Embed Size (px)
Citation preview

INFOMUSAINFOMUSALa Revue Internationale sur Bananiers et Plantains
INFOMUSA est publié avec le soutien du Centre Technique de Coopération Agricole et Rurale (CTA)
CTA
Vol. 9 N° 2Décembre 2000
DANS CE NUMÉRO
Criblage d’hybridesde bananiers résistantsà Radopholus similis
Variabilité des caractéristiquesdu système racinaire dubananier selon le groupegénomique et la ploïdie
Un nouveau nématicidebiologique pour le bananierplantain micropropagé
Mécanismes prédisposant les bananiers Cavendishà la fusariose durant l’hypoxie
Paramètres chimiques du sol et incidence et gravité de lamaladie de Panama
Sévérité des cercosporiosesnoire et jaune sur le cv.« Dominico hartón » soumis à irradiation par 60Co
Evaluation de l’éliminationsélective de mains de bananessur le cv. « Valery »
L’acceptabilité des bananesexotiques par leconsommateur ougandais
Méthode de multiplication des bananiers par décorticage de la souche
Évaluation préliminaired’introductions de bananiersau Kerala
Diversité morphologique de Musa balbisiana Colla aux Philippines
Nouvelles des Musa
Thèses
Livres etc.
Annonces
Nouvelles de l’INIBAP
Nouvelles de PROMUSA
FR
UITFUL N
ET
WORKING
�
• FIFTE
EN YEARS
O
F •
�
1985
inibap
2000

Vol. 9, N° 2Photo de couverture : Les bananes sont un aliment de base essentiel en Ouganda(Jean-Vincent Escalant, INIBAP).
Editeur : Réseau international pour l’améliorationde la banane et de la banane plantain (INIBAP)Rédacteur en chef :Claudine PicqComité de Rédaction :Emile Frison, Jean-Vincent Escalant,Suzanne Sharrock, Charlotte LustyImprimé en FranceISSN 1023-0068Rédaction :INFOMUSA, INIBAP, Parc Scientifique Agropolis II,34397 Montpellier Cedex 5, France. Téléphone : + 33-(0)4 67 61 13 02 ; Télécopie : + 33-(0)4 67 61 03 34 ; Courrier électronique : [email protected]’abonnement est gratuit pour les pays endéveloppement. Les lecteurs sont invités àenvoyer lettres et articles. La rédaction seréserve le droit d’abréger ou de reformulerles textes publiés pour des raisons de clartéet de concision. INFOMUSA ne peut s’enga-ger à répondre à toutes les lettres reçues,mais s’efforcera de le faire dans un délairaisonnable. La reproduction de tout extraitdu magazine est autorisée, à condition d’enspécifier l’origine. INFOMUSA est également publié en an-glais et en espagnol.
Changement d’adresse :Merci d’en informer la rédactiond’INFOMUSA à l’adresse indiquée ci-dessus,avec si possible six semaines de préavis,afin d’éviter toute interruption de réceptionde la revue.Les opinions émises dans les articles n’en-gagent que leurs auteurs et ne reflètentpas nécessairement le point de vue del’INIBAP.
INFOMUSA Vol. 9, N° 2
SOMMAIRE
Criblage d’hybrides de bananiers résistants à Radopholus similis. . . . . . . . . . . . 3
Évaluation de la variabilité des caractéristiques du système racinaire du bananier (Musa spp.) selon le groupe génomique et le degré de ploïdie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
Emploi d’un nouveau nématicide biologique pour la protection racinaire du bananier plantain (Musa AAB) multiplié par micropropagation . . . . . . . 8
Proposition d’explication des mécanismes prédisposant les bananiers Cavendish à la fusariose durant l’hypoxie. . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
Paramètres chimiques du sol en relation avec l’incidence et la gravité de la maladie de Panama . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
Sévérité de la cercosporiose noire (Mycosphaerella fijiensis Morelet) et de lacercosporiose jaune (Mycosphaerella musicola Leach) sur Musa AAB cv. « Dominico hartón » soumis à irradiation par 60Co . . . . . . . . . . . . . . . . . . . 16
Considérations méthodologiques pour l’évaluation de l’élimination sélective de mains de bananes (Musa AAA, cv. « Valery ») . . . . . . . . . . . . . . . . . . . . . 19
L’acceptabilité des bananes exotiques par le consommateur ougandais . . . . . 22
Méthode de multiplication des bananiers par décorticage de la souche. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
Évaluation préliminaire d’introductions de bananiers au Kerala . . . . . . . . . . . 27
Diversité morphologique de Musa balbisiana Colla aux Philippines. . . . . . . . . 28
Nouvelles des Musa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
Thèses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
Annonces. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
Nouvelles de l’INIBAP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
Livres etc . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
Nouvelles de PROMUSA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I à XVI
La mission de l’INIBAP est d’accroître de façon durable la productivité des bananiers et desbananiers plantain cultivés sur de petites exploitations pour la consommation locale et pourles marchés d’exportation.Le programme de l’INIBAP a quatre objectifs principaux :
• organiser et coordonner un effort global de recherche sur la banane et la banane plantain vi-sant au développement, à l’évaluation et à la dissémination de matériel génétique de Musaamélioré ainsi qu’à la conservation et à l’utilisation de la diversité génétique des Musa ;
• promouvoir et renforcer la collaboration et le partenariat en matière de recherche sur lesbananiers au niveau national, régional et international ;
• renforcer la capacité des Systèmes nationaux de recherche agricole à conduire des re-cherches sur la banane et la banane plantain ;
• coordonner, faciliter et appuyer la production, la collecte et l’échange d’information et dedocumentation sur la banane et la banane plantain.
L’INIBAP est un programme de l’Institut international pour les ressources phytogénétiques(IPGRI), un centre “Future Harvest”.
INFOMUSAINFOMUSALa Revue Internationale sur Bananiers et Plantains
INFOMUSA est publié avec le soutien du Centre Technique de Coopération Agricole et Rurale (CTA)
CTA
Vol. 9 N° 2Décembre 2000
DANS CE NUMÉRO
Criblage d’hybridesde bananiers résistantsà Radopholus similis
Variabilité des caractéristiquesdu système racinaire dubananier selon le groupegénomique et la ploïdie
Un nouveau nématicidebiologique pour le bananierplantain micropropagé
Mécanismes prédisposant les bananiers Cavendishà la fusariose durant l’hypoxie
Paramètres chimiques du sol et incidence et gravité de lamaladie de Panama
Sévérité des cercosporiosesnoire et jaune sur le cv.« Dominico hartón » soumis à irradiation par 60Co
Evaluation de l’éliminationsélective de mains de bananessur le cv. « Valery »
L’acceptabilité des bananesexotiques par leconsommateur ougandais
Méthode de multiplication des bananiers par décorticage de la souche
Évaluation préliminaired’introductions de bananiersau Kerala
Diversité morphologique de Musa balbisiana Colla aux Philippines
Nouvelles des Musa
Thèses
Livres etc.
Annonces
Nouvelles de l’INIBAP
Nouvelles de PROMUSA
FR
UITFUL N
ET
WORKING
�
• FIFTE
EN YEARS
O
F •
�
1985
inibap
2000

Carine Dochez, Paul R. Speijer†, John Hartman†, Dirk Vuylsteke†
et Dirk De Waele
Les nématodes parasites des plantessont une contrainte majeure à uneproduction durable du genre Musa
(Stover et Simmonds 1987). Ils ont étéidentifiés comme l’une des causes ma-jeures de diminution de la production ba-nanière en Ouganda (Speijer et al. 1999),le plus grand producteur mondial de ba-nanes d’altitude d’Afrique de l’Est (Musaspp., groupe AAA) (Lescot 1998). Le néma-tode le plus dévastateur des bananiers sousles tropiques est Radopholus similis(Cobb) Thorne (Gowen 1993). Il a donc étéutilisé comme espèce-test dans cette pro-cédure d’évaluation.
Les nématodes peuvent être combattusavec des produits chimiques, mais ceux-cipeuvent avoir des effets négatifs sur l’envi-ronnement ; de plus, leur utilisation esttrop onéreuse et les produits trop dange-reux pour les petits agriculteurs. La sélec-tion de plantes-hôtes résistantes repré-sente une alternative prometteuse dans lalutte contre les nématodes (Speijer et DeWaele 1997). Cependant, le criblage denouveaux hybrides au champ est coûteuxen termes de temps et d’espace. C’estpourquoi une méthode d’évaluation pré-coce et rapide du matériel génétique debananier résistant aux nématodes, baséesur l’inoculation des racines individuelles(De Schutter et al., en cours de prépara-tion), a été utilisée dans cette étude.
Matériels et méthodesDes essais en serre ont été conduits dansla zone centrale de l’Ouganda, au Centrerégional pour l’Afrique orientale et aus-trale de l’Institut international d’agricul-ture tropicale (IITA-ESARC), ferme Sen-dusu, Namulonge. La station est située à1 150 m d’altitude et est représentative dela zone de culture des bananiers d’altituded’Afrique de l’Est.
Les cultivars utilisés comme témoinssont Yangambi km5 (Musa AAA, très résis-tant à Radopholus similis), Gros Michel(Musa AAA, partiellement résistant àR. similis) et Valery (Musa AAA, sensibleà R. similis).
Les hybrides choisis par le programmede sélection de l’IITA-ESARC pour le testcomprennent les hybrides diploïdes déri-vés du bananier plantain TMP2x 2521S-31,TMP2x-47 et TMP2x-50 ; les hybrides di-
ploïdes dérivés du bananier TMB2x 1411S-2, TMB2x 1411S-10, TMB2x 2559S-1et TMB2x 2559S-2 ; l’hybride tétraploïdeTMBx 2094S-1 dérivé de ‘Pisang Awak’ ; etl’hybride tétraploïde TMHx 660K-1 dérivédu bananier d’altitude d’Afrique de l’Est.
L’inoculum de nématodes provenait de cultures sur des disques de carottes (Daucus carota L.) (Pinochet et al. 1995).Celles-ci avaient préalablement été stérili-sées en surface par une pulvérisation àl’éthanol à 96 % suivie d’une exposition à laflamme, puis épluchées, coupées endisques de 3 mm d’épaisseur et placéesdans des boîtes de Pétri de 35 mm de dia-mètre. Les nématodes ont été stérilisés ensurface avec du sulfate de streptomycineaqueux (2 000 ppm) pendant 6 heures etrincés 3 fois à l’eau distillée stérile. Envi-ron 100 nématodes, contenus dans 10 µld’eau, ont été placés sur chaque disque decarotte. Les boîtes de Pétri ont été hermé-tiquement scellées par un film de paraffineet incubées à 28 °C dans l’obscurité. Lesnématodes ont ensuite été repiqués surdes carottes fraîches toutes les 5 à 7 se-maines. L’inoculum a été préparé en rin-çant à l’eau distillée stérile les boîtes dePétri contenant les disques de carottes eten recueillant les nématodes dans uneéprouvette.
Tous les génotypes de bananiers ont étéplantés dans des boîtes en bois contenantde la sciure de bois stérilisée à la vapeur.Chaque boîte contenait 9 rejets, pelés ettraités à l’eau chaude (Colbran 1967).Quatre semaines après la plantation, troisracines ont été sélectionnées sur chaquerejet. Chaque racine sélectionnée a été soi-
gneusement déterrée et entourée d’unpetit pot en plastique à 5 cm du rhizome.L’inoculation a été effectuée en versantune suspension de 50 femelles de R. similissur chaque racine. La racine et le pot en plastique ont ensuite été couverts desable stérilisé à la vapeur. Les racines ino-culées ont été récoltées huit semainesaprès l’inoculation.
Les racines récoltées ont alors été lavéeset mises à macérer dans un mixeur pen-dant 10 secondes. Les nématodes ont étéextraits le lendemain selon une variante dela technique de l’entonnoir de Baermann(Hooper 1990) puis recueillies sur un tamisde 20 µm. Un décompte des mâles et desfemelles des différents stades de dévelop-pement vermiformes a été effectué. Letaux de reproduction de R. similis (popu-lation finale/population initiale) a été cal-culé pour chaque cultivar et les contrastesorthogonaux (SAS 1997) des matériels tes-tés par rapport aux cultivars de référenceYangambi km5 et Valery déterminés afinde comparer les taux de reproductionmoyens.
Résultats et discussionLe tableau 1 montre le taux de reproduc-tion de R. similis chez les différents culti-vars, tandis que le tableau 2 indique lescontrastes orthogonaux entre les cultivarstestés et les témoins résistants et sen-sibles. Pour tous les cultivars, le taux de re-production des nématodes est plus faibleque celui observé chez Valery (tableaux 1et 2). Chez les génotypes Gros Michel,TMP2 2521S-31 et 47, TMB2 1411S-10, TMB 2094S-1, TMH 660K-1 et TMB2 2569S-2,
INFOMUSA — Vol 9, N° 2 3
Criblage d’hybrides de bananiers résistants à Radopholus similis
Ressources génétiques Tests précoces de sélection
Tableau 1. Taux de reproduction de Radopholus similis sur des racines de 12 génotypes de bananiers 8 semaines après inoculation par une suspensioncontenant 50 femelles de R. similis.
Génotype Parents Pf Rr1 = Pf2/Pi3
Yangambi km5 1 0.02
Gros Michel 82 1.64
Valery 883 17.66
TMB2x 1411S-2* TMB2x 7197-2 x TMB2x 9839-1 427 8.54
TMB2x 1411S-10* TMB2x 7197-2 x TMB2x 9839-1 10 0.20
TMB2x 2569S-1* TMB2x 7197-2 x TMB2x 9128-3 2 0.04
TMB2x 2569S-2* TMB2x 7197-2 x TMB2x 9128-3 491 9.82
TMBx 2094S-1* Kayinja x TMB2x 7197-2 73 1.46
TMP2x 2521S-31* TMP2x 1518 x TMB2x 8075-3 66 1.32
TMP2x 2521S-47* TMP2x 1518 x TMB2x 8075-3 60 1.20
TMP2x 2521S-50* TMP2x 1518 x TMB2x 8075-3 0.3 0.006
TMHx 660K-1 Enzirabahima x Calcutta 4 99 1.981 Rr = Taux de reproduction (population finale/population initiale).2 Pf = Population finale, tous stades de développement vermiformes et tous sexes inclus.3 Pi = Population initiale, 50 femelles de R. similis.
* Hybrides ayant Pisang Jari Buaya dans leur pedigree.

les taux de reproduction des nématodes nesont pas significativement différents decelui observé chez Yangambi km5. Les gé-notypes dont les taux de reproduction nesont pas statistiquement différents decelui observé chez Yangambi km5 ont mon-tré de faibles densités et sont donc des gé-notypes prometteurs pour lesquels uneévaluation plus poussée est recommandée.Le génotype Pisang Jari Buaya (Musa AA),hautement résistant à R. similis (Pinochet1988), se retrouve dans le pedigree de tousles hybrides, à l’exception de TMH 660K-1.
RemerciementsLes auteurs expriment leur reconnaissanceà l’Association flamande pour la coopéra-tion pour le développement et l’assistancetechnique (VVOB) et l’Administration gé-nérale de la coopération pour le développe-ment (AGCD) pour leur soutien financier.Leurs remerciements vont également àMme Pamela Mpirirwe et Mlle ChristineKajumba pour leur assistance technique.Cet article est le manuscrit IITA numéroIITA/00/JA/29. ■
RéférencesColbran R.C. 1967. Hot water tank for treatment of
banana planting material. Queensland Depart-ment of Primary Industries, Brisbane. Advisoryleaflets Division of Plant Industry 9(24): 4.
De Schutter B., P.R. Speijer, C. Dochez, A. Ten-kouano & D. De Waele. (en préparation). Scree-ning of Musa germplasm for resistance to nema-todes by inoculating individual roots.
Gowen S.R. 1993. Yield losses caused by nematodeson different banana varieties and some manage-ment techniques appropriate for farmers inAfrica. Pp. 199-208 in Biological and integratedcontrol of highland banana and plantain pestsand diseases. Proceedings of a research coordi-nation meeting. Cotonou, Bénin, 12-14 November
1991 (C.S. Gold & B. Gemmill, eds.). Internatio-nal Institute of Tropical Agriculture, Ibadan, Nigeria.
Hooper D.J. 1990. Extraction and processing ofplant soil nematodes. Pp. 137-180 in Plant para-sitic nematodes in subtropical and tropical agri-culture (M. Luc, R.A. Sikora & J. Bridge, eds).CAB International, Wallingford, UK.
Lescot T. 1998. Banana. Little-known wealth of variety. Fruitrop 51: 8-11.
Pinochet J. 1988. Comments on the difficulty inbreeding bananas and plantains for resistance tonematodes. Revue de Nématologie 11(1): 3-5.
Pinochet J., C. Fernandez & J.L. Sarah. 1995. In-fluence of temperature on in vitro reproductionof Pratylenchus coffeae, P . goodeyi and Radopholus similis. Fundamental and AppliedNematology 18(4): 391-392.
SAS. 1997. SAS guide for personal computers. 6th ed.SAS Institute Inc., Cary, North Carolina, USA.
Speijer P.R. & D. De Waele. 1997. Screening ofMusa germplasm for resistance and tolerance tonematodes. INIBAP Technical Guidelines 1. In-
ternational Plant Genetic Resources Institute,Rome, Italy ; International Network for the Im-provement of Banana and Plantain, Montpellier,France ; ACP-EU Technical Centre for Agricultu-ral and Rural Cooperation, Wageningen, The Netherlands. 47 p.
Speijer P.R., C. Kajumba & F. Ssango. 1999. EastAfrican highland banana production as influen-ced by nematodes and crop management inUganda. International Journal of Pest Manage-ment 45: 41-49.
Stover R.H. & N.W. Simmonds. 1987. Banana. 3rd ed.Longman Scientific and Technical, London, UK.
Ce travail a été réalisé par Carine Dochez, PaulR. Speijer, John Hartman et Dirk Vuylsteke del’Institut international d’agriculture tropicale-Centrerégional pour l’Afrique orientale et australe (IITA-ESARC), BP 7878, Kampala, Ouganda, et Dirk DeWaele du Laboratoire pour l’amélioration des culturestropicales de l’Université Catholique de Leuven (KUL),Kasteelpark Arenberg 13, 3001 Leuven, Belgique.
4 INFOMUSA — Vol 9, N° 2
Tableau 2. Contrastes orthogonaux entre Yangambi km5 (résistant à R. similis),Valery (sensible à R. similis) et les autres cultivars.
Cultivars Contraste avec Yangambi km5 Contraste avec Valery
Yangambi km5 ***
Gros Michel Ns ***
Valery ***
TMB2x 1411S-2 ** **
TMB2x 1411S-10 Ns ***
TMB2x 2569S-1 ** **
TMB2x 2569S-2 Ns ***
TMBx 2094S-1 Ns ***
TMP2x 2521S-31 Ns ***
TMP2x 2521S-47 Ns ***
TMP2x 2521S-50 * ***
TMHx 660K-1 Ns ***Ns Contraste non significatif pour P > 0.05.
*** Contraste significatif pour P = 0.001.
** Contraste significatif pour P = 0.01.
* Contraste significatif pour P = 0.05.
G. Blomme, R. Swennen et A. Tenkouano
Pour améliorer génétiquement les ra-cines d’espèces végétales, il fautconnaître la variabilité intraspéci-
fique de leurs caractéristiques (O’Toole etBland 1987). Des différences génotypiquesdans la dimension des racines ont étéconstatées chez le maïs (Zea mays L.)(Pan et al. 1985, Aina et Fapohunda 1986,
Mackay et Barber 1986), l’orge (Hordeumvulgare L.) (Hackett, 1968), le blé (Triti-cum aestivum L.) (Hurd 1968), la tomate(Lycopersicon esculentum Mill.) et le ha-ricot (Phaseolus vulgaris L.) (Zobel 1975),le riz (Oryza sativa L.) (Nicou et al. 1970,Reyniers et al. 1975, Ekanayake et al. 1985a, 1985 b) et plusieurs autres espèces(O’Toole et Bland 1987).
Des recherches sur les différences géno-typiques dans les caractéristiques des ra-cines de Musa spp. en condition hydropo-
niques (Swennen 1984, Swennen et al.1986) ont abouti à la conclusion que le sys-tème racinaire est plus important chez lesbananiers dessert que chez les plantains.Dans une autre étude similaire, on a aussiobservé des différences dans l’émission ra-cinaire latérale (Draye et al. 1999).
On pense que le degré de ploïdie influesur la dimension des différentes partiesdes plants de l’espèce Musa (Simmonds1962 et 1966, Vandenhout et al. 1995), maisaucune étude systématique n’a encore été
Évaluation de la variabilité des caractéristiquesdu système racinaire du bananier (Musa spp.)selon le groupe génomique et le degré de ploïdie
Physiologie Influences de la ploïdie

faite pour déterminer les effets du degréde ploïdie et du groupe génomique sur lescaractéristiques des racines chez desplants cultivés en champ.
L’objectif de la présente étude consistaità évaluer la contribution relative du degréde ploïdie et de la composition génomiqueà la variabilité des caractéristiques des ra-cines chez l’espèce Musa.
Matériel et méthodesCette expérimentation a été effectuée à lastation de l’IITA située à Onne, en zone deforte pluviométrie dans le sud-est du Nigeria(4°42’ N, 7°10’ E, altitude 5 mètres). Le solest un ultisol dérivé de sédiments côtiers,bien drainé mais pauvre en éléments nutri-tifs, dont le pH est de 4,3 dans 1:1 H2O. Lapluviométrie annuelle moyenne s’élève à2 400 mm répartis entre février et novembreen régime monomodal. Ce site a été décritde manière détaillée par Ortiz et al. (1997).
On a évalué 18 génotypes de bananierset de plantains (Musa spp.) appartenant à
cinq groupes génomiques et ayant trois de-grés de ploïdie (tableau 1) au stade de lafloraison. Pour ce faire, on a produit des vi-troplants à l’aide des techniques standardde culture de méristèmes (Vuylsteke 1989,Vuylsteke 1998). Les plantules racinées ontété transférées dans des sacs en polyéthylène(hauteur = 25 cm, circonférence = 44 cm)et placées en serre (Vuylsteke et Talengera1998, Vuylsteke 1998), puis transplantéesen champ en juin 1996, six semaines aprèsl’acclimatation.
On a préparé manuellement le site expé-rimental, qui était sous jachère herbacéedepuis huit ans, afin d’éviter toute pertur-bation du sol. On a fertilisé les plants avecdu chlorure de potassium (m.a. K20, 60 % K)à la dose de 600 g plant-1 an-1 et avec del’urée (47 % N) à la dose de 300 g plant-1
an-1, en six fractions égales durant la sai-son des pluies. Aucun paillis n’a été appli-qué. On a traité la parcelle expérimentaleavec le nématicide Némacur (m.a. fenami-phos) à la dose de 15 g plant-1 (trois traite-
ments par an) afin de réduire les infesta-tions de nématodes. On a appliqué le fongi-cide Bayfidan (m.a. triadiménol) trois foispar an à la dose de 3,6 ml plant-1 afin deprévenir la cercosporiose noire (Mycos-phaerella fijiensis Morelet). Pendant lasaison sèche, les plants ont été irrigués à ladose de 100 mm mois-1.
On a utilisé un dispositif en blocs de Fisher avec deux répétitions consistant endeux plants par génotype. Afin d’éviter queles systèmes racinaires adjacents ne serencontrent, on a espacé les plants de 4 mx 4 m. Pour l’évaluation, on a entièrementsorti les plants de terre et mesuré les ca-ractéristiques suivantes : hauteur de plant(HP, cm), nombre de feuilles (NF), circon-férence du pseudotronc au niveau du sol(CP, cm) et hauteur du plus grand rejet(HR, cm). On a aussi calculé la surface fo-liaire (SF, cm2) selon la méthode d’Obie-funa et Ndubizu (1979). On a déterminé lepoids frais de la souche (PS, g), la hauteurde la souche (HS, cm) et sa plus grandelargeur (PGL, cm). On a compté le nombrede rejets (NR) présents sur la souche. Àl’aide d’un pied à coulisse, on a mesuré lenombre de racines adventives (NRA), lepoids sec des racines (PSR) et le diamètremoyen à la base des racines (DM, mm). Ona déterminé la longueur des racines adven-tives (LRA, cm) en se servant de la mé-thode des intersections (Newman 1966,Tennant 1975). On a mesuré le poids sectotal (PST, g) et la longueur totale (LT, cm)des racines de la touffe (pied mère et re-jets). Enfin, on a déterminé la croissanceaérienne, le développement de la souche etles caractéristiques de la croissance dusystème racinaire chez le plus grand desrejets.
L’analyse statistique a été effectuée àl’aide du logiciel SAS (SAS, 1989). On aévalué la variabilité des différentes carac-téristiques de la croissance à l’aide de lafonction PROC GLM de ce logiciel. On a ré-parti la variance phénotypique totale en
INFOMUSA — Vol 9, N° 2 5
Tableau 1. Nom, génome, degré de ploïdie, type et mode de développement desrejets des génotypes évalués dans cette étude.
Nom Génome Degré de ploïdie Type Développement des rejets
Niyarma Yik AA 2 Musa acuminata banksii Non régulé
Calcutta 4 AA 2 Musa acuminata burmannica Non régulé
Pahang AA 2 Musa acuminata malaccensis Non régulé
Pisang J. Buaya AA 2 Musa acuminata microcarpa Non régulé
Pisang Madu AA 2 Musa acuminata microcarpa Non régulé
Tjau Lagada AA 2 Musa acuminata microcarpa Non régulé
Yangambi km5 AAA 3 Bananier dessert Régulé
Valery AAA 3 Bananier dessert Régulé
Obino l’Ewai AAB 3 Plantain Inhibé
Agbagba AAB 3 Plantain Inhibé
Pelipita ABB 3 Bananier à cuire Régulé
Cardaba ABB 3 Bananier à cuire Régulé
Fougamou ABB 3 Bananier à cuire Régulé
TMPx 2796-5 AAB x AA 4 Plantain hybride (Bobby Tannap x Pisang lilin) Régulé
TMPx 7152-2 AAB x AA 4 Plantain hybride (Mbi Egome 1 x Calcutta 4) Régulé
TMPx 548-9 AAB x AA 4 Plantain hybride (Obino l’Ewai x Calcutta 4) Régulé
TMPx 5511-2 AAB x AA 4 Plantain hybride (Obino l’Ewai x Calcutta 4) Inhibé
TMPx 1658-4 AAB x AA 4 Plantain hybride (Obino l’Ewai x Pisang lilin) Régulé
Tableau 2. Résultats du test du carré moyen des écarts et du test de signification pour différentes caractéristiques quantitativesdes plants au stade de la floraison.
Caractéristiques#
Source de variation ddl SF HP PS NR HR PSRA NRA
Répétition 1 1017416013 392 2737103 1 2088 15212 201
Degré de ploïdie 2 7547029458*** 5680*** 12051416*** 41** 28298*** 93565*** 20537***
Groupe génomique 2 2608843147** 6364*** 11752654*** 65*** 7595* 45845*** 8463**
Génotype 13 2106141878*** 3575*** 8429250*** 33*** 10571*** 18524*** 2874*
Variation résiduelle 50 363318642 321 1109510 6 1526 4442 1330
LRA DM PST LT % PSRAPM % LRAPM NJJF
Répétition 1 4563021 0,04 24291 9812 12 198 1197
Degré de ploïdie 2 53138430*** 7,83** 18217 54977396 4497*** 3503*** 48142***
Groupe génomique 2 4179499 1,49** 62806 34364239 1090*** 1217*** 2297
Génotype 13 10764435* 0,38* 128050***133333288*** 404** 287* 10774***
Variation résiduelle 50 4845881 0,19 23054 38834528 134 146 1741# : ddl : degré de liberté, SF : surface foliaire (m2), HP : hauteur de plant (cm), PS : poids de la souche (g), NR : nombre de rejets, HR : hauteur du plus grand rejet (cm), PSRA : poids sec des racines(g), NRA : nombre de racines adventives, LRA : longueur des racines adventives (cm), DM : diamètre moyen à la base des racines adventives (mm), PST : poids sec total des racines de la touffe (g), LT :longueur totale des racines de la touffe (cm), % PSRAPM : pourcentage du poids sec des racines de la touffe attribuable au pied mère, % LRAPM : pourcentage de la longueur des racines de latouffe attribuable au pied mère, NJJF : nombre de jours jusqu’à la floraison.
*, **, *** significatif au seuil P < 0.05, 0.01 et 0.001 respectivement.

fonction des sources de variation suivantes :répétition, degré de ploïdie, groupe géno-mique et génotype.
Résultats et discussionOn a constaté un effet significatif du degréde ploïdie sur les différentes caractéris-tiques, excepté le poids sec total et la lon-gueur totale des racines adventives de latouffe (tableau 2). De manière générale,plus le degré de ploïdie était élevé, plus lavaleur des différentes caractéristiques dela plante tendait à augmenter. Parexemple, les tétraploïdes ont donné les va-leurs les plus élevées pour la surface fo-liaire, la hauteur de plant, le poids frais dela souche, les caractéristiques des racines,ainsi que les taux de croissance journalièrecorrespondants (tableau 3). L’effet du gé-notype s’est révélé significatif pour l’en-semble des caractéristiques évaluées, tan-dis que l’effet du groupe génomique étaitsignificatif pour toutes les caractéristiquesdes parties aériennes et pour plusieurs ca-ractéristiques des racines.
Simmonds (1962) avait déjà signalé quela taille du fruit augmentait avec le degréde ploïdie, et Vandenhout et al. (1995)avaient fait la même observation en ce quiconcerne la dimension des stomates. Appa-remment, le nombre supérieur de chromo-somes entraîne une augmentation de la
taille des cellules, qui se traduit par un ac-croissement de la dimension des organesdu plant. D’après Vakili (1967), des tétra-ploïdes de M. balbisiana obtenus à l’aidede la colchicine étaient plus grands et plusrobustes que les diploïdes, mais avaient unrythme de croissance plus lent, un pluspetit nombre de rejets et un système raci-naire moins abondant. Contrairement àces observations, on a constaté dans la pré-sente étude que les taux de croissancejournalière et la dimension du système ra-cinaire augmentaient avec le degré de ploï-die (tableau 3). Plus le degré de ploïdieétait élevé, plus le diamètre des racines ad-ventives était important (tableau 3), ce quiconfirme les observations faites par Mon-net et Charpentier (1965).
On a enregistré un accroissement du dé-veloppement des rejets avec la diminutiondu degré de ploïdie. Chez tous les bana-niers diploïdes, le comportement des rejetsn’était pas régulé (tous les rejets ayant unecroissance vigoureuse), ce qui donnait lieuà un développement plus rapide, tandisque chez les triploïdes et tétraploïdes étu-diés, il était régulé (deux ou trois rejetspoussant vigoureusement) ou inhibé(aucun rejet ne poussant vigoureusement)(tableau 1). Ainsi, le pied mère ne repré-sentait que 45 % du poids sec des racinesde la touffe chez les bananiers diploïdes,
ce qui indique que la croissance des rejetsétait vigoureuse. En revanche, chez lesplantains, les bananiers à cuire et les plan-tains hybrides tétraploïdes, plus de 60 % dusystème racinaire venaient du pied mère.
Blomme et Ortiz (1996) ont mis en évi-dence des corrélations positives significa-tives entre les caractéristiques de la crois-sance des racines et des parties aériennesdurant le stade végétatif, montrant que lesplants à croissance vigoureuse avaientaussi un système racinaire plus important.Dans la présente étude, on a établi qu’ilexistait, entre la croissance des parties aé-riennes et celle des racines, une nette rela-tion qui dépendait du groupe génomique.Par exemple, les diploïdes (génome AA) etles bananiers dessert (AAA) avaient unefaible valeur pour pratiquement toutes lescaractéristiques de la croissance des ra-cines et des parties aériennes, tandis queles plantains (AAB), les bananiers à cuire(ABB) et les plantains hybrides tétra-ploïdes (AAAB) avaient des valeurs plusélevées (tableaux 3 et 4). Les faibles va-leurs enregistrées chez les bananiers des-sert s’expliquent probablement par le faitqu’on a inclus la variété semi-naine « Va-lery » dans ce groupe.
Cette étude montre donc que la crois-sance des racines et des parties aériennesdes plants de bananiers tend à être d’au-tant plus vigoureuse que le degré de ploï-die est plus élevé. En revanche, le dévelop-pement des rejets et donc la pérennitésont d’autant plus importants que le degréde ploïdie est plus faible.
RemerciementsLes auteurs remercient l’Agence flamandede coopération au développement et d’assis-tance technique (Vlaamse Vereniging voorOntwikkelingssamenwerking en Techni-sche Bijstand, VVOB) et la Direction géné-rale de la coopération internationale (Bel-gique) pour leur aide financière. Mlle LyndaOnyeukwu a contribué à cette étude en as-surant la collecte des données. ■
RéférencesAina P.O. & H.O. Fapohunda. 1986. Plant Soil 94:
257-265.Blomme G. & R. Ortiz. 1996. Preliminary evaluation
of variability in Musa root system development.Pp. 51-52 in Biology of root formation and deve-lopment (A. Altman ed.). Plenum PublishingCompany, New York, USA.
Draye X., B. Delvaux & R. Swennen. 1999. Distribu-tion of lateral root primordia in root tips ofMusa. Annals of Botany 84: 393-400
Ekanayake I.J., D.P. Garrity, T.M. Masajo &J.C. O’Toole. 1985 b. Root pulling resistance inrice: Inheritance and association with droughtresistance. Euphytica 34: 903-913.
Ekanayake I.J., J.C. O’Toole, D.P. Garrity & T.M.Masajo. 1985 a. Inheritance of root charactersand their relations to drought resistance in rice.Crop. Sci. 25: 927-933.
6 INFOMUSA — Vol 9, N° 2
Tableau 3. Caractéristiques de la croissance de Musa spp. au stade de la floraisonselon le degré de ploïdie.
Degré de ploïdie
Caractéristiques# 2 3 4
SF 92 635 ± 6 617 78 968 ± 5 290 115 547 ± 5 366
NF 13 ± 0,4 10 ± 0,6 14 ± 0,5
HP 228 ± 8 248 ± 7 260 ± 5
CP 53 ± 2 63 ± 2 63 ± 1
PS 4 135 ± 444 5 312 ± 274 5 498 ± 344
HS 24 ± 1 22 ± 1 20 ± 1
PGL 16 ± 1 20 ± 1 21 ± 1
NR 13 ± 1 11 ± 1 10 ± 0,5
HR 161 ± 15 141 ± 12 90 ± 8
PSRA 212 ± 15 281 ± 22 343 ± 19
NRA 122 ± 9 162 ± 9 182 ± 9
LRA 5 807 ± 571 6 136 ± 365 8 707 ± 618
DM 4,53 ± 0,13 5,34 ± 0,11 5,70 ± 0,07
PST 533 ± 59 513 ± 33 475 ± 31
LT 15 236 ± 2 173 12 468 ± 1 064 12 760 ± 1 098
% PSRAPM 45 ± 3 56 ± 3 74 ± 2
% LRAPM 46 ± 3 53 ± 3 71 ± 3
NJJF 381 ± 16 348 ± 12 288 ± 6
SF/NJJF 242 ± 14 237 ± 19 406 ± 22
HP/NJJF 0,61 ± 0,02 0,73 ± 0,03 0,91 ± 0,02
PS/NJJF 11 ± 1 16 ± 1 19 ± 1
HR/NJJF 0,44 ± 0,04 0,41 ± 0,04 0,31 ± 0,03
PSRA/NJJF 0,56 ± 0,04 0,84 ± 0,08 1,21 ± 0,07
NRA/NJJF 0,32 ± 0,02 0,49 ± 0,04 0,64 ± 0,04
LRA/NJJF 15 ± 1 18 ± 1 31 ± 2
PST/NJJF 1,39 ± 0,13 1,50 ± 0,10 1,67 ± 0,12
LT/NJJF 39 ± 5 36 ± 3 45 ± 4# voir tableau 1 ; NF : nombre de feuilles, CP : circonférence du pseudotronc (cm), HS : hauteur de la souche (cm), PGL : plusgrande largeur de la souche (cm).

Hackett C. 1968. A study of the root system of Bar-ley. I. Effects of nutrition on two varieties. NewPhytol. 67: 287-299.
Hurd E.A. 1968. Growth of roots of seven varieties ofspring wheat at high and low moisture levels.Agron. J. 60: 201-205.
Mackay A.D. & S.A. Barber. 1986. Effect of nitrogen onroot grow of two corn genotypes in the field. Agron.J. 78: 699-703.
Monnet J. & J.M. Charpentier. 1965. Le diamètredes racines adventives primaires des bananiersen fonction de leur degré de polyploidie. Fruits20: 171-173.
Newman E.I. 1966. A method of estimating the totallength of root in a sample. J. appl. Ecol. 3: 139-145.
Nicou R., L. Seguy & G. Haddad. 1970. Comparaisonde l’enracinement de quatre variétés de riz plu-vial en présence ou absence de travail du sol.L’Agronomie Tropicale 25: 639-659.
Obiefuna J.C. & T.O.C. Ndubizu. 1979. Estimatingleaf area of plantain. Sci. Hortic. 11: 31-36.
Ortiz R., P.D. Austin & D. Vuylsteke. 1997. IITA highrainfall station: Twenty years of research for sus-tainable agriculture in the West African HumidForest. HortScience 32(6): 969-972.
O’Toole J.C. & W.L. Bland. 1987. Genotypic varia-tion in crop plant root systems. Adv. Agron. 41:91-145.
Pan W.L., W.A. Jackson & R.H. Moll. 1985. J. Exp.Bot. 36: 1341-1351.
Reyniers F.N., J.M. Kalms & J. Ridders. 1975. Étudedu comportement de deux types de variétés de riz selon leur alimentation hydrique. I. Étudedes facteurs permettant d’esquiver la séche-resse. Rapp. Inst. Rech. Agron. Trop. (IRAT),Côte d’Ivoire.
SAS Institute, Inc. 1989. SAS/STAT user’s guide,version 6, 4th edition, volume 1. Cary, N.C.: SASInstitute Inc.
Simmonds N.W. 1962. The evolution of bananas.Longman, Londres, Royaume-Uni.
Simmonds N.W. 1966. Bananas. Tropical AgricultureSeries, Longman, Londres, Royaume-Uni.
Swennen R. 1984. A physiological study of the suc-kering behavior in plantain (Musa cv. AAB). Ph.D. thesis, Dissertationes de Agricultura n° 132,Faculty of Agriculture, Katholieke UniversiteitLeuven, 180 pp.
Swennen R., E.A. De Langhe, J. Janssen & D. Deco-ene. 1986. Study of the root development of someMusa cultivars in hydroponics. Fruits 41: 515-524.
Tennant D. 1975. A test of a modified line intersectmethod of estimating root length. J. Ecol. 63:995-1001.
Vakili N.G. 1967. The experimental formation of polyploidy and its effects in the genus Musa. Am.J. Bot. 54: 24-36.
Vandenhout H., R. Ortiz, D. Vuylsteke, R. Swennen& K.V. Bai. 1995. Effect of ploidy on stomatal andother quantitative traits in plantain and bananahybrids. Euphytica 83: 117-122.
Vuylsteke D. 1989. Shoot-tip culture for the propa-gation, conservation, and exchange of Musagermplasm. Practical manuals for handling cropgermplasm in vitro 2. International Board forPlant Genetic Resources, Rome, Italie. 56 pp.
Vuylsteke D. 1998. Shoot-tip culture for the propa-gation, conservation, and distribution of Musagermplasm. International Institute of TropicalAgriculture, Ibadan, Nigeria. 82 pp.
Vuylsteke D. & D. Talengera. 1998. Postflask Manage-ment of Micropropagated Bananas and Plantains.
A manual on how to handle tissue-cultured ba-nana and plantain plants. International Instituteof Tropical Agriculture, Ibadan, Nigeria. 15 pp.
Zobel R.W. 1975. The genetics of root development.Pp. 261-275 in The development and function ofroots (G. Torrey & D.C. Clarkson, eds.). Acade-mic Press, Londres and New York.
G. Blomme et A. Tenkouano ont effectué cette re-cherche à la Crop Improvement Division, Institut In-ternational d’Agriculture Tropicale (IITA), Onne HighRainfall Station, L. W. Lambourn & Co., CarolynHouse, 26 Dingwall Road, Croydon CR9 3EE,Royaume-Uni et R. Swennen au Laboratory of Tropi-cal Crop Improvement, Katholieke Universiteit Leuven(K.U.Leuven), Kasteelpark Arenberg 13, 3001 Leuven,Belgique.G. Blomme travaille actuellement à Kampala (Ou-ganda) comme Coordonnateur régional adjoint del’INIBAP pour l’Afrique orientale et australe.
INFOMUSA — Vol 9, N° 2 7
Tableau 4. Caractéristiques de la croissance chez les groupes génomiques triploïdes de Musa au stade de la floraison.
Groupe génomique
Caractéristiques # AAA AAB ABB
SF 58 208 ± 9 730 92 365 ± 5 888 85 513 ± 7 710
NF 8 ± 1 11 ± 0,5 12 ± 0,9
HP 215 ± 11 257 ± 8 268 ± 8
CP 55 ± 2 62 ± 1 69 ± 2
PS 3 945 ± 354 6 285 ± 290 5 662 ± 369
HS 23 ± 2 22 ± 1 22 ± 1
PGL 17 ± 1 22 ± 1 20 ± 1
NR 14 ± 1 13 ± 1 8 ± 1
HR 165 ± 23 106 ± 10 150 ± 22
PSRA 220 ± 30 254 ± 31 361 ± 39
NRA 135 ± 10 151 ± 9 195 ± 16
LRA 5 285 ± 793 6 465 ± 397 6 599 ± 618
DM 4,9 ± 0,1 5,8 ± 0,2 5,3 ± 0,1
PST 556 ± 81 409 ± 18 567 ± 45
LT 14 883 ± 2 647 11 278 ± 1 358 11 379 ± 1 266
% PSRAPM 43 ± 5 62 ± 7 64 ± 4
% LRAPM 39 ± 5 61 ± 5 60 ± 5
NJJF 368 ± 24 338 ± 9 340 ± 25
SF/NJJF 168 ± 35 276 ± 20 264 ± 32
HP/NJJF 0,61 ± 0,06 0,77 ± 0,04 0,82 ± 0,06
PS/NJJF 11 ± 1 19 ± 1 17 ± 2
HR/NJJF 0,45 ± 0,06 0,32 ± 0,03 0,46 ± 0,07
PSRA/NJJF 0,60 ± 0,07 0,77 ± 0,11 1,13 ± 0,15
NRA/NJJF 0,38 ± 0,05 0,45 ± 0,03 0,61 ± 0,08
LRA/NJJF 15 ± 2 19 ± 1 20 ± 2
PST/NJJF 1,50 ± 0,19 1,21 ± 0,05 1,75 ± 0,17
LT/NJJF 40 ± 7 33 ± 3 34 ± 3# voir tableaux 2 et 3.

