Embed Size (px)
Citation preview

1
Banque « Agro-Véto » PRO-12.09
BIOLOGIE
Epreuve B
Durée : 3 heures 30 minutes
L’usage de la calculatrice, d’abaques et de tables est interdit pour cette épreuve. Si, au cours de l’épreuve, un candidat repère ce qui lui semble être une erreur d’énoncé, il le signale sur sa copie et poursuit sa composition en expliquant les raisons des initiatives qu’il a été amené à prendre.
La β galactosidase d’Escherichia coli. A partir de l’exploitation des documents proposés et de vos connaissances, étudiez quelques-unes des propriétés de cette enzyme impliquée dans le catabolisme d’E. coli.
* L’exposé sera encadré par une introduction et une conclusion et sera structuré par un plan faisant apparaître explicitement les thèmes abordés et la progression suivie. * L’exposé doit se limiter aux trois thèmes abordés. * Le candidat ne doit pas rédiger de longs développements de ses connaissances sur le sujet indépendamment de l’exploitation des documents. * Les documents peuvent être découpés et intégrés à la copie à condition d’être exploités.
Le sujet présente 7 pages (recto uniquement).
2
Thème 1 : Relation structure – fonction de la ß-galactosidase
(D’après concours ENS Cachan 1996)
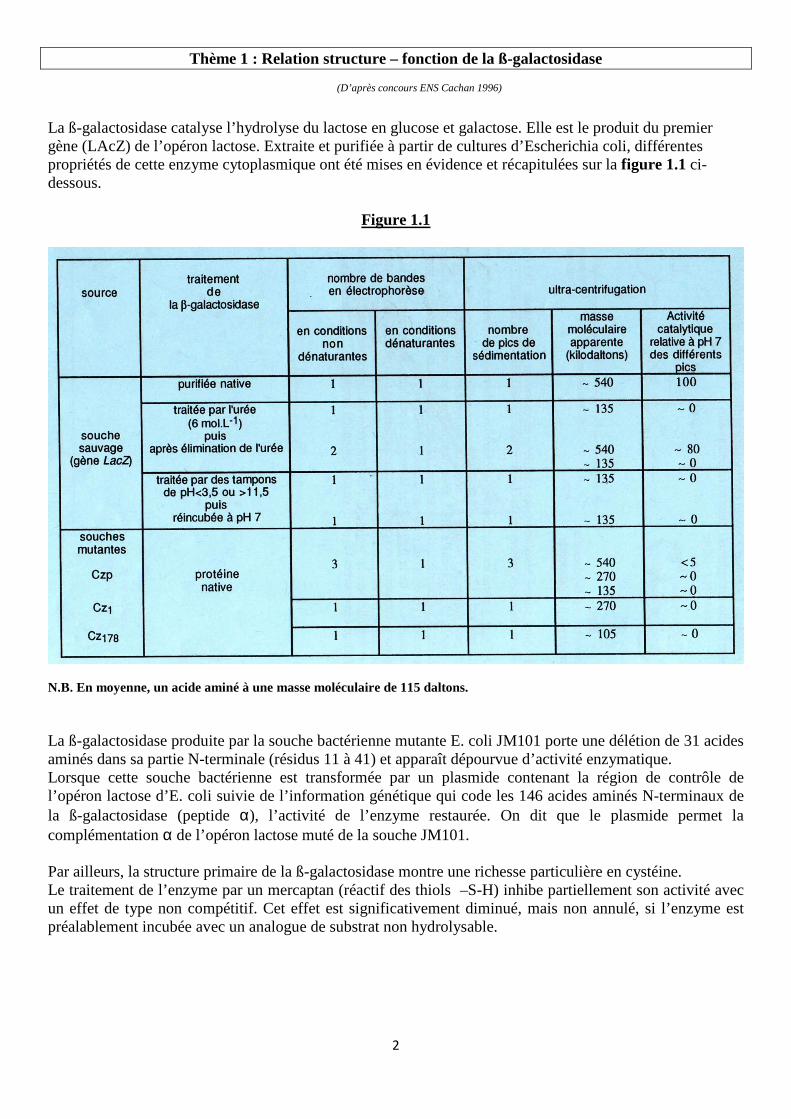
La ß-galactosidase catalyse l’hydrolyse du lactose en glucose et galactose. Elle est le produit du premier gène (LAcZ) de l’opéron lactose. Extraite et purifiée à partir de cultures d’Escherichia coli, différentes propriétés de cette enzyme cytoplasmique ont été mises en évidence et récapitulées sur la figure 1.1 ci-dessous.
