Embed Size (px)
Citation preview

www.neur-one.fr
[email protected] 1 12/01/2013
LA MEMBRANE PLASMIQUE… NEURONALE
http://www.ulysse.u-bordeaux.fr/atelier/ikramer/biocell_diffusion/gbb.cel.fa.101.b3/content/access.htm#D5 http://archimede.datacenter.dsi.upmc.fr/www_poly_interactif/neurophysio/chap01.html
Un NEURONE, ou cellule nerveuse, est une cellule excitable constituant l'unité fonctionnelle de base du système nerveux. Le terme de « neurone » fut introduit dans le vocabulaire médical en 1881 par l'anatomiste allemand Heinrich Wilhelm Waldeyer. Les neurones assurent la
transmission d'un signal bioélectrique appelé influx nerveux. Ils sont 10 à 50 fois moins nombreux que les cellules gliales, seconds composants
du tissu nerveux assurant plusieurs fonctions dont le soutien et la nutrition des neurones. Santiago Ramón y Cajal, à la même époque, eut l'idée
d'utiliser la coloration au nitrate d'argent mise au point par Golgi sur le tissu nerveux, ce qui lui avait permis d'étudier en détail les prolongements des neurones (axones et dendrites) et la façon dont ils semblaient se raccorder entre eux en formant des réseaux. Ces connexions
offraient une base pour expliquer les mécanismes physiologiques du transfert d'information entre neurones.
… Heinrich Wilhelm Gottfried von Waldeyer-Hartz Santiago Ramón y Cajal Né le 6 octobre 1836 à Hehlen an der Weser, Brunswick (Allemagne) et mort le 23 janvier 1921 (à 84 ans) à Berlin, Heinrich Wi lhelm Gottfried von Waldeyer-Hartz (ou von Waldeyer-Hartz à partir de 1916, après son anoblissement par Guillaume II) est un anatomiste allemand. Il est connu pour avoir contribué à la théorie neuronale de l'organisation du système nerveux et pour avoir proposé le nom de « neurone. Les historiens des neurosciences voient en Waldeyer l'un des fondateurs en 1891, de la « théorie du neurone » et lui rendent hommage comme le créateur du terme de « neurone » pour désigner la cellule nerveuse, unité structurelle fondamentale du système nerveux. Waldeyer étaya sa théorie sur les découvertes de deux neuroanatomistes lauréats du Prix Nobel en 1906, Camillo Golgi (1843-1926) et Santiago Ramón y Cajal (1852-1934). Santiago Ramón y Cajal (1er mai 1852 à Petilla de Aragón, Navarre, Espagne - 17 octobre 1934 à Madrid, Communauté de Madrid, Espagne) est un histologiste et neuroscientifique espagnol, colauréat du prix Nobel de physiologie ou médecine en 1906 avec Camillo Golgi « en reconnaissance de leurs travaux sur la structure du système nerveux». C'est en affinant la technique d'imprégnation argentique mise au point par Golgi (et qui porte aujourd'hui son nom) qu'il contribua de façon décisive à la théorie neuronale en opposition avec la théorie réticulaire, soutenue par Golgi. Il mit en effet en évidence que les neurones étaient des entités cellulaires séparées par de fins espaces (que Sherrington nommera « synapses »), et non les fibres d'un réseau ininterrompu.
Waldeyer apprit l'espagnol pour pouvoir assimiler directement l'œuvre de Cajal et il devint son ami et mentor. Il l'initia aux techniques
d'histologie alors dominées par les Allemands. La théorie de Waldeyer fut publiée dans une série d'articles dans le principal journal médical
d'Allemagne, le Deutsche Medizinische Wochenschrift, dont l'influence devint considérable. Toutefois Waldeyer ne contribua pas à la théorie
par ses propres recherches. Cajal déclara à ce sujet : « ... bien qu'il ait soutenu la théorie par le prestige de son autorité, il n'y a pas contribué par
une seule observation personnelle, se limitant à une exposition brillante courte des preuves objectives apportées par His, Kolliker, Retzius, Van Gehuchten et moi-même, et il a inventé le terme de neurone qui devait faire fortune. »1 Le travail obstiné de Cajal au cours de la décennie
suivante permit de valider un grand nombre d'aspects de la théorie du neurone, y compris le caractère unidirectionnel de la transmission des
signaux dans les neurones. Finalement Cajal réussit à convaincre Golgi de la contiguïté plutôt que de la continuité des contacts entre neurones.
Les neurones ont deux propriétés physiologiques : l'excitabilité, c'est-à-dire la capacité de répondre aux stimulations en les convertissant en impulsions nerveuses, et la conductivité, c'est-à-dire la capacité de transmettre les impulsions.
Le neurone est composé d'un corps appelé péricaryon ou corps cellulaire ou encore soma, et de deux types de prolongements : l'axone,
unique, qui conduit le potentiel d'action de manière centrifuge, et les dendrites qui conduisent les potentiels synaptiques en direction centripète.
La morphologie, la localisation et le nombre de ces prolongements, ainsi que la forme du soma, varient et contribuent à définir différentes familles morphologiques de neurones. De plus, certains neurones sont entourés d’une gaine de myéline formée par l’enroulement d’une cellule
de Schwann autour de l’axone, sauf au niveau des nœuds de Ranvier. Axones et dendrites de neurones différents entrent en contact et
transmettent l’information de cellule à cellule via des structures spécialisées : les synapses.

www.neur-one.fr
[email protected] 2 12/01/2013
Un neurone est une cellule du système nerveux spécialisée dans la communication et le traitement d'informations.
Du point de vue de leur fonction, les neurones ont pour rôle de faire circuler les informations entre l'environnement et l'organisme, ou
au sein de l'organisme. Un neurone est donc une cellule excitable, c'est-à-dire qu’un stimulus peut entraîner la formation dans la cellule d’un signal bioélectrique ou influx nerveux, qui pourra être transmis à d’autres neurones ou à d’autres tissus pour les activer (des muscles, des glandes
sécrétrices…). Certains neurones, lors de leur excitation, vont sécréter une neurohormone dans l’environnement des capillaires pour finalement
passer dans le sang.
Comme dans toute cellule, il existe de part et d'autre de la membrane une différence de potentiel électrique (polarisation), appelée potentiel de repos : l'extérieur de la cellule est positif par rapport à l'intérieur, négatif. Mais les cellules excitables telles que le neurone se
distinguent des autres par la capacité à créer un autre phénomène électrique, le potentiel d'action, ou influx nerveux : en quelques millièmes de
seconde et sur une infime longueur de membrane, un afflux de sodium (ion positif) entre dans la cellule, ce qui dépolarise la membrane. Ce
potentiel d'action se propage spontanément ensuite tout au long de la membrane. Il permet de transmettre un message, information sensitive ou ordre moteur. Les messages arrivent à un neurone donné par ses dendrites. Celles-ci amènent les messages au corps cellulaire (conduction
centripète). Le corps cellulaire analyse les messages et en produit de nouveaux, lesquels cheminent le long de l'axone (conduction centrifuge).
Un neurone est ainsi relié à d'autres neurones ou à des cellules musculaires. Le point de jonction entre l'axone d'un neurone et les dendrites d'un
autre neurone s'appelle la synapse. À ce niveau, la transmission du message d'un neurone à un autre fait intervenir diverses substances appelées neurotransmetteurs.
La membrane plasmique délimite le contour de tout le neurone, des épines dendritiques aux terminaisons axonales. C’est elle qui
permet de réguler les flux entrant et sortant d'électrolytes (sodium potassium entre autres) et de molécules grâce à la spécificité de sa
perméabilité sélective. De quelle manière l'activité électrique de neurones est-elle générée? L'activité électrique des neurones est le résultat de mouvements de particules électriques de part et d'autre de la membrane. Ce flux électrique est le résultat de l'entrée et de sortie d'ions à travers la
membrane de la cellule.
Le déplacement passif des ions suit un gradient de concentration qui va du plus concentré vers le moins concentré et un gradient de
potentiel qui va du plus chargé vers le moins chargé. La distribution des ions est également le résultat de l'ouverture de la fermeture de canaux ioniques que l'on constate à l'intérieur de la membrane du neurone, ainsi que de pompes membranaires qui rétablissent et maintiennent ces
différences de concentration ionique entre les deux milieux intra- et extra-cellulaire.
Quand un neurone est au repos, le côté intracellulaire, c'est-à-dire à l'intérieur du neurone, de la membrane,
possède une charge électrique d'environ - 70 mV par rapport au côté extracellulaire c'est-à-dire par rapport à l'autre
face de la membrane du neurone. Cette charge porte le nom de potentiels de repos. Elle est le résultat d'une
distribution en inégale des ions d'un côté de la membrane et de l'autre côté de la membrane du neurone. Une
distribution inégale des ions potassium et des ions chlore contribuent au potentiel de repos
Les grosses protéines qui sont chargées négativement sont trop importantes pour pouvoir sortir de la cellule. Quand le neurone est stimulé, les canaux ioniques de la membrane viennent modifier leur état ce qui aboutit à un
changement de la distribution des ions autour de la membrane et qui augmente ou qui diminue brutalement le
voltage à travers la membrane, de quelques millivolts. Une légère augmentation du voltage à travers la membrane
est appelée hyperpolarisation, alors qu'une légère diminution de ce voltage est appelée, au contraire, dépolarisation. On appelle cette modification des potentiels gradués.
Un potentiel d'action est un événement qui correspond un changement rapide et ample de la polarité de la
membrane d'un axone. Le déclenchement du potentiel d'action est le résultat de l'atteinte du seuil de dépolarisation
du potentiel de membrane d'environ -50 mV. Le voltage transmembranaire va s'inverser subitement c'est-à-dire que le côté intracellulaire va devenir positif par rapport au côté extracellulaire, puis s'inverser à une nouvelle fois
tout aussi brutalement
LA MEMBRANE PLASMIQUE NEURONALE : UNITÉ DE RÉCEPTION ET DE CONDUCTION DE
L'INFORMATION.
LES CONCEPTS DE CELLULE… PUIS DE NEURONE
Découverte et premières observations de la cellule.
D’après les philosophes anciens et les naturalistes, en particulier Aristote dans l’Antiquité et Paracelse à la Renaissance, « les animaux et les plantes, quelle que soit la complexité de leur organisation, ne sont constitués que d’un petit nombre d’éléments qui se répètent dans chacun
d’eux ». Plusieurs siècles plus tard, grâce à l’invention des lentilles grossissantes, on découvrit le monde des dimensions microscopiques. Les
observations révélèrent qu’un organisme entier peut être constitué d’une seule cellule, comme chez les protozoaires ou bien de plusieurs cellules
différenciées et agencées en tissus et organes comme c’est le cas chez les êtres multicellulaires.