Lazaro L. Castellanos Lopez, JorgeLopez Torrez, Julian Gonzalez
Rodriguez, Sergio Rodriguez Moraleset José De La C. Ventura Martín
La culture des bananiers et des bana-niers plantain représente d’ impor-tantes ressources sur le plan alimen-
taire et économique pour une grandepartie de la population mondiale, principa-lement dans les pays en voie de développe-ment d’Asie, d’Afrique et d’Amérique la-tine. Bien que dans beaucoup de ces payson continue à employer des méthodes depropagation conventionnelles, la culture invitro commence, depuis ces dernières an-nées, à être utilisée comme une alternativenovatrice pour la multiplication.
La culture in vitro permet en effet d’ob-tenir des plantes exemptes de parasites etde maladies avant de les transférer auchamp. Cependant, ces plantes sont en-core très fragiles lors de cette transplanta-tion ; cela les rend très sensibles aux at-taques de nématodes phytoparasites etprovoque parfois des pertes considérablescomme par exemple dans le cas d’ attaquedu nématode Meloydogine spp. Ces pertespeuvent presque complètement être sup-primées si l’on désinfecte le sol avant laplantation ou bien si l’on effectue la plan-tation sur des sols libres de nématodes.Pourtant, cela n’est pas si facile à obtenir :d’une part, l’application de nématicideschimiques affecte considérablement leprocessus de production et détruit l’équi-
libre écologique du sol et d’autre part, lesméthodes de détection des nématodesdans le sol ne sont pas totalement fiables.En effet, quand la population des parasitesdans le sol est très faible, il est pratique-ment impossible d’y détecter leur pré-sence. Par conséquent, on pensera dispo-ser de sols apparemment exempts denématodes dangereux pour les bananierset les bananiers plantain sur lesquels l’ap-plication de nématicides ne se justifieradonc pas. Cependant, au fil des mois, onpourra vérifier que les populations de né-matodes sont effectivement présentes etque les racines de ces plantules sans dé-fenses sont pratiquement inexistantes oubien recouvertes de nodules épais.
L’emploi de champignons destructeurs denématodes (nematode-trapping fungi) ap-partenant à différents genres comme Har-posporium sp., Dactylella spp., Stylopagesp., Dactylaria spp., Catenaria sp. et Arthrobotrys sp. (Duddington 1956, Cor-tado 1968, Generalao 1986, Stirling 1988,Persson 1997) paraît être une alternativeprometteuse pour faire face à ce problème.
Ces organismes offrent plusieurs typesd’avantages pour leur utilisation commecontrôle biologique des nématodes, parmilesquels :• Leur capacité d’attraper et d’éliminer un
grand nombre d’espèces de nématodescar ils possèdent des structures ou desorganes de capture spécialisés pour piéger les parasites en mouvement (anneaux, contractiles ou non ; filets ;structures adhésives et autres). Ceci est
particulièrement intéressant puisque lecontrôle se réalise donc avant que le né-matode ne pénètre dans la racine et yprovoque des dégâts.
• Leurs deux types de cycle biologique : 1) une phase saprophyte durant laquelleils n’utilisent que la matière organiquedu sol comme source de carbone (éner-gie) et d’acides aminés (azote) et 2) unephase parasitaire où ils se nourrissentuniquement de la matière organique desnématodes capturés (Stirling 1988, Pers-son 1997). On a pu observer qu’en pré-sence de nématodes ils sont capables depasser rapidement de la phase sapro-phyte à la parasitaire et qu’en outre celaprovoquait la germination de spores et ledéveloppement des organes de capture.
• Leur capacité à produire des substancesattractives pour les parasites ce qui aug-mente encore l’efficacité du contrôle deces derniers.
• L’émission chez certains d’une grandequantité de spores de résistance ce quipermet de les prescrire de différentes fa-çons.L’INIVIT conserve un stock de champi-
gnons nématophages et/ou parasites de né-matodes isolés à partir de sols cubainsplantés de bananiers et de bananiers plan-tain. Beaucoup d’entre eux ont déjà été ca-ractérisés et ont fait preuve d’une grandepathogénicité vis à vis des principales es-pèces de nématodes qui affectent les cul-tures bananières du pays.
L’introduction de champignons némato-phages dans la rhizosphère des vitroplantspermettrait de réduire ou d’éliminer lespertes de production, de diminuer les coûtsconsacrés à l’emploi des substances chi-miques et de préserver les sols puisque,dans cette optique, la protection racinairedes plantules se ferait de façon naturelle etécologique. C’est pourquoi ce travail a étéentrepris afin de vérifier l’efficacité de lasouche INIVIT 99 - 1 TPB de Arthrobotryssp. pour la protection racinaire des bana-niers plantains CEMSA 3/4 (Musa AAB)multipliés par micropropagation.
Matériel et méthodesLes recherches se sont déroulées à l’INIVIT,dans la zone de sevrage des vitroplants, aucours de l’année 1999.
On a utilisé des vitroplants du clone debananier plantain CEMSA 3/4 (Musa spp.AAB) et les travaux ont porté seulementsur la phase d’adaptation. Les traitements
8 INFOMUSA — Vol 9, N° 2
Figure 1. Effet du nématicide biologique pour la protection racinaire du plantain multiplié par micropropagation.
0% d
e p
aras
itis
me
A.TémoinB. Nématicide biologique + R. similisC. R. SimilisD. Nématicide biologique
80
60
40
20
0
A B C D
Emploi d’un nouveau nématicide biologique pour laprotection racinaire du bananier plantain(Musa AAB) multiplié par micropropagation
Lutte biologique Un champignon nématophage

sont répertoriés en : A = témoin, B = né-maticide biologique + inoculum de Radopholus similis, C = inoculum de R. si-milis et D = nématicide biologique seul.
On a utilisé des vitroplants prêts à êtresevrés. On les a semés en pots contenantun substrat stérilisé préparé à partir de solrouge, de compost et de bagasse. Dix joursaprès le semis, on a inoculé les traitementsB et D avec le nématicide. Au bout de cinqjours, on a appliqué sur chacun des potsdes traitements B et C, une suspension de5 x 103 nématodes (R. similis) préparéspar culture in vitro sur rondelles de ca-rottes (Daucus carota). Soixante joursaprès, on a évalué le pourcentage d’infec-tion des racines, on a relevé le poids total,le poids des racines et on a mesuré la hau-teur de chaque vitroplant.
Résultats et discussionContrairement au traitement C, les traite-ments B et D n’ont pas présenté de diffé-rences significatives par rapport au témoinA pour tous les paramètres évalués(figure 1). Ceci montre que, quand lecontrôle biologique est présent, R. similisne provoque pas de dommages importantssur les racines des vitroplants. Ces résul-tats confirment ceux rapportés pourd’autres nématicides biologiques (Jatala1986, Davide 1994) utilisés pour la protec-tion racinaire d’autres cultures.
Les plants sur lesquels on avait appliquéle nématicide seul ont une hauteur et unpoids racinaire supérieurs à ceux desautres traitements ; les différences signifi-catives étant minimes en ce qui concernele témoin et le traitement B mais élevéesavec le traitement C (tableau 1).
La taille plus élevée et le poids plus im-portant des vitroplants des traitements oùon a introduit la souche INIVIT 99 - 1 TPB
peuvent être dus au fait que cet organismeparticipe à la décomposition de la matièreorganique et libère des éléments nutritifsdans le sol, éléments qui peuvent alors êtreassimilés par les vitroplants, ce qui n’estpas le cas dans les traitements où l’on n’apas inclus de microorganismes. En outre, ilest possible que ces organismes élaborentdes substances stimulantes pour la crois-sance végétale comme cela se produitavec d’autres microorganismes du sol (Da-vide 1994).
Conclusions et recommandations• L’utilisation du nématicide biologique
(Cepa INIVIT 99 - 1 TPB de Arthrobotryssp.) protège efficacement les racines desvitroplants de bananiers plantain CEMSA3/4 contre les attaques de R. similis.
• Quand on l’emploie en phase d’adapta-tion, INIVIT 99 - 1 TPB associé à du com-post et à de la bagasse, augmente la tailleet le poids des vitroplants de CEMSA 3/4.
• Il est recommandé d’utiliser le nouveaunématicide biologique INIVIT 99 – 1 TPBpour protéger les racines des vitroplantsde bananiers plantain.
• Il est recommandé de vérifier l’efficacitédu nématicide sur d’autres clones de ba-naniers ou de plantains sensibles aux at-taques de phytonématodes.
• Il est recommandé de vérifier l’efficacitédu nématicide sur d’autres espèces de nématodes de grande importance écono-mique comme Meloydogine spp., Pratylenchus coffeae et Helicotylenchusmulticinctus. ■
RéférencesCortado R & R.G. Davide. 1968. Nematode-trapping
fungi in the Philippines (abstr). Phil. Phytopath.4: 4.
Davide R.G. 1994. Biological control of banana ne-matodes: development of BIOCON I (BIOACT)and BIOCON II technologies. Pp. 139-146 in Ba-nana nematodes and weevil borers in Asia andthe Pacific. Proceedings of a conference-work-shop on nematodes and weevil borers affectingbananas in Asia and the Pacific. 18-22 April 1994,Serdang, Malaysia (R.V. Valmayor, R.G. Davide,J.M. Stanton, N.L. Treverrow and V.N. Roa, eds.).ASPNET Book Series 5. INIBAP/ASPNET, LosBaños, Philippines.
Duddington C.L. 1956. The friendly fungi. Faber andFaber Ltd., London. 168 pp.
Generalao L. & R.G. Davide. 1986. Biological controlof Radopholus similis with three nematopha-gous fungi. Phil. Phytopath. 22: 36-41.
Jatala P. 1986. Biological Control of Plant ParasiticNematodes. Ann. Rev. Phytopathol. 459-489.
Persson C & H.B. Jansson. 1997. Colonization of soilby nematophagous fungi. Tercer SeminarioCientífico Internacional sobre Sanidad Vegetal.Ciudad Habana. Resúmenes. 127 pp.
Stirling G.R. 1988. Biological Control of Plant Parasi-tic Nematodes. Pp. 93-139 in Diseases of nema-todes. Vol II. CRC Press Inc., Boca Raton, Florida.
Les auteurs travaillent à l’Instituto de Investigacionesen Viandas Tropicales, INIVIT, Santo Domingo, VillaClara, Cuba, CP. 53000, courrier électronique : [email protected]
INFOMUSA — Vol 9, N° 2 9
Tableau 1. Effet des différents traitements sur le poids racinaire des vitroplants debananiers plantain.
Traitement Témoin A B C D
Poids des racines (g.) 5.9a 6.2a 1.3b 7.3a* Différence significative pour p>0.05.
Edna A. Aguilar, David W. Turner et K. Sivasithamparam
On a observé le flétrissement deplants de bananiers Cavendish auxPhilippines (Stover 1990) et à Car-
narvon dans l’ouest de l’Australie (Pegg etal. 1995) de manière localisée, dans des
conditions sous-optimales comme l’insuffi-sance de drainage et l’engorgement. Cephénomène a été signalé occasionnelle-ment, en dehors d’un cadre expérimental.
L’excès d’eau peut créer des problèmesdans les bananeraies, en particulier aprèsde fortes pluies ou quand on irrigue un sollourd. Si les champs sont inondés ou res-tent longtemps engorgés, le système raci-
naire est parfois endommagé et les bana-niers peuvent devenir plus sensibles à lafusariose.
L’engorgement réduit la concentrationd’O2 et augmente la concentration de CO2et d’éthylène dans le sol (Ponnamperuma1984). La diffusivité de l’O2 dans l’eauétant égale au 1/10 000e de sa diffusivitédans l’air, l’O2 dissous dans la solution dusol se trouve épuisé en quelques heures ouquelques jours du fait de la consommationdes racines des plants et des microorga-nismes du sol (Drew 1990). Or, l’O2 est es-sentiel pour la respiration, processus parlequel les organismes aérobies produisentde l’énergie sous forme d’ATP. Nous exami-nerons ici les études récentes sur la ré-ponse à court terme des racines des bana-niers et celle de l’agent causal de lafusariose (Fusarium oxysporum f. sp.
Proposition d’explicationdes mécanismes prédisposantles bananiers Cavendish à la fusariose durant l’hypoxie
Physiologie Effet de l’engorgement

cubense – Foc (E.F. Smith) Snyder et Han-sen) à la carence en O2 et nous nous effor-cerons de démontrer comment ces ré-ponses jouent un rôle dans la prédispo-sition à la fusariose chez des cultivars de bananiers réputés résistants à cette maladie.
Rôle possible des aérenchymesLa présence de lacunes aérifères lysigènes(les aérenchymes) dans les racines a étémise en évidence chez un certain nombrede cultivars de bananiers dans des étudesantérieures (Acquarone 1930, Riopel etSteeves 1964, Aguilar et al. 1999). Les aé-renchymes assurent la continuité de la dif-fusion de l’O2 des parties aériennes duplant aux racines, en augmentant le fluxd’O2 à travers le cortex. Nous avons quanti-fié la porosité des racines de différents cul-tivars de bananiers et mesuré les diffé-rences inhérentes dans la facilité dupassage de l’O2 à travers les racines. Les
aérenchymes représentent 10 % du volumedes racines matures des cultivars Caven-dish (AAA), tandis que chez Goldfinger(AAAB), cette proportion est de 5 % (Agui-lar et al. 1999). L’hypoxie accroît la poro-sité et l’épaisseur des racines (figure 1).En comparant la résistance physique à ladiffusion interne des gaz, nous avonsconstaté la disparition des différencesentre les quatre cultivars étudiés (Aguilaret al. 1998), ce qui montre que leurs ra-cines sont également adaptées aux condi-tions stagnantes, dans lesquelles elles ontla capacité de conduire l’O2 gazeux trois àcinq fois plus facilement que les racinesaérées. La concentration d’O2 dans les tis-sus racinaires est sensible aux change-ments dans la concentration extérieured’O2. En ce qui concerne les aérenchymescorticaux, la stèle, qui est le site où l’agentpathogène initie la maladie, a de faiblesconcentrations d’O2 (1,3-2,6 kPa) même sile milieu extérieur aux racines est entière-
ment aéré (21 kPa) (Aguilar et al. 1998)(figure 2). L’hypoxie (4 kPa O2) à l’exté-rieur des racines induit une anoxie dans lastèle des racines de bananier excisées(Aguilar 1998). Si la concentration d’O2 àla surface des racines est réduite à environ18 kPa, on estime que cela crée déjà unnoyau anoxique dans la stèle. Cette obser-vation a des implications en ce quiconcerne le développement de la fusariose,car c’est l’interaction entre l’hôte et l’agentpathogène dans la stèle qui joue un rôle cri-tique dans le développement de la maladie.Quand la stèle est en conditions d’anoxie,la mobilisation des mécanismes de dé-fense dans les racines de l’hôte infectépeut se trouver ralentie, voire stoppée,puisque la plupart de ces processus néces-sitent de l’énergie.
Si l’agent de la fusariose parvenait àmieux tolérer les faibles concentrationsd’O2, il aurait la possibilité de coloniser lesracines affectées et d’être systématique-ment distribué sur toute leur longueur.Dans des études in vitro, la croissance my-célienne est demeurée inaffectée même à1 % d’O2, mais s’est trouvée stoppée enconditions d’anoxie (0 % d’O2). On a ob-servé qu’elle reprenait quand l’aération re-commençait (Aguilar 1998). Les conidiesen germination de Foc avaient une faibledensité d’hyphes quand l’O2 devenait limi-tant et cessaient fréquemment leur acti-vité, produisant des chlamydospores ouhyphes au repos (figure 3). Nos études ontaussi montré que l’agent pathogène pou-vait exploiter la présence des aérenchymeset des concentrations plus élevées d’O2qu’ils contiennent (Aguilar 1998) (figure 4).
Ainsi, les aérenchymes, bien qu’ilsconstituent un avantage pour permettre àl’hôte de survivre en conditions hy-poxiques, pourraient bien être le « talond’Achille » de certains cultivars de bana-niers, qu’ils empêcheraient de résister à lafusariose. Il apparaît que les aérenchymesoffrent à l’agent pathogène une autre voie,en dehors du système vasculaire, pour en-vahir la racine longitudinalement. On peutenvisager un scénario dans lequel l’agentpathogène, se servant des aérenchymescomme base, pénètre occasionnellement àl’intérieur de la stèle pour avoir accès auxéléments nutritifs, puis ramène ceux-ci parl’intermédiaire du flux cytoplasmique versle mycélium à l’intérieur des aérenchymes,où il trouve les conditions requises pour sarespiration. L’agent pathogène est ainsi enmesure d’étendre sa croissance jusquedans la stèle qui, en elle-même, est norma-lement un environnement défavorable. Onpeut supposer que, si les conditions dans lastèle lui sont inhospitalières, l’agent patho-gène peut avoir recours à une phase de la-tence. Le processus de passage de l’agentpathogène par les aérenchymes pourraitcontribuer à l’invasion rapide de la souche(Aguilar 1998).
10 INFOMUSA — Vol 9, N° 2
Figure 2. Profil d’oxygène d’une racine de bananier (à 28 mm de l’apex), mesuré à l’aide d’une microélectrode dans une solution en écoulement.
Figure 1. Développement des aérenchymes dans les racines du cv. Williams cultivé en milieu nutritifstagnant ou aéré. Les racines mesuraient environ 200 mm de long. Les coupes ont été faites à 50 mmet 100 mm de l’extrémité des racines. (échelle 0,1 mm).
Racines en milieu stagnant(50mm de l’extrémité)
Couche intermédiaire
Cortex non-poreux
Cortex poreux
stele
Dif
fusi
on
d’o
xyg
ène
Co
ncen
tration
d’o
xygèn
e
Racines en milieu aéré(50mm de l’extrémité)
Racines en milieu stagnant(100mm de l’extrémité)
Racines en milieu aéré(100mm de l’extrémité)

Rôle de la réduction del’élongation des racines et de la mort des extrémités des racines en cas d’anoxieEn établissant l’anoxie (0 % d’O2) dans lemilieu, on a constaté que les racines arrê-taient de s’allonger dans un délai de 30 mi-nutes. Le retour de l ’aération après4 heures d’anoxie entraînait la reprise del’élongation, mais seulement dans une pro-portion de 50 % par rapport aux racines aé-rées de manière continue. Une anoxie deplus de 6 heures avait pour effet de tuer lesextrémités des racines (Aguilar 1998) (fi-gure 5). La réduction de la croissance raci-naire peut favoriser l’infection en augmen-tant le temps pendant lequel les partiessensibles des racines sont exposées à l’ino-culum, tandis que la mort des extrémitésdes racines ouvre à l’agent pathogène unevoie d’infection qui lui donne directementaccès à la stèle. Ainsi, une inondation,même temporaire, semble faciliter l’entréeet peut-être aussi l’activité de l’agent pa-thogène dans les racines, où il ne ren-contre pas les mécanismes de résistancenormalement présents dans des tissus nonaffectés. Foc semble être un nécrotrophefacultatif, capable d’être un endophytechez les hôtes sensibles comme chez leshôtes résistants.
Exposées à plusieurs épisodes d’anoxie decourte durée, dont chacun n’était pas assezsévère pour tuer leurs extrémités, les ra-cines ont été en mesure de survivre et ontcontinué à s’allonger, mais à un rythme ré-duit (Aguilar et al. 1998). Cela montre queles racines ont la capacité de s’acclimater àl’hypoxie. On a signalé par ailleurs qu’unprétraitement hypoxique améliorait la tolé-rance du maïs à l’anoxie (Gibbs et al. 1998).
Rôle des enzymes péroxydase(PER) et phénylalanine ammonia-lyase (PAL)Les enzymes PER et PAL jouent un rôlefondamental dans l’induction de la résis-
tance. La production de PER est en corréla-tion avec l’accroissement de la lignificationet la résistance à la maladie, par exemplechez le chou vis-à-vis de Fusarium oxysporum f. sp. conglutinans (Heitefusset al. 1960), chez la pomme de terre vis-à-vis de Phytophthora infestans (Friend et al. 1973) et chez les bananiers diploïdes(Musa acuminata Colla) vis-à-vis de Foc(Morpugo et al. 1994). Quant à la PAL, sonactivité est en corrélation avec la résis-tance de plantes telles que le niébé vis-à-vis de Phytophthora vignae (Ralton et al.1988) et le soja vis-à-vis de P. mega-sperma f. sp. glycinea (Bhattacharya etWard 1988).
Nous avons étudié les effets de l’infec-tion par Foc et de l’hypoxie sur l’activité dela PER et de la PAL dans les racines decultivars réputés résistants ou sensibles àcet agent pathogène (Aguilar et al. 2000).
Il s’est avéré que l’hypoxie stimulait l’ac-tivité de la PAL et de la PER. On ne sait sil’accroissement de l’activité de ces en-zymes résultant du stress hypoxique peut
offrir une protection à l’hôte. L’infectionpar Foc n’a accru que l’activité de la PER.Quand l’hypoxie et l’infection par l’agentpathogène étaient combinées, les diffé-rences enregistrées dans la rapidité et ledegré d’accroissement de l’activité de laPAL et de la PER semblaient associées à larésistance à la fusariose, et en particulier àla rupture de résistance chez Williams(cultivar Cavendish). Goldfinger, réputéplus résistant à la fusariose, a répondu parune activité de la PER et de la PAL plusimportante et plus soutenue que celle en-registrée chez les cultivars Williams etGros Michel (figures 6 et 7). Ces diffé-rences quantitatives pourraient être l’undes facteurs qui font que Williams suc-combe aux attaques de la fusariose enconditions d’engorgement (Aguilar et al.2000). Ce sont là des observations extrême-ment intéressantes, qui nécessitent des re-cherches plus approfondies. Si le traite-ment hypoxique peut être un moyen destimuler les mécanismes de défense des ra-cines, il doit se situer à un niveau, à un mo-ment et pendant une durée de nature à ac-croître l’activité de la PER et de la PALsans porter atteinte de manière irrémé-diable aux fonctions des racines. Il devraitêtre possible de trouver chez les cultivarsde bananiers des marqueurs génétiquespour ces enzymes, dont on pourrait se ser-vir afin d’accroître la résistance quantita-tive à la fusariose.
Le site de la synthèse et de l’activité deces enzymes dans la racine est d’une im-portance déterminante pour la résistanceà la fusariose. Pour être efficace, l’activitéenzymatique accrue doit se concentrerdans ou autour du système vasculaire, oùopèrent les mécanismes actifs de défensede l’hôte, ainsi que dans les tissus adja-cents à l’extrémité de la racine, qui peu-vent être des sites d’infection, particulière-ment en cas de mort de l’extrémité de laracine. Le métabolisme des phénols est un
INFOMUSA — Vol 9, N° 2 11
Figure 4. Croissance des hyphes de l’agent de la fusariose dans les aérenchymes corticaux de racinesde bananier.
Den
sité
des
hyp
hes
(m
m/m
m2 )
Longueur du flux de diffusion (mm)
40
35
30
25
20
15
10
5
00 2 4 6 8 10 12
Figure 3. Effet de l’accroissement de la longueur du flux de diffusion (mm) de l’O2 sur la densité deshyphes (mm/mm2) de Fusarium oxysporum f. sp. cubense (Foc) 24 heures après l’inoculation.

processus d’oxydation et l’hypoxie devraitavoir rendu des parties de la stèleanoxiques, tandis que le cortex sera relati-vement plus aéré. Il est possible que laPAL soit synthétisée dans les parties plusaérées de la racine, mais il reste à établirsi elle peut avoir une activité dans la stèle,où elle est fortement nécessaire (Aguilar etal. 2000).
On a constaté que l’activité des enzymesPAL et PER diminuait après le retour del’aération, tandis que l’agent pathogène(qui forme des spores au repos en condi-tion d’hypoxie) reprenait aisément unecroissance mycélienne normale. En consé-quence, dans une interaction avec un hôterésistant, la reprise d’activité de Foc
quelques heures après le retour de l’aéra-tion devrait aller de pair avec la poursuitede l’activité enzymatique. La solution opti-male, pour que l’hôte fasse preuve de résis-tance, serait d’accroître suffisamment l’ac-tivité enzymatique durant l’hypoxie pourcontenir l’agent pathogène, qui est alorsinactif, avant la reprise de l’aération. Uneblessure post-anoxie, alors que le stress de-vient trop sévère, pourrait priver l’hôte deson aptitude à maintenir la résistance(Aguilar et al. 2000).
ConclusionC’est l’état physiologique de l’hôte au mo-ment de la reprise de l’aération qui déter-mine le résultat de l’interaction hôte-agent
pathogène lorsqu’il y a carence en oxygène.Un endommagement irréparable des fonc-tions racinaires ne peut que favoriserl’agent pathogène, et donc le développe-ment de la maladie, voire son intensifica-tion. Il semble que si la racine s’acclimateet produit des réactions de défense suffi-samment rapides et fortes durant l’hy-poxie, elle puisse prendre l’avantage surl’agent pathogène. Lorsqu’un nouveaustress aura lieu, sa durée et sa sévérité dé-termineront très probablement la suite dela dynamique hôte-agent pathogène (Agui-lar 1998). ■
RéférencesAcquarone P. 1930. The roots of Musa sapientum
L. Rep. No. 26. United Fruit Co., Research Dept.Boston, Mass.
Aguilar E.A. 1998. Responses of banana roots to oxy-gen deficiency and implications for Fusariumwilt infection. PhD Thesis, The University of Wes-tern Australia. 148 pp.
Aguilar E.A., D.W. Turner, D.J. Gibbs, W. Armstrong& K. Sivasithamparam. 1998. Response of banana(Musa sp.) roots to oxygen deficiency and its im-plications for Fusarium Wilt. Acta Horticulturae490: 223-228.
Aguilar E.A., D.W. Turner & K. Sivasithamparam.1999. Aerenchyma formation in roots of four ba-nana (Musa spp.) cultivars. Scientia Horticultu-rae 80: 52-72.
Aguilar E.A., D.W. Turner & K. Sivasithamparam.2000. Fusarium oxysporum f.sp. cubense inocu-lation and hypoxia alter peroxidase and phenyla-lanine ammonia lyase enzyme activities in nodalroots of banana cultivars (Musa sp.) differing in
12 INFOMUSA — Vol 9, N° 2
Figure 6. Effet de l’inoculation avec Fusarium oxysporum f. sp.cubense (Foc) et de l’hypoxie sur l’activité de l’enzymephénylalanine ammonia-lyase (PAL) dans les racines des cultivars a) Williams, b) Goldfinger, c) Gros Michel et d) Sugar. Les traitements étaient les suivants : sans Foc, continuellementaéré (NFA) ; inoculé avec Foc, continuellement aéré (FA) ; inoculéavec Foc, continuellement en hypoxie (FHH) ; sans Foc,continuellement en hypoxie (NFHH) ; sans Foc, en hypoxie pendant48 heures, puis de nouveau aéré (NFHA) ; et inoculé avec Foc, enhypoxie pendant 48 heures, puis de nouveau aéré (FHA). Les barresverticales placées sur les courbes indiquent les erreurs types quandelles ont une dimension supérieure à celle des symboles. Les barresverticales placées en dehors des courbes indiquent les différencesmoyennes significatives au seuil p = 0,05 (test d’amplitude multiplede Duncan). Du haut à la première division, on a la comparaisonentre les points adjacents, tandis que du haut à la division du bas,on a la comparaison entre le point supérieur et le point inférieur.
a) Williams
25
20
15
10
5
00 24 48 72 96 120 144
0 24 48 72 96 120 144
c) Gros Michel25
20
15
10
5
0
b) Goldfinger25
20
15
10
5
00 24 48 72 96 120 144
Temps (heures)
Temps (heures)
Temps (heures)
Temps (heures)
d) Sugar25
20
15
10
5
00 24 48 72 96 120 144
NFA FA FHHNFHH NFHA FHA
Act
ivite
PA
L (
mol
CA
/h/g
Foc
)A
ctiv
ite P
AL
( m
ol C
A/h
/g F
oc)
Act
ivite
PA
L (
mol
CA
/h/g
Foc
)A
ctiv
ite P
AL
( m
ol C
A/h
/g F
oc)
Figure 7. Effet de l’inoculation avec Fusarium oxysporum f. sp. cubense (Foc)et de l’hypoxie sur l’activité de l’enzyme péroxydase (PER) dans les racinesdes cultivars a) Williams, b) Goldfinger, c) Gros Michel et d) Sugar. Lestraitements étaient les suivants : sans Foc, continuellement aéré (NFA) ;inoculé avec Foc, continuellement aéré (FA) ; inoculé avec Foc,continuellement en hypoxie (FHH) ; sans Foc, continuellement en hypoxie(NFHH) ; sans Foc, en hypoxie pendant 48 heures, puis de nouveau aéré(NFHA) ; et inoculé avec Foc, en hypoxie pendant 48 heures, puis de nouveauaéré (FHA). Les barres verticales placées sur les courbes indiquent les erreurstypes quand elles ont une dimension supérieure à celle des symboles. Lesbarres verticales placées en dehors des courbes indiquent les différencesmoyennes significatives au seuil p = 0,05 (test d’amplitude multiple deDuncan). Du haut à la première division, on a la comparaison entre les pointsadjacents, tandis que du haut à la division du bas, on a la comparaison entrele point supérieur et le point inférieur. La ligne horizontale HH indique ladurée du traitement hypoxique continu (120 heures au total) et la lignehorizontale HA indique la durée de l’épisode d’hypoxie (48 heures) et lemoment de reprise de l’aération (72 heures).
0 24 48 72 96 120 144
a) Williams10
8
6
4
2
0
Act
ivite
PE
R (
AO
D/s
/g F
oc)
Act
ivite
PE
R (
AO
D/s
/g F
oc)
Temps (heures)
HH
HA
0 24 48 72 96 120 144
b) Goldfinger10
8
6
4
2
0
Temps (heures)
0 24 48 72 96 120 144
c) Gros Michel10
8
6
4
2
0
Temps (heures) Temps (heures)
d) Sugar10
8
6
4
2
00 24 48 72 96 120 144
NFA FA FHH NFHH NFHA FHA
Act
ivite
PE
R (
AO
D/s
/g F
oc)
Act
ivite
PE
R (
AO
D/s
/g F
oc)
Figure 5. Élongation des racines du cv. Williams dans une solution nutritive aérée (témoin) et après2,4 et 6,5 heures d’anoxie.
0
50
40
30
20
10
050
40
30
20
10
0
50
40
30
20
10
050
40
30
20
10
15 30 45 60 0 15 30 45 60
0 15 30 45 60 0 15 30 45 60
Temps (heures)
Elo
ng
atio
n (
mm
)
Témoin aéré Reprise de l'élongation
Reprise de l'élongation
Reprise de l'élongationan
oxi
e
ano
xie
ano
xie
aéré
aéré
aéré
2h
4h 6.5h
1.02 = 0.08
0.45 = 0.04
0.88 = 0.15 0.02 = 0.02
0.47 = 0.060.83 = 0.14 0.42 = 0.01

their susceptibility to Fusarium wilt. AustralianJournal of Botany 48: 589-596.
Bhattacharya M.K. & E.W.B. Ward. 1988. Phenylala-nine ammonia-lyase activity in soybean hypoco-tyls and leaves following infection with Phytoph-thora megasperma f.sp. glycinea. CanadianJournal of Botany 66: 18-23.
Drew M.C. 1990. Sensing soil oxygen. Plant, Cell andEnvironment 13: 681-693.
Friend J., S.B. Reynolds & M.A. Aveyard. 1973. Phe-nylalanine ammonia-lyase, chlorogenic acid andlignin in potato tuber tissue inoculated with Phytophthora infestans. Physiological Plant Pa-thology 3: 495-507.
Gibbs J., D. W. Turner, W. Armstrong, M.J. Darwent& H. Greenway. 1998. Response to oxygen defi-ciency in primary roots of maize. 1. Developmentof oxygen deficiency in the stele reduces radialsolute transport to the xylem. Australian Journalof Plant Physiology 25: 745-758.
Heitefuss R., M.A. Stahmann & J.C. Walker. 1960.Oxidative enzymes in cabbage infected by Fusa-rium oxysporum f. conglutinans. Phytopatho-logy 50: 370-375.
Morpugo R.S., V. Lopato & F.J. Novak. 1994. Selec-tion parameters for resistance to Fusarium oxysporum f.sp. cubense race 1 and race 4 on diploid banana (Musa acuminata Colla). Euphytica 75: 121-129.
Pegg K.G., R.G. Shivas, N.Y. Moore & S. Bentley.1995. Characterisation of a unique population ofFusarium oxysporum f.sp. cubense causing Fu-sarium wilt in Cavendish bananas at Carnarvon,Western Australia. Australian Journal of Agricul-tural Research 46: 167-178.
Ponnamperuma F.N. 1984. Effects of flooding onsoils. Pp. 10-46 in Flooding and Plant Growth(T.T. Kozlowski, ed.). Academic Press, Inc., Florida.
Ralton J.E., B.J. Howlett, A.E. Clarke, J.A.G. Irwin &B. Imrie. 1988. Interaction of cowpea with Phy-tophthora vignae: inheritance of resistance andproduction of phenylalanine ammonia-lyase as aresistance response. Physiological and MolecularPlant Pathology 32: 89-103.
Riopel J.L. & T.A. Steeves. 1964. Studies on theroots of Musa acuminata cv. Gros Michel: theanatomy and development of main roots. Annalsof Botany 28: 475-494.
Stover R.H. 1990. Fusarium wilt of banana: somehistory and current status of the disease. Pp. 1-18in First International Conference on FusarialWilt of Banana (R.C. Ploetz, ed.). American Phy-topathological Society Press, St. Paul, Minn.
Edna A. Aguilar travaille au Farming Systems and
Soil Resources Institute, College of Agricuture, Uni-
versity of the Philippines at Los Baños, College, La-
guna 4031, Philippines. David W. Turner est spécia-
liste en sciences végétales et K. Sivasithamparam
en sciences du sol et nutrition des plantes à la Faculté
d’agriculture, University of Western Australia, Ned-
lands, Western Australia 6907, Australie
INFOMUSA — Vol 9, N° 2 13
Josué Francisco da Silva Junior, Zilton José Maciel Cordeiro et Arlene Maria Gomes Oliveira
La maladie de Panama ou flétrisse-ment dû à la fusariose du bananier,provoquée par le champignon
Fusarium oxysporum f.sp. cubense (Foc),est un des problèmes les plus sérieux de la culture bananière puisqu’elle est res-ponsable de graves dommages dans leszones qu’elle frappe. On connaît le rôle dé-cisif que joue le génotype des cultivars debananier pour l’expression de leur résis-tance ou de leur sensibilité à la maladie.Cependant, on considère également quel’incidence et la gravité de la maladie dePanama seraient plus ou moins directe-ment liées à des facteurs édaphiques et àla nutrition de la plante et que ceux-ci agi-raient sur les mécanismes de résistancetels que la formation de gel et tylosis(Stover 1962, Borges Perez et al. 1983,Beckman 1990).
Aux îles Canaries, Alvarez et al. (1981),Gutierrez Jerez et al. (1983), Borges Perezet al. (1983) et Trujillo Jacinto del Castilloet al. (1983) ont effectué des observationssur des sols de zones saines ou infectées eten ont conclu que le pH, la teneur en ma-tière organique (MO), les taux de calcium(Ca), de magnésium (Mg) et de zinc (Zn),ainsi que les relations Ca/Mg et K/Mgétaient étroitement corrélés à l’apparitionde la maladie. A Taiwan, Tu et Cheng(1982) ; Hwang (1985) ; Sun et Huang(1985) et Su et al. (1986), ont obtenu desrésultats prometteurs lorsqu’ils entrepri-rent des essais en vue de contrôler la mala-die de Panama grâce à des sols suppressifset conductifs et à l’ajout de divers compo-sés organiques ou inorganiques à des solsoù avaient lieu de sévères attaques de lamaladie. Dans l’Etat de Bahia, au Brésil,des observations ont mis en évidence quela teneur en matière organique était plusélevée dans les sols des zones saines (EM-BRAPA 1987).
Toujours au Brésil, Malburg et al. (1984)ont rapporté que les faibles valeurs du pHet des taux de Ca, de Mg et de Zn dans dessols de l’Etat de Santa Catarina plantés debananiers « Enxerto » (« Prata Anã » AAB)et « Branca »(AAB), étaient reliés à unhaut niveau d’incidence du flétrissement.Des observations effectuées à Bahia et Es-pirito Santo, toujours au Brésil, ont révélé
que, dans les zones infectées, le pH et lestaux de Ca, de Mg et de Zn étaient basalors que les relations K/Ca et K/Mgétaient hautes (EMBRAPA 1987, EMCAPA1988). Dans l’île de Tenerife, Borges Perezet al. (1991) ont constaté que la fertilisa-tion au Zn, pendant trois ans, avait réduitde façon significative l’apparition de la ma-ladie de Panama, au champ, chez le bana-nier « Dwarf Cavendish ».
En considérant ce qui précède, le travailprésenté ici a cherché à évaluer l’effet decaractéristiques chimiques du sol tellesque la teneur en matière organique, le ni-veau du pH et les taux de Ca, de Mg et deZn, sur l’incidence et la gravité de la mala-die de Panama chez le bananier « PrataAnã » (AAB).
Materiel et méthodesL’essai a été conduit sur le champ expéri-mental du Centro Nacional de Pesquisade Mandioca e Fruticultura Tropical(CNPMF), de la Empresa Brasileira dePesquisa Agropecuária (EMBRAPA),situé à Cruz das Almas, Etat de Bahia, auBrésil. Dans des trous de 0,38 m3 (0,70 mde diamètre et 0,30 m de profondeur) revê-tus sur les bords de plastique polyéthylène,on a placé comme substrat soit un sol mi-néral de type latosol alique jaune de cohé-sion Tb, de texture moyenne à argileuse ;soit un sol organique prélevé de 0 à 30 cmde profondeur. Le tableau 1 montre les ca-ractéristiques chimiques originelles dessols utilisés. On y a planté des bulbes d’en-viron 2 kg du cultivar « Prata Anã » (AAB),considéré comme sensible à la maladie dePanama (Cordeiro et al. 1991).
On a suivi un protocole statistiquecomplètement aléatoire comportant dixtraitements et dix répétitions, chaqueplante représentant une parcelle expéri-mentale. Afin d’évaluer l’effet du Ca seulou combiné avec du Mg, du pH, de la MO,de la stérilisation du sol et de l’additiondu Zn, on a réalisé les traitements sui-vants : 1) Sol organique + chaulage ;2) Sol organique stérilisé + chaulage ;3) Sol minéral stérilisé + chaulage ;4) Sol minéral sans chaulage ; 5) Sol mi-néral + ZnSO4 + chaulage ; 6) Sol minéral+ chaulage ; 7) Sol minéral + chaulageavec CaCO3 (pH d’environ 7,5) ; 8) Sol mi-néral + chaulage avec CaCO3.MgCO3 (pHd’environ 7,5) ; 9) Sol minéral + chaulageavec MgO (pH d’environ 7,5) ; 10) Sol mi-néral + CaSO4.2H2O.
Paramètres chimiques du sol en relation avec l’incidence et lagravité de la maladie de Panama
Maladies Influence de la composition du sol

On a stérilisé les sols minéral et orga-nique par fumigation au bromure de mé-thylène dans la proportion de 340,55 cm3de produit par m3 de sol.
Pour les traitements 1, 2, 3, 5 et 6, on achaulé avec de la chaux dolomitique (Pou-voir Relatif de Neutralisation Total = 99 %)jusqu’à des teneurs de Ca + Mg de 40mmolc/dm3, conformément aux indicationsfournies par l’analyse du sol (Comissáo Es-tadual de Fertilidade do Solo 1989). C’est àdire que, pour les traitements 1 et 2, on aajouté 3,13 t/ha de chaux et pour les traite-ments 3, 5 et 6 : 2,93 t/ha. Le traitement 4représente le témoin puisqu’il ne comportepas de chaulage.
Pour les traitements 7, 8 et 9, les sourcesde Ca et de Mg (respectivement chaux cal-citique, chaux dolomitique et oxyde de ma-gnésium) ont élevé le pH du sol à environ7,5. Conformément aux recommandationsde la méthode d’incubation, on a ajouté9,48 t/ha de chaux calcitique pour le traite-ment 7 ; 7,46 t/ha de chaux dolomitiquepour le traitement 8 et 3,23 t/ha d’oxyde demagnésium pour le traitement 9.
Pour le traitement 10, on a employé13,70 t/ha de plâtre agricole correspondantà 277,12 g de Ca/trou, soit la même quan-tité de Ca ajoutée au traitement 7 sousforme de CaCO3.
Au traitement 5, deux mois après lesemis, au même moment que la premièrefertilisation azotée, on a ajouté19,05 g/trou de sulfate de zinc, équivalent à4,0 g de Zn/plante.
Les plantes ont été fertilisées avec duNPK, à la dose de 100 kg de N/ha, 40 kg deP2O5/ha et de 450 kg de K2O/ha, provenantrespectivement de sulfate d’ammonium, desuper phosphate simple et de chlorure depotassium.
On a analysé les sols selon la méthodolo-gie adoptée par le Serviço Nacional de Le-vantamento e Conservação de Solos -SNLCS (EMBRAPA 1979). Les échantillonsont été prélevés pendant les phases sui-vantes de l’expérimentation : avant l’ouver-
ture des trous, à l’époque de la plantation(deux mois après les corrections chi-miques) et au bout de onze mois de crois-sance végétale.
On a inoculé les plantes quatre moisaprès la plantation avec 6,0 ml d’une sus-pension de spores de Foc, correspondant à8,3 x 107 conidies/trou ou 700 conidies/g desol. L’inoculum avait été réparti dans 100 gd’un mélange de farine de maïs et de sable.On a effectué l’inoculation en ouvrant dessillons profonds de 10 cm autour de chaqueplante et que l’on a recouvert de sol aussi-tôt après avoir répandu l’inoculum (EM-BRAPA 1991).
Sept mois après l’inoculation, on a réa-lisé l’évaluation finale de l’infection en ar-rachant les plantes âgées de onze mois afind’observer le degré d’infection du rhizomesur une série de coupes transversales al-lant de la base vers l’apex et en la notantde 0 à 6, selon l’échelle proposée par Cor-deiro et al. (1993) :0. rizhome complètement indemne ;1. points isolés d’infection ;2. infection sur plus du 1/3 de l’anneau vas-
culaire ;3. infection allant du 1/3 aux 2/3 de l’an-
neau ;4. plus des 2/3 de l’anneau infectés ;5. infection généralisée ;6. plante présentant des symptômes ex-
ternes et/ou visibles sur le pseudotronc. On a fait une analyse de variance, à par-
tir des données obtenues, en appliquant lapreuve de Tukey (P < 0,05) pour comparerles moyennes des différents traitements.On a aussi calculé les coefficients de corré-lation de Pearce.
Résultats et discussion
Effet de la matière organique La teneur élévée en MO du sol organique,comparativement au sol minéral, a une in-fluence sur les niveaux de gravité de la ma-ladie. En effet, les traitements 1 et 2 pré-sentent des indices réduits d’infection parrapport aux autres traitements (figure 1).La forte corrélation (r = 0,61) entre la notemoyenne d’infection par Foc et la matièreorganique (tableau 2) confirme l’effet posi-tif de la MO sur le contrôle de Foc, avec undegré de signification de 1 %. De plus,quand il n’y a pas eu de stérilisation (trai-tement 1), l’utilisation du sol organiqueprésente des différences significatives vis àvis du témoin (traitement 4) pour lapreuve de Tukey (5 %).
Ces résultats démontrent que l’agent pa-thogène provoque des dommages insigni-fiants aux plantes et impliquent que ce solpourrait posséder un caractère suppressifqui se maintiendrait tant que durent lesconditions déterminant sa possible sup-pressivité. Selon Hornby (1983), la majoritédes mécanismes proposés pour expliquer lasuppression de la maladie sur ce type desols n’a pas été prouvée. On suppose que lesfacteurs abiotiques comme le climat, les ca-ractéristiques chimiques (acidité, type d’ar-gile) et physiques (humidité) du sol agis-sent en synergie avec les facteurs biotiques.
Parmi les facteurs biotiques, on supposeque le phénomène d’antagonisme entre lamicrobiosphère originelle du sol et l’agentpathogène serait la cause de la réductionde l’infection. Ce phénomène est courantdans les sols très riches en MO et est im-portant pour l’équilibre microbiologique dusol et le contrôle biologique des orga-nismes pathogènes (Clark 1965, Siqueira etFranco 1988). Le haut pourcentage de MOest généralement en relation avec une po-pulation microbienne élevée (Warcup1965) et augmente l’effet de stase fongiquedu sol, ce qui ainsi rend difficile l’implan-tation d’un microorganisme nouvellementintroduit. Par conséquent, il se produit uneréduction ou un contrôle de la maladie.
Il n’y a pas de différences significativesentre les niveaux d’infection en ce quiconcerne la stérilisation ou non des sols or-ganique (traitements 1 et 2) et minéral(traitement 3 et 6) (figure 1). Selon Hem-wall (1960), des sols de haute teneur enMO sont relativement difficiles à fumiger.Plus tard, Kreutzer (1965) fait remarquerque la MO peut également protéger les mi-
14 INFOMUSA — Vol 9, N° 2
Tableau 1. Caractéristiques chimiques initiales du Latosol alique jaune et du Solorganique (0-30 cm de prof.) utilisés dans l’essai de Cruz das Almas, BA, Brésil.
Caractéristiques Latosol alique jaune Sol organique
pH - H2O 4,80 4,70
P (mg/dm3) 0,90 27,50
K (mmolc/dm3) 0,50 0,90
Ca (mmolc/dm3) 6,50 6,50
Mg (mmolc/dm3) 4,50 2,50
Al (mmolc/dm3) 1,00 1,20
Na (mmolc/dm3) 0,50 0,90
H + Al (mmolc/dm3) 54,00 188,00
CTC (mmolc/dm3) 66,00 198,00
Matière Organique (g/dm3) 21,00 173,00
Mn (mg/dm3) 3,94 1,10
Fe (mg/dm3) 89,05 27,49
Zn (mg/dm3) 3,50 3,17
Cu (mg/dm3) 0,31 0,71
Tableau 2. Coefficients de corrélation entre la matière organique, le pH et les notesd’infection par Foc.
Variables pH Matière organique
Matière organique -0,22*
Note d’infection 0,43** -0,61*** Significatif à 0,05 de probabilité.
** Significatif à 0,01 de probabilité.

