroksizomlarda bulunan enzimler,
7- Yaşlanma (53).
Eksojen serbest oksijen radikali oluşumuna neden olan kaynaklar:

13
1- İyonize ve non –iyonize radyasyon,
2- Isı, güneş ışığı,
3- Alkol uyuşturucu ve anestezik ilaçlar,
4- Stres: Stres ile kanda katekolamin seviyesi yükselmekte ve oksidasyona
uğraması ile de serbest radikal üretimi artmaktadır (54).
Meydana gelen serbest oksijen radikalleri (ROS) yine hücrede süperoksit
dismutaz (SOD), katalaz (KAT), glutatyon peroksidaz (GSH-Px) gibi antioksidan
enzimlerle veya C vit, E vit, transferin, beta- karoten, seruloplazmin, meletonin gibi
enzim olmayan antioksidan maddelerle reaksiyona girerek hücreye zararsız
bileşiklere dönüştürülmektedir. Eğer bir hücrede ROS üretimi yıkımından fazla ise
hücre hasarı var demektir. Hücrelerin membranalarında bulunan poliansetüre yağ
asitleri (PUFA) okside edici serbest radikaller tarafından kolaylıkla
etkilenebilmektedir. PUFA’nın oksidatif hasarı lipid peroksidasyonu olarak
adlandırılır. Lipid peroksidasyonu sonucu hücrede kendiliğinden devam eden zincir
reaksiyonları başlamaktadır (55). Zincir reaksiyonlarının en sonunda üretilen
hidroksialkenoller hücrenin farklı bölgelerine giderek hasar oluşturabilmektedir.
Nükleik asitler ve proteinler serbest radikallere karşı biraz daha dayanıklıdır. Ancak
serbest radikaller DNA’nın çok yakınında meydana gelirse bu molekülü de
kolaylıkla hasara uğratabilir (54).
İn vivo çalışmalarda PDT’yi takiben kan akışında tıkanma, damarsal
kollaps, damar dışına sızıntı gözlenmiştir. PDT, sitokinler ve diğer düzenleyicilerin
salıverilmesine neden olur ve bu da takiben inflamatif bir cevap ortaya çıkar. Çeşitli
tümör hücrelerinin PDT’ye cevabında doku tipi farklılığından ve fotosensitizerin
farklı tutulumundan kaynaklanan farklılıklar vardır (56).
1. PDT ile Hücre Ölüm Mekanizmaları
PDT’nin tümör hücrelerini iki şekilde etkileyip yok ettiği bilinmektedir.
Bunlar: Apipitozis ve nekrozis.

14
1-Apoptozis: Programlı hücre ölümüdür. Biyolojik görevini tamamlamış,
yaşlanmış veya DNA hasarı meydana gelmiş hücrelerin ortada bir iz bırakmadan
yok olmasıdır ( 57).
2-Nekrozis: Fiziksel veya kimyasal bir tahribat sonucu oluşur ve ortamda
bir inflamasyon bırakır (57).
Apopitozisin ekstrinsik veya intrinsik sinyal mekanizmasının işlemesi ile
gerçekleştiği bilinmektedir. Her iki sinyal yolunda da sistein içeren kaspas proteazlar
önemli rol oynamaktadır. Normal halde kaspaslar aktif hallerinden daha uzun bir
zincir içerirler, aktifleşince özel bölgelerinden iki alt birime ayrılırlar. Kaspaslar,
başlatıcılar (initatör) ve uygulayıcılar (efektör) olmak üzere ikiye ayrılmaktadırlar.
Başlatıcı kaspasların (kas-8,9,10) adaptör proteinlere bağlanabilen ölüm zinciri
içeren domainleri vardır. Efektör kaspasların (kas-3,6,7) özel ölüm domainleri yoktur
ve aktifleşince adaptör proteinlerle birleşmezler. Polipolimeraz (PARP), laminler,
topoizomeraz I gibi hücre içi vital substratları parçalarlar (58).
Ekstrinsik yolda mutlaka hücre zarında bulunan Fas, TNFR (tümör nekroz
faktör reseptörü) TRAIL (TNF-related apopitozis induced ligand) gibi ölüm
reseptörlerinin ligandlarınca uyarılması ile sinyal yolu aktiflenir. Uyarılan reseptörün
hücre içinde bulunan ölüm domainine hücre içinde özel adaptör proteinler
bağlanarak aktiflenir ve sonrasında başlatıcı kaspaslar aktifleşir, başlatıcı kaspaslar
efektör kaspasları aktifler ve efektör kaspaslar da vital substratları parçalayarak
apoptozisi gerçekleştirirler (59).
İntrinsik yolda sinyal ya hücrenin genomundan gelir ya da reseptöre
bağlanmaya ihtiyaç göstermeyen ilaç, ışın etkisi ile sağlanır. Eğer DNA hasarı oluştu
ise p53, p21 aracılığı ile siklin/CDK(siklin depended kinaz) inhibisyonu ve mitoz
bloğu sonrasında apopitozis gerçekleşir. İster DNA hasarı olsun ister ilaç etkisi ile
olsun intrinsink yolda proteaz aktivasyonunu başlatan ilk sinyaller mitokondri
üzerinden olmaktadır. Normalde proton ve diğer iyonlar mitokondri iç zarında
elektriksel bir gradient oluşturacak şekilde dağılmıştır. Katyonik lipofilik boyalar
kullanıldığında boya membran içine girerek, membran matriksi boyunca
yayılmaktadır. Apopitozise giden bir hücrede ise transmembran potansiyelinin
düşmesinden dolayı boya girişi azalmaktadır. Bu azalmaya permabiliti transisyon

15
(PT) denilmektedir. Spermin, Bcl-2 gibi proteinler ve Zn (çinko) PT inhibitörü olup
hücreyi apopitozisten korumaktadır (59).
2. Hyperisin’in Neden Olduğu Apoptozis
Hy nedenli apoptozis ilk olarak insan kötü huylu glioma hücrelerinde
tanımlanmıştır. Bu hücreler Hy ve ışığa maruz bırakıldığında DNA fragmentasyonu
ve hızlı bir apoptotik hücre ölümü olduğu gösterilmiştir. Bu olay diğer kanser hücre
türlerinde daha geç meydana gelmiştir. Bu tür Hy aracılı apoptoziste protein kinaz C
(PKC) inhibisyonunun tetikleyici olduğu ileri sürülmüştür (60). Daha sonraki
çalışmalarla Hy etkili apoptozisin sinyalizasyonunda PKC inhibisyonun yeterli
olmadığı ortaya çıkmıştır (61). Apoptozisin gerçekleşmesindeki ekstrinsik ve
intrinsik olmak üzere iki sinyal yolunun da etkili olabileceği, hatta ekstrinsik yol
intrinsik yol arasında bağlantı olan bir yolun da olduğu belirtilmiştir (62). Çeşitli
çevresel uyarılar veya DNA hasarından kaynaklanan uyarılar mitokondri seviyesinde
gelişim programlarını uyararak intrinsik yolu tetikler. Mitokondri iç zarında sit-c ve
diğer apoptotik proteinler sitosole salınır. Sit-c, apaf-1, prokaspas-9, dATP/ATP
apoptozom kompleksinin oluşumu ile kaspas-9 aktive olur. Aktive kaspas-9 da
efektör kaspasların (kas-3,6,7) aktivasyonunu neden olur. Bu kaspaslar da vital
proteinleri parçalayarak klasik apoptotik görünümlerin oluşması sağlar. Gerek
intrinsik gerekse ekstrinsik yolda kaspas proteazlarının aktivasyonu hücreyi
apoptozise sokmaktadır (63).
2- Antiviral Etkileri
Hyperisin ve kimyasal benzeri pseudohyperisin hem laboratuvar hem de
hayvan deneylerinde:
HIV-1 (64), sindbus virüs (18), murin sitomegalo virüs (MCMV) (18),
influenze A, herpes simplex I ve II (65) ve radyasyon lösemi virüsü (rad LV) (66)
gibi virüslerin replikasyonunu inhibe ettiği ve AZT (3-azido-3-deoksitimidin) ve
diğer nükleosid analoglarından farklı bir mekanizma ile bu antiviral etkiyi gösterdiği
ortaya çıkmıştır (18).

16
Hy HIV sinsitiyum oluşumunu bloke (67) etmesine rağmen revers
transkriptaz (RT) aktivitesinde direkt olarak bir değişiklik yapmaz (68).
Hy ile tedavi edilen hücrelerde immatür veya normal olmayan öz yapılar
oluşmaktadır. Bu gag (p24 geni) tarafından kodlanan öncü poliproteinlerin işleyişinin
bloke edildiğini gösterir. Hy’nin ister hücre içinde olsun ister hücre membranına
bağlanmış olsun protein sentezini engelleme yoluyla revers transkriptaz aktivitesini
düşürdüğü sanılmaktadır. gag geni tarafından kodlanan poliproteinlerin klevajı veya
sentezi engellendiğinde viral partiküller oluşamaz. Birleşmiş (assemble) virüs
özyapısı içinde revers transkriptaz muhtemelen proenzim veya inaktif bir enzim
formunu alır. Virüsler tarafından kodlanan proteazlar veya kinazların işe karıştığı
mekanizmalar RT’yi tekrar aktif hale getirebilir. Bu proteaz ve kinazlar
etkilendiğinde RT aktivitesi etkilenebilir. Hyperisin ve pseudohyperisin’nin proteaz
aktivitesini etkilediği sanılmaktadır. Sıra ile değişen proteaz aktivitesi gag tarafından
kodlanan poliproteinlerin sentezi ve klevajının bozulmasına sebep olabilir. Sonuç
olarak immatür viral özyapı oluşabilir (14,69).
Son bulgular bu bileşiğin hem RNA hem de DNA virüslerinden sadece
lipid zarfı olanlara etkili olduğu yönündedir (70,71).
Alternatif olarak Hy viral proteinlere seçici olarak bağlanarak virüs
birleşimi (assemble) için gerekli olan gag ve gag poliproteinlerinin işleyişine engel
olabilir. Bu yüzden Hy RNA’nın viral kapsid içine paketlenmesi sürecinde blok
yapabilir (14). Bazı araştırmacılar Hy’nin gag tarafından kodlanan prekürsör
poliproteinler veya diğer proteinleri bozmak yerine zarflı virüslerle direkt etkileşime
girerek infeksiyöz virüsü lize ettiğini savunmaktadırlar (72).
Özetle:
1- Zarflı virüslerin lipid olan zarf yapısı direkt olarak hyperisin tarafından
parçalanır, antijenik özelliğini kaybeden virüs inaktive olur (72).
2- Hyperisin proteaz aktivitesini etkileyerek gag tarafından poliproteinlerin
klevajlanması ve sentezine engel olarak veya bu ptoteinlere bağlanarak viral
nükleikasidin özyapı içine paketlenmesini engelleyip bozuk veya immatür özyapı
oluşumuna sebep olur (72).

17
3- Antidepresan Etkileri
Hypericum perforatum bitkisi antik çağlardan beri antidepresan özellikleri
için kullanılmaktadır. Eski Yunan kültüründe bu bitkinin özellikle kötü ruhları
kovduğuna inanılırmış (20).
Tarih boyunca insanlar H. perforatumu ruhsal sıkıntı, hafif depresyon,
yorgunluk, bitkinlik tedavisi için kullanmışlardır. Bu bitkinin günümüz tıbbında
bilinçli bir şekilde kullanılması tesadüf değildir. Geçmiş bugüne ışık tutmuştur (20).
Hypericum ekstrakları antidepresif etkilerinin bitkinin hangi bileşeni
tarafından gerçekleştiği tam olarak belli olmamakla birlikte hypersin’nin de bu etkide
önemli rol oynadığı sanılmaktadır (73,74).Hypericum ekstrakları son zamanlarda
dünyanın her yerinde özellikle de Almanya, diğer Avrupa ülkelerinde ve Amerika’da
antidepresan olarak doğal, güvenli ve daha az yan etkiye sahip olduğu için geleneksel
antidepresanlara tercih edilmektedir (75). H. perforatum’dan hazırlanan antidepresan
etkisi kanıtlanmış ekstraklar Tablo 2’de verilmiştir.
Tablo 2: Hypericum perforatum’un antidepresan etkisi kanıtlanmış extraktları.
Extraktlar Kaynaklar
Hyperisin (76)
Total ekstrakt (77)
Total ekstrakt fraksiyon II (77)
Total ekstrakt fraksiyon IIIc (77)
ZE117 (78)
LI160 (79)
Hyperiforce ® (80)
Jarsin 300 ® (81)
® işaretli olanlar ticari ürünlerdir.

