Embed Size (px)
Citation preview

Lactate Dehydrogenase (LDH) Gene Duplication During Chordate Evolution: The cDNA Sequence of the LDH of the Tunicate StyeZa plicata
David W. Stock, *icl Joseph M. Quattro, *2 Gregory S. Whitt,t_ and Dennis A. Powers* *Department of Biological Sciences, Hopkins Marine Station, Stanford University; and TDepartment of Ecology, Ethology and Evolution, University of Illinois at Urbana-Champaign
L-Lactate dehydrogenase (L-LDH, E.C. 1.1.1.27) is encoded by two or three loci in all vertebrates examined, with the exception of lampreys, which have a single LDH locus. Biochemical characterizations of LDH proteins have suggested that a gene duplication early in vertebrate evolution gave rise to Ldh-A and Ldh-B and that an additional locus, Ldh-C arose in a number of lineages more recently. Although some phylogenetic studies of LDH protein sequences have supported this pattern of gene duplication, others have contradicted it. In particular, a number of studies have suggested that Ldh-C represents the earliest divergence among vertebrate LDHs and that it may have diverged from the other loci well before the origin of vertebrates. Such hypotheses make explicit statements about the relationship of vertebrate and invertebrate LDHs, but to date, no closely related invertebrate LDH sequences have been available for comparison. We have attempted to provide further data on the timing of gene duplications leading to multiple vertebrate LDHs by determining the cDNA sequence of the LDH of the tunicate Styelu plicatu. Phylogenetic analyses of this and other LDH sequences provide strong support for the duplications giving rise to multiple vertebrate LDHs having occurred after vertebrates diverged from tunicates. The timing of these LDH duplications is consistent with data from a number of other gene families suggesting widespread gene duplication near the origin of vertebrates. With respect to the relationships among vertebrate LDHs, our data are not consistent with previous claims that Ldh-C represented the earliest divergence. However, the precise relationships among some of the main lineages of vertebrate LDHs were not resolved in our analyses.
Introduction
Comparisons of nuclear genome sizes and numbers of loci encoding isozymes and other proteins in a variety of chordates led Ohno (1970) to propose that episodes of large-scale gene duplication had occurred during the evolution of the group. The more recent discovery that many genes involved in the control of development are members of multigene families with more representa- tives in vertebrates than in invertebrates has led to in- creased interest in documenting the patterns of gene du- plication in chordate evolution (Holland 1992; Holland et al. 1994). The sizes of some of these families, coupled with the difficulty in determining gene number in a large sample of taxa, suggest the value of continued exami- nation of smaller, more extensively studied gene fami- lies in detecting episodes of gene duplication in chordate evolution. One such gene family encodes L-lactate de- hydrogenase (L-LDH, E.C. 1.1.1.27), which catalyzes the interconversion of pyruvate (the end product of gly- colysis) and L-lactate with the concomitant interconver- sion of NADH and NAD+ (Markert 1984). Extensive surveys of LDH locus number, expression pattern, ki- netic properties, and immunochemical similarities in representatives of the major taxa of jawed vertebrates (Marker& Shaklee, and Whitt 1975; Fisher et al 1980; Whitt 1984) have suggested that they all possess or- thologs of Ldh-A and Ldh-B. LDH-A is expressed pre-
’ Present address: Department of Anthropology, The Pennsylvania State University.
* Present address: Department of Biology, Program in Marine Sci- ence, University of South Carolina.
Key words: tunicate, molecular phylogeny, gene duplication.
Address for correspondence and reprints: David W. Stock, De- partment of Anthropology, The Pennsylvania State University, Uni- versity Park, Pennsylvania 16802. E-mail: [email protected].