croorganismes du sol contre l’action desagents fumigants.
De plus, on sait que, en stérilisant dessols au champ comme dans cette expéri-mentation, il se produit une récupérationrapide de la microbiosphère originelle caril n’y a pas d’effet résiduel dû au bromurede méthylène (Kreutzer 1965, Vanachter1979). Certains microorganismes qui pos-sèdent des structures de survie et résistentà la fumigation, surtout certains champi-gnons, peuvent être capables d’initier lerepeuplement (Kreutzer, 1965).
Effet du pH, du Calcium et du Magnésium L’utilisation de quantités élevées de Caseul, en utilisant du plâtre agricole, sansaltérer le pH initial du sol minéral (traite-ment 10), n’a pas eu d’effet positif pour lecontrôle de la maladie (figure 1). Ceci estconforme aux expérimentations faites parKnudson entre 1923 et 1927 (Stover 1962),où les composés de Ca utilisés n’avaientégalement pas modifié le pH du sol.
On constate que le pH élevé du soldonne lieu à une infection de taux égale-ment élevés. Le chaulage réalisé pour éle-ver le pH aux environs de 7,5 pour les trai-tements 7, 8 et 9 en utilisant des quantitésélevées de, respectivement : Ca, Ca + Mget Mg, n’a pas d’effet positif en ce quiconcerne le contrôle de la maladie de Pa-nama puisque l’on y rencontre des niveauxd’infection plus hauts que pour le témoin(figure 1).
Des résultats semblables ont été obtenusdans certaines régions productrices dumonde (Stover 1962, Blesa Rodríguez etFernandez Caldas 1973, García 1977), où
des sols de pH élevé et de hautes teneursen Ca et Mg étaient affectés par la mala-die. Il y a même eu des cas, comme à Tai-wan (Hwang 1985), pour lesquels les plusforts indices d’attaque étaient relevés dansdes zones où se pratiquait le chaulage(zones de pH supérieur à 7,0). D’autre partpourtant, l’effet positif d’un pH élevé, jointau Ca et au Mg, sur la maladie de Panama,est largement décrit dans la littérature, àtravers les observations et les essais réali-sés par Knudson (1923-1927), Volk (1930 a,b), Volk et Gallatin (1930), Wardlaw(1935), cités par Stover (1962), Scarseth(1945), Rishbeth (1957), Stover et Malo(1972), Alvarez et al. (1981), Malburg et al.(1984) et EMCAPA (1988).
Pour les traitements 7, 8 et 9, les fortesdoses de Ca et de Mg appliquées, seuls ouen combinaison, associées à un pH élevé,provoquent probablement un déséquilibredu sol minéral en influençant non seule-ment ses caractéristiques chimiques maisaussi sa microbiosphère originelle dontsont ainsi éliminés des microorganismesimportants pour la compétition avecl’agent pathogène. Ce déséquilibre, joint àla sensibilité du cultivar utilisé, peut avoircontribué au fait que les plantes aient pré-senté de plus forts taux d’infection.
Pour le traitement 4 (figure 1), l’équi-libre des caractéristiques du sol n’est pra-tiquement pas altéré et ceci peut être unedes raisons de l’incidence mineure de lamaladie. Outre cela, les mauvaises condi-tions de fertilité du sol de ce traitementpeuvent avoir provoqué un moindre déve-loppement des racines ce qui a entraîné unindice réduit d’infection. Car la pénétra-tion de l’agent pathogène dans la plante sefait par les racines secondaires et ter-tiaires (Stover 1962). Et, plus le systèmeradiculaire est développé, plus il y a dechances qu’il y ait de nouvelles infectionsqui s’y installent.
Effet du ZincIl n’y a pas de différences significatives ence qui concerne les niveaux d’infectionquand on ajoute du Zn au sol minéral(traitement 5) et que l’on compare avecles traitements 4 (témoin) et 6 (traite-ment sans Zn, mais avec chaulage)(figure 1). Il est probable que le cultivar« Prata Anã, » en raison de son comporte-ment sensible, ne soit pas efficace pourdévelopper dans le temps et dans l’espacedes mécanismes de résistance opérants(Beckman et al. 1962, Borges Perez et al.1983, Beckman 1990). Ou peut être auraitil été nécessaire d’ajouter des quantitésplus élevées de cet élément nutritifcomme cela s’est fait aux îles Canaries sur« Dwarf Cavendish » (Borges Perez et al.1991). Une troisième hypothèse serait quele Zn n’a que peu ou aucun effet sur lecontrôle de la maladie de Panama,contrairement à ce que l’on supposait.
Conclusions• Le sol organique s’est révélé être un sub-
strat suppressif quant au développementde la maladie de Panama sur le bananier« Prata Anã » ;
• Le chaulage réalisé pour élever le pH dusol à environ 7,5 en utilisant Ca et Mg,aussi bien que l’addition de quantitésélevées de Ca seul sans modifier le pHdu sol, n’ont eu aucun effet positif pourle contrôle le la maladie de Panama. ■
RemerciementsÀ Ana Lucia Borges et Aristóteles Pires deMatos, chercheurs à la Embrapa Mandiocae Fruticultura, pour leurs suggestions.
Références Alexander M. 1961. Introduction to soil microbio-
logy. John Wiley & Sons, New York. 472pp.Alvarez C.E., V. García, J. Robles & A. Díaz. 1981.
Influence des caractéristiques du sol sur l’inci-dence de la maladie de Panama. Fruits 36(2):71-81.
Beckman C.H. 1990. Host responses to the patho-gen. Pp. 93-105 in Fusarium wilt of banana (R.C.Ploetz, ed.). The American Phytopathological So-ciety, Saint Paul, Minnesota.
Beckman C.H., S. Halmos & M.E. Mace. 1962. Theinteraction of host, pathogen, and soil tempera-ture in relation to susceptibility to Fusarium wiltof bananas. Phytopathology 52(2): 134-140.
Blesa Rodríguez A.C. & E. Fernandez Caldas. 1973.Enfermedad de Panamá. Anales de Edafología yAgrobiología 32(3-4): 232-259.
Borges Perez A., I. Trujillo Jacinto del Castillo,F. Gutierrez Jerez & D. Angulo Rodríguez. 1983.Estudio sobre el mal de Panamá en las Islas Ca-narias. II. Influencia de los desequilibrios nutri-tivos P-Zn y K-Mg del suelo en la alteración delos mecanismos de resistencia de la platanera(Cavendish enana) al mal de Panamá. Fruits38(11): 755-758.
Borges Perez A., M. Fernández Falcón, J.J. BravoRodríguez, J.F. Pérez Francés & I. López-Car-reño. 1991. Enhanced resistance of bananaplants (Dwarf Cavendish) to Fusarium oxyspo-rum f.sp. cubense by controlled Zn nutritionunder field conditions. Banana Newsletter 14:24-26.
Clark F.E. 1965. The concept of competition in mi-crobial ecology. Pp. 339-347 in Ecology of soil-borne plant pathogens: prelude to biologicalcontrol (K.F. Baker & W.C. Snyder, eds.). JohnMurray, London, UK.
Comissão Estadual de Fertilidade do Solo. 1989. Ma-nual de adubação e calagem para o Estado daBahia. 2nd edition. CEPLAC/EMATERBA/EMBRAPA/EPABA/NITROFÉRTIL, Salvador, Brasil.
Cordeiro Z.J.M., K. Shepherd, W. dos S. Soares Filho& J.L.L. Dantas. 1991. Reação de cultivares eclones de banana ao mal-do-panamá. RevistaBrasileira de Fruticultura 13(4): 197-203.
Cordeiro Z.J.M., K. Shepherd & J.L.L. Dantas. 1993.Rating bananas for reaction to Fusarium wilt inBrazil. Pp. 84-88 in International symposium onrecent developments in banana cultivation tech-
INFOMUSA — Vol 9, N° 2 15
Figure 1. Evaluation de l’infection par le Foc surle bananier “Prata Anã” soumis à différentstraitements. Cruz das Almas, Brésil, 1992-1993.(Des moyennes suivies par la même lettre nediffèrent pas significativement entre elles, auniveau de 5 % par la preuve de Tukey).
Traitements1. S.O. + chaulage2. S.O. stérilisé + chaulage3. S.M. stérilisé + chaulage4. S.M. sans chaulage5. S.M. + ZnSO4 + chaulage6. S.M. + chaulage7. S.M. + chaulage avec CaCO3. (pH = 7,5)8. S.M. + chaulage avec CaCO3. MgCO3(pH = 7,5)9. S.M. + chaulage avec MgO (pH = 7,5)10. S.M. + CaSO4.2H2O
d cd
a
bc
abab
aa
a
ab
No
tes
d'in
fect
ion
s
4,54
3,53
2,52
1,51
0,50
1 2 3 4 5 6 7 8 9 10
0,80,9
1,5
2,6
2,9
3,8
4,14,4
2,73,9

nology. INIBAP/ASPNET, Los Baños, Laguna,Philippines.
EMBRAPA. Centro Nacional de Pesquisa de Man-dioca e Fruticultura Tropical. 1987. Relatório téc-nico do Centro Nacional de Pesquisa de Mandiocae Fruticultura – 1986. Cruz das Almas, Brasil.
EMBRAPA. Centro Nacional de Pesquisa de Man-dioca e Fruticultura. 1991. Relatório técnico doCentro Nacional de Pesquisa de Mandioca e Fru-ticultura Tropical – 1987/1990. Cruz das Almas,Brasil.
EMBRAPA. Serviço Nacional de Levantamento eConservação de Solos (SNLCS). 1979. Manual demétodos de análise de solo. Rio de Janeiro.
EMCAPA. Empresa Capixaba de Pesquisa Agrope-cuária. 1988. Relatório técnico anual da EmpresaCapixaba de Pesquisa Agropecuária – 1987. Vitó-ria, Brasil.
García V. 1997. État actuel des études de nutritionet fertilité en culture bananière à Ténérife.Fruits 32(1): 15-23.
Gutierrez Jerez F., I. Trujillo Jacinto Del Castillo &A. Borges Pérez. 1983. Estudio sobre el mal depanamá en las Islas Canarias. I. Característicasfísicas y químicas de los suelos y su relación conla aparición de la enfermedad. Fruits 38(10):677-682.
Hemwall J.B. 1960. Theoretical considerations ofseveral factors influencing the effectivity of soilfumigants under field conditions. Soil Science90(3): 157-168.
Hornby D. 1983. Suppressive soils. Annual Review ofPhytopathology 21: 65-85.
Hwang S.C. 1985. Ecology and control of fusarialwilt of banana. Plant Protection Bulletin 27(3):233-245.
Kreutzer W.A. 1965. The reinfestation of treatedsoil. Pp. 495-508 in Ecology of soil-borne plantpathogens: prelude to biological control (K.F.Baker & W.C. Snyder, eds.). John Murray, Lon-don, UK.
Malburg J.L., L.A. Lichtemberg, J.T. dos Anjos &A.A.A. Uberti. 1984. Levantamento do estado nu-tricional dos bananais catarinenses. Pp. 256-275in Anais do 7° Congresso Brasileiro de Fruticul-tura. Sociedade Brasileira de Fruticultura, Flo-rianópolis, Brasil.
Rishbeth J. 1957. Fusarium wilt of bananas in Ja-maica. II. Some aspects of host-parasite relation-ships. Annals of Botany 21(82): 215-245.
Scarseth G.D. 1945. Reported control of Panama di-sease (abst.). Review of Applied Mycology 24: 198.
Siqueira J.O. &, A.A. Franco. 1988. Biotecnologia dosolo. MEC-ABEAS, Brasília, Brasil/ESAL/FAEPE,Lavras, Brasil.
Stover R.H. 1962. Fusarial wilt (Panama disease) ofbananas and other Musa species. Common-wealth Mycological Institute, Kew, England.
Stover R.H. & S.E. Malo. 1972. The occurrence of fu-sarial wilt in normally resistant ‘Dwarf Caven-dish’ banana. Plant Disease Reporter 56(11):1000-1003.
Su H.J., S.C. Hwang & W.H. Ko. 1986. Fusarial wilt ofCavendish bananas in Taiwan. Plant Disease70(9): 814-818.
Sun S.K. & V.W. Huang. 1985. Formulated soilamendment for controlling Fusarium wilt andother soil-borne diseases. Plant Disease 69(11):917-920.
Trujillo Jacinto del Castillo I., F. Gutierrez Jerez &A. Borges Pérez. 1983. Estudio sobre el mal dePanamá en las Islas Canarias. III. Relacionesentre parámetros físicos y químicos de los suelos.Fruits 38(11): 759-764.
Tu C.C. & Y.H. Cheng. 1982. Soil microbial activityin relation to Fusarium wilt-suppressive soil andconducive soil. Tainan DAIS Scientific MeetingReport 1981, Taiwan.
Vanachter A. 1979. Fumigation against fungi. Pp.163-183 in Soil disinfestation (D. Mulder, ed.).Elsevier, Amsterdam.
Warcup J.H. 1965. Growth and reproduction of soilmicrorganisms in relation to substrate. Pp. 509-518 in Ecology of soil-borne plant pathogens:prelude to biological control (K.F. Baker & W.C.Snyder, eds.). John Murray, London, UK.
Josué Francisco da Silva Junior est chercheur à laEmpresa Pernambucana de Pesquisa Agropecuária(IPA), Caixa Postal 1022, CEP 50761-000, Recife, PE,Brasil. Courrier électronique : [email protected] ; ZiltonJosé Maciel Cordeiro et Arlene Maria Gomes Oli-veira travaillent comme chercheurs à la Embrapa Man-dioca e Fruticultura, Caixa Postal 007, 44380-000,Cruz das Almas, BA, Brasil. Courrier électronique : [email protected] ; [email protected].
16 INFOMUSA — Vol 9, N° 2
Claudia Milena Cardona Torres,Gustavo Adolfo Yepes González
et Jairo Castaño Zapata
L’industrie des Musacées (bananes etbananes plantain) est l’une des acti-vités agricoles qui a le plus fort im-
pact social dans les régions où elle est dé-veloppée. La maladie des raies noires oucercosporiose noire (Mycosphaerella fijiensis) est le principal problème phyto-sanitaire qui menace la production de cesressources à la fois alimentaire et finan-cière (Jacome 1998). La maladie de Siga-toka ou cercosporiose jaune (M. musicola)est en train d’être remplacée peu à peu parla cercosporiose noire sur des aires de cul-ture étendues ; or cette dernière peut pro-
voquer jusqu’à 50 % de réduction de la pro-duction, ce qui affecte sensiblement les re-venus du producteur (Burt et al. 1997).D’après les derniers relevés épidémiolo-giques, la cercosporiose noire peut toucherles cultures depuis le niveau de la merjusqu’à 1 940 m d’altitude. On avait cru parailleurs possible que son incidence dansles zones situées à plus de 1 000 m ne soitque de peu d’importance ; cependant lesobservations effectués en Colombie ont dé-montré le contraire (Aguirre et al. 1998).
Le contrôle chimique et la sélection deplantes résistantes continuent d’être parexcellence l’unique stratégie pour com-battre la cercosporiose noire. Les cultivarspossédant un quelconque degré de tolé-rance génétique pourraient ne pas paraîtreprioritaires pour les grands producteurs
alors que pour les petits agriculteurs, lesclones résistants ou tolérants sont lemoyen de contrôle le mieux adapté à leurformation technique et au contexte socioé-conomique de la culture (Wesseling et al.1996, cités par Riveros et Lepoivre 1998).
Le travail de recherche présenté ici a eucomme principal objectif d’évaluer l’effet dela mutagenèse provoquée par du 60Co sur letype de résistance ou de tolérance quepourrait développer le cultivar « Dominicohartón » vis à vis de la cercosporiose noire(M. fijiensis) ou jaune (M. musicola).
Matériel et méthodesLe projet a été mis en place dans le dépar-tement de Tolima, à 7 km de la localité deFresno, sur la route Manizales (Caldas) –Mariquita, sentier de La Ceiba, sur la plan-
Sévérité de la cercosporiose noire (Mycosphaerellafijiensis Morelet) et de la cercosporiose jaune(Mycosphaerella musicola Leach) sur Musa AAB cv. « Dominico hartón » soumis à irradiation par 60Co
Amélioration 60CO, un agent mutagène

tation de Campoalegre, située à 1 250 md’altitude, présentant des températuresmoyennes comprises entre 18 et 25 °C, unehumidité relative de 65 à 100 % et des pré-cipitations annuelles de 1 800 mm. La zonea une topographie de versant et on y dé-tecte la présence des cercosporioses noireet jaune. On a planté 1 600 plants du clone« Dominico hartón », provenant de culturein vitro, qui avaient été soumis à une irra-diation au 60Co à une dose de 25 gray àl’INEA (Bogotá) et qui, multipliés au labo-ratoire de culture de tissus de la Facultédes Sciences Agronomiques, ont été en-suite sevrés à la Granja Montelindo, pro-priété de l’Université de Caldas et enfin,transportés sur le site définitif de planta-tion. Les clones issus de mutagenèseétaient entourés de plants non irradiés dumême clone (témoins) afin d’obtenir uneplus grande pression d’inoculum. Les éva-luations ont été réalisées du 24 mai 1998au 10 avril 1999. Toutes les semaines, on aévalué 58 clones et 5 témoins, chaqueclone étant considéré comme une entitéexpérimentale, pour les paramètres va-riables suivants :
Paramètres de l’hôte : Nombre defeuilles fonctionnelles sur le pseudotronc,date de l’émission de l’inflorescence etpoids du régime (kg).
Paramètres de la maladie : Plus jeunefeuille nécrosée (PJFN) en considérant lestaches de stade 4 pour la cercosporiosejaune et de stade 5 pour la cercosporiosenoire ; sévérité des attaques des maladiesselon l’échelle de Stover modifiée par
Gauhl (1984) et indice de l’infection desmaladies.
Échelle de sévérité des attaques des cercosporioses jaune et noire :0. Feuille saine, sans symptômes1. Jusqu’à 5 % de surface foliaire affectée2. 6 à 15 % de surface foliaire affectée3. 16 à 33 % de surface foliaire affectée4. 34 à 50 % de surface foliaire affectée5. 51 à 100 % de surface foliaire affectée
Indice de sévérité, calculé grâce à la for-mule de Towsend et Heuberguer, selon Un-terstenhuefer (1963), cité par Orjeda et al.(1998) :où :
IS = indice de sévéritén = nombre de feuilles de chaque stadeb = stadeN = nombre de stades employés dansl’échelleT = nombre total de feuilles évaluées.
Indice de la plus jeune feuille nécrosée,calculé en utilisant la formule suivante :où :
IPJFN = indice de la plus jeune feuille né-croséePJFN = plus jeune feuille nécrosée au mo-
ment de l’évaluationT = nombre total de feuilles évaluées.
Paramètres du climat : température(maximum, minimum et moyenne), humi-dité relative et précipitations.
Les données ont été entrées, à partird’un protocole complètement aléatoire enfonction du temps, dans un programmeSAS (System Analysis Statistical) et ana-lysées grâce aux estimations obtenues parrégressions, intervalles de confiance, ana-lyses de variance et procédés descriptifs.
Résultats et discussionLes conditions climatiques de la zoned’étude ont consisté en une températuremoyenne de 21,5 °C, une humidité relativede 81 % et un cumul des précipitations de340,5 mm. En analysant ces conditions cli-matiques en détail et en relation avec ledéveloppement des maladies, on remarqueque les précipitations sont un paramètredéterminant pour l’apparition de l’infec-tion. La température et l’humidité relativependant l’étude ont, elles, favorisé le déve-loppement de l’épidémie en sachant quedes températures de 20 à 35 °C permettentla germination des ascospores et des coni-dies et que cette germination est maxi-mum pour des températures de l’ordre de25 à 28 °C en présence d’une forte humi-dité relative, en particulier lorsqu’il existeune pellicule humide sur la feuille (Ja-come et Schuh 1992). Les maladies possè-dent une dynamique saisonnière détermi-née par les variations de température et deprécipitations tout au long de l’année étantdonné que les structures reproductrices sedéveloppent par inoculation croisée, pro-cessus facilité quand il y a de l’eau libre surles feuilles (Mourichon et Zapater 1990).
La courbe de développement des mala-dies présente deux maxima atteints aux297e et 477e jours après plantation, datesqui coïncident avec des périodes de préci-pitations élevées (figure 1). Au bout de 297jours après plantation, la culture en est en-core à la phase végétative mais très prochede l’époque de la floraison, d’où l’impor-tance de conserver un bon nombre defeuilles. En effet, comme le rapportentOrtiz et Vuylsteke (1994), cités par Crae-nen (1998), il faut un minimum de huitfeuilles fonctionnelles pendant tout lecycle pour obtenir un bon rendement. Deplus, des plantes qui possèdent moins dehuit feuilles sans taches avant la floraisonsont répertoriées comme sensibles à la cer-cosporiose noire. Au bout de 477 joursaprès plantation, les clones en sont austade du développement du régime. C’estla raison pour laquelle, à la fin de l’expéri-mentation, on n’a pas obtenu de meilleurspoids de régime puisque 80 % des sujetsévalués présentaient leur plus jeune feuillenécrosée (PJFN) en position 1 et avec deforts indices de sévérité de la maladie, at-teignants jusqu’à 100 %.
Seuls 45 % des clones évalués, y comprisles témoins, parviennent au stade de
INFOMUSA — Vol 9, N° 2 17
Figure 1. Comportement des cercosporioses jaune et noire en fonction des conditions climatiques(mai 1998 - avril 1999).
Préc
ipit
atio
n (
mm
)Sé
véri
té
Tem
pér
atu
re (
°C)
et h
um
idit
ére
lati
ve (
%)
0
50
100
150
200
250
300
350
400
100
80
60
40
20
0207 237 267 297 327 357 387 417 447 477 507
010
20
30
40
50
60
7080
90
Jours après plantation
Sévérité maximum Température moyenne Humidité relative

l’émission florale avec plus de huit feuillesfonctionnelles. Parmi eux se distinguent,pour avoir conservé le plus grand nombrede feuilles fonctionnelles (11), les clones 4,17, 18, 21, 38 et 44 ; les autres ont fini leurcycle avec un nombre inférieur à onzefeuilles car la plupart d’entre elles étaienttotalement affectées par les maladies.
Au début de l’expérimentation, les ni-veaux les plus élevés de sévérité sont obser-vés sur les feuilles inférieures des plantes.Au fil du temps, la maladie gagne lesfeuilles supérieures indiquant que la sévé-rité de la maladie est directement en rela-tion avec le taux d’émission foliaire.
À l’époque de l’apparition de l’inflores-cence, vers 342 jours après plantation, lesfeuilles inférieures, et surtout celles en po-sition 11, 12, 13 et 14, présentaient les plusforts indices de sévérité de la maladie. Ceciest dû à la taille des plantes et à l’existenced’une plus grande surface foliaire ce qui en-traîne donc un plus grand risque de récep-tion de l’inoculum d’une future infection. Ala fin de l’étude, 50 % des clones avaientmoins de quatre feuilles fonctionnelles.Parmi ceux-ci, au moment de la récolte, lessujets 8, 15, 16, 41 et les témoins n’avaientplus qu’une seule feuille, au stade 5 de sé-vérité de la maladie (>50 %), sévérité étroi-tement liée aux conditions climatiques opé-rantes durant cette période.
En général, on n’a pas décelé de clonesprometteurs quant à leur résistance aux
cercosporioses jaune ou noire. Sur le ta-bleau 1, sont réunies les informationsconcernant la sévérité des maladies rela-tives à six clones ayant présenté les indicesles plus bas pour les positions 6, 7, 8 et 9de la feuille. La sévérité finale se situe au-dessus de 40 % pour les clones irradiés etne montre pas de différences significativespar rapport au témoin. Parmi ces sujets, leclone 57 a présenté le plus grand pourcen-tage de sévérité finale de la maladie :70,48 % sur la feuille en position 6. Aprèsl’émission de l’inflorescence, la grande ma-jorité des clones avaient de forts indices desévérité de la maladie. Parmi eux, ce sontles clones 9 et 57 qui présentent la sévéritémaximum (100 %) sur la feuille en position9 ; et ce, pendant que la sévérité moyenneaugmentait sur les plus vieilles feuilles deposition 8 et 9. Conformément à Belalcázar
et al. (1994), l’analyse des pourcentagesenregistrés pour la sévérité maximum de lamaladie permet d’établir clairement si lesclones sont résistants ou sensibles, toutparticulièrement en évaluant la feuille enposition 9.
Dans ce travail, les clones se classentdonc comme « hautement sensibles ».
Tant l’indice de sévérité que celui de laplus jeune feuille nécrosée ont eu un com-portement très hétérogène à l’époque del’émission de l’inflorescence (342-432 joursaprès la plantation), époque à laquelle ona observé la plus grande variation en fonc-tion du temps. A partir de ce moment, lesplantes ont cessé d’émettre des feuillespour commencer le processus de transloca-tion des nutriments vers l’inflorescence.Par conséquent, les feuilles se sont affai-blies et sont devenues plus sensibles auxmaladies. Ceci est renforcé par des condi-tions climatiques (température de 21 ºC,humidité relative de 75 % et précipitationsaccumulées de 100 mm) favorables à leurdéveloppement et permettant leur dissémi-nation sur toute la surface foliaire desplantes. Tout cela, ajouté au fait de ne pasconserver le nombre optimum de feuillesnécessaires pour la production d’un régimeconvenable (8 feuilles au minimum), s’esttraduit par des régimes de poids et de qua-lité fortement diminués (moins de 12 kg enmoyenne) (tableau 2).
On montre sur ce même tableau le com-portement des clones les plus résistantsaux cercosporioses et son influence sur leparamètre production. Chez ces sujets, ilest apparu quelques différences entre lesparamètres épidémiologiques, lesquels ontfinalement des répercussions sur le rende-ment de la culture. De façon générale, iln’y a pas de différences significatives entreeux et le témoin quant à la variable de laplus jeune feuille nécrosée (PJFN). Les su-jets 5, 16, 42, 68, 81 et 107 ont les plus fortsindices de sévérité de la maladie pendantl’étude.
ConclusionLes résultats présentés dans cette publi-cation indiquent que les mutagenèses,provoquées pour cette étude par l’irradia-tion au 60Co, n’ont conféré au cultivar« Dominico hartón » aucun type de tolé-
18 INFOMUSA — Vol 9, N° 2
Tableau 1. Relation entre les pourcentages de sévérité finale des cercosporiosesnoire ou jaune sur les clones présentant la plus petite sévérité moyenne en prenanten compte la position de la feuille après formation de l’inflorescence.
Clone* Position de la feuille Sévérité finale (%)
9 6 44,44
7 48,64
8 66,38
9 100,00
15 6 52,42
7 48,69
8 52,98
9 63,41
39 6 47,84
7 49,58
8 47,26
9 46,98
57 6 70,48
7 46,18
8 54,49
9 100,00
59 6 48,03
7 48,98
8 46,95
9 51,84
82 6 47,66
7 44,32
8 40,95
9 43,33
Témoin absolu 6 48,24
7 48,50
8 47,79
9 45,96* Clones récoltés à des dates différentes.
Tableau 2. Comportement de six clones de bananier plantain « Dominico hartón »irradiés, par rapport au cycle, à la plus jeune feuille nécrosée, au nombre de feuillesau moment de l’évaluation et au poids du régime (kg).
Clone Cycle (jours après plantation) PJFN NFME Poids du régime (kg)
9 528 5 8 8,00
15 493 5 10 12,50
39 528 4 10 5,60
57 528 4 8 8,70
59 493 5 10 8,00
82 473 4 10 10,00
Témoin 486 5 9 10,60PJFN = plus jeune feuille nécrosée.
NFME = nombre de feuilles au moment de l’évaluation.

rance vis à vis des attaques des cercospo-riose jaune et noire.
Des études réalisées par Cervantes(1997) indiquent que les irradiations au60Co de niveaux ou de degrés différents ontinduit chez le bananier Williams des muta-tions qui ont des répercussions négativessur les qualités phénotypiques de la planteet du fruit, rendant leur commercialisationimpossible. Cela diminue quelque peu l’es-poir d’obtenir une amélioration génétiquepar le biais de cet agent mutagène.
RemerciementsLes auteurs remercient l’Agence Interna-tionale de l’Énergie Atomique, AIEA,Vienne, Autriche et COLCIENCIAS, Colom-bie, pour l’aide apportée à la réalisation decette recherche menée dans le cadre del’Unité de Recherche sur Bananier, du Dé-partement de Phytotechnologie de la Fa-culté des Sciences Agronomiques de l’Uni-versité de Caldas, Colombie. ■
RéférencesAguirre M.C., J. Castaño-Zapata, J.A. Valencia, L.E.
Zuluaga & C. Arce. 1998. Interacción de Mycos-phaerella fijiensis Morelet y M. musicola Leachen siete genotipos de Musa sp., en un área límitede expansión de la Sigatoka negra en la zona ca-fetera colombiana. Pp. 192-220 in Memorias delSeminario Internacional sobre Producción dePlátano (M.J. Giraldo, S.L. Belalcázar, D.G. Cayóny R.G. Botero, eds). Universidad del Quindío ; Co-mité de Cafeteros del Quindío ; SENA ; INIBAP ;CORPOICA.
Belalcázar S., F. Salazar, J.A. Valencia, C.H. Silva,M.I. Arcila & R. Jaramillo. 1994. Reacción de va-riedades mejoradas al ataque de Sigatoka negra(Mycosphaerella fijiensis Morelet). Pp. 192-214in Mejoramiento de la Producción del Cultivo delPlátano. ICA-CORPOICA ; CIID ; INIBAP ; INPO-FOS ; CCQ.
Burt J. A., J. Rutter & H. Gonzalez. 1997. Short dis-tance wind dispersal of the fungal pathogenscausing Sigatoka diseases in banana and plan-tain. Plant Pathology 46(4): 451–458.
Cervantes A. & A. Rodríguez. 1997. Algunas carac-terísticas fenotípicas de las mutaciones induci-das en banano Williams (Musa AAA) por las irra-diaciones de Cobalto 60 in Memorias V Congresode la Sociedad Colombiana de Fitomejoramientoy Producción de Cultivos. Santa Marta. 109 pp.
Craenen K. 1998. Black Sigatoka disease of bananaand plantain: a Reference Manual. IITA, Ibadan,Nigeria.
Gauhl F. 1994. Epidemiolgy and ecology of black Si-gatoka (Mycosphaerella fijiensis Morelet) onplantain and banana in Costa Rica. INIBAP,Montpellier, France. 120 pp.
Jácome L. 1998. Sigatoka negra, la situación enAmérica Latina y el Caribe. Pp. 18-23 in Memo-rias Primer Simposio Internacional sobre Siga-toka Negra (M.M. Robles et al., compil.). Manza-nillo, Colima, Mexico, 8-10 de Julio 1998. SAGAR ;INIFAP ; INIBAP ; Universidad de Colima ; IICA.
Jacome L. H. & W Schuh. 1992. Effects of leaf wet-ness duration and temperature on development
Alfonso Vargas et Fabio Blanco
L’élimination sélective de mains estpratiquée dans les systèmes de pro-duction de bananes d’exportation
(Musa AAA). Pourtant, les informations deréférence sont rares, contradictoires etfragmentaires. A ce propos, l’augmentationdes dimensions des fruits résultant de l’éli-mination sélective de mains est mention-née par Irizarry et al. (1989, 1992 et 1994),Johns (1996), Boncato (1969), Calvo y Soto(1984) et Meyer (1975). Cependant, l’aug-mentation de la taille des fruits n’a pascompensé la diminution du poids du ré-gime (Irizarry et al. 1989, 1992 et 1994,Meyer 1985). Cette perte de rendement aété également constatée par Daniels(1987) et s’est produite sans que l’on soitparvenu à améliorer les dimensions du dia-mètre ou de la longueur des fruits.
Des résultats de Vargas et al. (1999)montrent que, dans les évaluations d’élimi-nation sélective, c’est le nombre de mainset leur position sur le régime qui doiventêtre pris en compte avant tout. Dans cetteoptique, ces auteurs n’ont pas trouvé de dif-férences entre les diverses intensités d’éli-mination sélective pour ce qui concerne lesdimensions des fruits provenant de mainscomparables, situées sur la partie supé-rieure ou inférieure de régimes ayant lemême nombre de mains à la floraison.
Les différentes méthodologies utiliséespour évaluer l’élimination sélective pour-raient expliquer les résultats discordants,
relatifs aux dimensions des fruits, que l’onmentionne fréquemment dans la littérature.
Le but du travail présenté est de compa-rer, à l’aide du même ensemble de don-nées, deux méthodologies d’évaluation del’effet de l’élimination sélective de mainssur les régimes de bananes.
Matériel et méthodesLes données ont été collectées sur unezone bananière (Musa AAA, cv. « Valery »)considérée comme hautement productive.Il s’agit d’une plantation âgée de 29 ans, si-tuée sur la côte caraïbe du Costa Rica etayant actuellement une densité de 1 750 plants/ha.
L’élimination sélective a été pratiquéedeux semaines après la sortie de l’inflores-cence et a inclus dans tous les cas la sup-pression du groupe de fleurs à étamines etde la fausse main.
Les traitements réalisés sur des régimesde huit, neuf et dix vraies mains à la florai-son sont les suivants : 1) élimination d’unevraie main et 2) élimination de deux vraiesmains.
On considère comme fausse main un en-semble de fleurs à pistil dont une ou plu-sieurs se développent anormalement(comme une réminiscence de l’ovaire) ; oubien, où il se trouve des fleurs à étamines.On considère comme vraie main un groupede fleurs à pistil dont les gynécées se déve-loppent normalement.
Les traitements d’élimination sélectiveont été faits au hasard sur une aire de cinqhectares, en accord avec l’apparition de
INFOMUSA — Vol 9, N° 2 19
Considérations méthodologiquespour l’évaluation de l’éliminationsélective de mains de bananes(Musa AAA, cv. « Valery »)
Agronomie Nombre de mains et productivité
of black Sigatoka disease on banana infected byMycosphaerella fijiensis var. difformis. Phytopa-thology 82(5): 515-520.
Mourichon X. & M.F. Zapater. 1990. Obtention invitro du stade M. fijiensis, forme parfaite de Cercospora fijiensis. Fruits 45(6): 553–557.
Orjeda G. en collaboration avec les groupes de tra-vail de PROMUSA sur la fusariose et les cercospo-rioses. 1998. Evaluation de la résistance des bananiers aux cercosporioses et à la fusariose.Guides techniques INIBAP 3. INIBAP, Montpel-lier, France.
Riveros A. S. & P. Lepoivre. 1998. Inductores exóge-nos asociados con los mecanismos de defensa a
la Sigatoka negra del banano. Pp. 126-132 in Me-morias Primer Simposio Internacional sobre Siga-toka Negra (M.M. Robles et al., compil.). Manza-nillo, Colima, Mexico, 8-10 de Julio 1998. SAGAR ;INIFAP ; INIBAP ; Universitad de Colima ; IICA.
Claudia Milena Cardona Torres et GustavoAdolfo Yepes González sont étudiants à la Facultadde Ciencias Agropecuarias, Programa Agronomía,Universidad de Caldas. Jairo Castaño Zapata estProfesseur Titulaire du Departamento de Fitotecnía,Facultad de Ciencias Agropecuarias. Universidad deCaldas, A.A. 275, Manizales, Caldas, Colombie.