18
H. perforatum ekstraktının beyinde etkilediği yerler:
Diensefalon (82)
Serebal korteks (82 )
Prefrontal kortex (83)
Lokus serulus (84)
Hipokappus (85)
Hipotalamus (85)
Etki mekanizması
H. perforatumun serotonin norepinefrin ve dopamin geri salımında blokaj;
serotonerjik ve dopaminerjik reseptörlerin yoğunluğunda bir artma; ayrıca MAO
inhibisyonu gibi geleneksel antidepresanların etkilerine benzer etkiler gösterdiği
düşünülmektedir (86,87).
Araştırılmakta olan birkaç teori olmasına rağmen bir çok depresyon çeşidini
tedavi kabiliyeti olan H. perforatum ’un nasıl işlev gördüğü açık değildir.
Bir teoriye göre H. perforatum monoaminoksidaz (MAO) ve katekolamin
metil transferaz (COMT) enzimlerini inhibe etmektedir (88). MAO kaynağı tiramin
oksidaz olan vücudun değişik yerlerinde ve beyinde sinaptik aralıkta dopamin,
noradrenalin, serotonin, gibi primer ve sekonder aminlerin terminal fazında
aminoalkol grubunun yıkımını sağlayarak nöromedyatör etkinliği baskı altında tutan
mitokondrial bir enzimdir (89).
Diğer bir teori serotonin ve norepinefrin nörotransimiterlerinin seviyesini
yükselmektedir. Bunu SSRI (selective serotonin re-uptake inhibitors)’lar ve SNRI
(selective norepinephrine re-uptake inhibitors)’lar serotonin ve norepinefrinin geri
salınımını engelleyerek sinaptik aralıktaki etki süresi ve sayısının artışını sağlayarak
yaptığı ileri sürülmüştür (90).
Bir diğerine göre ise H. perforatum ekstraktı stres hormonu kortizolün
seviyesini düşürür veya beyindeki gama amino bütirik asit (GABA) reseptörlerini
etkiler. GABA beyin ana inhibitör mediyatörüdür (91).
19
Bahsedilen teorilerden sonra bu etkileri yapan bileşiklerinin hangisi veya
hangileri olduğu sorusu akla gelmektedir ki bu konuda çelişkili raporlar mevcuttur
(92). Çalışmalar sürmektedir.
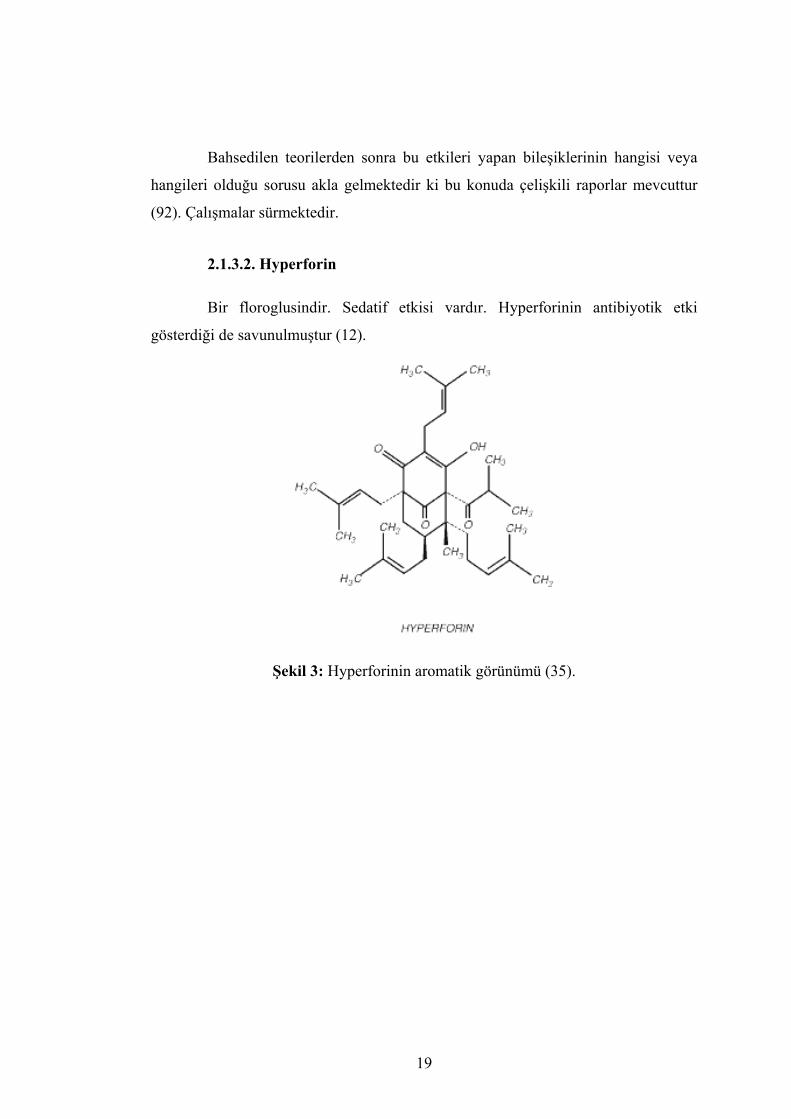
2.1.3.2. Hyperforin
Bir floroglusindir. Sedatif etkisi vardır. Hyperforinin antibiyotik etki
gösterdiği de savunulmuştur (12).
Şekil 3: Hyperforinin aromatik görünümü (35).

20
2.2. Kardeş Kromatid Değişimi (KKD)
Kardeş kromatid değişimi (KKD), DNA replikasyonu sırasında kardeş
kromatidler arasında özdeş segmentlerin karşılıklı simetrik yer değiştirmesidir (93).
Bu olay kromozomun her iki kromatidinin birden kırılması ile ortaya çıkmaktadır
(94). KKD’de yeni duplike olmuş kromatid ve eski kendi kardeş kromatidi arasında,
kromozom morfolojisi değişmeden simetrik olarak özdeş segmentlerin değişimi söz
konusudur. KKD memelilerde hücre bölünmesinin normal bir özelliği olarak spontan
meydana gelebilir (95). Her kırık sonrasında devreye giren tamir mekanizmaları,
zaman zaman iki homolog parçanın karşılıklı yer değiştirerek kardeş kromatide
yeniden bağlanması ile sonuçlanmaktadır. Bu değişim sonucunda kromatidlerin
morfolojisinde bir değişiklik ortaya çıkmamaktadır. Olayın moleküler mekanizması
ve biyolojik anlamı tam olarak bilinmemektedir (94).
Günümüzde artık insanların günlük yaşamlarında ve çalışma ortamlarında
genotoksik ajanlara mutajenik ve karsinojenik maddelere maruz kalmadan
yaşamaları mümkün olmamaktadır. Gelişen teknoloji ile üretilen yeni kimyasal
maddeler, ilaçlar, gıda katkı maddeleri, çevreye verilen atıklar, tarımda kullanılan
kimyasal maddeler ve manyetik alan oluşturan elektronik cihazlar, canlıların genetik
yapısında mutasyon oluşturma ihtimali taşımaktadırlar. DNA oluşan hasarı
kromozom düzeyinde tespit etmemizi sağlayan doğrudan metodların birisi de kardeş
kromatid değişimi (KKD) analizidir (24). KKD, DNA’daki çok küçük harabiyetlerin
bile hassas göstergesi olarak kabul edilir. DNA hasarının ve indüklenmiş DNA
tamirinin gösterilmesinde en basit, duyarlı ve kısa zamanda sonuç veren bir yöntem
olarak kullanılmaktadır (25). Vücuttaki total DNA’nın %1’inden daha az bir
bölümdeki kayıp bile KKD ile rahatlıkla tespit edilebilir (93).
Bir çok mitojen ve karsinojen çalışmasında KKD, güçlü bir indikatör tekniği
olarak hizmet etmektedir. Bazı kimyasallarda aynı çalışmada KKD ve KA
(kromozon aberasyon testi) yöntemleri birlikte yapıldığında, KKD yönteminin KA
yönteminden daha fazla hassas olduğu gösterilmiştir (93).
Yapılan çalışmlarda, Bloom sendromu, Ataksi telenjaktazi, Werner
sendromu, Kseroderme pigmentozum, Fanconi anemisi, Cockayne sendromu,

21
Duchenne ve Becker tipi kas distrofilerinde ve down sendromunda KKD değerleri
anlamlı derecede artmış olduğu rapor edilmiştir (96).
2.2.1. KKD’nin Tarihçesi
Mc Clintock 1938-1957 yılları arsında mısır bitkisinin monosentrik halka
kromozomlarının kaybolurken, disentrik halka kromozomlarının oluşumunun
arttığını gözlemiştir (97).
Kardeş kromatid değişimi ilk kez 1957 yılında Taylor ve arkadaşları bitki
mitotik kromozomlarında (Vicia faba ve Bellenalia romana) DNA replikasyonu ve
reğresyonu konusunda yaptığı otoradyografik çalışmalarla gözlemlemiştir (25).
Zakharov ve Egolina 1972’de Chinese Hamster hücrelerinde uzun süreli
kültür kurmuş timin analoğu olan 5-Bromo-2 deoxiuridine (BrdU) ile replike olan
kromozomların morfolojisini incelemiştir (98).
1973 yılında Latt DNA çift sarmalına BrdU bağlandığında, Bisbenzamid
(Hoecst 33258) boyasının fluoresansını azalttığını (99), Perry ve Wolf, 1974 yılında
BrdU’nin kromatin yapısına Giemsa boyasının girişini kısıtladığını göstermişlerdir.
Bu bulgular ışığında yeni bir teknik geliştirerek (Fluoresans boya + Giemsa tekniği=
FPG), otoradyografiye gerek kalmaksızın gösterilebilmişlerdir (97).
Latt’ın 1974’te mitomisin-C’nin KKD frekansında artışlara neden olduğu
(100) göstermesinden sonra günümüze kadar, KKD genotoksisitenin sensitif bir
indikatörü olarak kabul edilmekte ve çalışmalarda yaygın olarak kullanılmaktadır.
2.2.2. KKD’nin Kullanım Alanları
KKD analizi hem laboratuar hayvanlarında hem de insan populasyonlarında
çalışılabilir. KKD analizi, kültürde replikasyon yapabilen, filogenetik spektrumu
Drozofiladan insana kadar uzanan her hücre hattında yapılabilir. Çin Hamster
fibroblast hatları, Hela gibi insan hücre hatları, deri ve akciğerden alınmış insan
hücreleri, fare, sıçan, tavşan gibi laboratuar hayvanlarının ve insanların lenfosit
kültürleri yaygın olarak çalışılmaktadır. İnsan ve fare amniyotik sıvısında da
çalışılmıştır. Bitki, böcek, balık ve memelilere kadar in vivo KKD indüklenmesi hem

22
fiziksel hem de kimyasal ajanların değerlendirilmesinde kullanılmaktadır (96). KKD,
bireylerde somatik hücrelerde bilinen ya da potansiyel mutajen ve karsinojenlere
maruz kalmanın etkilerini saptamada da kullanılanır (25).
KKD analizi, daha çok çeşitli kimyasalların genotoksik etkilerini
araştırmakta kullanılmakla birlikte, kromozom instabilitesi ile seyreden Bloom
Sendromu ve Fankoni Aplastik Anemisi gibi bazı hastalıklarda da araştırma ve tanı
amaçlı olarak kullanılmaktadır. İn vivo kardeş kromatid değişiminin klinik açıdan
önemini belirten çalışmalarda dengesiz kardeş kromatid değişimi, Duchenne ve
Becker tipi kas distrofilerinin kalıtımında rol oynadığı ortaya çıkmıştır. Bu tür
değişikliklerin duplikasyonlara yol açarak bazı hastalıkların beklenenden farklı
kalıtım yolları izlemesine neden olabileceği bildirilmektedir (94).
2.2.3. KKD Mekanizmaları
Kardeş kromatid değişimi çoğalan hücrelerde spontan meydana gelebilir.
Çeşitli fiziksel ve kimyasal etkenlerle kromozom DNA’sında meydana gelen,
replikasyon esnasında onarılamayan hatalar KKD’nin ortaya çıkmasını ya da
artmasını sağlar. Yani DNA’nın hasarına neden olan pek çok ajanın KKD sıklığını
artırdığı bilinmektedir. Bu nedenle pek çok mutajenik yada karsinojenik etkileri
göstermede duyarlı bir parametre olarak kabul edilir (25).
KKD oluş mekanizması üzerine çeşitli modeller ileri sürülmekle birlikte
KKD’nin moleküler mekanizması henüz tam olarak bilinmemektedir. Şekil 4’de bir
kromozomda oluşabilecek KKD’ler görülmektedir.

23
Şekil 4: Kromozomlarda oluşabilecek KKD’lerin şematik gösterimi (24).
a) KKD oluşumu yok. BrdU iki tam döngü veya iki tam döngünün ilkinde
DNA sentezine girmiştir.
b) Tek KKD oluşumu (→)
c) İki KKD oluşumu
d) Üç KKD oluşumu
e) Sentromerde oluşan KKD
f) Boya kalıntısı veya BrdU’nun DNA ile tam birleşmemesi KKD olarak
sayılmaz.
g-h) DNA sentezi sırasında BrdU’nun iki replikasyon döngüsünden fazla
DNA’ya girmesi, KKD olarak değerlendirilmez.
i) Bir kromatin genişliğinden daha dar olan boyanma KKD olarak
değerlendirilebilir (24).