www.neur-one.fr
[email protected] 3 12/01/2013
Théorie tissulaire
Les premières lunettes ont été fabriquées en Europe au treizième siècle, et les premiers microscopes (composés de deux lentilles), ont été construits à la fin du seizième siècle.Au milieu du dix-septième siècle, une poignée de scientifiques d’avant-garde ont utilisé des microscopes
qu’ils avaient construits eux-mêmes et ont découvert un monde qui n’aurait jamais été révélé à l’œil nu. Le microscope composé, constitué de
deux lentilles, a été mis au point à la fin du XVIe siècle. Son grossissement atteignait alors 150 à 200 fois.
La découverte des cellules est généralement attribuée à Robert Hooke, microscopiste anglais qui, à l’âge de 27 ans, s’était vu attribuer un poste de conservateur à la Royal Society, académie scientifique la plus renommée d’Angleterre.. L’utilisation du microscope composé a le
permis à HOOKE de décrire dans Micrographia en 1665 pour la première fois l’organisation alvéolaire de fins copeaux de liège : il propose le
mot «cellule» (du latin cellula : petite chambre), pour désigner les minuscules cavités géométriques qu’il découvre (il s’agit en fait de cellules
végétales mortes, dont il a observé les parois). En effet, une des nombreuses questions que Hooke essayait de résoudre était celle-ci : pourquoi les bouchons de liège (faits d’écorce d’arbre) convenaient-ils si bien pour conserver l’air dans une bouteille ? « Je pris un morceau d’écorce bien
propre et, avec un couteau tranchant comme un rasoir, j’en coupai un morceau et … l’examinant avec un microscope, je pus voir qu’il était un
peu poreux, ressemblant à un rayon de miel ». Hooke appela les pores cellules, parce qu’ils lui rappelaient les cellules occupées par les moines
dans un monastère. En réalité Hooke avait observé les parois des cellules vides dans un tissu végétal mort, parois qui avaient été, à l’origine, produites par les cellules vivantes qu’elles entouraient.
Microscope de Hooke
Le microscope composé utilisé par Hooke. Une rotule sphérique et un anneau permettent le positionnement de l’instrument. Le système optique est constitué de trois lentilles (objectif, oculaire et lentille de champ). Le système d’illumination comporte une lentille boule qui focalise la lumière d’une flamme sur l’objet observé. b) Observation d’une fine lamelle de liège à l’aide de ce même microscope.
Sur ces entrefaites, Anton van Leeuwenhoek, un Hollandais qui vivait de la vente de vêtements et boutons, occupait son temps libre à polir des lentilles et construire des microscopes d’une qualité remarquable. Pendant 50 ans, Leeuwenheok envoya à la Royal Society de Londres,
des lettres décrivant ses observations microscopiques. Leeuwenhoek fut le premier à examiner une goutte d’eau provenant d’un étang et, à son
grand étonnement, il observait quantité d’ « animalcules » microscopiques qui filaient çà et là sous ses yeux. Il fut aussi le premier à décrire
diverses formes de bactéries qu’il trouvait dans de l’eau où du poivre avait été trempé ou dans les raclures de ses propres dents. Ses premières lettres à Royal Society, décrivant un monde encore inconnu, furent accueillies avec un tel scepticisme que la société chargea son conservateur,
Robert Hooke, de confirmer les observations. Hooke le fit, et Leeuwenhoek devint bientôt une célébrité mondiale, recevant en Hollande la visite
de Pierre le Grand de Russie et de la reine d’Angleterre.
Ces structures sont ensuite observées dans de très nombreux autres tissus végétaux : MALPIGHI et GREW y décrivent, entre 1670 et 1680, de nombreux «tubes et vésicules» ; ce sont les débuts de l’anatomie végétale. À partir de 1674, LEEUWENHOEK, au moyen d’une
simple loupe (ou microscope simple), mais de qualité optique remarquable et bien supérieure à celle des microscopes composés de ses
contemporains (grossissant près de 300 fois), décrit une multitude de micro-organismes vivants : Protistes et même Bactéries ! Au début du
XIXe siècle, l’idée d’une organisation en tissus des organismes végétaux commence à se faire jour ; DUTROCHET réintroduit, en 1820, le terme de cellule, qui avait disparu depuis 150 ans.
C’est ainsi que l’apparition de la notion de membrane cellulaire remonte donc aux premiers travaux de cytologie du XVIIe siècle et
accompagne la mise en œuvre d’un des premiers microscopes (Hooke publie les observations réalisées à l’aide de ce microscope dans
Micrographia en 1665) : « (. . . ) I could exceedingly plainly perceive it to be all perforated and porous, much like a Honey-comb, but that the pores of it were not
regular (. . . ) these pores, or cells, (. . . ) were indeed the first microscopical pores I ever saw, and perhaps, that were ever seen, for I had not met with
any Writer or Person, that had made any mention of them before this (. . . ) »
Théorie cellulaire
Un progrès technologique important survient en 1827, lorsque AMICI apporte des corrections à la construction des microscopes
composés et supprime les aberrations chromatiques, qui en limitaient beaucoup les possibilités. Une foule d’observations nouvelles s’en suit et,
en 1833, BROWN décrit le noyau de cellules vivantes (dans des cellules d’orchidées) et l’interprète comme une structure cellulaire constante. Ces observations débouchent en 1838-1839 sur la formulation de la théorie cellulaire, élaborée par SCHLEIDEN et SCHWANN
(respectivement anatomistes animal et végétal), selon laquelle tout être vivant complexe est constitué de cellules, qui représentent l’unité de base
structurale et fonctionnelle de la vie. Cependant, ces auteurs imaginent encore que les cellules peuvent apparaître par génération spontanée, à la
suite d’une coagulation de vésicules formées à partir d’un « liquide primordial», comme des «bulles dans le pain». Le concept de protoplasme, substance fondamentale vivante, est dégagé par DUJARDIN, chez les Protistes (1835), puis généralisé entre 1840 et 1845. En 1855, VIRCHOW
démontre, par des observations vitales, que toute cellule est issue d’une cellule préexistante (omnis cellula e cellula) ; la cellule est alors
envisagée sous son aspect actuel.
En résumé, ne fut pas avant les années 1830 qu’on réalisa l’importance universelle des cellules. En 1838, Matthias Schleiden, homme
de loi allemand converti à la botanique, conclut que, en dépit de différences dans la structure de divers tissus, les plantes sont constituées de
cellules et que l’embryon végétal provint d’une seule cellule. En 1839, Theodor Schwann, zoologiste allemand et collègue de Schleiden, publia
une étude d’ensemble sur la base cellulaire de la vie animale. Schwann arrivait à la conclusion que les cellules des plantes et des animaux ont une structure semblable et proposait les trois premiers principes de la théorie cellulaire :
Tous les organismes sont composés d’une ou plusieurs cellules.

