Embed Size (px)
Citation preview

ΚΥΤΤΑΡΙΚΟΣ ΚΥΚΛΟΣ - ΓΗΡΑΝΣΗ
Ιωάννης Π. ΤρουγκάκοςΤομέας Βιολογίας Κυττάρου & ΒιοφυσικήςΤμήμα Βιολογίας, Παν/μιο Αθηνών

M
G2
S
G1
Βλάβη στο DNA(G1)
Μη διπλασιασμένο DNA(S)
Βλάβη στο DNA(G2)
ΕΚΚΙΝΗΣΗ (R)
Μη φυσιολογικήΜιτωτική άτρακτος (Μ)
Πρόφαση: ΥψηλάΕπίπεδα MPF
ΕΛΕΓΚΤΗΣ
Είναι το μέγεθος του κύτταρου αρκετά μεγάλο?Είναι το περιβάλλον ευνοϊκό?Μπορεί το DNA να διπλασιαστεί με ασφάλεια?
Είναι όλα τα χρωμοσώματα ευθυγραμμισμέναστην άτρακτο?
∆ιπλασιάστηκε όλο το DNA?Είναι το περιβάλλον ευνοϊκό?Είναι το μέγεθος του κυττάρουικανοποιητικό?
Η εξέλιξη του κυτταρικούκύκλου εξαρτάται απόπολλαπλά σημείαελέγχου (check points)
τόσο κατά τη μεσόφαση όσοκαι κατά τη μίτωση. Στα
σημεία αυτά ελέγχονται μιασειρά από παραμέτρουςπου σχετίζονται με τηδραστηριότητα που
λαμβάνει χώρα σε κάθεφάση του κυτταρικού κύκλουόπως για παράδειγμα ηπαρουσία μιτογόνων, ήβλαβών στο DNA στο
σημείο G1S, η ακεραιότητατου διπλασιασμένου DNAμετά το τέλος της S φάσης, ή η σωστή διαμόρφωση τηςμιτωτικής ατράκτου κατά τημίτωση. Ο “εντοπισμός”βλαβών από τον ελεγκτή
οδηγεί σε αναστολή εξέλιξηςτου κυτταρικού κύκλου.
Σημεία ελέγχου του Κυτταρικού κύκλου (check points)

Ρυθμιστική υπομονάδα(Κυκλίνη-Cyc)
Καταλυτικη υπομονάδα(Κινάση εξαρτώμενη απόκυκλίνη-Cdk)
ΟλοένζυμοΣυμπλέγματος
ΚινάσηςΕξαρτώμενηςαπό κυκλίνηCdk-Cyc
Σύνδεση Cdk μεδιαφορετικές Cycs
Φωσφορυλίωσηδιαφορετικών πρωτεϊνών
Βασίζεται κατά κύριο λόγο σε μιαοικογένεια κινασών (ετεροδιμερή) πουαποτελούνται από μια ρυθμιστικήυπομονάδα (κυκλίνη-Cyc) και μιακαταλυτική υπομονάδα (κυκλινο-
εξαρτώμενη κινάση-Cdk). Ηενεργοποιημένη μορφή της κινάσηςμπορεί να φωσφορυλιώσει (και να
ενεργοποιήσει) διάφορα υποστρώματα. Στους μύκητες η ίδια Cdk μπορεί να
συνδεθεί με διάφορες Cyc.
Μοριακός μηχανισμός (βασίζεται σε ενεργότητα πρωτεϊνικήςκινάσης) που ελέγχει την πρόοδο του κυτταρικού κύκλου