24
2.2.4. KKD Oluş Mekanizması ile İlgili İleri Sürülen Modeller
a) Taylor modeli: 1957’de Taylor’un çalışmalarında kromozomların
radyoaktif timidin (3 HdTh=3H-deoksi-timidin) varlığında bir defa kendilerini
eşlemelerine ve sonraki replikasyonda da izotopun yokluğunda da kendilerini
eşlemelerine izin verilmiştir. Otoradyografide DNA’nın semikonservatif eşlenmesi
sonucunda, her bir kromozomun sadece bir kromatidinin işaretlendiği görülmüştür.
Bu işaretleme sonucunda kardeş kromatidler arasında simetrik değişmeler gözlenmiş
ve Taylor bunları “kardeş kromatid değişimi”olarak adlandırmıştır (24)(Şekil 5).
Şekil 5: KKD oluşum mekanizması (24).
b) Zakharov-Egolina modeli: Zakharov ve Egolina adındaki araştırmacılar
Çin Hamster hücrelerinde, timinin analoğu olan 5- bromo-2 deoksi üridin kullanarak
kromozom yapısını incelemişler. BrdU’nin yeni sentez edilen DNA’ya girişi
uygulanışını takip eden ilk S sırasında olmaktadır. Replikasyon sırasında timinin
yerine BrdU’ni alan zincir, diğer DNA sırasında replike olur. Timinin analoğu olan
BrdU, replikasyon sırasında DNA’ya girer timinindeki metil grubunun yerini brom
atomunun almasıyla DNA molekülünde oluşan bir seri fiziko-kimyasal değişikler
sonucu, ikincil grupların şekillenmesinde güçlüklere sebep olur (98) (Şekil 6).
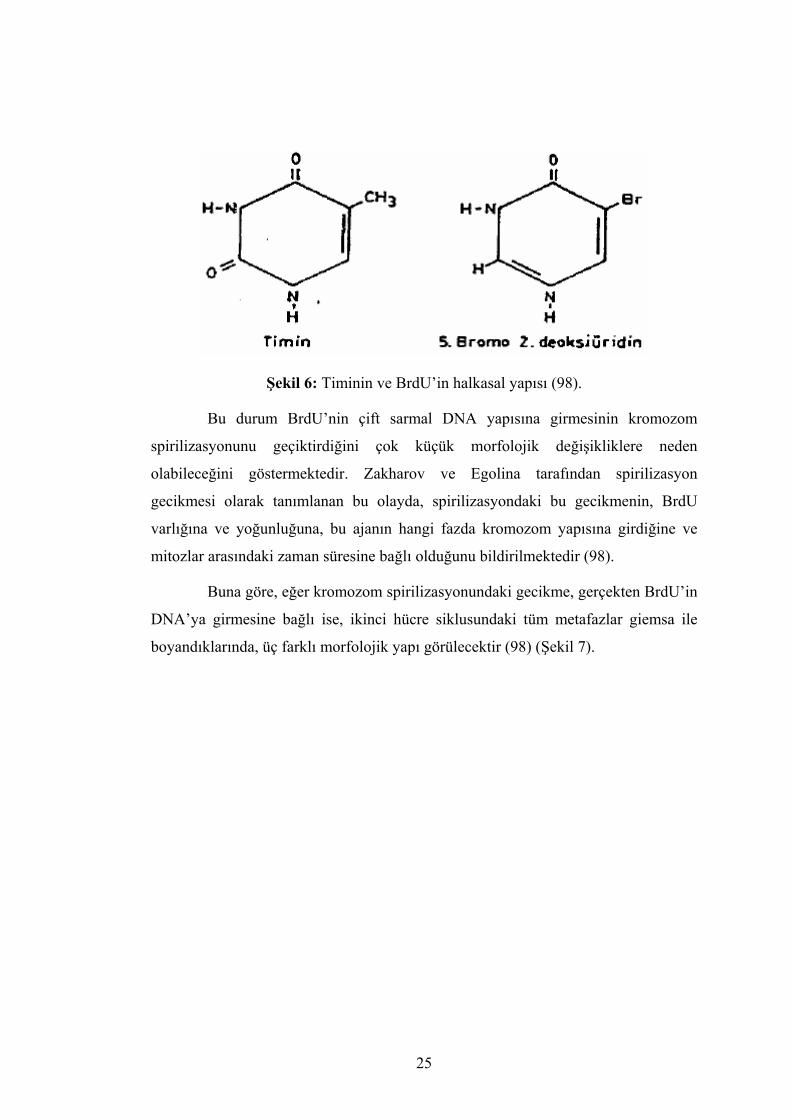
25
Şekil 6: Timinin ve BrdU’in halkasal yapısı (98).
Bu durum BrdU’nin çift sarmal DNA yapısına girmesinin kromozom
spirilizasyonunu geçiktirdiğini çok küçük morfolojik değişikliklere neden
olabileceğini göstermektedir. Zakharov ve Egolina tarafından spirilizasyon
gecikmesi olarak tanımlanan bu olayda, spirilizasyondaki bu gecikmenin, BrdU
varlığına ve yoğunluğuna, bu ajanın hangi fazda kromozom yapısına girdiğine ve
mitozlar arasındaki zaman süresine bağlı olduğunu bildirilmektedir (98).
Buna göre, eğer kromozom spirilizasyonundaki gecikme, gerçekten BrdU’in
DNA’ya girmesine bağlı ise, ikinci hücre siklusundaki tüm metafazlar giemsa ile
boyandıklarında, üç farklı morfolojik yapı görülecektir (98) (Şekil 7).

26
Şekil 7: BrdU varlığına bağlı spirilizasyon gecikmesi (98).
Her iki replikasyonda veya mitozda ortamda BrdU bulunması halinde
kromatidlerden birinin, her iki DNA zincirinde BrdU bulunur, diğer kromatidinin ise
yalnızca bir zincirinde BrdU, diğerinde timinin yer alır ve her iki kromatidde de
spirilizasyon gecikmesi gözlenir. Yalnız, iki DNA zincirinde de BrdU bulunduran
kromatidde spirilizasyon gecikmesi daha fazla olacağından diğerinden daha uzun ve
daha soluk boyanacaktır (Şekil 7a). Yalnız ikinci replikasyon ortamında BrdU
bulunması durumunda kromozomun her iki kromotidinde eş zamanlı spirilizasyon
gecikmesi olacak, ama kromozomlar aynı boyanacaktır (Şekil 7b). Her iki
replikasyon siklusunda ortamda BrdU bulunmaması veya sadece birinci replikasyon
siklusunda ortamda Brdu bulunması durumunda spirilizasyon gecikmesi görülmez,
morfolojik değişim oluşmaz ve normal boyanır (Şekil 7 d-e) (98,100).
Kromozom morfolojisindeki bu farklılıklar DNA replikasyonunda farklılığa
ve belki de genetik aktivitede değişikliklere neden olabilir (92). Kromatidlerdeki bu
uzunluk farkı, protein ile BrdU içeren DNA arasındaki etkileşimin farklı olması
nedeniyle ortaya çıkmaktadır. Proteinler kromozomların kondenzasyonda ve
spirilizasyonunda etkili olmaktadır. Proteinler BrdU içermeyen DNA’dan daha sıkı
bağlanmakta ve kromozomların spirilizasyonunu ve yoğunlaşmasını

27
zorlaştırmaktadır. BrdU’nin kromozomlardaki esas etkisinin paketlenme esansında
olabileceği de belirtilmiştir (101).
c) Evens isimli araştırıcının öne sürdüğü KKD oluş modeli: Yalnızca
replikasyon süresince mevcut etkenlerin KKD’leri ortaya çıkarabildiğini, bu nedenle
değiş tokuşun replikasyon çentiğinden başlaması gerektiği ileri sürülmüştür. KKD
oluşumunun bir kromatid üzerinden yeni sentezlenmiş zincir ile onun kardeşinin
atasal zinciri arasında değiş-tokuşu içerdiğini, bu esnada zincirlerden birinde veya
her ikisinde kırığın gerçekleştiğini öne süren bir modeldir (25)(Şekil 8).
Şekil 8: Evens isimli araştırıcının ileri sürdüğü KKD oluş mekanizması (25).
1- G1 evresinde timin yerine BrdU içeren DNA ve S evresinde sentezlenen
DNA’da herhangi bir etkenle hata oluşması.
2- 3- BrdU varlığında S evresinde ikinci replikasyon devam ederken hatalı
bölgeye gelindiğinde bir veya her iki zincirde kırılmalar meydana gelmektedir.
4- Hatanın çıkarılması, ancak kırık zincirlerin birbiri ile değil, kardeşleri ile
birleşmeleri sonucunda zincirler arasında değiş tokuş meydana gelmesi.
5- Replikasyon tamamlandığında molekülde, her iki zincirde de timin ve
BrdU içeren bölgeler ortaya çıkması.
6- Her iki zincirde de BrdU içeren bölgelerin soluk, timin içeren bölgelerin
koyu boyanması ve oluşan KKD’lerin gözlenmesi (102).

28
d) Loveday ve Latt isimli araştırmacıların ileri sürdüğü model: 1981
yılında Loveday ve Latt isimli araştrıcılar Potter ve Dressler’in önerdiği modelden
yola çıkarak yeni bir model öne sürmüşlerdir. Bu modele göre kalıp DNA’dan
komplementer olarak timin yerine BrdU’in girdiği tamamlayıcı DNA zinciri
sentezlenir. Her bir çift sarmalda tek zincir kırığı oluşur ve bir dupleksteki bir zincir
ile diğer dupleksteki DNA’nın tamamlayıcısı, kardeş zinciri arasında krosingover
gerçekleşir. Rekombinasyon sonucu hem timin, hem de BrdU’in yer aldığı
heterodupleks zincirler oluşur. DNA’nın krosingover noktası etrafındaki rotasyonu
ile X formu meydana gelirken, kırıklar oluşur ve kırılan zincirler birbiri ile değil,
kardeş zincirlerin parçaları ile DNA ligaz ile birleştirilerek her iki zincirde BrdU ve
her iki zincirde timinin içeren 2 rekombinant yapı oluşur. Her iki zincirde BrdU
içeren bölgeler soluk, her iki zincirin de timin veya bir zincirinde BrdU, diğerinde
timin içeren bölgelerin koyu boyanması ile KKD değerlendirilir (99) (Şekil 9).

29
Şekil 9: KKD oluş mekanizması (99).
2.2.5. KKD Yöntemini Etkileyen Faktörler
1- Biyolojik Faktörler
Başta genetik faktörler olmak üzere yaş, cinsiyet, hastalıklar, diyet, ilaç
kullanımı, alkol ve sigara alışkanlığının KKD sıklığını etkilediği belirtilmektedir
(101-103). Sigara kullanımının KKD sıklığını artırdığı yapılan bir çok çalışmayla
kanıtlanmıştır (104).

