Embed Size (px)
Citation preview

Indian Journal of Biochcmistry & Biophys ics Vol. 37. December 2000. pp. 470-476
-) I
·l(-)o·6'
Induction of polyunsaturated fatty-acid synthesis enhances tolerance of a cyanobact;[ium,-(Cylindrospennopsis raciborskii, to low-temperature
photoinhibition
(;0 ' z· 1* Z y ' k · 1* - 3 2 - I I ---, \ ' tto slros , suzsanna ar 'onyl , Att ila Kovacs, Tibor Farkas, Zoltan Gombos and Gyozo Garab
, Ilnstitute of Pl ant Biology and 21nstitute of Biochemislry,. Biological Research Center, Hungarian Academy of Sciences. S zeg~nd ' Inst itute of Limnology, Hungarian Academy of Sciences.Tihany. Hungary
Accepted all 31 May 2000
( Acyl-lip id desaturation introduces double bonds (unsa turated bonds) at specifically defined pos itions of fatt y !rcros that are esterified to the glycerol backbone of membrane glycerolipids. Desa turation pattern of the glycerolipids of Cy/il/drospe/'ll/opsis raciborskii (c. raciborskii) , a fi lamentous cyanobacterial stra in , was determined in cell s grown at 35°C and 25 °C. The lowering of the growth temperature from 35°C to 25°C resulted in a cons iderable accumulation of polyunsaturated oc tadecanoic fatty acids in all lipid classes. Lipid unsaturation of C. m ciborskii was also compared to Syl/ echocyslis PCC6803. In C. raciborskii ce ll s, a shi ft in growth temperature induced a much more pronounced alteration in the desaturation pattern of all lipid classes than in Syl/ echocyslis PCC6803 . The tolerance to low-temperature photoinhibition of the C. raciborskii cell s grown at 25 uC and 35°C was al so compared to the tolerance of Syl/ ecl/Ocyslis cel ls grown at the sa me temperatures. Lower growth temperature increased the tolerance of C. raciborsk ii cell s but not that of Syl/echocyslis ce ll s. These results strengthen the importance of polyunsaturated glycerolipids in the tolerance to environmental stresses and may give a physiological explanation for the determinative role of C. ra ciborskii strain in algal blooming in the Lake Balaton (Hung~ <" I • ' .... ..,-'
The physical and biochemical properties of glycerolipids depend on the degree of un saturation of the fatty acids I. In the membranes, the level of unsaturation is mod ulated by spec ific enzy me system", fatty-acid desaturases, which introduce doubl e bonds at defin ed positions in the alkyl chain of fatry acids3
. [n cyanobacterial strains acy l-lipid desaturases are presen t, which act on fatty ac ids that are esterifi ed to the glycerol backbone of membrane glycerolipids3
. Poi kil othermic organisms, wh ich are not able to adapt to the changes in environmental conditi ons in other way, modulate the level of unsatu rat ion of membrane lipids according to environmental temperature". Thus, acy l-lipid desaturases are the most efficient regul ators of the extent of lipid un satura ti on in the response to changes in ambient temperature.
Cyanobacteri a are ideal objects for stud ies of temperature-induced changes in the membrane since their fatty acid composi ti on is well known '. On
Author for correspondence: Zsuzsanna Varkonyi *These authors have contributed equall y to thi s article.
lowering the temperature the leve l of un saturated fatty acids in several cyanobacterial strain s, such as Anabaena variabilis, SynechococclIs PCC7002. Synechocystis PCC6714 and Synechocyslis PCC680l markedly increases-l.
The photosynthetic apparatus of photosyntheti c orga ni sms is embedded in thylakoid membrane and surrounded by a lipid matri x. The main constituents of the lipid matrix are glycerolip ids which prov ide the necessary structural background for the functi oning of protein complexes5
.G
, the in serti on of proteins, and their trans location across the membranes 7. In cell ul ar physiology, the unsaturation of membrane glycerolipids plays a key role in the to lerance of I· . . I 8· 10 I IVlng organi sms to ow-temperature stress . n our earlier studies we demonstrated the importance of polyunsaturated glycerolipids in the tolerance of photosynthetic organisms to low-temperature photoi nhibiti on by using transgenic cya nobacterial
. 11·1l d . I' I I 1-1 stralll s . an transgenic ll g le I' p ants .
Cyanobacteria are evolutiona rily adapted to changes of environmental temperatures and cont rol the phys ico-chemical propert ies of thei r membranes.

ZS IROS el. al .. TOLERANCE O F C. RACIBORSKII TO LOW-TEMP ERAT URE PHOTOI NHlBITI ON 471
Thus, Synechocystis PCC6803, a we ll-characteri zed system9
, can be used as reference organi sm in studies of temperature dependence. Cylindrospernwpsis raciborskii is a fil amentous, tox ic blue-green alga inhabiting sub-tropic/tropic wa ter bodies with optimal
growth temperature above 25°C (ref. 15). It appeared in the temperate water of Lake Ba laton (Hungary) a few years ago, where it became the most dominant strain in the seasonal alga bloom.