www.neur-one.fr
[email protected] 4 12/01/2013
Chaque cellule est entourée d'une membrane cytoplasmique.
Chaque cellule dérive d’une cellule préexistante.
Découverte des principaux organites des cellules eucaryotiques
Dans les années qui suivent, avec le développement des techniques histologiques, et les perfectionnements considérables apportés au
microscope composé par ABBE et ZEISS (1870-1885), les principales structures cellulaires sont décrites. Tous les organites cellulaires, à
l’exception des lysosomes et des peroxysomes, étaient identifiés dès la fin du XIX e siècle. Le microscope optique ayant atteint sa limite théorique de résolution, l’organisation précise de ces structures ne sera décrite qu’après la mise au point du microscope électronique (1931), dont
le développement nécessitait de nouvelles avancées en physique. L’histoire du microscope photonique avait duré plus de 250 ans ! Il faut
souligner, dans toute cette histoire, l’importance des biologistes végétaux, en raison des caractéristiques propres à leur matériel : cellules de
grande taille, présence d’une paroi épaisse vue et d’organites colorés.
Schémas simplifiés de cellules
La cellule est une unité vivante qui a sa vie propre, c'est-à-dire qui a sa propre homéostasie (biochimie), mais en même temps doit répondre aux besoins de l'organisme, c'est-à-dire doit être réceptive. Les cellules sont caractérisées par leur membrane, leur noyau et leur cytoplasme. Une cellule a un diamètre de 2 à 100µm et contient environ un milliard de molécules protéiques, constituant à peu près 60% de sa masse sèche. On pense qu'il y a environ 10.000 types différents de protéines dans une cellule. Pour bien fonctionner, les cellules ont compartimenté leur processus biochimiques dans le cytoplasme et ces compartiments sont les organites cellulaires (ou organelles)
Il est désormais possible d’isoler les constituants cellulaires, de comparer les images obtenues en microscopie électronique sur les constituants isolés et les coupes de cellules, et surtout de déterminer les activités biologiques de ces constituants.
Découverte et premières observations de neurones
La découverte selon laquelle le système nerveux est constitué de cellules qui lui sont propres résulte de l’expansion notable de la
microscopie et des techniques histologiques sur coupes fines à la fin du XIXème siècle mais aussi de la découverte de deux colorations extraordinaires pour visualiser les cellules nerveuses qui ont été nommées à cette occasion neurones. La première est due à Nissl qui montra que
le crésyl violet était susceptible de se fixer sur des corpuscules (nommés depuis corps de Nissl) présents dans le cytoplasme des neurones autour
du noyau. Les corps des cellules nerveuses et eux seuls sont donc colorées. Vers la même époque Golgi découvre qu’en plongeant un tissu
nerveux dans une solution de chrome argentique un nombre très limité de neurones se colorent dans leur totalité en noir. Cette technique sera mise à profit par son contemporain Ramon y Cajal pour décrire les neurones de tout le système nerveux, leur développement et leur régénération
Tout deux auront le prix Nobel en 1906 bien que Golgi défende l’idée que les neurones reliés entre eux ne forment qu’un seul ensemble comme
le système vasculaire, alors que Ramon y Cajal est sûr d’avoir vu des contacts entre cellules et défend l’idée de synapse déjà proposée à l’époque
par Claude Bernard. Le débat sera tranché en 1950 avec la visualisation de la jonction neuromusculaire en microscopie électronique par Couteaux.
Portraits à la fin du XIXème siècle des trois découvreurs des neurones, Nissl, le pionnier, Golgi et Santiago y Cajal respectivement découvreur et utilisateur de la coloration argentique. (Nobel lectures, Physiology and Medicine, 1901-1921, Elsevier, 1967).
La microscopie optique a permis de découvrir la diversité des neurones et de savoir que la plupart d’entre eux ont des corps cellulaires
minuscules de 5 à 10 µm, des dendrites de 1mm et moins. Mais aussi la surprise a été grande de découvrir des axones de plus d’un mètre de long
dans les cordons postérieurs de la moelle. Les neurones sont constitués de trois parties : un arbre dendritique plus ou moins touffu, un corps cellulaire, et un prolongement unique ramifié à son extrémité nommé axone (figure ci-dessous). Ces trois éléments sont polarisés. En effet le
neurone reçoit toutes les informations de l’extérieur grâce aux synapses dendritiques et somatiques. C’est la zone réceptrice. Le corps cellulaire
contenant le noyau a un rôle supplémentaire en synthétisant la majorité des protéines : il assure la maintenance des expansions cytoplasmiques
surtout de l’axone. L’axone synthétise et transporte des transmetteurs chimiques qu’il délivre à son extrémité dans des fentes synaptiques grâce au potentiel d’action qui parcourt sa membrane. C’est la partie émettrice du neurone. La découverte des fluctuations de leur potentiel de
membrane dans les années 40 puis la proposition par Weiss en 1948, qu’il existait un trafic intra axonal a relancé l’intérêt pour la biologie des
neurones en particulier pour leur régénération.
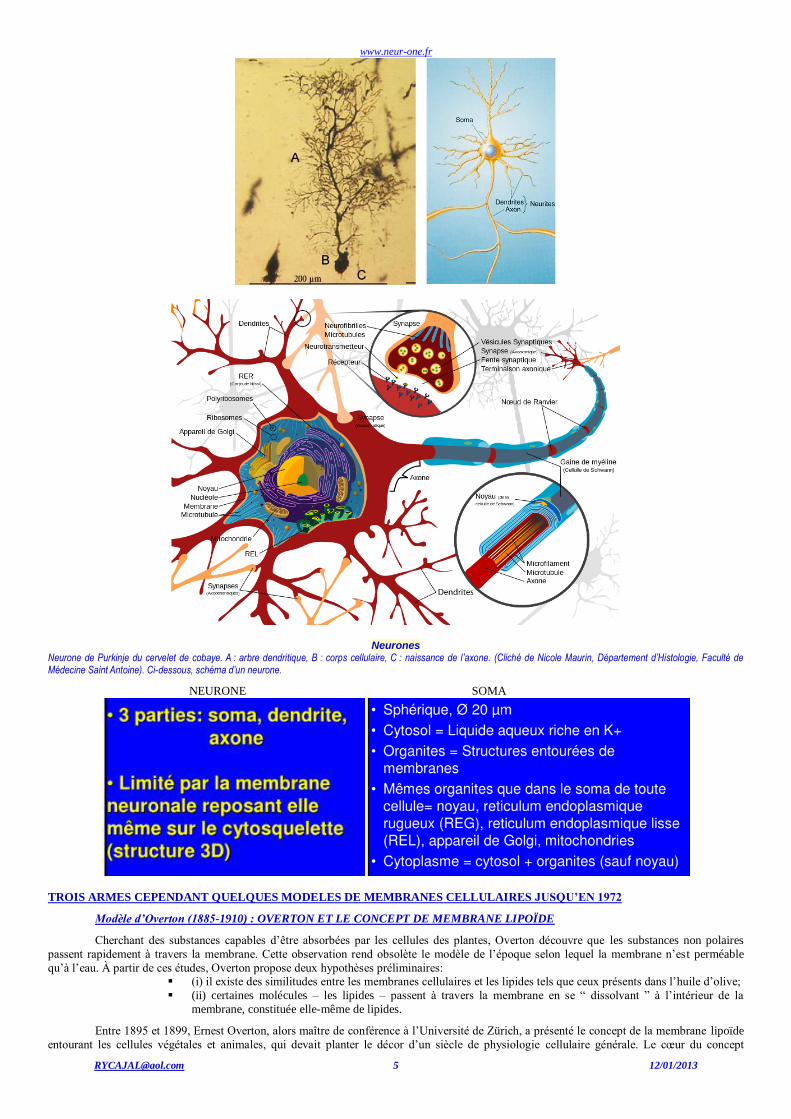
www.neur-one.fr
[email protected] 5 12/01/2013
Neurones Neurone de Purkinje du cervelet de cobaye. A : arbre dendritique, B : corps cellulaire, C : naissance de l’axone. (Cliché de Nicole Maurin, Département d’Histologie, Faculté de Médecine Saint Antoine). Ci-dessous, schéma d’un neurone.
NEURONE SOMA
TROIS ARMES CEPENDANT QUELQUES MODELES DE MEMBRANES CELLULAIRES JUSQU’EN 1972
Modèle d’Overton (1885-1910) : OVERTON ET LE CONCEPT DE MEMBRANE LIPOÏDE
Cherchant des substances capables d’être absorbées par les cellules des plantes, Overton découvre que les substances non polaires
passent rapidement à travers la membrane. Cette observation rend obsolète le modèle de l’époque selon lequel la membrane n’est perméable
qu’à l’eau. À partir de ces études, Overton propose deux hypothèses préliminaires: (i) il existe des similitudes entre les membranes cellulaires et les lipides tels que ceux présents dans l’huile d’olive;
(ii) certaines molécules – les lipides – passent à travers la membrane en se “ dissolvant ” à l’intérieur de la
membrane, constituée elle-même de lipides.
Entre 1895 et 1899, Ernest Overton, alors maître de conférence à l’Université de Zürich, a présenté le concept de la membrane lipoïde
entourant les cellules végétales et animales, qui devait planter le décor d’un siècle de physiologie cellulaire générale. Le cœur du concept

