Embed Size (px)
Citation preview

VESCICOLACOMPLETAMENTE
FORMATA
VESCICOLA DIIMMAGAZZINAMENTODEL SECRETO
LE SCIENZE n. 333, maggio 1996
39
NUCLEOCELLULARE
RETICOLOENDOPLASMATICO
CITOSOL
VESCICOLA
MEMBRANACELLULAREESTERNA
LISOSOMA
T
utte le cellule nucleate - che ap-partengano a colonie di lieviti, oa piante, o all'organismo umano
- hanno un'organizzazione interna com-plessa, che ricorda quella di una me-tropoli ben amministrata. La cosa forsepiù degna di rilievo è che sia la cellulasia la metropoli dipendono dall'attivitàcoordinata di distretti specializzati: nel-le cellule questi ultimi sono delimitatida membrane e si chiamano organelli.
Una breve escursione in alcuni deidistretti più importanti della «metropo-li» cellulare potrebbe avere inizio a li-vello della membrana esterna, che è es-sa stessa un organello. Questa strutturaricorda le mura di cinta, provviste diporte, delle città antiche, nel senso checontrolla l'ingresso delle sostanze nu-tritive e di altri materiali e l'esportazio-ne di prodotti sintetizzati all'interno.Un altro compartimento d'importanzafondamentale si trova nell'interno del-la cellula e funge da centro manifattu-riero. È qui, nel reticolo endoplasmati-co, che vengono prodotte molte protei-ne, le principali componenti operativedella cellula. Le proteine appena sinte-tizzate sono trasportate in un ulteriorecompartimento, l'apparato di Golgi, do-ve vengono modificate (spesso per ag-giunta di glucidi) per poi essere avviatead altre destinazioni, sia all'interno siaall'esterno della cellula. L'apparato diGolgi, quindi, è un importante centro didistribuzione per le nostre microscopi-che metropoli.
Le cellule includono perfino i propricentri di riciclaggio, i lisosomi. Questidemoliscono le vecchie proteine e certealtre molecole, in modo che i loro ele-menti costitutivi possano essere ristrut-turati e riutilizzati; inoltre i lisosomi in-corporano materiali portati nella celluladall'esterno.
Non sorprende, dunque, che le cellu-le abbiano sviluppato un sistema di tra-
38 LE SCIENZE n. 333, maggio 1996
sporto complesso per spostare le protei-ne da un organello all'altro. Esso è or-ganizzato come i sistemi di spedizionein container adottati negli aeroporti.Tutte le merci che hanno una destina-zione comune vengono caricate in unostesso contenitore - una vescicola di tra-sporto nel caso della cellula - il qualeviene aperto solo quando giunge a de-stinazione. Ogni cellula produce moltitipi di vescicole di trasporto e ciascuntipo è specializzato per un dato percor-so intracellulare o per un dato carico.La cellula produce anche vescicole ditrasporto che immagazzinano sostanzeimportanti per la comunicazione cellu-lare (come i neurotrasmettitori e l'insu-lina); questi vettori liberano il loro cari-co in risposta a segnali ben precisi.
E chiaro che le vescicole di trasportohanno un'importanza fondamentale permolti aspetti del funzionamento dellacellula e dell'organismo intero; eppureper parecchi anni nessuno è stato in gra-do di stabilire come si formino. Di re-cente, però, noi due e i nostri collabora-tori siamo riusciti a delineare molti par-ticolari di questo processo a livello mo-lecolare. Oltre ad avere un interesse in-
trinseco, i nostri risultati potrebbero ungiorno rivelarsi utili in campo medico.Per esempio, le cellule tumorali posso-no moltiplicarsi solo se le loro vescico-le di trasporto funzionano adeguata-mente; pertanto si può pensare che so-stanze che impediscono la formazio-ne delle vescicole possano prima o poiaggiungersi ai farmaci antitumorali og-gi in commercio.
La nostra storia è per molti aspettiesemplare nel rappresentare la biologiacellulare contemporanea, nel senso cheaffonda le proprie radici nella micro-scopia (la quale ha rivelato per primal'esistenza delle vescicole di trasporto)e si risolve nella biochimica (dato chenella formazione delle vescicole sonointeressate molte reazioni molecolari).Essa illustra anche la vera natura dellaricerca scientifica. Chi non è scienziatopensa spesso che una scoperta scientifi-ca sia un processo impersonale in cuil'intelletto puro conduce, per mezzo diuna logica inesorabile, alla soluzionechiara di un problema. Quest'opinionesottovaluta il ruolo che hanno i passifalsi, gli occasionali colpi di fortuna eun'ostinazione caparbia. E le descrizio-
ni riportate sui libri di testo non tra-smettono mai quel senso di ebbrezzache viene dall'inseguire un dato! Pernoi gran parte del divertimento è con-sistito nel lavorare in stretta collabora-zione nonostante fossimo separati dal-l'Atlantico, il che ci consentiva soltantoincontri annuali.