chacun des types de régimes souhaités. Ona réalisé cinq cycles bimensuels d’élimina-tion : en avril, juin, août, octobre et dé-cembre. Conformément à la programma-tion de la plantation, on a récolté les fruitsdans le courant de chaque onzième se-maine, après l’élimination sélective, pourles trois premiers cycles et dans le courantdes semaines 11 et 12 pour les deux der-niers. On a considéré que chaque régimeétait une répétition. On a évalué 416 ré-gimes.
Les mesures des dimensions des fruitsde chaque régime ont été relevées sur ladeuxième, l’avant-dernière et la dernièremain. Pour les régimes n’ayant pas lemême nombre de vraies mains (régimesnon équivalents), on a comparé les traite-ments au niveau de la dernière main réma-nente (DMR) en plus de la deuxième main.Pour les régimes ayant le même nombre devraies mains (régimes équivalents), on a
effectué la comparaison sur la dernièremain comparable (DMC) en plus de ladeuxième main.
On définit comme DMR la main distaleprésente après avoir pratiqué un des deuxtraitements d’élimination sélective (fi-gure 1). On définit comme DMC la dernièremain distale présente après avoir pratiquéle traitement d’élimination sélective le plusintense : c’est la cinquième sur les régimesà huit mains, la sixième sur ceux de neuf etla septième sur ceux de dix.Les paramètres évalués ont été :a. le poids (kg) du régime.b. le diamètre (1/32”) du fruit central (ran-
gée externe) de la deuxième main, de ladernière main rémanente (DMR) et dela dernière main comparable.
c. la longueur (cm) du fruit central (ran-gée externe) de la deuxième main, de laDMR et de la DMC. La comparaison desdeux méthodologies a été menée en uti-
lisant le même ensemble de données.Pour la première, on a utilisé des ré-gimes non équivalents et la DMR commecomparateur, en plus de la deuxièmemain. Pour la seconde, on a utilisé desgroupes de régimes équivalents et laDMC comme comparateur, en plus de ladeuxième main. Dans les deux cas, on aréalisé une analyse de variance des don-nées afin de comparer les effets des trai-tements.
RésultatsL’analyse de l’ensemble des données où onn’a pas pris en compte le nombre de mainsdu régime et où on a utilisé la DMR, enplus de la deuxième main, comme compa-rateur des dimensions du fruit (tableau 1),montre des différences significatives (P < 0,01) entre les traitements d’élimina-tion sélective pour ce qui concerne le dia-mètre et la longueur du fruit central de laDMR. À mesure que l’intensité de l’élimi-nation augmente, le poids du régime dimi-nue (P < 0,01). Les dimensions du fruitcentral de la deuxième main ne sont pasaffectées (P > 0,36) par les intensités d’éli-mination sélective.
L’analyse de l’ensemble des données oùon a pris en compte le nombre de mains durégime et où on a utilisé la DMC, en plusde la deuxième main, pour évaluer le dia-mètre et la longueur du fruit (tableau 2),ne montre pas de différences significatives(P > 0,40) entre les traitements ; et ce, in-dépendamment de la taille du régime. Enrevanche, quand on fait l’analyse en pre-nant en compte le nombre de mains du ré-gime et la DMR comme comparateur (ta-bleau 2), on peut noter des différencessignificatives (P < 0,02) entre les traite-ments ; et ce, indépendamment de la tailledu régime. Le poids du régime diminue aufur et à mesure que l’intensité de l’élimina-tion augmente pour les régimes de 9 et 10 mains (P < 0,01), mais pas plus pourceux de 8 mains (P > 0,15). Les dimen-sions du fruit central de la seconde maindes régimes de 8, 9 et 10 mains ne se sontpas différenciées (P > 0,09) en fonctiondes intensités d’élimination.
DiscussionUne expérimentation peut être définiecomme « une étude de recherche où l’onmanipule délibérément une ou plusieurs
20 INFOMUSA — Vol 9, N° 2
Figure 1. Représentation schématique de régimes équivalents avec les mesures des dimensions du fruit(diamètre et longueur) sur la dernière main rémanente (DMR) et sur la dernière main comparable (DMC).
Tableau 1. Mesures ± erreur standard du poids du régime, du diamètre et de la longueur du doigt central inférieur de banane(Musa AAA, cv. Valery) de la deuxième et dernière main rémanente (DMR) en fonction de l’intensité de l’élimination sélective (I ES). On a pris en compte la totalité de l’ensemble des régimes comportant 8, 9 et 10 mains à la floraison.
I ES1 n Poids du régime Diamètre du fruit2 Longueur du fruit Diamètre du fruit2 Longueur du fruit (cm)
(kg) 2e main (cm) 2e main DMR DMR
2 191 25,53 ± 0,35 43,26 ± 0,12 25,92 ± 0,11 39,50 ± 0,12 21,70 ± 0,09
3 225 23,84 ± 0,29 43,40 ± 0,12 26,00 ± 0,09 40,48 ± 0,14 22,49 ± 0,09
Prob > F 0,0001 0,3661 0,6507 0,0001 0,00011 Correspond au nombre de mains éliminées.2 Une unité de diamètre = 1/32 de pouce (0,79375 mm).
Régimes équivalents de 10 mains
DMCDMR
Fausse main
Groupe de fleurs à étamines
Vraies mains éliminées

variables indépendantes (causes suppo-sées) afin d’analyser les conséquences quecette manipulation a sur une ou plusieursvariables liées (effets supposés) dans unesituation contrôlée par l’expérimentateur »(Hernández et al. 1998).
Les variables indépendantes (causes)représentent les traitements expérimen-taux, lesquels se réfèrent à des entités ex-périmentales. Les entités expérimentalesattribuées aux traitements doivent êtreéquivalentes en tout (homogènes) ou alorsleur variabilité doit être dûment contrôléedans le cadre du protocole expérimental.
En général, l’effet de l’élimination sélec-tive sur régimes de bananes a fait l’objet derecherches sans considérer le nombre devraies mains présentes lors de la floraison.Dans de telles conditions, il est très pro-bable que des groupes non équivalentsaient reçu des traitements différents, cequi invalide la comparaison à cause del’approximation commise. Les consé-quences négatives sur la qualité de l’expé-rimentation sont encore plus marquéesdans ce cas par l’utilisation de la DMRpour relever les mesures des dimensionsdu fruit (diamètre et longueur). La posi-tion de cette dernière sur le régime dé-pend en effet de la taille de celui-ci et del’intensité de l’élimination sélective. Onsait que la longueur des fruits sur un ré-gime de bananes (cv. « Valery ») décroîtdes premières aux dernières mains (Jara-millo 1982), avec une diminution plus ac-centuée à partir de la cinquième main. Parconséquent, la DMR ne peut procurerl’équivalence des mains nécessaire pourque les différences entre les traitementspuissent être estimées objectivement.C’est pourquoi elle est inadéquate pourcomparer des intensités distinctes d’élimi-nation sélective.
Les sources d’invalidité mentionnées ci-dessus peuvent être éliminées de façon ap-propriée en considérant chaque régimecomme une entité expérimentale, en appli-quant au hasard les traitements au champ,
en comparant des régimes comportant lemême nombre de mains à la floraison et encomparant des mains de position identiquesur le régime (régimes et mains respective-ment équivalents), ce à quoi se sontconformé Vargas et al. (1999).
D’après les résultats obtenus, il n’appa-raît pas que la plus forte intensité d’élimi-nation sélective (suppression de troisvraies mains) provoque, par rapport à l’in-tensité la plus faible (suppression de deuxvraies mains), l’augmentation des dimen-sions des fruits du cultivar « Valery ». Demême, en accord avec l’analyse antérieure,il est confirmé que la diminution du poidsdu régime est une conséquence de l’élimi-nation sélective.
L’analyse conceptuelle différente d’unmême ensemble de données permet d’ex-pliquer la différence des résultats présen-tés par les travaux de recherche s’ap-puyant sur l’un ou l’autre des concepts. Parconséquent, la méthodologie d’évaluationet d’analyse communément employée, fon-dée sur l’utilisation de régimes et de mainsnon équivalents, ne devrait plus l’être pourdes travaux de recherche relatifs à l’élimi-nation de mains sur régimes de bananes.Son usage pourrait amener des résultatserronés comme l’a montré cette étude. Asa place, on doit utiliser la deuxième mé-thode fondée sur l’emploi de régimes et demains équivalents. ■
RéférencesBoncato A. 1969. Effects of reducing the number of
hands in a bunch of Lacatan banana. Journal ofPlant Industry 32(3-4) : 243-251.
Calvo J. & M. Soto. 1984. Efecto del desmane en lacalidad del fruto del banano (Musa AAA, Subgrupo Cavendish ‘Gran enano’). Pp. 128-129 in VICongreso Agronómico Nacional. Colegio de Inge-nieros Agrónomos de Costa Rica. San José, CostaRica.
Daniels J., P. O’Farrel, C. Mulder & S. Campbell.1987. Effects of bunch covering and bunch trim-ming on bananas in North Queensland. Queens-
land Journal of Agricultural and Animal Sciences44(2) : 101-105.
Hernández R., C. Fernández & P. Baptista, P. 1998.Metodología de la Investigacíon. (2da edición).McGraw-Hill Interamerican Editors, S.A. de C.V.México, D.F. 501 pp.
Irizarry H., E. Hernández & J. Rodríguez. 1994.Yield of five dwarf banana cultivars grown withminimun tillage in Puerto Rico’s mountain re-gion. The Journal of Agriculture of the Universityof Puerto Rico 78(1-2) : 1-7.
Irizarry H., E. Hernández & J. Rodríguez. 1992.Bunch and ratoon management for profitableproduction of high quality bananas (Musa acu-minata, AAA). The Journal of Agriculture of theUniversity of Puerto Rico 76(3-4) : 119-129.
Irizarry H., E. Hernández, E. Rivera, D.E. Beu-champ, E. Caloni, & R. Guadalupe. 1989. Perfor-mance of elite bananas (Musa acuminata AAA)cultivars in four locations of Puerto Rico. TheJournal of Agriculture of the University of PuertoRico 73(3) : 209-221.
Jaramillo R. 1982. Las principales característicasmorfológicas del fruto de banano, variedad Ca-vendish Gigante (Musa AAA) en Costa Rica.Unión de Países Exportadores de Banano(UPEB). 42 pp.
Johns G. 1996. Effects of bunch trimming anddouble bunch covering on yield of bananas du-ring winter in New South Wales. Australian Jour-nal of Experimental Agriculture 36 : 229-235.
Meyer J. 1975. Influence de l’ablation de mains surle rendement en poids des régimes de bananespar catégories de conditionnement aux Antilles.Fruits 30(11) : 663-668.
Vargas A, J. Sandoval & F. Blanco. 1999. Efecto dela intensidad de desmane sobre las dimensionesdel fruto y el peso del racimo de banano (MusaAAA, cv. Valery). CORBANA 24(51) : 85-92.
Alfonso Vargas travaille à la Dirección de Investiga-ciones de la Corporación Bananera Nacional (COR-BANA) Apdo. 390-7210, Guápiles, Costa Rica. FabioA. Blanco travaille à la Escuela de Ciencias Agrarias,Universidad Nacional, Apdo. 86-300, Heredia,Costa Rica.
INFOMUSA — Vol 9, N° 2 21
Tableau 2. Mesures ± erreur standard du poids du régime, du diamètre et de la longueur du fruit central inférieur de bananier(Musa AAA, cv. Valery), de la deuxième et dernière main comparable (DMC) en fonction de l’intensité de l’élimination sélective(IES) et du nombre de mains totales (MT) du régime. DMR = 6e et 5e si MT = 8 ; 7e et 6e si MT = 9 ; 8e et 7e si MT = 10 ; DMC = 5esi MT = 8, 6e si MT = 9 et 7e si MT = 10.
MT n IES Poids régime Diamètre1 Longueur Diamètre1 Longueur Diamètre1 Longueur DMC
(kg) 2e main 2e main (cm) DMR DMR (cm) DMC (cm)
8 40 2 20,41 ± 0,47 43,19 ± 0,22 25,71 ± 0,22 39,60 ± 0,26 21,92 ± 1,18 41,01 ± 0,21 23,00 ± 0,17
51 3 19,43 ± 0,47 43,47 ± 0,25 25,63 ± 0,19 41,34 ± 0,42 23,21 ± 0,18 41,34 ± 0,43 23,21 ± 0,18
Prob > F 0,1510 0,4072 0,7710 0,0016 0,0000 0,5250 0,4033
9 73 2 24,24 ± 0,39 43,11 ± 0,18 26,64 ± 0,18 39,56 ± 0,18 21,68 ± 0,16 40,46 ± 0,16 22,55 ± 0,19
89 3 22,90 ± 0,28 43,48 ± 0,20 26,02 ± 0,14 40,46 ± 0,19 22,44 ± 0,14 40,46 ± 0,19 22,44 ± 0,14
Prob > F 0,0063 0,1916 0,0888 0,0001 0,0004 0,9945 0,6511
10 78 2 29,37 ± 0,42 43,44 ± 0,19 26,29 ± 0,18 39,38 ± 0,20 21,59 ± 0,14 39,97 ± 0,17 22,14 ± 0,15
85 3 27,49 ± 0,34 43,28 ± 0,20 26,18 ± 0,16 39,99 ± 0,17 22,10 ± 0,15 39,99 ± 0,17 22,10 ± 0,15
Prob > F 0,0006 0,5558 0,6606 0,0206 0,0138 0,9770 0,87021 Une unité de diamètre = 1/32 de pouce.

K. Nowakunda, P.R. Rubaihayo, M.A. Ameny et W. Tushemereirwe
En Ouganda, la productivité des bana-niers décline depuis les années 1970(Hartmanns 1989), principalement
du fait de la baisse de fertilité des sols, del’augmentation de l’incidence des rava-geurs et des maladies, d’une diversité trèslimitée du matériel génétique et denombre de contraintes socioéconomiques(Rubaihayo et Gold 1991). Toutes les ba-nanes à cuire AAA d’Afrique de l’Est (AAA-EA) sont sensibles à la cercosporiosenoire, aux charançons et aux nématodes(Tushemereirwe 1996).
L’utilisation de cultivars résistants auxmaladies et ravageurs est apparue commel’approche la plus réalisable pour solution-ner ce problème (Ortiz et Vuylsteke 1998).En conséquence, un bon nombre d’hy-brides de bananiers et de cultivars locauxrésistants ou tolérants principalement à lacercosporiose noire et/ou à la fusariose ontété introduits en Ouganda.
Les caractéristiques physiques, chi-miques et sensorielles de ces cultivars ontété déterminées pour établir leur profil dequalité. Les attributs agronomiques inté-ressant les utilisateurs finaux de bananessont la période de maturité et l’obtentionde la qualité de consommation à un stagede développement précoce.
Cet article présente les résultats de testsportant sur le degré d’acceptabilité de 14 cultivars de bananiers et bananiersplantain introduits en Ouganda pour lacuisson et l’extraction de jus.
Matériels et méthodesLes caractéristiques de 14 cultivars etcelles de bananes d’altitude d’Afrique del’Est utilisés dans cette étude sont présen-tées dans le tableau 1. Les cultivars locauxles plus communs sont Mbwazirume, lesbananes à cuire Kisansa (AAA-EA), Ndizi(AB), Musa-Kayinja (ABB) et la banane àbière Entundu (AAA-EA) ; ils ont été utili-sés comme témoins dans un dispositif ex-périmental en blocs en randomisation to-tale.
La période de remplissage de la bananea été considérée comme celle allant del‘émission de l’inflorescence au jour où aumoins un doigt manifeste des signes de ma-turation, jour où le régime a été récolté(Palmer 1971). Le régime a été pesé (en
kg) à la récolte en utilisant une balance(BFK-265-030B, 60, Fisher, UK). Le poidsdu doigt a été déterminé en pesant sur unebalance électronique Mettler (P1200, Fi-sher, UK) cinq doigts prélevés à partir dudeuxième doigt du régime. Les mêmesdoigts ont été utilisés pour déterminer lalongueur moyenne du doigt, sa circonfé-rence ainsi que le poids de la peau et de lapulpe. Avant d’être pesées, la peau et lapulpe ont été séparées manuellement avecun couteau inoxydable. La période pendantlaquelle le fruit reste vert a été déterminéepar la différence en nombre de jours entrela récolte et le moment où la deuxièmemain montre les premiers signes de matu-ration (passage du vert sombre au vertclair) (Dadzie et Orchard 1997). Les échantillons utilisés pour vérifier la pé-riode pendant laquelle le fruit reste vertont été récoltés une semaine avant la pé-riode de maturation physiologique escomp-tée telle que définie par Palmer (1971).Les teneurs en tanin ont été déterminéesen utilisant le titrage à base de vanillinetel que décrit par Broadhurst et Jones(1978).
Les caractéristiques sensorielles et l’ac-ceptabilité des bananes ont été détermi-nées en faisant bouillir les bananes éplu-chées dans un litre d’eau. Six grammes de
sel de table ont été ajoutés aux bananes,lesquelles ont été ensuite bouillies jusqu’àcuisson complète. Cette méthode de prépa-ration a été préférée à la méthode tradi-tionnelle de cuisson consistant à cuire lesbananes à l’étuvée avant de les écraser enpurée (muwumbo) pour éviter d’éven-tuelles erreurs dues aux préjugés de pré-sentation.
Quinze panélistes ont été choisis surbase de leur jugement de solutions diluéesdes goûts de base : sucré (sucrose à 8 g/l),salé (chlorure de sodium 1,5 g/l), aigreur(acide citrique 0,25 g/l), amertume (ca-féine 0,05 g/l) (Bainbridge et al. 1996).
Les panélistes sélectionnés ont été sou-mis à une série de tests sur des bananesnon utilisées dans l’expérience afin de leurpermettre d’apprendre les différents quali-ficatifs à attribuer pour caractériser laqualité des bananes. Avant d’être présen-tés aux panélistes, les échantillons expéri-mentaux ont été numérotés avec quatrechiffres pour éviter les erreurs de noms.Les panélistes ont été ensuite priés decoter les échantillons en utilisant uneéchelle hédonique à 6 points allant de 1pour une approbation maximum et 6 pourune désapprobation maximum d’une qua-lité donnée (Larmond 1987, Jellinek 1985).Les caractéristiques évaluées par les pané-listes sont le goût, la texture, la couleur etl’acceptabilité. L’extraction de jus a étéfaite en utilisant les méthodes décrites parKyamuhangire (1990).
Les données ont été analysées en utili-sant le Modèle Linéaire Généralisé (GLM)et l’analyse des variances (Mead et al.
22 INFOMUSA — Vol 9, N° 2
L’acceptabilité des bananesexotiques par le consommateurougandais
Amélioration Tests de qualité
Tableau 1. Caractéristiques des cultivars/hybrides utilisés dans l’étude.
Nom Génome Origine Caractéristiques
Cercosporiose Fusariose Charançons Poids moyen du régime (kg)
FHIA-01 AAAA FHIA R R T 45,6
FHIA-02 AAAA FHIA R R T 25,8
FHIA-03 AAAA FHIA R R T 35,1
FHIA-17 AAAA FHIA R R T 43,9
FHIA-23 AAAA FHIA R R T 45,4
TMPx582/4 AAAB IITA T R T 16,9
TMPx5511/2 AABB IITA T R T 17,9
TMPx548/9 AAAB IITA T R T 20,8
PV 03-44 - IITA T R T 25,7
TMPx548/4 AAAB IITA T R T 15,2
TMPx7002/1 AAAB IITA T R T 25,4
Pisang Ceylan AA IITA T R T 17,2
Yangambi km5 AAA IITA R R T 25,1
Saba ABB IITA T R T 16,7
*Mbwazirume AAA Local S R S 15,6
*Kisansa AAA Local S R S 16,5
+Entundu AAA Local S R S 16,1
+Kisubi AB Local S S T 12,6
+Pisang awak ABB Local S S T 14,2S = sensible, T = tolérant, R = résistant.
* Cultivars utilisés comme témoins pour les tests de cuisson.
+ Cultivars utilisés comme témoins pour les tests de rendement en jus.
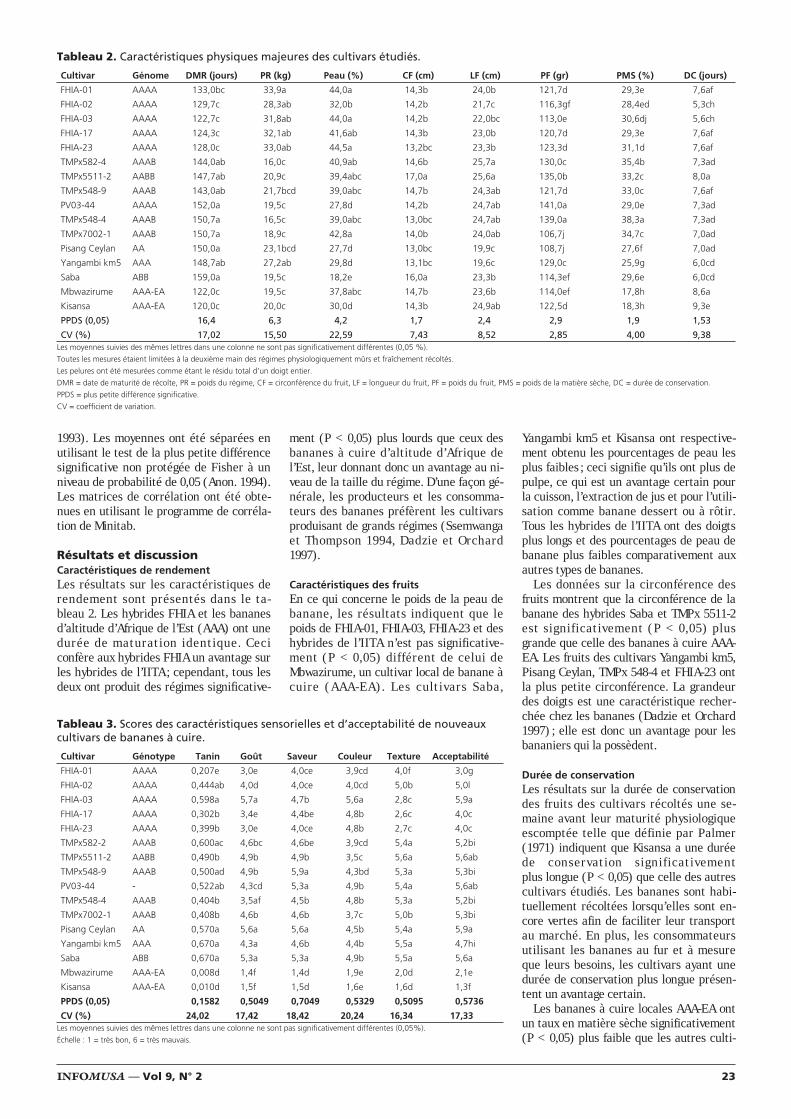
1993). Les moyennes ont été séparées enutilisant le test de la plus petite différencesignificative non protégée de Fisher à unniveau de probabilité de 0,05 (Anon. 1994).Les matrices de corrélation ont été obte-nues en utilisant le programme de corréla-tion de Minitab.
Résultats et discussionCaractéristiques de rendementLes résultats sur les caractéristiques derendement sont présentés dans le ta-bleau 2. Les hybrides FHIA et les bananesd’altitude d’Afrique de l’Est (AAA) ont unedurée de maturation identique. Ceciconfère aux hybrides FHIA un avantage surles hybrides de l’IITA ; cependant, tous lesdeux ont produit des régimes significative-
ment (P < 0,05) plus lourds que ceux desbananes à cuire d’altitude d’Afrique del’Est, leur donnant donc un avantage au ni-veau de la taille du régime. D’une façon gé-nérale, les producteurs et les consomma-teurs des bananes préfèrent les cultivarsproduisant de grands régimes (Ssemwangaet Thompson 1994, Dadzie et Orchard1997).
Caractéristiques des fruitsEn ce qui concerne le poids de la peau debanane, les résultats indiquent que lepoids de FHIA-01, FHIA-03, FHIA-23 et deshybrides de l’IITA n’est pas significative-ment (P < 0,05) différent de celui de Mbwazirume, un cultivar local de banane àcuire (AAA-EA). Les cultivars Saba,
Yangambi km5 et Kisansa ont respective-ment obtenu les pourcentages de peau lesplus faibles ; ceci signifie qu’ils ont plus depulpe, ce qui est un avantage certain pourla cuisson, l’extraction de jus et pour l’utili-sation comme banane dessert ou à rôtir.Tous les hybrides de l’IITA ont des doigtsplus longs et des pourcentages de peau debanane plus faibles comparativement auxautres types de bananes.
Les données sur la circonférence desfruits montrent que la circonférence de labanane des hybrides Saba et TMPx 5511-2est significativement (P < 0,05) plusgrande que celle des bananes à cuire AAA-EA. Les fruits des cultivars Yangambi km5,Pisang Ceylan, TMPx 548-4 et FHIA-23 ontla plus petite circonférence. La grandeurdes doigts est une caractéristique recher-chée chez les bananes (Dadzie et Orchard1997) ; elle est donc un avantage pour lesbananiers qui la possèdent.
Durée de conservationLes résultats sur la durée de conservationdes fruits des cultivars récoltés une se-maine avant leur maturité physiologiqueescomptée telle que définie par Palmer(1971) indiquent que Kisansa a une duréede conservation significativement plus longue (P < 0,05) que celle des autrescultivars étudiés. Les bananes sont habi-tuellement récoltées lorsqu’elles sont en-core vertes afin de faciliter leur transportau marché. En plus, les consommateursutilisant les bananes au fur et à mesureque leurs besoins, les cultivars ayant unedurée de conservation plus longue présen-tent un avantage certain.
Les bananes à cuire locales AAA-EA ontun taux en matière sèche significativement(P < 0,05) plus faible que les autres culti-
INFOMUSA — Vol 9, N° 2 23
Tableau 2. Caractéristiques physiques majeures des cultivars étudiés.
Cultivar Génome DMR (jours) PR (kg) Peau (%) CF (cm) LF (cm) PF (gr) PMS (%) DC (jours)
FHIA-01 AAAA 133,0bc 33,9a 44,0a 14,3b 24,0b 121,7d 29,3e 7,6af
FHIA-02 AAAA 129,7c 28,3ab 32,0b 14,2b 21,7c 116,3gf 28,4ed 5,3ch
FHIA-03 AAAA 122,7c 31,8ab 44,0a 14,2b 22,0bc 113,0e 30,6dj 5,6ch
FHIA-17 AAAA 124,3c 32,1ab 41,6ab 14,3b 23,0b 120,7d 29,3e 7,6af
FHIA-23 AAAA 128,0c 33,0ab 44,5a 13,2bc 23,3b 123,3d 31,1d 7,6af
TMPx582-4 AAAB 144,0ab 16,0c 40,9ab 14,6b 25,7a 130,0c 35,4b 7,3ad
TMPx5511-2 AABB 147,7ab 20,9c 39,4abc 17,0a 25,6a 135,0b 33,2c 8,0a
TMPx548-9 AAAB 143,0ab 21,7bcd 39,0abc 14,7b 24,3ab 121,7d 33,0c 7,6af
PV03-44 AAAA 152,0a 19,5c 27,8d 14,2b 24,7ab 141,0a 29,0e 7,3ad
TMPx548-4 AAAB 150,7a 16,5c 39,0abc 13,0bc 24,7ab 139,0a 38,3a 7,3ad
TMPx7002-1 AAAB 150,7a 18,9c 42,8a 14,0b 24,0ab 106,7j 34,7c 7,0ad
Pisang Ceylan AA 150,0a 23,1bcd 27,7d 13,0bc 19,9c 108,7j 27,6f 7,0ad
Yangambi km5 AAA 148,7ab 27,2ab 29,8d 13,1bc 19,6c 129,0c 25,9g 6,0cd
Saba ABB 159,0a 19,5c 18,2e 16,0a 23,3b 114,3ef 29,6e 6,0cd
Mbwazirume AAA-EA 122,0c 19,5c 37,8abc 14,7b 23,6b 114,0ef 17,8h 8,6a
Kisansa AAA-EA 120,0c 20,0c 30,0d 14,3b 24,9ab 122,5d 18,3h 9,3e
PPDS (0,05) 16,4 6,3 4,2 1,7 2,4 2,9 1,9 1,53
CV (%) 17,02 15,50 22,59 7,43 8,52 2,85 4,00 9,38Les moyennes suivies des mêmes lettres dans une colonne ne sont pas significativement différentes (0,05 %).
Toutes les mesures étaient limitées à la deuxième main des régimes physiologiquement mûrs et fraîchement récoltés.
Les pelures ont été mesurées comme étant le résidu total d’un doigt entier.
DMR = date de maturité de récolte, PR = poids du régime, CF = circonférence du fruit, LF = longueur du fruit, PF = poids du fruit, PMS = poids de la matière sèche, DC = durée de conservation.
PPDS = plus petite différence significative.
CV = coefficient de variation.
Tableau 3. Scores des caractéristiques sensorielles et d’acceptabilité de nouveauxcultivars de bananes à cuire.
Cultivar Génotype Tanin Goût Saveur Couleur Texture Acceptabilité
FHIA-01 AAAA 0,207e 3,0e 4,0ce 3,9cd 4,0f 3,0g
FHIA-02 AAAA 0,444ab 4,0d 4,0ce 4,0cd 5,0b 5,0l
FHIA-03 AAAA 0,598a 5,7a 4,7b 5,6a 2,8c 5,9a
FHIA-17 AAAA 0,302b 3,4e 4,4be 4,8b 2,6c 4,0c
FHIA-23 AAAA 0,399b 3,0e 4,0ce 4,8b 2,7c 4,0c
TMPx582-2 AAAB 0,600ac 4,6bc 4,6be 3,9cd 5,4a 5,2bi
TMPx5511-2 AABB 0,490b 4,9b 4,9b 3,5c 5,6a 5,6ab
TMPx548-9 AAAB 0,500ad 4,9b 5,9a 4,3bd 5,3a 5,3bi
PV03-44 - 0,522ab 4,3cd 5,3a 4,9b 5,4a 5,6ab
TMPx548-4 AAAB 0,404b 3,5af 4,5b 4,8b 5,3a 5,2bi
TMPx7002-1 AAAB 0,408b 4,6b 4,6b 3,7c 5,0b 5,3bi
Pisang Ceylan AA 0,570a 5,6a 5,6a 4,5b 5,4a 5,9a
Yangambi km5 AAA 0,670a 4,3a 4,6b 4,4b 5,5a 4,7hi
Saba ABB 0,670a 5,3a 5,3a 4,9b 5,5a 5,6a
Mbwazirume AAA-EA 0,008d 1,4f 1,4d 1,9e 2,0d 2,1e
Kisansa AAA-EA 0,010d 1,5f 1,5d 1,6e 1,6d 1,3f
PPDS (0,05) 0,1582 0,5049 0,7049 0,5329 0,5095 0,5736
CV (%) 24,02 17,42 18,42 20,24 16,34 17,33Les moyennes suivies des mêmes lettres dans une colonne ne sont pas significativement différentes (0,05%).
Échelle : 1 = très bon, 6 = très mauvais.

vars étudiés ; ceci constitue un désavan-tage énorme en termes d’amélioration derendement pendant la sélection (Anon.1993). Les résultats sur le taux en tanin(polyphénols) et les analyses sensoriellesdes cultivars indiquent que les bananes àcuire locales AAA-EA ont un taux en taninplus faible que les cultivars introduits, les-quels ont un goût astringent désagréable(tableau 3).
Analyse sensorielleLes résultats de l’évaluation sensoriellepour le goût, la texture, la couleur, l’odeuret l’acceptabilité des bananes cuites indi-quent que les bananes introduites et lesbananes plantain ont des scores significati-vement (P < 0,05) plus faibles que les ba-nanes d’altitude d’Afrique de l ’Est(tableau 3). La ‘texture’ des bananes a étéqualifiée en termes sensoriels commeétant ‘dure’, ‘moyenne’ ou ‘molle’ (Ssem-wanga et al. 1996). Les panélistes ont re-jeté la texture des hybrides la décrivantcomme ‘dure’ et donc inacceptable. Lesconsommateurs ougandais n’aiment pas lesbananes cuites dont la texture n’est pasmolle (Ssemwanga et Thompson 1994).
Les résultats de l’étude ont aussi montréque la couleur et le goût des bananes intro-duites sont significativement (P < 0,05)moins appréciés que ceux des bananes àcuire AAA-EA. Les panélistes ont aussiémis des plaintes concernant une sensa-tion piquante dans la bouche et un manquede saveur des bananes des cultivars intro-duits, les rendant ainsi inférieures aux ba-nanes à cuire.
Acceptabilité généraleLe jugement final des cultivars par les pa-nélistes (acceptabilité générale), englo-bant toutes les perceptions, montre quetous les cultivars introduits sont significati-
vement (P < 0,05) inférieurs aux bananes àcuire AAA-EA. Tous les hybrides de l’IITAet FHIA-03 ont obtenu des scores de 5 etplus, indiquant ainsi une désapprobationtotale. FHIA-01 obtient un score de 3, leplus proche des bananes à cuire AAA-EA.Les résultats d’acceptabilité sont conformesaux rapports antérieurs selon lesquels lesbananiers hybrides sont supérieurs auxcultivars locaux en ce qui concerne les ca-ractéristiques du fruit et du régime, maisleur sont inférieurs en ce qui concerne lesqualités d’utilisation (Anon. 1993, Ssem-wanga et Thompson 1994, Sebasigari 1996,Mwenebanda et Banda 1996).
Les résultats de la matrice de corréla-tion entre les caractéristiques des cultivarsétudiés montrent qu’il y a une relationétroite entre le taux en tanin et la couleur(r = 0,81) tel que signalé par Macheix etal. (1990) (tableau 4). Cette corrélationhautement positive entre la teneur entanin et la couleur suggère qu’une teneurélevée en tanin affecte négativement l’ap-parence des bananes cuites. Le coefficientde corrélation entre la texture et l’accepta-bilité (r = 0,77) fait supposer que les culti-vars ayant une texture dure ne seraient pasacceptées par les consommateurs. Le coef-ficient de corrélation entre le goût et l’ac-ceptabilité de 0,89 suggère que le goût in-fluence significativement (P < 0,05)l’acceptabilité, confirmant l’observationfaite dans le même sens par Ssemwanga etThompson (1994) et Sebasigari (1996).
La corrélation (r = 0,59) significative (P < 0,05) entre la teneur en fibres et l’ac-ceptabilité suggère que la quantité desfibres crues est un critère important pourl’acceptabilité. La corrélation entre la ma-tière sèche et la texture est aussi significa-tive (P < 0,05), suggérant que les variétésayant une haute teneur en matière sècheauraient une texture dure et seraient donc
inacceptables. Ceci expliquerait le faibletaux d’acceptabilité des hybrides de l’IITA.La relation entre la teneur en tanin et latexture suggère que les cultivars ayant unehaute teneur en tanin ont une texture in-acceptable. La relation entre la date dematurité et l’acceptabilité est aussi signifi-cative (r = 0,68) (P < 0,05), suggérant queles cultivars tardifs sont moins accep-tables.
Les résultats sur le rendement en jus in-diquent que seuls FHIA-03, FHIA-01, Yangambi km5 et Saba ont produit du jus(tableau 5). Le pourcentage de jus produitpar FHIA-03 est significativement plusélevé que celui du cultivar local Entunduet des cultivars Musa-Kayinja et Kisubi. Lescultivars Yangambi km5, FHIA-01 et Sabaont eu des rendements en jus similaires àEntundu (AAA-EA), un cultivar à jus tradi-tionnel. Les cultivars ayant un faible ren-dement en jus ont produit un jus contenantune quantité de brix significativement (P < 0,05) plus importante que celle descultivars ayant un rendement en jus plusélevé. Normalement, le jus est directementconsommé après extraction ou dilué selonla douceur voulue. La douceur d’un jus ac-ceptable correspond normalement à untaux de brix compris entre 12 et 14 %(Pekke, communication personnelle) ; ceciexpliquerait pourquoi le jus de banane est normalement dilué avant d’êtreconsommé. Le désavantage d’avoir unfaible taux de brix est compensé par desrendements en jus plus élevés. FHIA-01 etEntundu ont un pH de jus identique et si-gnificativement (P < 0,05) plus élevé quecelui des autres cultivars. Le pH des jus estun attribut important parce qu’il influenceles types et les niveaux de contaminationmicrobienne et donc le type de conserva-tion requis (Jay 1987). Un pH faible en-traîne des niveaux de contamination mi-
24 INFOMUSA — Vol 9, N° 2
Tableau 4. Corrélation entre les caractéristiques physiques, chimiques et sensorielles des cultivars étudiés.
DM PR PF LF CF PP % PMS % TSS % TNN PRo CT FB GT SV CL TX AC
DMPR -0,533* -
PR 0,263 -0,241 -
LF 0,019 -0,463 0,447 -
CF 0,104 -0,229 0,064 0,506* -
PP % -0,446 0,358 0,013 0,297 -0,113 -
PMS % 0,532* -0,108 0,292 0,268 0,008 0,357 -
TSS % -0,121 0,186 -0,016 0,171 0,306 0,151 0,394 -
TNN 0,663* -0,087 0,282 -0,180 0,130 -0,230 0,640* 0,274 -
PRo -0,566 0,104 0,074 0,392 0,102 0,139 -0,432 0,174 -0,528* -
CT -0,371 0,061 0,187 0,146 -0,227 0,213 -0,326 -0,408 -0,307 0,233 -
FB 0,601* -0,432 0,219 0,260 0,304 -0,093 0,567* 0,034 0,754* -0,365 -0,008 -
GT 0,614* -0,046 -0,098 -0,267 0,151 -0,171 0,598* 0,263 0,832* -0,587* -0,408 0,603* -
SV 0,699* 0,008 0,131 -0,162 0,035 -0,112 0,725* 0,220 0,793* -0,636* -0,310 0,508* 0,891* -
CL 0,499 -0,007 -0,098 -0,377 -0,076 -0,120 0,595* 0,230 0,810* -0,535* -0,424 0,560* 0,839* 0,719* -
TX 0,901* -0,391 0,292 -0,052 0,115 -0,329 0,614* 0,095 0,728* -0,556* -0,431 0,489 0,700* 0,771* 0,595* -
AC 0,678* -0,135 0,086 -0,184 0,064 -0,123 0,725* 0,376 0,861* -0,580* -0,438 0,594* 0,938* 0,901* 0,818* 0,766* -DM = date de maturité (période de remplissage), PR = poids du régime, PF = poids du fruit, LF = longueur du fruit, CF = circonférence du fruit, PP % = pourcentages des pelures, PMS %= pourcentage en matière sèche, TSS % = pourcentage de tous les solides solubles, TNN = tanin (valeurs d’absorbance), PRo = protéine, CT = cendre totale, FB = fibres brutes, GT = goût,SV = saveur, TX = texture, CL = couleur, AC = acceptabilité.
* Coefficients significatifs.

crobienne plus faibles et donc un besoinmoindre de recourir à des méthodes depréservation caustique (Jay 1987).
La saveur du jus du cultivar Kasubi a étécoté comme significativement (P < 0,05)meilleur que celle du jus de la plupart desautres cultivars étudiés. Ce cultivar a aussile pH le plus faible (4,5). Les cultivarsSaba et FHIA-01 ont produit des jus ayantune douceur au palais significativement (P < 0,05) meilleure que celle des bananesà bière locales. La douceur au palais du jusest une caractéristique de qualité très im-portante (Koffi et al. 1991).
Un bon rendement en jus, un jus de bonnequalité, des régimes de grande taille et unebonne résistance à la cercosporiose noire età la fusariose font de FHIA-01, FHIA-03,Yangambi km5 et Saba des bons candidatspour la production de jus/bière dans les en-droits du pays où le brassage est une activitééconomique importante et où les cultivarstraditionnels sont en train d’être déciméspar les ravageurs et les maladies.
ConclusionLes résultats de l’étude indiquent que,bien que les cultivars introduits portent degros régimes et de beaux fruits, leurs quali-tés culinaires sont inacceptables pour lesconsommateurs du fait de leur taux élevéen tanin, de leur texture dure et de leurmanque de saveur, comparés aux bananesà cuire AAA-EA traditionnelles.
Cependant, les scores obtenues par lescultivars FHIA-01, FHIA-17 et FHIA-23pourraient les rendre acceptables commebananes à cuire dans certaines parties dupays où les cultivars locaux sont en trainde disparaître. A cause de leur jus debonne qualité, les cultivars FHIA-01, FHIA-03, Yangambi km5 et Saba peuvent rempla-cer les cultivars à jus locaux traditionnelslà où ceux-ci sont en train de disparaître. ■
RéférencesAnonymous. 1993. Plantain/Banana improvement.
Annual Report, IITA.Anonymous. 1994. SAS/STAT users’ guide. Ver-
sion 6, 4th ed., Vol. 2. Cary CN. SAS InstituteInc. 846 pp.
Bainbridge Z, K. Tomlins, K. Wellings & A. Westby.1996. Methods for assessing quality characteris-tics of non-grain starchy staples. Part 3: Labora-tory methods. NRI, UK. 70 pp.
Broadhurst R.B. & W.T. Jones. 1978. Analysis ofcondensed tannins using acidified Vanillin. Jour.Food Sc. & Agric. 29: 77-79.
Dadzie B.K. 1998. Postharvest characteristics ofblack Sigatoka resistant banana, cooking bananaand plantain hybrids. Inibap technical guidelinesNo 4. 75 pp.
Dadzie B.K. & J. E. Orchard. 1997. Routine posthar-vest screening of banana/plantain hybrids: crite-ria and methods. Inibap technical guidelinesNo 2. 63 pp.
Hartmanns E. 1989. Five-year food crops researchplan (1989-1994) and recommendations forstrengthening research and extension linkages.MAAIF, Uganda.
Jay M. J. 1987. Modern food microbiology. 3rd ed.,CBS Publ., India.
Jellinek G. 1985. Sensory evaluation of food. Theoryand practice. Food Tech Journ. 14: 33-60.
Koffi E.K., C.A. Sims & R.P. Bates. 1991. Viscosityreduction and prevention of browning in the pre-paration of clarified banana juice. Journ. of foodquality, University of Florida, Gainesville, USA.14 (3): 209-218.
Kyamuhangire W. 1990. Banana juice extractionand processing. M.Sc. Thesis. University of NewSouth Wales, Kingston, Australia.
Larmond E. 1987. Sensory evaluation can be objec-tive. Pp. 3-14. in Objective methods in food qua-lity assessment No 1. CRC Press Inc., BocaRaton, Florida.
Macheix J.J., A. Fleuriet & J. Billot. 1990. Fruit phe-nolics. CRC Press Inc., Boca Raton, Florida. Pp 10-22.
Mead R., R.N. Curnow & A.M. Harsted. 1993. Statis-tical methods in agriculture and experimentalbiology. Chapman and Hall, London. 415 pp.
Mwenebanda B.M.L & D.L. Banda. 1996. Perfor-mance of the advanced Musa yield trial in Ma-lawi. MusAfrica 10: 11.
Ortiz R. and D. Vuylsteke. 1998. Bita-3, a starchy ba-nana with a partial resistance to black Sigatokaand tolerant to streak virus. Hort. Sci. 33: 358-359.
Palmer J.K. 1971. The biochemistry of fruits andtheir products. 11: 65-105. Academic press, London.
Rubaihayo P.R. & C. Gold. 1991. A Report on RapidRural Appraisal Survey of Banana Production.Banana Cropping Systems Research. Dpt. of CropSci. Makerere University. Research Bulletin 2.
Sebasigari K. 1996. Characterisation of exotic bana-nas to determine their fruit utilisation accordingto food habits in East Africa. MusAfrica 10: 7.
Ssemwanga J.K., A.K. Thompson & J.A. Aked. 1996.Quality and acceptability studies of the bananahybrid FHIA-03 as compared to the indigenouscultivars for matooke preparation. Ph.D thesispresented to University of Reading, U.K.
Ssemwanga J.K. & A.K. Thompson. 1994. Investiga-tion of postharvest and eating qualities likely toinfluence acceptability of matooke banana culti-vars to be introduced in Uganda. Aspects of ap-plied biol. 39: 207-213.
Tushmereirwe W. K. 1996. Factors influencing theexpression of leafspot diseases of highland bana-nas in Uganda. A Ph.D thesis submitted to theUniversity of Reading, Department of Agricul-ture, UK.
K. Nowakunda et W. Tushemereirwe travaillent auProgramme Bananier du Kawanda Agricultural Re-search Institute, PO BOX 7065, Kampala ; P. R. Rubaihayo et M. A. Ameny travaillent à la Fa-culté d’Agriculture de l’Université Makerere, PO Box7062, Kampala, Ouganda.
INFOMUSA — Vol 9, N° 2 25
Tableau 5. Rendement en jus et caractéristiques du jus des cultivars introduits.
Cultivar Génotype Poids de la pulpe (kg) Rendements en jus (%) Brix (%) pH Goût Couleur Douceur
FHIA-01 AAAA 2,0 63,4bc 20,08c 5,1a 2,5a 2,0 1,8cd
FHIA-02 AAAA 2,0 0,0 - - - - -
FHIA-03 AAAA 2,0 76,5a 21,70bc 4,7bc 2,8a 2,3 2,8ab
FHIA-17 AAAA 2,0 0,0 - - - - -
FHIA-23 AAAA 2,0 0,0 - - - - -
TMPx582-2 AAAB 2,0 0,0 - - - - -
TMPx5511-2 AABB 2,0 0,0 - - - - -
TMPx548-9 AAAB 2,0 0,0 - - - - -
TMPx548-4 AAAB 2,0 0,0 - - - - -
TMPx7002-1 AAAB 2,0 0,0 - - - - -
Pisang Ceylan AA 2,0 0,0 - - - - -
Yangambi km5 AAA 2,0 67,1b 20,92c 4,8b 2,3ab 2,3 2,0c
Saba ABB 2,0 65,3bc 22,00bc 4,6cd 2,5a 2,3 1,5d
Kisubi AB 2,0 52,7d 25,58a 4,5d 1,5b 2,3 2,8ab
Entundu AAA-EA 2,0 68,5b 19,83c 5,1a 2,8a 2,8 3,0b
Musa-Kayinja ABB 2,0 60,5c 24,00ab 4,8b 2,3ab 2,3 2,5b
PPDS (0,05) 5,77 2,808 0,02 0,85 NS 0,46
CV (%) 33,7 0,76 2,86 24,78 21,17 44,59* Les données analysées proviennent de 7 cultivars qui avaient produit du jus.