www.neur-one.fr
[email protected] 6 12/01/2013
d’Overton était que la membrane cellulaire est constituée essentiellement de substances lipidiques et que le coefficient de perméation d’une
molécule à travers la membrane est directement proportionnel à sa lipophilie. La règle d’Overton stipule que la perméabilité (P) d’une molécule à travers la membrane cellulaire peut être exprimée selon la formule suivante :
P = KD/dx
où K est le coefficient de partition entre les phases huileuse et aqueuse (lipophilie), D le coefficient de diffusion et dx l’épaisseur de la
membrane.
Overton avait également observé que les lipides empêchent la migration des particules chargées, en prédisant que les ions et les
substances non lipophiles doivent utiliser des voies spécifiques à travers la membrane. Le concept de membrane lipoïde a évolué en modèle de la
mosaïque fluide incluant des domaines lipidiques et des régions de pores constitués de protéines, tandis que la notion que les substances
traversent la membrane cellulaire proportionnellement à leur solubilité lipidique a résisté à l’épreuve du temps.
En 1917, Langmuir formule l’hypothèse selon laquelle les lipides forment une monocouche sur l’eau en s’orientant verticalement,
leurs chaînes carbonées étant hors de l’eau alors que leurs groupements polaires restent en contact avec la surface de l’eau. En résumé :
Modèle de Gorter et Grendel (1937)
En 1925, Gorter et Grendel solubilisent les lipides d’un globule rouge à l’aide d’acétone. En utilisant une cuve de Langmuir, ils
mesurent la superficie de la monocouche de lipides ainsi formée. Parallèlement, ils évaluent la surface d’un globule rouge en supposant qu’il a une forme sphérique. La surface calculée d’un globule rouge est alors égale à la moitié de la superficie mesurée de la monocouche de lipides
provenant de ce globule rouge ; ils concluent que la membrane est une double couche de lipides (figure ci-dessous). La sous-estimation de la
surface du globule rouge liée à l’hypothèse erronée sur sa forme n’a été sans conséquence que parce que l’extraction des lipides par l’acétone
était incomplète : les deux erreurs se compensaient !
Modèles de Danielli et Davson et modèle de Robertson (1935-1970)
Le modèle de bicouche lipidique ne permet pas d’expliquer que la tension superficielle de l’interface membrane-eau est toujours
inférieure à celle d’une interface huile-eau. La faible tension superficielle de l’interface membrane-eau ne peut s’expliquer si la membrane ne
contient que des lipides ; au contraire, lorsque des protéines sont ajoutées à l’huile ou l’eau, elles adsorbent à l’interface et induisent une réduction de la tension superficielle de l’interface huile-eau. Ces différentes observations ont conduit Danielli et Davson à proposer un nouveau
modèle en 1935 dans lequel les protéines sont parties intégrantes de la membrane. Chaque côté de la bicouche lipidique est recouvert d’un
manteau de protéines globulaires, les protéines étant attachées par des liaisons ioniques aux têtes polaires des lipides. Ce modèle est étendu
quelques années plus tard à des protéines dépliées et qui peuvent constituer un fin manteau protéique en contact étroit avec les lipides.
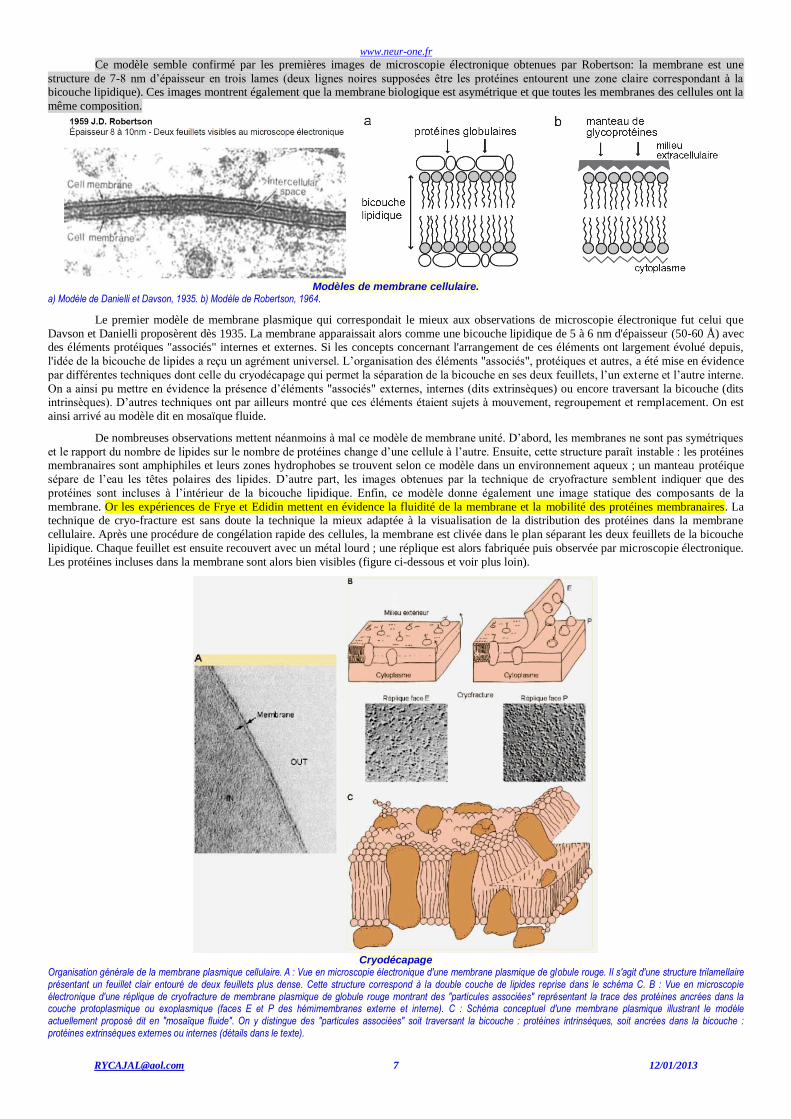
www.neur-one.fr
[email protected] 7 12/01/2013
Ce modèle semble confirmé par les premières images de microscopie électronique obtenues par Robertson: la membrane est une
structure de 7-8 nm d’épaisseur en trois lames (deux lignes noires supposées être les protéines entourent une zone claire correspondant à la bicouche lipidique). Ces images montrent également que la membrane biologique est asymétrique et que toutes les membranes des cellules ont la
même composition.
Modèles de membrane cellulaire.
a) Modèle de Danielli et Davson, 1935. b) Modèle de Robertson, 1964.
Le premier modèle de membrane plasmique qui correspondait le mieux aux observations de microscopie électronique fut celui que
Davson et Danielli proposèrent dès 1935. La membrane apparaissait alors comme une bicouche lipidique de 5 à 6 nm d'épaisseur (50-60 Å) avec des éléments protéiques "associés" internes et externes. Si les concepts concernant l'arrangement de ces éléments ont largement évolué depuis,
l'idée de la bicouche de lipides a reçu un agrément universel. L’organisation des éléments "associés", protéiques et autres, a été mise en évidence
par différentes techniques dont celle du cryodécapage qui permet la séparation de la bicouche en ses deux feuillets, l’un externe et l’autre interne.
On a ainsi pu mettre en évidence la présence d’éléments "associés" externes, internes (dits extrinsèques) ou encore traversant la bicouche (dits intrinsèques). D’autres techniques ont par ailleurs montré que ces éléments étaient sujets à mouvement, regroupement et remplacement. On est
ainsi arrivé au modèle dit en mosaïque fluide.
De nombreuses observations mettent néanmoins à mal ce modèle de membrane unité. D’abord, les membranes ne sont pas symétriques
et le rapport du nombre de lipides sur le nombre de protéines change d’une cellule à l’autre. Ensuite, cette structure paraît instable : les protéines membranaires sont amphiphiles et leurs zones hydrophobes se trouvent selon ce modèle dans un environnement aqueux ; un manteau protéique
sépare de l’eau les têtes polaires des lipides. D’autre part, les images obtenues par la technique de cryofracture semblent indiquer que des
protéines sont incluses à l’intérieur de la bicouche lipidique. Enfin, ce modèle donne également une image statique des composants de la
membrane. Or les expériences de Frye et Edidin mettent en évidence la fluidité de la membrane et la mobilité des protéines membranaires. La technique de cryo-fracture est sans doute la technique la mieux adaptée à la visualisation de la distribution des protéines dans la membrane
cellulaire. Après une procédure de congélation rapide des cellules, la membrane est clivée dans le plan séparant les deux feuillets de la bicouche
lipidique. Chaque feuillet est ensuite recouvert avec un métal lourd ; une réplique est alors fabriquée puis observée par microscopie électronique.
Les protéines incluses dans la membrane sont alors bien visibles (figure ci-dessous et voir plus loin).
Cryodécapage
Organisation générale de la membrane plasmique cellulaire. A : Vue en microscopie électronique d'une membrane plasmique de globule rouge. Il s'agit d'une structure trilamellaire présentant un feuillet clair entouré de deux feuillets plus dense. Cette structure correspond à la double couche de lipides reprise dans le schéma C. B : Vue en microscopie électronique d'une réplique de cryofracture de membrane plasmique de globule rouge montrant des "particules associées" représentant la trace des protéines ancrées dans la couche protoplasmique ou exoplasmique (faces E et P des hémimembranes externe et interne). C : Schéma conceptuel d'une membrane plasmique illustrant le modèle actuellement proposé dit en "mosaïque fluide". On y distingue des "particules associées" soit traversant la bicouche : protéines intrinsèques, soit ancrées dans la bicouche : protéines extrinsèques externes ou internes (détails dans le texte).

www.neur-one.fr
[email protected] 8 12/01/2013
Modèle de Singer et Nicolson (1972)
Le premier modèle de membrane plasmique qui correspondait le mieux aux observations de microscopie électronique fut celui que Davson et Danielli proposèrent dès 1935. La membrane apparaissait alors comme une bicouche lipidique de 5 à 6 nm d'épaisseur (50-60 Å) avec
des éléments protéiques "associés" internes et externes. Si les concepts concernant l'arrangement de ces éléments ont largement évolué depuis,
l'idée de la bicouche de lipides a reçu un agrément universel. L’organisation des éléments "associés", protéiques et autres, a été mise en évidence
par différentes techniques dont celle du cryodécapage qui permet la séparation de la bicouche en ses deux feuillets, l’un externe et l’autre interne. On a ainsi pu mettre en évidence la présence d’éléments "associés" externes, internes (dits extrinsèques) ou encore traversant la bicouche (dits
intrinsèques). D’autres techniques ont par ailleurs montré que ces éléments étaient sujets à mouvement, regroupement et remplacement. On est
ainsi arrivé au modèle dit en mosaïque fluide.
Singer et Nicolson partent du modèle de la bicouche lipidique de Gorter et Grendel et utilisent les découvertes de Frye et Edidin en matière de fluidité de la membrane et de mobilité des protéines. Ce modèle de la mosaïque fluide reste le modèle de référence, même s’il a subi
quelques aménagements. La membrane plasmique y est décrite comme une bicouche lipidique fluide dans laquelle flottent des protéines :
« a two dimensional oriented solution of integral proteins (. . . ) in the viscous phospholipid bilayer. »
Dans le modèle original de Singer et Nicolson, lipides et protéines sont distribués plus ou moins aléatoirement. Les protéines sont
insérées plus ou moins profondément dans la bicouche lipidique sous forme compacte : les protéines peuvent être intégrales (protéines
transmembranaires) ou adsorbées à la surface de la bicouche. Les chaînes polypeptidiques, le plus souvent organisées sous forme d’hélices α
[Unwin 1984] et contenant de nombreux résidus d’acides aminés hydrophobes, prennent la place des lipides et assurent ainsi la continuité de la partie hydrophobe de la membrane. Les parties les plus hydrophiles des protéines émergent sur au moins une des deux faces de la bicouche. La
face externe de la membrane est rendue encore plus hydrophile par la présence de résidus osidiques. L’agitation thermique est responsable de la
diffusion et de la rotation de toutes les molécules dans le plan de la membrane.
Modèle de Singer et Nicolson, 1972.
a) Schéma 2D. b) Schéma 3D tiré de l’article [Singer 72]. c) Hélices α proposées par Unwin et Henderson en 1984. d) Modèle amélioré de la mosaïque fluide, développé dans les années 80. Les oses sont tous situés sur le côté extracellulaire de la membrane. D’après [Bretscher 85].
Dans les décennies suivantes, de nombreux travaux montrent que la plupart des protéines membranaires ne diffusent pas aussi
librement que ne le suppose le modèle de Singer et Nicolson. Quelques mécanismes de diffusion sont recensés par Jacobson en 1995. Jacobson
insiste particulièrement sur le rôle du cytosquelette dans le confinement des protéines, s’appuyant pour cela sur les travaux de Kusumi. En
parallèle, des travaux ont continué à porter sur les hétérogénéités de composition moléculaire dans la membrane, donnant lieu au modèle des radeaux lipidiques de Simons. L’ensemble de ces travaux sont développées dans :
http://tel.archives-ouvertes.fr/docs/00/09/70/10/PDF/2006_PhDthesis_wawrezinieck.pdf
Dès 1970, les expériences de Frye et Edidin mettent en évidence la fluidité de la membrane et la mobilité des protéines membranaires. Dans l’expérience de Frye et Edidin, les antigènes de surface de cellules humaines et de souris sont marqués par double immunofluorescence.
Juste après la fusion des deux cellules humaines et de souris, deux hémisphères de couleurs différentes sont visibles ; pendant les minutes
suivantes, les antigènes diffusent dans la membrane de la cellule hybride issue de la fusion, jusqu’à l’obtention d’une distribution aléatoire des
protéines humaines et de souris.
Diffusion confinée dans la membrane cellulaire
L’hypothèse selon laquelle les molécules s’ordonnent aléatoirement dans la membrane est néanmoins rapidement remise en question.
Dès 1974, de nombreuses données sur la distribution et la fonction des protéines membranaires indiquent que, dans une même membrane,

www.neur-one.fr
[email protected] 9 12/01/2013
différentes protéines peuvent être localisées dans des environnements différents. D’abord, des résultats de mesure de résonance magnétique ont
mis en évidence une possible séparation de phase des lipides dans le plan de la membrane.
Jain et White (1974) proposent alors un modèle de membrane possédant une organisation lipidique à longue distance. La membrane est
composée d’un certain nombre de plateaux lipidiques (plate-model) en mouvement les uns par rapport aux autres (figure a ci-dessous). Des
bicouches ayant des compositions et des organisations différentes peuvent coexister dans le plan de la membrane (figure b ci-dessous). De telles
différences sont supposées provenir de conformations moléculaires différentes et de la spécificité de certaines interactions moléculaires.
Modèles de membrane non homogène.
a et b) Modèle de Jain et White, 1977. c) Modèle de Jacobson, 1995 : différents mécanismes de diffusion latérale. Diffusion gênée (A) par des agrégats de molécules ou (B) par le
cytosquelette. (C) Diffusion dirigée le long d’un microtubule de cytosquelette. (D) Diffusion libre.
Propriétés des membranes biologiques
COMPOSITION CHIMIQUE DES MEMBRANES