«Ma chi è lei?»
A metà degli anni ottanta, prima chenoi due cominciassimo a collaborare, imicroscopisti che esaminavano sezionidi cellule intere avevano già delinea-to certe fasi fondamentali del trasportodi proteine da parte delle vescicole. E-ra noto, soprattutto grazie alle ricerchecondotte negli anni sessanta dal NobelGeorge E. Palade, allora alla Rockefel-ler University, che le vescicole si for-mano, probabilmente per gemmazione,dalla membrana di un organello, diven-tando così minuscoli sacchetti delimita-ti da una membrana. Successivamenteciascuno di questi sacchetti migra attra-verso i citosol (il «succo» cellulare, adalto contenuto di proteine, in cui galleg-giano gli organelli) e si fissa sulla mem-brana esterna di un organello bersagliospecifico, fondendo la propria membra-na con essa e riversando nell'organel-
lo le proteine contenute nel proprio in-terno. In altre parole, il container vieneaperto a destinazione e il suo contenutoscaricato.
Per chiarire come avviene la gemma-zione, sarebbe necessario studiare ilprocesso in provetta, cioè ricreare laformazione delle vescicole all'esternodella cellula eliminando elementi estra-nei. In questo modo si potrebbero mani-polare o comunque analizzare i compo-nenti del macchinario che produce levescicole, per determinare quali mo-lecole agiscano per prime, quali inter-vengano successivamente e così via.Ma nessuno aveva cercato di progetta-re un simile sistema privo di cellule fi-no al 1980, quando uno di noi (Roth-
man) riuscì nell'impresa alla StanfordUniversity.
Nel 1984, Rothman e collaboratoripubblicarono un lavoro in cui annuncia-vano che il loro sistema acellulare erain grado di ricreare il processo di for-mazione di vescicole che avviene nellecellule intere. Esso aveva consentito avescicole di trasporto di formarsi pergemmazione da membrane dell'appa-rato di Golgi, che possiede parecchicompartimenti annessi. La maggior par-te delle vescicole derivate da questemembrane trasporta il proprio carico daun compartimento all'altro dell'appara-to di Golgi ed è proprio questo il tipo divescicole che è stato osservato. Alcu-ne vescicole di Golgi, però, trasportano
La gemmazione di vescicolenelle cellule viventi
Una collaborazione tra scienziati sulle due sponde dell'Atlantico hachiarito il meccanismo responsabile della formazione dei minuscoli, ma
essenziali, contenitori che immagazzinano e trasportano le proteine
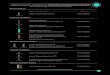
di James E. Rothman e Lelio OrciLe vescicole di trasporto (sfere in colore) abbondano nelle cellule. Alcune (in giallo)trasportano proteine prodotte nel reticolo endoplasmatico verso l'apparato di Gol-gi, dove vengono modificate. Altre (in blu) veicolano le proteine da un comparti-mento all'altro dell'apparato di Golgi. Da questo emergono tre tipi di vescicole: uno(in arancione) esporta immediatamente le proteine dalla cellula; un altro - la vesci-cola di immagazzinamento del secreto (in rosa scuro) - libera il proprio contenutoquando riceve un appropriato segnale. Un terzo tipo (in rosa chiaro) trasporta enzi-mi digestivi in apposite celle, i lisosomi, che demoliscono varie molecole, compresequelle portate nella cellula da ulteriori vescicole (in verde). Una vescicola si forma(nella pagina a fronte) per gemmazione da un organello; essa cede il carico traspor-tato (in rosso) fondendo la propria membrana con quella dell'organello bersaglio.

Il rivestimento di clatrina è stato il primo tipo di involucro a essere identificato nel-le vescicole di trasporto; si trova prevalentemente sulle vescicole che trasportanoproteine dall'esterno della cellula ai lisosomi. Il microscopio elettronico, che for-nisce un'immagine di queste vescicole sia in sezione (in alto) sia alla superficie (alcentro), rivela che il rivestimento possiede una struttura a gabbia. La sua organiz-zazione a esagoni e pentagoni ricorda quella di una cupola geodetica (in basso).
Il rivestimento COP, identificato dagli autori e dai loro collaboratori, avviluppa levescicole di trasporto che hanno origine nell'apparato di Golgi. La differenza ri-spetto al rivestimento di clatrina è ben evidente nelle microfotografie elettronichea sinistra. La microfotografia qui sopra mostra due vescicole con rivestimentoCOP mentre stanno formandosi per gemmazione da una membrana di Golgi.
proteine verso destinazioni più lontane(si veda l'articolo La compartimenta-zione dell'apparato di Golgi di JamesE. Rothman in «Le Scienze» n. 207, no-vembre 1985).