30
2- Kültür Koşulları ile İlgili Faktörler
a) BrdU konsantrasyonu: BrdU toksik sınırlar dışında yüksek dozda hücre
kültürüne eklenirse, mitotik indekste azalmalara ve KKD frekansında umulandan
fazla artışlara neden olmaktadır. BrdU konsantrasyonu %10 artırıldığında KKD
frekansının %50 artığı gösterilmiştir (102).
b) Sıcaklık: Kültür ortamının optimum sıcaklık 37 oC olup 36-38 oC
aralığına tekabül eder. Sıcaklığın geçiçi düşmesi hücreleri öldürmez gelişimlerini
duraklatıp hasat zamanını geçiktirirken sıcaklık 39 oC’nin üzerine çıkarsa hücreler
ölür (94).
c) Besiyeri: Farklı besiyerinde üretilen insan lenfositlerinin KKD sıklığının
farklı olduğu gözlenmiştir (102).
d) Antibiyotik: Kültür ortamında antibiyotik kullanılması KKD sıklığını
azaltır (102).
e) Serum: Besiyerine konulan serumun konsantrasyonu KKD oluşumunu
etkiler (102).
f) Mitoz Uyarıcı: Günümüzde in vitro koşullarda kullanılan mitotik ajanla,
TTP (tüberkülin pürifiye protein), tetenoz ekstresi, konkavanalin A, PWM
(prokeweed mitojen) ve PHA (fitohemaglutinin)’dır. PHA, barbunya fasulyesi
(phaseolus vrilganis) ekstrası olan bir mukopolisakkarittir. En çok kullanılan
PHA’nın işlergesi bilinmemekte olup indüksiyon sürecinin ilk 24 saatinde gereklidir.
Kültürde ilk 24 saatten sonra RNA sentezini artırır. İkinci 24 saat süresince çekirdek
genişlemekte ve DNA sentezi başlamaktadır (94).
g) Kültür Süresi: İlk mitoz 48 saatte gözlenir. Her 24 saatte bir mitoz dalgası
gerçekleşir. Bu nedenle hasat 72. saatte veye 96. saatte yapılır. Rutin amaçlı
kültürlerde önerilen süre 72 saattir (94).
h) Karanlık Ortam: Hücrelerin kültüre edildiği sürece flourans ışığa maruz
kalmaları BrdU içeren DNA’nın fotolizine neden olarak hareketli alkali gruplarının
oluşumuna neden olur. Bu oluşum guanin ile reaksiyona girerek DNA’nın depürine
olmasına ve bu da tek sarmalda kırılmalara neden olarak KKD’nin oluşumunu

31
indükler. Bu neden KKD analizi için kullanılacak kültürlerin ışıktan koruması
gerekir. (94)
i) Kolşisin Konsantrasyonu: Mitozu metafazda durdurmak için mekik
inhibitörleri kullanılır. Kolşisin de bunlardan biridir ve 10 μg/ml olacak şekilde
hazırlanır. Kolşisinde bırakma süresi ile metafaz indeksinin sayısı birbiri ile
orantılıdır. Fakat uzun süre bırakmalarda kromozom boylarında kısalma gözlenir. Bu
nedenle süre iyi ayarlanmalıdır (94).
j) Hipotonik solüsyon: Hipotonik çözeltilerin yoğunluğu hücre plazma
yoğunluğundan az olduğundan hücreler içine su alır, şişer ve eritrositlerin çoğu
patlar. Hipotonik uygulaması kritik bir zamanı gerektirir. Kolşisinin etkisini
sonlandırıp kromozomların dağılmasını sağlar. Hipotonik çözelti olarak % 1’lik
sodyum sitrat veya 0,075 M KCl kullanılabilir. Kromozomda en az hasar yaptığından
en çok kullanılan hipotonik çözelti KCl’dir (94).
k) Fiksasyon: Fiksasyonda 3:1 alkol ve glasiyal asetik asit oranı önemlidir.
Asetik asitin fazla olması hücrelerin erken yırtılmasına neden olur. Bu da kromozom
kayıplarına yol açar. Kan kültürlerinde ilk fiksasyonda karıştırma çok önemlidir,
kalan eritrositler hemoliz olur. Kırmızı renkli hemoglobin koyu kahve renkli
hematine dönüşür. Yeterli karıştırma yapılmazsa koyu kahve renkli çökeltiler
gözlenirken mitotik indeks düşer ve kalitesiz preperatlar elde edilir (94).

32
3. MATERYAL VE METOD
3.1. Materyal
3.1.1. Kimyasallar
Fitohemaglutinin (Biologcal Industries BI12-006-1H), Fetal calf serum
(Biologcal Industries BI04-001-1B), Leguamine 25.000IU (Roche HEP-5), RPMI
1640 Medium (Biologcal IndustriesB101-106-1B), Bis Benzimide 15090 (Serva
II33258), Giemsa (Merck 109204), Potasyum dihydrogen phosphate (KH2PO4,
Merck 104873), Di-sodium hydrogen phosphate anhydrous (Na2HPO4.2H2O,
Merck 106586), Colchicine (Serva), Penisilin/Streptomisin (Irvine Scientific), L-
Glutamin (Irvine Scientific ), 5-Bromo-2 deoxyuridine (Sigma), Asetic asit glacial
(Merck), Methanol (Merck), Potasyum chloride (KCl) (Merck), PBS tablets
(phosphate buffered saline, amresco E404-100TABS), Hypericin 1 mg (Sigma),
Dimetilsülfoksit (Sigma).
3.1.2. Diğer Malzemeler
Kapaklı falkon tüp (greiner), cam pastör pipeti 3 ml (Isolab), lam (rodajlı
Superior marienfeld ), plastik pastör pipet 3ml’lik (disposebl), 5cc’lik enjektör,
plastik pipet, şale, balon joje, beher, etüv, benmari, santrifüj, vortex.
3.1.3. Deney Gruplarının Oluşturulması
Deney, Süleyman Demirel Üniversitesi Tıbbi Biyoloji Sitogenetik
laboratuvarında yürütülmüştür. Kan örnekleri 10 (5 bayan ve 5 bay) gönüllü, sağlıklı,
sigara kullanmayan bireyden alındı. Birbirine bağlı üç grup oluşturuldu: Grup 1
(kontrol grubu), grup 2 (5μM hyperisin), grup 3 (10 μM hyperisin) (33). Her bireye
ait kan örneğinden 3 ekim yapıldı.

33
3.2. Metod
3.2.1. Hücre Kültürlerinin Kurulması
5 cc’lik heparinli enjektörlere yaklaşık 5 cc periferel kan örnekleri alındı.
Besiyeri hazırlanması:
100 ml RPMI 1640 Medium (Biological Industries,BİO1-106-1B)
2 ml L-glutamin (Biological Industries,BİO4-001-1B)
2 ml penisilin-streptomisin (Biological Industries,BİO12-006-1H)
3,3 ml fitohemaglutinin (Biological Industries,BİO3-031-1B)
30 ml fetal bovin serum (Biological Industries,BİO3-020-1B) karıştırılır.
Fitohemaglutinin hazırlanması:
Ticari olarak 5 ml gelen fitohemaglutinin 5 ml RPMI 1640 Medium ile
sulandırıldı.
Fetal bovin serumun hazırlanması:
Fetal bovin serum çözdürüldükten sonra 65 oC’de 1 saat inaktive edildi.
Hyperisinin çözülmesi:
1 mg’lık toz Hyperisin (Sigma), 2 ml DMSO (dimetilsülfoksit) ile ışık
geçirmeyen kendi ambalajında çözüldü (105). Sonra distile-noniyonize su ile 10
ml’ye tamamlandı. Işık geçirmeyen cam şişe içersinde –20 oC’de saklandı.
5-Bromo-2 deoxyuridine (BrdU) solüsyonunun hazırlanması:
15,37 mg BrdU (Sigma, B5002) 10 ml distile su ile çözüldü, filtre edildi.
Steril ağzı kapaklı bir tüp içersinde üzeri aluminyum folyo il sarılıp +4 oC’de stok
olarak saklandı.

34
İşlemler:
Hazırlanan besiyerinden plastik ağzı kapaklı konik santrifüj tüpüne kontrol
için 9 ml üzerine 1 ml kan ilave edildi. Deney grularında da aynı işlem yapılıp
hyperisin dozlarının da eklenmesi ile toplam hacim 10 ml’ye tamamlandı.
Bütün tüpler alt-üst edildikten sonra 0,1 ml 5-Bromo-2 deoxyuridine (BrdU)
stok solüsyonu eklendi. Tüplerin üzeri aliminyum folyo ile kapatıldı. Tüpler yavaş
vortekslendi. Tüpler 37 oC’ye ayarlanmış etüve 45 derecelik açı ile kondu. Her gün
sabah tüpler alt üst edildi. 70. saatte tüpler etüvde çıkarılıp 2 damla kolşisin (2’lik
enjektörle) damlatıldı. 1 saat 10 dakika 37 oC ye ayarlanmış etüvde bekletildi.
3.2.2. Kromozom Eldesi
Hipotonik solüsyonun hazırlanması:
0,075 M KCl (Merck 4935) olacak şekilde 0,56 gr tartılır distile su ile 100
ml’ye tamamlanır. Kullanımdan en az 1 saat önce hazırlanır ve 37 oC de bekletilir.
Fiksatifin hazırlanması:
Bir kısım glasial asetik asit 3 kısım metanol (30 ml GAA+90 ml metanol)
karıştırıldı.
Lamların hazırlanışı:
Lamlar önce distile sudan geçirilip şaleye yerleştirilir, yarıya kadar distile su
ile doldurulup üstüne alkol ile tamamlanır. Kullanılacağı gün sabah alkol boşaltılıp
distile su konur. Uygulama sırasında buzlu şale içersinden alınarak kullanılır.
İşlemler:
Kromozom preperasyonu için modifiye Moorheard ve arkadaşlarının tekniği
uygulandı (105). Süre bitiminde tüpler 1200 rpm de 10 dak. santrifüj edildi.
Süpernatan kısım atıldı. Sonra tüpler bir taraftan vortekslenirken bir yandan da
hipotonik yavaş yavaş 6 ml çigisine kadar üzerine ilave edildi. Biraz daha
vortekslenip 37 oC ye ayarlanmış etüvde 1 saat bekletildi.
Süre sonunda tüpler 1200 rpm de 10 dak. santrüfüj edildi, süpernatan kısmı
1,5-2 ml kalıncaya kadar atıldıktan sonra tüpler vortekslendi ve vortekslenirken
fiksatif pipet ile tüpün kenarından ilave edildi. İlk uygulamada tüpte bir renk

35
değişimi gözlendi (eritrositler patlayıp hemoglobin açığa çıktığı için) koyu
kahverengi siyah bir görümün aldı. Fiksatif 6 çizgisine kadar ilave edildi. İlk fiksatif
uygulamasından sonra tüpler –18 oC de bekletildi. Diğer fiksatif uygulamalarında
bekletmeye gerek yoktur. Tüplerdeki renk berrak oluncaya kadar önce santrifüj sonra
fiksatif uygulandı. Genellikle 3 –4 uygulamadan sonra istenilen renk elde edildi. Son
fiksasyondan sonra santrifüj edilip süpernatan kısım atılarak 1,5-2 ml bırakıldı ve
cam pastör pipetlerle birkaç kez pipetaj yapıldı. En az bir gün önceden şale içine
hazırlanıp -18 oC’ye konulan lamlar buzlu şale içersinden çıkarıldı (lamların soğuk
olması gerekir). Pipetlere çekilen homojenat önce 45 derece eğimli tutulan lama
mümkün olan en yüksek mesafeden damlatıldı. 15 damladan sonra bu eğim kaldırıldı
lam düz tutularak 15 damla daha damlatıldı. Aynı şekilde her bir örnekten paralel
ikinci lam da yayıldı. Yayılan lamlara isim ve tarih yazılarak ya 24 saat 65 oC’de ya
da 3 gün 37 oC’de bekletilerek yaşlandırıldı.
3.2.3. Kardeş Kromatid Değişimi Boyama Tekniği
Korenberg ve arkadaşlarının önerdikleri modifiye boyama yöntemi
kullanıldı (106).
Kullanılan Solüsyonlar:
2xSSC solüsyonu:
0,87 gr sodyum sitrat (0,03 M Mecrk,104873) ve 0,44 gr sodyum klorür
(0,3 M Merck,106404) 100 ml bidistile suda çözüldü .
PBS solüsyonu:
5 adet PBS tablet ( amresco AIE 404-100)) 500 ml distile suda çözüldü. 100
ml ,100 ml ve 99 ml şeklinde şalelere hazırlandı geri kalan da balon jojede bırakıldı
(UV’de kullanmak için).
Bis Benzimide (hoechst) stok solüsyonu: 0,5 mg Bis Benzimide (Serva,
15090) tartıldı ve 20 ml deiyonize suda çözüldü +4 oC’de saklandı.

36
Söranson tamponu:
A solüsyonu: Potasyum dihydrogen phosphate (KH2PO4, Merck 104873) 4,
537 gr 500 ml distile suda çözüldü.
B solüsyonu: Di-sodium hydrogen phosphate anhydrous (Na2HPO4.2H2O,
Merck 106586) 5,936 gr 500 ml distile suda çözüldü. Balon jojeye A solüsyonundan
alınarak pH 6,8’e gelinceye kadar B solüsyonu eklendi.
Giemza boya solüsyonu:
3 ml Giemsa (Merck 109204) + 97 ml Söranson tamponu 100 ml’ye
tamamlandı.
İşlemler:
Preperatlar,
- 100 ml PBS solüsyonunda 5 dakika bekletildi.
- 9 ml PBS solüsyonu + 1 ml Bis Benzimide’de 20 dakika bekletildi.
- 100 ml PBS solüsyonunda çalkalandı.
- preperatlar tepsiye dizildi geri kalan PBS üzerine döküldü ve UV altıda 25
dakika bekletildi.
- Distile suda çalkalandı.
- 2xSSC solüsyonunda (benmaride) 15 dakika bekletildi.
- Distile suda çalkalandı.
- Giemza boya solüsyonunda 20 dakika bekletilerek boyandı.
- Distile suda çalkalandı.
- Son olarak preperatlar kurutma kağıdı ile kurutuldu. Mikroskopta
immersiyon yağı ile 100’lük objektifle incelendi.
3.2.4. KKD’nin Değerlendirilmesi
Her birey için iyi dağılmış 20 metafazdaki KKD oranları 100X’lik
mikroskop (Olimpus) altında incelendi. Birli, ikili, üçlü ve dörtlü değişimlerdeki

37
kırık noktaları gözlendi, her bir metafazdaki toplam değişim değeri belirlendi ve her
birey için 20 metafaz değerlendirilerek değişimlerin ortalaması alındı.
3.2.5. İstatistiksel İşlemler
Kontrol grubu ve vaka gruplarının her bir bireyi için KKD ortalamaları
alındı. Kontrol grubu ile vaka grupları KKD değerleri Paired-t Testi kullanılarak,
istatistik açıdan değerlendirildi. İstatistik analizler SPSS for Windows istatistik
programı ile bilgisayarda yapıldı. İstatistik analizlerde p<0,05 anlamlı kabul edildi.
Değerler, ortalama ± standart sapma olarak verildi.
3.2.6. Fotoğrafik İşlemler
Tezde üç gruba ait önreklerden kaliteli olan birkaç mefazın 100X objektifle
immersiyon yağı altında Zeis marka mikroskop ve Canon marka digital fotograf
makinesi kullanılarak fotografları çekildi.

