Embed Size (px)
Citation preview

-- ---------.
GENETIC ENGINEERING OF INSECT TOLERANCE IN VAUruT: IMPROVED TRANSFORMATIONEFFICIENCY.
Abhaya M. Dandekar, Gale H. McGranahan, Patrick V. Vail, Sandra L. Uratsu,Charles Leslie, J. Steven Tebbets and Darlene Hoffman
ABSTRACT
The genes encoding the insecticidal crystal proteins (ICPs) of BacIllusthuringiensis are good candidates for protecting walnut plants against insectpests. Ve have estimated the dose response of three important walnut pests(codling moth, navel orangeworm and Indianmeal moth) with two highly purifiedinsecticidal proteins of Bacillus thurJ.nglensis and show that these three insectsare highly sensitive to these proteins. Sublethal effects of the insecticidalproteins on development and reproduction are also being determined. In order thatthe genes encoding these proteins be transferred to walnut we have developed agene transfer system.
The Agrobacterium-mediated gene transfer systemthat relieson repetitiveembryogenesis to regenerate transgenic walnut plants was made more efficient byusing a combination of a selectable (kanamycin resistance) and a scorable
(beta-glucuronidase, GUS) marker gene. The transformation with DNA containing
these two genes greatly facilitated early screening and selection. Introduction
of the GUS marker gene allows positive identification of transformed secondaryembryos as early as 5-6 weeks after inoculation. DNA analysis of arepresentative subset of lines (n-13) derived from secondary embryos confirmed
transformation and provided evidence for multiple insertion events in singleinoculated primary embryos.
Ve have now used this gene transfer system to introduce a gene [cryIA (c)gene of BacIllus thurlnglensIs] conferring tolerance to insect pests into walnut
embryos. These embryos have been regenerated to form shoots which are beinggrafted onto seedling rootstock in the greenhouse. Ve have applied for a permitto grow these transgenic plants in a field test plot.
OBJECTIVES:
1. Feeding studies with ICPs incorporated into the diet2. Analysis of transgenic GUS embryos and plant tissue3. Transformation with Agrobacterium containing genes encoding ICPs4. Induction of somatic embryos from nucellus and ovules of Chandler5. System improvements
L. Feeding Studies withBT incoI'porated into the diet.
The insecticidal activityof BacIllus thuringiensis(BT) resides in thespore of this bacterium and occurs as a crystalline inclusion body containingone or more insecticidalcrystal proteins (ICPs) also known as the delta-endotoxin (reviews: Aronson et al., 1986; Whiteley and Schnepf, 1986; H6fte and
Whiteley, 1989). Typically the insecticidal crystal is ingested by a feedinginsect. The ingested crystal is first solubilized in the alkaline gut of the
insect and then acted upon by protease(s) inside the midgut to release a fragment
12
- --- --

of the insecticidal crystal protein (ICPF) which kills the insect (H6fte andWhiteley, 1989). Recent evidence indicates that the ICPF binds to specific cellreceptors on the surface of cells that line the midgut (epithelium cells) ofsusceptible insects (Hofmann et al. 1988). Non-target organisms presumably donot possess the specific cell receptors that are required to bind ICPF and arethus unaffected by this protein.
Different strains of BacIllus thuringiensis make ICPs that are effective
against different groups of insect pests (Aronson et al., 1986; Whiteley andSchnepf, 1986; H6fte and Whiteley, 1989). At least 40 genes encoding ICPs
effective against Lepidopteran, Dipteran and Coleopteran insects have been
isolated and their DNA sequence compared (H6fte and Whiteley, 1989). Genes of
BacIllus thuringiensis encoding ICP active against Lepidoptera can be classified
into 8 groups based upon differences in their gene structure and that of theencoded protein products i.e., cryIA(a), cryIA(b) , cryIA(c), cryIB, cryIC, cryID,
cryl1A and cryl1B (H6fte and Whiteley, 1989). We have obtained from Dave Fischoffat Monsanto, purified preparations of ICPF from E. colI expressing the productsfrom cryIA(b) and cryIA(c) genes, encoding an ICP of BT strains HD-1 and HD-73
respectively. Last year we showed preliminary data on the incorporation of theseproteins into diet to test dose response of the target insects
1... Analysis of transgenic GUS embryos and plant tissue
One of the major drawbacks of our gene transfer system has been the lowefficiency of transformation and the time and labor involved in identifyingtrans formants . Last year we reported the improvement of the system using vectors
that carried two marker genes. We used a bacterial gene encoding the enzymebeta-glucuronidase (GUS) (Jefferson et al., 1987) in addition to the geneencoding resistance to the antibiotic kanamycin in a binary vector system in
Agrobacterium. Last year we presented data that showed that exposure oftransformed embryos to kanamycin shortly after inoculation with the Agrobacterlumenhanced the frequency of transgenic embryos recovered. The GUS gene product wasexpressed and reliably detected in transformed walnut tissue and could be used
as an excellent scorab1e marker. Since the observed transformation frequencieswe reported were quite high (15-30%) we were interested to determine if thetransformed embryos represented independent transformation events.
1. Transformation with Agrobacterium containing BT genes
As mentionedabove the genes of Bacillus thurlnglensls encoding ICP activeagainst Lepidoptera can be classified into 8 groups based upon differences intheir gene structure and that of the encoded protein products. We have evaluatedtwo of these i.e., cryIA(b) and cryIA(c) in the feeding studies with walnuttarget pest species and found both to be effective. We focused our effort to test
one such vector (pWAG139) carrying the cryIA(c) in a construct similar to that
described by Vaeck et al., 1987. The plasmid vector pWB139 is described below.In this first generationvector the cryIA(c)gene sequencesare fused to thecoding region of the gene encoding APH(3')11 making a fusion proteinproductthatgives the kanamycin resistance phenotype as well as insect toxicity as describedearlier (Vaeck et al., 1987).
13
- --

The gene transfer system we have developed for walnuts relies onrepetitive somatic embryogenesis to regenerate embryos and plants containing atransferred gene Le., transgenic plants (McGranahan et a1., 1988). Theembryogenic tissues we used to develop the system were originally obtained fromthe cotyledons of developing zygotic embryos and therefore were not geneticallyidentical to the parent cultivar. For our system to be applied to geneticimprovement of walnut cultivars it is important that we have the means to inserta desired gene into a cultivar of choice and not into its offspring. One meansof achieving this goal is to initiate repetitively embryogenic cultures directlyfrom the cultivar of choice. In other crops, nucellus has been used to initiateembryogenic cultures (George and Sherrington, 1984).
Our objective was to determine if embryogenic cultures could be initiatedfrom nucellus or ovule tissues of the walnut cultivar.
~ System imnrovements
Work directed towards improvement of the gene transfer system includedimprovements in the KAN screening procedure for embryos and development of a KANscreening procedure for shoots.
Although use of the GUS marker gene (Jefferson, 1987) vastly improves theefficiency of transformant selection (Dandekar et sl., 1989; McGranahan et sl.,1990), the GUS marker is not available with all Agrobscterium vectors. Theinsertion of the BT encoded ICP reported here uses the KAN marker instead of theGUS marker. Kanamycin screening conducted to date has employed the lowestkanamycin concentrations at which differences were detectable in order to recoverall possible trans formants , even those with low expression, and also because theupper limits of kanamycin tolerance could not be discerned until knowntransformants were obtained and could be used for testing. Screening trialsreported previously (McGranahan et sl., 1988) employed 75 mg/l kanamycin and workreported here employs 100 mg/l kanamycin. The disadvantage of low kanamycinconcentrations is that some non-transformants continue to emerge, requiring timeand costly repetitive screening. We therefore examined the suitability of usingincreased kanamycin concentrations for transformant screening.