Mol. Biol. Evol. 14(12): 1273-1284. 1997 0 1997 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
dominantly in white skeletal muscle and is kinetically suited to the reduction of pyruvate to lactate under con- ditions of low oxygen availability, while LDH-B is pre- dominantly expressed in more aerobic tissues and is tai- lored to the oxidation of lactate to pyruvate (Markert 1984). An additional locus encoding an isozyme with more variable kinetic properties (LDH-C) is expressed in the sperm of mammals and columbid birds and in a variety of tissues or the liver or eye of ray-finned fishes (Markert, Shaklee, and Whitt 1975). Jawless fishes pos- sess either a single locus (lampreys) or, in the case of hagfishes, two loci with tissue expression patterns sim- ilar to those of jawed vertebrate Ldh-A and Ldh-B (Mar- kert, Shaklee, and Whitt 1975). Markert, Shaklee, and Whitt (1975) proposed that an initial gene duplication gave rise to Ldh-A and Ldh-B early in vertebrate evo- lution and that Ldh-C is the result of a more recent du- plication.
Subsequent phylogenetic analyses of vertebrate LDH sequence data have supported some aspects of this “traditional” scenario for LDH evolution and chal- lenged others (Li et al. 1983; Hendriks et al. 1988; Crawford, Constantino, and Powers 1989; Stock and Whitt 1992; Quattro, Woods, and Powers 1993; Tsuji et al. 1994; Quattro et al. 1995). Some points that remain controversial or poorly resolved are the phylogenetic po- sition of the LDH-Cs, the relationship of cyclostome LDHs to gnathostome LDHs, and the monophyly of gnathostome LDH-As. The former two issues involve the earliest duplication events leading to the multigene family in vertebrates. Such duplications would be fur- ther elucidated by an examination of the relationship of invertebrate and protochordate LDHs to those of verte- brates. To date, the only complete LDH sequence avail- able from an invertebrate animal is that of the nematode Caenorhabditis elegans (Tsoi and Li 1994), a represen-
1273
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1274 Stock et al.
tative of a group that is quite distant from vertebrates. Among protochordates, tunicates and amphioxus have been reported to possess single LDH loci (Fisher et al. 1980; Baldwin, Mortimer, and Patak 1988). Immuno- chemical analyses and comparisons of amino acid com- position were interpreted as evidence that tunicate LDH was more closely related to the LDH-C of a teleost fish than to either LDH-A or LDH-B (Baldwin, Mortimer, and Patak 1988), while Fisher et al. (1980) suggested that the single-locus condition of amphioxus and tuni- cates predated the divergence of these groups from ver- tebrates. We have attempted to provide additional reso- lution of the early duplication history of chordate LDHs by cDNA sequence determination and phylogenetic analysis of an LDH locus from the tunicate Sty& pli- cata.
Materials and Methods RNA Extraction
Adult specimens of the tunicate Styelu plicata were obtained from Gulf Specimen Company (Panacea, Fla.). A section of the body wall was cut from a single spec- imen, and the inner and outer surfaces were removed with a scalpel. The remaining portion was ground to a fine powder in liquid N2 using a mortar and pestle. Total RNA was extracted from 2 g of powdered tissue by homogenization in the presence of sodium tri-isopro- pylnaphthalene sulfonate followed by phenol/chloro- form extraction, ethanol precipitation, and precipitation from sodium acetate, as previously described (Stock et al. 1991).
Reverse Transcription
Total RNA was reverse transcribed with MMLV re- verse transcriptase and either random hexamers or a dT adaptor primer (5’-GATGCTGCAGAGCTCAAGC(T)is- 3’) as described by Stock and Powers (1995).