La nostra collaborazione cominciò aquesto punto, in modo memorabile. Untardo pomeriggio, meno di una settima-na dopo la pubblicazione dell'articolo,avvenuta nel dicembre 1984, il telefonosquillò nell'ufficio di Rothman. Inter-rompendo il proprio lavoro, Rothmansollevò il ricevitore, ma, prima che po-tesse pronunciare un semplice «Pron-to!», una voce profonda avviò un mo-nologo fitto fitto, rapido e totalmenteincomprensibile, in inglese con accentoitaliano. Rothman, benché a sua voltaben noto per la sua verbosità, rimasetramortito da un simile eloquio. Tra-scorsi parecchi minuti, completamentesconcertato, tornò alle sue carte, la-sciando che il monologo andasse avan-ti; solo dopo vari altri minuti, durante iquali le uniche parole comprensibili deldiscorso furono «collaborare», «com-prendere meglio il modello» e «saremofratelli», urlò finalmente, in totale fru-strazione: «Ma chi è lei?».
La voce tacque e quindi, molto paca-tamente e distintamente, addirittura conmodestia, riprese dicendo: «Sono LelioOrci. Insegno all'Università di Ginevrae lavoro su...».
Rothman riconobbe subito il nome(Orci era un notissimo esperto di micro-scopia elettronica) e si scusò con lui.Orci cominciò a parlare più lentamente,ed ebbe così luogo un lungo e proficuocolloquio, anche se a Ginevra era già damolto tempo passata la mezzanotte.
Un'idea sbagliata, ma un risultato utile
Nel giro di poche settimane, Roth-man giunse a Ginevra. Con molto en-tusiasmo, entrambi elaborammo un pia-no per sfruttare le tecniche di Orci al fi-ne di determinare se le vescicole for-matesi in provetta da membrane di Gol-gi fossero dotate di una particolareproprietà. Specificamente, desiderava-mo sapere se erano racchiuse in un rive-stimento di clatrina: uno strato spessoche formava una specie di gabbia ed eracomposto in primo luogo da clatrina,una proteina filamentosa. Per la suastruttura, questo rivestimento ricordavauna sorta di cupola geodetica.
Ci eravamo posti questa domandaperché sapevamo che certe vescicole ditrasporto possiedono rivestimenti diquesto tipo, l'unico noto a quell'epoca.(Vescicole rivestite di clatrina traspor-
tano proteine dall'esterno della cellulaal sistema lisosomale perché siano de-molite; esse si formano per gemmazio-ne dalla membrana cellulare, per essereliberate nel citosol.) Non era stato peròpossibile decifrare il meccanismo dellagemmazione per mancanza di un siste-ma sperimentale appropriato. Se Orciavesse confermato che le vescicole de-rivate dalle membrane di Golgi eranorivestite da clatrina, Rothman avreb-be potuto avviare immediatamente stu-di biochimici per validare un modelloaffascinante della formazione di vesci-cole rivestite da clatrina.
Questo modello era stato sviluppatonel corso degli anni sessanta in ricercheavviate da Toku Kanaseki e Ken Ka-dota dell'Università di Osaka. Nel 1969Kanaseki e Kadota purificarono vesci-cole dotate di rivestimento e scoprironoche quest'ultimo possedeva una struttu-ra sorprendentemente regolare. Il moti-vo di questa regolarità divenne chiarosei anni dopo, quando Barbara M. F.Pearse del Medical Research CouncilLaboratory di Cambridge, in Inghilter-ra, scoprì che il rivestimento consiste-va in primo luogo di molte copie diun'unica proteina, che chiamò clatrina.Secondo Kanaseki e Kadota, il rivesti-mento stesso era responsabile dellagemmazione. Via via che si sviluppavala gabbia di clatrina, con tutta probabi-lità a partire da componenti della mem-brana, la sottostante membrana (che èuna struttura molto plastica) si defor-mava in una specie di cupola, risuc-chiando via letteralmente dalla mem-brana una vescicola sferica con tutte leproteine a essa fissate.
Nel 1984 era ormai chiaro che questaipotesi era un po' troppo semplicistica.Quando Rothman diede una dimostra-zione della gemmazione con il suo si-stema in provetta, trovò che il processonon poteva aver luogo a partire soltantoda membrane isolate. Per formarsi levescicole richiedono la presenza di ci-tosol e di una fonte di energia; pertantoil rivestimento deve in qualche modoprovenire da sostanze presenti nel cito-sol. Il modello di base di Kanaseki eKadota era comunque attraente e meri-tava di essere verificato.