Bakelana-ba-Kufimfutu et Mpanda
Le bananier est une culture impor-tante en République Démocratiquedu Congo, où elle arrive en deuxième
position après le manioc. Ses fruits jouentun rôle très important dans l’alimentationde la population. La banane douce estconsommée dès maturité, et la bananeplantain est transformée en pâte appelée« Lituma », ou bouillie et mélangée auxautres aliments.
Le développement de la culture du bana-nier passe par une amélioration des pra-tiques culturales et du matériel de planta-tion (rejets). Les agriculteurs ont degrandes difficultés à se procurer du maté-riel de plantation en quantité suffisantepour augmenter les superficies cultivées.
Il existe plusieurs techniques de multi-plication des rejets. Avec la multiplicationnaturelle, pratiquée par la majorité despaysans, il est difficile d’obtenir des rejetsen temps voulu et en quantité suffisante.La fausse décapitation qui s’est révéléeperformante dans les essais expérimen-taux ne satisfait pas les besoins des pay-sans qui ne veulent pas voir retarder leursrécoltes pour cause de multiplication ra-pide des rejets.
Les bananiers issus de multiplication na-turelle par rejets ont des rendementsfaibles en régimes par rapport à ceux obte-nus par culture de tissus (Adelaja 1995).Cependant la culture de tissus reste inac-cessible aux paysans compte tenu de sescoûts et l’infrastructure requise.
Noupadja (1996) a comparé trois mé-thodes de multiplication rapide des rejets :fausse décapitation, décapitation totale etinclinaison du pseudotronc. Les meilleursrésultats sont obtenus avec la fausse déca-pitation : une moyenne de 12 rejets étaitobtenue 9 mois après plantation. Parallèle-ment, le Centre de recherches régionalessur bananiers et plantains (CRBP) établi
au Cameroun a obtenu une production de 8à 14 rejets, 10 mois après plantation avecla fausse décapitation (CRBP 1996).
D’autres méthodes de multiplication ra-pide par fragmentation ou décorticage ontété testées au CRBP. Le décorticage de lasouche a permis d’obtenir de 30 à 80 rejetsen peu de temps (CRBP 1996). Le décorti-cage des souches qui avaient déjà fleuriétait plus performant que le décorticagedes souches n’ayant pas encore fleuri(Bonté et al. 1995, CRBP 1996).
Une évaluation de la méthode de décorti-cage des souches a été entreprise à l’Univer-sité Protestante de Kimpese pour infirmerou confirmer les résultats antérieurs. Seulesles souches-mères étaient décortiquées ; lesrejets de premier rang étaient immédiate-ment sevrés à 3 feuilles épanouies.
Matériel et méthodesLocalisationL’essai de décorticage des souches a étéconduit à l ’Université de Kimpese, à222 km sur la route nationale Kinshasa-Ma-tadi, à une altitude de 337 m. La moyenneannuelle des précipitations varie entre 900et 1 500 mm et la température moyenne an-nuelle oscille autour de 25 °C.
TraitementsDix-huit pieds de bananier dessert ont étédessouchés : neuf pieds non fleuris et neufpieds fleuris (c.à.d. ayant produit un ré-gime de bananes). Les traitements appli-qués étaient les suivants :1. Souche du pied non fleuri et non fendu2. Souche du pied non fleuri et fendu en
deux sections3. Souche du pied non fleuri et fendu en
quatre sections4. Souche du pied fleuri et non fendu5. Souche du pied fleuri et fendu en deux
sections6. Souche du pied fleuri et fendu en quatre
sections
Dispositionpieds non fleuris 1 2 3pieds fleuris 4 5 6
Le dispositif expérimental utilisé étaitcelui des blocs en randomisation totaleavec deux répétitions. Les bulbes isolés deleurs fausses tiges étaient soigneusementdéshabillés des gaines foliaires.
Deux gaines superposées déterminentun bourgeon latéral au point creux ou en‘V’ de leur rencontre. Le nombre de bour-geons est donc égal au nombre de feuillesque le pied contient. Les bourgeons defeuilles âgées sont plus visibles que ceuxdes très jeunes feuilles. Des précautionsont été prises pour ne pas endommager lesbourgeons visibles ayant plus de potentia-lité de développement en conditions favo-rables. Après décorticage, le méristèmeapical des bulbes des pieds non fleurisétait endommagé pour activer la crois-sance des bourgeons latéraux. Les bulbesétaient régulièrement arrosés pendanttoute la durée de l’évaluation. Les para-mètres suivants ont été observés :1)nombre de jours au démarrage du bour-
geon2)nombre des rejets de premier rang pro-
duit par chaque souche3)hauteur des rejets au sevrage
Chaque rejet était sevré de la souche-mère au stade trois feuilles. Toutes lessouches étaient repiquées après décorti-cage dans un substrat constitué d’argile.
Résultats et discussionLes résultats sont présentés dans le ta-bleau 1.
L’analyse statistique des données met enévidence des différences significativespour le nombre de jours au démarrage desbourgeons latéraux et la hauteur des re-jets.• Influence de l’âge : les bourgeons laté-
raux se développent plus rapidement surles pieds fleuris. Ceci peut s’expliquerpar le fait que les vieux bulbes possèdent
Méthode de multiplication des bananiers par décorticage de la souche
Agronomie Obtention simple de rejets
Tableau 1. Influence de l’âge de la souche et du nombre de fentes sur le démarrage des bourgeons latéraux, le nombre de rejets produits et la hauteur des rejets.
Age du bulbe Nombre de fentes Nombre de jours au démarrage du bourgeon Nombre de rejets Hauteur des rejets (cm)
Pied non fleuri 0 fente 47 2 18
2 fentes 39 2 20
4 fentes 33 7 22
Pied fleuri 0 fente 31 5 28
2 fentes 27 8 33
4 fentes 18 7 35
PPDS (0,5) 4,50 NS 3,4
CV (%) 9,36 52,7 8,3
26 INFOMUSA — Vol 9, N° 2

des bourgeons mûrs et des réserves nu-tritives favorisant leur développement.
• Influence du nombre de fentes : le dé-marrage des bourgeons est plus rapidepour le traitement à 4 fentes. La compé-tition entre les bourgeons en croissanceest moins forte sur la souche fendue. Ladominance est plus active sur la souchenon fendue.
• Nombre de rejets produits : les piedsfleuris ont produit plus de rejets mais ladifférence n’est pas significative. Ceciconfirme les résultats obtenus dans lepassé (Bonté et al. 1995, CRBP 1996). Lenombre de rejets obtenus est inférieur àcelui obtenu par le CRBP. Mais notreétude s’est limitée au décorticage de lasouche-mère alors que les essais duCRBP portaient aussi sur le décorticagedes rejets de premier et peut-être dedeuxième rang avant le sevrage des re-jets produits. Les rejets produits sur lessouches des pieds fleuris étaient detaille supérieure à ceux produits sur lespieds non fleuris. La croissance des re-jets des souches des pieds fleuris et fen-dues 4 fois est supérieure à ceux desautres rejets.
Conclusion et recommandationsLa méthode de multiplication des rejets àpartir de souches décortiquées de bana-niers est très simple et peu coûteuse. Lessouches saines abandonnées dans lechamp après la récolte peuvent être récu-pérées pour servir à la production des re-jets dont les paysans ont besoin. ■
RéférencesAdelaja B.A. 1995. Rapid on-farm multiplication tech-
nique for plantain and banana. MusAfrica 8: 6.Bonté E., R. Verdonck & L. Grégoire. 1995. La multi-
plication rapide du bananier et du bananier plan-tain au Cameroun. Tropicultura 13(3): 109-116.
CRBP. 1997. Rapport annuel 1996. Centre de Re-cherches Régionales sur Bananiers et Plantains,Douala, Cameroun.
Noupadja P. 1995. Study of three field multiplicationtechniques for generating planting material of in vitro propagated plantain (Musa cv. AAB). MusAfrica 8: 7-8.
Bakelana-ba-Kufimfutu travaille à l’Institut Nationalpour l’Étude et la Recherche Agronomique, BP 2007,Kinshasa I, République Démocratique du Congo, etMpanda à l’Université Protestante de Kimpese, Bas-Congo, République Démocratique du Congo.
INFOMUSA — Vol 9, N° 2 27
Rema Menon
La banane est la principale culturefruitière tropicale de l’État du Ke-rala, qui occupe le sud-ouest de la
péninsule du sous-continent indien. Loca-lisé dans le centre de diversité des bana-niers cultivés, le Kerala est, comme deuxautres États du sud de l’Inde – le TamilNadu et le Karnataka –, réputé pour la di-versité de ses ressources génétiques, no-tamment au sein des groupes génomiquesAAB, AB et ABB de la section Eumusa desbananiers comestibles (Stover et Sim-monds 1987). Si l’on trouve une multitudede cultivars au Kerala, le plus communé-ment cultivé est Nendran (French plan-tain, AAB), qui jouit d’une grande popula-rité à la fois comme banane dessert etbanane à cuire. La culture commerciale decette variété représente près de 50 % de laproduction bananière de l’État. Les autrescultivars importants sont Poovan (AAB),Rasthali (AAB), Red Banana (AAA), DwarfCavendish (AAA), Robusta (AAA), Kunnan(AB) et Monthan (ABB), qui sont cultivésuniquement en jardin et constituent unesource de revenus importante pour la po-pulation rurale.
La station de recherche bananière deKannara dépend de la Kerala AgriculturalUniversity. L’un de ses principaux objec-tifs est de collecter, conserver et évaluer lematériel génétique de bananier. Elle a unecollection au champ de 225 accessions,comprenant du matériel indigène et exo-tique. Ces accessions ont été caractériséeset évaluées à l’aide des descripteurs IN-IBAP/IPGRI. Ci-après sont présentés lesrésultats d’essais d’évaluation prélimi-naires effectués de 1994 à 1997 sur un cer-tain nombre d’introductions.
Matériel et méthodesLe matériel testé était constitué de 22 ac-cessions de la collection de l’INIBAP (série
ITC) reçues en 1994 par le canal du Natio-nal Bureau of Plant Genetic Resources(NBPGR) de New Delhi. Ces accessions ontété reçues sous la forme de petits rejets detaille standard un à deux, obtenus par cul-ture de tissus. Les rejets ont été toutd’abord plantés dans des pots, puis transfé-rés dans une parcelle de la station de re-cherche bananière. Celle-ci est située à58 m d’altitude, dans une région où la tem-pérature moyenne s’élève à 28 °C et où lapluviométrie varie entre 2 700 et 3 000 mmpar an, répartis entre deux saisons despluies. L’humidité relative se situe entre 77et 94 %. D’après les observations effectuéessur le premier cycle de culture, il s’estavéré que 14 des 22 accessions étaient déjàreprésentées dans le matériel génétiquede la station. Parmi les huit accessions restantes, trois – Three Hands Planty(ITC1132), Three Vert (ITC1127) et Foconah (ITC649) – n’ont pu s’établir. Ona sélectionné les cinq accessions exotiquesfigurant au tableau 1 pour les soumettre àune nouvelle évaluation.
On a planté cinq rejets de chaque culti-var, chacun sur une rangée, en un disposi-tif compact avec un espacement de 2,5 mentre les rangées et de 2 m entre lesplants de chaque rangée. Ayant constatéque trois de ces cultivars étaient de typeplantain, on a utilisé comme témoins deuxcultivars de plantains indigènes, Nendranet Myndoli. On a enregistré les paramètresde la croissance : hauteur de plant, circon-férence du pseudotronc, nombre defeuilles fonctionnelles à la floraison etdurée du cycle. D’autres variables ont étéobservées : poids du régime, nombre demains et nombre de doigts. On a égale-ment mesuré le rapport pulpe/peau et letaux de conversion en chips chez les culti-vars de type plantain.
Résultats et discussionLe French plantain Nendran, indigène dusud-ouest de l’Inde et représenté par plu-
Évaluation préliminaired’introductions de bananiers au Kerala
Amélioration nouvelles évaluations en Inde
Tableau 1. Accessions sélectionnées pour une nouvelle évaluation.
N° Nom du cultivar Génome Code ITC
1 Big Ebanga AAB 1129
2 Njock kon AAB 1133
3 Nyombe AAB 1124
4 Yangambi km5 AAA 1123
5 Popoulu AAB 1135
6 Nendran* AAB
7 Myndoli* AAB* Cultivars French plantains indigènes.

sieurs écotypes (Jacob 1952), est le princi-pal cultivar de bananier du Kerala et contri-bue de manière fondamentale à la sécuritédes revenus des populations rurales. Ontrouve aussi, de manière limitée, des plan-tains Vrais Cornes, tandis que les FauxCornes ne sont cultivés nulle part en Inde.
Parmi les cultivars exotiques, les deuxFrench plantains Njock kon et Nyombe et leplantain French Big Ebanga ont fait preuved’une très bonne adaptabilité et sont appa-rus comme des introductions utiles pour cequi concerne le Kerala.
Les données sur les caractères végétatifsdes introductions et des cultivars locauxprésentées au tableau 2 font ressortir desvariations dans la hauteur de plant et la cir-conférence du pseudotronc. Njock kon, dotéd’un pseudotronc robuste (76,3 cm), étaitde taille moins haute (250 cm) que Nendran qui mesurait en moyenne 271 cm.Son port, caractérisé par des feuilles trèsimbriquées, suggère le nanisme, qui est uncaractère utile car il améliore la résistanceau vent et facilite la récolte. Big Ebanga etNyombe ont produit des plants de plushaute taille que ceux de Nendran. Nyombes’est caractérisé par un pseudotronc ro-buste de couleur rouge violacé et par unehauteur de 400 cm, dépassant en taille lecultivar de plantain géant local Myndoli. Lecycle de ces trois introductions de plantainsétait plus long que celui des cultivars lo-caux. Néanmoins, Big Ebanga, pour lequelon a enregistré un cycle de 335 jours, pour-rait s’insérer dans le cycle de culture an-nuel pratiqué pour les cultivars de plantainsau Kerala. La longueur du cycle de Nyombea favorisé son infestation par le charançondu pseudotronc (Odoiporus longicollis) quis’attaque depuis quelques années aux plan-tations de bananiers du Kerala.
En ce qui concerne le rendement, BigEbanga a produit en moyenne un régimed’un poids de 14,3 kg, supérieur à celui deNendran (10,9 kg). Le nombre de fruits parrégime était plus élevé chez Nendran. Njockkon et Nyombe, avec un poids moyen de res-pectivement 25 et 32 kg, ont dépassé Nen-dran. Les plantains étant communémentutilisés à l’état vert pour préparer des chips,on a évalué l’adéquation des cultivars deplantains exotiques pour cet usage. BigEbanga a donné un taux de conversion enchips de 34,7 %, dépassant Nendran et les
autres cultivars de plantains pour lesquelsce taux s’est situé entre 29 et 30 %.
Le cultivar Popoulu, qui a produit desfruits courts, épais et tronqués, s’est révéléêtre une introduction intéressante. Il n’estjusqu’à présent cultivé dans aucune régionde l’Inde. Il s’est caractérisé par une hau-teur de plant moyenne de 262 cm et uncycle de 314 jours. Son régime, portant 6mains et 51 doigts, pesait en moyenne9,5 kg. La pulpe de son fruit mûr ressem-blait de manière frappante à celle de Nendran. Yangambi km5, décrit comme un cultivar à flaveur acidulée (Daniells etBryde 1995), a fait preuve d’une très bonnecapacité d’établissement et représente unapport intéressant pour la collection de ma-tériel génétique en raison de sa grande ré-sistance à la cercosporiose jaune, l’une desmaladies qui limite le plus les rendementsau Kerala. Son plant était de hauteur inter-médiaire (en moyenne 226 cm). Son régime,portant de beaux fruits bien remplis de cou-leur vert clair et de flaveur agréable, pesait13,5 kg et se composait de 9 mains et 132doigts. Son rendement était comparable àcelui de plusieurs cultivars de bananiersdessert populaires dans la région.
D’après les résultats de cette évaluationpréliminaire, Big Ebanga et Njock kon of-frent de bonnes possibilités pour la culturecommerciale au Kerala. Étant donné lesdifférences observées dans leur potentielde production, il s’avère nécessaire d’étu-dier leur performance avec différents ni-veaux d’intrants et méthodes de gestion.Ils font actuellement l’objet d’essais multi-locaux en milieu paysan. Popoulu, qui res-semble beaucoup à Nendran, et Yangambikm5, qui a combiné un rendement et unequalité acceptables avec la résistance à lacercosporiose jaune, ont été jugés promet-teurs pour la culture de jardin. ■
RéférencesJacob. 1952. Madras banana: a monograph. Superin-
tendent, Govt. Press, Madras.Stover R.H. & N.W. Simmonds. 1987. Banana
(3rd edition). Longman, London, Royaume-Uni.Daniells J. & N. Bryde. 1995. Un mutant semi-nain
de Yangambi km5. InfoMusa 4(2): 16-17.
L’auteur travaille à la Kerala Agricultural University,Banana Research Station, Marakkal, Kannara, Thrissur 680 652, Inde (courrier électronique : [email protected]).
Tableau 2. Caractères du plant et du régime d’introductions et de cultivars indigènes de bananiers.
Cultivar Hauteur Circonférence du Nb de feuilles Cycle (jours) Poids du régime Nb de mains Nb de doigts Rapport Conversionde plant (cm) pseudotronc (cm) fonctionnelles (kg) pulpe/peau en chips (%)
Big Ebanga 281 53,2 13,5 335 14,3 5,1 44,2 3,3 34,7
Njock kon 250 76,3 12,5 426 25,0 8,7 125,6 3,2 30,0
Nyombe 400 70,5 10,3 480 32,0 10,2 150,3 3,0 29,0
Popoulu 262 52,0 11,0 314 9,5 6 51,0 4,2 -
Yangambi km5 226 51,5 12,0 320 13,5 9,0 132,0 4,0 -
Nendran 271 52,6 12,3 310 10,3 5,0 52,0 3,2 30,0
Myndoli 350 73,2 12,5 422 28,2 8,8 120,3 3,1 29,0
28 INFOMUSA — Vol 9, N° 2
Rachel C. Sotto et Roel C. Rabara
Les bananiers et bananiers plantaincomestibles cultivés dans le mondesont étroitement apparentés à deux
espèces sauvages, dont ils sont en fait ori-ginaires. L’une de ces espèces est Musabalbisiana Colla. Espèce distincte au seindu genre Musa, celle-ci est considéréecomme indigène des Philippines où elle estlargement répandue. Sa large distributionpeut être attribuée à sa grande vigueur et àsa résistance apparemment très élevée auxravageurs et aux maladies. Mais faute d’in-
térêt ou de ressources, on n’a encore en-trepris aucune étude systématique sur ladiversité génétique de M. balbisiana ni dé-crit ses sous-espèces (Shepherd 1988).Face à l’appauvrissement rapide de la bio-diversité des espèces végétales indigènesdes Philippines, il est urgent d’évaluer lavariation génétique et de conserver la di-versité de M. balbisiana dans ce pays.C’est dans cette perspective qu’a été effec-tuée la recherche décrite ci-dessous.
Un projet de recherche de deux ans sur« la diversité génétique et la variation mor-photaxinomique de Musa balbisiana Collaaux Philippines », financé par le Philip-
Diversité morphologique de Musabalbisiana Colla aux Philippines
Ressources génétiques Caractérisation

pine Council for Agriculture, Forestryand Natural Resources Research and De-velopment (PCARRD), a été entrepris le15 juillet 1998. À ce jour, on a collecté 105accessions dans 24 provinces appartenantà 10 régions géopolitiques du pays. Ce ma-tériel est maintenu dans la banque de ma-tériel génétique fruitier de l’Institute ofPlant Breeding (IPB), avec trois touffespar accession, soit un total de 306 touffes.Un double de cette collection a été établi auBureau of Plant Industry - Davao NationalCrop Research and Development Center (BPI-DNCRDC), qui abrite aussi le
Southeast Asian Banana and PlantainGermplasm Resource Center. Sur les 105accessions, on en a caractérisé 76 pourl’ensemble de la plante, le pseudotronc, lafeuille, l’inflorescence, les bractées, lesfleurs mâles et le fruit à l’aide des descrip-teurs de l’IPGRI pour le bananier (Musaspp.) (IPGRI-INIBAP/CIRAD 1996). Cesobservations initiales ont mis en évidenceune vaste diversité de M. balbisiana sur leplan des caractères morphologiques. L’as-pect du régime variait de lâche à très com-pact, et la couleur du fruit avant maturitéde vert à vert clair. On a trouvé une forme
argentée (cireuse) de M. balbisiana quiest particulièrement intéressante. En cequi concerne la forme du fruit, on a ob-servé une grande variation allant d’uneforme ellipsoïde (figure 1a) à une forme ar-rondie (figure 1b), l’apex du fruit variantd’une forme effilée à une forme fortementen goulot de bouteille (figure 1c). Le bour-geon mâle était également variable, allantde l’ellipsoïde (figure 2a) à l’ovale (figure2b), la majorité des accessions exhibantune forme ellipsoïde. Quant à la couleurdes fleurs mâles, elle se situait entre leblanc crémeux et diverses nuances de rose.
INFOMUSA — Vol 9, N° 2 29
Figure 1. Variation de la forme du fruit de M. balbisiana : a) ellipsoïde; b) arrondie ; c) fortement en goulot de bouteille.
Figure 2. Variation de la forme du bourgeon mâle de M. balbisiana : a) ellipsoïde; b) ovale.
a
a b
b c

Nouvelles des Musa
Asie et PacifiqueMise au point d’une nouvelle approchepour l’amélioration des triploïdes à TaiwanLa plupart des cultivars de bananiers sontdes triploïdes. Ils sont normalement vigou-reux et entièrement stériles en conditionsnaturelles. Il existe traditionnellementdeux approches pour les améliorer. L’uneconsiste à croiser des plants triploïdesavec le pollen d’un diploïde pour produireun tétraploïde (3x X 2x ➔ 4x), puis à re-croiser celui-ci avec le géniteur diploïdepour obtenir un triploïde secondaire (4x X2x ➔ 3x) (Rowe et Richardson 1975). Laseconde approche consiste à produire un au-totétraploïde à partir d’un plant diploïde entraitant celui-ci à la colchicine (2x ➔ 4x),puis à le croiser avec un diploïde pour ob-tenir un triploïde (4x X 2x ➔ 3x) (Bakry etHorry, 1994). L’année dernière, le Dr C.T. Shii et Mlle C.F. Liu, chercheurs àl’université nationale de Taiwan, ontréussi, à l’aide de la technique de culturede tissus, à obtenir des plants triploïdes àpartir de l’endosperme de graines en déve-loppement résultant de la fertilisation dediploïdes (Liu 1999, Liu et Shii 1999). On adonc là une nouvelle méthode, à la foissimple et rapide, pour produire des bana-niers triploïdes : 2x X 2x ➔ culture de l’en-dosperme ➔ 3 x.
Un autre avantage de cette approche estqu’on peut utiliser toute combinaison dediploïdes, du moment qu’on obtient un en-dosperme après pollinisation. Les re-cherches se poursuivent sur l’applicationpratique de cette méthode pour l’améliora-tion des bananiers.
RéférencesBakry F. & J.P. Horry. 1994. Musa breeding at
CIRAD-FLHOR. Pp 169-175 in The Improvement
and testing of Musa: a Global Partnership. Pro-ceedings of the First Global Conference of theInternational Musa testing Program (D. Jones,ed.), held at FHIA, Honduras, 27-30 April, 1994.INIBAP, Montpellier.
Liu C.F. 1999. Studies on regeneration capacity inendosperm tissue culture of diploid bananas(Musa spp.). Master thesis. The National TaiwanUniversity, Taipei. 72 pp (en chinois avec résuméen anglais).
Liu C.F. & C.T. Shii. 1999. Induction of embryogene-sis from the endosperm culture of diploid bana-nas. J. Chinese Soc. of Hort. Sci. (Taiwan) 45: 471(résumé en chinois).
Rowe P.R. & D.L. Richardson. 1975. Breeding bana-nas for disease resistance, fruit quality and yield.Bulletin 2. SIATSA. La Lima, Honduras, 41 pp.
Pour plus de détails, contacter : C.Y. Tang, TBRI,PO Box 18, Chiuju, Pingtung 90403, Taiwan
Recensement des plantations de bananiersen AustralieLe Queensland’s Banana Industry Protec-tion Board a récemment publié les résul-tats d’un recensement des plantations debananiers à la date du 30 juin 2000. On adénombré 1 013 exploitations représentant10 501 hectares de plantations. Sur cettesuperficie, les variétés Cavendish occupent9 680 hectares (soit 92 %). Les maladies lesplus courantes sont la cercosporiose jaune(1 013 exploitations), la moucheture fo-liaire (1 013 exploitations), la fusariose(162 exploitations) et le bunchy top (89 ex-ploitations). La cercosporiose noire, le flé-trissement bactérien et la mosaïque desbractées n’ont pas été rencontrés.Pour toute information complémentaire,contacter : N. Janetzki, Secretary, Banana IndustryProtection Board, c/o Department of PrimaryIndustry, GPO Box 46, Brisbane Q 4001, Australie
Effets de la pollution par les fluorures sur la culture des bananiers en ChineUne étude a été effectuée dans le comté deDongguan, en Chine, afin de déterminer les
30 INFOMUSA — Vol 9, N° 2
Figure 3. Variation des caractères des feuilles de M. balbisiana : a) canal pétiolaire étroit avec marges érigées, base du limbe non équilatérale ; b) canal pétiolaireavec marges recourbées vers l’intérieur, base du limbe équilatérale.
On a aussi noté une variation distincte descaractères des feuilles, notamment en cequi concerne le canal pétiolaire qui étaitétroit avec des marges érigées (figure 3a)ou présentait des marges recourbées versl’intérieur (figure 3b), sans que celles-cisoient jamais closes ou se chevauchent. Labase du limbe était équilatérale (figure 3b)ou non équilatérale (figure 3a). On a ob-servé d’autres variations, qui feront l’objetd’un rapport dès que le travail de caracté-risation aura été achevé.
Une caractérisation morphologique etbiochimique à l’aide d’isoenzymes est encours. En complément, il faudra procéder à une caractérisation moléculaire à l’aidede microsatellites spécifiques de locus oude répétitions de séquences simples (SSR),ce qui permettra d’affiner la description de la variation observée au sein de la col-lection de matériel de M. balbisianades Philippines. ■
RéférencesIPGRI-INIBAP/CIRAD. 1996. Descripteurs pour
le bananier (Musa spp.). Institut internationaldes ressources phytogénétiques, Rome, Italie ;Réseau international pour l’amélioration de labanane et de la banane plantain, Montpellier,France ; Centre de coopération internationale enrecherche agronomique pour le développement,Montpellier, France.
Shepherd K. 1988. Observations on Musa taxo-nomy : a note on Musa germplasm of the Philip-pines. Pp. 158-165 in Identification of GeneticDiversity in the Genus Musa : Proceedings of anInternational Workshop held at Los Baños, Phi-lippines, 5-10 September 1988 (R. L. Jarret, ed.). INIBAP, Montpellier, France.
Les auteurs travaillent à l’Institute of Plant Breeding,College of Agriculture, University of the PhilippinesLos Baños, 4031 College, Laguna, Philippines.
a b

causes d’un phénomène de brûlure desmarges des limbes foliaires des bananiers.Ce phénomène, reconnu depuis 1987comme une contrainte majeure pour la pro-duction dans cette zone, semble être res-ponsable de pertes de rendement de l’ordrede 20 %. L’étude a démontré que la causede cette brûlure des feuilles est la pollutionpar des fluorures d’origine atmosphérique,qui proviennent essentiellement des bri-queteries et des usines de ciment. On aconstaté que les Cavendish nains sont plussensibles à cette pollution que les plantainset les autres variétés Cavendish.Pour tous renseignements complémentaires,contacter : Zhang Hailan et Wu Dinyao, Tropicaland Subtropical Fruit Research Lab, South ChinaAgricultural University, Guangzhou 510642, Chine
Études sur les bananiers séminifères de l’Assam, en IndeDans le nord-est de l’Inde, on se sert com-munément de deux types de bananiers sé-minifères comme aliment pour les bébés.Ces bananiers, appelés localement « Bhim-kal » et « Athiakal », sont très robustes ; ré-sistant aux ravageurs, aux maladies et à lasécheresse, ils fournissent des rendementsélevés. Bhimkal, en particulier, est une ex-cellente source d’énergie (114,4 kcal/100 g).On a étudié la croissance, le développementet le génome de ces deux bananiers à l’uni-versité agricole de l’Assam. Leur caractéri-sation morphotaxinomique a révélé queBhimkal et Athiakal ressemblent fortementà Musa balbisiana. Selon le système de no-tation de Simmonds et Shepherd (1955), onn’a trouvé chez eux aucune caractéristique« acuminata » et les notes obtenues indi-quent que ces bananiers ont tous deux ungénome « balbisiana » pur. En comptant leschromosomes dans des cellules de l’extré-mité des racines et des fleurs mâles au mo-ment de la métaphase I, on a trouvé 22chromosomes chez l’un et l’autre bananiers,en 11 paires. Il est donc clair que Bhimkalet Athiakal sont des bananiers diploïdes.
RéférenceSimmonds N. W. & K. Shepherd. 1955. The taxo-
nomy and origins of the cultivated bananas.J. Linn. Soc. Bot. 58: 302-312.
Pour plus de détails, contacter : U. Kotoky,Associate Professor, Department of Horticulture,Faculty of Agriculture, Assam AgriculturalUniversity, Jorhat, Assam, Inde
Le bunchy top : une sérieuse menace pour la culture du bananier d’altitude au Tamil Nadu, en IndeEn Inde, la culture du bananier occupe unesuperficie d’environ 4 millions d’ha et pro-duit des rendements variant entre 4,0 et28,7 tonnes de fruits par ha. Le bananierd’altitude sirumalai/vellavazhai se ren-contre principalement entre 800 et 1 400mètres au Tamil Nadu où il est cultivé ensec, notamment sur les bas de versant descollines de la Palani, ainsi que dans le Ke-rala. Couvrant 15 000 ha avec des rende-
ments de 7 tonnes/ha, cette culture rap-porte 26,5 millions de roupies par an (1000roupies = 23 US$).
Dans cette région, le bunchy top du ba-nanier cause de sérieuses pertes de pro-duction. La forte incidence de cette mala-die s’explique à la fois par la naturepérenne de la culture, la présence conti-nue de l’agent pathogène et le climat douxfavorable au puceron qui en est le vecteur.Lors d’une enquête sur les pertes écono-miques subies à différentes altitudes, on aconstaté que le bunchy top réduit la renta-bilité de la culture du bananier dans uneproportion de 60 % à 1 200 m et de 50 % à1 000 m d’altitude. Au-dessus de 1 200 m oùl’humidité est plus forte, et en dessous de400 m où la température est plus élevée,les pertes sont moins importantes car lesconditions sont moins favorables à la re-production du puceron vecteur.
Cette forte incidence du bunchy top setraduit par des pertes économiques sévèresdans une grande partie de la zone de cul-ture du bananier d’altitude. Si l’on veut mi-nimiser l’impact de la maladie de façon àredynamiser la culture du bananier d’alti-tude et l’économie agricole, il importe deprocéder à l’indexation des rejets (à l’aidedu test ELISA ou de la PCR) afin de détec-ter la présence du bunchy top dès ce stadeet de fournir des rejets sains.Pour toute information complémentaire,contacter : K. Manickam, T. Ganapathy etS. Doraiswamy, Department of Plant Pathology,Centre for Plant Protection Studies, Tamil NaduAgricultural University, Coimbatore 641 003, Inde
Afrique de l’OuestAu Ghana, une nouvelle technologieappropriée permet de multiplierrapidement les bananiers plantainLes plantains et les bananes sont des den-rées amylacées de base pour les Ghanéens.Outre leur importance socioéconomique etnutritionnelle, ils sont aussi une sourced’emploi et de revenu pour les ruraux.
Les principales contraintes de productionsont les ravageurs (charançons et néma-todes), les maladies (en particulier la cerco-sporiose noire), le manque de matériel végé-tal et le déclin de la fertilité des sols.
On a introduit des hybrides tétraploïdesde bananiers et plantains qui sont dotés derésistance ou de tolérance aux maladies.Afin de disposer de davantage de matérielvégétal, on a mis au point une technique demultiplication rapide à partir de fragmentsde souche. Cette technique permet de pro-duire, à partir d’une souche de dimensionmoyenne, 8 à 10 rejets qui peuvent êtretransplantés au bout de 8 à 12 semaines.
Les chercheurs viennent de mettre aupoint une nouvelle technique grâce à la-quelle il est possible de produire davantagede rejets en un temps plus court.
La procédure est la suivante :1. Fabriquer une colonne de bois évidée
mesurant environ 1,2 m de large, 30 cm
de haut et de longueur indifférente, et laremplir de sciure.
2. Déraciner une souche, en couper toutesles racines et la peler afin d’éliminer lesnématodes et les charançons.
3. Couper la souche en deux ou quatre mor-ceaux selon sa dimension.
4. Traiter les morceaux de souche avec unesolution de fongicide (par exemple, 1 %de Benlate).
5. Laisser sécher les morceaux de soucheainsi traités pendant 24 heures à l’airlibre dans un endroit ombragé.
6. Planter les morceaux de souche à 15 cmde profondeur dans la sciure, en plaçantla face coupée vers le bas (de préférenceà proximité d’une source d’eau afin defaciliter l’irrigation).
7. À l’aide de feuilles de palmier ou d’unefeuille de polyéthylène, faire un abri au-dessus de la colonne afin d’y maintenirl’humidité.
8. Irriguer chaque jour.9. La germination commence au bout de 8
jours.Exciser les nouvelles pousses de lasouche lorsqu’elles ont deux feuillesvertes et les planter dans un sac en poly-éthylène noir de 7 x 9 cm rempli de 7parties de sol loameux, 2 parties desable de rivière et 1 partie de fumier devolaille bien décomposé. Après un moisd’acclimatation, on peut transférer lesplants dans le champ.Continuer d’irriguer la colonne et enle-ver les pousses au fur et à mesure deleur apparition.La souche mère continuera de produiredes rejets sains pendant environ 5 se-maines.Avantages :
1. Les rejets sont produits à proximité del’exploitation, ce qui réduit les frais detransport.
2. Le matériel ainsi obtenu est indemne demaladies et de ravageurs.
3. On obtient plusieurs rejets en une pé-riode de temps très courte.
4. Tous les bourgeons présents sur la souchesont en mesure de s’exprimer (tandisqu’une partie risque d’être perdue avec latechnique des fragments de souche).
5. Le matériel obtenu est génétiquement pur.Pour tous renseignements complémentaires,contacter : B.M. Dzomeku, B. Banful, D.K.Yeboah et S.K. Darkey, Crops Research Institute,PO Box 3785, Kumasi, Ghana
Amérique Latine et CaraïbesAmélioration des clones de bananiers« Manzano » (AAB) et « Gros Michel »(AAA) pour la résistance à la fusariosegrâce à la culture de tissus et à la mutagenèse in vitroAvec les progrès que les biotechnologiesont permis de réaliser dans l’améliorationgénétique au cours de ces dernières an-nées, il est devenu plus facile de combinerefficacement la culture in vitro avec l’in-
INFOMUSA — Vol 9, N° 2 31
10.
11.
12.

32 INFOMUSA — Vol 9, N° 2
duction d’une mutation somatique. Dans lecadre de ce projet, il s’agissait de sélec-tionner, en laboratoire puis au champ, dessomaclones issus des clones sensibles« Manzano » (AAB) et « Gros Michel »(AAA) qui soient dotés de résistance ou to-lérance aux races 1 et 2 de la fusariose(Fusarium oxysporum var. cubense). Aprèsavoir initié des vitroplants à partir de bour-geons de ces deux clones, on a induit la for-mation de bourgeons multiples qu’on atraités avec des agents mutagènes (rayonsgamma) sources de Co60. On a ensuite pro-cédé à leur inoculation avec différents iso-lats fongiques en provenance de l’INIFAT –en appliquant pour chaque souche surquatre cycles de développement in vitroune solution de 3 x 105 spores/ml durant30 minutes, méthode qui s’était révélée
être la plus pathogénique dans des étudesantérieures. Une fois ce processus achevé,on a planté les vitroplants dans du sol orga-nique contaminé avec des résidus deplants infectés par Fusarium oxysporum.Au bout de 60 jours, on a réinoculé les vi-troplants qui ne présentaient pas de symp-tômes de jaunissement des feuilles, en in-jectant à la base du pseudotronc unesolution de spores de la même souche. Sixmois plus tard, on a transplanté les vitro-plants sans symptômes dans le champ in-fecté par l’agent pathogène afin d’évaluerl’incidence de la maladie. On a ainsi sélec-tionné 42 somaclones pour en faire unenouvelle multiplication in vitro, de façon àobtenir 100 individus pour chacun d’entreeux (lignée clonale), puis on a de nouveauévalué leur réaction à la maladie en labora-
toire et au champ. Enfin, on a sélectionnéneuf somaclones de « Gros Michel » quiprésentaient au champ des signes d’infec-tion inférieure à 30 %. Parmi le matérieldérivé du clone « Manzano », toutes les sé-lections se sont révélées sensibles après lepremier cycle. On a procédé à la micropro-pagation in vitro des somaclones résistantsqui avaient des caractères agronomiquesfavorables, et une étude est en cours pourévaluer leur rendement.
Informations fournies par : I. Bermúdez,P. Orellana, N. Veitía, C. Romero, J. Clavelo,L. García et M. Acosta, Instituto de Biotecnologíade las Plantas, Universidad Central de las Villas,Carretera a Camajuaní Km 5 1/2, UniversidadCentral « Martha Abreu » de las Villas, SantaClara, Villa Clara, Cuba (courrier électronique :[email protected] ; tél. : 81360 – 81693) etL. Herrera I., Facultad de Ciencias Agropecuarias,UCLV, Santa Clara, Villa Clara, Cuba.
Pauline Osiro Omoaka
Il n’existe pas encore de méthodes ob-jectives et satisfaisantes pour évaluerla qualité post-récolte de la banane.
Pareilles méthodes sont indispensables àla réussite du commerce de la banane et àla protection de la santé du consommateur.
La possibilité d’utiliser la spectrostoco-pie de l’impédance électrique dans l’éva-luation de la qualité post-récolte et la ma-turation de la banane est présentée danscette dissertation. Les résultats de l’étudeindiquent qu’une résistance de faible fré-quence et ‘Py’ ont augmenté avec la matu-rité de la banane alors que les valeurs‘faibles’ (dépendant de la couleur de lapeau) ont surtout été associées à unepulpe de mauvaise qualité. La techniqued’impédance électrique permet d’établirun rapport entre la qualité interne et lacouleur de la peau de la banane pendant lamaturation.
La qualité de la banane peut, à tout mo-ment, être définie par des paramètres debase intercorrelés. Un degré donné de qua-lité de la banane est normalement le fruitd’une combinaison particulière de para-mètres de qualité internes et externes. Ce-pendant, il est difficile de définir les carac-téristiques exactes pouvant permettre defaire la différence entre les degrés de qua-lité de la banane.
L’application des techniques post-ré-coltes appropriées a permis de prolonger la
vie verte, la période de maturation ainsique la durée de conservation du fruit ; ce-pendant la qualité du fruit après récolteest restée inchangée. Le degré de maturitédu fruit à la récolte est donc indispensablepour obtenir une qualité de banane dési-rée. La manipulation de la banane, lesconditions de stockage et les opérationsannexes influencent les changements dansla pulpe et dans la peau et affectent aussila durée de stockage du fruit. Cependant,les changements observés ont complète-ment été dépendants de la physiologie etde la biochimie de la banane.
L’étude a examiné les changements phy-sio-chimiques produits dans des bananessoumises à des pratiques de manipulationet des milieux différents. Les indicateurspotentiels de maturation, de qualité et dedurée de conservation ont été la produc-tion d’éthylène (initiateur), le taux de res-piration, la couleur de la peau, le contenuen solides solubles, la fermeté de la pulpeet l’impédance électrique.
Les changements induits dans les ba-nanes avec l’éthylène sont le résultat d’uneffet significatif de la température sur lamaturation. Cependant, un retard impor-tant dans le ramollissement de la pulpe etla décoloration de la peau a été observédans les fruits ayant mûri ‘naturellement’.Les paramètres tels que l’acide carboxy-lique-1-aminocyclopropane-1 (ACA), lecontenu en chlorophylle et la fermeté de lapulpe tendent à confirmer les change-ments observés. La température et l’induc-
tion d’éthylène ont parfois conduit à desmaturations incontrôlées. L’éthylène exo-gène a induit la production d’éthylène au-tocatalytique. Les bananes soumises à unetempérature comprise entre 14 et 18 °C etune humidité relative comprise entre 85 et95 % ont mûri entre 4 et 8 jours.
Des dégâts causés par le froid se sontmanifestés sur les bananes stockées endessous de 12 °C. L’importance des dégâtsobservés à une température donnée dé-pend de la durée d’exposition du fruit àcelle-ci. Les symptômes sont devenus vi-sibles lorsque les fruits ont été transférés àdes températures favorables ou pendant lapériode de maturation subséquente. Latempérature de la pulpe seule n’a pas étésuffisante pour expliquer la qualité finaledu fruit observée.
Un traitement à l’air chaud des bananesvertes a permis d’induire des changementsnormalement associés au processus de ma-turation de la banane. Sans aucun doute,les résultats obtenus dans cette étudecontribueront à propager l’utilisation dutraitement à l’air chaud comme alternativeà l’induction à l’éthylène pour provoquer lamaturation des bananes.
L’utilisation de la couleur de la peau dela banane comme unique indicateur de ma-turation est erronée à cause des diffé-rences liées au temps, au lieu et à l’évalua-teur. Une mesure plus objective del’attribut ‘couleur de peau’ pourrait consti-tuer une étape ultérieure dans l’évaluationde la qualité des bananes. ■
Physiologie post-récolte, maturation et évaluationde la qualité des bananes (Musa spp.)Thèse de doctorat soumise en août 2000 à la Faculté des Sciences Agricoles et Biologiques Appliquées, Université Catholique de Leuven (KUL), Belgique
Thèses

Pour chaque poster, une présentation de 5 mn suivie de 5 mn de discussion seraplanifiée.
Les résumés devront être en anglais(250 mots maximum) et seront publiésdans African Plant Protection. Ils devrontêtre envoyés à l’adresse électronique sui-vante : [email protected] dans la der-nière version de Microsoft Word ou WordPerfect. Des copies imprimées peuventégalement être envoyées à M. Daneel,ARC-Institute for Tropical and SubtropicalCrops, Private Bag X11208, Nelspruit 1200,Afrique du Sud.Dates importantes :Soumission des résumés : 15 janvier 2001Date limite d’inscription : 15 janvier 2001Accusé de réception des résumés : 28 février 2001Adresse du congrès :Mieke Daneel - NSSA Organizing CommitteeARC-Institute for Tropical and SubtropicalCropsPrivate Bag X11208, Nelspruit, 1200,Afrique du SudTél. : (27) 13 7 532 071Fax : (27) 13 7 523 854Courrier électronique : [email protected] informations plus détaillées et des for-mulaires d’inscription sont disponibles à :http://www.inibap.org/actualites/nssa_eng.htm
IVe séminaire international sur la protection des plantes (2e annonce)Varadero, Cuba, 11–15 juin 2001Le Centro Nacional de Sanidad Agrope-cuaria (CENSA) et l’Instituto de Investi-gaciones de Sanidad Vegetal (INISAV) or-ganisent, en collaboration avec plusieursorganisations régionales, le IVe séminaireinternational sur la protection des planteset d’autres réunions sur ce thème qui setiendront à Varadero, Cuba, du 11 au15 juin 2001 au Palais des conventions« Plaza América ». Les langues officiellesseront l’anglais et l’espagnol.
Ce forum permettra de discuter des pro-blèmes, des résultats les plus récents etdes tendances dans le domaine de la phy-topathologie pour le prochain millénaire.Huit réunions scientifiques auront lieu etparmi elles, un atelier sur « les maladies etles ravageurs des bananiers : situation ac-tuelle et challenges pour le futur ».
Scientifiques, spécialistes et étudiantstravaillant dans ce domaine sont invités àsoumettre dès maintenant des résumés deleurs communications ou de leurs posters.Les résumés doivent être envoyés avant le30 mars 2001 par courrier électronique à :[email protected] avec copie au Pré-sident du comité d’organisation de la ré-union à laquelle ils souhaitent participer.En ce qui concerne l’atelier sur les mala-
INFOMUSA — Vol 9, N° 2 33
Ruth Stoffelen
La recherche a porté sur le criblage debananiers (Musa spp.) pour leur ré-sistance aux nématodes à lésions et à
galles. Le travail a été réalisé en troisétapes : 1) la standardisation de la procé-dure de criblage, 2) le criblage de 68 varié-tés Musa pour évaluer leur résistance auxnématodes pendant le premier stade decroissance végétative et 3) l’évaluation detrois aspects importants concernant les ex-périence de criblage : la variabilité de la ca-pacité de reproduction des populations desnématodes, la variabilité de l’organisation(architecture) des racines des différentesvariétés de bananier et l’interaction entrela reproduction des nématodes et le déve-loppement des racines.
Dans une première étape, une procédurede criblage pour l’évaluation de la résis-tance des bananiers aux nématodes à lé-sions et à galles pendant leur premier stadede croissance végétative a été développée etadaptée aux conditions de laboratoire (invitro). Pendant la deuxième étape, la résis-tance de 68 bananiers Eumusa et Australi-musa à Radopholus similis, Pratylenchuscoffeae et Meloidogyne spp. a été évaluéependant leur premier stade de croissancevégétative. Deux nouvelles sources de résis-tance à R. similis ont été identifiées dans lasection Australimusa (bananiers Fe’i). Au-cune source de résistance à P. coffeae etMeloidogyne spp. n’a été trouvée.
Dans la troisième étape, la reproductiondes nématodes, le développement des ra-cines et l’interaction entre ces deux para-mètres ont été examinés. Le pouvoir de re-production de quatre populations deR. similis cultivés sur des disques de ca-rotte a présenté une certaine variabilitécontrairement à celui de trois populationsde P. coffeae. Une certaine diversité dansl’architecture des racines (longueur de tousles types de racines) a été observée sur 11variétés de bananiers cultivés en milieu hy-droponique. Finalement, l’interaction entrela croissance des racines des bananiers etla reproduction des nématodes a été exami-
née pendant 4 mois. La croissance des ra-cines a été un processus dynamique consis-tant en une phase de croissance nulle sui-vie de trois phases de grande croissancedes racines. Les nématodes se sont repro-duites dans tous les types de racines ; leurreproduction a été conditionnée par la pré-sence de tissus racinaires frais. Cette étudea aussi révélé que la croissance des racinesdes bananiers et la reproduction des néma-todes sont influencées par l’environnement.
La procédure d’identification peut êtreoptimisée 1) en utilisant la même popula-tion des nématodes pour éviter l’influencede la variabilité de la capacité de repro-duction, 2) en incluant un cultivar de ba-nanier de référence susceptible aux néma-todes pour détecter les effetsenvironnementaux et 3) en adaptant l’ino-culation et l’analyse à la croissance des ra-cines des bananiers. ■
Annonces
15e congrès de nématologie sur la lutte intégrée contre les nématodesSkukuza, Afrique du Sud, 20-24 mai 2001La Nematological Society of SouthernAfrica (NSSA) annonce le 15e congrès denématologie sur la lutte intégrée contre lesnématodes qui se tiendra à Skukuza, dansle Parc national Kruger en Afrique du Suddu 20 au 24 mai 2001. Le congrès compren-dra des communications et des sessions deposters. Des ateliers sur de nouveauxthèmes peuvent également être organisés.Une visite dans les zones de production ba-nanière sera organisée ainsi qu’une ré-union de PROMUSA qui aura lieu à la suitele congrès du 24 au 26 mai 2001. La ré-union se tiendra à Cybele Guest House,Kiepersol, Mpumalanga, Afrique du Sud.
Le comité d’organisation invite les parti-cipants à présenter une ou plusieurs com-munications ou posters sur tous les aspectsde la recherche en nématologie. Lescontributions devront être présentées depréférence en anglais.
Les posters (110 cm x 90 cm) seront ex-posés pendant toute la durée du congrès.
Criblage précoce de bananiers Eumusaet Australimusa résistants aux nématodes à lésions et à gallesThèse de doctorat soumise en mars 2000 à la Faculté des Sciences Agricoles et Biologiques Appliquées, Université Catholique de Leuven (KUL), Belgique

dies et ravageurs des bananiers, contacterle Dr Luis Pérez Vicente à l ’une desadresses électroniques suivantes :[email protected] ; [email protected] ;[email protected] ou par courrier à :INISAVPO Box 634, 11300, PlayaLa Habana, CubaFax : (53 7) 332501Pour plus d’information générale,contactez :Dr Esther Lilia PeraltaPlant Protection DivisionCENSAPO Box 10, San José de las LajasLa Habana, CubaTél. : (53 64) 63014, Ext. 45Fax : (53 64) 63897Courrier électronique : [email protected] visitez les sites web suivants :http://www2.cuba.cu/ciencia/inisav/h t t p : / / w w w . i n i b a p . o r g / a c t u a l i t e s /varadero_eng.htm
Nouvelles de l’INIBAP
Réunions et exposition en ThaïlandeDu 4 au 12 novembre, une série de ré-unions et d’expositions a eu lieu à Bang-kok, en Thaïlande, dans le cadre d’un sym-posium international exceptionnel sur lesbananes qui était coorganisé par l’INIBAP,la Direction de la vulgarisation agricole etl’université Naresuan. Son Altesse RoyaleMaha Chakri Sirindhorn, Princesse deThaïlande, a ouvert les sessions techniquesle 6 novembre, après un week-end d’exposi-tions et de concours pour identifier lameilleure banane dessert, la meilleure ba-nane à cuire et les meilleurs produits àbase de banane. Parmi ceux qui étaientvenus exposer leurs travaux et leurs produits au Queen Sirikit’s NationalConvention Center se trouvaient, aux côtésdes principales institutions de rechercheagricole et entreprises industrielles thaï-landaises, des représentants d’organismesde toutes les régions du monde. L’INIBAPavait préparé des panneaux d’expositionspécialement pour la circonstance. Il aégalement mis cette occasion à profit pourprésenter le cédérom multimédia adaptéde la brochure sur les bananes et récem-ment traduit en anglais. Le centre de Transit de l’INIBAP et le réseau ASPNETont fait particulièrement bonne figure. De plus, chaque membre d’ASPNET avaitapporté des posters décrivant le travail de recherche-développement mené dans son pays.
Ensuite ont eu lieu trois jours de ré-unions structurées en sessions parallèles.Une session sur le transfert de technologiea rassemblé plus de 300 chercheurs ettechniciens de la région. Des présentationsont été faites sur les avancées et le poten-
34 INFOMUSA — Vol 9, N° 2
Vue partielle de l’exposition au Queen Sirikit’s National Convention Center.
Les produits présentés sur le stand de l’INIBAP ont été très appréciés.
a, b et c. Exposition des régimes de bananes ayant participé au concours. Au centre, la banane MotoEbanga (CRBP), primée pour son originalité.
S y m p o s i u m I n t e
a b