38
4. BULGULAR
Türkiye genelinde yaygın olarak yetişen özellikle de Isparta yöresinde
kantaron otu olarak tanınan bu bitkinin çiçeğinin zeytinyağı içerisine katılarak
güneşte bekletilip elde edilen karışımın folklorik olarak haricen yara iyileştirici,
dahilen sindirim sistemi hastalıkları için kullanıldığı bilinmektedir. Bu çalışmamızda,
H. perforatum bitkisinin aktif maddesi olan hyperisinin insan lenfosit kültürlerinde
KKD sıklığına olan etkisi araştırıldı.
Gönüllü ve sağlıklı (son 6 ay içinde radyasyon almamış, herhangi bir
sistemik hastalığı olmayan, herhangi bir ilaç, sigara ve alkol kullanmayan) yaşları
23-30 arasında kadınlarda ortalama 26.8 ± 2.9, erkeklerde 25.4 ± 2.5 olan 5 erkek
5 kadın toplam 10 kişiden alınan kan örnekleri her grup için bir adet olmak üzere 3
ekim yapıldı. Birici grup kontrol grubu olarak seçildi. İkinci ve üçüncü gruba
hyperisinin’nin farklı iki dozu (5 μM/L ve 10 μM/L) uygulanarak KKD sıklığı
değişimi araştırıldı.
Çalışılan gruplara ait preperatlarda ortalama 20 metafazda sayım
yapılmıştır. Her olgu için metafazlarda 1’li, 2’li, 3’lü ve 4’lü kırıklar sayılmış KKD
değeri olarak kaydedilmiştir.
Grup I için KKD ortalama 6.5 ± 1.2 , grup II ortalaması 7.2 ± 0.9 grup III
için 8.2 ± 0.4 bulunmuştur (Tablo 3). Grup I ile grup II arasındaki fark istatistiksel
olarak anlamsız bulunurken grup I ile grup III arasındaki arasındaki fark Paired-t
testine göre anlamlı bulunmuştur (p<0,05).
Tablo 3: Gruplar arasındaki KKD değerleri
Gruplar
N KKD ortalama ±ss
Grup I (kntrol grubu) 10 6.5 ± 1.2 Grup II (5 μM/L Hy) 10 7.2 ± 0.9 Grup III (10 μM/L Hy) 10 8.2 ±0.4
ss: standart sapma N: Birey sayısı
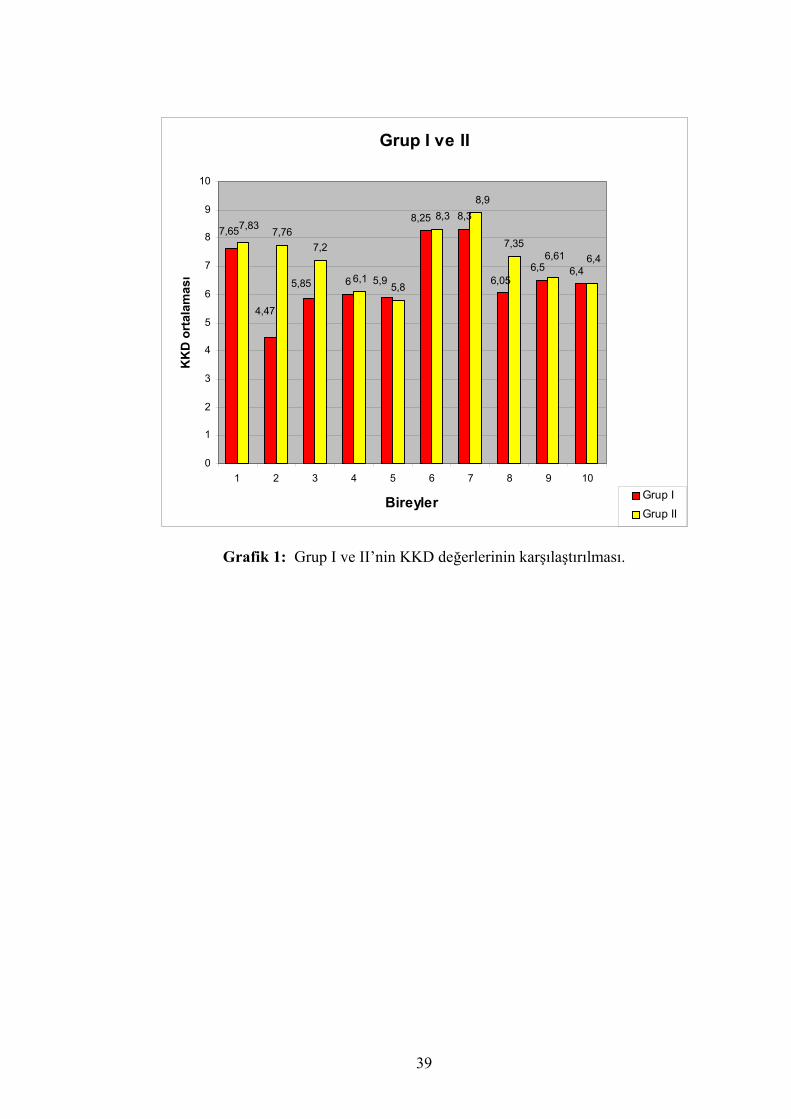
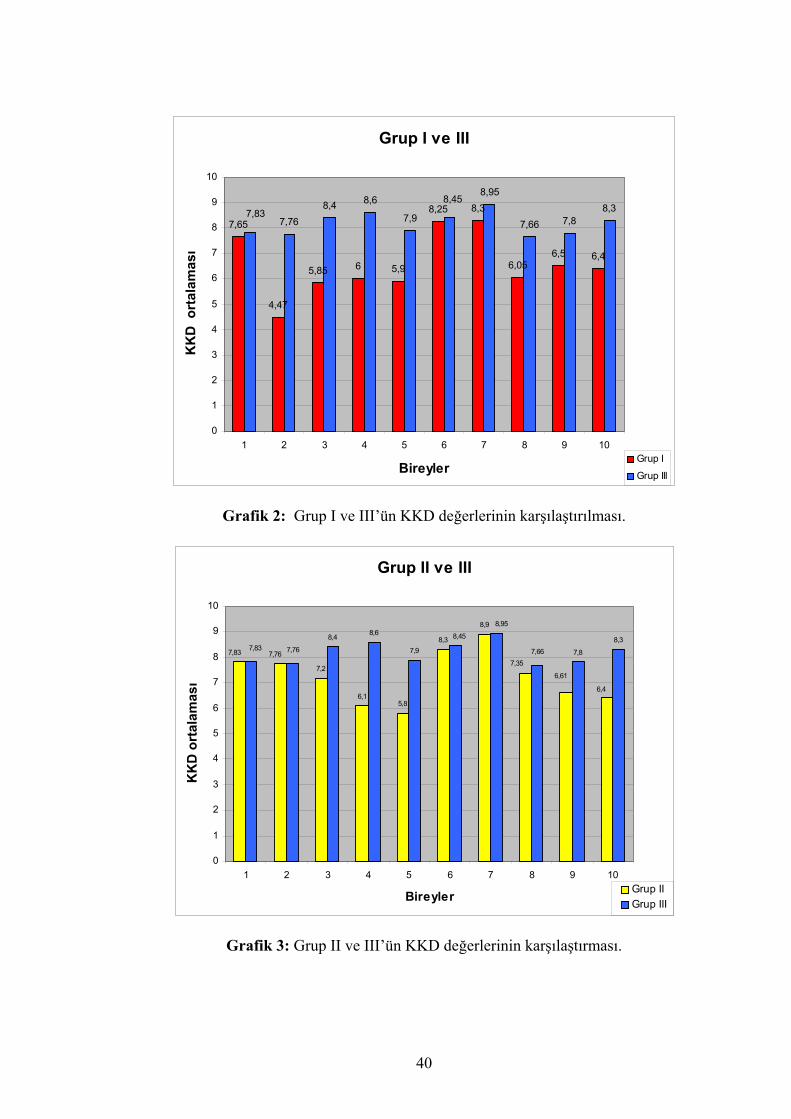
Ayrıca grup I ve II KKD karşılaştırması grafik 1’de, grup I ve grup III KKD
karşılaştırması grafik 2’de, grup II ve III karşılaştırması da grafik 3’te gösterilmiştir.
39
Grup I ve II
6
8,37,76
7,2
6,15,8
7,35
8,25
6,056,5 6,4
5,95,85
4,47
7,65
6,46,61
8,98,3
7,83
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10
Bireyler
KK
D o
rtal
aması
Grup I
Grup II
Grafik 1: Grup I ve II’nin KKD değerlerinin karşılaştırılması.
40
Grup I ve III
7,65
4,47
5,85 6 5,9
8,25 8,3
6,056,5 6,4
7,768,4 8,6
7,9
8,95
7,66 7,88,37,83
8,45
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10
Bireyler
KK
D o
rtal
aması
Grup I
Grup III
Grafik 2: Grup I ve III’ün KKD değerlerinin karşılaştırılması.
Grup II ve III
7,2
6,15,8
8,3
8,9
8,4 8,6
7,9 7,8
8,3
7,83 7,767,35
6,61
6,4
7,83 7,76
8,45
8,95
7,66
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10
Bireyler
KK
D o
rtal
aması
Grup IIGrup III
Grafik 3: Grup II ve III’ün KKD değerlerinin karşılaştırması.
41
Tüm gruplarda kadın ve erkek KKD ortalamalarını karşılaştırıldığında grup
I, grup II, grup III’de erkeklerin KKD oranları kadınlardan yüksek bulunmuştur
(Tablo 4). Grup I ve II’deki fark istatisitksel olarak anlamlı (p<0,05) iken grup
III’deki fark anlamsız bulunmuştur.
Tablo 4: Kadın ve erkek KKD değerleri
CİNSİYET
Grup I
KKD
ortalaması ± ss
Grup II
KKD
ortalaması ± ss
Grup III
KKD
ortalaması ± ss
Kadın (N:5) 5.9 ± 1.1 6.9 ± 0.9 8.1 ± 0.4
Erkek (N:5) 7.1 ± 1.1 7.5 ± 1.1 8.2 ± 0.5
ss: standart sapma N: Birey sayısı

42
Resim 1: Grup I’e ait bir metafaz örneği.
Resim 2: Grup II’e ait metafaz örneği.

43
Resim 3: Grup III’e ait bir metafaz örneği.

44
5. TARTIŞMA VE SONUÇ
H. perforatum bitkisinin en önemli bileşeni hyperisinin antitümör, antiviral
etkileri bilimsel bir gerçektir. Hy’nin bu özellikleri fotosensitizer özelliklerinden
kaynaklandığı yapılan çalışmalarla ortaya çıkarılmıştır (107). PDT kanser
tedavisinde son zamanlarda sık kullanılan bir yöntemdir. PDT’de doğal fotosensitizer
olarak Hyperisin tercih edilmektedir (52). Hy’nin antitümör etkisi in vivo ve in
vitroda meme (46), kolon (115), glioma (60), prostat (5), tiroid (33), nazofarenks
(18,108,17), özafagus (18) kanserlerinde çalışılıp etki mekanizması araştırılmıştır.
Antiviral etkileri 1988’den sonra çok çalışılmış ve Hy’nin sadece zarflı virüsler ve
retrovirüslere etki ettiği ve bu etkinin ışık uygulandığı zaman ortaya çıktığı
belirtilmiştir (14). HIV tedavisi için kullanılabilirliği için çeşitli araştırmalar
yapılmıştır. Etki mekanizmasının tam açıklanamaması, AIDS hastalarına sistemik
uygulamasında görülen fototoksititeden (109) ve klinik uygulama azlığından dolayı
tedavide kullanım için daha çok erken gözükmektedir.
Hy ışıkla aktive olduktan sonra tümör hücrelerinde apopitozis ve nekrozise
yol açmaktadır. Apopitozisin ekstrinsik veya intrinsik sinyal mekanizmasının
işlemesi ile gerçekleştiği bilinmektedir. Her iki sinyal yolunda da sistein içeren
kaspas proteazlar önemli rol oynamaktadır (19). Ekstrinsik yolda mutlaka hücre
zarında bulunan Fas, TNFR (tümör nekroz faktör) gibi ölüm reseptörlerinin
ligandlarınca uyarılması ile sinyal yolu aktiflenir. Hy aracılı apoptoziste protein
kinaz C (PKC) inhibisyonunun tetikleyici olduğu görüşü extrinsik sinyal yolunun
etkili olabileceği fikrini desteklemektedir. İntrinsik yolda sinyal ya hücrenin
genomundan gelir ya da reseptöre bağlanmaya ihtiyaç göstermeyen ilaç, ışın etkisi
ile sağlanır (15,19). Hyperisinin hücre kültürlerine eklendiğinde 3,5 saatte nükleus
içinde biriktiği, hatta hücre DNA’sının pürinlerinin 7. azotu ile etkileşime girdiği
belirlenmiştir (110). Bu da Hyperisinin genom üzerinden mitoz bloğu oluşturduğu
ve apopitotik sinyallerin genomdan gelebileceği fikrini desteklemektedir. Nitekim
son yapılan bir çalışmada topoizomerz IIα’nın Hyperisin tarafından inhibe edilerek
transkripsiyon ve DNA replikasyon bloğu oluşturduğu ileri sürülmüştür (111).
Hyperisinin düşük dozlarda antiproliferatif etki yüksek dozlarda apopitozisi
indükleyici etki gösterdiği yapılan çalışmalarla ortaya konmuştur (35). Marekova ve