Subito dopo il ritorno di Rothman aStanford, uno dei suoi allievi, BenjaminS. Glick, cominciò a mettere in atto ilnostro piano producendo vescicole ditrasporto in provetta mediante incuba-zione dell'apparato di Golgi con il cito-sol e con una fonte di energia. Inviò poii campioni a Ginevra per farli analiz-zare al microscopio. Entro pochi giorni
Orci stabilì che le vescicole erano in ef-fetti dotate di rivestimento, ma non deltipo previsto. La sua struttura fine dif-feriva da quella dei rivestimenti di cla-trina, e il materiale di cui esso era co-stituito non veniva riconosciuto daglianticorpi che si legano in modo speci-fico alla clatrina. Questo risultato sor-prendente indicava che le cellule pro-ducono almeno due tipi di vescicole ditrasporto, le quali presentano rivesti-menti diversi. Oggi è evidente che e-sistono molti tipi di vescicole di tra-sporto, ciascuno dei quali ha un diver-so rivestimento.
Il risultato di questo studio è un ma-gnifico esempio di quanto possa essereprovvidenziale un passo falso. Un'ipo-tesi sbagliata (ossia che i rivestimentidi clatrina inducano la formazione divescicole di trasporto nell'apparato diGolgi) ci ha spinto a eseguire un espe-
rimento che ha spostato la nostra atten-zione dalle vescicole rivestite di clatri-na a quelle che avevamo appena iden-tificato. Il nostro errore ci ha così porta-to verso un nuovo e fruttuoso indiriz-zo di ricerca, che ci ha rivelato moltidettagli della formazione delle vesci-cole di trasporto. (Viceversa, le cono-scenze su come si sviluppino le vesci-cole rivestite da clatrina sono progredi-te molto più lentamente. Fino a nonmolto tempo fa nessuno era riuscito astudiare queste strutture al di fuori del-le cellule.)
Per scoprire in che modo si forminole vescicole derivate dall'apparato diGolgi, dovevamo in primo luogo deter-minare la composizione del loro rive-stimento. Questa sfida ci avrebbe tenu-ti occupati per parecchi anni. Per co-minciare, dovevamo procurarci un cer-to quantitativo delle minuscole sfere informa pura, compito davvero formida-bile a quell'epoca. Dopo sforzi accaniti,nel 1989 Vivek Malhotra e Tito A. Se-rafini, giovani ricercatori del laborato-rio di Rothman, ne isolarono una quan-tità minuscola, ma sufficiente per l'ana-lisi. Essi poterono così dimostrare che ilrivestimento conteneva otto proteine,da noi denominate COP (dall'inglesecoat protein); ben presto le vescicoleformatesi dall'apparato di Golgi prese-ro il nome di vescicole COP.
Allo scopo di chiarire il meccanismo
della gemmazione, dovevamo scoprirealtri aspetti delle proteine COP e delleloro interazioni reciproche. La nostraprima importante intuizione riguardo aqueste interazioni si ebbe nel 1990,quando scoprimmo che sette delle pro-teine del rivestimento si assemblavanoin un complesso di grandi dimensioniprima di fissarsi come unità singola al-la membrana di Golgi; questa unità èstata battezzata «coatomero» (dall'in-glese coat, rivestimento). Una sola pro-teina si legava per proprio conto. Quin-di il rivestimento consisteva essenzial-mente di due elementi principali: il coa-tomero e l'ottava proteina. Questa sco-perta semplificava profondamente lecose, dato che potevamo indagare le fa-si principali della gemmazione senzadover studiare separatamente ogni pro-teina del coatomero.
Determinanti in questa fase sono sta-te fortuna e perspicacia, dato che la sco-perta è stata fatta in maniera del tuttoaccidentale. Una sera del 1990 M. Ge-rard Waters, un giovane ricercatore delgruppo di Rothman (allora alla Prince-ton University), stava esaminando i ri-sultati dei suoi tentativi di purificareuna sostanza necessaria alle vescicoledi trasporto per fondersi con le mem-brane degli organelli bersaglio. Per pu-ra coincidenza, Serafini capitò nel la-boratorio e, guardando da sopra le spal-le di Waters, rimase meravigliato da ciòche vedeva.
Per purificare una proteina si suddi-vidono i componenti di un estratto cel-lulare in gruppi differenti in base alleloro proprietà fisiche e chimiche. Quin-di questi gruppi, o frazioni, sono sotto-posti a prove per vedere se presentanol'attività che interessa (per esempio seinducono la fusione tra una vescicola ela membrana). I campioni positivi sonoulteriormente suddivisi in un nuovo in-sieme di frazioni purificate e risottopo-sti ad analisi, mentre le frazioni che nonmostrano attività vengono in generescartate. Alla fine si ottiene un campio-ne che contiene esclusivamente la pro-teina responsabile dell'attività biologi-ca che interessa: una proteina pura.