The method we have pursued in developing a system for transforming cultivarmaterial is induction of embryogenesis from parental tissues. The embryogenesissystem is advantageous in that it eliminates possible chimeras but other methods,including direct DNA injection with a particle gun (Klien et sl., 1987) andAgrobscterium vectored transformationof shoot tips (Ulianet sl., 1988) couldbe used to transform shoot material directly. Transformation by these methodsusing available gene constructswould require a kanamycin selection methodapplicable to shoot cultures. Our objectivewas to develop such a screeningprocedure.
14
- -

PROCEDURES:
1...
The ICPs encoded by the cryIA(b) and cryIA(c) genes of Bacillusthuringiensis RD-1 and RD-73 were used in the dose response experiments. Thehighly toxic ICPF corresponding to the N-termina1 1/3 of the protein wereobtained from Dave Fischoff at Monsanto. The purified ICPFswere obtained aftera column chromatographic separation of a tryptic digestion (with the proteaseTrypsin) of the individual ICP preparations. Purified ICP preparations wereobtained by over expression of the cryIA(b) and cryIA(c) genes in E. coli. Thepurified ICPFs were stored in 100 mK Sodium Carbonate (pH 10.0), 50 , Glyceroland 10 mK DTT (dithiothreito1) frozen at -SO.C. Dilutions were made in 100 mKSodium Carbonate buffer (pH 10.0) containing 10 mK DTT. Two controls were used1) plain buffer and 2) buffer with DTT to ensure that neither the buffer or DTTwere toxic to the insects.
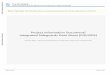
The test insects Cydia pomonella, Tortricidae - codling moth (CM), Amyeloistransitella, Pyra1idae - navel orange worm (NOW) and Plodia interpunctella,Pyra1idae - Indian meal moth (IHK) were reared and the bioassays conducted atthe Horticultural Crops Research Laboratory at Fresno, California. Thesusceptibility of each of these insect species to the BT encoded ICPs CryIA(b)and CryIA(c) were determined by feeding (per os) experiments. Serial dilutionsof each protein were either layered onto the surface or incorporated into anagar-based diet (Bioserv #9370) and the dose was expressed as nanograms per sq.cm or cm3 of diet.
a.) Surface inoculations: The agar based dietwas distributedin 2 m1 vials(analytical beakers) containing snap caps and into each was introduced one m1of diet. Between 5 to S dilutionsof each ICPF was made in the buffer (100 mKSodium Carbonate buffer (pH 10.0) containing 10 mK DTT) and 10 1£1 of eachdilution was spread over the surface of the agar (approximately 100sq mm). Oneneonatal larvawas placed in each vial (containingca. one m1 of surfaceinoculated diet) and about 55 to 160 larvae were tested for each dilution and
for each insect species. Dose response evaluations were made daily by observingthe larvae for mortality and extent of development. The controls were larvae
incubated on pure diet and on diet containing 101£1 of buffer without ICPF. Thelarvae were incubated at 80.F for a period of 2 to 3 weeks for theseobservations. The data obtained from replicated tests was analyzed by Probit
analysis to determine the dose (LD) required to reach 50 or 95% mortality (Table1).
b.) Diet incorporation: These tests were carried out essentially asdescribed above the only difference being that the dilution of ICPF wasincorporated into the diet prior to solidification (while'the diet was still a
warm liquid). Observations, replicates, data acquisition and analysis were alsoidentical to that described above; however, the dose as expressed asnanograms/cm3(Table2a) or I£glm1 (Table 2b).
c.) Sublethal effects: The insect Amyelois transitella, Pyralidae - navelorange worm (NOW) was used for these studies as was the surface inoculationtechnique described above. The dilutionsof the two ICPFs. CryIA(b) and CryIA( c) ,
15--- -

were prepared as described above and 10 pl amounts were spread on the surfaceof the solidified diet to give the following sublethal doses; 0, 0.1, 0.5, 1.0
ng per sq. mID. A single neonate larvae was introduced per vial. 'ft1esetests arein progress to determine effects of sublethal exposure (per os) on insect growth,rate of development, and mating and fecundity (Table 3).
L. Analysis of transgenic GUS embryos and nlant tissue
a.) DBA analysisof transformedembryos: Embryo DNA was isolated andanalyzed by Southern blot analysis as previously described (Dandekar et al.,1988, McGranahan et al., 1988). The Southern blots were hybridized sequentially
with two probes, one for detecting the right border and the other for detectingthe chimeric kanamycin resistance gene. The probe used to detect the kanamycin
resistance region has been previously described (Dandekar et al., 1988,McGranahan et al., 1988). To detect the right border sequences a 450 bp fragment
from pCGN154l obtained after digestion with the restriction endonucleases HindIIIand BglII (K. McBride, Calgene Inc., unpublished) containing bp 13990-14273 of
pTIA6 (Barker et al.,1983) was used as a probe. Apart from the 283 bp border
region this fragment contains, 284 bp of DNA from pUC18 (bp 396 to 680).Processing of T-DNA occurs at position 14062 (Albright et al., 1987) leaving onlya 72 bp (13990-14602, Barker et al. ,1983) homology region with the right borderof T-DNA from pCGN7001. The right border of T-DNA from pCGN7314 has 356 bp
region of homology upon processing, that would include in addition to the 72 bpborder region described above, a 284 bp region from pUC18. DNA fragments usedas probes were obtained by removing the appropriate DNA fragments from low melt
agarose gels. The DNA fragments were separated from the agarose by phenolextraction followed by alcohol precipitation as described by Ausube1 et al.,
1988. Purified DNA fragments were then labelled using the random primed reaction
(Feinberg and Vogelstein, 1983 and Feinberg and Voge1stein, 1984) using a
Boehringer Mannheim kit. Hybridization, washing and autoradiography were asdescribed earlier (Dandekar et al., 1988, McGranahan ~ 41., 1988).
w o
a.) Vector.: The two binary p1asmids pWB139 and pWB149 were constructed by
Dr. Wayne M. Barnes, Dept. of Biological Chemistry, Washington University Schoolof Medicine, 660 S. Euclid, St. Louis, Missouri 63110. The complete DNA sequence
analysis of the T-DNA segments of these two p1asmids has been completed (Barnesunpublished data). A map locating various fragments of the T-DNA region is shown
in Fig. 1. The plasmid backbone of both pWB139 and pWB149 is the plasmid pRK252
constructed in Dr. Don Helinski's laboratory at University of California, SanDiego. pRK252 is a broad host range plasmid capable of replication in both E.
coli and Agrobacterium and contains a gene for tetracyclineresistance forgenetic selection.The plasmid pWB139 contains a wound inducible gene thatencodes for a fusion protein containing insecticidal activity of the ICP from
the RD-73 strain of Bacillus thuringiensis with the kanamycin resistance gene
product APH(3')II. The 6.2 kb long T-DNA segment of pWB139 contains, the right
border (bp 005-646), nopa1ine synthase gene 5' (bp 1638-1671) and 3' (bp 5038-5721, Fig. 1) DNA fragments from the Ti plasmid pTiT37. The DNA fragments locatedbetween bp 646-1632 and 6492-7377 were obtained from the 35S promoter of the
cauliflower mosaic virus (CaKV184l). The sequences in the region of bp 4308-5023are a fragment of the tomato protease inhibitor I gene and contain the sequencesof exon 3 and part of the 3' region. This region is responsible for the induction
of this gene by wounding and is a response that is mediated through PIIF
16
- - --

(protease inhibitor inducing factor) in plant tissue. The insecticidal protein
coding region is located between bp 1678-3527 (Fig. 1) and this regioncorresponds to the N-termina1 portion of the cryIA(c) gene of Bacillus
thuringiensis strain RD-73 (H6fte and Whiteley, 1989). This N-termina1 regionof the Bt-ICP has been fused (translation fusion) to the coding region of the
Tn5 kan gene (bp 1566-2345; Beck et al., 1982) and these sequences are locatedat the following position of the T-DNA of pVB139 , bp 3529-4308. The plasmid
pVB149 is identical to pVB139 with one exception it does not contain the codons
encoding ICP but only those encoding APH(3')II and serves as a control in ourexperiments. The T-DNA in pVB149 is about 4.4 kb long and structurally isidentical to pVB139 with a deletion of the cryIA(c) coding region. Both pVB139
and pVB149 were introduced into WAgll via the triparenta1 mating procedure (Dittaet al., 1980).