INFOMUSA — Vol 9, N° 2 35
Une jeune fille thai prépare des bonbons à labanane.
e r n a t i o n a l e n T h a ï l a n d e
Les produits à base de banane étaientégalement en compétition.
c

tiel des nouvelles technologies pour pro-duire des vitroplants, la gestion des affec-tions virales, l’amélioration variétale et lesbiotechnologies. Des représentants deChine, des Philippines, de Malaisie etd’Australie ont aussi présenté des exposés sur la production et le commercedes bananes dans leur pays.
L’ampleur de ces événements a offert uncadre idéal pour célébrer le 15e anniver-
saire de l’INIBAP et jeter un regard en arrière sur les acquis qui ont jalonné sonexistence.
10e réunion du Comité consultatifd’ASPNETLa 10e réunion du Comité consultatifd’ASPNET s’est déroulée les 10 et 11 no-vembre à Bangkok à l’invitation du Prof.Sujin Jinahyon, Président de l’Université
Naresuan et du Dr Ananta Dalodom, Direc-teur Général du Département de l’agricul-ture (DOA/Thailande). Émile Frison étaitinvité d’honneur. Des invités spéciaux duQUT, de l’IITA, de la KUL et du CIRAD sontintervenus sur les thèmes suivants : l’avan-cée de la recherche-développement desbiotechnologies dans le domaine bananier,l’application des biotechnologies en virolo-gie, le point sur l’amélioration des bana-
36 INFOMUSA — Vol 9, N° 2
Les 15 ans de l’INIBAP à Montpellier
A l’occasion de son quinzième anniversaire, l’INIBAP a organisé le 14 décembre dernier à Agropolis International à Montpellier, un séminaire d’une demi-journée sur le thème « Bananes et sécurité alimentaire ». Parmi les intervenants, on peut citer Messieurs Emil Javier, Président du “Technical Advisory Committee” du GCRAI, Gilles Saint-Martin, Secrétaire Éxécutifdu Comité interministériel pour la recherche agricole internationale, Jacky Ganry, Directeur adjoint du Cirad-Flhor, Henri Rouillé d’Orfeuil, Vice-Président du Forum mondial pour la recherche agronomique (GFAR), Louis Thaler, Professeur emeritus à l’Université de Montpellier II, Michel de Nucé de la Mothe,Président d’Agropolis et Emile Frison, Directeur de l’Inibap.L’ensemble des participants s’est accordé sur le fait que le mode de fonctionnement novateur et audacieux de l’INIBAP s’est avéré un réel succès. Ayant fait ses preuves au cours des quinze dernières années, il est maintenant repris comme modèle pour les orientations futures de la rechercheagronomique internationale.
Parallèlement au séminaire, Jean-Vincent Escalant animait un atelier pour enfants sur la diversité desbananiers à Agropolis Museum.
Jean Champion, bien connu des chercheursbananiers, a assisté au séminaire du 14 décembreen compagnie de son épouse Arlette.

niers, les avancées de la recherche en né-matologie, des études sur les populationsde Mycosphaerella fijiensis et le niveau derésistance des bananiers, et le point sur larecherche-développement sur le charan-çon du bananier.
Agustín Molina, Coordinateur régionaldu réseau ASPNET, a ensuite exposé lesdifférents points forts de l’année, entreautres les activités de recherche-dévelop-pement aux Philippines, la recherche ennématologie au Viêt-Nam, le projetICAR/NRCB-INIBAP en Inde, l’évaluationet l’adoption des hybrides de l’INIBAP auSecrétariat de la Communauté du Paci-fique, l’évaluation des variétés et la gestiondes maladies au Sri Lanka et l’annonced’un clone prometteur à Taiwan, cette der-nière information ayant suscité un grandintérêt parmi les participants.
L’IMTP et les activités des programmesnationaux d’évaluation, la propositionconcernant le centre national de dépôt,multiplication et distribution, le pro-gramme d’amélioration des Musa et les di-verses activités et collaborations ont étédiscutés au cours de la session de planifi-cation. J.V Escalant et S. Sharrock, dusiège de l’INIBAP, ont discuté avec lesmembres du Comité des deux niveauxd’évaluation de la phase III de l’IMTP :sites d’évaluation de la performance (10 accessions minimum) et sites d’évalua-tion approfondie (évaluation complète du matériel). Une étude préliminaire a étémenée pour évaluer l’implication des diffé-rents pays représentés à cette réunion. La participation du secteur privé dans lesévaluations a été évoquée. L’éventualitéd’intégrer un groupe de travail IMTP dansPROMUSA a également été soulevée. Unatelier pourrait être conduit afin d’homo-généiser les formats et le regroupementdes données.
A. Molina a présenté la propositionconcernant le centre national de dépôt,multiplication et distribution. En accordavec le plan, les gouvernements de chaquepays désignera une institution responsablede l’acquisition du matériel végétal del’ITC, de la multiplication et de la distribu-tion à l’intérieur du pays. Quelques repré-sentants ont indiqué qu’une telle disposi-tion existait déjà dans leur pays mais queles volets multiplication et distribution de-vaient être améliorés. Une recherche defonds pourrait être entreprise pour les paysqui ont besoin d’aide dans ce domaine.
Des nouvelles récentes du RISBAP ontété présentées par V. Roa. L’Australie, lesPhilippines, le Sri Lanka et Taiwan contri-buent activement à l’alimentation de labase de données. L’Australie a été félicitéepour sa campagne de promotion des basesde données de l’INIBAP. Il a également étéindiqué qu’une lettre d’agrément existeavec la Malaisie afin d’aider ce pays à ras-sembler les données.
En dernier point, une clarification desresponsabilités des membres du Comitéconsultatif a été demandée. Après délibé-ration, il a été décidé qu’un petit Comitéserait formé pour établir une liste decelles-ci qui sera diffusée auprès desmembres pour commentaires puis soumiseau Coordinateur régional.
Au cours de la réunion, le Prof. Det Wattanachaiyingcharoen a été élu Prési-dent du Comité consultatif d’ASPNET enremplacement du Dr Chen Houbin de laSouth China Agricultural University(SCAU), qui exerçait ce mandat depuis no-vembre 1999. Le Prof. Wattanachaiyingcha-roen exercera les fonctions de présidentjusqu’à la 11e réunion d’ASPNET qui setiendra au Sri Lanka en août 2001.
Prix Pisang RajaLors de la réunion du comité de pilotagedu réseau ASPNET, le prix Pisang Raja aété décerné à Rony Swennen en reconnais-sance de 21 années de travail remarquableen amélioration génétique des bananierset biotechnologies, au cours desquelles il aapporté une précieuse contribution à l’INIBAP et au réseau. Le prix a été égale-ment attribué à Benchamas Silayoi pourson immense contribution à la recherche-développement – dont témoignent plus de 20 articles techniques et livres sur la taxi-nomie, la génétique et l’amélioration desbananiers – et pour son association delongue date (depuis 1986) avec l’INIBAP et ses activités.
Gestion des viroses au BengladeshLes maladies représentent une contraintemajeure à la production bananière au Bengladesh. Des virus, tels que le BBTV, leBBrMV, le BSV et le CMV, sont particuliè-rement problématiques. Afin d’aider àtraiter ce problème, l’INIBAP a financé unatelier sur les maladies des bananiers etune formation sur l’indexation pour lesvirus. Ces réunions, les troisième du genreorganisées par ASPNET, se sont tenues enseptembre à l’Horticulture ResearchCentre, Bangladesh Agricultural Research Institute (HRC-BARI), Joydeb-pur, Bangladesh. Les précédentes ré-unions s’étaient déroulées aux Philippineset au Sri Lanka.
Cinquante deux participants ont assisté àl’atelier avec des intervenants de Taiwan,des Philippines et du Bengladesh. La nature, l’épidémiologie et la gestion desprincipaux ravageurs et maladies des bana-niers ont été passés en revue. L’accent aété mis sur l’importance de l’utilisation de culture de tissus indexées pour les virus,dont l’efficacité a été prouvée dans la ges-tion des viroses. Le Directeur général deBARI, Dr M.A. Razzque, a remercié l’INIBAP, le réseau ASPNET et le Prof.Hong-Ji Su de la National Taiwan
University, pour leur appui et leur volontéde réduire les problèmes liés aux maladieset aux ravageurs au Bengladesh. Il a af-firmé qu’un programme de gestion desvirus serait mis en place. Une formationpratique, conduite par le Prof. Hong-Ji Su,a été dispensée à neuf chercheurs du BARI.Deux procédures d’indexation, les tech-niques ELISA et PCR, ont été enseignées.
Nouvelles approches pour améliorer la productionbananière en Afrique de l’EstDepuis une vingtaine d’années, les rende-ments des plantations de bananiersconnaissent un déclin persistant enAfrique de l’Est. Ce phénomène sembledevoir être attribué à la pression crois-sante des ravageurs et des maladies (notamment les nématodes, les charan-çons et la cercosporiose noire) et à la dé-gradation des ressources naturelles. Lestentatives faites pour produire des variétésaméliorées dotées de caractéristiquespost-récolte localement acceptables àl’aide des méthodes conventionnelles seheurtent au degré élevé de stérilité de laplupart des variétés de bananiers d’alti-tude d’Afrique de l’Est. Cependant, lestechniques du génie génétique offrent uneautre voie pour améliorer les rendements.
Le Gouvernement ougandais finance,dans le cadre de sa contribution auGCRAI, un projet intitulé « Nouvelles approches pour améliorer la productionbananière en Afrique de l’Est – les appli-cations des biotechnologies » exécuté encollaboration avec l’IITA, la NARO, l’uni-versité de Makerere, le CIRAD, la KUL et l’INIBAP, qui en assure la coordinationet la supervision.
Ce projet a pour objectif d’améliorer lerendement des variétés existantes en lesdotant d’une meilleure résistance aux af-fections fongiques, aux nématodes et auxcharançons, tout en maintenant leurs ca-ractéristiques post-récolte et culinairesdésirables. Des approches complémen-taires seront mises en œuvre. Pour la résistance à la cercosporiose noire et auxnématodes, on se servira des techniquesde transformation à l’aide de diversesconstructions génétiques pour introduiredans des cultivars de bananiers d’altituded’Afrique de l’Est des gènes codant pourdes protéines antifongiques et des gènesoffrant un potentiel élevé pour contrôlerles nématodes. On identifiera les gènes les plus efficaces tout d’abord chez desplantes modèles transgéniques, puis dansdes essais en champ. Le pyramidage des gènes permettra d’obtenir une résis-tance multiple.
Pour la résistance aux charançons, deuxapproches seront utilisées. En collabora-tion avec l’IITA et la NARO, on étudiera lesmécanismes génétiques de la résistance àces ravageurs chez les bananiers d’altitude
INFOMUSA — Vol 9, N° 2 37

38 INFOMUSA — Vol 9, N° 2
d’Afrique de l’Est, afin d’identifier desgènes de résistance et des marqueurs gé-nétiques. Dans le même temps, on testera,en collaboration avec le CIRAD, les toxinesdes nouvelles souches de Bt qui agissentsur les Coleoptera afin de déterminer sielles ont une action efficace sur les cha-rançons. Si cette phase débouche sur desrésultats positifs, on recherchera un finan-cement en vue de transférer la résistance
aux charançons dans des cultivars de bana-niers d’altitude.
Une composante essentielle du projetconsistera à établir un centre d’expertiseen biotechnologies et à améliorer les ins-tallations de recherche en biologie molécu-laire qui existent déjà en Ouganda. Deschercheurs ougandais bénéficieront d’uneformation et d’une aide pour participer àdes réunions et séminaires internationaux.
Une réunion de planification et un ate-lier technique ont rassemblé tous les par-tenaires du projet au mois de septembreen Ouganda. Afin d’élaborer des plans
Partenaires du projet « Nouvelles approches pour améliorer la production bananière en Afrique del’Est – les applications des biotechnologies » lors de leur rencontre en septembre en Ouganda.
Visite au champ en Ouganda.
Le multimédia
BANANESCe document multimédia sur lesbananes vous est présenté par leRéseau international pourl’amélioration de la banane et de labanane plantain (Inibap) et leCentre de coopération internationaleen recherche agronomique pour ledéveloppement (Cirad).
Organisé autour de sept chapitres, ce CD-ROMprésente, grâce à une très riche iconographie(reportage, film d’époque, photos, cartes, bandesdessinées, etc.), l’ensemble des caractéristiques des filières bananes et plantain.
Présenté en démonstration à l’Exposition universelle deHanovre (Août 2000) et à la Journée professionnelle Banane-Ananas organisée par le Cirad-flhor à Montpellier (Septembre 2000).
Commandez dèsmaintenant votre CD-Rommultimedia !
Tarifs :Jusqu’à 50 exemplaires 80 F l’unitéDe 51 à 200 exemplaires 60 F l’unitéPlus de 200 exemplaires 40 F l’unitéTarif unitaire 150 F
Commande et règlement à renvoyer à :INIBAP
Parc Scientifique Agropolis II, 34397 Montpellier Cedex 5, FranceTél. : 33 - (0)4 67611302Fax : 33 - (0)4 67610334Email : [email protected]
• La plus grande herbe du
Monde
• L’histoire d’un fruit
voyageur
• Une nourriture de base
• La filière industrielle
de la banane dessert
• Une culture
sous surveillance
• Défendre et protéger
les cultures
• La banane,
légume ou dessert?

pour la mise en œuvre du projet, quatregroupes de travail ont été constitués surles suspensions cellulaires et la culture detissus, la recherche moléculaire, l’évalua-tion du matériel génétique et la recherchesur les charançons. Chacun de ces groupesa préparé un plan d’action détaillé, qui définit clairement les responsabilités dechaque partenaire. Des visites ont étéfaites aux laboratoires de biotechnologiede l’université de Makerere, à la station derecherche de la NARO à Kawanda et à lastation de recherche de l ’IITA à Namulonge afin d’examiner les facilitésdisponibles. On a engagé un processus derecrutement, identifié les étudiants endoctorat qui travailleront sur le projet, et commencé à établir des plantations. Leprojet sera entièrement opérationnel enjanvier 2001.
Site web de l’INIBAPDepuis juin dernier, la fréquentation dusite web de l’INIBAP a pratiquement dou-blé. Chaque jour sont enregistrées près de100 visites qui durent en moyenne plus detrois minutes chacune. Les bases de don-nées et les publications sont parmi les élé-ments les plus populaires. De nouvellespages web seront lancés prochainementpour les réseaux régionaux : MUSACO enAfrique occidentale et centrale, BARNESAen Afrique orientale et australe, MUSALACdans la région Amérique latine et Caraïbes,et ASPNET dans la région Asie et Paci-fique. Ces pages fourniront des informa-tions plus détaillées sur les activités, lesobjectifs et la composition des réseaux,ainsi que des statistiques mettant en lu-mière l’importance des bananes et des ba-nanes plantain dans ces régions. On peutactuellement télécharger à partir du site ledernier numéro d’INFOMUSA (vol. 9, 1),les actes du symposium sur « les produc-tions bananières : un enjeu majeur pour lasécurité alimentaire » (« Bananas andfood security »), « Organic/environmen-tally friendly banana production » (en version espagnole) et « Organic bana-nas 2000 : towards an organic bananainitiative in the Caribbean ». Égalementdisponibles sont le remarquable cataloguedes bananiers sauvages et cultivés de PaulAllen, et les résultats de la phase II del’IMTP (« Evaluating bananas : a globalpartnership »). Enfin, on a donné le « look » INIBAP ausite web de la banque de matériel géné-tique, qui est accessible à l ’adressehttp: / /www.agr .kuleuven.ac .be/dtp/tro/itc.htm
Initiative conjointe de l’IITA et de l’INIBAPLes deux centres Future Harvest menantdes activités de recherche-développementsur les Musa – l’IITA et l’IPGRI, au tra-vers de l’INIBAP – ont récemment décidé
d’intégrer leurs activités en Afrique. Enseptembre 2000, l’accord d’établissementd’un programme conjoint pour les bana-niers et bananiers plantain en Afrique aété finalisé au cours d’une réunion en Ouganda.
Bien que travaillant déjà en étroite col-laboration en Afrique, l’IITA et l’INIBAPont jugé nécessaire de combiner leurs pro-grammes complémentaires en un seul effort de recherche bien focalisé. Ils enten-dent ainsi maximiser l’impact de leurs pro-grammes sur les petits producteurs afri-cains de bananes et bananes plantain. Lenouveau programme conjoint sera mis enœuvre dans le cadre des deux réseaux derecherche bananière sous-régionaux diri-gés par les SNRA : BARNESA qui opèresous l’égide de l’ASARECA et MUSACO quiopère sous l’égide du CORAF.
Planification stratégiqueLa planification annuelle du programmeaura lieu lors des réunions des comités depilotage des réseaux BARNESA et MUSACO. À cette occasion, les parties pre-nantes procéderont à un examen globaldes activités de recherche et de tous les as-pects liés au développement de la produc-tion bananière dans chaque sous-région.Ces réunions serviront de cadre pour éta-blir des priorités et pour définir clairementles rôles et responsabilités des différentsacteurs de la recherche.
Conservation du matériel génétiqueL’IITA et l’INIBAP assumeront conjointe-ment la responsabilité de la conservation àlong terme du matériel génétique de Musad’origine africaine. Cela englobe la désigna-tion du matériel à inclure dans la collectiontenue en fidéicommis sous les auspices dela FAO, la duplication de l’ensemble du ma-tériel conservé et la mise à disposition del’information sur ce matériel au travers dela base de données SINGER.
Évaluation du matériel génétiqueDans le domaine de l’évaluation du maté-riel génétique, l’IITA et l’INIBAP ont desrôles clairement complémentaires. L’IITAcontinuera donc de privilégier l’évaluationprécoce des lignées dans le cadre d’essaismultilocaux, tandis que l’INIBAP axera da-vantage ses activités sur l’évaluation enamont dans le cadre du Programme inter-national d’évaluation des Musa (IMTP).
InformationLe bulletin MusAfrica, produit jusqu’àprésent par l’IITA, deviendra une co-pu-blication IITA-INIBAP. Il continuera deservir de forum pour permettre auxmembres des réseaux sous-régionauxd’échanger des informations sur la re-cherche bananière dans la région. D’autre
part, l’INIBAP et l’IITA rendront comptedes activités menées dans le cadre du pro-gramme conjoint dans leurs rapports an-nuels respectifs.
SensibilisationL’INIBAP et l’IITA conjugueront leurs ef-forts pour sensibiliser les décideurs et lepublic, afin de mobiliser davantage de res-sources pour les activités de recherche vi-sant à développer la production bananièredans la région.
Personnel de l’INIBAPAlberto VilarinhosDans le cadre de la Plate-Forme de re-cherche avancée d’Agropolis à Montpel-lier, l ’INIBAP et le CIRAD ont lancé
un projet conjointqui bénéficie d’unfinancement duG o u v e r n e m e n tfrançais. AlbertoVilarinhos, cher-cheur de l ’EM-BRAPA (Brésil)spécialisé dans lestechniques molé-culaires et la géné-tique du bananier,
mènera cette recherche pendant quatreans dans le cadre de la préparation de sonPhD. Le projet, intitulé « Cartographie despoints de césure des translocations chezMusa acuminata à l’aide de la cytogéné-tique moléculaire », consiste à utiliser denouvelles méthodes mises au point pourles animaux ainsi que pour le blé et leseigle, afin d’identifier les points où dessegments d’ADN, constituant des translo-cations, subissent une césure dans leschromosomes. Cela contribuera à ré-pondre à des questions concernant lesmodes de ségrégation, tout en facilitant la cartographie génétique et donc l’amélio-ration des bananiers. Alberto, après s’êtreinstallé en France avec sa famille, a commencé à travailler sur le projet en septembre.
Gaston BoussouGaston Boussou a été recruté temporaire-ment comme assistant pour la base de données MGIS. Il doit actualiser cette base
de données afin de préparer la pu-blication d’unedeuxième versionde Musalogue. Titulaire d’une maî-trise en informa-tion et documenta-tion de l’universitéde Montpellier, il aaussi des compé-tences en cartogra-
phie, gestion et qualité de l’eau, ainsi qu’enbiologie. Auparavant, il a travaillé sur la
INFOMUSA — Vol 9, N° 2 39

base de données Agritrop au CIRAD et afourni des informations bibliographiquespour MUSALIT, toujours au CIRAD.
Stijn MessiaenDepuis juillet 1998, Stijn Messiaen était dé-taché par l’INIBAP au Centre de recherchesrégionales sur bananiers et plantains(CRBP) basé à Nyombe, au Cameroun, entant qu’expert associé en entomologie del’Agence flamande de coopération au déve-loppement et d’assistance technique(Vlaamse Vereniging voor Ontwikkeling-ssamenwerking en Technische Bijstand,VVOB). Pendant deux ans, il a apporté unecontribution significative aux stratégies delutte intégrée contre le charançon Cosmo-polites sordidus, ravageur à l’origine de dé-gâts économiquement importants danstoutes les régions du monde où l’on produitdes bananes et des bananes plantain. Stijna travaillé sur les applications de la géné-tique (résistance de la plante hôte), les bioinsecticides (tourteaux de neem,[Azadirachta indica]), cendres ménagèreset feuilles de Tithonia incognita), lesagents microbiens (Beauveria bassiana),ainsi que les méthodes de lutte physiques(piégeage) et chimiques. Dans le cadre dela mise au point de techniques de luttecontre le charançon, il a effectué des re-cherches pour élucider le cycle biologiqueet le comportement de ce ravageur. Stijn adéjà publié une partie des résultats de sontravail dans des revues scientifiques et aplusieurs autres manuscrits en préparation.Parallèlement à ses recherches, il a aussiparticipé à la formation d’étudiants enstage au CRBP. Roger Fogain a supervisé letravail de Stijn au CRBP avec le concours deCliff Gold de l’IITA (Kampala, Ouganda) etde Dirk De Waele de la KUL (Belgique).Stijn rentre à présent en Belgique pour pré-parer un PhD à la KUL et nous lui souhai-tons de réussir dans cette entreprise.
Julie SchurgersDe novembre 1998 à juillet 2000, JulieSchurgers a travaillé comme stagiaire del’INIBAP au CRBP, où elle s’est intégréedans l’équipe qui a entrepris des re-cherches sur les suspensions cellulaires etl’embryogenèse somatique. Dans un pre-mier temps, elle a coordonné les réponsesaux demandes d’envoi de plantules raci-nées et de cultures proliférantes ducentre régional de distribution de maté-riel génétique mandaté par l’INIBAP pourl’Afrique occidentale et centrale. Au coursdes sept derniers mois, elle a étudié lestechniques d’utilisation des marqueursmoléculaires dans l’amélioration des ba-naniers pour la résistance aux nématodesau laboratoire d’agronomie tropicale de laKUL, en Belgique, sous la direction duprofesseur Dirk De Waele. Nous lui sou-haitons plein succès dans la poursuite desa carrière en Belgique.
Livres, etc.
Banana cultivar names andsynonyms in Southeast AsiaR.V. Valmayor, S.H. Jamaluddin, B. Silayoi,S. Kusumo, L.D. Danh, O.C. Pascua etR.R.C. EspinoISBN : 971-91751-2-5
La région de l’Asie du Sud-Est abrite la plusgrande richesse de la diversité bananièredu monde, aussi bien en formes sauvagesqu’en formes cultivées ; elle comprendaussi quelques variétés très rares dans lemonde. Mais, leur classification ainsi queleur nomenclature ont toujours été un pro-blème très compliqué. Dans la plupart descas, les mêmes variétés portent des nomsdifférents en des endroits différents alorsque dans d’autres les mêmes noms sontdonnés à des variétés différentes.
En septembre 1999 une réunion décisiveincluant les conservateurs de différentescollections nationales de matériel géné-tique bananier a été organisée aux Philip-pines. Les participants sont arrivés à ré-soudre le problème de la nomenclatured’environ 300 variétés de bananiers et sesont mis d’accord sur un système de classi-fication à l’instar de celui de Cheeseman etdu Code International de Nomenclaturepour les Plantes Cultivées. Ce livret de 24pages fournit une liste finale des noms etsynonymes des variétés acceptés à la ré-union. Ce livret est un compagnon indis-pensable pour les collecteurs, les conserva-teurs, les améliorateurs et les chercheursdu monde entier !
Disponible en anglais auprès du bureaurégional de l’INIBAP pour l’Asie et le Paci-fique, c/o IRRI Collaborators Centre, Col-lege, Laguna 4031, Philippines.
Advancing banana and plantain R & D in Asia and the PacificCompte-rendu de la 9e réunion du ComitéConsultatif Régional INIBAP – ASPNETorganisée à l’Université Agricole de laChine du Sud, Guangzhou, Chine, 2-5 novembre 1999Edité par A.B. Molina et V.N. RoaISBN : 971-91751-3-3Pour la première fois le compte-rendu dela réunion du Comité Consultatif Régional
de l’ASPNET est publié sous la forme d’unlivre de 154 pages. Cette publication pré-sente les informations les plus récentes surla recherche sur Musa, le développementet les perspectives d’avenir en Malaisie, enIndonésie, dans les Iles du Pacifique, auSri Lanka, au Bengladesh, aux Philippineset un rapport approfondi sur la situationbananière en Chine. Il fournit aussi lesdernières informations sur l’état des nomset des synonymes des bananiers cultivés enAsie du sud-est ; les résultats des étudessur les nématodes des bananiers couram-ment cultivés au Vietnam, la biodiversitédes bananiers sauvages dans le nord de la Thaïlande et les progrès de rechercherécents sur la fusariose à Taiwan.
Des copies peuvent être obtenues au-près du bureau régional de l’INIBAP auxPhilippines.
Managing banana and citrusdiseasesCompte-rendu d’un atelier de travailrégional sur la gestion des maladies des bananiers et des agrumes par l’utilisation du matériel de plantation sain, organisé à Davao City, Philippines, 14-16 Octobre 1988Edité par A.B. Molina, V.N. Roa, J. Bay-Petersen, A.T. Carpio et J.E.A. JovenISBN : 971-91751-1-7Les agrumes et les bananiers sont les prin-cipales cultures de rente des paysans del’Asie du Sud-Est. Cependant, leurs rende-ments sont en train de diminuer de ma-nière dramatique à cause de diverses mala-dies ; ces infections sont tellementrépandues que même les pépinières et lespieds-mères généralement utilisés pour re-peupler les plantations détruites en sontatteints et le matériel végétal sain doit êtrechercher ailleurs. Les progrès dans la ges-tion des maladies des agrumes et des bana-niers ont permis de développer des nou-velles voies permettant de produire et dedistribuer efficacement des plantules noninfectées et de développer des méthodesde détection très sensibles. Celles-ci per-mettent même de détecter des infectionslatentes et celles qui se développent sans
40 INFOMUSA — Vol 9, N° 2

INFOMUSA — Vol 9, N° 2 41
symptômes extérieurs. Dans le cas des ba-naniers, le matériel indemne est obtenupar la culture de tissus. Quant auxagrumes, la distribution des plantules cer-tifiées indemnes de maladies provenantdes pépinières saines est encouragée. Les leçons tirées du développement desprogrammes de gestion intégrée des mala-dies des agrumes et des bananiers sontaussi d’une grande utilité aux paysans pratiquant d’autres types de cultures.
L’atelier de travail sur la ‘gestion desmaladies des bananiers et des agrumes’ a
été conjointement organisé par l’INIBAP etle Food and Fertilizer Technology Centerbasé à Taiwan. Il a rassemblé les cher-cheurs engagés au premier plan dans lalutte contre les maladies des bananiers etdes agrumes en Asie du Sud-Est. Leur tra-vail sur l’épidémiologie, l’écologie, la dé-tection et les méthodes de gestion des ma-ladies virales des bananiers et des agrumesest présenté dans ce livre de 164 pages. Celivre contient aussi spécifiquement les ré-sultats des études sur l’épidémiologie, lacaractérisation et la gestion des virus dubunchy top et de la mosaïque en tirets. Lesrecommandations pour la recherche, le développement et la politique découlant de cet atelier unique en son genre y sontégalement incluses.
Cette publication est disponible enanglais auprès du bureau régional del’INIBAP aux Philippines.
Organic banana 2000 :towards an organic bananainitiative in the CaribbeanRapport de l’atelier international sur la production et la commercialisation des bananes biologiques par les petits producteurs, 31 octobre-4 novembre 1999, Santo Domingo, République DominicaineEdité par M. Holderness, S. Sharrock,E. Frison et M. KairoISBN : 2-910810-40-2La diversification n’a jamais été aussi im-portante qu’elle ne l’est aujourd’hui pour
les paysans engagés dans l’économie demarché. Les pays des Caraïbes ont décou-vert le créneau du marché des bananesbiologiques, marché dans lequel la Répu-blique Dominicaine est actuellement laplus grande exportatrice.
Cette réunion, organisée conjointementpar l’INIBAP, CAB International et leCTA, a été un forum de discussion etd’échange d’informations pour diversgroupes d’intérêt, allant des paysans auxdétaillants. L’objectif de la réunion étantd’initier une action visant à soutenir la pro-duction et l’exportation des bananes biolo-giques dans les Caraïbes.
Ce rapport de 174 pages, publié en an-glais, présente des articles sur l’état actuelde la production de la banane et en parti-culier le développement de la productionbiologique aussi bien dans les Caraïbesqu’en Amérique Centrale, en Amérique La-tine et au Cameroun ; il présente aussi lesperspectives du marché de la banane biolo-gique en Amérique du nord et en Europe,les aspects de certification et d’assurancede qualité et les contraintes de production.Les résultats des groupes de travail et lesconclusions de la réunion y sont aussi in-
clus. En bref, cette publication présentedes débats importants sur les derniers pro-blèmes concernant la production biolo-gique des bananes et formule un plan d’ac-tion détaillé pour l’avenir.
Contacter l’Unité Information/Communi-cation au siège de l’INIBAP pour recevoirune copie du livre.
Nouvelles IPM : Biocontrol Newsand InformationISSN : 0143-1404Le numéro de septembre 2000 de BiocontrolNews and Information (Vol. 21, n° 3) publié par CABI Bioscience a une trèsforte saveur de banane. Presque tout lenuméro de ce bulletin d’information estconsacré à la gestion intégrée des rava-geurs des bananiers. Les articles donnentdes informations sur la recherche sur lalutte intégrée contre les ravageurs (IPM)en Ouganda, au Brésil, au Ghana, auPérou, au Cameroun, en Guadeloupe, en
Australie et au Costa Rica. Tout en cou-vrant d’autres aspects de l’IPM, beaucoupde pays se concentrent surtout sur l’IPMdes charançons du bananier et le contrôledes nématodes. Les aspects additionnelscouverts par ces pays comprennent la ges-tion intégrée des maladies des bananiers,la gestion de la mosaïque en tirets du ba-nanier, les systèmes de prévision visantune utilisation réduite des pesticides etdes méthodes de contrôle non chimiquesdes maladies post-récoltes.
Ce numéro inclut deux contributions dela part du personnel de l’INIBAP. ‘IPM andINIBAP’ décrit l’importance de l’IPM dansles activités de l’INIBAP et la contributionfaite par l’INIBAP pour promouvoir l’IPM àtravers des ateliers, des publications,l’appui apporté à la recherche, la conser-vation et la distribution du matériel géné-tique. Les aspects suivants résument bienl’engagement spécifique de l’INIBAP dansles différentes stratégies d’IPM : le travailen Asie pour développer des programmesde contrôle des virus, le succès dans lalutte contre la maladie du sang en Indoné-sie, la mobilisation de l’IPM pour une pro-duction durable en Afrique et les essais enchamp en Afrique de l’Est. ‘L’approche in-tégrée pour contrôler les charançons auCameroun’ (Integrated approach for weevil control in Cameroon) esquisse lesdivers tests IPM en cours dans ce pays. Leneem et la cendre de bois, les champi-gnons entomopathogènes, les insecticideschimiques et les pièges aux phéromonessont parmi les mécanismes actuellementtestés. Le CRBP inclut aussi la résistancecontre les charançons dans son pro-gramme d’amélioration.
Internet Round-UpToujours dans la dernière édition de Bio-control News and Information, la section‘Internet Round-up’ présente une liste desites web sur les bananiers et la luttecontre les ravageurs que nous publions ci-dessous avec la permission de l’éditeur : Lesite du Consortium for International CropProtection (CICP) présente une compila-tion d’informations et de sites Internet surla lutte intégrée contre les ravageurs quicouvrent un grand éventail de questionssur l’IPM, incluant la lutte biologique :http:/ /www.ippc.orst .edu/cicp/fruit/banana.html
Le site de l’INIBAP lui-même est également recommandé pour la gestion des ra-vageurs et pour les liens intéressants qu’ilpropose : http://www.inibap.org
Le Department of Primary Industries,Queensland, publie les “notes DPI” qui don-nent des conseils sur la gestion des rava-geurs, ainsi que des informations pratiquesindiquant comment gérer divers problèmesliés aux ravageurs et aux maladies concer-nant diverses plantes alimentaires, in-

42 INFOMUSA — Vol 9, N° 2
cluant les bananes. L’adresse du site est :http://www.dpi.qld.gov.au/dpinotes
Des informations sur les aspects poli-tiques concernant les bananes, une sectionsur les bananes biologiques et une liste dedocuments utiles sont actuellement compi-lés et seront bientôt disponibles en ligneau site de Banana Link : http://www.geocites.com/NapaValley/1702/
L’Australian Banana Growers Council(ABCG) présente des activités en cours,ses programmes de recherche et des lienssur d’autres sites bananiers à : http://www.abgc.org.au
Des rapports de projets et des résumésissus du Programme mondial du CGIAR surl’IPM sont disponibles en ligne à :http://www.cgiar.org/spipm
L’Institut International d’AgricultureTropicale (IITA) gère actuellement un pro-jet intitulé ‘Improving plantain- and ba-nana-based systems’, dont un résumé estdisponible à : http://www.cgiar.org/spipm/dbase/projects/iitaipd.html
Le DfID’s Crop Protection Programmepropose également le résumé d’un projetsur le développement de la résistance auxnématodes chez les bananiers et les bana-niers plantain à : http://vwww.netcom.net.uk/~n/nri/pcpp/r6391.htm
Des détails sur le travail de CABI Bios-cience sur les nouvelles techniques de dé-tection et de caractérisation des champi-gnons causant la fusariose et lescercosporioses sont disponibles à : http://www.cabi.org/bioscience/annualreport_projects_egham.htmpdn
Le Florida Entomologist présente un ar-ticle sur le charançon du bananier enAfrique de l’est intitulé ‘Timing and dis-tribution of attack by the banana weevil(Coleoptera: Curculionnidae) in EastAfrican highland banana (Musa spp.)’ àl’adresse suivante : http://www.fcla.edu/FlaEnt/fe82p631.htm
On trouve un article d’Hort Digest sur lamouche du bananier Opogona sacchari(Bojer). à : http://www.hortdigest.com/archives/2-2000/bananamoth.htmet uneprésentation des recherches du Cirad surce thème sur le site d’Agropolis à http://www.agropolis.fr/actualiteevenements/lettre/spe1099gb/integraprotec.html
Cette série de liens publié dans la sec-tion ‘Internet Round-up’ de ‘BiocontrolNews and Information’ est reproduite avecla permission de CAB International. Bio-control News and information est dispo-nible sur le site de CABI Bioscience :www.cabi.org
CD-Rom MusaDocLa deuxième édition du CD-Rom MusaDoc,MusaDoc 2000 est sortie ! Les versions actualisées des bases de données de l’INIBAP, MUSALIT (contenant les résu-més et les données bibliographiques despublications sur Musa) et BRIS (la base de
données sur les chercheurs travaillant surle bananier) sont disponibles. Sont égale-ment présentes sur MusaDoc 2000 toutesles publications récentes dont la très popu-laire brochure ‘Les bananes’, les nouvellesfiches techniques et les publications pro-duites cette année telles que le RapportAnnuel 1999, ‘Banana cultivar names andsynonyms in Southeast Asia’, ‘Evaluatingbananas : a global partnership’ et ‘Lesproductions bananières : un enjeu écono-mique majeur pour la sécurité alimen-taire’. Le CD-Rom fournit aussi un résuméillustré des activités de l’INIBAP. Mainte-nant, n’importe qui peut être connectémême s‘il n’est pas sur le web !
Les demandes devront être adressées di-rectement à l’Unité Information/Communi-cation au siège de l’INIBAP.
IMTP 2000Autre CD-Rom disponible : le CD-RomIMTP 2000 qui sera bientôt égalementconsultable sur le web. IMTP 2000 est lefruit des premiers efforts de l’INIBAP vi-sant à rendre largement disponible la tota-
lité des résultats des phases I et II del’IMTP sous forme de base de données. LeCD-Rom contient aussi des copies desguides techniques et la publication ‘Eva-luating bananas : a global partnership’laquelle donne une analyse détaillée desrésultats de la Phase II. Il contient aussi lecatalogue des clones candidats et les
clones de référence (y compris ceux quisont disponibles pour les essais de la PhaseIII) ainsi que l’accord sur le transfert dematériel génétique. En somme, tous les ou-tils indispensables pour participer à laPhase III de l’IMTP sont disponibles.
Les demandes des copies de l’IMTP 2000devraient être adressées au Coordinateurde l’IMTP au siège de l’INIBAP.
Postcosecha y agroindustría del plátano en el eje cafetero de ColombiaEdité par D.G. Cayón Salinas, G.A. GiraldoGiraldo et M.I. Arcila PulgarínISBN : 958-96885-0-0
Ce livre de 265 pages, rédigé en espagnol,présente les résultats de recherches sur lapost-récolte et l’utilisation des sous-pro-duits du bananier plantain dans la zone ca-féière centrale de Colombie, travaux réali-sés au travers de collaborations entreCorpoica, la Universidad del Quindío, leComité de Cafeteros del Quindío et Col-ciencias.
La première partie fait le point sur lesprincipaux processus biochimiques et phy-siologiques qui régulent et contrôlent lapost-récolte des végétaux, en mettant plusparticulièrement l’accent sur les fruits cli-matériques comme la banane plantain. Ony discute également des principales tech-nologies de transformation du produit. Sui-vent 20 articles scientifiques et techniquesrésultant des recherches sur la post-ré-colte de la banane plantain réalisées parles différents instituts de la zone caféièrecentrale de Colombie. La publication decet ouvrage a été rendue possible grâce àl’appui technique et financier de la Asocia-ción para la Investigación en Plátano –ASIPLAT et des instituts ayant participéaux recherches.
Pour obtenir un exemplaire de cet ou-vrage, contacter Gerardo Cayón Salinas,Corpoica – Armenia. Avenida Bolívar Sec-tor Regivit 28 Norte, Armenia, Quindío, Co-lombie. Courrier électronique : [email protected]. Prix : US$10plus frais de port.