As part of our e fforts to understand the phys iologica l ro les of the unsaturation of membrane glycerolipids, we studied the e ffects of enhanced level of polyunsaturated g lycerolipids in the to lerance to low-temperature photo inh ibiti on of Cylindrospenllopsis r{lciborskii in comparI son to Synechocystis PCC6803 cell s.
Materials and Methods
Orgallisms and culture conditiollS
Cylindrospen llopsis mciborskii was culti vated in modi fied BG-II medium containing 10% of the ori ginal microe lements. SYllechocystis PCC6803 was grown photoautotrophyca ll y in BG-II medium supplemented with 20 mM HEPES-NaOH (pH 7 .5) as described by Wada and Murata l6. The stra in s we re illuminated with incandescent lamps providing 40 )..lmollm2/sec intensity and aerated by steri le a ir containing I % CO2,
Analysis of/clfry acid m Id chlorophyll Lipids were ex tracted fro m the cell s by the method
of Bligh and Dyer l7. The fatty-acid compos it ion was analyzed as desc ribed by Wada and Murata l6. The lipid extracts were transmethylated in the presence of
methanol containing 5% HCI at 80°C for 3 hr. Fatty ac id methyl esters were separated by gas chromatograph on a FFAP (Supe lco, Bellefonte, PA) capillary column (30 mxO.25 mm i.d .). Chlorophyll concentrati ons were determined by the method of Arnon et 0 11 8.
Measllremellt of photosynthetic {lctivity Photosynthetic oxygen evolving acti vity was
measured in intac t ce ll s with Clark-type e lec trode according to Gombos et {II. 19 without arti fic ial e lec tron donor or acceptor. Actinic li ght was provided by an incandescent lamp in combination with a red optical fi lter (R-62; Hoya G lass, Tokyo, Japan) and an infrared-absorbing fil ter (HA-50 ; Hoya Glass) at an intensity of 600 )..lmoLlml/sec . Photosystem Il activity was measured in the presence of 1.0 mM 1,4-benzoqu inone as e lec tron acceptor.
Photoinhibitiollllleasurelllell ts The opt ical density of the cell s at 750 nm was 0.5 .
The suspension of the ce ll s was illuminated at vari ous temperatures in a thermostated reaction vessel w ith white light of various intensities suppli ed by two incandescent lamps, by aerati on with sterile ai r conta ining I % CO2.
Results
Lipid compositioll of C. m ciborskii Four major lipid classes of C. mciborskii were
identified : monogalactosy l-d iacy lglycerol (MGDG), the major lipid component, diga lac tosyld iacylglycerol (DGDG), sul fo lipid (SL), and phosphatidylglycero l (PG). The di stribution of the indi vidual lip ids in the total lipid ex trac t was simi lar to other cyanobacteria
4
or thylakoids of higher plants20 (data not shown). The growth temperature had no effec t on the di stribution of lipid c lasses.
Effect of growth tempem ture 0 11 fat ty {lc id compositioll (i) SYllechocystis PCC6803
The fatty ac id composition o f total lipid c lasses is summarized in Table I comparing cell cultures grown
at 25 °C and 35°e. The ce ll s growing at lower temperature conta ins a- 18:3 and 18:4 fatty acids. The synthes is of these fatty acids is induced under the growth temperature of 30°e. Thi s is in good agreement with the resul ts of Wada and Murat}l.
Table I-Fatly acid compositi on of glycerolipids from SYllechocyslis PCC6S03 cells grown at 25°C and 35°C, respectively
Growth Fally ac id co mposition, reI. % temperature 16: 0 16: 1 IS:O IS: I IS:2 y- IS:3 a- IS:3 IS:4
25"C 52 4 9 10 IS 5 4 35"C 55 3 9 14 IS tr tr
tr. trace amou nt « 0.5%). The results arc fro m three independent ex peri ments; standard deviations are within ± 2% of indi vidual values.
472 INDI AN J BIOCH EM. BIOPHYS., VOL. 37, DECEMBER 2000
The relati ve percentage of tota l polyunsaturated fatty ac ids was 34% and 32% at 25°C and 35°C, respectively. This resu lt shows that the adaptation to lower growth tempera ture resulted in a relatively small change in the fatty acid compositi on. The level of mono unsaturated fatty acid deri vati ves (palmitoleic ac id + oleic ac id) was not affected by the change in the growth temperature. The proportion of saturated fatty acids (palmitic acid + stearic acid) slightl y increased in the cell s grown at hi gher growth temperature.
(ii ) Cylilldrosperlllopsis raciborskii In C. raciborskii simi larly to the fatty-ac id
compos it ion of Synechocystis cell s the major fatty ac id spec ies were, in that order, palmitic, linoleic , ulinolenic, oleic, y-linolenic, stearic and
octadecatetraenoic acids (Table 2). According to the mode of fatty-acid desaturation , C. raciborskii can be classified as a strain belonging to group 4 (ref. 4). As expected simultaneous ly with a significant dec rease in the saturated-to-unsaturated ratio (sa t%/unsat%) of fatty ac ids, on lowering the temperature the double bond index increased. The level of polyunsaturated fatty acids increased from 34% to 53 % in cells grown at 35°C and 25°C, respectively. The slight variation between cultures has to be noted. With decreas ing growth temperature the level of I inoleic acid dec reased from 19% to 2%, while the levels of ulinolenic acid and octadecatetraenoic acid increased from 11 % to 49%. Th is adaptive alterati on of fatty acid composition was remarkably higher in C. raciborskii cell s than in SYlleclwcystis PCC6803 cells.