b. ) Inoculation of embryos and selection of transformants: Somatic embryos
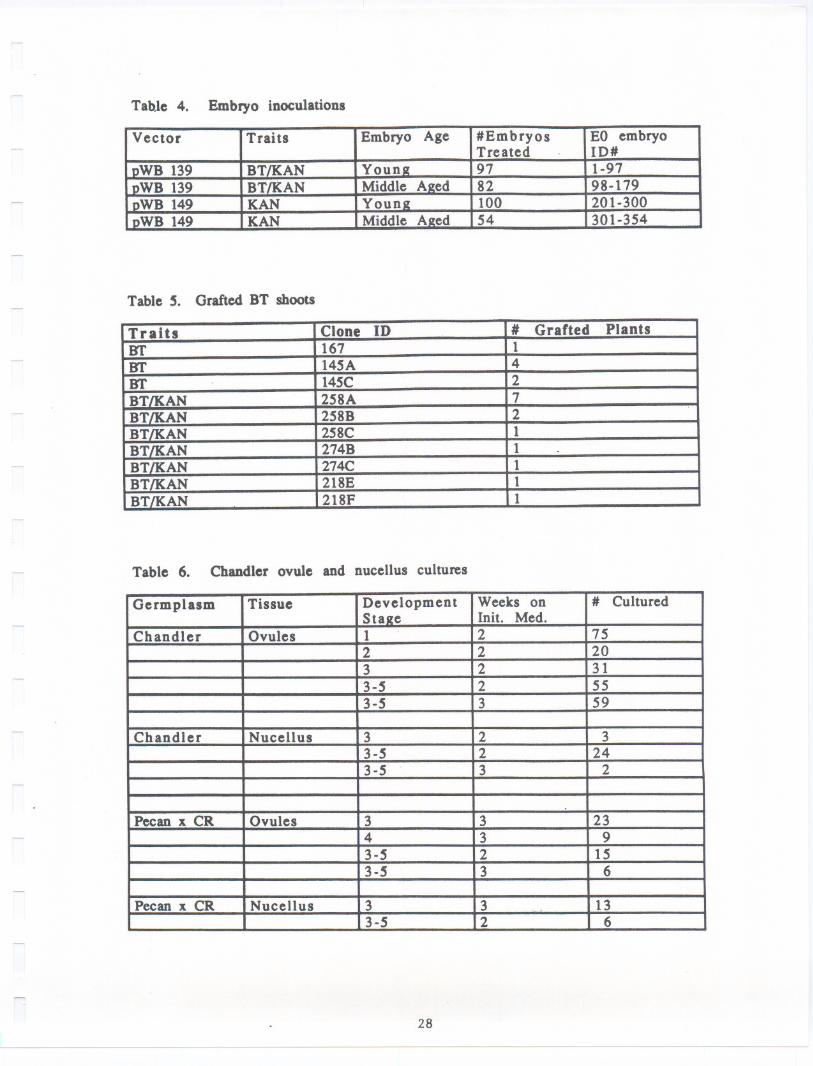
of a repetitively embryogenic culture line designated "SU2", originally derivedfrom the cotyledons of an open pollinated nut of Sun1and were used forinoculations. Two types of embryos, young white embryos and middle aged cream
colored embryos, were chosen for treatment (Table 4). Older, browning embryoswere not used. The embryos were inoculated by immersion for 10-20 minutes in
523 medium (Rodriguez and Tait, 1983) containing 100 pM acetosyringone and 2.5x 108 cel18/m1 Agrobacterium inoculum. Embryos were then blotted briefly on
sterile filter paper to remove excess inoculum and plated to DKW basal medium
(Driver and Kuniyuki, 1984) containing 100 pM acetosyringone (AS medium). After
24 hours, embryos were transferred to DKW basal medium containing 500 mg/Lcefotaxime (CEF medium). Four days later they were transferred to fresh plates
of the same medium. Twelve days after the initial exposure to inoculum the
embryos were transferred to DKW basal medium containing 500 mg/L cefotaxime and
100 mg/L kanamycin (KAN/CEF medium) and subsequently transferred on this medium
every 7-10 days.
Embryos which showed no further production of new somatic embryos onKAN/CEF medium were discarded between two and five months post-inoculation.Embryo clones which still showed growth and multiplication at 5 months wereretained and embryo samples from these clones were analyzed biochemically toconfirm transformation.
Several embryos were then germinated from each of these surviving embryo
clones. The shoot tip was cut from each germinated embryo and micropropagated,
resulting in individual shoot cultures, each derived from a single germinated
embryo. Each of these shoot cultures was labeled with the EO# of the embryo
clone from which it originated and a letter designation (A,B, C, etc.) indicating
the individual germinated embryo from which it was derived.
In order to provide transformed material for greenhouse and field testing
a method for grafting tissue cultured shoots onto seedling rootstock was
developed. Young seedling rootstocks were germinated in pots in the greenhouseand grafted soon after emergence. Tissue cultured shoots were washed carefully
to remove all medium and sugars, cut with razor blades, and cleft grafted tothe seedling rootstocks. Grafts were wrapped in parafi1m and kept in closed
containers or plastic bags at high humidity for several weeks. Tissue cultured
shoots desiccate easily so lids on the containers were gradually opened over a
17
- --- - ---

period of several weeks to allow the plants to successfully acclimate togreenhouse conditions (Table 5).
c.) Analysis of transformed embryo.: DNAwas isolated from the 7 embryolines, 3 that were transformed with pYB139 and 4 that were transformed withpYB149. DNAwas also isolated from an untransformed SU2embryo line to serve asa control. The DNAsamples were then digested with the restriction endonucleaseBgl II and Southern blots prepared as previously described (Dandekar et al.,1988, 89). In pYB139 Bgl II sites are located at positions 646, 1632 and 5018and in pYB149 at bp 646, 1632 and 3194. In the case of transformants containingT-DNA from pVB139, a probe containing either the APH(3')11 coding region orcryIA(c) coding region for ICP should hybridize to a band 3386 bp in length.However in the case of pVB149 trans formants APH(3')II coding sequences shouldhybridize to a 1.54 kb fragment, with no hybridization to the cryIA(c) probe.The APH(3')II probe was isolated and prepared as described earlier (Dandekar etal., 1988, 1989). The cryIA(c) coding region that was used as a probe wasobtained from pYB139. Purified pYB139 plasmid DNAwas digested with the tworestriction endonuclease BamHI and XhoI in succession and the digested productsseparated on a 0.8% agarose gel. The cryIA(c) coding region was isolated as a1.8 kb XhoI/BamHI band from the gel using a silica matrix called glassmilk(BiolOl Inc., La Jolla, CA). Radioactive labelling of the DNAfragments to beused as probes and the hybridization of the blots, washing and autoradiographicprocedures were identical to that described earlier (Dandekar et al., 1989).
andler
Bags were placed on 250 flower bearing branches of Chandler before theflowers became receptive (4/8/89-4/15/89). Seventy were pollinated with pecanpollen to stimulate embryogenesis. The others were kept isolated from anypollen.
Flowers were collected at various stages of development throughout thespring from 4/10/89 (before receptivity, unbagged) through 5/18/89 (last dateof attached flowers in the bags). Prior to receptivity, flower bearing brancheswere simply clipped. Later flowers which were bagged were collected by clippingthe branch proximal to the bag, leaving the flowers bagged during transport tothe lab in a chest cooler. Flowers were removed from the branches andimmediately rinsed under running tap water for 15-30 minutes. They were thengiven a 10-15 second dip in 95% ethanol followed by sterilization for 15 minutesin 10% Clorox with 2 drops of Tween 80.