INFOMUSA — Vol 9, N° 2 43
SiègeParc Scientifique Agropolis II34397 Montpellier Cedex 5 - FRANCEE-mail : [email protected]://www.inibap.orgDirecteur :Dr Émile FRISONE-mail : [email protected] des Recherches sur les Ressources GénétiquesDr Jean-Vincent ESCALANTE-mail : [email protected] de la Conservation du Matériel GénétiqueMelle Suzanne SHARROCKE-mail : [email protected] de l’Information et de la Communication :Melle Claudine PICQE-mail : [email protected] du MGISMelle Élizabeth ARNAUDE-mail : [email protected] Financier :Mr Thomas THORNTONE-mail : [email protected]
Bureau Régional pour l’Amérique latine et les CaraïbesCoordinateur Régional : Dr Franklin E. ROSALES
Expert associé, transfert de technologies :Luis POCASANGREC/o CATIEApdo 60-7170 Turrialba, COSTA RICATel/Fax : (506) 556 2431E-mail : [email protected]
Bureau Régional pour l’Asie et le PacifiqueCoordinateur Régional : Dr Agustín MOLINAC/o IRRI Collaborators Center3rd FloorLos Baños, Laguna 4031PHILIPPINESFax : (63 2) 845 05 63E-mail : [email protected]
Bureau Régional pour l’Afrique occidentale et centraleCoordinateur Régional : Dr Ekow AKYEAMPONGC/o CRBP - BP 12438Douala, CAMEROUNTel/Fax : (237) 42 91 56E-mail : [email protected]
Bureau Régional pour l’Afrique orientale et australeCoordinateur Régional : Dr Eldad KARAMURAExpert associé, transfert de technologies :
Guy BLOMME PO Box 24384 Kampala, OUGANDAFax : (256 41) 28 69 49E-mail : [email protected]
Centre de Transit INIBAP (ITC)Responsable :Melle Ines VAN DEN HOUWEKatholieke Universiteit LeuvenLaboratory of Tropical Crop ImprovementKasteelpark Arenberg 13,B-3001 Leuven, BELGIQUEFax : (32 16) 32 19 93E-mail : [email protected]
Experts associés, NématologieInge VAN DEN BERGHC/o VASIVan Diem, Than TriHanoi, VIET-NAMFax : (84 4) 86 13 937E-mail : [email protected] MOENSC/o CORBANAStation de recherche La RitaApdo 390-7210Guápiles, COSTA RICAFax : (506) 763 30 55E-mail : [email protected]
Les textes dactylographiés seront préparésen français, anglais ou espagnol et envoyésau rédacteur en chef de la revue. Ils serontprésentés en double interligne. Toutes lespages seront numérotées (y compris les ta-bleaux, figures, légendes et références) àpartir de la page de titre. Le titre sera leplus court possible. Mentionnez le nomcomplet de tous les auteurs ainsi que leuradresse au moment de l’étude. Indiquezégalement l’auteur auquel doivent êtreadressées les correspondances.Si le texte a été saisi sur un système informa-tisé, merci d’envoyer avec votre version im-primée une copie sur disquette ou par cour-rier électronique en indiquant les référencesdu logiciel de traitement de texte utilisé.RésumésUn résumé dans la langue du texte et éven-tuellement dans les deux autres langues dela revue devra accompagner la contribution.Il ne devra pas excéder 200 à 250 mots.SiglesIls seront développés lors de leur premièreapparition dans le texte et suivis du sigleentre parenthèses.
BibliographieLes références bibliographiques serontprésentées par ordre alphabétique d’au-teurs. L’appel à référence dans le texte in-diquera le nom de l’auteur et l’année depublication (ex : Sarah et al. 1992).Vous trouverez ci-dessous trois exemplesde références parmi les plus courantes :Articles de périodiques : Sarah J.L., C. Bla-vignac & M. Boisseau. 1992. Une méthodede laboratoire pour le criblage variétal desbananiers vis-à-vis de la résistance aux né-matodes. Fruits 47(5): 559-564.Livres : Stover R.H. & N.W. Simmonds.1987. Bananas (3rd edition). Longman,Londres, Royaume Uni. Articles (ou chapitres) de publicationsnon-périodiques : Bakry F. & J.P. Horry.1994. Musa breeding at CIRAD-FLHOR. Pp.169-175 in The Improvement and Testingof Musa: a Global Partnership (D.R. Jones,ed.). INIBAP, Montpellier, France. TableauxNumérotez-les et faites référence à ces nu-méros dans le texte. Chaque tableau seraaccompagné d’un titre.
IllustrationsNumérotez-les et faites référence à ces nu-méros dans le texte. N’oubliez pas d’indi-quer les légendes.Graphiques : Merci de fournir avec le gra-phique les données brutes correspon-dantes.Dessins : dans la mesure du possible, four-nir des originaux.Photographies noir et blanc : elles doiventêtre tirées sur papier brillant et trèscontrastées.Photographies en couleur : fournir un trèsbon tirage papier ou des diapositives debonne qualité.
Note : Les auteurs citant dans leur articledu matériel végétal originaire du Centre detransit de l’INIBAP (ITC) à Leuven ou in-dexé dans ce centre indiqueront les numé-ros de code ITC des accessions citées.
Merci de suivre ces conseils.Cela facilitera et accélérera le travail
d’édition.
Conseils aux auteurs
Les adresses de l’INIBAP

Disponibles au Siège central à Montpellier :
INIBAP 2000. M. Holderness, S. Sharrock, E. Frison & M. Kairo (eds). Organic banana 2000:Towards an organic banana initiative in the Caribbean. Report of the internationalworkshop on the production and marketing of organic bananas by smallholder farmers.31 October-4 November 1999, Santo Domingo, Dominican Republic.
INIBAP 2000. Annual Report 1999.
INIBAP 2000. G. Orjeda (compil.). Evaluating bananas: a global partnership. Results ofIMTP Phase II.
INIBAP/EARTH/IDRC 1999. F.E. Rosales, S.C. Tripon & J. Cerna (eds).Organic/environmentally friendly banana production. Proceedings of a workshop held at EARTH, Guácimo, Costa Rica, 27-29 July 1998 (in press).
INIBAP/CRBP/CTA/CF 1999. C. Picq, E. Fouré & E.A. Frison (eds). Bananas and FoodSecurity/Les productions bananières : un enjeu économique majeur pour la sécuritéalimentaire. Proceedings of an International Symposium held in Douala, Cameroon, 10-14 November 1998.
INIBAP/FHIA 1999. F.E. Rosales, E. Arnaud & J. Coto (eds). A tribute to the work of PaulAllen : a catalogue of wild and cultivated bananas.
INIBAP/RF/SDC 1999. E.A. Frison, C.S. Gold, E.B. Karamura & R.A. Sikora (eds). MobilizingIPM for sustainable banana production in Africa. Proceedings of a workshop on bananaIPM held in Nelspruit, South Africa, 23-28 November 1998.
INIBAP 1999. E. Akyeampong (ed.). Musa Network for West and Central Africa. Report ofthe second Steering Committee meeting held at Douala, Cameroon, 15-16 November1998.
INIBAP 1999. Annual Report 1998.
INIBAP 1999. K. Shepherd. Cytogenetics of the genus Musa.
INIBAP 1998. E. Akyeampong (ed.) Musa Network for West and Central Africa. Report ofthe first Steering Committee meeting held at Douala, Cameroun, 8-10 Decembre 1998.
INIBAP 1998. E.A. Frison & S.L. Sharrock (eds). Banana streak virus: a unique virus-Musainteraction? Proceedings of a workshop of the PROMUSA virology working group held inMontpellier, France, 19-21 January 1998.
INIBAP 1998. C. Picq (ed.). Segundo seminario/taller de la Red regional de informaciónsobre banano y plátano de America Latina y el Caribe. San José, Costa Rica, 10-11 deJulio 1997.
INIBAP 1998. B.K. Dadzie. Post-harvest characteristics of black Sigatoka resistant banana,cooking banana and plants hybrids. INIBAP Technical Guidelines 4.
INIBAP 1998. G. Orjeda, en collaboration avec les groupes de travail de PROMUSA sur lafusariose et les cercosporioses. Évaluation de la résistance des bananiers auxcercosporioses et à la fusariose. Guides techniques INIBAP 3.
CIRAD/INIBAP 1998. Les bananes.
INIBAP/ACIAR 1997. E. Arnaud & J.P. Horry (eds). Musalogue, a catalogue of Musagermplasm: Papua New Guinea collecting missions 1988-1989.
INIBAP/CTA/FHIA/NRI/ODA 1997. B.K. Dadzie & J.E. Orchard. Evaluation post-récolte deshybrides de bananiers et bananiers plantain : critères et méthodes. Guides techniquesINIBAP 2.
INIBAP/CTA 1997. P.R. Speijer & D. De Waele. Evaluation du matériel génétique de Musapour la résistance aux nématodes. Guides techniques INIBAP 1.
INIBAP/The World Bank 1997. E.A. Frison, G. Orjeda & S. Sharrock (eds). PROMUSA: AGlobal Programme for Musa Improvement. Proceedings of a meeting held in Gosier,Guadeloupe, March 5 and 9, 1997.
INIBAP-IPGRI/CIRAD 1996. Descripteurs pour le bananier (Musa spp.).
Disponibles directement auprès du bureau régionald’Asie/Pacifique
INIBAP-ASPNET 2000. R.V. Valmayor, S.H. Jamaluddin, B. Silayoi, S. Kusumo, L.D. Danh,O.C. Pascua & R.R.C. Espino. Banana cultivar names and synonyms in Southeast Asia.
INIBAP-ASPNET 2000. A.B. Molina & V.N. Roa (eds). Advancing banana and plantain R & Din Asia and the Pacific. Proceedings of the 9th INIBAP-ASPNET Regional AdvisoryCommittee meeting held at South China Agricultural University, Guangzhou, China, 2-5November 1999.
INIBAP-ASPNET/FFTC 2000. A.B. Molina, V.N. Roa, J. Bay-Petersen, A.T. Carpio and J.E.A.Joven(eds). Managing banana and citrus diseases. Proceedings of a regional workshopon disease management of banana and citrus through the use of disease-free plantingmaterials held in Davao City, Philippines, 14-16 October 1998.
INIBAP-ASPNET 1999. V.N. Roa & A.B. Molina (eds). Minutes: Eighth meeting of INIBAP-ASPNET Regional Advisory Committee (RAC) hosted by the Queensland HorticultureInstitute (DPI) in Brisbane, Australia, 21-23 October 1998.
INIBAP-ASPNET 1998. Minutes: Seventh meeting of INIBAP-ASPNET Regional AdvisoryCommittee (RAC) hosted by the Vietnam Agricultural Science Institute (VASI) in Hanoi,Vietnam, 21-23 October 1997.
INIBAP-ASPNET 1997. V.N. Roa & R.V. Valmayor (eds). Minutes: Sixth meeting of INIBAP-ASPNET Regional Advisory Committee (RAC) hosted by National Research Center onBanana (ICAR) in Tiruchirapalli, India, 26-28 September 1996.
INIBAP-ASPNET 1996. R.V. Valmayor, V.N. Roa & V.F. Cabangbang (eds). RegionalInformation System for Banana and Plantain – Asia and the Pacific (RISBAP):Proceedings of a consultation/workshop held at Los Baños, Philippines, 1-3 April 1996.(ASPNET Book Series No. 6).
Les publications de l’INIBAP
ww
w.inib
ap.o
rg

PROMUSA IINFOMUSA — Vol 9, N° 2
PROMUSA N° 6
Sommaire
Procès-verbal de la deuxième réunion du Comité de pilotage de PROMUSA tenueles 7 et 8 novembre 2000 à Bangkok (Thaïlande) . . . . . . . . . . . . . . . . . . . . .p. I
Réunion du Groupe de travail de PROMUSAsur l’amélioration génétique . . . . . . .p. IV
Réunion du Groupe de travail de PROMUSAsur les cercosporioses . . . . . . . . . . .p. VII
Réunion du Groupe de travail de PROMUSAsur la fusariose . . . . . . . . . . . . . . . . .p. X
Réunion du Groupe de travail de PROMUSAsur la nématologie . . . . . . . . . . . . . .p. XII
Réunion du Groupe de travail de PROMUSAsur la virologie . . . . . . . . . . . . . . . .p. XIV
Qu’est-ce que PROMUSA ?
Le programme mondial pour l’améliorationdes bananiers (PROMUSA) est un pro-gramme qui cherche à impliquer les princi-paux acteurs de l’amélioration des bananiers.Il est un moyen de relier le travail mené surles problèmes des producteurs travaillantpour l’exportation et les initiatives dans le do-maine de l’amélioration de la production d’au-tosubsistance et à petite échelle pour les mar-chés locaux. Le programme mondial est basésur les acquis de la recherche et se construitsur les recherche en cours. PROMUSA estdonc un mécanisme qui permet de maximiserles résultats et d’accélérer l’impact de l’effortmondial en matière d’amélioration des bana-niers. Ce mécanisme novateur, qui permet decatalyser les recherches menées tant à l’inté-rieur qu’à l’extérieur du GCRAI, favorise lacréation de nouveaux partenariats entre lesSystèmes nationaux de recherche agricole(SNRA) et les instituts de recherche dans lespays développés et dans les pays en voie dedéveloppement. La création de tels partena-riats contribue aussi à renforcer la capacitédes SNRA à conduire des recherches sur lesbananiers.
L’une des initiatives majeures de PRO-MUSA est le développement d’un largeéventail de nouveaux hybrides de bananiercorrespondant aux différentes attentes despetits producteurs du monde entier. Le pro-gramme rassemble à la fois les acteurs del’amélioration conventionnelle, basée sur lestechniques d’hybridation et ceux travaillantsur des approches liées au génie génétiqueet aux biotechnologies. Cet effort en matièred’amélioration génétique s’appuie sur les re-cherches menées sur des ravageurs et desmaladies spécifiques dans le cadre des diffé-rents groupes de travail de PROMUSA. Lemécanisme efficace mis en place pour éva-luer les nouvelles variétés produites dans lecadre de PROMUSA est une autre compo-sante essentielle du programme.
PROMUSAUn programme mondial pour l’amélioration des Musa
Présents : Émile Frison (INIBAP) – mo-dérateur, Abdou Tenkouano (IITA), MaryWabule (SNRA d’Afrique orientale etaustrale), Luis Sequeira (IRA d’Amériquedu Nord), Adiko Amoncho (SNRAd’Afrique occidentale et centrale), Eliza-beth Aitken (IRA d’Australie – région duPacifique), Philippe Lepoivre (IRA d’Eu-rope), David Berroa (SNRA d’Amériquelatine et des Caraïbes), Luc Sas (prési-dent du Groupe de soutien à PRO-MUSA), Jean-Vincent Escalant (secré-taire de PROMUSA), Suzanne Sharrock(rapporteur). Observateurs : Cliff Gold (IITA), EldadKaramura (INIBAP). Excusé : P. Faylon (SNRA d’Asie et duPacifique).
Groupe de soutien à PROMUSALe rôle du Groupe de soutien à PRO-MUSA a été expliqué aux membres duComité de pilotage. Il s’agit d’un groupeinformel de donateurs et autres partiesprenantes, avec lesquels des réunionsont lieu pour discuter des différents as-pects du développement de la productionbananière et maintenir un intérêt pourcelle-ci. Ces réunions servent avant toutà échanger des informations et ne don-nent normalement lieu à aucun engage-ment financier.
Rôle du Comité de pilotageIl a été souligné que cette réunion devaitêtre considérée comme la première
réunion officielle du Comité de pilotagede PROMUSA, celui-ci étant à présentconstitué de membres nommés en bonneet due forme par les différents groupesqu’ils représentent. Le rôle et les respon-sabilités du Comité, tels qu’ils ont été dé-finis et publiés précédemment, ont étéconfirmés. De même, il a été confirméque le secrétariat de PROMUSA estchargé d’établir les rapports d’activité etde les soumettre au Comité de pilotage.
Durée du mandat etparticipation aux réunionsEtant donné la nécessité d’une continuitédans la composition du Comité de pilo-tage, il a été décidé que ses membresauront un mandat de deux ans, pouvantêtre renouvelé trois fois. Il a été proposéque la représentation de l’IITA et del’IPGRI soit institutionnelle plutôt que no-minative, de façon que le même repré-sentant puisse siéger au sein du Comitéplus longtemps que la durée maximalenormale de six ans. Il a été convenuqu’en cas d’ impossibi l i té pour unmembre d’assister à une réunion, il (ouelle) devra désigner une personne pourle (ou la) remplacer.
Financement des projets de PROMUSAIl a semblé qu’il y avait une certaineconfusion chez les membres de PRO-MUSA en ce qui concerne le rôle du Se-crétariat et du Comité de pilotage dans larecherche de financements pour les pro-
Procès-verbal de la deuxième réunion du Comité de pilotage de PROMUSA tenue les 7 et 8 novembre 2000 à Bangkok (Thaïlande)

II PROMUSA INFOMUSA — Vol 9, N° 2
jets. Les membres du Comité ont doncjugé nécessaire de préciser que le rôleprincipal de PROMUSA est de faciliter lacollaboration et les partenariats, et nonde trouver des fonds. Ils ont toutefoisnoté que le Secrétariat peut, sur de-mande, aider des membres de PROMUSA à élaborer des requêtes definancement pour des projets. S’agissantdes projets conçus dans le cadre de PROMUSA, ils ont estimé que ceux-ci nedoivent normalement pas être évaluéspar le Comité de pilotage, car cela ajou-terait un degré de bureaucratie supplé-mentaire au processus d’élaboration des projets.
Rapports sur les activités de PROMUSARapports au Comité de pilotageIl a été constaté, durant la séance d’ou-verture de la réunion de PROMUSA, quecertains des rapports des responsablesde liaison des groupes de travail étaientde nature très générale et ne faisaientpas véritablement ressortir les progrèsaccomplis dans les deux dernières an-nées. Le secrétaire de PROMUSA a si-gnalé que les responsables de liaison onteu des difficultés à obtenir un feedbackdes membres des groupes de travail etque, de manière générale au cours deces deux années, ils n’ont pas joué unrôle suffisamment actif. À cet égard, lesmembres du Comité ont estimé que lerôle du facilitateur n’avait pas été définide manière assez précise et ils ont notéque les groupes de travail devaient discu-ter de cette question durant la réunion encours. Concernant les rapports, il a étéconvenu que le Secrétariat soumettra unrapport au Comité de pilotage tous lessix mois. Son travail serait facilité si l’ondemandait aux responsables de liaisondes groupes de travail de fournir un rap-por t au Secrétar iat avec la même fréquence.
Nouvelles de PROMUSALes membres du Comité ont félicité l’INIBAP pour les nouvelles de PRO-MUSA publiées deux fois par an dans INFOMUSA. Ils ont noté qu’on est entrain d’actualiser le site Web de PRO-MUSA, qui deviendra ainsi un autre canalimportant pour partager l’information et
communiquer les résultats du pro-gramme.
Partage de l’information au sein de PROMUSALes membres du Comité ont soulignél’importance des listes de messagerieélectronique pour partager l’information,notamment avec les chercheurs qui n’ontpas facilement accès à Internet. Cepen-dant, ils ont observé que, pour l’instant,ces messageries ne sont pas utiliséescorrectement. Cela tient à plusieurs rai-sons, et notamment au fait que les cher-cheurs ne sont pas habitués à travaillerde cette manière, de sorte que les com-munications ne se font souvent qued’une personne à une autre. Il a été re-connu qu’il faudrait encourager les res-ponsables de liaison et les membres desgroupes de travail à se servir davantagede ces messageries.
Contacts entre les groupes de travailIl faudrait s’efforcer d’améliorer les inter-actions entre les groupes de travail. À ceteffet, il a été décidé de créer un « Comitédes facilitateurs » de l’ensemble desgroupes de travail. Ce Comité tiendraune réunion formelle avant chaque ré-union mondiale de PROMUSA. Il a étéjugé également nécessaire de dévelop-per les contacts informels entre les res-ponsables de liaison.
Feedback des réseaux régionauxLes membres du Comité ont convenuque le Secrétariat de PROMUSA doitprendre des initiatives pour stimuler leséchanges d’information entre PROMUSAet les réseaux régionaux. Il s’agit ainsid’assurer que PROMUSA réponde véri-tablement aux besoins existant à l’éche-lon régional.
Financement de PROMUSAIl a été souligné que PROMUSA n’a pasété créé pour servir de mécanisme de fi-nancement. Cependant, le fonctionne-ment du programme suppose desmoyens financiers et il convient donc detrouver les fonds nécessaires. À cetégard, il ne semble guère probable qu’unfinancement suffisant soit assuré à brèveéchéance par les donateurs.
Les membres du Comité se sont ac-cordés à reconnaître que les participantsà PROMUSA doivent s’associer au finan-
cement du programme, par exemple encouvrant les coûts de leur participationaux réunions. Ils ont estimé que la priseen charge de ces coûts contribueraitaussi à démontrer aux donateurs poten-tiels la valeur du programme. Ils ont rap-pelé que certains participants l’avaientdéjà fait pour la présente réunion et pourla précédente. Il a été décidé que, pourles grands projets mis en place dans lecadre de PROMUSA, les coûts des ré-unions pourraient être inclus dans le bud-get du projet.
Il a été convenu que tous les membresdu Comité devaient faire des efforts pourassurer le suivi des contacts avec les do-nateurs, de façon à contribuer à trouverdes financements pour PROMUSA. À cetégard, il a été souligné que le Secrétariatdevra tenir le Comité informé des re-quêtes préparées pour des projets et desprogrès accomplis dans la recherche deleur financement.
Réunions de PROMUSAIl a été noté que le symposium internatio-nal sur la biologie moléculaire et cellu-laire des bananiers, qui doit avoir lieu

INFOMUSA — Vol 9, N° 2 PROMUSA III
tous les deux ans, offrirait un contexteapproprié pour les réunions mondialesde PROMUSA. Le coût de l’organisationdes réunions mondiales a été discuté et ila été proposé de faire des efforts pourcontacter des fondations et des entre-prises privées susceptibles d’apporterune aide pour les prochaines réunions.
Les membres du Comité ont estiméqu’en combinant les réunions futures dePROMUSA avec le symposium en ques-tion, on faciliterait l’identification de fi-nancements par les participants, quipourraient bénéficier par exemple desubventions couvrant les frais de voyageà une réunion scientifique. La liaison desdeux événements aurait aussi pour avan-tage de permettre aux membres dePROMUSA de prendre connaissancedes résultats scientifiques récents pré-sentés au symposium.
Nouvelles initiatives sous l’égide de PROMUSAConsortium sur la génomique du bananierLe Secrétariat de PROMUSA a présentéun rapport au Comité de pilotage au
sujet du Consortium sur la génomique dubananier. Créé à Montpellier au début del’année, celui-ci vise à élaborer un « plandirecteur » pour la génomique du bana-nier et à instaurer de libres échangesd’ informations entre les membres du Consor tium, tout en l imitant leséchanges d’informations en dehors decette structure. Le travail du Consortiumdoit être axé sur la recherche pré-compé-titive et tous les résultats des rechercheseffectuées par ses membres seront mis àdisposition gratuitement. Le Consortiuma demandé au Comité de pilotage d’enté-riner le placement de cette initiative sousl’égide de PROMUSA. Il a été préciséque le Consortium possède son propre « comité scientifique » et qu’un certaindegré de confidentialité entoure le dérou-lement de ses activités. Les membres du Consortium appartiennent tous augroupe de travail de PROMUSA surl’amélioration génétique. Les membresdu Comité de pilotage ont pris note dufait que le travail du Consortium est axésur l’amélioration génétique des bana-niers, et que sa finalité et ses objectifscoïncident avec ceux de PROMUSA. Ilsont reconnu que le Consortium assure
aux chercheurs un mécanisme leur per-mettant d’avoir un certain degré de confi-dentialité autour de leur travail, tout en seconformant à l’esprit de collaboration quiest celui de PROMUSA. Ils ont donc ac-cepté que le Consortium sur la géno-mique du bananier opère sous l’égide dePROMUSA.
Les charançons du bananier : une nouvelle priorité pour PROMUSALors de la création de PROMUSA, lescharançons du bananier n’étaient pasencore reconnus comme une contraintepour la production bananière à l’échellemondiale et l’amélioration génétiquen’était pas considérée comme la techno-logie la plus appropriée pour combattreces ravageurs. Cependant, la situation aévolué au cours de ces dernières an-nées. Les outils de l’amélioration géné-tique ont considérablement progressé etl’on a identifié des sources de résistanceaux charançons. L’amélioration génétiquepour la résistance aux charançons, àl’aide des méthodes conventionnelles etdes biotechnologies, apparaît donc au-jourd’hui comme une option viable. Dansce contexte, le Secrétariat de PROMUSA

IV PROMUSA INFOMUSA — Vol 9, N° 2
a reçu plusieurs demandes pour qu’oninscrive la résistance contre les charan-çons au programme du groupe de travailsur l’amélioration génétique et qu’un nou-veau groupe de travail sur les charan-çons soit constitué.
Après avoir débattu de la question, lesmembres du Comité ont décidé de créerun groupe informel sur les charançons.Ce groupe devra rassembler des infor-mations plus précises sur les thèmes etles sites des recherches actuellementmenées dans ce domaine, afin de pré-senter un rapport à la prochaine réuniondu Comité de pilotage.
Réseau sur les biotechnologies en Amérique latineDes informations ont été données sur unréseau qui a été créé en Amérique latinedans le cadre de MUSALAC pour déve-lopper l’application des biotechnologiesau bananier. Ce réseau est axé sur latransformation génétique et la génétiquemoléculaire. Les membres du Comitéont estimé qu’il fallait encourager ce ré-seau à s’intégrer à PROMUSA dans lecadre du groupe de travail sur l’améliora-tion génétique.
SensibilisationIl a été noté que les membres de PRO-MUSA participent à un grand nombre deréunions internationales. Il a donc étéproposé que les groupes de travail élabo-rent des posters que les membres pour-ront présenter à ces réunions. Le Secré-tariat fournira une aide pour préparerces posters.
Élection du modérateurde PROMUSAÉmile Frison a été réélu modérateur dePROMUSA pour 2000-2001.
SecrétariatLes par t icipants ont confirmé que l’INIBAP continuera d’assurer le Secréta-riat de PROMUSA.
Fréquence des réunions du Comité de pilotageIl a été confirmé que le Comité de pilo-tage doit s’efforcer de se réunir une foispar an. Des réunions supplémentaires
pourront être organisées en cas de besoin. La date et le lieu de la pro-chaine réunion du Comité seront fixésultérieurement.
Réunion du groupe detravail de PROMUSAsur l’améliorationgénétiquePar t icipants : Mar ia Elena Agui lar(CATIE, Costa Rica), Françoise Carreel(CIRAD, Guadeloupe), François Cote(CIRAD, France), James Dale (QUT, Aus-tralie), Jaroslav Dolezel (IEB, Tchèquie),Antonio Figueira (CENA USP, Brésil),Rafael Gomez Kosky (IBP, Cuba), PeterGresshoff (CRCTPP, Australie), Pat Hes-lop-Harrison (JIC, Grande-Bretagne),Christophe Jenny (CIRAD, Guadeloupe),Dieter Kaemmer (Frankfurt Univ. Biocen-ter, Allemagne), Dale Krigsvold (FHIA,Honduras), Gomez Lim (CINVESTAV,Mexique), Do Nang Vinh (AGI, Viêt-nam),Sebastiao de Oliveira e Si lva (EM-BRAPA, Brésil), Luis Pérez Vicente (IN-IVIT, Cuba), Michael Pillay (IITA, Nigeria),Nicolas Roux (IAEA, Autriche), LazloSagi (KUL, Belgique), S. Sathiamoorthy(NRCB, Inde), Jorge Sandoval (COR-BANA, Costa Rica), Rony Swennen(KUL, Belgique), Abdou Tenkouano (IITA,Nigeria), Kodjo Tomekpe (CRBP, Came-roun), S. Uma (NRCB, Inde), P. K. Valsa-lakumari (KAU, Inde).Rapporteurs : Jaroslav Dolezel et S. Sathiamoorthy.
Questions générales et administrativesS. Sathiamoorthy (NRCB) et J. Dolezel(IEB) ont été nommés facilitateurs ausein du groupe de travail, le premierétant chargé du sous-groupe sur l’amé-lioration et la génétique, et le second dusous-groupe sur le génie génétique.
Les participants ont évoqué le pro-blème de l’insuffisance des fonds mis àla disposition de la recherche sur les ba-naniers et les bananiers plantain, comptetenu de l’importance de ces culturesdans le monde. Le développement de larecherche dans le domaine de la géno-mique, qui peut avoir un impact sur tous
les aspects de l’amélioration des bana-niers, ne pourra se faire sans des inves-tissements considérables. D’autre part, legroupe a noté qu’il entre dans les fonc-tions de PROMUSA de :• contribuer à établir des priorités
internationales/mondiales pour larecherche et l’amélioration ;
• être une plate-forme decommunication/interaction ;
• être un dépositaire d’informations etaider à les diffuser (nouvellespublications, outils, matériel) ;
• organiser des réunions – réunionsmondiales de PROMUSA et réunionssatellites spécialisées ;
• identifier et contacter des bailleurs defonds potentiels ;
• sélectionner les appels d’offres pourdes projets et diffuser l’information ;
• aider au montage de projets.Les participants ont défini comme suit lerôle des facilitateurs : • préparer l’ordre du jour des réunions
du groupe ; • diriger les discussions durant les
réunions ; • stimuler les interactions entre les
membres du groupe dans l’intervalleentre deux réunions ;
• accompagner le secrétariat dePROMUSA pour rencontrer lesdonateurs potentiels.La dimension du groupe et la nature
approfondie des discussions n’ont paslaissé de temps pour présenter les rap-ports scientifiques ou avoir des échangesde vues avec les autres groupes de tra-vail. Il faudrait envisager de diviser le groupe de travail sur l’amélioration gé-nétique en deux groupes de dimensionplus réduite.
Sous-groupe sur la génétiqueet l’hybridationLes priorités de recherche suivantes ontété retenues pour le sous-groupe sur lagénétique et l’hybridation : • établir un programme de collaboration
à l’échelon régional pour l’améliorationdes bananiers en Asie, principal centred’origine et de diversité de cette plante;
• effectuer des prospections dans leszones de diversité naturelle,notamment en Asie du Sud et du Sud-

INFOMUSA — Vol 9, N° 2 PROMUSA V
Est, en vue de collecter de nouveauxtypes sauvages/natifs ;
• caractériser et évaluer les variétés afind’identifier de nouvelles sources derésistance aux principaux ravageurs etmaladies – cercosporioses,nématodes, fusariose et charançon(s’attaquant à la souche et aupseudotronc) ;
• rassembler les informationsdisponibles sur la diversité génétiquedes bananiers dans le monde ;
• mettre l’accent sur l’obtention dediploïdes afin de créer de nouvelleslignées à l’aide des méthodesclassiques et des outils desbiotechnologies ;
• élargir la base génétique à l’aide desapproches conventionnelles et desbiotechnologies.
Sous-groupe sur le géniegénétiqueAu sein du sous-groupe sur le génie gé-nétique, il n’a pas été jugé nécessaire demodifier les priorités globales et les stra-tégies établies lors des deux précé-dentes réunions de PROMUSA. Toute-fois, les participants ont noté qu’il est deplus en plus urgent de stimuler le déve-loppement de la génomique du bananieret que cet objectif doit recevoir la plushaute priorité. Le Consortium sur la gé-nomique du bananier, créé en avril 2000à l’initiative de PROMUSA, est appelé àjouer un rôle majeur dans ce domaine.On a aussi signalé la création en Amé-rique latine d’un réseau pour l’applicationdes biotechnologies au bananier. Il fau-drait envisager d’établir des liens plusétroits entre les deux groupes.
Cultures de tissus et de cellules in vitroUne méthodologie pour le développe-ment de suspensions cellulaires embryo-gènes a été mise au point ; elle est encours de transfert dans différents labora-toires. Les cultures embryogènes sontconsidérées comme un matériel expéri-mental approprié pour la transformationgénétique, la mutagenèse et l’isolementde protoplastes chez le bananier. La prin-cipale contrainte étant l’instabilité géné-tique des cellules cultivées en suspen-
sion, il a été fortement recommandé detester en champ les plants régénérés. S’ilest jugé irréaliste d’étudier la nature etles mécanismes de la variation génétiquein vitro, il est urgent d’isoler des mar-queurs pour au moins quelques-uns deshors-types les plus fréquemment régéné-rés à partir des suspensions. Ces mar-queurs seraient précieux pour effectuerdes criblages précoces et ils pourraientégalement servir à optimiser les condi-tions de culture afin de réduire l’instabilitégénétique. La culture de tissus doit aussiêtre considérée comme un outil dansl’obtention d’hybrides améliorés, l’éradi-cation des virus et la cryoconservation àlong terme.
Transformation génétiqueDes progrès importants ont été accom-plis dans la mise au point de systèmesde transformation efficaces pour les ba-naniers. Cette technologie est actuelle-ment transférée dans différents labora-toires, et au moins cinq laboratoires dusecteur public ont déjà obtenu des plantsde bananiers transgéniques. De manièregénérale, les protocoles utilisant Agro-bacterium reçoivent la préférence parrapport à la biolistique, car ils sont plusefficaces et produisent moins de copiesintégrées et une moindre fréquence duphénomène de mise en silence destransgènes. Si la plupart des projets encours visent à transformer des cultivarsexistants, il a été proposé d’étendre l’ap-plication de cette technologie aux géni-teurs diploïdes et aux espèces natives,et d’intégrer les plants transgéniquesdans les programmes d’améliorationexistants. La principale contrainte tientau fait que les systèmes de sélectionfont intervenir la résistance aux herbi-cides et aux antibiotiques, ce qui risquede rendre difficile l’acceptation des bana-niers et bananiers plantain transgé-niques par le public. Il existe un intérêtgénéral pour collaborer sur les questionsde biosécurité ainsi que sur l’utilisationdes cultures de suspensions cellulairesembryogènes pour la transformation. Enrevanche, dans le domaine des promo-teurs et des vecteurs, il n’est prévuqu’une collaboration limitée en raisond’éventuels problèmes relatifs aux droitsde propriété intellectuelle.
MutagenèseLa mutagenèse physique, par irradiationgamma, a été appliquée avec succès parquelques projets de recherche qui signa-lent l’obtention de plusieurs clones dotésde caractères agronomiques intéressants(précocité, hauteur réduite, résistanceaux maladies, rendement accru). Lesprincipales contraintes sont le chimé-risme et la nécessité de cribler des popu-lations de plantes importantes. Il est ur-gent de mettre au point des systèmes invitro pour dissocier rapidement les chi-mères, ainsi que des systèmes de cri-blage rapide des caractères désirés. Si laplupart des projets en cours sont axéssur l’induction de mutations chez des cul-tivars triploïdes existants, il a été proposéde produire davantage de mutants à par-tir de géniteurs diploïdes et d’espèces na-tives. Outre l’irradiation gamma, il faudraitemployer la technique de l’irradiation parneutrons rapides. D’autre part, l’utilisationde produits chimiques et la mutagenèseinsertionnelle doivent être considéréescomme des alternatives à l’irradiation. Lamutagenèse se révèle être un outil inté-ressant pour obtenir des plantes possé-dant des caractères qui ne sont pas pré-sents dans la nature. Elle devrait jouer unrôle de plus en plus important dans lesprojets de cartographie du génome dubananier, dans le cadre desquels elle ser-vira à produire des stocks de délétions etdes mutants par élimination.
CytogénétiqueDes progrès importants ont été réalisésdans le domaine de la cytogénétique desbananiers. La cytométrie en flux del’ADN est aujourd’hui largement accep-tée pour déterminer rapidement le degréde ploïdie et cette méthode a été trans-férée à deux stations d’amélioration gé-nétique. Elle a également été utiliséepour vérifier le degré de ploïdie des ac-cessions de la banque de gènes (ITC)de l’INIBAP à Leuven. Étant donné laproportion relativement élevée des ac-cessions dont le degré de ploïdie n’a pasété confirmé (≈ 9 %), il est fortement re-commandé d’analyser les autres collec-tions. D’après des résultats récents, lacytométrie en flux est une méthode ap-propriée pour évaluer rapidement laconstitution génomique d’accessions ethybrides inconnus, et pour détecter

VI PROMUSA INFOMUSA — Vol 9, N° 2
l’aneuploïdie. Deux laboratoires (AIEA etIEB) offrent ce service aux institutionsqui ne possèdent pas les équipementsnécessaires. Des méthodes de cytogé-nétique moléculaire (hybridation in situen fluorescence – FISH, et hybridationgénomique in situ – GISH) ont été misesau point. La FISH sert à analyser lastructure des chromosomes de bananierau niveau moléculaire, y compris la dis-tribution génomique d’éléments géné-tiques mobiles et d’intégrants du BSV. LaGISH est extrêmement utile pour déter-miner la constitution génomique des hy-brides. La caractérisation détaillée ducaryotype de bananier, incluant l’identifi-cation des différents chromosomes et ladétection des remaniements structurelsdes chromosomes, sera une priorité àcour t terme dans le cadre des re-cherches en cytogénétique.
AneuploïdieL’aneuploïdie pourrait devenir un outil im-portant dans les études génétiques surles bananiers. Des résultats récents ontmontré qu’il est possible de produire degrandes quantités d’aneuploïdes (pour laplupart des hypoploïdes, du fait de laper te d’un ou de plusieurs chromo-somes). On a obtenu des aneuploïdesaprès régénération de cultures de sus-pensions cellulaires embryogènes, aprèsirradiation et après traitement avec dessubstances qui sont des poisons pour lefuseau mitotique. Il s’avère que la cyto-métrie en flux de l’ADN permet de détec-ter rapidement les plantes ayant unnombre de chromosomes qui ne corres-pond pas au nombre de base. Une foisobtenus, les plantes aneuploïdes peu-vent être maintenues par multiplicationvégétative. Jusqu’ici, on a obtenu desaneuploïdes à partir de matériel triploïde.Il a été proposé de produire une séried’aneuploïdes à partir de génotypes di-ploïdes sélectionnés et de procéder àleur caractérisation.
Mise en silence de gènes et interaction des génomesIl importe d’effectuer des études pluspoussées sur les phénomènes de miseen silence de gènes et d’interaction desgénomes, car ils pourraient avoir des ré-percussions importantes sur les pro-
grammes d’amélioration des bananiersen cours. La plupart de ces programmescomportant la création d’hybrides, lamise en silence de gènes et l’interactiondes génomes peuvent influer sur les ca-ractéristiques du matériel nouvellementobtenu. L’altération de l’expression d’ungène peut constituer un mécanisme im-portant pour engendrer une variabilitégénétique in vitro, tandis que la mise ensilence d’un gène peut modifier l’expres-sion des transgènes. D’après des résul-tats récents, il s’avère que le génome Ade Musa intervient dans l’activation desséquences du BSV qui sont intégréesdans le génome B. L’élucidation des mé-canismes sous-jacents à ce phénomènepourrait fournir des outils pour créer dumatériel résistant au BSV.
GénomiqueLe groupe de travail a identifié la géno-mique du bananier comme sa premièrepriorité de recherche. Malgré l’impor-tance des bananiers et des bananiersplantain dans le monde, les études dansce domaine accusent du retard par rap-port à celles entreprises sur d’autresgrandes cultures. Les informations déri-vées de ces études devraient contribuerà la création de cultivars capables defournir une performance optimale dansdes conditions biologiques, écologiqueset culturales différentes.
Les discussions ont abordé les troisgrands aspects de la génomique du bananier :
Cartes génétiquesIl est urgent de parvenir à une carte gé-nétique saturée du bananier. Trois popu-lations sont actuellement disponiblespour la cartographie au CIRAD/CRBP, àl’université du Queensland et à l’IITA. Denouvelles populations en ségrégeantesdoivent être établies par l’INIBAP en col-laboration avec le CIRAD. Il a été recom-mandé que les laboratoires échangentdes marqueurs moléculaires et collabo-rent pour augmenter la densité descartes. Il faudrait s’efforcer de développerdes marqueurs STMS (sites microsatel-lites à séquences répertoriées) qui sontspécifiques de locus, codominants et for-tement polymorphes. Bien que les STMSsoient onéreux, une fois qu’ils sont éta-blis, la méthodologie est facilement trans-
férable. Le groupe a également soulignéla nécessité de transférer dans d’autreslaboratoires les populations utilisées pourla cartographie.
Cartes physiquesIl est indispensable de disposer rapide-ment de cartes physiques, ainsi que decartes génétiques et physiques intégréespour le séquençage. Les ressources sui-vantes sont nécessaires pour la cartogra-phique physique : banques de BAC(chromosomes bactériens artificiels),banques d’ADNc (ADN complémentaire)et EST (étiquettes de séquences expri-mées). Bien qu’aucun investissementstratégique n’ait été effectué dans la gé-nétique du bananier au cours des der-nières années, on dispose déjà d’unecertaine quantité de matériel : deuxbanques de BAC (Calcutta 4) sont dispo-nibles (contre paiement) au Texas A&MUniversity BAC Centre, de nouvellesbanques de BAC sont actuellementconstituées par le CIRAD, le CICY etl’IEB, et le CINVESTAV a établi unebanque d’ADNc de feuilles de bananier.En outre, Zenecca a développé plus de80 000 EST. Si elle est une nécessité ur-gente, la cartographie physique exigerades investissements considérables enmatériel, produits consommables et per-sonnel qualif ié. Actuellement, deux laboratoires (CIRAD et IEB) sont équipésdu matér iel automatisé requis pourconstruire et manipuler les banquesd’ADN et pour préparer des puces àADN, et un laboratoire (UL) est en traind’acheter le matériel nécessaire pourproduire des microarrays. Afin de rationa-liser l’utilisation des ressources exis-tantes, il importe de coordonner les ef-forts de recherche en cours.
Découverte de gènesSi le clonage des gènes sur la base descartes demeure une voie intéressantepour isoler des gènes, les projets de re-cherche doivent aussi envisager d’autresapproches : génomique comparative, cri-blage différentiel et étiquetage des gènes.
Sélection assistée par des marqueursMalgré son intérêt pour l’amélioration desbananiers, la sélection assistée par desmarqueurs apparaît encore essentielle-ment comme une aspiration pour l’avenir.

INFOMUSA — Vol 9, N° 2 PROMUSA VII
Cela s’explique par le retard enregistrédans le développement de la génomiquedu bananier et par le manque de mar-queurs moléculaires liés à des carac-tères intéressants comme la résistance àdes maladies ou la parthénocarpie. Ac-tuellement, on dispose seulement demarqueurs permettant de détecter rapi-dement la présence des génomes A et B.
Évaluation du matérielgénétique (biodiversité)Le nouveau matériel collecté lors desmissions de prospection, ainsi que le ma-tériel des collections existantes (notam-ment celles d’Asie) devront faire l’objetd’une caractérisation approfondie à diffé-rents niveaux (morphologique, agrono-mique, moléculaire). Ce faisant, il faudradéterminer le nombre de chromosomes,la dimension du génome et la constitu-tion génomique. On dispose de mar-queurs moléculaires (RFLP, STMS,CAPS, AFLP) pour classer et identifier lematériel génétique de bananiers. LeCIRAD s’en est servi pour caractériserdes accessions de la banque de gènesde l’INIBAP à Leuven. La classificationd’une proportion relativement élevée deces accessions (≈ 10 %) n’a pas étéconfirmée, ce qui montre qu’il est indis-pensable d’analyser aussi les autres col-lections. Les membres du groupe de tra-
vail ont proposé que l’évaluation des col-lections in vitro soit faite avec des dupli-cations en champ. Ils ont également sug-géré d’améliorer l’accès à la base dedonnées MGIS.
Réunion du groupe detravail de PROMUSAsur les cercosporiosesParticipants : E. Aitken (CRCTPP, Austra-lie), C. Abadie (CRBP/CIRAD, Came-roun), J.-P. Busogoro (Faculté de Gem-bloux, Belgique), J. Carlier (CIRAD,France), B. Fullerton (HortResearch,Nouvelle-Zélande), A. Gutierrez Rojas(CORPOICA, Colombie), M. Guzman(CORBANA, Costa Rica), Z. Jiang(SCAU, Chine), N. Masdek (MARDI, Ma-laisie), G. Molina (INIBAP), W. Tusheme-reirwe (NARO, Ouganda), L. Pérez Vi-cente (INISAV, Cuba), G. Rivas (CATIE,Costa Rica), R. Selvarajan (NRCB, Inde),B. Williams (QDPI, Australie).Faci l i tateurs : Jean Car l ier et Bob Fullerton.Rapporteur : Jean Carlier
IntroductionChaque participant a tout d’abord pré-senté un bref bilan des activités de son
propre groupe de recherche. Puis desdiscussions ont eu lieu sur l’identificationet la distribution des agents pathogènesdes bananiers en Asie. La durabilité de larésistance aux maladies a également faitl’objet d’un débat. Les participants ontensuite passé en revue et actualisé lespriorités de recherche. Celles-ci sont pré-sentées ci-dessous. Les sources d’infor-mation et des commentaires figurententre parenthèses, afin de faciliter lapoursuite des échanges au sein dugroupe et avec les autres groupes de tra-vail. Enfin, les participants ont discutédes avantages apportés par PROMUSAet ont proposé des activités à incluredans le programme.
Priorités de recherche
1. Détermination de la distribution etdes fréquences relatives de Mycos-phaerella eumusae, M. fijiensiset M. musicola dans différents paysd’Asie du Sud-EstÀ la suite de l’article paru dans Phytopa-thology (Carlier et al. 2000), on a adopté« Septoria leaf spot » (en français, septo-riose du bananier) comme nom communde la maladie causée par Mycosphae-rella eumusae (stade anamorphe Septoriaeumusae). Cette maladie a peut-être étéobservée précédemment en Inde (DrSelvarajan, NRCB). Il est prévu de pu-
Réunion du groupe de travail sur l’amélioration génétique.