NH2
COOH
Κουτί καταστροφής
Πρωτεόσωμα
G1 S G2 M
Συγκέντρωση κυκλινών
Ενεργότητα MPF
NH2
COOH
APCΠρωτεόλυση
Ρύθμιση σχετικήςσυγκέντρωσης κυκλινώνμέσω αποδόσης στο
πρωτεάσωμα
Σχετική συγκέντρωσηκυκλινών και ενεργότηταςMPF στις διάφορες φάσειςτου κυτταρικού κύκλου
Παράλληλα, κατά την εξέλιξη του κυτταρικού κύκλου μεγάλα πρωτεϊνικά σύμπλοκα είναι δυνατόν ναμαρκάρουν είτε κάποιους ειδικούς αναστολείς των διαφόρων γεγονότων του κυτταρικού κύκλου είτετις ίδιες τις κυκλίνες (εναλακτικός μηχανισμός ρύθμισης της ενεργότητας των Cyc-Cdk συμπλόκων)
προωθόντας έτσι την αποδόμηση τους στο πρωτεόσωμα. Όταν η συγκέντρωση της Cyc είναιχαμηλή η Cdk λόγω απώλειας της Cyc είναι ανενεργή και ενεργοποιείται μόνο όταν η συγκέντρωση
της Cyc υπερβεί κάποια όρια.

MG2
S
G1
ΣημείοΠεριορισμού
G0
Cdk2-CycΕ
Cdk1-Cycs Α,B
6-12 hrs
6-8 hrs
3-4 hrs1 hr
Cdk2-CycA
Cdks 4, 6- (5?)Cycs D1,2,3
E2F1,2,3
E2F4,5
Μεγάληενεργότητακινάσης
Μικρήενεργότητακινάσης
Ενεργότητα τωνσυμπλόκων κυκλινών καικυλινο-εξαρτώμενωνκινασών (τόξα) κατά τηδιάρκεια του κυτταρικού
κύκλου σε κύτταραθηλαστικών (τα οποία
είναι δυνατόν ναβρίσκονταν σε φάση G0και έχουν διεγερθεί μέσωμιτογόνων να εισέλθουνστον κυτταρικό κύκλο).
Το πλάτος των γραμμών (τόξα) απεικονίζει χονδρικά την
ενεργότητα των συμπλόκωνCdk-Cyc που αναφέρονται σεκάθε φάση του κυτταρικού
κύκλου. Τα σκιασμένα βέλη στηφάση G1 υποδεικνύουν την
ενεργοποίηση των μελών τηςοικογένειας του μεταγραφικού
παράγοντα E2F από τοαντίστοιχο Cdk-Cyc σύμπλοκο.

Γενικά η προώθηση του κυτταρικού κύκλου πέρα από το σημείο ελέγχου R(restriction point) εξαρτάται από την ενεργότητα των μεταγραφικώνπαραγόντων της οικογένειας E2F. Οι μεταγραφικοί αυτοί παράγοντες
απαιτούνται για την μεταγραφή αρκετών γονιδίων όπως είναι οι Cyc-Α,-Ε καιCdk2-, ενώ επιπλέον οι E2F ενεργοποιούν και τα ίδια τους τα γονίδια
(αυτοενεργοποίηση).
Η μεταγραφική ενεργότητα των E2F παραγόντων αναστέλεται είτε από τηπρόσδεση της πρωτεϊνης του ρετινοβλαστώματος (pRb) είτε από την πρόσδεση
δύο σχετίζομενων πρωτεϊνών των p107 και p130.

Cyc-A,BCyc-A,BCyc-A,B
Cyc-A,B
Rb
Φωσφορυλίωσηκινάσης
Cyc-A,B
Cdk1
Cyc-A,B
Αυτο-φωσφορυλίωσηκυκλίνης
ΣύνδεσηΚυκλίνης-Κινάσης
Απο-φωσφορυλίωσηκινάσης
Φωσφορυλίωσηκινάσης
ΣύνθεσηΚυκλίνης
Αποδόμησησηκυκλίνης
Αποφωσφορυλίωσηκινάσης
Cdk1
Cdk1Cdk1
Cdk1Cdk1
Κινάση Wee1
Κινάση CAK
Φωσφατάση Cdc25
Tyr1
5
Thr1
61
Ενεργοποίηση
ΑΝΕΝΕΡΓΗΑΝΕΝΕΡΓΗ ΕΝΕΡΓΗ
MFPMFP
πρόσδεσηυποστρώματος
APC
Η ενεργότητα τηςκινάσης της φάσης τηςμίτωσης (MPK ή MPF)
(σύμπλοκο CycA ή B-Cdk1) ρυθμίζεται στα θηλαστικά
(όπως και στονσακχαρομύκητα) μέσω
φωσφορυλιώσεων (κινάσεςWee1, CAK) και
αποφωσφορυριώσεων(φωσφατάση Cdc25) της
Cdk1 και πρωτεϊνικήςαποδόμησης των κυκλινών Α, Β κατά το τέλος της μίτωσης. Οι κινάσες που ενεργοποιούντο σύμπλοκο είναι αντίστοιχεςμε αυτές που έχουν βρεθείστους μύκητες. Τα αμινοξέαπου φωσφορυλιώνονται στηνCdk2 είναι η Tyr15 και η Thr161. Ο σχετικό χρονισμός για κάθε
γεγονός δεν είναι ακόμηαπόλυτα γνωστός.