Prior to dissection the developmental stage of flowers was noted as follows:
Flower Development Stages
StaJte12345
em diameter
18
-- - ---

__u___ ___ _._ ____________
Flowers were dissecteditndersterLle conditio~susing a microscope toremove either intact ovules or nucellus tissues. Isolated tissues were placedon the somatic embryo initiation medium that has been successfully used forinitiating somatic embryos from cotyledon pieces (Tuleke and KcGranahan, 1985).After 2-3 weeks on this medium, embryos were switched to DKVbasal medium, thestandard somatic embryo growing mediUJI, transferred every 1-2 weeks, and observedfor development of somatic embryos.
5. System improvements
We tested the ability of a variety of trans formant embryo clones expressingthe kanamycin resistance marker gene to sustain somatic embryo production onmedia containing increased concentrations of kanamycin. Nine trans formant embryoclones derived from separate insertion events using seven different bacterialvectors were tested on 0, 100, 200, 300, and 500 mg/L kanamycin along with foursets of non-transformed controls. Tests employed 5 initial embryos per cloneper medium, or a total of 25 embryos per clone and 6S embryos per medium.Embryos were transferred and evaluated weekly for their ability to produce newsomatic embryos.
We also tested the ability of several non-embryo tissues to grow on variousconcentrations of kanamycin and G4l8 ("Geneticin", Sigma Co., St. Louis, KO),a kanamycin analog. Shoot stem segments of a transformed shoot clone (TG29) anda non-transformed clone (SU2) , each with at least one auxilIary bud, werecultured on DKVshoot medium (Driver and Kuniyuki, 1984) supplemented with either0, 100, 250, 500 or 750 mg/L kanamycin. Cultures were grown in both the dark andin 24 hour light. Leaf and rachis pieces were also cultured in the light onkanamycin media supplemented with 1.0 mg/L 2,4-D. Additional stem segments werecultured in the light on DKVshoot medium supplemented with 0, 10, 25, 50 or 100mg/L G4l8. Tissues were observed for growth, leaf emergence, callus formation,and color changes.
RESULTS AND CONCWSIONS:
~ Feeding Studies with BT incoroorated into the diet
In last year's report we showed some of the preliminary dose response dataobtained with the three walnut target insect species in surface inoculation typefeeding studies. This year we have been able to complete these studies bycarrying out additional replications and the data has been statisticallyanalyzed.
The data that we have obtained on feeding studies with ICPFs from CryIA(b)and CryIA(c) isolated from RD-l and HD-73 strains of Bacillus thuringiensisclearly show that both proteins are toxic to the target insects at relativelylow concentrations (doses are expressed as nanogram of proteinpersqemof diet;Table 1). We found no significantdifferencein toxicitybetween CryIA(b) andCryIA(c) with respectto any of the three insectstested,with the exceptionofNOW where CryIA(b)was marginallymore toxic compared to CryIA(c). Comparinginsects we found that IMK was the most susceptible species and NOW the leastsusceptibleto either protein. Furthermore surface-inoculated diet thathad beenat 80° F for eight days showed no loss of activity when infested with neonatal
larvae, indicating that the protein remained active during the duration of the
19
- - --

study. Results from the diet-incorporation studies resembled those from thesurface-inoculated tests. All these insect species were susceptible to both
CryIA(b) and CryIA(c) (Tables 2a and 2b). The dose has been shown as both ng/cm3
(Table 2a) or pg/ml (Table 2b) as both are used in the literature. IMK was the
most susceptible and NOW the least susceptible of the three species tested. A
comparison of the LDSO'8 showed no significant difference between the two
insecticidal proteins with these insects.
Sublethal effects when NOW larvae are exposed to sublethal doses of the
two insecticidal crystal proteins CryIA(b) and CryIA(c) are in progress.Preliminary results on the effects of exposure to sublethal doses ondevelopmental times are shown in Table 3. The time required for neonatal larvaeto develop to adult increased with increasing dosages of the insecticidal
proteins (Table 3; time differences compared to control). The protein, CryIA(c),
caused an even greater delays in developmental time compared to the same doses
of CryIA(b) (Table 3; time differences between proteins or ICPFs). We had somemethod problems during the first series of these tests. The insect diet began
drying before the surviving insects had completed their development that was notrelated to the direct effects of ingestion of the proteins. Therefore, we are
conducting these experiments again with changes in methodology. As a result, themethods and results of these tests will be presented in next year's report.
2 Analysis of transeenic GUS embryos and Dlant tissue
From our previous results we found that the percent of GUS positive E1
embryos emerging from transformant producing Eo embryos increased with time in