Tablc 2-Fatly acid co mposition of glyccro li pids from Cy/il/drospel'l/wpsis raciborskii cell s grown at 25"C and 35"C, respectivel y
Growth Fatly acid composition, rel. % tcmperatu re 16:0 16: 1 18:0 18: I 18:2 y- 18:3 a- 18:3 18:4
25"C 39 4 I 3 2 2 23 26 35"C 45 7 3 I I 19 4 9 2
The results are from three indepcnden t expcrimcnts: standard deviations are wi thin ± 2% of indi vidual val ucs.
~ 20 °C A
~1 30°C B
100
- \ ! ~ c:
80 ~~!~ ~!~! 0
~~~---!--!--i ;:
\ i --i--i - i :I "0 ~2------2 >
~~ CI:I c: 60 ---2 CIJI 0'1 >- 2~ >< 0 -0 40 CIJI
~ ... ~ ~
CIJI > :0:: 20 .!l! (I)
0::
0L-0L---~~20--~---4~0--~---6LO-L~0--~---2~0~~--~4~0--~--~6~0
Duration of incubat ion in light at 1.0 mmollm2/sec (min )
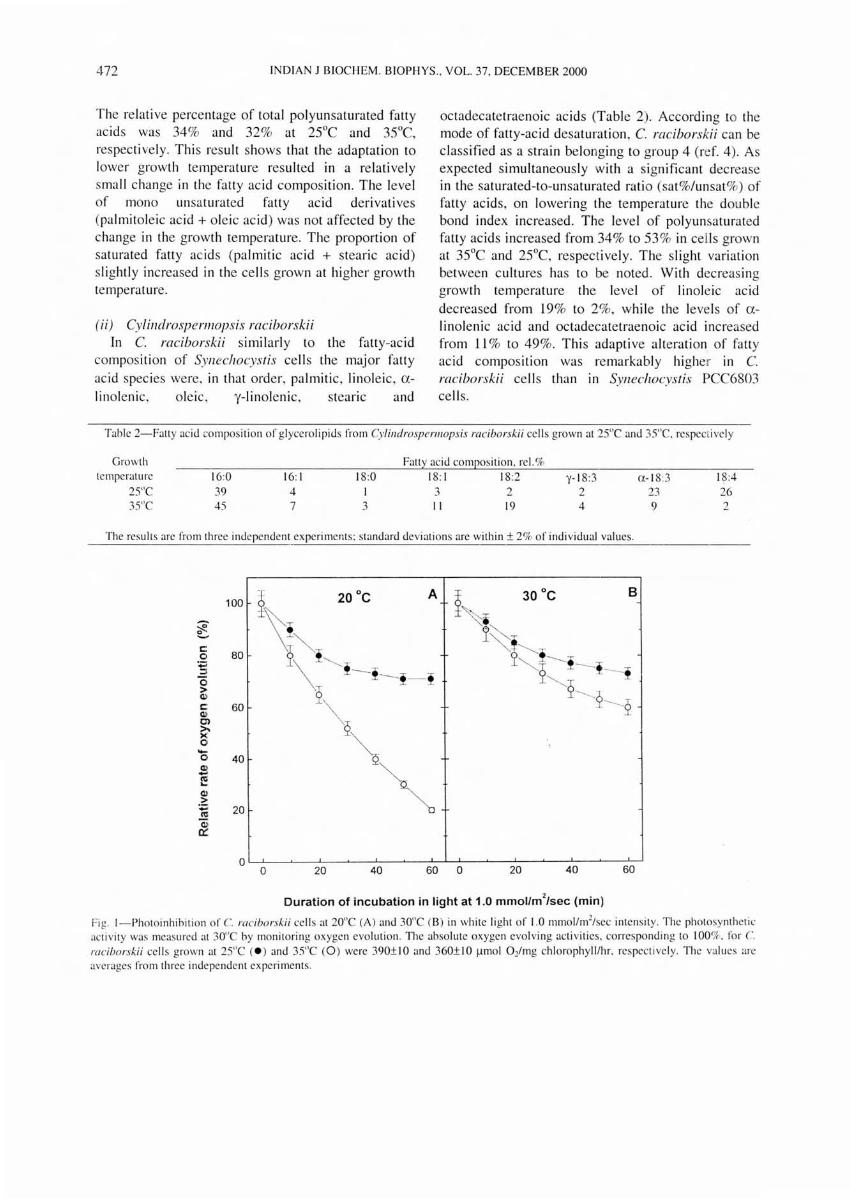
Fic . I-Photoinhibition of C. raciborskii cells at 20"C (A) and 30"C (B) in white li ght of 1.0 mmol /m2/scc intcnsi ty. The photosynthet ic ac~ivi ty was measured at 30"C by monitoring oxyge n evolu tion. The absolute oxygen evolving ac ti vities, correspond ing to I 00%. for C. raciborskii ce ll s grown at 25"C (e ) and 35"C (0 ) were 390± 10 and 360± 10 ~mol O/ mg chlorophyll /hr. respective ly. The values are averages from three independen t experiments.