RT-PCR Amplification of Internal Fragment of Styela Ldh
A number of degenerate PCR primers (designed from an alignment of bacterial and vertebrate LDH pro- tein sequences-Stock and Whitt 1992; Stock and Pow- ers 1995) were tested for their ability to amplify a prod- uct of the expected size from Styela first-strand cDNA. Of these degenerate primers, only Ldhll3FhF (5’- GCGAATTCGTNGGNATGGCNTGYGCNRT-3’, where the underlined sequences indicate restriction sites, and ambiguous bases are indicated by the appropriate IUPAC codes) and LdhB736FhR (S’-GCCGCTGCA- GCCCANBWNGTRTANCCYTT-3’) amplified a frag- ment of Styela L+dh. The sequence obtained from this fragment was used to design gene-specific primers for additional RT-PCR (S’-GCCGGAGCTCAAGGT- GAAGTAATGGACT-3 ‘, used in combination with primer LdhB736FhR) and RACE amplifications (see be- low). Conditions for RT-PCR were as described by Stock and Powers (1995). Weak bands were reamplified in a similar PCR reaction using as a template either a dilution of a band cut from a low-melting-point agarose gel (NuSieve-FMC) or the DNA adhering to a syringe
needle stabbed into a band of the expected size (band stab technique of Bjourson and Cooper 1992).
RACE Amplification of the 3’ End of Styela Ldh
The 3’ end of the Styelu LDH cDNA was amplified by 3’ RACE, essentially as described by Frohman (1990). A dT-adaptor reverse transcription was used as the tem- plate for an initial PCR with an adaptor primer (5’-GA- TGCTGCAGAGCTCAAGCTT-3’) and the gene-specific primer 5’-GCCGGAGCTCGRTGGMCCWGAAGGY- TGGGA-3’. PCR conditions were as described by Stock and Powers (1995). The initial PCR was then used as a template for a second PCR with the adaptor primer 5’- GATGCTGCAGAGCTCAAG-3’ and the gene-specific primer 5’-GCCGGAGCTCACAYAARCARGTYRTY- GATG-3’.
RACE Amplification of the 5’ End of Styela Ldh
5’ RACE amplification followed Frohman (1990). RNA was reverse transcribed with the degenerate LDH primer 5’-GCCGCTGCAGTCNACNACYTGYTTRTG- 3’. The first-strand cDNA was tailed with dATP using terminal deoxynucleotidyl transferase. PCR amplifica- tion of the dA-tailed template was with the gene-specific primer 5’-GCCGCTGCAGTGGCCGATACGGAG- TAATCT-3’, the dT adaptor primer, and one of the adap- tor primers used above. The PCR reaction was subjected to electrophoresis on an agarose gel, and the band stab technique (Bjourson and Cooper 1992) was used to reampufy a faint band of the expected size with the nest- ed LDH primer 5’-GCCGCTGCAGCCRTGYTGNA- RRTCCAT-3’ and the same adaptor primer as in the first reaction.
Cloning
PCR products were cloned into Ml3 mp18 and mp 19 or pBluescript KS+ and KS - (Stratagene) using standard procedures (Sambrook, Fritsch, and Maniatis 1989). PCR products were either digested with restric- tion enzymes (recognizing sites at the 5’ ends of the primers or in the region between the primers), made blunt-ended with the Klenow fragment of DNA poly- merase I, or cloned directly using the t-vector technique (Marchuk et al. 1991).
Sequencing
Single-stranded DNA for sequencing was either ex- tracted from Ml 3 phage particles or obtained from pBluescript-containing cultures infected with VCSM13 helper phage (Stratagene) according to Sambrook, Fritsch, and Maniatis (1989). DNA sequencing was by dideoxy chain termination using a modified T7 DNA polymerase (Sequenase, version 2.0, United States Bio- chemicals). In order to minimize errors associated with the sequencing of cloned PCR products, each nucleotide was determined from at least six clones, with at least three sequences representing each strand.