the presence of kanamycin (McGranahan et al., 1990). From the GUS activity datawe were able to identify those embryos that were transformed but it was not clearwhether the GUS-positive E1 embryos derived from a single Eo embryo were the
result of multiplication at a single site of insertion and therefore products
of one transformation event or whether they resulted from several independent
transformation events on the surface of the Eo embryo. In order to distinguish
between these two possibilities DNA was isolated from 5 E1 embryo lines derived
from different Eo embryos (Fig. 2) as well as 8 E1 embryo lines derived from a
single Eo embryo (Fig. 3).
The DNA was digested with the restriction endonuclease Eco RI which cuts
near the right border within the T-DNA and into the flanking plant DNA, yieldingunique fragments corresponding to each integration event. This enzyme alsoyields internal T-DNA fragments of -4.5kb in pCGN700l and -0.9 kb in pCGN73l4
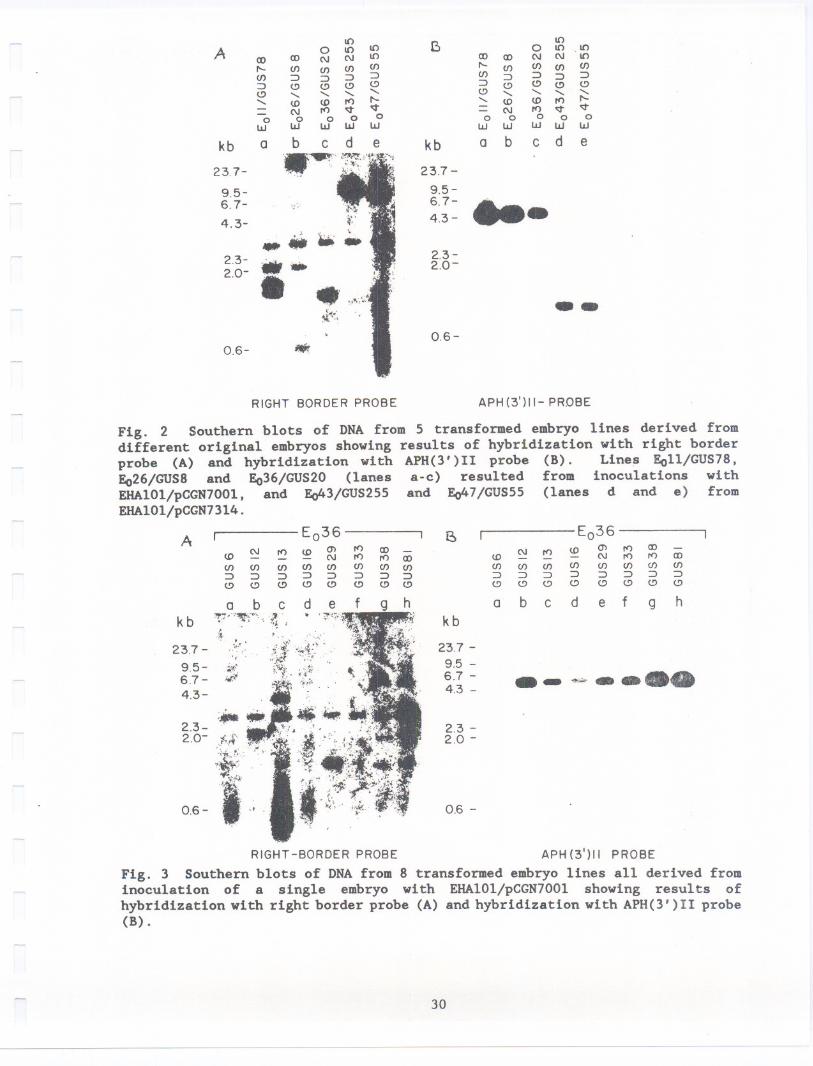
containing the coding region of the APH(3')II enzyme. Southern blots of theDNA were first hybridized with a right border (RB) probe (Fig. 2A and 3A). After
autoradiography the blots were washed free of the RB probe and rehybridized with
an APH(3')II probe (Figs. 2B and 3B). When hybridized with the RB probe eachof the lines originating from different Eo embryos displays a unique pattern of
fragments confirming separate transformation events. The patterns indicate that
from one (GUS 20, 255, and 55; Fig. 2A) to three (GUS 78; Fig. 2A) copies of
T-DNA have been incorporated in the embryos. The fragment about 3 kb long thatappears in all lanes is due to hybridization of an endogenous sequence in walnut
DNA and therefore was not counted. The differences in intensity of hybridization
observed in the RB hybridizations in Fig. 2A are due to differences in the regionof homology of the RB probe with respect to T-DNA from each of the vectors as
described in the methods section. The RB probe had a greater homology (356 bp,
20

see Methods) to T-DNA from pCGN7314 and therefore gives a more intensehybridization (Fig. 2A, lanes d and e; GUS 255 and 55) as compared to thecorresponding pCGN700l where the region of homology is only 72 bp (Fig. 2A lanesa,b and c; GUS 78,8, and 20). In contrast, the APH(3')11 probe hybridizes tothe two different T-DNA fragments in an identical manner with the result thatthe single copy insertions from pCGN7314 (Fig. 2B lanes d and e; GUS 255 and 55)are identical in intensity to the single copy insertion from pCGN7001 (Fig. 2Blane c; GUS 20). The single copy insertions can be distinguished by their lowerintensity of hybridization when compared with two copies (Fig. 2B lane b; GUS8) and 3 copies (Fig. 2B lane a; GUS 78).
The blots in Fig. 3A and B represent the DNAof E1 lines obtained from asingle EO embryo inoculated with EHA101/pCGN7001. At least 4 of the 8 linestested display unique patterns (Fig. 3A lanes b, c, e and h), 2 have similarpatterns (Fig. 3A lanes g and h) and for three there is no detectablehybridization (Fig. 3A lanes a, d and f). The unique patterns confirm independentinsertion events from which E1 embryos are derived (Fig. 3A). Variation in bandintensity (Fig. 3B) again corresponds to the copy number of T-DNA moleculesinserted per genome. The samples GUS6, GUS16 and GUS33 that do not displaydetectable hybridization with the right border probe, display GUS activi~y andhave a single T-DNA as judged by hybridization with the APH(3')11 probe (Fig.3B lanes a, d and f). A possible explanation for the lack of hybridization inthe three samples could be due to further loss of the 72 bp homology region withthe probe (see methods) caused by processing of T-DNA during its insertion andincorporation into walnut genomic DNA. The sample E036/GUS12 possibly containstwo insertions and these run very close together around 2 kb (Fig 3A lane b).This could explain the more intense hybridization observed with the APH(3')11probe (Fig 3A, lane b). The band above the major hybridization band in lane bis due to partial digestion. These results indicate that individual E1 embryosgenerated from the surface of a single inoculated EO embryo may come fromdifferent transformed cells and represent separate transformation events.
Embryos from 19 of these transformed embryo clones were germinated(McGranahan et al., 1988) and propagated In vItro as shoot cultures. Stem andleaf tissue from these shoots exhibited GUS activity in X-gluc assays. Shootsare now being grafted to seedling rootstocks in the greenhouse in preparationfor field planting next spring.
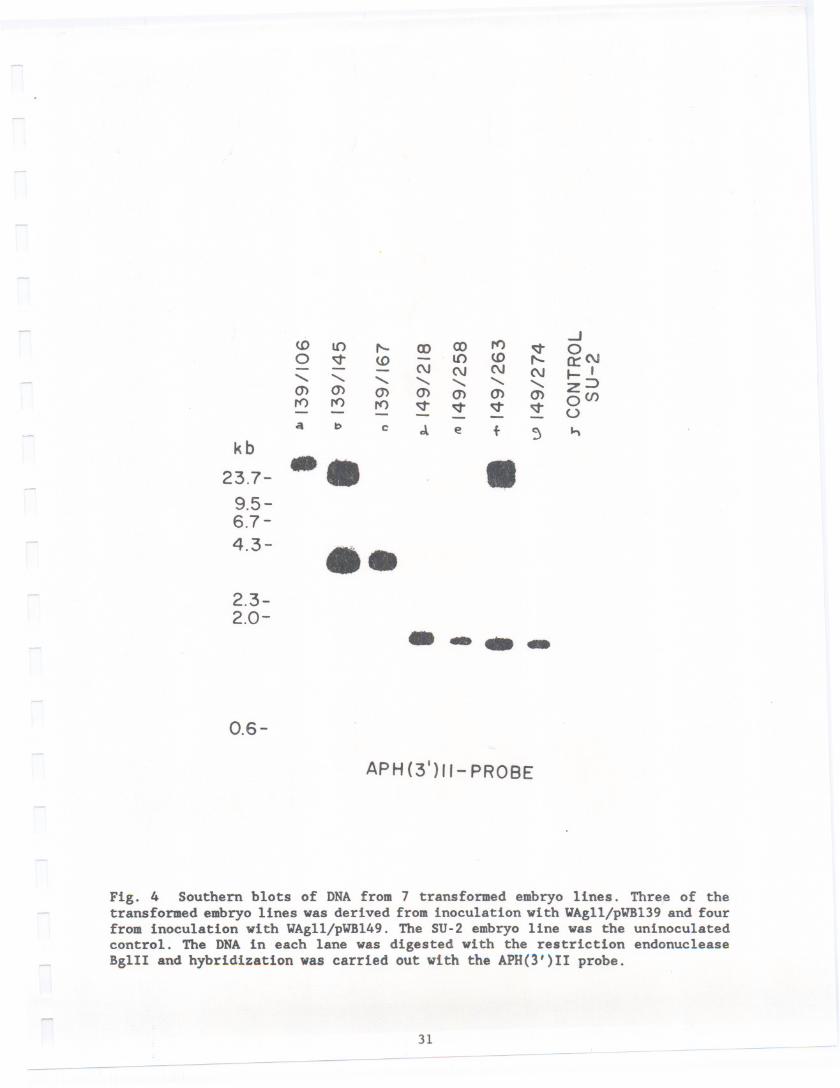
After 5 months of repetitive transfer on KAN/CEFmedium, selectionswerenarrowed to three BT/KAN clones (designated 139/106, 139/145 and 139/167) and4 KANonly clones (designated149/218, 149/258, 149/263 and 149/274) based ontheir ability to multiply on the KAN/CEF medium. Of these, one KAN clone(149/218) was clearly the best growing on this medium. Samples of each of theseembryo clones were then assayed to confirm insertion of the genes.