www.neur-one.fr
[email protected] 10 12/01/2013
Premières données expérimentales
Dès le début du siècle, à la suite de travaux sur la perméabilité cellulaire vis-à-vis de diverses substances organiques, plusieurs auteurs ont proposé que la membrane plasmique soit essentiellement constituée de lipides. Ils avaient en effet observé une relation étroite entre le
caractère liposoluble d’un composé et sa capacité à pénétrer dans les cellules. Certaines substances, telles que les cétones ou les éthers (qui sont
des solvants des lipides), ou d’autres qui sont elles-mêmes solubles dans ces solvants, s’accumulent beaucoup plus rapidement dans le
cytoplasme que celles plus solubles dans l’eau. Ce résultat semblait paradoxal dans la mesure où les cellules présentent surtout des besoins nutritifs en composés hydrosolubles tels que les sucres ou les acides aminés. De fait, certaines exceptions (le méthanol ou l’éthylèneglycol, par
exemple), conduisaient à penser que la membrane n’était pas homogène sur toute sa surface et qu’elle devait pouvoir laisser passer de tels
composés, peu liposolubles.
Les lipides forment une double couche (épaisse de 5 à 6 nm) qui est relativement imperméable au passage de la plupart des molécules hydrosolubles (protéines, hormones, ions). Cette propriété est bien illustrée par une expérience de diffusion des ions potassium (K+). Alors que
les ions K+ arrivent à un équilibre de diffusion dans l'eau sur une distance de 6 nm en 5 millisecondes, il leur faut 12 jours pour arriver à
l'équilibre à travers une double couche de lipides. Même les petites molécules comme le glucose ou l'adrénaline mettent un temps considérable
pour traverser. La membrane est donc une barrière très efficace ! Mais elle peut facilement être franchie par des molécules hydrophobes telles que les alcools, les stéroïdes et les anesthésiques généraux (pentobarbital). Les propriétés de perméabilité d'une double couche lipidique
artificielle sans protéines sont indiquées ci-dessous.
Perméabilité d'une double couche lipidique
La structure en double couche est due aux propriétés amphiphiles des molécules lipidiques. Celles-ci possèdent ainsi une extrémité hydrophile (aimant l'eau ou polaire) et une extrémité hydrophobe (craignant l'eau ou apolaire). Diagramme montrant l’étendue des valeurs des coefficients de perméabilité (cm.s-1) pour le passage de diverses catégories de molécules et d’ions à travers les bicouches lipidiques artificielles.
En 1926, un autre type de résultats était obtenu par une approche chimique directe sur une catégorie de cellules qui s’avère être, depuis
lors, un « système-modèle » pour l’étude des membranes : les hématies de Mammifères. Ces cellules, non nucléées, sont également dépourvues de tout système membranaire interne et la membrane cytoplasmique entoure un hyaloplasme essentiellement constitué d’hémoglobine.
L’hémolyse de ces cellules permet de les vider de leur contenu et de purifier aisément, après centrifugation, leur seule membrane limitante ; on
obtient alors ce qu’on appelle des fantômes d’hématies.
L’analyse chimique de ces structures a montré qu’elles contenaient une proportion importante de lipides (40 % de la masse), associés à
des protéines et des glucides. Des expériences élégantes, utilisant la propriété propre aux lipides de constituer des monocouches moléculaires à
la surface de l’eau, ont conduit à suggérer que la membrane était constituée d’une double couche lipidique. En effet, la surface de la monocouche, obtenue à partir de la totalité des lipides membranaires purifiés provenant d’une quantité connue d’hématies, correspondait à peu
près au double de la surface de leurs membranes cytoplasmiques !
D’autres arguments, tels qu’une forte résistance électrique (mesurée sur des axones géants de calmar), ou une grande sensibilité des
membranes vis-à-vis des agents lipolytiques : solvants, détergents, certaines enzymes (phospholipases), renforçaient l’idée qu’une composante lipidique jouait un rôle majeur dans l’organisation membranaire.
Les constituants membranaires
A. Les lipides
Ils sont tous amphiphiles et organisés en bicouche lipidique. La partie hydrophile (polaire) de la molécule est la tête, la partie hydrophobe (apolaire) est la queue.
Les acides gras présents sont saturés et insaturés.

www.neur-one.fr
[email protected] 11 12/01/2013
Les lipides présents dans les différentes classes de lipides membranaires sont faits de :
Phospholipides (glycérophospholipides et sphingomyélines),
Cholestérol : l’ensemble de la structure est apolaire à l’exception du OH alcoolique qui est polaire. Il
participe à la fluidité membranaire
Glycolipides : les cérébroglucosides, cérébrogalactosides et gangliosides sont localisés sur la face
externe de la membrane plasmique
B. Les protéines
Elles sont en nombre très variables (25 à 75 % de la masse membranaire) et ont des fonctions spécifiques : protéines structurales,
enzymes, protéines de transport, récepteurs...
C. Les glucides
Ils sont associés aux protéines sous forme de glycoprotéines ou aux lipides sous forme de glycolipides.
D. Propriétés des membranes biologiques
Bien qu’elles aient des fonctions et des structures différentes, les membranes biologiques ont un certain nombre de propriétés
communes : Ce sont des structures en forme de feuillets
Elles sont constituées surtout de lipides, protéines et glucides
Elles sont asymétriques c’est-à-dire que la composition des faces externe et interne sont différentes
(glycérophospholipides ; sphingolipides ; protéines) Les lipides membranaires sont amphiphiles (amphipathiques) avec une partie hydrophobe (apolaire) et une partie
hydrophile (polaire). Ils forment spontanément des bicouches lipidiques, véritables barrières s’opposant au flux des
molécules polaires
Les membranes ont une structure fluide. Cette fluidité est maintenue par la nature des acides gras (insaturés ou non, longeur de la chaîne hydrocarbonée) et le cholestérol.
LES LIPIDES ; RAPPEL BIOCHIMIQUE
Généralités
* Ce sont des molécules organiques insolubles dans l’eau mais facilement solubles dans les solvants organiques (benzène,
chloroforme, éther,…). C’est une famille hétérogène de composés de faible teneur en oxygène.
* Les lipides sont des esters d’acides gras et d’alcool : Réaction d’estérification
Acide gras Alcool Ester = "lipide"
Ils sont caractérisés par la présence dans la molécule d’au moins un acide gras ou chaîne grasse.
Sont rattachés aux lipides, en raison de leur insolubilité dans l’eau, le cholestérol, les stéroïdes, la vitamine D, qui
sont des dérivés polyisopréniques.
Ils représentent environ 20 % du poids du corps.
Ils sont une réserve énergétique mobilisable : 1g lipides → 9 Kcal Ils ont un rôle de précurseurs : stéroïdes, vitamines, prostaglandines.
www.neur-one.fr
[email protected] 12 12/01/2013
Deux acides gras polyinsaturés sont des facteurs nutritionnels essentiels car ils ne sont pas synthétisés par
l’organisme et doivent lui être apportés par l’alimentation. Ce sont des acides gras indispensables : acide linoléique et acide linolénique.
Les membranes ont une structure lipidique.
Les lipides sont synthétisés dans le réticulum endoplasmique lisse et c'est là que l'asymétrie des couches est engendrée par des
protéines d'échange des phospholipides (lipoprotéines intracellulaires). L'asymétrie lipidique est importante sur le plan fonctionnel surtout dans
la localisation des protéines liées à la membrane et intervenant dans la transduction du signal. 1001
La bio synthèse des lipides s’effectue à partir de :
Classification des lipides Synthétisés par les organismes vivants Peu ou pas solubles dans l’eau Solubles dans les solvants organiques non polaires Contiennent un ou plusieurs acides gras ou dérivés
d’acides gras Constitués de longues chaînes hydrocarbonées.

www.neur-one.fr
[email protected] 13 12/01/2013
Les acides gras
Les acides gras
Ils sont monoacides, linéaires, à nombre pair de carbone, soit saturés, soit insaturés.
Les acides gras sont les principaux constituants des lipides. La formule générale des acides gras est : R–COOH
R : chaîne latérale hydrocarbonée (C et H)
–COOH : fonction carboxylique (acide)
On classe les AG selon la nature de la chaîne R c’est-à-dire selon :
- sa longueur
- présence ou absence des doubles liaisons - les positions des doubles liaisons

www.neur-one.fr
[email protected] 14 12/01/2013
Les acides gras monoinsaturés
Dans les acides gras insaturés, la position de la première double liaison peut s’exprimer :
soit en partant du carboxyle (1er carbone) ; le symbole est ∆
soit en partant du méthyl (dernier carbone) ; le symbole est oméga ω. En médecine clinique et en biologie, la
désignation des acides gras insaturés la plus courante est celle qui fait appel au symbole oméga (ω).
L’acide oléique C18 : 1 ω9
L’acide oléique possède 18C, une double liaison en oméga 9 (ω9) ce qui s’écrit C18 :1 ω9.