Per essere certo di non trascurarenulla, Waters aveva analizzato il conte-
nuto proteico di una frazione che nonaveva dato segno di indurre la fusione.Per mesi egli aveva, in maniera del tut-to ovvia, scartato come inutile questafrazione. Ma Serafini rimase colpitodalla somiglianza tra le proteine dellafrazione eliminata e quelle che lui eMalhotra avevano in precedenza identi-ficato come COP.
Una rapida analisi dimostrò che ilmateriale scartato consisteva quasi e-sclusivamente di sette COP, legatestrettamente l'una all'altra. Sapevamoche le proteine dovevano essere asso-ciate perché la frazione opponeva unastrenua resistenza a ulteriori suddivisio-ni. Per ironia della sorte, dunque, men-tre una parte del gruppo di Rothman eraimpegnata in sforzi estenuanti per cer-care di isolare vescicole con rivesti-mento (per poter ottenere minuscolequantità delle proteine che lo costitui-scono), un'altra parte non aveva fattoaltro che gettare via in enormi quanti-tà le stesse proteine. Grazie alla pron-tezza di Serafini e Waters, si poté bloc-care lo spreco, riconoscere l'esistenzadel coatomero e risparmiare anni di fa-tiche. Ormai avevamo a nostra dispo-sizione una fonte illimitata di protei-ne del coatomero necessarie per i nostriesperimenti.
Ben presto ci dedicammo a stabilirel'identità dell'ottava proteina. Per molteragioni, sospettavamo che si trattasse diuna molecola chiamata fattore di ribosi-lazione dell'ADP (ARF) e descritta perla prima volta da Richard A. Kahn e Al-fred G. Gilman della Southwestern Me-dical School dell'Università del Texas.Si sapeva che questa molecola consen-te alla tossina del colera di esercitare ipropri effetti patogeni, ma il suo ruolonormale nell'organismo era misterioso.Fortunatamente è stato facile confer-mare il nostro sospetto. Da Kahn abbia-mo ottenuto anticorpi che riconosce-vano specificamente l'ARF: quando liabbiamo mescolati con la nostra pro-teina, essi hanno stabilito rapidamenteun legame, indicando che la molecolaera realmente il fattore di ribosilazionedell'ADP.
Dopo sei anni di arduo lavoro, aveva-mo quindi ottenuto tutte le proteine del
P
oo
LE SCIENZE n. 333, maggio 1996 4140 LE SCIENZE n. 333, maggio 1996

2 3 4
GTP
1!!!N
COATOM ERO
CO
CO
oo
VESCICOLAPRONTA
PER FONDERSICON UN ORGANELLO
BERSAGLIO
65 VESCICOLACON RIVESTIMENTO
FOSFATO
7
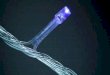
La microscopia elettronica rivela che il coatomero controlla lagemmazione delle vescicole con rivestimento COP. Quando ilsolo ARF si lega alle membrane di Golgi, non si formano gem-
me e le membrane restano piatte (a sinistra). Il fissaggio delcoatomero a membrane a cui è legato l'ARF porta invece a unamassiccia produzione di vescicole con rivestimento (a destra).
La formazione di una vescicola con rivestimento COP da una membrana di Golgi èseguita dopo breve tempo dalla rimozione del rivestimento. La vescicola inizia a svi-lupparsi (1) quando una proteina chiamata ARF, unita a una molecola più piccola, ilGDP, entra in contatto con la membrana. Poco dopo (2) un enzima addiziona ungruppo fosfato al GDP trasformandolo in GTP; questo mutamento consente all'ARFdi reclutare un complesso di proteine, il coatomero (3). L'assemblaggio dell'ARF edelle unità del coatomero sulla membrana provoca la formazione di una gemma (4),che poi si distacca (5) come vescicola munita di rivestimento. Quest'ultimo deve esse-re rimosso perché la vescicola possa fondersi con l'organello bersaglio. La rimozioneha luogo quando la conversione del GTP in GDP, con liberazione di un gruppo fosfa-to (a destra in 5), fa sì che l'ARF e le molecole del coatomero si stacchino (6 e 7).
rivestimento in forma pura ed erava-mo finalmente pronti per decifrare ilmeccanismo della gemmazione. Questocompito era più semplice. Abbiamoavviato il progetto nel 1991, dopo cheRothman aveva trasferito il propriogruppo al Memorial Sloan-KetteringCancer Center, e in appena due anniabbiamo ottenuto notevoli risultati.