Three embryo colonies from pVB139 transformation and four colonies frompVB149 were shown to be positive by Southern analysis (Fig. 4). DNAextractedfrom each of the 7 embryo colonies in addition DNAwas also isolated from theSU-2 control. The DNAwas then digested with the restriction endonuclease BglII. In pVB139 Bgl II sites are locatedat positions646, 1632 and 5018 and inpVB149at bp 646,1632 and 3194. In the case of transformants containingT-DNA
21
- ---

from pWBl39, a probe containing either the APH(3')II coding region or cryIA(c)
coding region for ICP should hybridize to a band 3386 bp in length, the datacertainly shows this to be the case (Fig. 4 lanes b and c) a 3.4 kb band is
clearly visible. No hybridization is detectable in untransformed (Fig. 4 laneh). Embryos transformed with pWBl49 (Fig. 4 lane d-g) hybridize to the lean probe
encoding APH(3')II (described above) revealing a band of hybridization 1562 bpin length. No hybridization was detected in these samples when the cryIA(c)
coding sequences were used as a probe as expected (data not shown).
Embryos from each of these lines have now been successfully germinated andmicropropagated as shoot cultures. Three to five shoot cultures, each derivedfrom an individual embryo, were established for each of the selected embryoclones. These are presently being assayed for the presence of the insertedgenes.
Transformed shoots containing the BT/KAN genes and controls containingeither the KAN gene alone or no insert are currently being grafted onto seedlingrootstocks in the greenhouse. Some of these are now in a screen house for winterchilling in preparation for spring field planting pending permit approval.Plants currently grafted and acclimated are shown in Table 5
Both intact ovules and isolated nucellus tissues were placed in culture
over a six week period. A total of 269 Chandler flowers from which pollen hadbeen excluded and 72 Chandler flowers exposed to Pecan pollen were dissected and
placed in culture (Table 6).
To date no embryos have emerged from cultured ovules or nucellus. After
several months some of these cultures were transferred to DIal medium supplementedwith 1 mg/l 2,4-D and then returned to basal DIal. Although several of these
developed a small amount of callus, none have yet produced somatic embryos.
2 IlDDrovementsin the iene transfer system
Embryo production on increased kanamycin concentrations was rather variable
but evaluations showed that most transformants could sustain satisfactory embryoproduction on 200 mg/L kanamycin. Embryo production by some transformants
declined on higher concentrations of kanamycin although several clones exhibited
high production even on 500 mg/L kanamycin. The number of embryos produced bynon-transformants declined to nearly zero on 200 mg/L kanamycin and none were
observed at higher concentrations. Because 200 mg/L kanamycin allowedmultiplication of most or all transformants while more completely suppressinggrowth of non-transformants than concentrations used in the past, thisconcentration of kanamycin should prove more efficient for use in futurescreening.
Screening of transformed and non-transformed shoot tissues on kanamycinand the kanamycin analog G4l8 showed that the growth of non-transformed shoots
and leaf emergence from non-transformed buds was markedly inhibited by 100 mg/L
kanamycin or 25 mg/L G4l8 and very strongly or completely inhibited by 250 mg/Lkanamycin or 50 mg/L G4l8. Transformed material continued to grow well onkanamycin and was not noticeably inhibited by the concentrations of G4l8 tested.
Observations of swelling or callusing of rachis and leaf samples and bleaching
22

of leaves matched the results obtained for growth and leaf emergence. Results
of these tests indicate shoots or leaves expressing the kanamycin resistance
marker gene can be selected on media containing 250 mg/L kanamycin or 50 mg/LG418.
CONCLUSIONS
The susceptibility of NOW, CM and IMM to two insecticidal crystal proteins
of Bacillus thuringiensis was determined by feeding tests. The data showed that
both proteins were toxic to all the target insects tested at relatively lowconcentrations. IMM was the most susceptible and NOW the least susceptible toeither protein. Significant delays in the time required for development to adult
were observed when NOW was exposed (per os) to very low (sublethal)concentrations. Early kanamycin exposure enhanced transgenic embryo recovery andthe GUS marker served as an efficient, reliable and easily detected marker for
transformation. Screening for GUS expression in the E1 embryo generation reduced
the time and work involved in maintaining large embryo populations for screeningin later generations as was recommended earlier (McGranahan et al., 1988). Most
transgenic E1 embryos represent a unique transformation event. DNA analysis ofa larger population of E1 embryos from a single Eo is needed to confirm this.
Transformation of embryos with a vector containing cryIA(c) gene of Bacillusthuring~ensis has been confirmed by Southern analysis.
REFERENCES
Aronson, A.I., W. Beckman and P. Dunn. 1986. Bacillus thuringiensis and relatedinsect pathogens. Microbiol. Rev. 50: 1-24.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith,J.A., Strubl, K. eds. Current Protocols in Molecular Biology. Vol. 1 & 2. NewYork: Greene Publishing Associates and Wiley-Interscience; 1987.
Barker, R.F., Idler, K.B., Thompson, D.V., Kemp, J.D. 1983. Nucleotide sequenceof the T-DNA region from the Agrobacterium tumefaciens octopine Ti plasmidpTi15955. Plant Mol. BioI. 2: 335-350.
Beck, E., Ludwig, G., Auerswald, E.A., Reiss, B., Shaller, H. 1983. Nucleotide
sequence and exact localization of the neomycin phospho transferase gene fromtransposon Tn5. Gene 19: 327-336.
Dandekar, A.M., S.L. Uratsu, G.H. McGranahan, C.A. Leslie, P. Vail, S. Tebbits.
1989. Genetic Engineering of Insect Tolerance in Walnut: Susceptibility to BT.Walnut Research Reports 1988: 26-41.
Dandekar, A.M., G.H. McGranahan, C.A. Leslie and S.L. Uratsu. 1989.Agrobacterium-mediated transformation of somatic embryos as a method for theproduction of transgenic plants. J. Tissue Cult. Meth. (In Press)
Dellaporta, S.L., J. Wood, J .B. Hicks. 1983. A plant DNA minipreparation: VersionII. Plant Mol. BioI. Rep. 1(2): 19.
23

Ditta, G., G. Stanfield, D. Corbin, D.R. Helsinki. 1980. Broad host range DNAcloning system for gram negative bacteria. Construction of . gene bank ofRhizobium melilotii. Proc. Natl. Acad Sci. U.S.A. 17: 7347-7351.
Driver, J.A., and A.H. Kuniyuki. 1984. In vitro propagation of Paradox walnutrootstock. HortScience 19(4): 507-509.
Feinberg, A.P., B. Vogelstein. 1983. A technique for radiolabelUng DNArestriction endonuclease fragments to high specific activity. Anal. Biochem. 132:6-13.
Feinberg, A.P., B. Vogelstein. 1984. AddendUII to a technique for radiolabellingDNA restriction endonuclease fragments to high specific activity. Anal. Biochem.137: 266-267.
Fischhoff, D.A., K.S. Bowdish, F.J. Perlak, P.G. Marrone, S.M.
Niedermeyer, D.A. Dean, K. Kusano-Kretzmer, E.J. Mayer, D.E.
Rogers and R.T. Fraley. 1987. Insect tolerant transgenic
Bio/Technology 5: 807-813.
McCormick, J.G.
Rochester, S.G.
tomato plants.
George EF and PD Sherrington. 1984. Plant propagation by tissue culture. pub.
Eastern Press, Reading, U.K.
Hofmann, C., H. Vanderbruggen, H. H6fte, J. Van Rie, S. Jansens and H. VanMellaert. 1988. Specificity of Bacillus thuringiensis delta endotoxins iscorrelated with the presence of high affinity binding sites in brush bordermembrane of target insect midguts. Proc. Natl. Acad. Sci. USA 85: 7844-7848.
H6fte, H., and H.R. Whiteley. 1989. Insecticidal crystal proteins of Bacillusthuringiensis. Microbiol. Rev. 53: 242-255.
Holsters, M., D. De Waele, A. Depicker, E. Messens, M. Van Montagu, J. Schell.
1978. Transfection and Transformation of Agrobacterium tumefaciens. Mol. Gen.Genet. 163: 181-187; 1978.
Hood, E.H., G.L. Helmer, R.T. Fraley, M-D. Chilton. 1986. Tbe hypervirulence of
Agrobacterium tumefaciens A28l is encoded in a region of pTiBo542 outside T-DNA.J. Bacteriol. 168: 1291-1301; 1986.