Sequence Assembly and Alignment
The cDNA sequence of the LDH of Styelu plicata was assembled from the consensus of overlapping clones using the SEQMAN contig assembly program in
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

Tunicate LDH 1275
the DNASTAR package (DNASTAR, Inc.). The se- quence described in this paper has been deposited in GenBank under accession number AF023168 (fig. 1). Additional lactate dehydrogenase sequences were ob- tained from the GenBank (release 92) or PIR database. D-LDHs (E.C. 1.1.1.28) were not included in the anal- yses, as they represent an, at best, distantly related pro- tein family or families (Kochhar et al. 1992). The L-
LDH sequences used and their accession numbers (GenBank, unless otherwise noted) are: lamprey (Pet- romyzon marinus) LDH (M74064); dogfish shark (Squa- lus acanthias) LDH-A (U38893); killifish (Fundulus heteroclitus) LDH-A (I-43525), LDH-B (M33969), and LDH-C (L07336); frog (Xenopus Zaevis) LDH-A (UO7179), LDH-B (UO7176), and LDH-C (U07175); duck LDH-B (503869); chicken LDH-A (X53828) and LDH-B (PIR accession A00346); cattle LDH-A (D90143); pig LDH-A (U07178) and LDH-B (UO7180); rabbit LDH-A (M22585); human LDH-A (X02152), LDH-B (YOO711), and LDH-C (502938); mouse LDH- A (YOO309), LDH-B (X51905), and LDH-C (X04752); rat LDH-A (X01964) and LDH-C (U07177); fox (Vul- pes vulpes) LDH-C (U19868); nematode (C. elegans) LDH (U15420); barley LDH-A (M55685) and LDH-B (M55684); rice LDH (D13817); maize LDH (211754); the LDHs of the apicomplexan protists ToxopZusma gon- dii (U35 118) and Plasmodium falciparum (M93720); and the L-LDHs of the eubacteria Streptococcus mutuns (M72545), S. thermophilus (D 13405), Luctococcus lac- tis (M88490), Luctobacillus casei (M76708), Luctoba- cillus sake (U26688), Lactobacillus plantarum (D90340), Pediococcus acidilactici (X70927), Bacillus stearothermophilus (M19396), B. caldolyticus (M19394), B. caldotenax (M19395), B. psychrosacchar- olyticus (LDH-A-X55 118 and LDH-B-X55 119), B. subtilis (PIR accession A25805), B. megaterium (M22305), Mycoplasma hyopneumoniae (X67286), M. genitalium (U39733), Thermus aquaticus (D00585), T. caldophilus (X045 19), Deinococcus radiodurans (D63899), Thermotoga maritima (X74302), BiJidobac- terium longum (M33585), and Synechocystis sp. (D64003).
Phylogenetic Analyses
Positions l-44 in the alignment of LDH sequences (fig. 2) were omitted from all analyses because of the difficulty in obtaining a reliable alignment of this region. Phylogenetic analyses were conducted using both neigh- bor-joining and parsimony methods. Amino acids were used as characters in both types of analysis because of the large amount of sequence divergence among some of the taxa and the fact that we were interested in early branching events in LDH evolution (Kumar, Tamura, and Nei 1993). Neighbor-joining trees were constructed from pairwise amino acid distances (gamma corrected with a = 2.0) using the MEGA package of programs (Kumar, Tamura, and Nei 1993). Parsimony analyses were conducted with PAUP, version 3.0s (Swofford 1991), and in most analyses, amino acids were treated as unordered characters with all possible transformations weighted equally. Preliminary analyses with amino acid replacements weighted according to the minimum num- ber of nonsynonymous nucleotide substitutions required (PROTPARS weighting-Felsenstein 1993) gave simi- lar results but were not pursued extensively because of the computational time required. Parsimony analyses with equal weighting were performed with 100 random addition sequences of taxa and TBR branch swapping (Swofford 1991), while those with PROTPARS weight- ing used 20 random addition sequences. The CON- STRAINTS feature of PAUP was used to determine the lengths of the shortest trees matching a number of al- ternative phylogenetic hypotheses, In the case of con- strained analyses using PROTPARS weighting, the PROTPARS program of the PHYLIP package (Felsen- stein 1993) was used to test whether the length differ- ences between constrained trees and the best uncon- strained tree were significant according to the method of Templeton (1983). Bootstrapping was used to assess the degree of support for particular nodes. Neighbor- joining analyses were conducted with 500-1,000 boot- strap replications, and bootstrap parsimony analyses were conducted with 10 replications of a random addi- tion sequence for each of 100 bootstrap replications.