45
arkadaşlarını yaptığı bir çalışmada hipofiz kanserinde At20 / D16v-F2 hücrelerinin
büyümesinin fotoaktif hyperisine bağlı inhibisyonunu araştırılmıştır. Işıkla uyarılarak
aktif hale getirilen hyperisin 4-8 μmol/1 konsantrasyonunda HL-60 hücrelerinin
büyümesini büyük oranda inhibe ettiği, ışığa maruz bırakılmadan uygulandığı zaman
ise inhibisyonun önemsiz derecede gerçekleştiği bulunmuştur. Işıkla aktive edilen
hyperisinin 32 μmol/1 konsantrasyonunda At20/D16v-F2 hücrelerinin
proliferasyonun durduğu, 16 μmol/1 konsantrasyonda bu hücrelerin büyümesinin
inhibe olduğu, 1-8 μmol/1 konsantrasyonunda ışık uygulanması halinde apoptozis
görülmüş, apopitoziste protein kinaz C inhibisyonun rolu olduğu fikri üzerinde
durulmuştur (33).
Hypericum perforatum ekstraklarının Avrupa ve Kuzey Amerikada
menepoz sıkıntıları, ılımlı depresyon belirtileri, premenstüral sıkıntılar, anksiyete,
uykusuzluk tedavisi için kullanımı her geçen gün artmaktadır (20). Antidepresan etki
mekanizması henüz tam açıklığa kavuşturulamamış olan H. perforatum
ekstraklarının serotonin, norepinefrin, dopamin, gama amino bitürik asit (GABA), L-
glutamatın sinoptosomal geri salımını inhibe ederek etki gösterdiği ileri sürülmüştür
(112). Bu konuda araştırmalar devam etmekte olup, antidepresan etkiyi, önceleri
hyperisin bileşiğinin gerçekleştirdiği sanılmış daha sonra bu etkiyi hyperisin,
hyperforin ve flovonoitlerin birlikte yaptığı düşünülmüştür (21). Son yapılan
çalışmalarla antidepresan etkiden hyperisinden çok hyperforin bileşiğinin sorumlu
olduğu fikri ağırlık kazanmıştır (113). Klinik çalışmalar H. perforatum ekstraklarını
günde üç kez, her doz 300 mg (%0.3 hypericin) olmak üzere toplam 900 mg’lık dozu
ılımlı depresyon tedavisi için kullanımı önerilmektedir. Bu dozlarda yan etki ve
fotosensitizasyon görülmediği belirtilmektedir (114).
Hunt ve arkadaşalarının yaptığı çalışmada sebest radikal üretimi ölçülmüş
bir serbest hücre kültürü, bir de insan damar doku hücre kültürü H. perforatum
metanol extraktının farklı konsantrasyonları uygulanmış. 1:1 konsantrasyonda
hücrelerde bir prooksidan etki görülmüş olmasına rağmen 1:10 ve 1:20 gibi düşük
konsantrasyonda uygulanan hücrelerde ise yüksek antioksidan özellik gösterdiği
saptanmıştır. 1:1’in üstündeki bütün dilüsyonlarda süperoksit inhibisyonu ile ilişkili
olan antioksidan özelliklere sahip olduğu ileri sürülmüştür. Bu antioksidan etkinin
hangi bileşenden kaynaklandığı ise anlaşılamamıştır (115).

46
Yapılan literatür araştırmalarında hyperisin maddesinin karanlıkta düşük
toksik etkili olduğu rapor edilmiş (116-118) fakat genotoksik etkisi için hiç çalışma
yapılmadığı belirlenmiştir. Bu çalışmamızda KKD sıklığı analizi kullanıldı. KKD
analizi, DNA’daki çok küçük harabiyetlerin bile hassas göstergesi olarak kabul
edildiği, DNA hasarının ve indüklenmiş DNA tamirinin gösterilmesinde en basit,
duyarlı ve kısa zamanda sonuç veren bir yöntem (26) olduğu için tercih edildi.
Küçük yaşalarda KKD oranı düşük, ergenlik çağında ve ileri yaşlarda normalden
yüksek olduğu bildirilmiş (93), sigara içenlerde içmeyenlerden, alkol kullananlarda
kullanmayanlardan daha yüksek bulunmuştur (103,104). Ayrıca Ataksi Telenjektazi,
Bloom Sendromu, Fankoni Anemisi gibi genetik hastalıklarda KKD oranı normalin
10-13 katı artmaktadır (96).
Çalışmamızda birbirine bağlı üç grup oluşturuldu. Çocuk, adolesan çağında,
yaşlı, hamile veya emziren, sürekli ilaç kullanan ve 6 ay içinde röntgen ışığına maruz
kalmış olan bireylerin KKD ortalamaları değişebileceğinden grup bireylerini sağlıklı
yetişkin bay ve bayanlardan seçildi. KKD sıklığını etkileyecek etmenleri minimalize
etmek için tüm grupların ekimleri aynı anda yapıp değerlendirme işlemi için aynı
mikroskop kullanıldı.
Çalışmamızda birinci grup (kontrol grubu) KKD ortalaması bayanlarda
5,9±1,1, erkeklerde 7,1±1,1 olarak bulunmuştur. Bu sonuçlar birey sayısı az
olmasına rağmen, Akbaş’ın yaş ortalaması 27,9 olan, sigara içmeyen, sağlıklı 25 bay
ve 25 bayanın katıldığı çalışmasındaki (104) bay ve bayan KKD ortalamaları ile
uyumludur.
Hy uygulanan gruplar ve Hy uygulanmayan grup arasındaki KKD oranı
farklılığının hücre kültürüne uygulanan Hy miktarına bağlı olduğu sonucuna varıldı.
Çünkü düşük dozda (5 μM) Hy uygulanan grup ile kontrol grubu KKD oranında
anlamlı bir değişiklik yok iken ve yüksek doz (10 μM) uygulanan grubun KKD
oranında anlamlı bir artış söz konusudur. Bu sonuçlarla, doza bağlı olarak insan
lenfosit kültürlerine Hy uygulamasının ışık uyaranı kullanmadan da genotoksik etki
gösterebileceği gözlenmiştir. Kanser tedavisi için kullanılan Hy dozu çok iyi
ayarlanmalı ve yan etkileri yönünden denekler veya normal hücre kültürleri iyi

47
gözlenmelidir. Bu konuda kesin bir sonuca ulaşmak için daha detaylı moleküler
çalışmaların yapılması gerekmektedir.
H. perforatum yurdumuz Isparta yöresinde geleneksel olarak bir çok
hastalık için tedavi edici olarak kullanılmaktadır. Hazırlanan zeytinyağı
ekstraklarında Hy bileşiğinin standardize edilmeme ihtimali yüksek olduğundan
fototoksik etkileri açısından tehlikeli olabilir. Hy maddesi ve dolayısı ile H.
perforatum ekstraklarının dozunun iyi ayarlanması gerekmektedir. Gelişi güzel
kullanımlar zararlı olabilir.

48
Kantaron Otundan (Hypericum perforatum) Elde Edilen Hyperisin Maddesinin
İnsan Lenfosit Kültürlerinde Kardeş Kromatid Değişimi (KKD) Üzerine Etkisi
ÖZET
Hypericin (Hy) kanser tedavisinde son zamanlarda umut verici bir madde
olarak bilimsel çevrelerce ilgi görmektedir. Bu madde hypericum perforatum
bitkisinden elde edilen doğal fotsensitizer olup fotodinamik terapi ile kanser
tedavisine yönelik çalışmalarda kullanılmaktadır. Bu çalışmada, Hy molekülünün
karanlıkta genotoksik etkisinin olup olmadığı araştırıldı. Bunun için sağlıklı ve
gönüllü 10 kişiden (5 bay ve 5 bayan) periferal kan örnekleri alındı. Biri kontrol ve
ikisi deney olmak üzere üç grup oluşturuldu. İlk gruba kardeş kromatid değişimi
(KKD) prosedürü diğer iki gruba düşük doz (5 μM/L) ve yüksek doz (10μM/L) Hy
10 ml’lik insan lenfosit kültürlerine ekleyip KKD prosedürü uygulandı. Elde edilen
KKD ortalamalarını karşılaştırıldı. Birinci grup (kontrol) ile ikinci grup KKD oranı
karşılaştırıldığında anlamlı bir değişiklik gözlenmedi. Birinci grup ile üçüncü grup
karşılaştırıldığında fark istatistiksel olarak anlamlı bulundu (p<0,05). Bu çalışmada,
düşük dozlarda genotoksik etkisi olmayan Hy’nin yüksek dozlarda genotoksik
olabileceği gösterildi.
Anahtar kelimeler: Kardeş Kromatid Değişimi, Hyperisin, Sarı kantaron

49
The efficacy of hypericin derived from Hypericum perforatum on Sister
Chromatid Exchange (SCE) in human lymphocyte culture
SUMMARY
Hypericin is throught to be a promising material in cancer treatment. It is
natural sensitizer extracted from Hypericum perforatum plant and used for the cancer
treatment studies with photodynamic therapy. In this study, we aimed to search
genotoxic effect of hypericin in the darkness peripheral blood samples were obtained
from ten (5 male, 5 Famale) healthy volunteers. Three groups were performed (1
control, 2 test groups). Sister Chromatid Exchange (SCE) procerdure were performed
for control groups and low dose (5 µM/L) and high dose (10 µM/L) of hypericin
were added to human lymphocyte cultures and the SCE procedure was performed
also there two groups. And the mean SCE values were compered. No diffrence as
found between control group and the 5 µM/L hypericin added group. The mean SCE
values between controls and the 10µM/L hypericin added group was found
statistically significant (p<0,05). Our results showed that hypericin may have
genotoxic effect in high doses, in contrast no genotoxic effect shown in low doses.
Keywords: Sister Chromatid Exchange (SCE), Hypericum perforatum, Hypericin

50
KAYNAKLAR
1. Kırbay S. Hypericum perforatum L.’un değişik ekstratlarının antimikrobiyal etkileri.
Journal of Qafgaz University (2 ):121-136.1999.
2. Tanker M, Tanker N. Farmakopnazi cilt 1 S:114 –115 İstanbul,1973.
3. Davis P.H. Flora of Turkey and the East Aegean Islands II. p:355 and P:400 (1978-
1988) Edinburg University Press, Edinburg.
4. Baytop T. Farmakognazi ders kitabı İst.Ünv.Yay.No:2003. Ecz.Fak.No:19 İstanbul,
1974.
5. Colasanti A, Kisslinger A, Liuzzi R, Quarto M, Riccio P, Roberti G, Tramontano D,
Villani F. Hypericin photosensitization of tumor and metastatic cell lines of human
prostate. J Photochem Photobiol B. 54(2-3):103-7, 2000.
6. Tang J, Colacino J.M, Larsen S.H. Virusidal activity of hypericin against envoloped
and non-enveloped DNA and RNA viruse. Antiviral Res. 13 (6): 313-325, 1990.
7. Thiele B, Brink L, Ploch M. Modülasyon of sitokine expression by hypericum extract.
J.Ger.Psychiatry and Neurology 7:60-62, 1993.
8. Reichling J, Weseler A, Saller R.A current review of the antimicrobial activity of
Hypericum perforatum L.Pharmacopsychiatry. 34 Suppl 1:S116-8, 2001.
9. Çubuklu B, Meriçli A.H, Mar A, Sarıyar G, Sütlüpınar N, Meriçli F, İst.Üni.Yayını.
Fitoterapi Yard.Ders Kitabı İst. Üni.Yay No:4311 Ezc.Fak.Yay No:79 İstanbul, 2002.
10. Önder S. Hypericum perforatum L. Bitkisinin Annaljezik Etkisinin Mekanizması.
Anadolu Üniversitesi Sağlık Bilimleri Enst. Yüsek Lisans Tezi. Eskişehir,1995.
11. Herakman T. Hypericum perforatum fraksiyonlarının hepatoprotektif etkilerinin
araştırılması. Anadolu Üniversitesi Sağlık Bilimleri Enst. Yüksek Lisans Tezi.
Eskişehir, 1996.
12. Potacka J. The chemistry, pharmacology and toxicology of the biologicallyactive
constituens of the herb Hypericum perforatum L. J.Appl.Biomed.1,61-73, 2003.
13. Betty R.C, Trikojus V.M, Hypericin and non- flourescent photosensitive pigment from
St. John Wort. J.Exp.Biol.Med.Sci. 21:175-182, 1943.