www.neur-one.fr
[email protected] 15 12/01/2013
C’est un acide gras très abondant dans les graisses végétales et animales.
La présence d’une double liaison dans un acide gras entraîne une isomèrie cis-trans. Les acides gras naturels sont cis :
Les acides gras polyinsaturés
1. Famille linoléique (ω6)
Acide linoléique C18 : 2 ω6
L’acide linoléique est un acide gras indispensable (besoins quotidiens : 3-4 g). C’est un acide gras en C18 avec 2 doubles liaisons (ω6,
9)
Il conduit par voie enzymatique à l’acide arachidonique dans l’organisme.
Acide arachidonique C20 : 4 ω6
Il possède 4 doubles liaisons en ω6, 9, 12, 15
L’acide linoléique donne naissance dans l’organisme à l’acide arachidonique à 20 C et 4 doubles liaisons. En
l’absence d’acide linoléique dans l’alimentation, l’acide arachido-nique devient indispensable.
2. Famille linolénique (ω3)
Acide α linolénique C18 : 3 ω3 Il possède 3 doubles liaisons en ω3, 6, 9
Propriétés des acides gras
A. Propriétés physiques : 1. Solubilité
L’acide butyrique à 4C est soluble dans l’eau, puis la solubilité des acides gras baisse progressivement et ils sont insolubles à
partir de 10C.
Ils sont solubles dans les solvants organiques apolaires : benzène, chloroforme, …
2. Le point de fusion
augmente avec le nombre de C.
diminue quand le nombre de doubles liaisons augmente. Ils sont liquides à 20° C si n <10 C solides si n = 10 C
B. Propriétés chimiques :
1. Oxydation des doubles liaisons
L’oxydation par l’oxygène de l’air conduit au rancissement des graisses
L’oxydation enzymatique intracellulaire de l’acide arachidonique par la cyclooxygénase (cyclisation + oxydation) conduit
aux prostaglandines qui sont des médiateurs très actifs, très rapidement dégradés.
Action biologique des prostaglandines. Elles interviennent
dans la contraction des muscles lisses (intestin, utérus, vaisseaux) ;
dans la régulation des métabolismes ;
dans l’agrégation plaquettaire. L’inhibition de la cyclooxygénase des plaquettes par l’aspirine est utile en
thérapeutique (antiagrégant plaquettaire).
2 Formation de sels de sodium ou potassium
www.neur-one.fr
[email protected] 16 12/01/2013
Ce sont des savons à propriétés moussantes, mouillantes et émulsionnantes. Dans les savons se dissocient en Na +R-COO L’anion a 2
pôles
Ces molécules appelées amphiphiles ou amphipathiques, sont tensioactives : elles abaissent la tension superficielle de l’eau d’où leurs
propriétés.
3 Formation d’ester (avec Glycérol et Cholestérol) et de thioester (avec le Coenzyme A)
Les lipides
CLASSIFICATION DES LIPIDES
On distingue :
Les lipides simples : Glycérides et Stérides
Les lipides complexes : Glycérophospholipides et Sphingolipides
LES LIPIDES SIMPLES : GLYCERIDES ET STERIDES
Ce sont des lipides simples, composés ternaires constitués de C, H, O
Ce sont des esters d’acides gras + Alcool
3 types d’alcool sont estérifiés par des acides gras :
— Glycérol → Glycérides
— Cholestérol → Stérides
— Alcool à PM élevé → Cérides (non étudiés ici).

www.neur-one.fr
[email protected] 17 12/01/2013
1 LES GLYCERIDES
Ce sont des esters d’Acides Gras et de Glycérol
Si les 3 AG sont identiques, le triglycéride est homogène ; s’ils sont différents, il est hétérogène.
Ce sont les lipides naturels les plus nombreux, présents dans le tissu adipeux (graisses de réserve) et dans de
nombreuses huiles végétales. Ils représentent une réserve énergétique importante chez l’homme. Ils sont solubles dans l’acétone ce qui les différencie des phospholipides (ils sont très apolaires).
Hydrolyse des triglycérides
2 LES STERIDES
Ce sont des esters du cholestérol. Le cholestérol est une structure composée de 3 cycles hexagonaux + un cycle pentagonal correspondant au cyclopentanoperhydrophénanthène.
Il possède une fonction alcool secondaire en C3 et une double liaison en ∆5.
Le stéride est formé par estérification d’un AG sur la fonction alcool en 3 du cholestérol. Le cholestérol est apporté dans l’alimentation et synthétisé par le foie ; il est transporté dans le sang dans les
lipoprotéines.
C’est un constituant des membranes (rôle dans la fluidité).
Le cholestérol sert dans l’organisme à la synthèse de 3 groupes de molécules :
Les hormones stéroïdes (cortisol, testostérone...)
La vitamine D3
Les acides biliaires
3 LA VITAMINE D3 OU CHOLECALCIFEROL
Formule de la Vitamine D3
Elle est synthétisée à partir d’un précurseur le 7-déhydrocholestérol, présent dans la peau, qui se transforme en vitamine D 3 (qui est une prohormone), sous l’effet des UV.
Elle est métabolisée dans le foie où une 25-hydroxylase la transforme en 25-OH-vitamine D 3
puis cette dernière est hydroxylée dans le rein par une 1-hydroxylase pour donner la 1,25-di- hydroxyvitamine D 3
ou calcitriol qui est une hormone. Le calcitriol est responsable de toutes les propriétés de la vitamine D 3. La vitamine D 3 est une vitamine liposoluble qui prévient le rachitisme en favorisant la fixa- tion du calcium sur
l’os.
GLYCEROPHOSPHOLIPIDES
1 L’ACIDE PHOSPHATIDIQUE
C’est l’élément de base des glycérophospholipides. Acide phosphatidique = Glycérol + 2 Acides Gras + H3PO4

www.neur-one.fr
[email protected] 18 12/01/2013
Les deux acides gras ont une chaîne longue (≥ 14C), l’acide gras en position 2 est souvent insaturé.
L’acidité de la molécule provient des 2 H mobiles libres de l’acide phosphorique.
Au pH sanguin (7,35 - 7,45) les 2 fonctions acides sont ionisées.
L’acide phosphatidique est un second messager intracellulaire.
2 LES GLYCEROPHOSPHOLIPIDES
Ils sont constitués d’acide phosphatidique + alcool
A ; Nature de l’alcool
B. Les différentes classes de glycérophospholipides
Le lipide se forme par fixation d’un alcool sur l’acide phosphatidique. Selon l’alcool, on obtient des classes différentes de lipides. Phosphatidylsérines = Acides Phosphatidiques + Sérine
Phosphatidyléthanolamines = Acides Phosphatidiques + Ethanolamine
Phosphatidylcholines = Acides Phosphatidiques + Choline
Phosphatidylinositols = Acides Phosphatidiques + Inositol
Les glycérophospholipides
Présents chez les animaux, les plantes et les microorganismes. Acide phosphorique estérifié en sn-3 du glycérol ± aminoalcool ou sucre alcool. Grand nombre de molécules différentes
3 LES PHOSPHATIDYLETHANOLAMINES ET PHOSPHATIDYLSERINES

www.neur-one.fr
[email protected] 19 12/01/2013
4 LES PHOSPHATIDYLCHOLINES OU LECITHINES
5 LES PHOSPHATIDYLINOSITOLS
Structure de l’inositol
L’inositol est un hexaalcool cyclique qui a 9 isomères possibles. Le myoinositol est le plus fréquent dans les lipides.
6 PROPRIETES DES GLYCEROPHOSPHOLIPIDES
Ce sont des molécules amphipathiques (ou amphiphiles) car elles présentent 2 pôles :
l’un hydrophobe dû aux AG ; l’autre hydrophile dû à l’ester phosphorique.
Elles ont donc des propriétés identiques à celles des savons (émulsionnants, ...).
Ce sont des molécules amphotères car elles possèdent à la fois :
une fonction acide apportée par H3PO4 une fonction basique apportée par l’AA alcool (sérine, thréonine) ou par la choline.
SPHINGOLIPIDES
Ce sont des amides de la sphingosine qui se forment par liaison du carboxyle de l’AG sur le -NH2 de la sphingosine :
Les sphingolipides
Les sphingomyélines
Les plus abondants des sphingolipides membranaires. Absents dans les végétaux et les microorganismes. Feuillet externe de la membrane plasmique. 10% des lipides du cerveau.

www.neur-one.fr
[email protected] 20 12/01/2013
LES SPHINGOMYELINES
Elles sont constituées de l’association Sphingosine + AG + Phosphorylcholine
L’acide gras le plus fréquent est l’acide lignocérique (C24:O). Au pH du sang, la molécule est ionisée.
On les trouve dans le tissu nerveux (graines de myéline) et dans les membranes.
La déficience en sphingomyélinase entraîne leur accumulation dans le cerveau, la rate et le foie.
LES GLYCOLIPIDES
A. Cérébrogalactosides ou Galactosylcéramides
Ils sont constitués de : Sphingosine + AG + βD Galactose
B. Les Sphingomyélines
Elles sont constituées de l’association : Sphingosine + AG + Phosphorylcholine
L’acide gras le plus fréquent est l’acide lignocérique (C24:O).
Au pH du sang, la molécule est ionisée.
On les trouve dans le tissu nerveux (graines de myéline) et dans les membranes.
La déficience en sphingomyélinase entraîne leur accumulation dans le cerveau, la rate et le foie.
Les lipides membranaires
Les lipides de la membrane cellulaire sont tous des amphiphiles possédant des groupements aliphatiques ou aromatiques (comme le
cholestérol) et des groupements polaires variés. La géométrie de chaque chaîne aliphatique dépend de l'existence ou non d'insaturation. Les
chaînes d'acides gras non saturées sont très flexibles et possèdent un très grand nombre de conformations possibles, chaque liaison possédant 3 degrés de liberté de rotation. La conformation la plus probable est celle de l'extension maximale de la chaîne, conduisant à une énergie minimale.
Une chaîne insaturée possède au moins une double liaison. La configuration cis, largement plus probable que la configuration trans, conduit à un
coude dans la chaîne carbonée de l'ordre de 30°.
La tête polaire est attachée aux deux chaînes carbonées par l'intermédiaire d'une molécule servant de lien entre les trois groupements.
Ce lien, qui joue le rôle de squelette pour l'ensemble de la molécule, est généralement réalisé par un alcool, comme le glycérol, ou une
sphingosine. Les groupements polaires sont le plus souvent basés sur un groupe phospho ou glyco, d'autres molécules venant s'y greffer.
Les lipides sont classés en fonction de la nature de leur molécule de liaison et de celle de leurs chaînes carbonées : on distingue donc les glycérophospholipides, les glycéroglycolipides, les sphingophospholipides et les sphingoglycolipides.
On distingue trois catégories principales de lipides membranaires : les phospholipides, les glycolipides et les stérols ; ces
derniers ne répondent pas exactement à la définition classique des lipides mais sont des molécules apparentées au plan physicochimique.
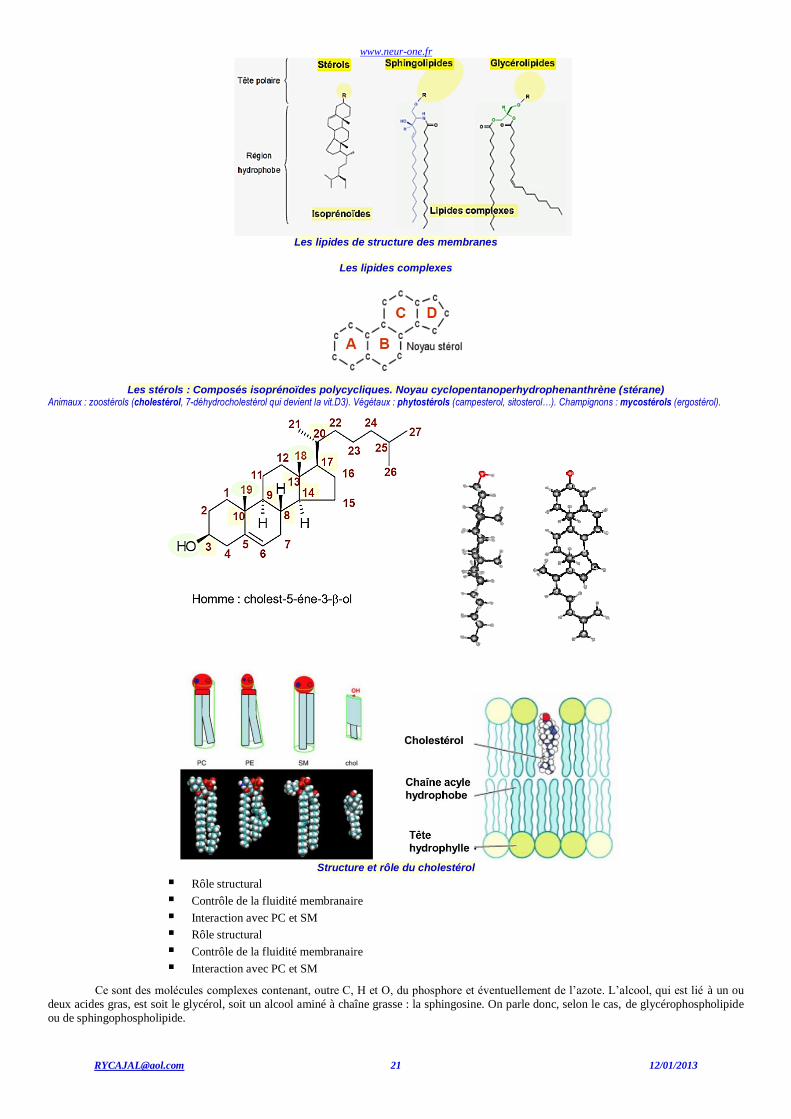
www.neur-one.fr
[email protected] 21 12/01/2013
Les lipides de structure des membranes
Les lipides complexes
Les stérols : Composés isoprénoïdes polycycliques. Noyau cyclopentanoperhydrophenanthrène (stérane)
Animaux : zoostérols (cholestérol, 7-déhydrocholestérol qui devient la vit.D3). Végétaux : phytostérols (campesterol, sitosterol…). Champignons : mycostérols (ergostérol).
Structure et rôle du cholestérol
Rôle structural Contrôle de la fluidité membranaire Interaction avec PC et SM Rôle structural Contrôle de la fluidité membranaire Interaction avec PC et SM
Ce sont des molécules complexes contenant, outre C, H et O, du phosphore et éventuellement de l’azote. L’alcool, qui est lié à un ou
deux acides gras, est soit le glycérol, soit un alcool aminé à chaîne grasse : la sphingosine. On parle donc, selon le cas, de glycérophospholipide
ou de sphingophospholipide.