La gemmazione svelata
Ci siamo chiesti innanzifutto se ilcoatomero e l'ARF fossero le unicheproteine necessarie per la gemmazionedelle vescicole COP dall'apparato diGolgi. Avevano forse bisogno di assi-stenza da parte di altre proteine? I dueelementi da soli si sono rivelati suf-ficienti. Benché fossimo partiti da unestratto grezzo contenente molte mi-gliaia di proteine diverse del citosol, lapotenza della biochimica ci aveva con-sentito di selezionare le due moleco-le che avevano realmente importanza -il coatomero e l'ARF - nonché di dimo-strare che erano in grado di funziona-re da sole.
Nella fase successiva abbiamo addi-zionato le proteine una alla volta, inve-ce che tutte insieme, per poter decifrareil ruolo svolto dalle due componenti.Abbiamo visto così che l'ARF si legaalla membrana dell'apparato di Golgi inassenza del coatomero, reclutandolo poiper fissarlo alla membrana. Il coatome-
ro non può, però, legarsi da solo al-l'apparato di Golgi (come è stato an-che dimostrato indipendentemente daRichard D. Klausner e collaboratori deiNational Institutes of Health). In segui-to l'esame con il microscopio elettroni-co ha rivelato che, quando ARF si legaalla membrana di Golgi, questa rimanepiatta; ma quando però con l'ARF siaggiunge anche il coatomero, si formauna gemma. Queste osservazioni hannopermesso di stabilire che l'ARF e ilcoatomero hanno ruoli distinti nellaformazione delle vescicole di trasporto.Il coatomero induce la curvatura del-la membrana e la gemmazione, mentrel'ARF determina il momento in cui haluogo la gemmazione stessa (ordinandoal coatomero quando e dove svolgere ilsuo compito).
Il vero e proprio processo di gemma-zione è conosciuto in maggiore detta-glio. L'ARF libero nel citosol è di solitoassociato a una molecola di guanosin-difosfato (GDP), una base azotata unitaa un glucide e a due gruppi fosfato.Quando questa unità incontra l'appara-to di Golgi, un enzima sulla superficiedella membrana sostituisce il GDP conuna molecola affine proveniente dal ci-tosol: il guanosintrifosfato (GTP). Que-sta modificazione è significativa perdue ragioni: in primo luogo, solo l'ARFlegato al GTP può agganciarsi salda-mente alla superficie della membrana,permettendo così al complesso del coa-
tomero di «salire a bordo». In secondoluogo, il legame del GTP con l'ARF èuna fase importante del processo chefornisce energia per la formazione dellevescicole.
Come dovremmo immaginare il pro-cesso di gemmazione? I singoli coato-meri sono ancorati alla membrana dauna o più molecole di ARF; il nume-ro esatto probabilmente non è né fissoné decisivo. Noi pensiamo (ma non loabbiamo ancora dimostrato) che questicomplessi finiscano per legarsi, a fiancoa fianco, in una schiera regolare, che dàorigine a una cupola sulla superficiedella membrana (qualcosa di simile aquanto fa la clatrina). Via via che laschiera cresce e si curva, la parte dellamembrana di Golgi fissata all'internodi questo rivestimento viene «scolpita»in una vescicola che sporge dalla super-ficie e, alla fine, si chiude e si separa.Pertanto le vescicole COP si formanoproprio come predetto dal modello diKanaseki e Kadota, che era stato propo-sto per le vescicole rivestite da clatri-na. La nostra ricerca porta tuttavia amodificare questo modello, indicandoche, ogni volta che una vescicola si for-ma per gemmazione, il nuovo rivesti-mento deve costituirsi pezzo per pez-zo a partire da componenti del citosol.L'assemblaggio è, inoltre, guidato dal-l'ARF e ha bisogno di GTP come fontedi energia.
Benché sia essenziale per la gemma-
zione delle vescicole COP, il rivesti-mento intralcia la fusione della mem-brana di queste vescicole con quelladell'organello bersaglio; la fusione esi-ge che le membrane entrino diretta-mente in contatto e quindi, perché ilcontenuto delle vescicole sia liberato, ilrivestimento deve cadere. Ciò avvienemediante una semplice inversione delmeccanismo di assemblaggio. In unavescicola con rivestimento completa-mente formata le molecole ARF sonoancora legate al GTP, ma quest'ul-timo perde ben presto un gruppo fo-sfato, trasformandosi in GDP e libe-rando l'energia immagazzinata nel le-game. Questa conversione fa sì che lemolecole ARF perdano la propria affi-nità per la membrana delle vescicole esi distacchino. Poiché le unità del coa-tomero hanno bisogno di queste mole-cole per il fissaggio, anch'esse si dis-sociano e lasciano dietro di sé una ve-scicola priva di rivestimento, libera difondersi con l'organello bersaglio. Inun certo qual modo, dunque, la cellu-la, durante la gemmazione, colloca nelproprio rivestimento piccole «bombe aorologeria» sotto forma di molecole diGTP; a gemmazione conclusa, le cari-che vengono fatte detonare per elimi-nare il rivestimento.