Jefferson, R.A. 1987. Assaying chimeric genes in plants: Tbe GUS gene fusionsystem. Plant Mol. BioI. Rep. 5: 387-405.
Klein, T.M., E.D. Wolf, R. Wu, and J.C. Sanford. 1987. High velocitymicroprojectiles for delivering nucleic acids into living cells. Nature 327: 70-73.
Maniatis, T., E.F. Fritsch, J. Sambrook. 1982. Molecular Cloning: A laboratorymanual. New York: Cold Spring Harbor Laboratory.
McGranahan, G.H., C.A. Leslie, S.L. Uratsu, L.A. Martin, A.M. Dandekar. 1988.
Agrobacterium-mediated transformation of walnut somatic embryos and regenerationof transgenic plants. Bio/Technology 6: 800-804.
24
---

KcGranahan. G.B.. C.A. Leslie. S.L. Uratsu. and A.K. Dandekar. 1990. Improvedefficiency of the walnut somatic embryo gene transfer system. Plant Cell Rep.(In Press).
Rodriguez. R.L.. and R.C. Tait. 1983. Recombinant DNA Techniques.Benjamin/Cummings Pub. Co.. Kenlo Park. CA. 184 pp.
Southern. E.K. 1975. Detection of specific sequences among DNA fragmentsseparated by gel electrophoresis. J. Kol. BioI. 98: 503-507.
Tuleke. W.. and G.B. KcGranahan. 1985. Somatic embryogenesis and plantregeneration from cotyledon tissue of walnut. Juglans regia. Plant Sci. 40:57-63.
Uliano E.C.. R.B. Smith. J.H. Gould. and T.D. KcKnight. 1988. Transformation of
plants via the shoot apex. In Vitro Cellule Develop. BioI. 24: 951-954.
Vaeck. K.. A. Reynaerts. H. Hofte. S. Jansens, K. DeBeuckeleer. C. Dean. K.
Zabeau. K. Van Kontagu and J. Leemans. 1987. Transgenic plants protected frominsect attack. Nature 328: 33-37.
Wahl. G.K.. K. Stern. R.G. Stark. 1977. Efficient transfer of large DNA fragments
from agarose gels to diazobenzloxymethal paper and rapid hybridization usingdextran sulfate. Proc. Natl. Acad. Sci. U.S.A. 76: 3683-3687.
Whiteley. H.R.. and H.E. Schnepf. 1986. The molecular biology of parasporalcrystal body formation in BacIllus thuringiensis. Ann. Rev. Kicrobiol. 40: 549-576.
25
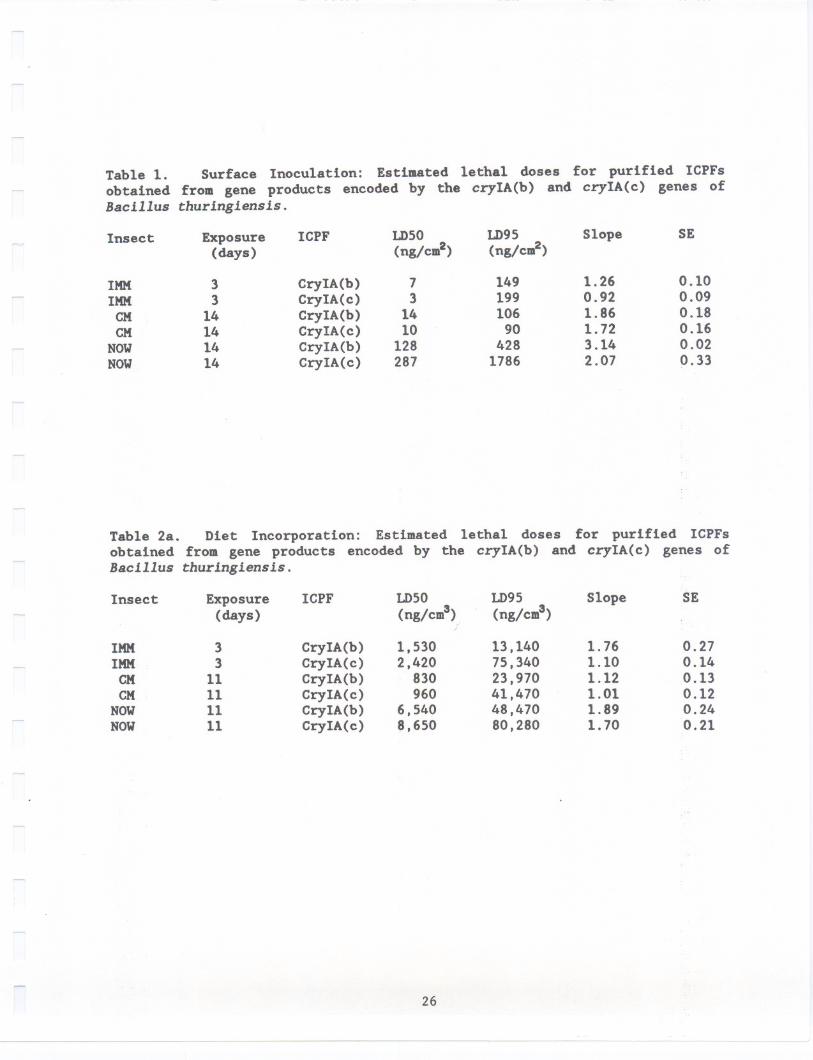
Table 2a. Diet Incorporation: Estimated lethal doses for purified ICPFsobtained from gene products encoded by the cryIA(b) and cryIA(c) genes ofBac1llus thur1ng1ens1s.
Insect Exposure(days)
33
11111111
ICPF LD50(ng/cm3)
1,5302,420830960
6,5408,650
IMMIMMCMCMNOWNOW
CryIA(b)CryIA(c)CryIA(b)CryIA(c)CryIA(b)CryIA(c)
26
-- ------
LD95(ng/cm3)
13,14075,34023,97041,47048,47080,280
Slope SE
1.761.101.121.011.891.70
0.270.140.130.120.240.21
Table 1. Surface Inoculation: Estimated lethal doses for purified ICPFsobtained from gene products encoded by the cryIA(b) and cryIA(c) genes ofBac1llus thur1ng1ens1s.
Insect Exposure ICPF LD50 LD95 Slope SE
(days) (ng/cm2) (ng/c)
IMM 3 CryIA(b) 7 149 1.26 0.10
IMM 3 CryIA(c) 3 199 0.92 0.09CM 14 CryIA(b) 14 106 1.86 0.18CM 14 CryIA(c) 10 90 1. 72 0.16
NOW 14 CryIA(b) 128 428 3.14 0.02NOW 14 CryIA(c) 287 1786 2.07 0.33
Table 2b. Diet Incorporation: Estimated lethal doses for purified ICPFsobtained from gene products encoded by the cryIA(b) and cryIA(c) genes ofBacIllus thuringiensis.
Insect
IMMIMMCMCMNOWNOW
Exposure(days)
3311111111
ICPF LD50
(pg/m1)
1.52.40.81.06.58.7
LD95
(pg/m1)
13.175.324.041.548.580.3
Slope SE
CryIA(b)CryIA(c)
CryIA(b)CryIA(c)
CryIA(b)
CryIA(c)
1.761.101.121.011.891.70
0.270.140.130.120.240.21
Table 3. Developmental times from neonate larvae to 50' adult emergence forNOW raised on media surface contaminated with sublethal concentrations of ICPFs
of CryIA(b) and CryIA(c) insecticidal crystal proteins.