Φωσφορυλίωση του pRbΑποδέσμευση του E2F
M
G2
S
G1
Κυκλίνη D
Cdk4/6
Rb E2F
E2F
Κυκλίνη E
Cdk2
E2F
Απο-φωσφορυλίωση του pRbπρόσδεση στον E2FΌμοια σε G0 κύτταρα
ΜΙΤΟΓΟΝΑ
ή p107, p130
PP-1
Τα μιτογόνα μέσω διακριτώνμονοπατιών μεταγωγής σήματος
ενεργοποιούν τα σύμπλοκακυκλινών-κινασών CycD-Cdk4,
CycD-Cdk6 και CycΕ-Cdk2 τα οποίαμε τη σειρά τους φωσφορυλιώνουντην πρωτεϊνη pRb που έχει προσδέσειτους μεταγραφικούς παράγοντες τηςοικογένειας E2F (ή τις πρωτεϊνες p107,
p130 που επίσης προσδένουν τουςE2F παράγοντες) Η φωσφορυλίωσητης pRb έχει σαν αποτέλεσμα τηναπελευθέρωση των μεταγραφικώνπαραγόντων E2F που ενεργοποιούνστη συνέχεια τα ίδια τους τα γονίδια(αυτορύθμιση), τις κυκλίνες και τιςκυκλινο-εξαρτώμενες κινάσες που
εμπλέκονται στη φωσφορυλίωση τουpRb αλλά και μια σειρά άλλων γονιδίωνπου συμμετέχουν στο διπλασιασμό τουDNA επιτρέποντας έτσι την προώθησητου κυτταρικού κύκλου στο σημείο
G1S. Η αποφωσφορυλίωση του E2F(από τη φωσφατάση PP-1) κατά το
τέλος της μίτωσης έχει σαν αποτέλεσματην αναστολή της κυτταρικής αύξησηςμέχρι να παρουσιαστεί και πάλι τοκατάλληλο ερέθισμα (παρουσία
μιτογόνων).
Μοριακά γεγονότα που απαιτούνται για τηνπροώθηση του κυτταρικού κύκλου στο σημείο
μετάπτωσης G1S μετά από ενεργοποίηση εισόδουστον κυτταρικό κύκλο παρουσία μιτογόνων