Dato che quest'ultimo viene perdutoprima che la vescicola raggiunga la pro-pria destinazione, è improbabile che siacoinvolto nell'orientazione della vesci-cola verso il bersaglio. La membranadelle vescicole prive di rivestimentocontiene invece un insieme di proteineche eseguono questa funzione; esse at-traggono anche altre proteine che pro-muovono la fusione della vescicola conla membrana bersaglio.
Un meccanismo comune
Tutte le vescicole di trasporto hannolo stesso meccanismo di formazione diquelle rivestite dalle proteine COP? Ilprimo indizio a favore di questa ipotesiè emerso quando si è scoperto che an-che l'assemblaggio del rivestimento diclatrina è innescato dal fissaggio allamembrana di molecole ARF legate alGTP. Questo risultato è stato ottenutonel laboratorio di Rothman e ci ha fatto
tornare, dopo un giro completo, al pun-to da dove eravamo partiti.
Eppure, quando si osservano le ve-scicole al microscopio, la maggior par-te di esse non sembra possedere alcunrivestimento. Questa osservazione po-trebbe far pensare che il meccanismodi gemmazione che abbiamo descrittonon si applichi alla formazione di ve-scicole diverse da quelle rivestite dalleCOP e dalla clatrina. Una simile con-clusione sarebbe però prematura. Datoche il rivestimento viene rimosso ra-pidamente dopo la gemmazione di u-na vescicola da un organello, di solitonon si riesce a individuarlo con i meto-di standard di osservazione. Effettiva-mente, la storia di questo settore di ri-cerca può essere riassunta come se-gue: i ricercatori ipotizzano dapprimache un dato tipo di vescicola sia pri-vo di rivestimento e quindi si formi pergemmazione secondo un meccanismonuovo. Successivamente il rivestimen-to viene scoperto dopo che la gem-mazione di quel tipo di vescicola è sta-ta riprodotta in un sistema sperimen-tale, che consente di «catturare» e i-dentificare lo stato transitorio munitodi rivestimento.
Tra le vescicole per le quali si è re-centemente dimostrata la necessità diun rivestimento vi sono quelle chetrasportano proteine dal reticolo en-doplasmatico all'apparato di Golgi.Esperimenti effettuati da Randy W.Schekman dell'Università della Cali-fornia a Berkeley e da Orci e collabo-ratori hanno dimostrato che una mo-lecola molto affine all'ARF svolge lostesso ruolo di quest'ultimo, in quan-to promuove il processo di gemmazio-
ne. Inoltre un diverso insieme di pro-teine, dette COP II, prende il posto delcoatomero.
Vi sono ormai ben pochi dubbi chetutte le cellule nucleate utilizzino unaserie comune di princìpi per formare levescicole di trasporto. Il processo è ditale efficacia che è stato conservato du-rante l'evoluzione dalle semplici specieunicellulari agli organismi pluricellula-ri sempre più complessi. In primo luo-go l'ARF, o una molecola a esso mol-to affine, avvia la formazione del rive-stimento legandosi al GTP e fissando-si alla membrana. In seguito questo ini-ziatore recluta ulteriori componenti, chesi riuniscono a formare un rivestimen-to a forma di cupola. Così facendo, in-ducono la regione della membrana fis-sata a questi componenti a estrofletter-si, dando origine a una vescicola. Infi-ne le molecole di GTP innescano la ri-mozione del rivestimento per consenti-re la fusione della vescicola con l'orga-nello bersaglio.
Se il rivestimento serve soltanto perla gemmazione - un processo che èfondamentalmente lo stesso in ogni or-ganello - per quale motivo la celluladeve possederne molti tipi? Il motivopiù probabile è che esso abbia il com-pito di selezionare il carico che deveessere immagazzinato in una vescico-la. In alcuni casi, le proteine da im-magazzinare possono risiedere nellamembrana stessa e legarsi direttamen-te al rivestimento; in altri casi, essepossono legarsi al rivestimento attra-verso un intermediario (un recettore),che è localizzato nello spessore dellamembrana. L'impiego di rivestimentidiversi consentirebbe a differenti tipi
42 LE SCIENZE n. 333, maggio 1996
LE SCIENZE n. 333, maggio 1996 43

LE SCIENZEraccoglitori
1996Questi raccoglitori, ora disponibili, corrispondonoai volumi LVI e LVII e rispettivamente ai fascicoli
da gennaio (n. 329) a giugno (n. 334)e da luglio (n. 335) a dicembre (n. 340).