51
14. Meruelo D, Lavie G, Lavie D. Therapeutic agents with dramatic antiretroviral activity
and little toxicity at effective doses: aromatic polycyclic diones hypericin and
pseudohypericin. Proc Natl Acad Sci. U S A. 85(14):5230-4, 1988.
15. Agostinis P, Vantieghem A, Merlevede W, de Witte PA. Hypericin in cancer treatment:
more light on the way. Int J Biochem Cell Biol. 34(3):221-41,2002.
16. Seyed M.A, Malini O, Gan Y.Y, Bn Soo K.C, İnduction of apoptosis by Hypericin
through activation of caspase-3 in human carcinoma cells. Int. J Mol. Med. 8:521-530,
2001.
17. Du H.Y, Olivo M, Tan B.K, Bay B.H. Hypericin-mediated photodynamic therapy
induces lipid peroxidation and necrosis in nasopharyngeal cancer. Int. J Oncol.
23(5):1401-5, 2003.
18. Hudson J.B, Lopez-Bazzocchi I, Towers G.H. Antiviral activities of hypericin.
Antiviral Research.15:101-112,1991.
19. Fadeel B, Zhivotovsky B, Orrenius S.All along the watchtower: on the regulation of
apoptosis regulators.FASEB J. 13(13):1647-57, 1999.
19. Vantieghem A, Xu Y, Declercq W, Vandenabeele P, Vandenheede J.R, et al. Different
pathways mediate cytochrome c release after photodynamic therapy with hypericin,
Photochem. Photobiol. 74 133–142, 2001.
20. Üstün Ç. Santral sinir sistemi üzerine etkili tıbbi bitkilerin tarihsel süreç içinde ve
günümüzde tedavideki yeri. Ege Üniversitesi Sağlık Bilimleri Enst. Doktora Tezi. İzmir
1998.
21. Wirz A, Simmen V, Herlmann J, Calis I, Meier B, Sticher D. Bisanthraguinone
plycosides of Hypericum perforatum with binding inhibition to CRH-I receptors.
Phytochemistry; 55:941-947, 2000.
22. Baytop T. Farmakognazi ders kitabı İst.Ünv.Yay.No:2003. Ecz.Fak.No:19 İstanbul,
1988.
23. Mukherjee P.K, Verpoorte R, Suresh B. Evalution of in-vivo wound healing activity of
Hypericum patulum (Family:Hypericaceae) leaf extract on different wound model in
rats. J of Ethnopharm; 70:315-321, 2000.
24. Soysal Y. Mitomicin C ile indüklenmiş lenfosit hücre kültürlerinde melotonin ve
karotenin in vitro sistemde kardeş kromatid değişimi oranları ve mitotik indeks

52
değerleri üzerine olan etkilerinin karşılaştırılması. Gazi Üniversitesi Sağlık Bilimleri
Enst. Yüksek Lisans Tezi, 1999.
25. Koşar Aslan P. Mesane kanserli olguların lenfosit hücrelerinde kardeş kromatid
değişim sıklığı. Süleyman Demirel Üniversitesi Sağlık Bilimleri Enstitüsü Tıbbi
Biyoloi.Yüksek Lisans Tezi. Isparta, 1999.
26. Acar A. Ferrokrom fabrikasında çalışan işçilerde sitogetik çalışmalar. Akdeniz
Üniversitesi Tıbbi Biyoloji. Doktora Tezi, 1985.
27. Baytop T. Türkçe Bitki Adları Sözlüğü. Türk Tarih Kurumu basımevi s 130-131.
Ankara, 1994.
28. Çubukçu B, Oğuzoğlu Ş.A. Günümüz Uygur geleneksel bitkisel drogları. Geleneksel ve
Folklorik Droglar Dergisi. cilt:3 sayı 2 s.11 İstanbul, 1996.
29. Guedes R.C, Eriksson L.A. Effects of halogen substitution on the photochemical
properties of hypericin. J Photochem and photobiol. ChemistryA. 246 851-563, 2005.
30. Kitanov G.M. Hypericin and pseudohypericin in some Hy-pericums pecies, Biochem.
Syst. Ecol. 29 171–178,2001.
31. Falk H, Schmitzherger. On The Nature of soluble hypericin Hypericum Speicies.
Momatsh.Chem.123 731-739, 1999.
32. Delaey E.M, Kamuhabwa A.R, Vandenbogaerde A.L, de Witte P.A. Photocytotoxicity
of protohypericin after photoconversion to hypericin. Planta Med. 65(8):719-22, 1999.
33. Marekova M, Vavrova J, Vokurkova D, Cerman J. Light-induced photoactivation of
hypericin inhibits cellular growth in pituitary adenoma cell line At 20/D16v-f2
(Hypericin inhibits cellular growth of AtT20/d16v-F2). Acta Medica 44(1):7-13, 2001.
34. Guedes R.C, Eriksson L.A. Theoretical study of hypericin. J of Photochem and
PhotobilA.Chemistry 172; 293-299, 2005.
35. Crnolatac I, Huygens A, van Aerschot A, Busson R, Rozenski J, de Witte PA.
Synthesis, in vitro cellular uptake and photo-induced antiproliferative effects of
lipophilic hypericin acid derivatives. Bioorg Med Chem. 1;13(23):6347-53, 2005.
36. Mazur Y, Meruelo D. Ion pairs of hypericin compounds having antiviral activity, WO
93/15607,1993.
37. Gerhardt J.J, fowkes S.W. Hypericin Forefront Palo Alto. MegaHealty society 6(6)1-5,
1991.

53
38. Duran N, Song P.S. Hypericin and its photodynamic action. Photochem Photobiol. 198.
43(6):677-80, 1998.
39. Lavie G, Mazur Y, Lavie D, Prince A.M, Pascual D, Liebes L, Levin B, Meruelo D.
Hypericin as an inactivator of infectious viruses in blood components. Transfusion.
35(5):392-400, 1995.
40. Agostinis P, Vandenbogaerde A, Donella-Deana A, Pinna L.A, Lee K.T, Goris J,
Merlevede W, Vandenheede J.R, De Witte P. Photosensitized inhibition of growth
factor-regulated protein kinases by hypericin. Biochem Pharmacol. 49(11):1615-22,
1995.
41. Chaloupka R, Obsil T, Plasek J, Sureau F. The effect of hypericin and hypocrellin-A on
lipid membranes and membrane potential of 3T3 fibroblasts. Biochim Biophys Acta.
1418(1):39-45, 1999.
42. Hadjur C, Richard M.J, Parat M.O, Jardon P, Favier A. Photodynamic effects of
hypericin on lipid peroxidation and antioxidant status in melanoma cells. Photochem
Photobiol. 64(2):375-81, 1996.
43. Miccoli L, Beurdeley-Thomas A, De Pinieux G, Sureau F, Oudard S, Dutrillaux B,
Poupon M.F. Light-induced photoactivation of hypericin affects the energy metabolism
of human glioma cells by inhibiting hexokinase bound to mitochondria. Cancer Res.
58(24):5777-86, 1998.
44. Weiner L, Roth E, Mazur Y, Silman I. Targeted cross-linking of a molten globule form
of acetylcholinesterase by the virucidal agent hypericin. Biochemistry.
31;38(35):11401-5, 1999.
45. Schey K.L, Patat S, Chignell C, Datillo M, Wang R.H, Roberts J.E. Photooxidation of
lens alpha-crystallin by hypericin (active ingredient in St. John's Wort). Photochem
Photobiol. 72(2):200-203, 2000.
46. Thomas C, Pardini R.S. Oxygen dependence of hypericin-induced phototoxicity to
EMT6 mouse mammary carcinoma cells. Photochem Photobiol. 55(6):831-7, 1992.
47. English D.S, Doyle R.T, Petrich J.W, Haydon P.G. Subcellular distributions and
excited-state processes of hypericin in neurons. Photochem Photobiol. 69(3):301-5,
1999.

54
48. Kocisova E, Chinsky L, Miskovsky P, Sequence specificinteraction of the
antiretrovirally active drug hypericin with5_ ATGGCAGGATAT3_ oligonucleotide: a
resonance Ramanspectroscopy study. J. Biomol. Struct. Dyn. 15 1147–1154, 1998.
49. Kocisova E, Chinsky L, Miskovsky P, Sequence specificinteraction of the photoactive
drug hypericin depends on the structural arrangement and the stability of the
structurecontaining its specific 5AG3 target: a resonance Ramanspectroscopy study. J.
Biomol. Struct. Dyn. 17 51–59, 1999.
50. Oleinick N.L , Evans H.H , The photobiology of photo-dynamictherapy: cellular targets
and mechanisms. Rad.Res. 150 S146–S156, 1998.
51. Vandenbogaerde A.L, Delaey E.M, Vantieghem A.M, Himpens B.E, Merlevede W.J,
de Witte P.A. Cytotoxicity and antiproliferative effect of hypericin and derivatives after
photosensitization. Photochem Photobiol. 67(1):119-25, 1998.
52. Du H.Y, Olivo M, Tan B.K, Bay B.H. Photoactivation of hypericin down-regulates
glutathione S-transferase activity in nasopharyngeal cancer cells. Cancer Lett.
30;207(2):175-81, 2004.
53. Halliwel B, Gutteridge J.M.C. Free radicals in biology and medicine.Second editio,
Claren Pres. Oxford, 1989.
54. Mollaoğlu H. GSM 900 MHz Telefonlarının oluşturduğu manyetik alanın etkisiyle
ratlarda meydana gelen oksidatif değişiklikler üzerine melatoninin etkisi. Süleyman
Demirel Üniveristesi Tıp fakültesi fizyoloji AB. Uzmanlık Tezi. Isparta, 2003.
55. Akkuş İ. Serbest Radikaller ve Fizyopatolojik Etkiler. Mimoza Yayınları. s,1-132.
Konya, 1995.
56. Dougherty T.J, Gomer C.J, Henderson B.W, Jori G, Kessel D, Korbelik M, Moan J,
Peng Q. Photodynamic therapy. J Natl Cancer Inst. 17;90(12):889-905, 1998.
57. Fiers W, Beyaert R, Declercq W, Vandenabeele P. More than one way to die:
apoptosis, necrosis and reactive oxygen damage, Oncogene 18 7719–7730, 1999.
58. Ashkenazi A,. Dixit V.M. Apoptosis Control by Death Anddecoy Receptors, Curr.
Opin. Cell Biol. 11, 255–260, 1999.
59. Ustaçelebi Ş, Mutlu G, İmir T, Cengiz T, Tümbay E, Mete Ö. Temel ve klinik
mikrobiyoloji. Güneş Kitapevi. Ankara 1999.

55
60. Couldwell W.T, Gopalakrishna R, Hinton D.R, He S, Weiss M.H, Law R.E, Apuzzo
M.L, Law R.E. Hypericin:a potential antiglioma therapy. Neurosurgery 35 705–710,
1994.
61. Weller M, Trepel M, Grimmel C, Schabet M, Bremen D, Krajewski S, Reed J.C.
Hypericin induced apoptosisof human malignant glioma cells is light-dependent,
independent of Bcl-2 expression, and does not requirewild-type p53, Neurol. Res. 19,
459–470, 1997.
62. Fadeel B, Zhivotovsky B, Orrenius S. All along the watchtower: on the regulation of
apoptosis regulators, FASEB J. 13 1647–1657, 1999.
63. Green D.R, Reed J.C. Mitochondria and apoptosis. Science. 281 (1309–1312, 1998.
64. Hudson JB, Harris L, Towers GH. The importance of light in the anti-HIV effect of
hypericin.Antiviral Res. 20(2):173-8, 1993.
65. Zheng M.S. An experimental study of the anti-HSV-II action of 500 herbal drugs J
Tradit Chin Med. 9(2):113-6, 1989.
66. Degar S, Lavie G, Meruelo D. Photodynamic inactivation of radiation leukemia virus
produced from hypericin-treated cells. Virology. 197(2):796-80, 1993.
67. Lenard J, Rabson A, Vanderoef R. Photodynamic inactivation of infectivity of
humanimmunodeficiency virus and other enveloped viruses using hypericin and rose
bengal: inhibition of fusion and syncytia formation. Proc Natl Acad Sci U S A.
1;90(1):158-62, 1993.
68. Schinazi R.F, Chu C.K, Babu J.R, Oswald B.J, Saalmann V, Cannon D.L, Eriksson
B.F, Nasr M. Anthraquinones as a new class of antiviral agents against human
immunodeficiency virus. Antiviral Res. 13(5):265-72, 1990.
69. Lavie G, Valentine F, Levin B, Mazur Y, Gallo G, Lavie D, Weiner D, Meruelo D.
Studies of the mechanisms of action of the antiretroviral agents hypericin and
pseudohypericin. Proc Natl Acad Sci U S A. 86(15):5963-7, 1989.
70. Tang J, Colacino J.M, Larsen L.H, Spitzer W. Virucidalactivity of hypericin against
enveloped and non-enveloped DNA and RNA viruses, Antiviral Res. 13 313–325,
1990.
71. Prince A.M, Pascual D, Meruelo D, Liebes L, Mazur Y, Dubovi E, Mandel M, Lavie
G., Strategies for evaluation of enveloped virus inactivation in red cell concentrates
using hypericin, Photochem. Photobiol. 71 188–195, 2000.