Figure 1.1
N.B. En moyenne, un acide aminé à une masse moléculaire de 115 daltons. La ß-galactosidase produite par la souche bactérienne mutante E. coli JM101 porte une délétion de 31 acides aminés dans sa partie N-terminale (résidus 11 à 41) et apparaît dépourvue d’activité enzymatique. Lorsque cette souche bactérienne est transformée par un plasmide contenant la région de contrôle de l’opéron lactose d’E. coli suivie de l’information génétique qui code les 146 acides aminés N-terminaux de la ß-galactosidase (peptide α), l’activité de l’enzyme restaurée. On dit que le plasmide permet la complémentation α de l’opéron lactose muté de la souche JM101. Par ailleurs, la structure primaire de la ß-galactosidase montre une richesse particulière en cystéine. Le traitement de l’enzyme par un mercaptan (réactif des thiols –S-H) inhibe partiellement son activité avec un effet de type non compétitif. Cet effet est significativement diminué, mais non annulé, si l’enzyme est préalablement incubée avec un analogue de substrat non hydrolysable.
3
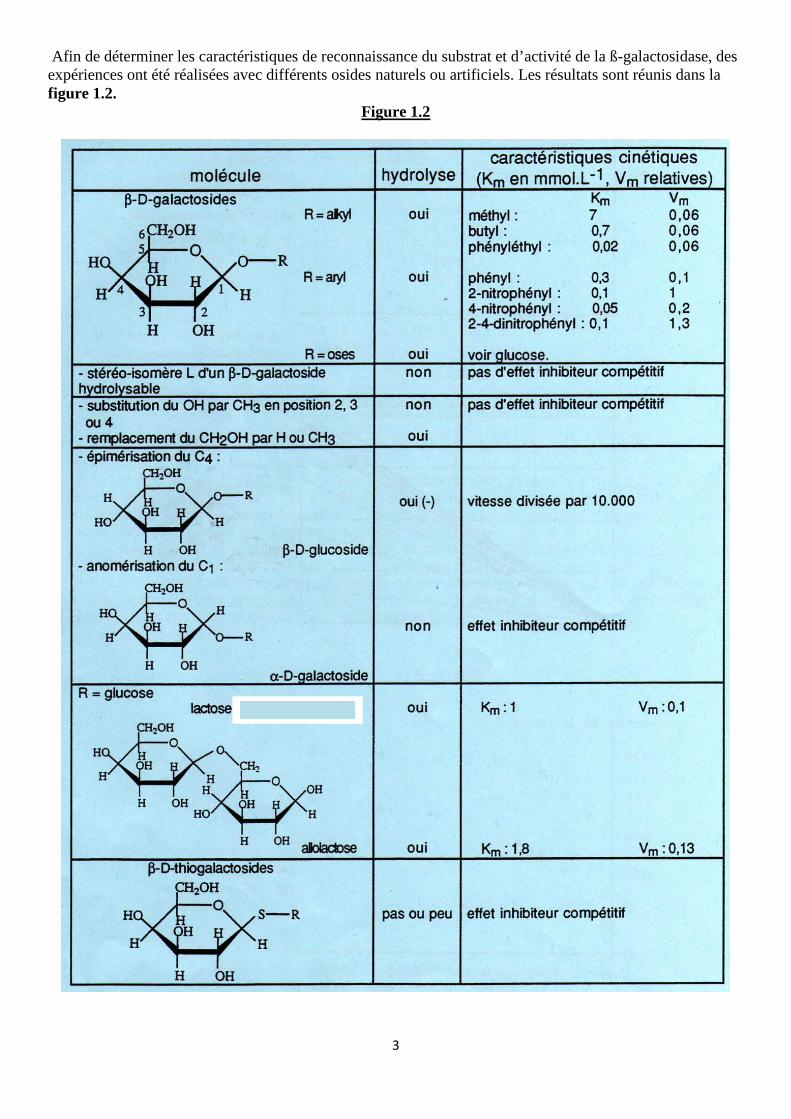
Afin de déterminer les caractéristiques de reconnaissance du substrat et d’activité de la ß-galactosidase, des expériences ont été réalisées avec différents osides naturels ou artificiels. Les résultats sont réunis dans la figure 1.2.