VIII PROMUSA INFOMUSA — Vol 9, N° 2
blier une description latine officielle deMycosphaerella eumusae afin d’établir lavalidité de cette espèce (QDPI, CIRAD).
La présence de cet agent pathogèneest actuellement confirmée dans le sudde l’Inde, au Sri Lanka, en Thaïlande, enMalaisie, à Maurice, à la Réunion et auNigeria (CIRAD). En revanche, elle n’apas encore été rencontrée en Chine (DrJiang, SCAU).
La distribution et l’importance relativedes trois agents pathogènes distincts quisont à l’origine des cercosporioses enAsie sont pour l’essentiel inconnues. Afinde répondre à ces questions, on a entre-pris une enquête en Inde (NRCB), enMalaisie (MARDI), au Sri Lanka (RARC),aux Philippines (IPB/UPLB) et en Chine(SCAU). Le CIRAD appor te sonconcours à ce projet en élaborant desprotocoles et en effectuant des analysescomplémentaires sur des échantillons defeuilles. Il est prévu d’étendre cette en-quête à d’autres pays asiatiques.
2. Mise au point d’outils de diagnosticadaptés à l’identification des agentspathogènes foliaires MycosphaerellaPour faciliter les enquêtes sur les diffé-rentes espèces de Mycosphaerella, il estnécessaire d’organiser un cours sur lesméthodes de base servant à identifier lesagents fongiques à partir des caractèresmorphologiques. Il serait particulièrementutile d’avoir un manuel décrivant lessymptômes et les caractères morpholo-giques pour les différentes espèces. Ils’avère également indispensable demettre au point des méthodes molécu-laires permettant de distinguer les es-pèces afin de confirmer ou de diag-nostiquer les infections mixtes ou les in-fections de matériel foliaire âgé. On adéjà mis au point une méthode de cegenre, reposant sur la digestion à l’aided’enzymes de restriction des régions ITSde l’ADNr amplifiées par PCR (Dr Carlier,CIRAD). Cependant, cette méthode né-cessite l’isolement et le clonage deschampignons présents dans les échan-tillons de feuilles. Pour pouvoir analyserde grandes quantités d’échantillons, il serait utile de disposer d’un test rapidepermettant d’identifier les agents patho-gènes localement, sans avoir besoin deles isoler. L’amorce que Johanson etJeger ont définie en 1993 dans les sé-quences ITS pour M. fijiensis et M. musi-
cola n’est pas suffisamment spécifiquepour les distinguer des autres espècesde Mycosphaerella qui peuvent être iso-lées dans les feuilles de bananier (10 es-pèces de Mycosphaerella ou apparte-nant à des genres anamorphesapparentés ont été détectées, Dr Carlier,CIRAD).
3. Réalisation d’une étude sur la biolo-gie fondamentale de M. eumusaeet sur l’épidémiologie de la septoriosedu bananierIl faudrait en savoir davantage sur lecycle biologique de M. eumusae, lesconditions d’infection et de sporulation,l’importance relative des différents typesde spores et la réponse de la septoriosedes bananiers aux différentes conditionsclimatiques.
4. Acquisition d’une connaissance approfondie de la structure des populations de M. fijiensis,M. musicola et M. eumusaeDes études sur la structure des popula-tions de M. fijiensis et M. musicola ontété entreprises à l’échelle continentale,régionale et locale en Afrique, dans la ré-gion Amérique latine/Caraïbes et en Aus-tralie (CRBP, CATIE, CIRAD, CRCTPP,CORPOICA). I l serait nécessaired’étendre ces études, en particulier àl’Asie du Sud et du Sud-Est. En Inde, lesbananiers sont cultivés sous des lati-tudes, altitudes et conditions climatiquestrès diverses et dans le cadre de sys-tèmes de production extrêmement variés.Il est probable qu’une diversité considé-rable s’est constituée avec le temps ausein des populations des différentsagents pathogènes (Dr Selvarajan,NRCB).
L’étude de la structure des populationsdoit inclure à la fois des analyses molé-culaires et des analyses biologiques.Cette structure peut différer entre lesgrandes exploitations commerciales etles petites exploitations en raison de ladifférence de pression de sélection (chi-mique ou génotypique). Il convient departager et de standardiser les proto-coles et les méthodes d’échantillonnageafin d’assurer que les résultats des diffé-rents laboratoires soient comparables.
5. Mise au point de méthodes poursuivre l’évolution des populations depathogènes en réponse à la pression
de sélection des nouveaux génotypesde bananiersIl est indispensable de suivre l’évolutiondes populations pathogènes dans leszones où de nouveaux hybrides résis-tants sont cultivés à grande échelle. Surla base de l’analyse de la structure despopulations et des études épidémiolo-giques, on pourra définir des stratégiespour assurer une gestion durable de larésistance. Ces stratégies feront certai-nement appel à l’amélioration classiqueet à la transformation génétique. Uneétude est en cours au Cameroun (CRBP,CIRAD). Elle doit permettre d’évaluerune méthodologie incluant l’analyse de lastructure des populations et des étudesépidémiologiques. La dimension de laparcelle et la durée de la culture pour-raient être des facteurs clés dans cetteétude. D’après un certain nombre d’ob-servations, on peut penser que les popu-lations de pathogènes évoluent pour sur-monter la résistance à la maladie. Larésistance de Yangambi km5 à M. fijiensisa été apparemment surmontée au Ca-meroun (CRBP) et probablement aussi àCuba (Dr Pérez Vicente, INISAV). La résistance de Paka a été surmontée enPolynésie (Dr Fullerton, HortResearch)et peut-être à Cuba (Dr Pérez Vicente) eten Chine (Dr Jiang, SCAU). Toutefois, il faudrait vérifier l’identité des clones. Il semblerait aussi que la résistance decertains hybrides de la FHIA diminue. On a signalé des niveaux d’infection si-gnificatifs chez FHIA-01 dans les îles duPacifique, en Inde et en Australie (Dr Fullerton, HortResearch ; Dr Selvarajan,NRCB ; Dr Aitken, CRCTPP) et chezFHIA-03 à Cuba (Dr Pérez Vicente, IN-ISAV). Cependant, il se pourrait que ceshybrides exhibent des taches de cerco-sporiose en raison de fortes chargesd’inoculum. Il convient donc d’établir unedistinction entre l’évolution des popula-tions pathogènes et des effets épidémio-logiques particuliers.
6. Identification de nouvelles sourcesde résistance aux affections foliairesdes bananiersLes programmes d’amélioration ne dis-posent actuellement que d’une base gé-nétique très étroite pour développer la ré-sistance (bilan par Dr Tenkouano, IITA). Ilexiste une grande diversité génétique,tant interspécifique qu’intraspécifique,

INFOMUSA — Vol 9, N° 2 PROMUSA IX
chez les bananiers asiatiques. Il faudraitcollecter du matériel nouveau en Asie duSud et du Sud-Est et évaluer sa résis-tance aux différents agents pathogènescausant des taches foliaires. Un travailde collecte important a déjà été effectuéau Viêt-nam et dans d’autres pays. Il fau-drait aussi évaluer les collections exis-tantes. Ces analyses aideront à détermi-ner les résistances aux différents agentspathogènes. Elles devront aussi fournirdes informations sur la réponse des diffé-rentes variétés de bananiers à des infec-tions artificielles ou naturelles de cesagents pathogènes, et inclure des étudesépidémiologiques.
7. Acquisition d’une meilleure compréhension des mécanismes de résistance, et en particulier de résistance partielleOn a identifié plusieurs « mécanismes »de résistance partielle à M. fijiensis etdes évaluations sont en cours pour dé-terminer leur efficacité relative dans lalutte contre la maladie (CRBP, CIRAD).La résistance partielle est probablementplus durable que la résistance totale. Lacombinaison de différents mécanismesde résistance pourrait améliorer leschances de durabilité. Il faudrait analyserla résistance partielle chez les diploïdesafin de pouvoir introgresser la résistancedans le cadre des programmes d’amélio-ration des diploïdes.
Des travaux portant sur la cartographieet l’identification des gènes contrôlant larésistance ont été réal isés (CRBP,CIRAD, CORPOICA). Ils devraient êtrepoursuivis pour permettre la mise aupoint de systèmes de sélection assistéepar des marqueurs et de transformationgénétique. Il faudrait effectuer parallèle-ment des études génétiques et molécu-laires sur la pathogénicité.
8. Mise au point de méthodes de criblage améliorées pour évaluer la résistance du matériel génétiqueaux maladiesIl faudrait publier la procédure du test surdes morceaux de feuilles, y compris lesméthodes d’isolement, culture et produc-tion d’inoculum pour les différents cham-pignons pathogènes (CIRAD), et la diffu-ser aux chercheurs concernés. Unpremier protocole a déjà été diffusé. Uneéchelle standardisée doit être établie pour
enregistrer les niveaux de résistance auxmaladies dans les criblages in vitro. L’utili-sation de toxines de M. fijiensis (ou de ju-glone) pourrait déboucher sur un systèmede criblage utile (Dr Busogoro, Faculté deGembloux), mais il convient de détermi-ner le rôle des toxines et la façon dontelles agissent dans la pathogénicité. Desétudes sont également nécessaires pourétablir si des toxines interviennent dans laseptoriose du bananier.
Autres maladies et actualités
Un nouveau projet est soumisLe CIRAD, en collaboration avec leNRCB, le MARDI et le NERI (Danemark),et avec l’aide de l’INIBAP, a soumis unprojet INCO-DEV à la Communauté eu-ropéenne. Ce projet est axé sur plusieursdes priorités décrites plus haut. Il per-mettra d’entreprendre un important tra-vail commun en Inde et en Malaisie. Ilfaudrait rechercher des fonds supplé-mentaires pour étendre ces études àd’autres pays asiatiques et aborder unplus grand nombre de priorités.
Maladie du sangCette affection létale est causée par unesouche de Ralstonia (Pseudomonas) so-lanacearum qui est étroitement apparen-tée à l’organisme responsable de la ma-ladie de Moko. L’agent pathogène esttransmis par le matériel végétal infecté,et peut-être aussi par des chauve-souriset des insectes qui se nourrissent defleurs. La maladie s’est répandue danstoute l’Indonésie et à Irian Jaya, et l’on arécemment constaté sa présence dansl’ouest de la Papouasie-Nouvelle-Guinée(Dr Williams, QDPI). Il semble que tousles types de bananiers soient sensibles,ce qui aurait des répercussions immé-diates sur la sécurité alimentaire en Pa-pouasie-Nouvelle-Guinée et entraîneraitune perte de diversité génétique.
Moucheture causée par Mycosphaerella spp.Dans la plupart des cas, cette maladieest considérée comme mineure. Cepen-dant, son incidence semble en accroisse-ment chez les hybrides. On soupçonneque plusieurs espèces d’agents patho-gènes en sont responsables (Dr Carlier,CIRAD). Il convient d’identifier ces es-pèces et de déterminer leur répartition. Il
faudrait aussi faire des recherches sur lastructure de leurs populations. Il apparaîtnécessaire d’intégrer la détection decette maladie dans les programmes decriblage. Pour l’instant, on ne disposed’informations que sur les infections na-turelles, et il faudrait donc mettre au pointun système d’inoculation artificielle.
Moucheture noire causée par Phyllosticta/GuignardiaCette affection est importante en Malai-sie et aux Philippines. Son incidencedans les autres pays de la région n’estpas connue.
ChineBeaucoup de maladies sont signalées enChine (Dr Jiang, SCAU). Colletotrichumet Deightoniella peuvent y prendre desdimensions très sévères. La présence dela cercosporiose noire est évidente, maisla cercosporiose jaune est cause de pré-occupations plus immédiates. La septo-riose du bananier n’a encore jamais étésignalée.
Résistance aux fongicidesL’accroissement de la résistance aux fon-gicides devient un sérieux problème dansla plupart des zones de production com-merciale. On signale en particulier unerésistance aux benzimidazoles, triazoleset strobilurines (Dr Guzman, CORBANA ;Dr Abadie, CIRAD, basé au CRBP). Desrecherches sont en cours pour identifierdes solutions de remplacement. En at-tendant, l’apparition de cette résistancecroissante aux fongicides met en relief lanécessité d’une variété de bananier quisoit à la fois résistante aux maladies etcommercialement acceptable.
Symposium sur les cercosporiosesLes participants ont discuté des avan-tages qu’il y aurait à organiser la réunionde PROMUSA juste après le symposiumsur les cercosporioses. Plusieurs ré-unions doivent avoir lieu au Costa Rica età Cuba en 2001. La possibilité de combi-ner certaines réunions est à l’étude et legroupe de travail de PROMUSA sur lescercosporioses décidera ensuite du lieude sa prochaine réunion.
PROMUSALes participants ont estimé que lesavantages apportés par PROMUSA ré-sident principalement dans la possibilitéde participer aux réunions, de pouvoir

X PROMUSA INFOMUSA — Vol 9, N° 2
interagir avec d’autres chercheurs deprofil similaire et d’obtenir des informa-tions récentes au sujet de l’incidencedes différentes maladies et des re-cherches menées sur celles-ci sansavoir à attendre leur publication. Ces as-pects sont particulièrement utiles pourles chercheurs exécutant des re-cherches sur le terrain, qui travaillentsouvent dans l’isolement. La réunion dePROMUSA leur permet de se mettre aucourant des priorités en dehors de leurrégion immédiate. Les échanges d’infor-mations ont été également considéréscomme une fonction très importante dePROMUSA, et les participants ont ap-précié le fait que les groupes puissentdécider de leurs modes de communica-tion et d’interaction.
Les réunions de PROMUSA doiventêtre le moyen d’organiser la recherche etde parvenir à une meilleure efficacité (endéfinissant des priorités). Les partici-pants ont proposé d’inclure les activitéssuivantes dans le programme : • créer et tenir à jour une base de
données sur les organisationssusceptibles d’être des institutionspartenaires pour les projets ;
• faciliter les réunions entre partenairespotentiels (y compris le financement oul’appui pour les visites de planification) ;
• établir des contacts étroits avec lessources de financement, PROMUSAfaisant fonction d’organe decoordination pour définir des prioritéset éviter des duplications ;
• bien que PROMUSA n’ait pas pour rôled’évaluer ou de cautionner lespropositions formulées pour desprojets, il pourrait donner son avis surla pertinence de celles-ci par rapportaux priorités mondiales.
Réunion du groupe detravail de PROMUSAsur la fusariose (GTF)Par t icipants : Afr icano Kangire(KARI/NARO, Ouganda), Altus Viljoen(FABI, Afrique du Sud), Aristoteles Piresde Matos (EMBRAPA/CNPMF, Brésil),Julio Hernandez (ICIA, Canaries), Mauri-cio Rivera (FHIA, Honduras), Mary Wa-bule (KARI, Kenya), Mike Rutherford(CABI, Grande-Bretagne), Randy Ploetz
(University of Florida, USA), Shin ChuanHwang (TBRI, Taiwan), Suzy Bentley(CRCTPP, Australie) et Zaag de Beer(ITSC, Afrique du Sud). Absents (excu-sés) : Ken Pegg (QDPI, Australie), LiewKon Wui (Universiti Sains Malaysia), MikeSmith (QDPI, Australie) et Natalie Moore(QDPI, Australie).Rapporteur : Suzy Bentley
Les discussions ont été articulées autourde quatre thèmes :• aspects fondamentaux pour la gestion
de la maladie et priorités actuelles dela recherche ;
• interactions entre le GTF et les autresgroupes de travail ;
• commentaires sur le fonctionnementde PROMUSA ;
• stratégie pour améliorer lacommunication.
Aspects fondamentaux pour la gestion de la maladie et priorités actuelles de la rechercheLes aspects fondamentaux pour la ges-tion de la fusariose et les priorités ac-tuelles de la recherche ont été définis
Réunion du groupe de travail sur les cercosporioses.

INFOMUSA — Vol 9, N° 2 PROMUSA XI
comme suit lors de la réunion du GTFdes 21 et 22 octobre 1999 à Kuala Lum-pur en Malaisie (voir PROMUSA n° 4, pu-blié dans INFOMUSA vol. 8, n° 2) :• diversité de l’agent pathogène ;• stratégies de gestion de la maladie ;• épidémiologie ;• autres domaines de recherche
prioritaires.Les participants ont fait le point sur lesprogrès accomplis dans chacun de cesdomaines, dont ils ont examiné les diffé-rents éléments afin de déterminer les ac-tions nécessaires et d’actualiser les prio-rités de recherche.
Priorités de rechercheLes participants ont estimé que la pre-mière priorité est la standardisation etl’évaluation du test de criblage des plan-tules pour la résistance à la fusariose. Ilest nécessaire d’élaborer un protocolestandard d’évaluation de la résistance,
de l’évaluer grâce à des essais dans différents laboratoires, puis de le mettreà la disposit ion des programmes d’amélioration.
La mise au point d’un système de dia-gnostic basé sur l’ADN, qui permette dedétecter et d’identifier toutes les races etsouches de Fusarium oxysporum f.sp.cubense (Foc) directement à par t ird’échantillons de plantes et de sol, a étéégalement jugée prioritaire. Il est néces-saire de disposer d’un test rapide et pré-cis pour déceler l’apparition de la mala-die, identifier/certifier le matériel végétalsain et mener des recherches sur l’épidé-miologie et l’écologie de la maladie.
Bases de donnéesIl faudrait créer une base de données re-censant tous les isolats de Foc dispo-nibles dans les principales collections decultures (CABI, Royaume-Uni ; DPI, Aus-tralie ; FABI, Afrique du Sud ; ICIA, îles
Canaries ; KARI, Ouganda ; TBRI, Tai-wan ; UF, États-Unis ; et USM, Malaisie).Il serait également utile d’établir une bi-bliographie contenant tous les docu-ments existants sur la fusariose (pourchacun des principaux domaines de re-cherche susmentionnés) et de la placersur le site Web de l’INIBAP.
Interactions entre le GTF et les autres groupes de travailEn ce qui concerne les interactions avecles autres groupes de travail de PRO-MUSA, les éléments suivants ont étéjugés prioritaires.
Groupe de travail sur l’améliorationgénétique• identification de nouvelles sources de
résistance chez les espèces natives etles hybrides ;
• standardisation et évaluation du test decriblage des plantules ;
Aspect Action recommandée/priorité de recherche1. Diversité de l’agent pathogène1.1 Diversité génétique • Programme de formation à l’analyse des GCV et à la technique DAF pour caractériser les isolats de Foc
• Base de données combinant les informations des principales collections de cultures1.2 Diversité pathogénique • Évaluation et standardisation du test de criblage des plantules1.3 Diagnostics • Mise au point d’un système de diagnostic basé sur l’ADN pour toutes les souches de Foc2. Stratégies de gestion de la maladie2.1 Éducation et sensibilisation • Production d’une fiche d’information générale en plusieurs langues sur la fusariose pour la diffuser largement (en préparation)
• Encouragement de la mise en place de parcelles de démonstration en milieu paysan2.2 Mesures de quarantaine pour prévenir la diffusion • Adaptation de la fiche d’information générale en fonction des conditions et réglementations locales, et la diffuser auprèsde l’agent pathogène du personnel chargé de la quarantaine
• Élaboration de protocoles standard pour la désinfection du matériel, des outils, des pédiluves, etc.2.3 Utilisation de matériel végétal sain • Sensibilisation des producteurs aux avantages des vitroplants2.4 Développement de variétés résistantes • Sélection et évaluation du matériel résistant
• Nécessité d’un test de criblage des plantules pour évaluer leur résistance• Programme d’amélioration des bananiers en Asie
2.5 Adoption de variétés résistantes • Formation à l’utilisation et à l’application des protocoles de l’IMTP III• Encouragement de la mise en place de parcelles de démonstration en milieu paysan
2.6 Lutte chimique/biologique • Utilisation d’agents SAR pour accroître la résistance de l’hôte• Utilisation de F. oxysporum non pathogènes, bactéries, etc.
2.7 Lutte culturale • Amendements du sol• Culture annuelle
3. ÉpidémiologiePlusieurs possibilités de recherche : développement temporel et spatial de la maladie, développement focal de la maladie, survie de l’agent pathogène dans le matériel végétal, le sol et les débris végétaux infectés,études sur les hôtes vicariants4. Autres domaines de recherche4.1 Test de criblage des plantules • Haute priorité !
• Standardisation du protocole de Liew et évaluation dans différents laboratoires de différentes régions dans le monde entier• Validation du test sur la base des données sur les réactions au champ• Création d’un centre de tests pour les isolats exotiques (au CABI)• Mise à disposition du protocole des programmes d’amélioration
4.2 Collecte de matériel génétique indigène et évaluation • Les programmes nationaux devront fournir à l’TC du matériel accompagné de l’accord d’acquisition de ce matériel de sa réaction à Foc
• Vérification de l’identité du matériel génétique4.3 Fausse maladie de Panama • Fausse maladie de Panama aux îles Canaries et en Afrique du Sud
• Flétrissement des Matooke en Ouganda• Production et diffusion d’une fiche technique combinée pour augmenter la sensibilisation à ce problème
Tableau 1. Mesures pour la gestion de la fusariose et priorités de recherche.

XII PROMUSA INFOMUSA — Vol 9, N° 2
• compilation des données existantessur la résistance des bananierssauvages détenus dans les collectionsde matériel génétique ;
• meilleure compréhension desmécanismes de résistance à lafusariose ;
• identification de marqueurs pour larésistance à la fusariose ;
• création d’un programmed’amélioration des bananiers en Asie.
Groupe de travail sur la nématologie• Informer le groupe de travail sur la
nématologie des progrès accomplisdans la mise au point du test decriblage des plantules pour larésistance à la fusariose –combinaison/coordination éventuelleavec la mise au point du test pour larésistance aux nématodes.
Commentaires sur lefonctionnement de PROMUSA• La structure de PROMUSA (relations
entre les groupes de travail, au sein dechacun d’entre eux, et rôle du comitéde pilotage) n’étant pas claire pourtous, il serait utile d’élaborer destermes de référence simples pourpréciser les objectifs des groupes detravail, le rôle du secrétariat et le rôledu facilitateur.
• Il a été suggéré d’établir unmécanisme pour examiner les activitésdes groupes de travail, ainsi qu’uneprocédure pour constituer de nouveauxgroupes de travail (par exemple, sur lavulgarisation).
• Il a aussi été proposé de mettre enplace un système plus rigoureux pourrendre compte des activités desgroupes de travail : par exemple,établissement de rapports semestrielssur la suite donnée auxrecommandations, et la mise en placed’indicateurs permettant le suivi desactions recommandées.
Stratégie pour améliorer la communicationAfin d’améliorer la communication ausein du groupe de travail, les mesuressuivantes ont été recommandées :• utilisation de la liste de messagerie
électronique de PROMUSAFUS ;
• soumission de bilans mensuels de larecherche par les différents membresdu GTF (diffusés par la liste demessagerie électronique dePROMUSAFUS) et compilation de cesbilans sous forme de bulletin annuel(publié sur le site Web de PROMUSA).
Il a été proposé que la prochaine réuniondu groupe de travail sur la fusariose, quiaura lieu en 2002, soit organisée enAfrique du Sud par Altus Viljoen (FABI).
Réunion du groupe detravail de PROMUSAsur la nématologie(GTN)Participants : Guy Blomme (INIBAP, Ou-ganda), Mieke Daneel (ITSC, Afrique duSud), Dirk De Waele (KUL, Belgique),Carine Dochez (IITA, Ouganda), RogerFogain (CRBP, Cameroun), SimonGowen ( University of Reading, Grande-Bretagne), Imelda Kashai ja (KARI/NARO, Ouganda), Thomas Moens(CORBANA, Costa Rica), Tony Pattison(QDPI, Australie), Jean-Louis Sarah(CIRAD, France), K. Soorianathasundaram(TNAU, Inde), Inge Van den Bergh (VASI,Viêt-nam). Excusée : Julie Stanton (QDPI, Australie).Rapporteur : Dirk De Waele
Résultats de rechercheChacun des membres du groupe de tra-vail sur la nématologie a exposé les ré-sultats des activités de son propregroupe de recherche. Les recherchesmenées depuis la précédente réunion auCameroun (1998) peuvent être classéesdans trois grandes catégories.
Biodiversité au sein des communau-tés de nématodesDes études sont en cours sur les diversi-tés interspécifique et intraspécifique deRadopholus similis et Pratylenchus cof-feae (Australie, Costa Rica, Viêt-nam,Cameroun et France). Les enquêtes surle terrain peuvent révéler l’existenced’espèces secondaires, qui constitue-raient une menace potentielle pour laproduction bananière. Par exemple, on atrouvé une nouvelle espèce, R. musicola,en Australie. On a identifié un total de 33
espèces chez 18 genres au Cameroun.En revanche, il semble que R. similis soitabsent des plantations bananières duViêt-nam. Le CIRAD a montré, à l’aidede différentes techniques moléculaires,qu’on peut répartir les populations de R.similis dans deux groupes génomiques :l’un couvrant le Cameroun, le Costa Ricaet l’Australie, l’autre s’étendant depuis laCôte d’Ivoire jusqu’en Ouganda.
Impact des nématodes sur la produc-tion bananièreDes études portant sur les corrélationsentre les dégâts des nématodes sur lesracines et les pertes de production sonten cours au Costa Rica, au Viêt-nam etau Cameroun. Des préoccupations ontété exprimées au sujet des résultatsd’une enquête récente, qui indique que laplupart des producteurs sud-africains quicultivent la banane pour leur subsistancese servent de la même variété. En outre,la production au Mozambique, qui repré-sente une source importante de matérielpour les exploitants sud-afr icains,connaît actuellement un déclin.
Criblages, mécanismes de résistance,transformationDes criblages sont effectués actuellementdans plusieurs pays afin de déterminer larésistance des variétés de bananiers. AuViêt-nam, les variétés récemment collec-tées sont soumises à des essais. En Ou-ganda, on a mis au point une nouvelletechnique de criblage reposant sur l’ino-culation d’une seule racine. Des criblagesin vitro sont en cours à la KUL. En Inde etau Cameroun, on étudie les composésphénoliques et l’histomorphologie des ra-cines afin de déterminer les mécanismesde résistance aux nématodes, en particu-lier chez Yangambi km5. D’autre part, laKUL étudie l’utilisation de plantes trans-géniques modèles, comme Arabidopsis,pour développer la résistance aux néma-todes chez les bananiers.
Priorités de rechercheLes participants ont identifié plusieursnouvelles priorités de recherche. Lespriorités précédemment établies ont étérestructurées autour de trois axes princi-paux : • communautés de nématodes, y
compris leur biodiversité ;

INFOMUSA — Vol 9, N° 2 PROMUSA XIII
• dégâts des nématodes et pertes deproduction potentielles ;
• criblage pour la résistance : méthodes,sources, mécanismes.
Liaison avec le groupede travail sur l’améliorationgénétiqueLe groupe de travail sur la nématologie aétabli le tableau 2 suivant des besoinsprésumés des sélectionneurs. Dans les
trois premières colonnes figurent les différentes catégories d’activités. La dernière colonne indique l’« offre » decontribution des nématologistes.
Rôle du groupe de travail sur la nématologie et de PROMUSADes discussions ont eu lieu sur le fonc-tionnement du GTN et de PROMUSAdans son ensemble. Bien que les
membres du GTN aient été très actifs, iln’y a guère eu de collaboration au seindu groupe de travail, ni entre celui-ci etles sélectionneurs. Il est clair qu’il fau-drait améliorer la communication. Lesparticipants se sont efforcés de trouverdes solutions à ces problèmes et de défi-nir les priorités actuelles de la rechercheet les besoins pour l’année à venir.
Le rôle du facilitateur a fait l’objet d’un débat. Dirk De Waele a été choisicomme nouveau titulaire de cette fonc-
Membres du groupe de travail sur la nématologie.
Lutte intégrée Méthodes classiques Méthodes moléculaires Connaissances actuellesDe quelle contribution du GTN les sélectionneurs ont-ils besoin ?➣ Statut de l’hôte :•résistance +++ +++ +•tolérance +++ -➣ Sources de résistance (utiles pour l’amélioration) +++ +➣ Mécanismes de résistance + (+) +++ +➣ Héritabilité à base génétique ++ ++ -➣ Marqueurs (moléculaires) +++ + -➣ Gènes (protéines) (+) +++ -Outils➣ Criblages in vitro +++ +++ + ++➣ Criblages en serre +++ +++ + ++➣ Criblages en champ +++ +++ + ++➣ Criblages hautement sensibles +++ -
Tableau 2. Liaisons avec le groupe de travail sur l’amélioration génétique

XIV PROMUSA INFOMUSA — Vol 9, N° 2
tion. Les participants ont estimé que le fa-cilitateur ne peut, à lui seul, faire fonction-ner le groupe de travail et que tous lesmembres doivent participer activement.Ils ont aussi souligné que le facilitateur nedoit pas constituer un « goulot d’étrangle-ment » pour la communication externe(avec le secrétariat/comité de pilotage ,ou avec les autres groupes de travail).
En ce qui concerne le rôle de PRO-MUSA, les participants sont parvenusaux conclusions suivantes : • PROMUSA doit faciliter la
communication (entre les chercheursqui veulent communiquer) de façon àaméliorer la collaboration et leséchanges d’informations ;
• PROMUSA doit faciliter l’accès desgroupes de travail à desfinancements (sans toutefois fournirlui-même de fonds) et améliorer lacommunication pour le montage desprojets.
Afin de contribuer à PROMUSA, le GTNdevra : • renforcer la communication en mettant
à jour le fichier d’adresses, enpartageant des informations surInternet (résumés de publications,méthodologies, description desrecherches en cours), en organisantdes échanges d’étudiants de cyclesupérieur et d’associés de recherche,en échangeant du « matériel », etc. ;
• recenser les connaissances actuelleset établir trois bases de données sur :les communautés de nématodes, leurbiodiversité (J.-L. Sarah), les dégâtsdes nématodes et les pertes deproduction potentielles (R. Fogain), lessources et les mécanismes derésistance (D. De Waele) ;
• participer à la phase III de l’IMTP enfournissant une liste de 10 génotypesà tester dans les différents sites (R. Swennen et tous les membres), enenvoyant des informations auxchercheurs figurant dans le fichierd’adresses (de façon à permettre àtout chercheur intéressé de participer)et en faisant des essais sur le terrain(15 répétitions, avec des plants enprovenance de champs naturellementinfestés et de champs traités avec desnématicides) ;
• organiser une réunion à la suite ducongrès de nématologie de la NSSA
(mai 2001, Afrique du Sud) – « Ateliersur l’amélioration génétique desbananiers pour la gestion desnématodes » (Workshop on geneticimprovement of Musa for nematodemanagement) (2-3 jours).
Réunion du groupe detravail de PROMUSAsur la virologiePar t icipants : Mar ie Line Caruana(CIRAD, France), James Dale (QUT, Aus-tralie), Martine Delanoy (FUSAGx, Bel-gique), Glyn Harper (JIC, UK), BertrandHelliot (FUSAGx, Belgique), John Hu(Université de Hawaii, USA), Jackie Hu-ghes (IITA, Nigeria), Roger Hull (JIC,UK), Lawrence Kenyon (NRI, UK), Phil-lipe Lepoivre (FUSAGx, Belgique), BenLockhart (Université du Minnesota, St.Paul, USA), Gerhard Pietersen (ARC-PPRI, Afrique du Sud), Helena Reichel(CORPOICA, Colombie), H.J. Su (Uni-versité nationale de Taïwan), John Tho-mas (DPI, Australie).Rapporteur : Jackie Hughes
Le BSV et les autres (potex)virus dans les transferts et échanges de matérielgénétiqueDes discussions ont eu lieu sur le fonc-tionnement de PROMUSA et du groupede travail, et sur les fonctions du facilita-teur. Un nouveau facilitateur a été dési-gné, Jackie Hughes, et les participantsont exprimé leur appréciation pour sacontribution à Roger Hull, qui assumaitprécédemment cette charge. Lesgroupes de recherche représentés ausein du groupe de travail ont fait uncompte-rendu des activités menées pourchacun des virus. Un tableau récapitulatifa été dressé sur les diagnostics actuelle-ment utilisés par les centres d’indexationde l’INIBAP, ainsi que ceux dont dispo-sent les chercheurs et les entreprisescommerciales.
Virus du bunchy top du bananier(BBTV)Il a été reconnu que la variabilité de laséquence du BBTV, particulièrement en
Asie, est plus importante qu’on ne l’avaitpensé jusqu’alors. D’après l’expressiondes symptômes, il existe différents va-riants biologiques. Des indicateurs de ré-sistance ont été identifiés chez desplants matures. On a aussi créé un cer-tain nombre de plants transgéniques. Aumoins une construction se montre résis-tante au virus, mais non immune. Laconnaissance de la biologie moléculairedu BBTV progresse et l’on a établi unesérie de séquences d’ADN microsatel-lites associées aux isolats asiatiques.
Virus de la mosaïque des bractées dubananier (BBrMV), genre PotyvirusLa Queensland University of Technology(QUT) a signalé que des variétés Caven-dish et Bluggoe génétiquement modifiéesavec des constructions de la capside pro-téique du BBrMV sont maintenant dispo-nibles pour des essais. Des expérimenta-tions en champ doivent avoir lieu auxPhilippines. Le séquençage des gènes dedifférents échantillons du virus (dont CPet Nib) a révélé que la variation génétiquen’est pas supérieure à 5 % au niveau desnucléotides de CP et Nib. On a créé uneconstruct ion ayant une var iat ion « moyenne ». Outre les sites où sa pré-sence est connue aux Philippines, en Indeet au Sri Lanka, le virus a été détecté unefois dans les Samoa occidentales et, ende rares occasions, au Viêt-nam. Toute-fois, ces isolats n’engendraient pas desymptômes typiques.
Le Queensland Department of PrimaryIndustry (QDPI) a identifié un isolat duBBrMV présentant plus de 10 % de va-riation génétique, ce qui a des implica-tions significatives pour le développe-ment de matériel transgénique. Un jeucomplet de diagnostics sérologiques etbasés sur la PCR est disponible.
Virus de la mosaïque du concombre(CMV)Bien que largement répandu, le CMVn’engendre généralement pas de mala-die sérieuse. Cependant, avec l’augmen-tation du nombre des vitroplants dans laproduction bananière à Taiwan et enChine, ce virus commence à devenirplus problématique. Des infectionsmixtes du CMV et du virus de la mo-saïque légère du bananier peuvent aussiproduire des symptômes plus sévères.Les souches du CMV infectant les bana-

INFOMUSA — Vol 9, N° 2 PROMUSA XV
niers ont été caractérisées à Hawaï, àTaiwan et en Chine. On est en voie decréer des plants transgéniques dotés derésistance à ce virus.
Virus de la mosaïque légère du bananier (BanMMV)La souche virale filamenteuse isoléechez le cv. « Ducasse » Pisang Awak(ABB) a été entièrement séquencée. Ilsemble qu’il s’agisse d’un nouveau typede virus, se situant entre les potex, fovéaet alexivirus.
Le virus filamenteux identifié en coin-fection avec le BanMMV est sérologique-ment apparenté à la particule filamen-teuse qui infecte une grande diversité debananiers, notamment les bananiers àcuire et 11 % de la collection du CIRADen Guadeloupe.
On utilise des amorces définies à partird’une zone conservée de CP du virusDucasse, ainsi que des poly-A, afind’étudier la diversité du virus au sein decette collection et dans une partie du ma-tériel génétique de l’INIBAP. Étant donnéqu’il n’y a pas de symptômes, ou seule-ment de légers symptômes associés à cevirus en infection simple, on ne disposed’aucune information véritable sur sonimpact et son épidémiologie.
Un projet conjoint FUSAGx/CIRAD/-CORPOICA-Université du Costa Rica/-
INCO a été soumis en septembre dans lecadre de PROMUSA en vue d’évaluer lesrisques associés à la diffusion de cevirus, seul ou en coinfection avec le BSVou le CMV. Un nouvel étudiant en PhDtravaille actuellement à Gembloux sur lacaractérisation de la souche colom-bienne et la mise au point d’un kit de dia-gnostic fiable.
Virus de la mosaïque en tirets du bananier (BSV)Les par t icipants ont examiné lespreuves montrant clairement que l’infec-tion par le BSV provient de séquencesvirales qui s’intègrent dans le génomede bananier. La culture de tissus est unfacteur qui déclenche l’expression épi-somale des séquences intégrées duBSV. Des séquences « activables » (ca-pables d’expression épisomale) aussibien que des séquences « non acti-vables » (incapables d’expression épi-somale) s’intègrent dans le génome debananier.
Un intégrant activable du BSV (BSV-OL) est associé au génome B de bana-nier, mais pas au génome A. Trois autresespèces épisomales du BSV (BSV-GF,BSV-IM et BSV-MYS) sont intégréesdans le génome de bananier. De mêmeque la BSV-OL, elles semblent associéesau génome B. D’après les premières ob-
servations, il semble qu’il existe une séried’autres intégrants du BSV chez les ba-naniers. Cependant, ils n’ont pas encoreété caractérisés.
L’intégrant à expression épisomaleBSV-OL et d’autres intégrants virauxpouvant avoir ce type d’expression (BSV-GF, BSV-IM) sont largement présentschez les bananiers plantain (AAB) et lesbananiers (AAA) d’Amérique centrale etd’Amérique du Sud. Les discussions ontabouti à la conclusion que ces espècesdu BSV ne sont pas introduites dans denouvelles zones chez les hybrides tétra-ploïdes AAAB produits par les différentsprogrammes d’amélioration. L’infectiondes bananiers dessert AAA par le BSV-OL et le BSV-GF au Costa Rica, enÉquateur et au Venezuela résulte d’uneinfection latente préexistante ou de latransmission du virus à partir de bana-niers plantain.Les recommandations suivantes ont été
formulées :• il est nécessaire de procéder
systématiquement à l’indexation duBSV dans le cadre de la productioncommerciale de vitroplants debananiers AAA ;
• des recherches doivent êtreimmédiatement entreprises sur le rôlede la cochenille farineuse dans latransmission du BSV en champ ;
Virus Diagnostic(s) ObservationsBBTV ELISA Les antisérums commerciaux détectent les souches connues
PCR Sensible, mais spécificité liée à la soucheBBrMV Miniprép + ISEM Antisérums polyclonaux et antisérums recombinants disponibles *
ELISAStandardisation avec miniprép + ISEM
BanMMV Miniprép + ISEM Antisérum polyclonal disponible *Méthode standardisée
AbaMV Miniprép + ISEM Antisérum polyclonal disponible *Méthode standardisée
CMV ELISA Antisérum commercial disponible, détectant les deux sérotypesMéthode standardisée – même antisérum
BSV (épisomal) Miniprép + ISEM Antisérum poly-polyclonal disponible* détectant le virus épisomal. Possibilité de problèmes en cas de variation des souches. Possibilité de problèmes avec les antisérums et les amorces en cas de variation.
IC-PCRMéthode standardisée miniprép + ISEM
BSV (intégrant activable) PCR Spécificité liée à la souche. Nécessite des informations sur la séquence de la souche.Autres virus Miniprép + EM Détecte la présence des virus en forme de bâtonnets et, en combinaison avec des antisérums0*,
détecte des virus précédemment non reconnus. Plus de difficultés avec les virus isométriques de faible concentration.
* Non disponible pour la détection commerciale.0 Quelques anticorps disponibles pour le BDBV.
Diagnostics actuellement disponibles pour les virus des bananiers

XVI PROMUSA INFOMUSA — Vol 9, N° 2
• l’expression épisomale des intégrantsdu BSV dans le génome B de bananiersemble nécessiter la présence dugénome A. Ce modèle est corroborépar des données sur des phénomènessimilaires chez le tabac et le pétunia.La PCR avec immunocapture (IC-PCR) est dix fois plus sensible quel’immunoélectroscopie (ISEM) pourdétecter le BSV. Il convient depoursuivre les recherches pourapporter une réponse aux questionsqui se posent au sujet de la fiabilité decette technique.
Autres virusUne brève discussion a eu l ieu surd’autres virus en forme de bâtonnets ren-contrés chez le matériel génétique de ba-naniers. On a trouvé en particulier desparticules semblables à des potyvirus auSri Lanka. On a aussi trouvé des parti-cules virales isométriques chez Musaspp. au Nigeria. Ce virus, dénommé pro-visoirement virus du dépérissement dubananier (BDBV, genre ? Népovirus), en-gendre de sévères symptômes de dépé-rissement et semble se diffuser lente-ment dans le champ. La production dediagnostics (polyclonal, monoclonal,amorces) est en cours.
Fonctionnement de PROMUSALa dernière partie des débats a porté surle fonctionnement de PROMUSA. Les
participants ont constaté que PROMUSAa facilité la communication au sein dugroupe, mais qu’il faudrait renforcer lesliaisons entre les différents groupes detravail. Ils ont proposé de constituer un « groupe des facilitateurs » pour aider àdiffuser l’information, les résultats de re-cherche et les besoins aux membres desgroupes appropriés. Ils ont aussi de-
mandé qu’on crée un site Internet àaccès sélectif, sur lequel les membrespourraient placer des informations(comme des données incomplètes surdes séquences) à l’intention des seulsgroupes de travail. Enfin, il conviendraitd’inviter des virologistes de pays commel’Inde et la Chine à participer au groupede travail.
Participants du groupe de travail sur la virologie.
Priorités de la recherche en virologie en relation avec l’amélioration génétique
• détection fiable du BSV grâce à l’utilisation rationnelle de diagnosticsappropriés, à l’aide d’une meilleure compréhension de la diversité du BSV etpar la mise en lumière de la signification des génomes A et B.
• production d’une brochure PROMUSA sur les procédures actuelles dediagnostic des virus.
• meilleure compréhension de l’hétérogénéité du génome B.• meilleure compréhension de la contribution du génome A à l’activation des
intégrants du virus chez les lignées avancées.• mécanismes de mise en silence des intégrants du BSV dans le génome.• recherche sur la diversité géographique du BSV au regard des transferts de
matériel génétique, afin d’améliorer l’information épidémiologique etl’évaluation des risques.
• mise au point de méthodes de criblage pour la résistance.• Approvisionnement en diagnostics.• Recherche sur la possibilité de produire des plantules indemnes de virus, et en
particulier du BanMMV et du BSV.





