ZSIROS el. al.: TOLERANCE OF C. RA CIBORS KIITO LOW-TEMPERATURE PHOTOINHIBITION 473
PhOfOillhibitioll of photosynthesis The inhibition of the activity of photosystem II of
C. raciborskii cells was measured following a long term incubation of the cell s grown at 25°e or 35°e at a light intensity of 1.0 mmollm2/sec (Fig. 1). The measurements were carried out by monitoring photosystem II medi ated e lectron transport from H20 to 1,4-benzoquinone. Photoinhibition occurred in both cu ltures grown at 25 De and 35 De and at both temperatures of photoinhibitory treatment, at 20De and 30De. However, cells grown at 25De were more tolerant to high light exposure than the cell s grown at 35 De. The difference was more pronounced when measured at 20De (Fig. LA) than at 30De (Fig. 1.8). 80th types of cells were more sensitive to light at 20De than at 30De. The photosynthetic act ivity was not altered when the cells were kept in the dark for 2 hr (data not shown). For comparison, the inhibition of photosystem II activity of SYllechocystis cells grown at 25De and 35De was also measured under similar conditions. These cells a lso showed higher susceptibility to photoinhibition at lower temperatures, but it was not affected by the growth temperature (data not shown).
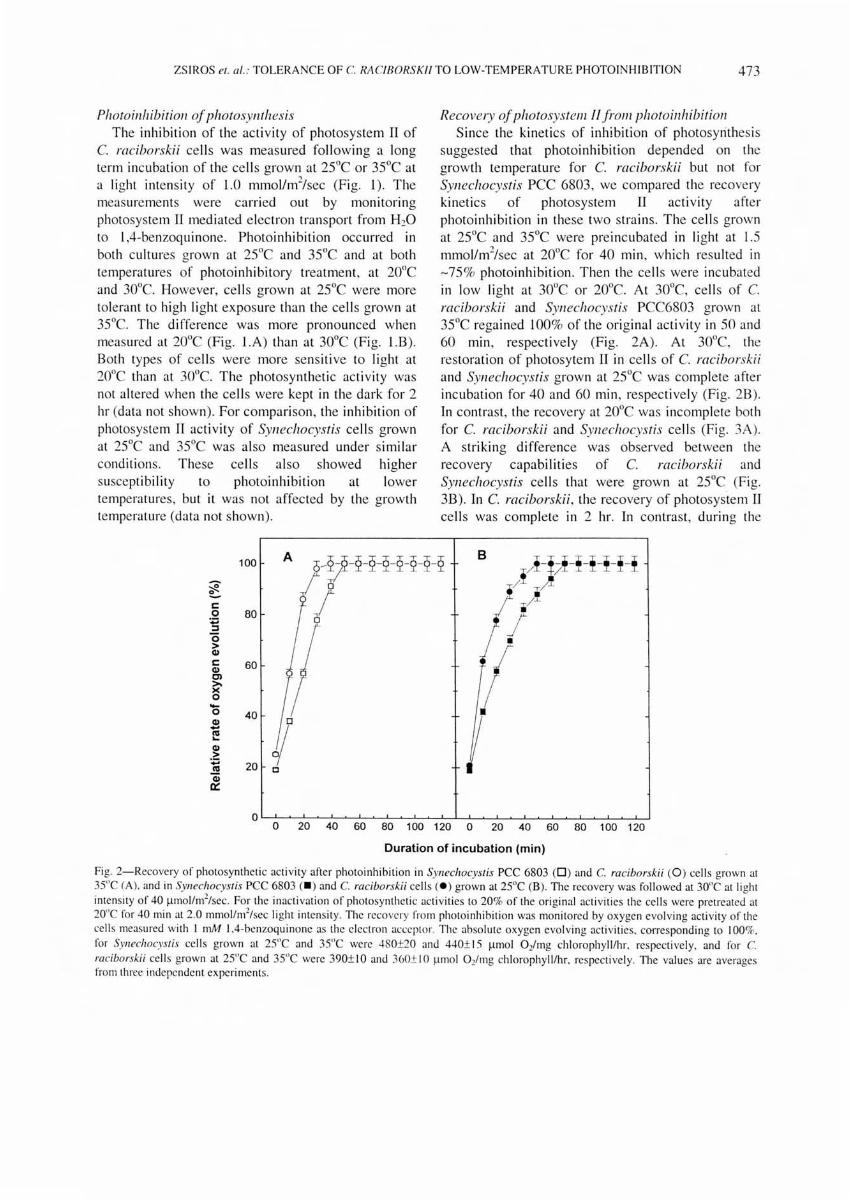
Recovery of photosystem If from photoinhibitioll Since the kinetics of inhibition of photosyrithes is
suggested that photoinhibition depended on the growth temperature for C. raciborskii but not for SYllechocystis pee 6803, we compared the recovery kinetics of photosystem II act IvIty after photoinhibition in these two strains. The cel ls grown at 25De and 35 De were preincubated in light at 1.5 mmollm2/sec at 20De for 40 min , which resulted in -75 % photoinhibition . Then the cell s were incubated in low light at 30De or 20De. At 30De, cells of C. raciborskii and Synechocystis pee6803 grown at 35 De regained 100% of the original ac tivity in 50 and 60 min , respectively (Fig. 2A). At 30De, the restoration of photosytem II in cells of C. raciborskii and Synechocystis grown at 25 De was complete after incubation for 40 and 60 min, respectively (Fig. 28). In contrast, the recovery at 20De was incomplete both for C. raciborskii and Synechocystis cell s (Fi g. 3A). A striking difference was observed between the recovery capabilities of C. raciborskii and Synechocystis cells that were grown at 25 DC (Fig. 38). In C. raciborskii , the recovery of photosystem II cells was complete in 2 hr. In contrast, during the