Ν CI II III IV V
ΠρόσδεσηSV40T
ΠρόσδεσηE6, E1bΠρόσδεση
MDM2Θέση τετραμερισμού
Θέση ενεργοποίησηςμεταγραφής
Περιοχή πρόσδεσης στο DNAμη-εξαρτώμενη από την ακολουθία
Περιοχή πρόσδεσης στο DNAεξαρτώμενη από την ακολουθία στόχο
Mdm2αποδόμηση p53αναστολή έκφρασης p21
Δομής της πρωτεΐνης p53
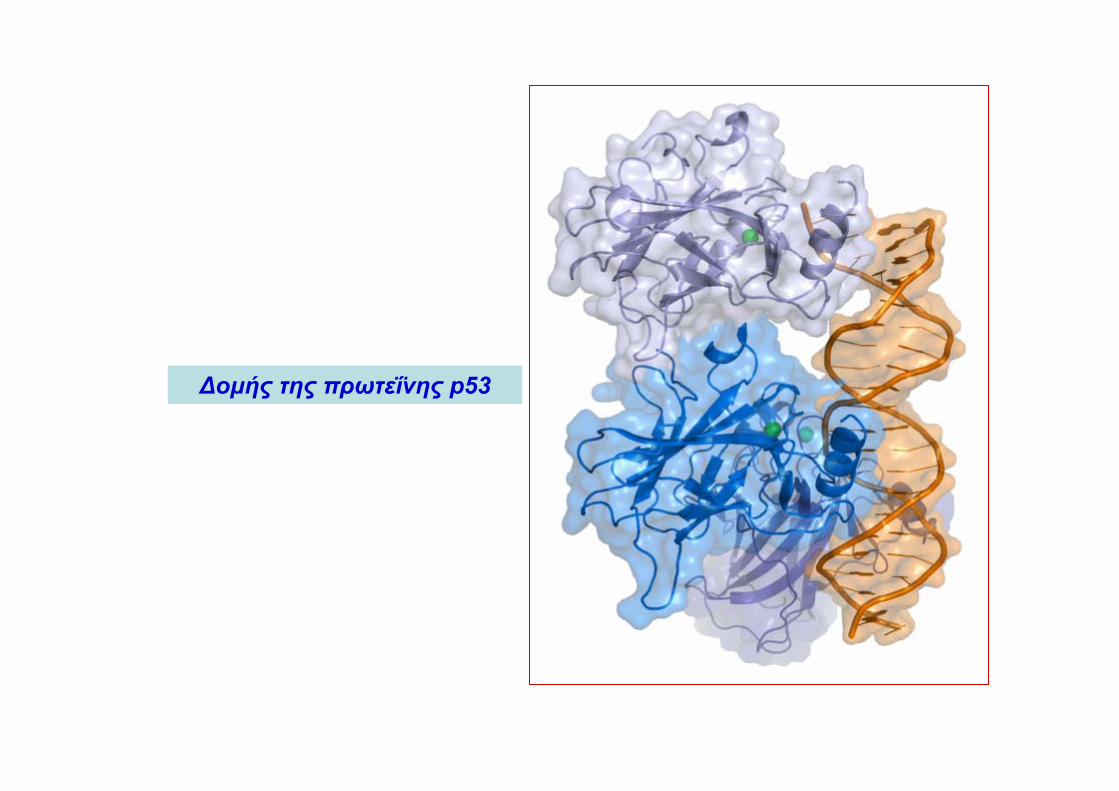
Δομής της πρωτεΐνης p53

p19p19 p19
Mdm2
p53
Mdm2
Mdm2
p53
Mdm2
p53
αποδόμηση του p53από το 26S πρωτεόσωμα
Mdm2Mdm2Mdm2
πυρηνίσκος
πυρήνας
Ρύθμισης της σταθερότητας και τοπολογίαςτης πρωτεΐνης p53

p53
Bax MDM2
p21
Cdk-Cyc, PCNA
GADD45
προσδένεται στο PCNAεπιδιόρθωση DNA
Cyclin Gάγνωστη λειτουργία
IGF-BP3
απόπτωση
σινιάλα από μιτογόνουςαναπτυξιακούς παράγοντες
Βλάβη στο DNA
ATM
p53
p14ARF ATM
Η ανακοπή του κυτταρικού κύκλου σε κάποιο από τα σημεία ελέγχου συνδέεται μετη σύνθεση των CdkIs η οποία ενεργοποιείται ειδκά μέσω ειδικών μονοπατιών
μεταγωγής σήματος που εντοπίζουν τις εν λόγω βλάβες.
Τα πιο πολλά τέτοια μονοπάτια μεταγωγής σήματος ελέγχονται από την πρωτεΐνη p53 (τον «φύλακα του γονιδιώματοσ»)