Figure 1.2
4
Thème 2 : Étude cinétique de la ß-galactosidase d'Escherichia coli
(D’après concours ENS Cachan 1996 & T.P. agrégation 2002(partiels) ; T.D. de Biochimie Gérard Coutouly & al. DOIN P.143)
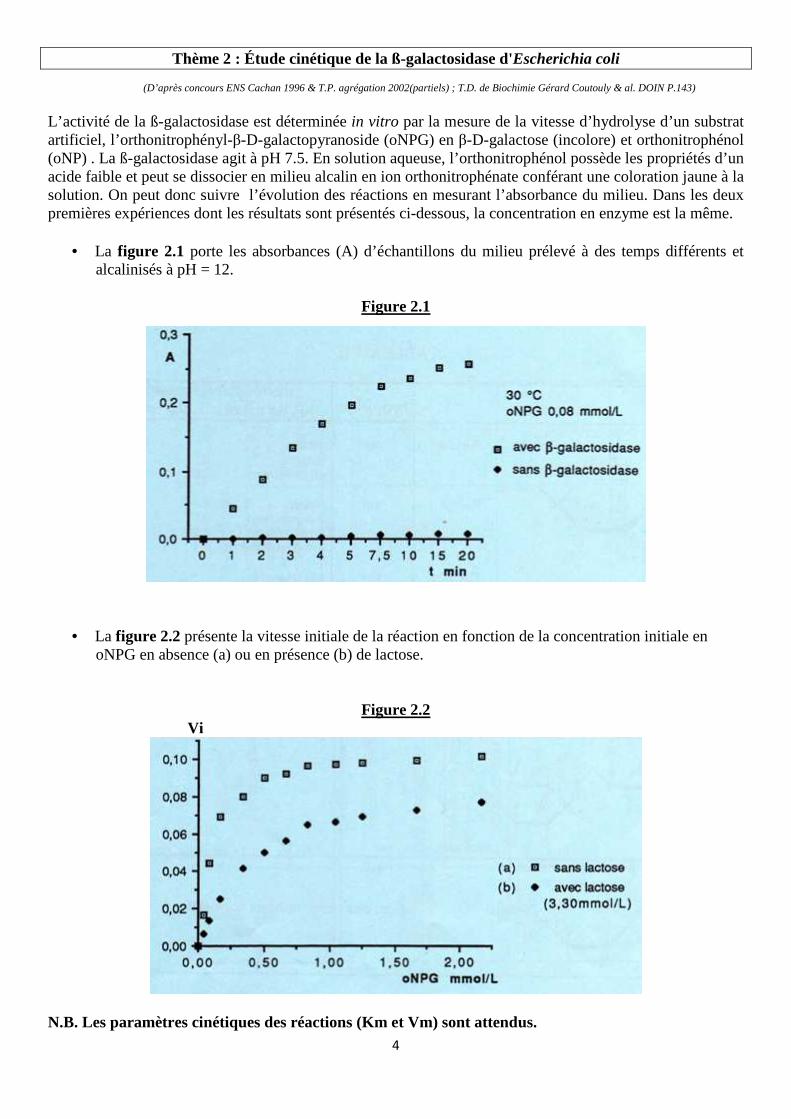
L’activité de la ß-galactosidase est déterminée in vitro par la mesure de la vitesse d’hydrolyse d’un substrat artificiel, l’orthonitrophényl-β-D-galactopyranoside (oNPG) en β-D-galactose (incolore) et orthonitrophénol (oNP) . La ß-galactosidase agit à pH 7.5. En solution aqueuse, l’orthonitrophénol possède les propriétés d’un acide faible et peut se dissocier en milieu alcalin en ion orthonitrophénate conférant une coloration jaune à la solution. On peut donc suivre l’évolution des réactions en mesurant l’absorbance du milieu. Dans les deux premières expériences dont les résultats sont présentés ci-dessous, la concentration en enzyme est la même.
• La figure 2.1 porte les absorbances (A) d’échantillons du milieu prélevé à des temps différents et alcalinisés à pH = 12.
Figure 2.1
• La figure 2.2 présente la vitesse initiale de la réaction en fonction de la concentration initiale en oNPG en absence (a) ou en présence (b) de lactose.
Figure 2.2 Vi
N.B. Les paramètres cinétiques des réactions (Km et Vm) sont attendus.