Sono ancora disponibili i raccoglitori dal Vol. Lal Vol. LV e raccoglitori non numerati
al posto di quelli esauriti.I raccoglitori si possono richiedere direttamenteall'editore usando l'apposita cartolina allegata
alla rivista e unendo il relativo importo;gli ordini vengono evasi solo a pagamento avvenuto.
I raccoglitori si trovano anche presso
i seguenti punti vendita:
BOLOGNA - Libreria Parolini -Via U. Bassi 14
FIRENZE - Libreria Marzocco - Via de' Martelli 22/R
GENOVA - Libreria Int. Di Stefano - Via R. Ceccardi 401R
MILANO - Le Scienze S.p.A. - Piazza della Repubblica 8
NAPOLI - Libreria Guida A. - Via Port'Alba 20121
PADOVA - Libreria Cortina - Via F. Marzolo 4
PALERMO - Libreria Dante - Quattro Canti di Città
ROMA - Libreria Feltrinelli - Via V. Emanuele Orlando 84
TORINO - Libreria Zanaboni - C.so Vittorio Emanuele 41
Ogni raccoglitore L. 9000
di carico di essere inoltrati, a partire daun unico punto di origine, verso distret-ti diversi.
Rimangono altri problemi da risolve-re, molti dei quali riguardano dettagliminuti della formazione del rivestimen-to. Per esempio, esattamente in che mo-do l'ARF e analoghe molecole si fissa-no alle membrane, e quali interazioniconsentono alle proteine dei coatomeridi ancorarsi a esse? Recenti risultati in-dicano che certi lipidi possono coadiu-vare l'ARF nel reclutare un coatomerosulla superficie della membrana, e chele stesse proteine che compongono ilcarico potrebbero influenzare l'assem-blaggio di un rivestimento.
Ci piacerebbe anche conoscere me-glio le attività specifiche delle varie su-bunità del coatomero. Cionondimeno, èappagante sapere che sono state ormaidefinite molte fasi fondamentali del tra-sporto delle proteine. Nel contempo, èuna salutare lezione di umiltà notareche nessuna metropoli organizzata dal-l'uomo può competere con l'eleganteefficienza con cui cellule di ogni generetrasportano i loro «lavoratori» da unpunto all'altro.
JAMES E. ROTHMAN e LELIOORCI sono collaboratori da oltre die-ci anni. Rothman è direttore del pro-gramma di biochimica cellulare e biofi-sica presso il Memorial Sloan-KetteringCancer Center e membro della Natio-nal Academy of Sciences. Orci è diret-tore del Dipartimento di morfologia eprofessore di istologia e biologia cel-lulare presso la Facoltà di medicina del-l'Università di Ginevra in Svizzera.
ORCI L., GLICK B. S. e ROTHMAN J. E., ANew Type of Coated Vesicular CarrierThat Appears Not to Contain Clathrin:Its Possible Role in Protein Transportwithin the Golgi Stack in «Cell», 46,n. 2, 1986.
WATERS M. G., SERAFINI T. e ROTH-MAN J. E., «Coatomer»: A CytosolicProteine Complex Containing Subunitsof Non-Clathrin-Coated Golgi Tran-sport Vesicles in «Nature», 349,pp. 248-251, 17 gennaio 1991.
SERAFINI T., ORCI L., AMHERDT M.,BRUNNER M., KAHN R. A. e ROTHMAN J.E., ADP-Ribosylation Factor Is a Subu-nit of the Coat of Golgi-Derived COP-Coated Vesicles in «Cell», 67, n. 2, 18ottobre 1991.
OSTERMANN J., ORCI L., TANI K.,AMHERDT M., RAVAZZOLA M. e ROTH-MAN J. E., Stepwise Assembly of Func-tionally Active Transport Vesicles in«Cell», 75, n. 5, 3 dicembre 1993.
ROTHMAN J. E., Mechanisms of Intra-cellular Protein Transport in «Nature»,372, pp. 55-63, 3 novembre 1994.
44 LE SCIENZE n. 333, maggio 1996

![CAPITOLO II [modalit compatibilit ] · deformabilità transito rallentato e maggiore distruzione nei sinusoidi splenici ( EMOLISI) ... vescicola , (5) reticolo endoplasmatico rugoso](https://img.pdfslide.net/doc/110x75/5c65b70409d3f2b26e8d2090/capitolo-ii-modalit-compatibilit-deformabilita-transito-rallentato-e-maggiore.jpg)