Το μονοπάτι σηματοδότησης του p53
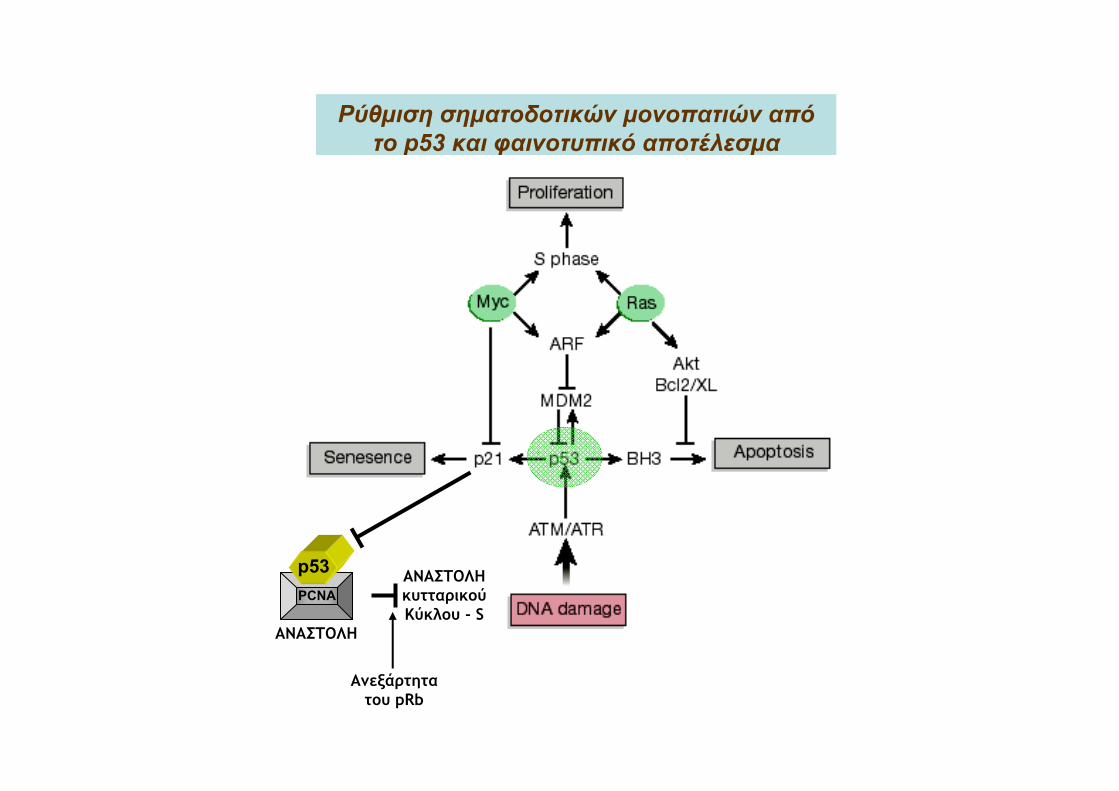
Ρύθμιση σηματοδοτικών μονοπατιών απότο p53 και φαινοτυπικό αποτέλεσμα
ΑΝΑΣΤΟΛΗ
ΑΝΑΣΤΟΛΗκυτταρικούΚύκλου - S
Ανεξάρτητατου pRb
p53PCNA

Οι CdkIs αποτελούν μέρος των μηχανισμών ανάδρασης των κυττάρων που ελέγχουν τα κατάσταση τωνγεγονότων που λαμβάνουν χώρα κατά τον κυτταρικό, κύκλο όπως η αντιγραφή του DNA ή η
συμπύκνωση των χρωμοσωμάτων. Η “ανάλυση” της πληροφορία αυτής στη συνέχειακαθορίζει αν ο κυτταρικός κύκλος θα συνεχιστεί. Κατά αυτήν την έννοια τα σημεία G1S καιG2M είναι ζωτικής σημασίας όσον αφορά τους πρωταρχικούς ελέγχους των μηχανισμών ανάδρασης
και αφορούν τη “δέσμευση” (που λαμβάνει χώρα κατά τη φάση G1) για τον διπλασιασμό τωνχρωμοσωμάτων και τη “δέσμευση” (που συμβαίνει στο τέλος της φάσης G2) για την είσοδο στη μίτωση(δευτερεύων σημείο ελέγχου). Αν ένα κύτταρο εκτεθεί σε παράγοντες που προκαλουν βλάβες στο DNA
το πέρασμα από το σημείο G1-S καθυστερεί μέχρι να διορθωθεί η βλάβη, ενώ αν διαπιστωθεί μησωστός σχηματισμός της μιτωτικής ατράκτου ο κύκλος διακόπτεται κατά την ανάφαση.
Η καταστολή της ενεργότητας των συμπλόκων Cyc-Cdk (ενεργότητα κινάσης) και κατ΄επέκταση η περαιτέρω ρύθμιση της εξέλιξης του κυτταρικού κύκλου επιτυγχάνεται μέσω
αρνητικών ρυθμιστών στους οποίους περιλαμβάνονται ομάδες πρωτεϊνών μικρού(συνήθως) μοριακού βάρους που ονομάζονται αναστολείς Cdk (CdkI, Cdk Inhibitors).
Οι ειδκοί αυτοί αναστολείς δρούν είτε μπλοκάροντας την ενεργοποίηση της Cdkκινάσης (λόγω πρόσδεσης τους στη Cyc), είτε παρεμποδίζοντας την πρόσβαση του
υποστρώματος στόχου ή του ATP στο σύμπλοκο Cyc-Cdk.