5
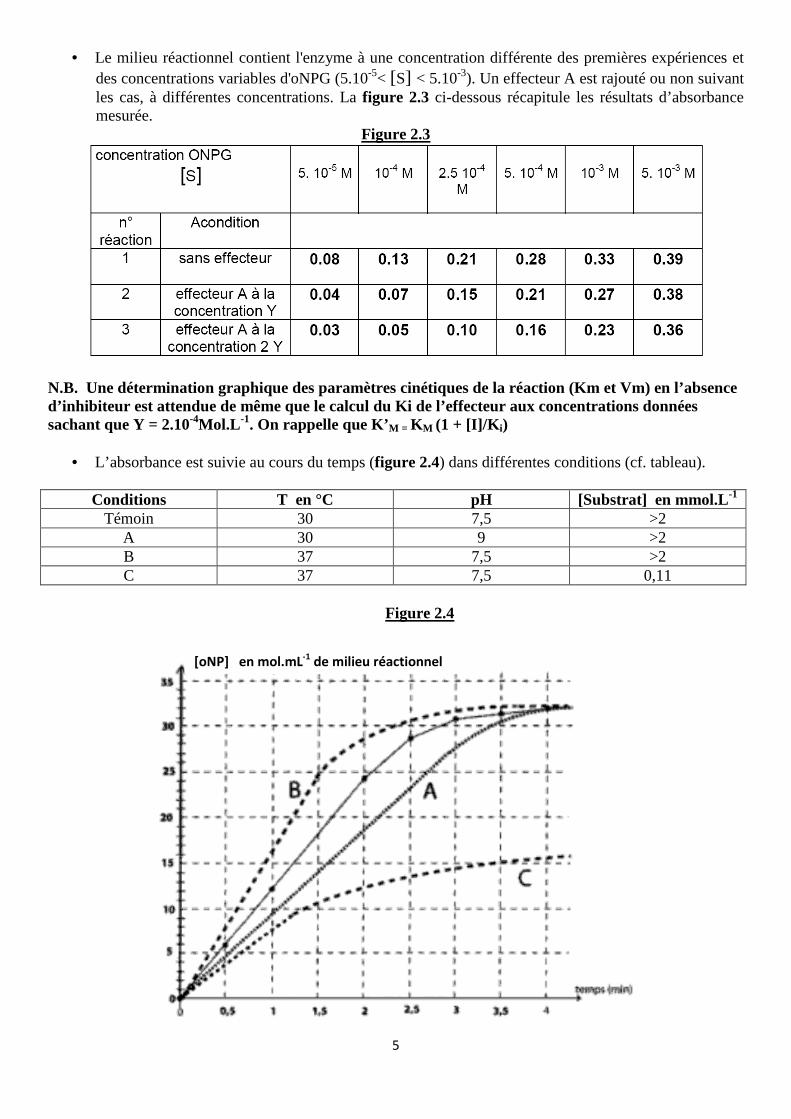
• Le milieu réactionnel contient l'enzyme à une concentration différente des premières expériences et des concentrations variables d'oNPG (5.10-5< [S] < 5.10-3). Un effecteur A est rajouté ou non suivant les cas, à différentes concentrations. La figure 2.3 ci-dessous récapitule les résultats d’absorbance mesurée.
Figure 2.3
N.B. Une détermination graphique des paramètres cinétiques de la réaction (Km et Vm) en l’absence d’inhibiteur est attendue de même que le calcul du Ki de l’effecteur aux concentrations données sachant que Y = 2.10-4Mol.L -1. On rappelle que K’M = KM (1 + [I]/K i)
• L’absorbance est suivie au cours du temps (figure 2.4) dans différentes conditions (cf. tableau).
Conditions T en °C pH [Substrat] en mmol.L-1 Témoin 30 7,5 >2
A 30 9 >2 B 37 7,5 >2 C 37 7,5 0,11
Figure 2.4
[oNP] en mol.mL-1
de milieu réactionnel
6
Thème 3 : Contrôle de l’expression du gène de la ß- galactosidase
(D’après Livre d’Exercices la Cellule (4ème édition) P.153 & T.P. agrégation 2002 partiel)
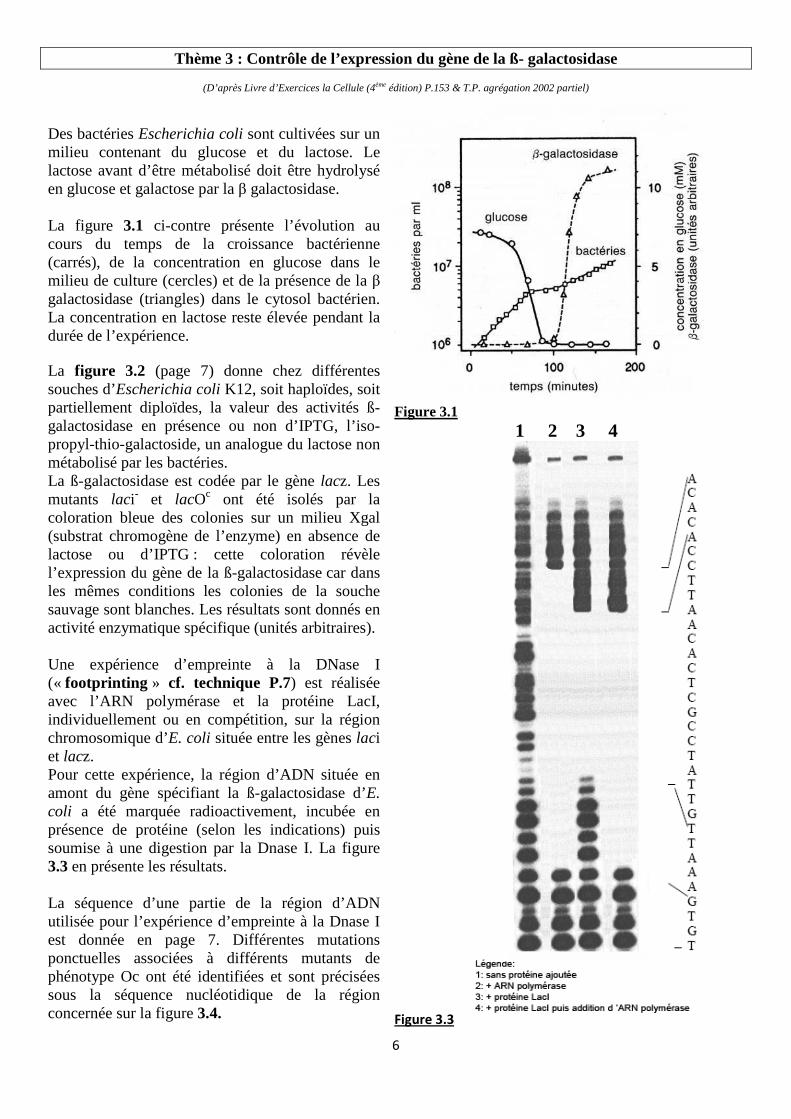