100 A ~2-~-2-2-2-2-2-2-2
c: o .. :l (5 > (I)
c: (I) Cl >. )( o -o 2 ~ (I)
> .. ..!!! (I)
Q::
/-L 80
60
40
20 f
O~~~~~~~~-L~-L~~~~~L-~~-J~~~-L~-L~
o 20 40 60 80 100 120 0 20 40 60 80 100 120
Duration of incubation (min)
Fig. 2-Recovery of photosynthetic activity after photoinhibition in Syl/echocyslis PCC 6803 (D ) and C. raciborskii (0 ) cells grown at 3S"C (A). and in Syl/echocyslis PCC 6803 (. ) and C. raciborskii cell s ( e ) grown at 2SoC (B). The recovery was fo llowed at 30°C at light intensity of 40 I1mo l/m2/sec. For the inac ti vati on of photosynthetic ac ti vities to 20% of the original activities the cells were pretreated at 20"C for 40 min at 2. 0 mmollnl/sec li ght intensity. The recovery from photoinhibition was monitored by oxygen evolving activity of the cells measured with I mM lA-benzoquino ne as the elec tron acceptor. The absolute oxygen evolving acti vities. corresponding to 100%.
for SYllechocyslis cells grown at 2S"C and 3S"C were 480±20 and 440± 15 I1mo l 0 2/mg chlorophyll/hr. respectivel y, and for C. raciborskii cell s grown at 2S"C and 3S"C we re 390± I 0 and 360± I 0 ~lIno l 0 2/mg chl orophylllhr. respect ive ly. The values are averages from three independent experiments.
474 INDI AN J BIOC HEM. BIOPHYS ., VOL. 37, DECEMB ER 2000
c: o ~ ::J
'0 > QJI
c: QJI 01
~ o .... o CI.l .... ~ CI.l > ~ 10
~
A 100
80
60
40
20
o 20 40 60 80 100 120 0
Duration of incubation (min)
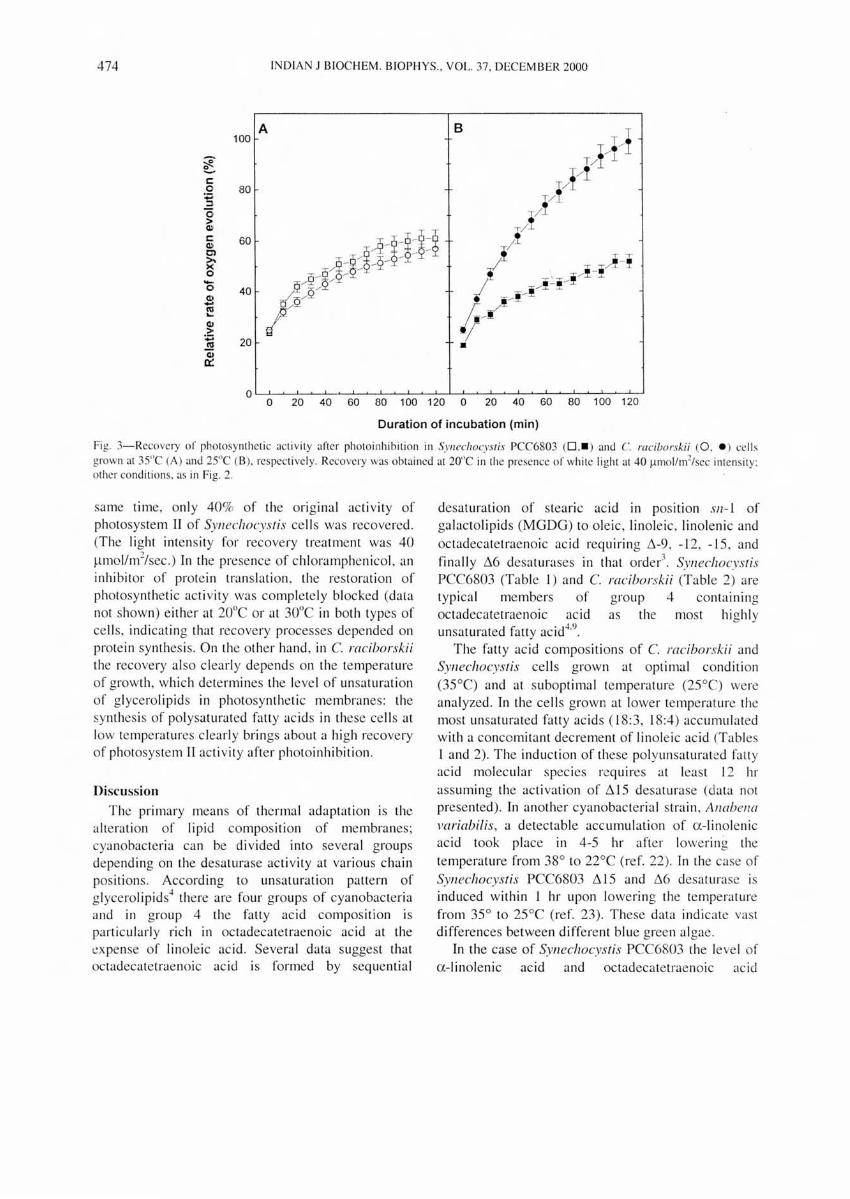
Fig. 3-Recovery of photosyntheti c acti vity after photo inhi bi tion in SYllechocyslis PCC6S03 (0 ,_ ) and C. raciborskii (0 . • ) ce lls grown at 35"C (A) and 25"C (B). respectively. Recovery was obtained at 20"C in the presence of white light at 40 ~.lJnollm2/sec illlensity; other conditi ons, as in Fig. 2 .
sa me time , onl y 40% of the o rig ina l ac ti vity of photosyste m II of Synechocystis ce ll s was recovered . (The light intensity for recovery treatme nt was 40