ICPF
CryIA(b)
CryIA(c)
CryIA(b)
CryIA(c)
CryIA(b)CryIA(c)
27
- ------
Dose Developmental Time Diff. between:
(ng/cm2) time (days) Control ICPF
29.510 38.5 + 910 42.5 +13 +450 46.0 +16.550 51.0 +21.5 + 5100 47.0 +17.5100 52.5 +23 + 5.5
j
Tab.le 4. Embryo inoculations
Table 5. Grafted BT shoots
Table 6. Chandler ovule and nucellus cultures
28--
Vector Traits Embryo Age #Embryos EO embryoTreated ID#
oWB 139 BTIKAN Younll 97 1-97oWB 139 BTIKAN Middle Ailed 82 98-179oWB 149 KAN Younll 100 201-300
IoWB 149 KAN Middle Ailed 54 301-354
Traits Clone ID # Grafted PlantsBT 167 1BT 145A 4BT 145C 2BT AN 258A 7BT AN 258B 2BT AN 258C 1BT AN 274B 1BT AN 274C 1BT AN 218E 1BT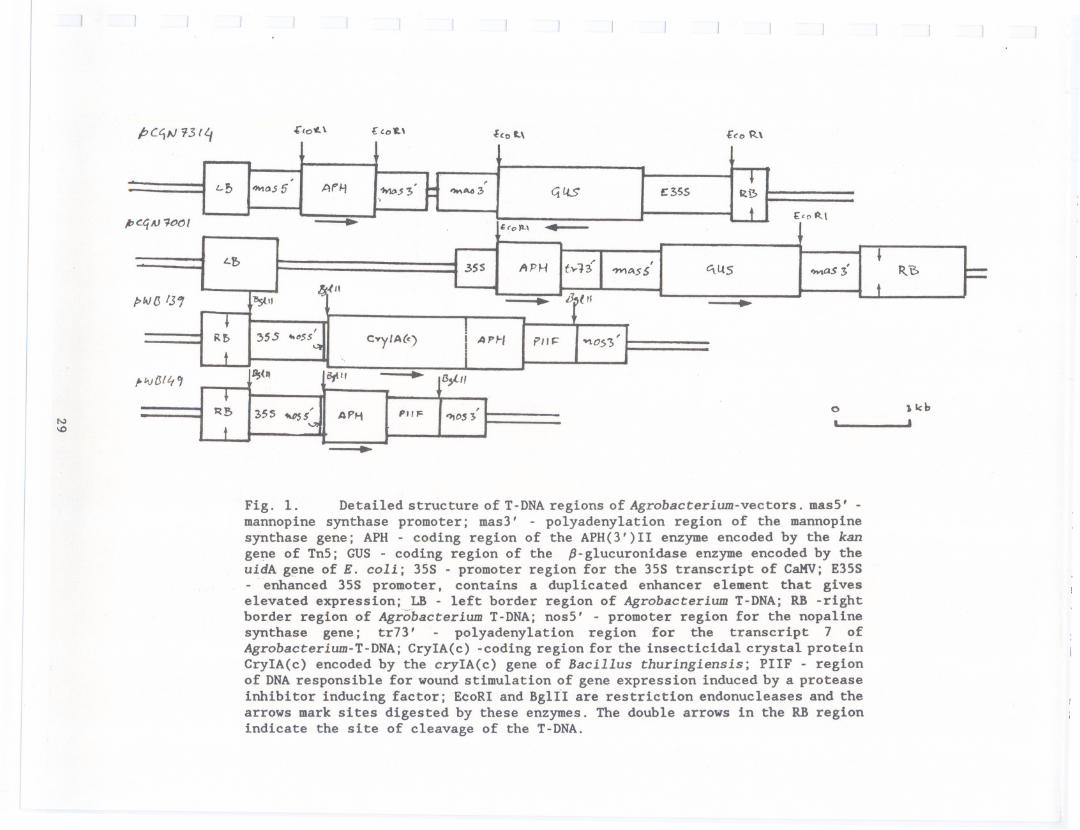
pcc,fJ 13/'1 "'tooL \ £ to t., ~to L\ £co (t\
l'
N'"
Fig. 1. Detailed structureof T-DNAregions of Agrobacterium-vectors. masS' -mannopine synthase promoter; mas3' - polyadenylation region of the mannopinesynthase gene; APH - coding region of the APH(3')11 enzyme encoded by the kangene of TnS; GUS - coding region of the p-glucuronidase enzyme encoded by theuidA gene of E. coli; 3SS - promoter region for the 3SS transcript of CaKV; E3SS- enhanced 3SS promoter, contains a duplicated enhancer element that giveselevated expression; LB - left border region of Agrobacterium T-DNA; RB -rightborder region of Agrobacterium T-DNA; nosS' - promoter region for the nopalinesynthase gene; tr73' - polyadenylation region for the transcript 7 ofAgrobacterium-T-DNA; CryIA(c) -coding region for the insecticidal crystal proteinCryIA(c) encoded by the cryIA(c) gene of Bacillus thuringlensls; PIIF - regionof DNAresponsible for wound stimulation of gene expression induced by a proteaseinhibitor inducing factor; EcoRI and BglII are restriction endonucleases and thearrows mark sites digested by these enzymes. The double arrows in the RB regionindicate the site of cleavage of the T-DNA.
, , .L-!; ""'os 5 Afli
.:t>Ila$>'IMA43 4 £355 5
t £'0 \qAJ 1COI .. 'oJt.\ .
, .f> APH t-..13,
ltS R..3SS 'W\sS 'MaS 3O(" t
f>fN(J131 l!:stI' .11tH
... I
Rr. 5.s ..O$S'C"'y'A(c)
I APf-!
PII '>\0$ '3 't
"IVtJIL, , 1st" Sflt, . B,l,'.
0 JlcbB Z>5S 'VS l ,." F ,APM D$J . .
t'-'I
RIGHT BORDER PROBE APH(3'jll- PROBE
Fig. 2 Southern blots of DNA from 5 transformed embryo lines derived fromdifferent original embryos showing results of hybridization with right borderprobe (A) and hybridization with APH(3') II probe (8). Lines Eoll/GUS78,Eo26/GUS8 and Eo36/GUS20 (lanes a-c) resulted from inoculations withEHA101/pCGN7001, and Eo43/GUS2SS and Eo47/GUSSS (lanes d and e) fromEHAI01/pCGN7314.
0.6 -
RIGHT-BORDER PROBE APH(3'jll PROBE
Fig. 3 Southern blots of DNA from 8 transformed embryo lines all derived frominoculation of a single embryo with EHA101/pCGN7001 showing results ofhybridization with right border probe (A) and hybridization with APH(3')11 probe(8).
30
A I E036 I B I E036C\J If) CD 0'1 If) CD - C\J If) CD 0'1 If) CD
CD - - - C\J If) If) CD CD - C\J If) If) CD(f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f) (f)::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::> ::><.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!) <.!)
b c d e f 9 h a b c d e f 9 hkb 1f" ':r"'-' '"l ..';.: kb,. .... .. '.4ir 'j;;"::::... . . , .,." ''". . ._-',
23.7 -9.5 -6.7 - -_........,...4.3-
2.3 -2.0 -
0.6-
APH(31)/I-PROBE
Fig. 4 Southern blots of DNA from 7 transformed embryo lines. Three of thetransformed embryo lines was derived from inoculation with VAgII/pWB139 and fourfrom inoculation with VAgll/pWB149. The SU-2 embryo line was the uninoculatedcontrol. The DNA in each lane was digested with the restriction endonucleaseBglll and hybridization was carried out with the APH(3')II probe.
31-- ------ -- ---- - - - -
-'<D 10 I"- CD CD rt) V-ON° V- <D - 10 <D I"- a:: ,- - N N N Nf-::::>"- "- "- "- "- "- "-en en en en en en en (f)rr> rr> rt) V- V- V- V- u- - -.. t> c J. e t '"'
kb -. .23.7-9.5-6.7-4.3- .-2.3-2.0- ----..