Des bactéries Escherichia coli sont cultivées sur un milieu contenant du glucose et du lactose. Le lactose avant d’être métabolisé doit être hydrolysé en glucose et galactose par la β galactosidase. La figure 3.1 ci-contre présente l’évolution au cours du temps de la croissance bactérienne (carrés), de la concentration en glucose dans le milieu de culture (cercles) et de la présence de la β galactosidase (triangles) dans le cytosol bactérien. La concentration en lactose reste élevée pendant la durée de l’expérience.
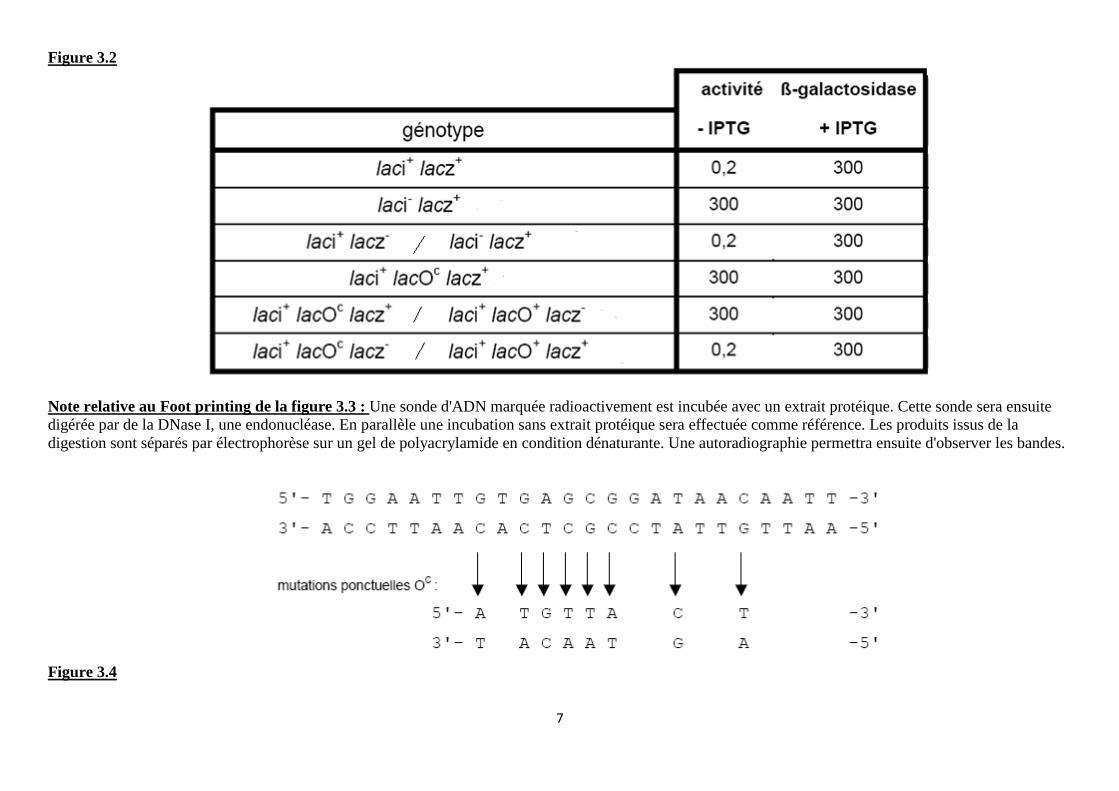
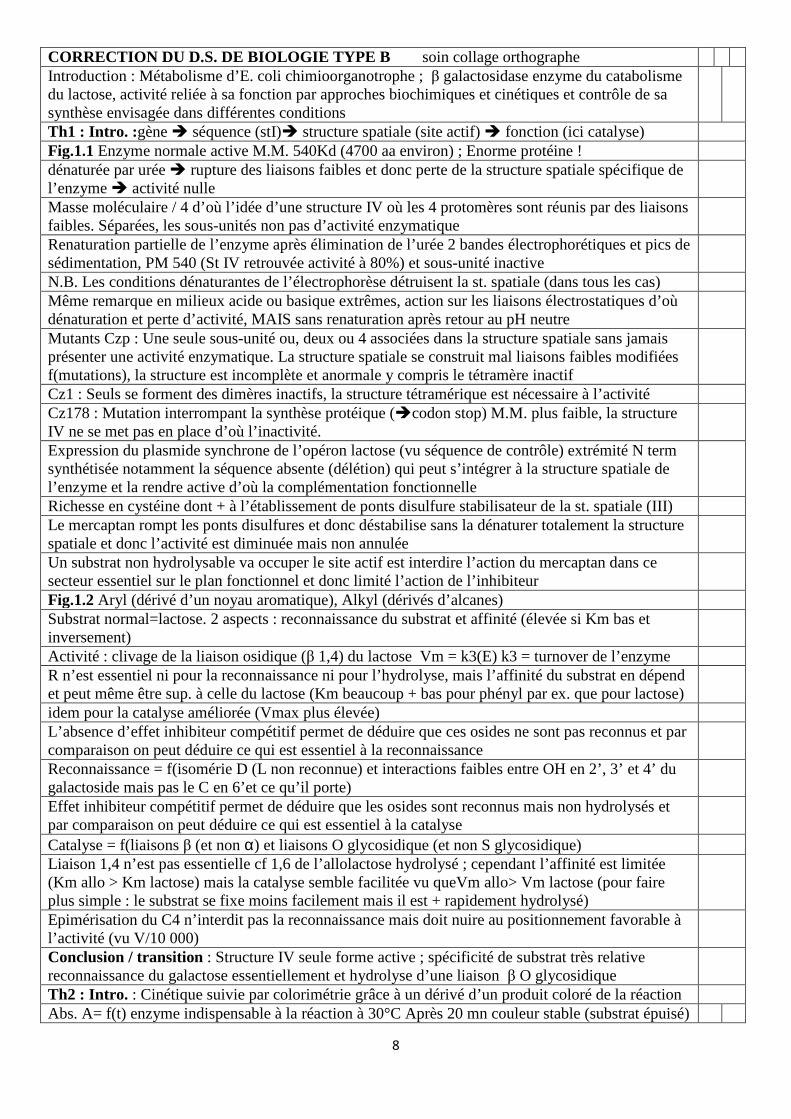
La figure 3.2 (page 7) donne chez différentes souches d’Escherichia coli K12, soit haploïdes, soit partiellement diploïdes, la valeur des activités ß-galactosidase en présence ou non d’IPTG, l’iso-propyl-thio-galactoside, un analogue du lactose non métabolisé par les bactéries. La ß-galactosidase est codée par le gène lacz. Les mutants laci- et lacOc ont été isolés par la coloration bleue des colonies sur un milieu Xgal (substrat chromogène de l’enzyme) en absence de lactose ou d’IPTG : cette coloration révèle l’expression du gène de la ß-galactosidase car dans les mêmes conditions les colonies de la souche sauvage sont blanches. Les résultats sont donnés en activité enzymatique spécifique (unités arbitraires). Une expérience d’empreinte à la DNase I (« footprinting » cf. technique P.7) est réalisée avec l’ARN polymérase et la protéine LacI, individuellement ou en compétition, sur la région chromosomique d’E. coli située entre les gènes laci et lacz. Pour cette expérience, la région d’ADN située en amont du gène spécifiant la ß-galactosidase d’E. coli a été marquée radioactivement, incubée en présence de protéine (selon les indications) puis soumise à une digestion par la Dnase I. La figure 3.3 en présente les résultats. La séquence d’une partie de la région d’ADN utilisée pour l’expérience d’empreinte à la Dnase I est donnée en page 7. Différentes mutations ponctuelles associées à différents mutants de phénotype Oc ont été identifiées et sont précisées sous la séquence nucléotidique de la région concernée sur la figure 3.4.
Figure 3.1
1 2 3 4
Figure 3.3
7
Figure 3.2
Note relative au Foot printing de la figure 3.3 : Une sonde d'ADN marquée radioactivement est incubée avec un extrait protéique. Cette sonde sera ensuite digérée par de la DNase I, une endonucléase. En parallèle une incubation sans extrait protéique sera effectuée comme référence. Les produits issus de la digestion sont séparés par électrophorèse sur un gel de polyacrylamide en condition dénaturante. Une autoradiographie permettra ensuite d'observer les bandes.