www.neur-one.fr
[email protected] 22 12/01/2013
LES PHOSPHOLIPIDES
La structure de base des premiers est la suivante: – une molécule de glycérol : CH2 OH-CHOH-CH2 OH ;
– deux molécules d’acide gras, saturés : (CH3 -(CH2 )n-COOH) ou insaturés ;
– une molécule d’acide phosphorique : H 3 PO4 ; – une molécule supplémentaire liée à l’acide phosphorique.
C’est cette dernière molécule qui confère son identité au lipide en question. Les trois fonctions alcool du glycérol sont estérifiées par
deux acides gras et l’acide phosphorique, constituant ainsi l’acide phosphatidique. La molécule supplémentaire peut être un alcool (le
glycérol, l’inositol, la choline, l’éthanolamine) ou un acide aminé (la sérine) ; ces trois derniers composés contiennent de l’azote. La molécule
finale porte, suivant les cas, le nom de phosphatidyl-glycérol, phosphatidyl-inositol… La phosphatidylcholine, trouvée en abondance dans le jaune d’œuf, est la plus connue et porte aussi le nom de lécithine ; le diphosphatidyl-glycérol (ou cardiolipide), caractéristique de la
membrane interne des mitochondries et de la membrane plasmique des Procaryotes est aussi une de ces molécules. On notera que la molécule
supplémentaire apporte, ou pas, selon sa nature, une ou deux charges électriques au phospholipide (en plus de celle liée à l’acide phosphorique).
La principale caractéristique physicochimique de tous ces phospholipides est l’amphiphilie, qui sera définie plus loin.
a) Structure des phospholipides. b), c), d) Les trois catégories de phospholipides. la phosphocholine est un glycérolipide, la sphingomyéline est un
sphingolipide, et le cholestérol est un stérol. Chaque lipide est représenté sous forme compacte (chaque sphère a pour rayon le rayon de Van der Waals de l'atome) et par sa formule semi-développée. D'après le catalogue Avanti.
Les phospholipides présentent tous une tête hydrophile (phosphate et groupement spécialisé) et une queue hydrophobe (glycérol et
acides gras). On distingue deux types de phospholipides :
Les glycérophospholipides correspondent à l’association de glycérol, de deux acides gras, d’un acide phosphorique et d’alcools ou d’acides aminés. Les alcools ou les acides aminés donnent l’identité et la
caractéristique du glycérophospholipides. Parmi les acides aminés on trouve la sérine et parmi les alcools on trouve l’inositol, l’éthanolamine et la choline ; on obtient ainsi la phosphatidyl-sérine, le phosphatidyl-inositol,
la phosphatidyl-éthanolamine et la phosphatidyl-choline.

www.neur-one.fr
[email protected] 23 12/01/2013
Les glycérolipides sont bâtis sur la structure du glycérol. Deux de ses trois fonctions alcool, dont la fonction secondaire, sont
estérifiées par des acides gras à longue chaîne (principalement 16 et 18 carbones). Une des deux chaînes d'acides gras est généralement
insaturée.
Si la troisième fonction alcool est substituée par un éther de mono- ou di-galactoside, le lipide est un glycéroglycolipide. Si la troisième fonction alcool est estérifiée par un acide phosphorique, le lipide est un glycérophospholipide. L'acide phosphorique peut lui-même
être estériné par différents groupements, tels que la choline, la serine, l'éthanolamine. Quelques-uns des lipides ainsi formés sont recensés ci-
dessous.
Les sphingophospholipides correspondent à l’association de sphingosine, d’acide gras, d’acide phosphorique et d’alcool ou
d’acides aminés ; on obtient ainsi la sphingomyéline (par association de la choline).