Nucleotide sequences were translated into protein sequences according to the authors’ feature specifica- tions. An alignment of amino acid sequences was con- structed using CLUSTAL W (Thompson, Higgins, and Gibson 1994) and the default settings. Length variability among chordate LDH sequences in this alignment was limited to the amino terminal arm (positions l-43 in fig. 2, as defined by Li et al. 1983) and the extreme carboxy terminus. No attempt was made to improve the align- ment of the former region because of extensive sequence variability, but the alignment of the later region was ad- justed manually. Invertebrate and plant LDH sequences require the addition of only a few additional gaps to the alignment of chordate LDHs, but the alignment of bac- terial and protist sequences appeared less reliable. Rath- er than attempting additional alignment methods or pa- rameters, we conducted phylogenetic analyses with and without the latter two groups of LDHs.
Results Features of Styelu Ldh
The 1,168-nt cDNA sequence of S. plicatu LDH is shown in figure 1. Three of 10 clones of the 3’ end had an additional 10 nucleotides before the poly-A tail (not shown). No consensus polyadenylation signal (Sheets, Ogg, and Wickens 1990) was found in either class of clone, suggesting that the reverse transcription primer may have bound upstream of the true poly-A tail. Of the sites at which nucleotide differences from the se- quence shown in figure 1 were found in individual clones of PCR products, only seven exhibited the same difference in more than one clone. Two of these differ- ences occurred in the 3’ untranslated region (positions 116 1 and 1164), three were silent (positions 923, 97 1, and 1037), and only two encoded different amino acids (N and P at nucleotide positions 454 and 583, respec- tively). These latter two differences were only found in
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1276 Stock et al.
GATTGGAGTATTTGGTTGTATTAAGAATTTTGATAATTAAACACAA 45 TCATGGCTGATCTGAAGGGATTCGATATAATGTCCGAGTTATTCA 90
MADLKGFDIMSELF 14 CGGAAGTCTGTCCAGACATTCCAAAATCTGGACCATCCAAAGTCA 135 TEVCPDIPKSGPSKV 29 CTGTAGTTGGAGTTGGGATGGTGGGGATGGCGTGTGGGATGAGTG 180 TVVGVGMVGMACGMS 44 TGGTGTTGAAGGGACTATGTACAGATCTTGTACTAGTCGATGTTG 225 VVLKGLCTDLVLVDV 59 TTCAGGACAAACTTCAAGGTGAAGTAATGGACTTGCAACATGGTA 270 VQDKLQGEVMDLQHG 74 GTTTGTTTCTAGAGAATATTCTATGGAGACAAAGATTACT 315 SLFLENIKVYGDKDY 89 CCGTATCGGCCAACAGTCGGATTGTCATAGTAACTGCTGGTGCTC 360 SVSANSRIVIVTAGA 104 GTCAACZUXCTGGGGAATCTAGATTGAGTTCAACGG 405 RQQPGESRLSLVQRN 119 TCAACATTTTCTTCCACAAATTGCGAAATATAGTC 450 VNIFKHIIPQIAKYS 134 CAAGn=CTATTCTTGTTATn=TTTCAAATCCAGTTGACTT 495 PSAILVIVSNPVDLM 149 CATATGTTGCCTGGAAATTGTCAAACTTTCCTCGTAATCGTGTGA 540 TYVAWKLSNFPRNRV 164 TTGGTTCTGGZUuZGAATCTTGATTCTGCJUiGATTTCGACATCTTA 585 IGSGTNLDSARFRHL 179 TCGCGGAAAAATIY3AATTTGTCACCTGTCAGTGTTCATG 630 IAEKLNLSPVSVHGW 194 TCATCGGCGAACATGGCGATTCAAGTGTTCCAATGTGGAG 675 IIGEHGDSSVPMWSG 209 TTAACGTGAGTGGGZUATGTCGATTCATCCAAGAATAG 720 VNVSGKCLNSIHPRI 224 GATATCCGGATGGACCAGAAGGTTGGGATAAAATACATACATAAACAAG 765 GYPDGPEGWDKIHKQ 239 TTGTCGATGGGGCATACGATGTAATAAGATTAAAAGGATACACAA 810 VVDGAYDVIRLKGYT 254 ACTGGGCTATTGGTCTGAGCTGTGCAGAATTGCTTGCA?iCTATTT 855 NWAIGLSCAELLATI 269 TACATCACAGACATAGAATCCATCCAGTTACTTGTTTTGTGAAGG 900 LHHRHRIHPVTCFVK 284 GCCGATATGGAATCACTGATGATGTATGCCTATCTCTACCTTGTG 945 GRYGITDDVCLSLPC 299 TTTTGZUXX'GTAATGGAGTCAACTCTATTGT~TGTTGATTTGA 990 VLNCNGVNSIVNVDL 314 CTGCTGAAGAAGAAGCAATTGCAATGACTATTG 1035 TAEEEAMIKKSAMTI 329 CTGATGTG CAAAAAGGCCTGAAATGGTAAAATTTACCAGAATACT 1080 ADVQKGLKW* 338 ACATAAAAAGCTTTCATTGTGCTCTTTCTCAAAGCGCTTTCAAGT 1125 CCCTTTAATGTCATGGCCGATATGCTAAAAGACCCAGACTTGT 1168