Ilmolim"/sec.) In the presence of c hl oramphenicol, an inhibitor of prote in translati on, the restorati on o f pho tosynthetic ac ti vity was comple te ly bl oc ked (data not shown) e ither at 20°C or at 30°C in both types of ce ll s, indi cating that recovery processes depended on prote in synthes is. On the other hand , in C. raciborskii the recovery a lso clearl y depe nd s on the temperatu re of growth , which dete rmines the level of unsa turati on of glycero lipid s in photosynthe tic me mbranes: the synthes is of po lysa turated fatty ac id s in these ce ll s at low te mperatures c lea rl y brings abo ut a hi gh recovery of photosyste m II ac ti vity afte r photo inhibitio n.
Discussion
The primary means o f thermal adaptati on is the a lte ration of li pid compos itio n o f me mbranes ; cyanobac te ri a ca n be divided into several gro ups depending on the desa turase acti vity at va rio us chain pos iti ons. According to un sa turation patte rn of glycero lipids4 the re a re fo ur g ro ups of cyanobac te ria and in gro up 4 the fatty ac id compos it ion is parti c ul a rl y ri ch in oc tadecate traeno ic ac id at the ex pense o f lino le ic ac id . Several data suggest tha t octadecatetraeno ic ac id is fo rmed by sequentia l
desa turation of stea ri c ac id in posi ti on sll- I of ga lacto lipids (MGDG) to o le ic, linole ic, linolenic and
oc tadecate traeno ic ac id requiring ~-9, -1 2, -I S, and
fin all y ~6 desa tura ses in that o rder). SYllerhocystis PCC6803 (Table 1) and C. r{/(;iborskii (Table 2) a re typi cal me mbers of group 4 conta ining oc tadecate traeno ic ac id as the IllOS t hi ghly unsa turated fa tty ac id4
.9
.
The fatty ac id compos iti ons of C. raciborskii and Synechocystis cell s grown at optima l cond iti on
(3S °C) and at suboptimal te mperature (2S °C) were analyzed. In the ce ll s grown at lower te mperature the most unsaturated fa tty ac ids (18:3 , 18:4) accumul ated w ith a concomitant decre ment of lino le ic ac id (Tab les 1 and 2). The inductio n of these po lyunsa turated fatty ac id molecular spec ies requires at leas t 12 hr assuming the ac ti va ti on of ~ lS desatu rase (data not presented). In anothe r cyanobac teria l stra in , AI/abel/a voriobiiis, a de tec table acc umul ati on of a -lino lenic ac id took pl ace in 4-S hr after loweri ng the
te mperature fro m 38° to 22°C (ref. 22). In the case of
SYllechocystis PCC6803 ~l S and ~6 desa turase is induced within 1 hr upon lowering the te mperature from 3So to 2SoC (re f. 23). These data ind ica te vast diffe re nces be tween di ffe rent blue g reen a lgae.