Figure 3.4
8
CORRECTION DU D.S. DE BIOLOGIE TYPE B soin collage orthographe Introduction : Métabolisme d’E. coli chimioorganotrophe ; β galactosidase enzyme du catabolisme du lactose, activité reliée à sa fonction par approches biochimiques et cinétiques et contrôle de sa synthèse envisagée dans différentes conditions
Th1 : Intro. :gène � séquence (stI)� structure spatiale (site actif) � fonction (ici catalyse) Fig.1.1 Enzyme normale active M.M. 540Kd (4700 aa environ) ; Enorme protéine ! dénaturée par urée � rupture des liaisons faibles et donc perte de la structure spatiale spécifique de l’enzyme � activité nulle
Masse moléculaire / 4 d’où l’idée d’une structure IV où les 4 protomères sont réunis par des liaisons faibles. Séparées, les sous-unités non pas d’activité enzymatique
Renaturation partielle de l’enzyme après élimination de l’urée 2 bandes électrophorétiques et pics de sédimentation, PM 540 (St IV retrouvée activité à 80%) et sous-unité inactive
N.B. Les conditions dénaturantes de l’électrophorèse détruisent la st. spatiale (dans tous les cas) Même remarque en milieux acide ou basique extrêmes, action sur les liaisons électrostatiques d’où dénaturation et perte d’activité, MAIS sans renaturation après retour au pH neutre
Mutants Czp : Une seule sous-unité ou, deux ou 4 associées dans la structure spatiale sans jamais présenter une activité enzymatique. La structure spatiale se construit mal liaisons faibles modifiées f(mutations), la structure est incomplète et anormale y compris le tétramère inactif
Cz1 : Seuls se forment des dimères inactifs, la structure tétramérique est nécessaire à l’activité Cz178 : Mutation interrompant la synthèse protéique (�codon stop) M.M. plus faible, la structure IV ne se met pas en place d’où l’inactivité.
Expression du plasmide synchrone de l’opéron lactose (vu séquence de contrôle) extrémité N term synthétisée notamment la séquence absente (délétion) qui peut s’intégrer à la structure spatiale de l’enzyme et la rendre active d’où la complémentation fonctionnelle
Richesse en cystéine dont + à l’établissement de ponts disulfure stabilisateur de la st. spatiale (III) Le mercaptan rompt les ponts disulfures et donc déstabilise sans la dénaturer totalement la structure spatiale et donc l’activité est diminuée mais non annulée
Un substrat non hydrolysable va occuper le site actif est interdire l’action du mercaptan dans ce secteur essentiel sur le plan fonctionnel et donc limité l’action de l’inhibiteur
Fig.1.2 Aryl (dérivé d’un noyau aromatique), Alkyl (dérivés d’alcanes) Substrat normal=lactose. 2 aspects : reconnaissance du substrat et affinité (élevée si Km bas et inversement)
Activité : clivage de la liaison osidique (β 1,4) du lactose Vm = k3(E) k3 = turnover de l’enzyme R n’est essentiel ni pour la reconnaissance ni pour l’hydrolyse, mais l’affinité du substrat en dépend et peut même être sup. à celle du lactose (Km beaucoup + bas pour phényl par ex. que pour lactose)
idem pour la catalyse améliorée (Vmax plus élevée) L’absence d’effet inhibiteur compétitif permet de déduire que ces osides ne sont pas reconnus et par comparaison on peut déduire ce qui est essentiel à la reconnaissance
Reconnaissance = f(isomérie D (L non reconnue) et interactions faibles entre OH en 2’, 3’ et 4’ du galactoside mais pas le C en 6’et ce qu’il porte)
Effet inhibiteur compétitif permet de déduire que les osides sont reconnus mais non hydrolysés et par comparaison on peut déduire ce qui est essentiel à la catalyse
Catalyse = f(liaisons β (et non α) et liaisons O glycosidique (et non S glycosidique) Liaison 1,4 n’est pas essentielle cf 1,6 de l’allolactose hydrolysé ; cependant l’affinité est limitée (Km allo > Km lactose) mais la catalyse semble facilitée vu queVm allo> Vm lactose (pour faire plus simple : le substrat se fixe moins facilement mais il est + rapidement hydrolysé)
Epimérisation du C4 n’interdit pas la reconnaissance mais doit nuire au positionnement favorable à l’activité (vu V/10 000)
Conclusion / transition : Structure IV seule forme active ; spécificité de substrat très relative reconnaissance du galactose essentiellement et hydrolyse d’une liaison β O glycosidique