www.neur-one.fr
[email protected] 24 12/01/2013
Les sphingolipides sont des dérivés de céramides, contenant à la fois des donneurs et des accepteurs de liaisons hydrogène.
Contrairement aux glycérolphospholipides, les deux chaînes carbonées sont souvent saturées.
Si le groupement polaire est la phosphorylcholine, le lipide obtenu est la sphingomyéline, un sphingophospholipide. Le groupe polaire
de la sphingomyéline est donc identique à celui de la phosphatidylcholine.
Parmi les sphingolipides, on peut également compter les gangliosides. Ce sont des sphingoglycolipides acides dont les chaînes
oligosaccharidiques sont terminées par des résidus d'acides sialiques. La nomenclature des gangliosides comporte deux lettres et un chiffre. Par exemple, dans le ganglioside GMI, le G signifie ganglioside, le M correspond au nombre de résidus d'acide sialique dans elle l’oligosaccharide
(M=l) et le chiffre est lié au nombre de résidus d'osés dans la chaîne à l'exception des acides sialiques. GMI est ainsi un monoacide sialique
comportant 4 résidus d'oses en sus du résidu d'acide sialique.
Exemples de sphingolipides.
La structure de base des sphingolipides est un arnino-alcool à longue chaîne, la sphingosine. Un acide gras amidifie la fonction aminé, tandis que la fonction alcool secondaire porte le groupement polaire. Suivant la nature de ce groupement polaire, on peut distinguer les sphin-gophospholipides (groupement polaire phosphorylé) et les sphingoglycolipides (groupement polaire glycosylé). a) structure des sphingophospholipides et exemple de la sphingomyéline. b) structure des sphingoglycolipides et exemple du ganglioside GMI •
La composition de base des sphingophospholipides est très différente de celle des molécules précédentes mais la structure finale
obtenue s’en rapproche étonnamment ; elle comprend :
– une molécule de sphingosine (alcool aminé à longue chaîne grasse) ;
– une molécule d’acide gras ;
– une molécule d’acide phosphorique ;
– une molécule supplémentaire (de même nature que celles vues plus haut et liée de la même façon à l’acide
phosphorique).
Formules de quelques glycérophospholipides (diacylphosphoglycérides)
L’acide phosphatidique constitue la partie commune à toutes ces molécules, tandis que la molécule supplémentaire permet de les distinguer les unes des autres. Les chaînes d’acides gras sont schématisées sous la forme d’un zig-zag.
www.neur-one.fr
[email protected] 25 12/01/2013
La liaison amide de l’acide gras sur la fonction NH2 de la sphingosine donne la céramide (voir la figure ci-dessous).
L’encombrement et les propriétés physicochimiques de ces molécules sont identiques à ceux des glycérophospholipides vus plus haut. La
sphingomyéline, qui contient de la choline, est un représentant typique de cette catégorie ; on la trouve en abondance dans le système
nerveux.
Formules de la sphingosine, de la céramide et d’un sphingophospholipide
Si X est constitué de choline, on obtient la sphingomyéline.
La cohésion des phospholipides membranaires est due aux Liaison de Van Der Waals entre les acides gras : La forces de Van der
Waals s'exercent lorsque 2 atomes adjacents sont suffisamment proches l'un de l'autre : Contact partiel entre les nuages d'électrons qui entourent
les atomes. Les interactions de Van Der Waals sont dues à des interactions entre des moments polaires instantanés
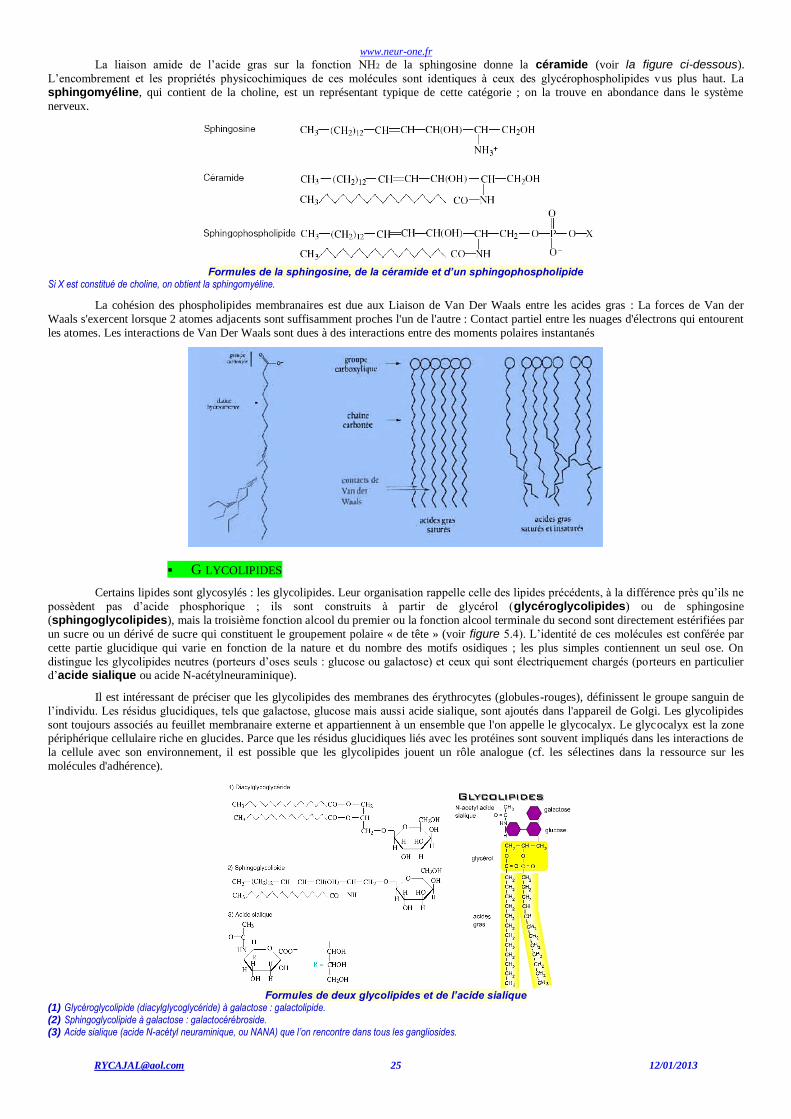
G LYCOLIPIDES
Certains lipides sont glycosylés : les glycolipides. Leur organisation rappelle celle des lipides précédents, à la différence près qu’ils ne
possèdent pas d’acide phosphorique ; ils sont construits à partir de glycérol (glycéroglycolipides) ou de sphingosine
(sphingoglycolipides), mais la troisième fonction alcool du premier ou la fonction alcool terminale du second sont directement estérifiées par
un sucre ou un dérivé de sucre qui constituent le groupement polaire « de tête » (voir figure 5.4). L’identité de ces molécules est conférée par
cette partie glucidique qui varie en fonction de la nature et du nombre des motifs osidiques ; les plus simples contiennent un seul ose. On
distingue les glycolipides neutres (porteurs d’oses seuls : glucose ou galactose) et ceux qui sont électriquement chargés (porteurs en particulier
d’acide sialique ou acide N-acétylneuraminique).
Il est intéressant de préciser que les glycolipides des membranes des érythrocytes (globules-rouges), définissent le groupe sanguin de
l’individu. Les résidus glucidiques, tels que galactose, glucose mais aussi acide sialique, sont ajoutés dans l'appareil de Golgi. Les glycolipides
sont toujours associés au feuillet membranaire externe et appartiennent à un ensemble que l'on appelle le glycocalyx. Le glycocalyx est la zone périphérique cellulaire riche en glucides. Parce que les résidus glucidiques liés avec les protéines sont souvent impliqués dans les interactions de
la cellule avec son environnement, il est possible que les glycolipides jouent un rôle analogue (cf. les sélectines dans la ressource sur les
molécules d'adhérence).
Formules de deux glycolipides et de l’acide sialique
(1) Glycéroglycolipide (diacylglycoglycéride) à galactose : galactolipide. (2) Sphingoglycolipide à galactose : galactocérébroside. (3) Acide sialique (acide N-acétyl neuraminique, ou NANA) que l’on rencontre dans tous les gangliosides.

www.neur-one.fr
[email protected] 26 12/01/2013
Chez les Bactéries ou les Végétaux, on ne trouve que des glycolipides neutres à glycérol, tandis que ceux à sphingosine sont très
abondants chez les Animaux, en particulier dans les cellules nerveuses. Parmi ceux-ci, on doit citer le galactocérébroside (neutre), contenant
du galactose seul et trouvé presque exclusivement dans la gaine de myéline entourant les axones des Vertébrés (cette gaine est formée par des couches concentriques de membrane plasmique de cellules spécialisées entrant en contact avec l’axone), et les gangliosides (chargés), qui
portent des chaînes plus ou moins ramifiées riches en acide sialique ; ces derniers sont abondants dans la membrane plasmique des cellules du
système nerveux, où l’on en a identifié plusieurs dizaines d’espèces différentes.
CHOLESTÉROL
Ce composé est typique des cellules animales ; totalement absent des membranes bactériennes et des membranes végétales, il est
cependant remplacé chez ces dernières par des stérols de nature différente. Sa formule montre un volumineux noyau hydrophobe constitué de
quatre cycles (hydrocarbure tétracyclique) portant une chaîne carbonée latérale ramifiée de huit carbones, elle-même hydrophobe. À l’opposé de cette chaîne, on trouve un groupement alcool (le seul motif hydrophile de la molécule). L’ensemble forme une sorte d’ellipsoïde aplati et rigide
présentant un caractère bipolaire amphiphile, typique des lipides (voir la figure). L’analyse de sa formule montre que le cholestérol n’est pas un
lipide « normal » (un ester) ; on le classe cependant parmi les lipides membranaires car, pour des raisons physicochimiques, il joue le même rôle
qu’eux au sein des membranes plasmique ou internes, où il est parfois très abondant.
Le cholestérol est composé d’un noyau stéroïde hydrophobe, d’une queue hydrophobe et d’une fonction alcool hydrophile. La molécule est donc amphiphile, représente environ un quart des lipides membranaires et influence la fluidité membranaire. Le cholestérol est un
lipide de structure distincte. Il joue un rôle particulier au sein de la membrane, en la rendant moins déformable (plus rigide) et en diminuant sa
perméabilité aux petites molécules hydrosolubles.
Les stérols Les stérols sont des molécules composées de 3 cycles à 6 carbones et 1 cycle à 5 carbones. Ces quatre cycles forment une structure plane et rigide, sur laquelle différents groupements peuvent être attachés. Les molécules de stérol ne peuvent former une bicouche à
elles seuls, mais peuvent s'insérer dans la membrane avec la même orientation que les phospholipides. Le cholestérol interagit pré-
férentiellement avec les sphingolipides plutôt qu'avec les phospholipides insaturés
Formule du cholestérol
Noter l’importance considérable de la partie hydrophobe de cette molécule (noyau stéroïde et queue hydrocarbonée ramifiée), en comparaison de la seule fonction polaire (-OH) qu’elle porte à l’opposé de la chaîne hydrocarbonée.
La figure ci-dessous montre l'insertion des molécules de cholestérol entre les chaînes des phospholipides.
a) Structure du cholestérol, b) Insertion dans la membrane.

www.neur-one.fr
[email protected] 27 12/01/2013
Variété de la composition lipidique des membranes
On observe de grandes variations dans la composition des membranes de différentes cellules ou des membranes de différentes
organelles d'une même cellule. La membrane d'une mitochondrie a notamment une composition lipidique assez spécifique.
La composition respective des deux feuillets de la membrane d'une cellule est également différente. Les lipides chargés sont ainsi très
présents dans le feuillet interne, tandis que les lipides PC et SM se trouvent surtout dans le feuillet externe. La perte d'asymétrie et notamment la présence de lipides PS sur le feuillet externe est un signal de mort cellulaire. La composition lipidique de la membrane des globules rouges
humains est détaillée dans la figure ci-dessous.
Composition lipidique de la membrane du globule rouge humain, a) Proportions relatives des différents lipides de la membrane du globule rouge humain. D'après [Tanford 80]. b) Composition lipidique des deux feuillets de la membrane du globule rouge humain.
Principaux glycérophospholipides présents dans la membrane.
Exemples de glycérophospholipides.
Propriétés physiques des membranes lipidiques
La plupart des lipides peuvent prendre des structures différentes selon leur état physicochimique et les conditions physiques
extérieures. La structure en bicouche lipidique est fréquemment rencontrée. Deux paramètres d'ordre peuvent alors être étudiés : la surface
moyenne occupée par molécule et la forme de la chaîne carbonée. La surface moyenne par lipide est quasiment indépendante de la longueur de la chaîne carbonée. Dans les phases ordonnées, elle est également relativement indépendante de la température : elle est de l'ordre de 0.4 à 0.6
nm2. L'épaisseur d'une bicouche dans les phases ordonnées est fonction du diamètre des têtes polaires de lipides, de l'inclinaison des chaînes
carbonées et de l'épaisseur de l'interface entre les deux monocouches.
L'inclinaison des chaînes carbonées est due à des contraintes de compactage des têtes polaires ; elle disparaît dans les phases désordonnées. La surface moyenne par lipide devient néanmoins plus grande (> 0.75 nm2), les chaînes carbonées se plaçant les unes par rapport
aux autres de façon plus désordonnée. Les chaînes étant agencées de façon moins compacte, l'épaisseur de la bicouche est réduite de 30 à 50%
par rapport aux bicouches en phase ordonnée. Cette réduction n'est qu'en partie compensée par l'hydratation supérieure des têtes polaires et par
l'augmentation des fluctuations perpendiculairement au plan de la bicouche. Les bicouches donc moins épaisses que les mêmes bicouches en phase ordonnée
Le deuxième paramètre d'ordre possible pour distinguer les différentes phases des bicouches lipidiques est l'ordre de la chaîne
carbonée. On distingue généralement les phases suivantes : la phase cristalline, la phase ordonnée (ou phase gel), la phase désordonnée (ou
phase fluide ou cristal liquide). Il est néanmoins possible d'affiner une telle distinction en ajoutant d'autres paramètres d'ordre tels que les ordres d'orientation et de rotation des chaînes carbonées et/ou des têtes polaires.





