In the case of SYllechocystis PCC6803 the level of a -lino le nic ac id and oc tadecate traeno ic acid

ZS IROS el. at.: TOLERA NCE OF C. RACIBORSKII TO LOW-TEMPERAT URE PHOTOI NHIBITION 475
increased parallel with lowering the temperature from 42°C to 22°C (ref. 9). In contrast, the transformation of Anacystis Ilidulalls R2-SPc, sensitive to chi lling temperatures, with the gene encoding fd2 desaturase resulted in an enhancement of linoleic ac id and an increased tolerance to low temperature stresses8
. In C. raciborskii cell s, the levels of a -linolenic acid and octadecatetraenoic acid increased, in stead of linoleic ac id, when the cell s were exposed to suboptimal growth temperatures. Some data show that in higher plants, a -linolenic ac id is involved in the cold tolerance'!.". St. John et al.25 demonstrated that the inhibition of the formation of linolenic acid led to increased chilling sensiti vity of several cerea ls. In addition, Sommerville and Browse26 suggested that an increase in the level octadecatetraenoic ac id in higher plants correlated with chilling tolerance. Kodama el al. '27 found that trienoic fa tty acids were acc umulated in tobacco following ~15 desaturase induced transformati on. In C. m cihorskii cell s grown at lower temperature, although the levels of octadecapolyenoic ac ids were increased, there was no indicat ion of increased chilling tolerance in the darkness.
The change in the rati o of unsaturated to saturated fa tty ac ids can result in certain modification in membrane structure, which can ex plain the alterati on of phys iologica l properties of the photosynthetic organi sms8
. In the glycerolipids of SYllechocystis cell s the unsatisat ratio showed very minor change when the cell s were transferred from 35°C to 25°C (Table 3) and the change in doubl e bond index (OBI) was negligible as well. In contrast, fo r C. racihorskii the growth temperature dramaticall y affected the unsatisat ratio and the DBI of glycerolipids (Table 3). These find ings suggest that while the unsaturation level of glycerolipids from Synechocyslis is unchanged C. m ciborskii cell s show a more fl exible adaptati on to the temperature of environment on the level of lipid unsaturati on.
Tn our ea rli er studies we in ves tigated the effec t of polyunsaturated glycerolipids on the susceptibility of photosyntheti c organ Isms to low-temperature photoinhibition using transgenic cyanobacteri al
. 8 10'8 I h d " d C strall1s' '-. n t e present stu y we II1 ves tl gate . mciborskii . a cyanobac teri al strain which was found to be ex tremely stress res istant in natural environmental conditions. Therefore, we compared their susceptibility to photoinhibition at low temperature with that of the well -c harac terized SYll echocyslis cell s; and also compared the
Table 3- Characte ri stics o f glycerolipid co mposi tion of the cyanobacterial st rains grown at d ifferent temperatu res.
Strains Growth unsallsat(% ) OBI temp.
Syllechocyslis 25"C 0.89 ± 0.05 1.09 ± 0.05 PCC6803 35"C 0.79 ± 0.04 0.94 ± 0.05
C. raciborskii 25°C 1.50 ± 0.07 1.90 ± 0.06 35"C 1.08 ± 0.05 1.03 ± 005
unsallsat(%), represents the rati o between unsaturated and saturated fa tty ac ids
OB I, double bond index was calcu lated fro m the percentage of unsatu rated fatty ac ius and mul tiplied with the number of double bonus
susceptibility of cells grown at different temperatures. Results of these experiments (Figs. 2 and 3) revealed that the extent of the photoinhibition was higher in the C. raciborskii cell s grown at higher temperature than in the cell s grown at lower temperature possessing considerably higher amount of polyunsaturated glycerolipid components. These findings indicated a good correlation between the amounts of polyunsaturated lipid molecules and the ability of cell s to protect photosystem II aga inst lowtemperature photoinhibition.
Recent studies on the photoinhibition and recover/ 9 have demonstrated that these processes are related to the degradati on of the D 1 protein and reconstituti on of the de novo synthes is of Dl protei n of photosystem II. The Dl prote in is embedded in the membrane. Thus, the incorporation of prec ursor of 01 to the photosystem II complex, and the process ing of the precursor of the 01 protein to the mature 01 protein depend on the membrane environment. Prev iously it was suggested that the processi ng of pre-O 1 is related to the unsaturati on level of glycerolipids in the membrane3o
.31
. Ex periments to determine how thi s process is modul ated by the level of unsaturation of membrane lipids in C. mciborskii ce lls is in progress .
In summary, we in vestigated the phys iologica l consequences of the altered growth temperatures in the adaptati on to hi gh li ght at low temperature. ]n the case of SYll echocyslis PCC6803 the growth temperature did not affect the tolerance to lowtemperature photoinhibition. We could reach alterati on of the tolerance to low-temperature photoinhibition by the complete elimination of polyunsaturated glycerolipids l3
. In C. mciborskii. a decrease in the growth temperature enhanced the

476 INDIAN J BIOCHEM. BIOPHYS. , VOL. 37, DECEMBER 2000
tolerance to low-temperature photoinhibition. The results of fatty acid analyses revealed that the variations 111 the level of polyunsaturated glycerolipids of the thylakoid membranes as a function of growth temperature were much higher in C. raciborskii than SYllechocystis PCC6803. These data show that 111 natural conditions the photosynthetic organisms can manipulate their tolerance to stress conditions by adjusting the lipid unsaturation; as demonstrated in this work, this influences significantly the degree of protection against low temperature photoinhibition. In particular, our data point to the importance of polyunsaturated glycerolipids in the recovery process after photoinhibition of photosynthesisl 3
.