Th2 : Intro. : Cinétique suivie par colorimétrie grâce à un dérivé d’un produit coloré de la réaction Abs. A= f(t) enzyme indispensable à la réaction à 30°C Après 20 mn couleur stable (substrat épuisé)

9
Fig.2.1 Le passage au pH 12 arrête l’hydrolyse vu l’optimum de pH de l’enzyme qui est de 7,5 Fig.2.2 Plateau de saturation de l’enzyme donne le Vmax (enzyme limitante) = 0,1 Km = 0,1mM/L Cinétique Michaelienne or la structure est quaternaire ; l’enzyme n’est donc pas allostérique Argh ! Lactose = inhibiteur compétitif de l’oNPG (vu que c’est le substrat naturel de l’enzyme) donc Vmax identique et Km plus élevée (affinité moindre…) = 0,5 mM/L
Figure 2.3 Construction graphique réalisée avec soin Vmax = 0,4 environ donc : KM témoin = 2,5 10-4M KMy = 5 10-4M K M2y = 7,5 10-4M environ Ki = (Km*Y)/(K’m-Km) = 2 *10 -4M
Fig.2.4 Pente traduit une transformation plus ou moins rapide du substrat en produit A/témoin plus lente facteur limitant le pH (optimum à 7,5) B/témoin plus rapide T optimale 37°C (T du corps qui héberge E.coli T.D. humain entre autres…) C/B qui réunit les conditions optimales T et pH pente environ 2 fois plus faible, concentration en substrat est limitante (elle correspond à peu près au K)
Conclusion/transition : Enzyme indispensable à l’hydrolyse du lactose dans les conditions biologiques T=37°C et pH=7,5 optimale pour le fonctionnement de l’enzyme ; exceptionnelle St IV mais cinétique Michaelienne avec inhibition compétitive ; pas d’allostérie! Conditions d’action ?
Th3 : Intro. Le milieu de vie est fluctuant avec des conséquences sur le catabolisme Fig.3.1 Entre 0 et 100 mn : Décroissance glucose d'où exploitation � croissance bactérienne, lactose non utilisé car pas de galactosidase
Autour de 70 à 100mn ralentissement à arrêt temporaire de croissance (manque de substrat glucose épuisé et pas encore d'enzyme pour hydrolyser le lactose)
Après 100mn reprise de la croissance, plus faible qu'avec le glucose (cf. pente), en présence de galactosidase, le lactose est utilisé par les bactéries
Démonstration d'une néosynthèse enzymatique en réponse à un facteur externe le lactose, contrôle adapté, à noter que la présence du glucose inhibe visiblement la synthèse de galactosidase.
Fig.3.2 En présence d’IPTG l’activité est toujours maximale. Ce qui n’est pas toujours le cas en son absence. L’expression dépendrait de la présence d’un substrat de l’enzyme
Laci+ /laci- montre l’action inhibitrice exercée par laci + en absence de substrat Le substrat semble par sa présence lever l’inhibition Le premier diploïde partiel montre l’action en trans de laci+ action inhibitrice sur lacz+ placé sur l’autre ADN . lacOc annule l’action inhibitrice de laci+ en absence de substrat
Le deuxième diploïde partiel montre l’action en cis de lacOc car lacO+ n’a pas d’effets sur l’activité de lacz+ situé sur l’autre ADN (expression en absence et présence de substrat)
Le troisième diploïde partiel présente l’inhibition en absence de substrat. Donc un inhibiteur protéique produit par le gène laci+ qui serait inhibé en présence de substrat mais qui en son absence se fixerait sur un segment lacO+ ce qui bloquerait la transcription de lacz
Fig.3.3 La fixation d’une protéine sur un secteur de l’ADN va le protéger et empêcher la coupure de l’ADN par les enzymes de restriction DNAse I d’où des zones non marquées dont la correspondance en séquence nucléotidique est donnée à droite de la figure 3.3.
La fixation de LacI dans une partie de la zone où se fixe l’ARNpolymérase interdit lors de la compétition une fixation correcte de l’enzyme sur la totalité de sa zone (ACAC non protégée !!!)
Mauvais positionnement qui ne permet pas la transcription Fig.3.4 Le secteur d’ADN où se fixe LacI est le siège de mutations ponctuelles diverses qui doivent empêcher une fixation correcte de l’inhibiteur sans interdire la fixation de la polymérase qui se fait sur une zone plus grande avec notamment un secteur non porteur de mutations.
Conclusion : Actions en cis et en trans sous contrôle de la présence substrat => Adaptation enzym. Conclusion générale : Enzyme originale à différentes points de vue structural, cinétique et synthèse, ouvrir sur les opérons contrôlés de diverses façons : inductibles ou répressibles

10





