56
72. Liebes L, Mazur Y, Freeman D, Lavie D, Lavie G, Kudler N, Mendoza S, Levin B,
Hochster H, Meruelo D. A method for the quantitation of hypericin, an antiviral agent,
in biological fluids by high-performance liquid chromatography. Anal Biochem.
15;195(1):77-85, 1991.
73. Chatterjee S.S, Bhattacharya S.K, Wonnemann M, Singer A & Muller W.E. Hyperforin
as a possible antidepressant component of Hypericum extracts. Life Sciences 63 (6):
499-510, 1998.
74. Linde K, Ramirez G, Mulrow C, Pauls DçA, Weidenhammer W, & Melchart D. St.
John's wort for depression- an overview and meta-analysis of randomized clinical trials.
BMJ 313 (7052): 253-258, 1996.
75. Butterweck V, Petereit F, Winterhoff H, Nahrstedt A. Solubilized hypericin and
pseudohypericin from Hypericum perforatum exert antidepressant activity in the forced
swimming test. Planta Med. 64(4):291-4, 1998.
76. Tucker J.D, Auletta A, Cimino M.C, Dearfield K.L. Jacobson- Kram D, Tice R.R,
Carrona A.V. Sister-chromatid Exchange: second report of the Gene-Tox program.
Mutation Research,1993: 101-180.
77. Butterweck V, Wall A, Lieflander-Wulf U, Winterhoff H, Nahrstedt A. Effects of the
total extract and fractions of Hypericum perforatum in animal assays for antidepressant
activity. Pharmacopsychiatry. 30 Suppl 2:117-24, 1997.
78. De Vry J, Maurel S, Schreiber R, de Beun R, Jentzsch K.R. Comparison of hypericum
extracts with imipramine and fluoxetine in animal models of depression and
alcoholism. Eur Neuropsychopharmacol. 9(6):461-8, 1999.
79. Vorbach E.U, Arnoldt K.H, Hubner W.D. Efficacy and tolerability of St. John's wort
extract LI 160 versus imipramine in patients with severe depressive episodes according
to ICD-10. Pharmacopsychiatry.30 Suppl 2:81-5, 1997.
80. Lenoir S, Degenring F.H, Saller R. A double-blind randomised trial to investigate three
different concentrations of a standardised fresh plant extract obtained from the shoot
tips of Hypericum perforatum L. Phytomedicine. 6(3):141-6, 1999.
81. Franklin M, Chi J, McGavin C, Hockney R, Reed A, Campling G, Whale R.W, Cowen
PJ. Neuroendocrine evidence for dopaminergic actions of hypericum extract (LI 160) in
healthy volunteers. Biol Psychiatry. 15;46(4):581-4, 1999.

57
82. Calapai G, Crupi A, Firenzuoli F, Costantino G, Inferrera G, Campo G.M, Caputi AP.
Effects of Hypericum perforatum on levels of 5-hydroxytryptamine, noradrenaline and
dopamine in the cortex, diencephalon and brainstem of the rat. J Pharm Pharmacol.
51(6):723-8, 1999
83. Muller W.E, Rolli M, Schafer C, Hafner U. Effects of hypericum extract (LI 160) in
biochemical models of antidepressant activity. Pharmacopsychiatry. 30 l 2:102-7, 1997.
84. Kaehler S.T, Sinner C, Chatterjee S.S, Philippu A. Hyperforin enhances the
extracellular concentrations of catecholamines, serotonin and glutamate in the rat locus
coeruleus. Neurosci Lett. 12;262(3):199-202, 1999.
85. Butterweck V, Bockers T, Korte B, Wittkowski W, Winterhoff H. Long-term effects of
St. John's wort and hypericin on monoamine levels in rat hypothalamus and
hippocampus. Brain Res. 15;930(1-2):21-9, 2002.
86. Rodriquez Landa J, Contresas F. A Rewiev of Clinical and Experimental Observations
About Antidepressant Actions and Site Effects Prodused by Hypericum Perforatum
Extrakts. Pyhtomedicine 10:688-699, 2003.
87. Chatterjee S.S, Noldner M,koch E,erdelmeier C. Antidepresant activity of hypericum
perforatum and hyperforin: the negleted possibility. Pharmacopsychiatr.31;7-15, 1998.
88. Thiede H.M and A. Walper. Inhibition of MAO and COMT by Hypericum Extracts and
Hypericin. J Geriatr Psychiatry Neurol 7(suppl 1): S54-S56, 1994.
89. Tuğlular I. Klinik Uygulamalı Farmakoloji Cilt:1 Psikofarmakoloji. MAS Ambalaj
Sanayi, İzmir 1985.
90. Neary J. T, & Bu Y. Hypericum LI 160 inhibits uptake of serotonin and norepinephrine
in astrocytes. Brain Research 816 (2): 358-363, 1999.
91. Öztürk M.O. Ruh Sağlığı ve Bozuklukları. Hacettepe Üniversitesi AB. Emekli Öğretim
Üyesi. 7. Baskı.s(25). Ankara 1997.
92. Mennini T, Gobbi M. The antidepressant mechanism of Hypericum perforatum. Life
Sci. 16;75(9):1021-7, 2004.
93. Tanrıverdi N. Kardeş Kromatid Değişimi. Çukurova Üniversitesi Sağlık Bilimleri
Enstitüsü Fiz. AB. Bilim Uzmanlığı Tezi, 1991.

58
94. Ceylaner Başkaya G. Şizofrenili olgularda kardeş kromatid değişimlerinin (Sister
Chromatid Exchange-SCE) araştırılması. Cumhuriyet Üniversitesi Sağlık Bilimleri
Enst. Doktora Tezi, 1997.
95. Paralı F. Ethidum bromidin (EtBr) in vitro insan lenfositlerine ekisisnin SCE yöntemi
ile gösterilmesi. Dokuz Eylül Üniversitesi Sağlık Bilimleri Enst.Yüksek Lisans Tezi,
1994.
96. Bal F, Şahin F.İ, Yirmibeş M, Balcı A, Menevşe S. The in vitro effect of beta-carotene
and mitomycin C on SCE frequency in Down’s Syndrome lymphocyte cultures.
Tohoku. J. Exp. Med. 184 (4): 295-300, 1998.
97. Perry P, Wolff S. New Giemsa method for the differential staining of sister chromatids.
Nature. 13;251(5471):156-8, 1974.
98. Zakharov ve Egolina N.A. Differantial spirilization along mammalian mitotic
chromosomes. Chromosoma; 38: 341-365. (Berl) 1972.
99. Lovadey K.S, Latt S.A. A high buyant density fraction in mammalian DNA.
Experimental Cell Res. 141: 127-137,1982.
100. Latt S.A. Sister chromatid exchanges, indices of human chromosome damage and
repair: detection by fluorescence and induction by mitomycin C. Proc Natl Acad Sci U
S A. 71(8):3162-6, 1974.
101. Sakallı Çetin E. Ökse Otunun Kardeş Kromatid Değişimi Üzerine Etkisi. Süleyman
Demirel Üniversitesi Sağlık Bilimleri Enst. Tıbbi Tiyoloji ve Genetik AB. Yüksek
Tisans Tezi. Isparta 2005.
102. Erdem N. Kontraplak ve yonga-levha fabrikalrında çalışan işçilarde formaldehit ve
odun tozuna maruziyetin solunum fonksiyonları üzreine etkisi ve genotoksik hasarın
araştırılması. Gazi Üniversitesi Sağlık Bilimleri Enst. Doktara Tezi, 1997.
103. Başaran Gönlüşen F. Alkolik bireylerdeki alkolve sigara tüketimine bağlı oluşan SCE
oranlarının sigara içen ve sigara içmeyen bireylerdeki SCE oranları ile karşılaştırılması.
Yüksek Lisans Tezi, 1998.
104. Akbaş E. Sigara Kullanımının Lenfosit Yaşam Süresi ve Genotoksik Etkilerinin
İncelenmesi. Mersin Üniversitesi Tıp Fakültesi Tıbbi Biyoloji ve Genetik AB. Geriatri
4(1):15-18, 2001.

59
105. Moorhead P, Nowell P.C, Mellman W.J, Batips D.M, and Hungerford D.A.
Chromosome preparations of leukocytes cultured from human peripheral blood. Exp.
Cell Res. 20; 613-616, 1960.
106. Korenberg J, Feedlender E.F. Giemsa technique for the detection of sister chromatid
exchanges. Chromosome (Berl). 48: 355-360, 1974.
107. Hostanska K, Reichling J, Bommer S, Weber M, Saller R. Hyperforin a constituent of
St John's wort (Hypericum perforatum L.) extract induces apoptosis by triggering
activation of caspases and with hypericin synergistically exerts cytotoxicity towards
human malignant cell lines. Eur J Pharm Biopharm. 56(1):121-32, 2003.
108. Johnson S.A, Dalton A.E, Pardini R.S. Time-course of hypericin phototoxicity and
effect on mitochondrial energies in EMT6 mouse mammary carcinoma cells. Free
Radic Biol Med. 15;25(2):144-52, 1998.
109. Gulick R.M, Mc Auliffe V, Holden-Wiltse J, Crumpacker C, Liebes L, Stein D.S,
Meehan P, Hussey S, Forcht J, Valentine F.T. Phase I studies of hypericin, the active
compound in St. John's Wort, as an antiretroviral agent in HIV-infected adults. AIDS
Clinical Trials Group Protocols 150 and 258. Ann Intern Med. 16;130(6):510, 1999.
110. Miskovsky P, Sureau F, Chinsky L, Turpin P.Y. Subcellular distribution of hypericin in
human cancer cells. Photochem Photobiol. 62(3):546-9. 1995.
111. Peebles K.A, Baker R.K, Kurz E.U, Schneider B.J, Kroll D.J. atalytic inhibition of
human DNA topoisomerase IIalpha by hypericin, a naphthodianthrone from St. John's
wort (Hypericum perforatum). Biochem Pharmacol. 15;62(8):1059-70, 2001.
112. Barnes J, Anderson L.A, Phillipson J.D. St. John's wort (Hypericum perforatum L.): a
review of its chemistry, pharmacology and clinical properties. J Pharm Pharmacol.
53(5):583-600, 2001.
113. Raffa R.B. Screen of receptor and uptake-site activity of hypericin component of St.
John's wort reveals sigma receptor binding. Life Sci. 62(16):PL265-70, 1998.
114. Biber A, Fischer H, Romer A, Chatterje S.S. Oral bioavailability of hyperforin from
hypericum extakts in rat and human volunteers. Pharmacopschiatry.31;36-43,1998.
115. Hunt E.J, Lester C.E, Lester E.A, Tackett R.L. Effect of St. John's wort on free radical
production. Life Sci. 1;69 (2):181-90, 2001.

60
116. Siboni G, Amit-Patito I, Weizman E, Waintraub-Porat M, Weitman H, Ehrenberg B,
Malik Z. Specificity of photosensitizer accumulation in undifferentiated versus
differentiated colon carcinoma cells. Cancer Lett. 30;196(1):57-64, 2003.
117. Jacobson J.M, Feinman L, Liebes L, Ostrow N, Koslowski V. Et al. Pharmacokinetics,
safety, and antiviral effects of hypericin, a derivative of St. John’s wort plant, in
patientswith chronichepatitis C virus infection, Antimicrob. Agents Chemother. 45;
517–524, 2001.
118. Fox E, Murphy R.F, McCully C.L, Adamson P.C. Plasmapharmacokinetics and
cerebrospinal fluid penetration ofhypericin in non-human primates, Cancer Chemother.
Pharmacol. 47; 41–44, 2001.





