Acknowledgement This work was supported by the Balaton
Secretariat of the Prime Minister's Office and by grants from the Hungarian Scienti fic Research Fund (OTKA T023794, T025804, T030631 and T020293 ).
References I Quinn P J ( 1988) in Physiolog ical Reg ulation of Melllbrane
Fluidity (A loia R C, Curtain C C & Gordon L M eds) Vol 3 pp 293-321 , Alan R Li ss Inc. New York
2 Cossins A R ( 1994) in Telllpera/llre Adaptation of Biological Melllbranes (Cossins A Red) pp 63-76, London, Portland
3 Murata N & Wada H ( 1995) Biochelll J 308, 1-8 4 Murata N, Wada H & Gombos Z (1992) Plant and Cell
Physiol 33, 933-941 5 Siegenthaler P-A ( 1998) in Lipids in Photosynrhesis:
Structure FUllctioll and Generics (Siegenthaler & Murata eds) pp 119-144, Kluwer Academic Publi shers, The Nederlands
6 Doyle M F & Yu C-A ( 1985) Biochilll Biophys Res COllI/nUll 13 1. 700-706
7 Van't Hof R, Van Klompeburg W, Pilon M, Kozubek A, De Korte-Kool G, Demel R A, Weisbeek P J & De Kruijff B ( 1994) J Bioi Chelll 268 4037-4042
8 Nishida I & Murata N (1996) Annll Rev Plant Physiol Plant Mol Bioi 47, 541-568
9 Wada H, Gombos Z & Murata N ( 1990) Nature, London 347. 200-203
10 Gombos Z & Murata N (1998) in Lipids ill Photosynthesis: StmCl/lre FlInction and Genetics (S iegenthaler & Murata eds) pp 249-262 Kluwer Academic Publi shers. The Nedcrlands
II Gombos Z, Wada H & Murata N ( 1992) Proc Natl Acad Sci USA 89,9959-9963
12 Gombos Z, Wada H & Murata N ( 1994) Proc Natl Acad Sci USA 91 , 8787-8791
13 Tasaka Y, Gombos Z, Nishiyama Y. Mohanty P. Ohba T, Ohki K & Murata N (1996) EMBO J 15, 6416-6425
14 Moon B Y, Higashi S-I , Gombos Z & Murata N ( 1995) Proc Natl Acad Sci USA 92, 6219-6223
15 Vinogradska T A (1974) Ukrainian Botanical Review 3 1. 733-739
16 Wada H & Murata N (1989) Plant Cell Physiol 30, 971-978
17 Bligh E G & Dyer W J ( 1959) Can J Biochelll Physiol 37. 911-917
18 Arnon 0 I, McSwain B D. Tsuji moto H Y & Wada K (1974) Biochilll Biophys Acta 357, 231-245
19 Gombos Z, Wada H & Murata N ( 1991 ) Plant Cell Physiol 32,205-211
20 Webb M S & Green B R (1991) Biochinl Biophys Acta 1060. 133-158
21 Wada H & Murata N (1990) Plallt Physiol 92, 1062-1069
22 Sato N & Murata N (1980) Biochim Biophys Acta 619, 353-366
23 Ki s M, Zsiros O. Farkas T. Wada H. Nagy F & Gombos Z ( 1998) Proc Natl Acad Sci USA 95. 4209-4214
24 Graham 0 & Patterson B 0 (1982) Anll Rev Plallt Physiol 33. 347-342
25 St John J B, Chri stiansen M N. Ashworth E N & Gentner W A (1979) Crop Sci 19, 65-69
26 Sommerville C & Browse J (199 r) Science 252, 80-87
27 Kodoma H, Hamada T, Horiguchi G, Nishimura M & Iba K ( 1994) Plant Physiol l05 , 601-605
28 Varkonyi Zs, Zsiros 0 & Gombos Z (1996) J Scientific & Indllstrial Res 55. 658-668
29 Aro E-M. Virgin I & Andersson B ( 1993) Biochilll Biophys Acta 1143 , 113-134
30 Kanervo E, Aro E-M & Murata N ( 1995 ) FEBS Lell 364. 239-242
31 Gombos Z, Kanervo E, Tsvetkova N, Sakamoto T, Aro E-M & Murata N (1997) Plant Physiol 115 , 551-559







![日本植物生理学会...S556 S557 S558 S559 S560 *422, sp. PCC6803 7 -f 7 & (Anabaena sp. PCC7120, "Synechococcus elongatus" Toray, G]oeobacter wolaceus PCC7421) Y C. Z Bchl Rb](https://img.pdfslide.net/doc/110x75/60ac83c79c38990c4d313409/oeccc-s556-s557-s558-s559-s560-422-sp-pcc6803-7-f-7-.jpg)

