Επαγωγέας
ΑΝΑΣΤΟΛΗ τηςενεργότητας κινάσης
Aναστολέας Cdk(CdkI)
ΑΝΑΣΤΟΛΗκυτταρικούκύκλου
Η επιτυγχάνεται μέσω καταστολήςτης ενεργότητας κινάσης των
συμπλόκων Cyc-Cdk. Μεταξύ τωναρνητικών ρυθμιστών
περιλαμβάνεται μια ομάδα απόμικρού μοριακού βάρους
πρωτεΐνες που ονομάζονταιαναστολείς Cdk (CdkI, Cdk
Inhibitors). Οι αναστολείς αυτοίδρουν είτε μπλοκάροντας τηνενεργοποίηση της Cdk κινάσης
(λόγω φυσικής τουςαλληλεπίδρασης με την Cyc), είτεμπλοκάροντας την πρόσβαση τουυποστρώματος στόχου ή του ATP
στο σύμπλοκο Cyc-Cdk.
Αναστολή ενεργότητας κινασών

(2) Την οικογένεια Cip/Kip που αποτελείται από τρία μέλη. Την p21Cip1, την p27Kip1 και τηνp57Kip2. Οι πρωτεΐνες αυτές αναστέλουν ισχυρά τα σύμπλοκα των Cdk2-CycE και Cdk2-CycA κινασών ενώ παρότι έχει βρεθεί ότι τα σύμπλοκα CycD-Cdk όχι μόνο δεν επηρεάζονται αλλάκαι απαιτούν τις Cip/Kip πρωτεϊνες για σωστή αυτοσυγκρότηση δεν μπορεί να αποκλειστεί ότι
κάτω από συγκεκρίμενες συνθήκες οι Cip/Kip πρωτεϊνες μπορούν να αναστείλουν τηνενεργότητα και των Cdk4-6/CycD.
Οι CdkIs ομαδοποιούνται σε δύο οικογένειες:
(1) Την οικογένεια ΙΝΚ4 (inhibitors of kinase 4) που αποτελείται από τέσσερα μέλη καιειδικότερα τις πρωτεϊνες p16Ink4a, p15Ink4b, p18Ink4c και p19Ink4d. (ή p14ARF το ανθρώπινο
ομόλογο γονίδιο) που αναστέλουν ειδικά τα σύμπλοκα CycD-Cdk4, CycD-Cdk6.

G1 και S Φάσεις του Κυτταρικού κύκλου

G2 και M Φάσεις του Κυτταρικού κύκλου

(p14ARF) p19ARF
INK4A/ARF
p16INK4A
Mdm2
p53pRB
σιγή E2F
Cdks4/6 Cyclin D?
p21
ΑΤΜ
ΑναστολήΚυτταρικού
Κύκλου-Γήρανση
Bmi-1
Bmi-1
Απόπτωση
p27kip1
PIP3-μείωση
PI3KOnco-Ras
Raf, Mek
Μειωμένηπολλαπλασιαστική
ικανότητα
Shc PTEN
ΣυσσώρευσηΚυτταρικώνδιαιρέσεων
? PCNA
?
Cdk2 Cyclins E/A
ΣυσσώρευσηΚυτταρικώνδιαιρέσεων
Τα μονοπάτιασηματοδότησηςτου p53 και Rb

Τα μονοπάτια σηματοδότησης του p53 και Rb