FIG. l.+DNA and deduced amino acid sequence of Styela pli- cata L_dh.
2 of 11 and 3 of 15 clones and may have been PCR errors. Two of the silent differences (positions 923 and 971) may be the result of heterozygosity, as two bands of relatively equal intensity were seen at these positions on autoradiographs of directly sequenced (uncloned) in- dependent PCR products (data not shown).
The Styelu LDH protein that we inferred from the nucleotide sequence (assigning the start codon according to the properties of vertebrate translation start sites listed by Kozak 1996) is 338 amino acids long including the initial M; vertebrate LDHs, in comparison, range from 332 to 334 amino acids in length. The additional length of the tunicate sequence is in the N-terminal arm (Abad- Zapatero et al. 1987 and included references), a region that has previously been shown to also exhibit the great- est degree of sequence variability among vertebrate LDHs and is reduced or absent in bacterial sequences (Li et al. 1983). The only other region of length differ- ence among chordate LDHs reported to date is near the carboxy terminus (fig. 2); at the penultimate position, all vertebrate LDH-Bs and the LDH-B-derived teleost fish
LDH-C (Quattro, Woods and Powers 1993) exhibit an extra amino acid that is missing in mammal LDH-Cs, lamprey LDH, and all vertebrate LDH-As. Styelu LDH, like that of the nematode and the less reliably alignable plant LDHs is also missing an amino acid at this posi- tion. The tunicate sequence, with its greater similarity to vertebrate sequences in this region, provides addi- tional evidence that the absence of the amino acid is the primitive condition and that the amino acid was added in the common ancestor of LDH-Bs and teleost fish LDH-C. Interestingly, there are no reversals of this char- acter evident in the vertebrate sequences examined to date.
Summaries of the pairwise sequence identities of LDHs are shown in table 1. Styelu LDH is approxi- mately equally similar to all vertebrate LDHs, with the highest similarities to certain LDH-As and the lowest to mammalian LDH-Cs. The lowest within-vertebrate sim- ilarity is lower than some of the similarities of vertebrate and Styela LDH. Compared to other groups outside the Vertebrata, Styela-to-vertebrate similarities are approxi- mately the same as those between nematode and verte- brate LDH, and greater than those between plants and vertebrates, apicomplexans and vertebrates, and bacteria and vertebrates.
Nonorthologous LDHs of Apicomplexan Protists and Cyanobacteria
A neighbor-joining analysis of 54 LDH sequences is shown in figure 3. The tree has been rooted at the midpoint; it cannot be rooted in a manner that makes eukaryotic LDHs a monophyletic group because of the clustering of the cyanobacterial Synechocystis LDH with the Apicomplexan protist LDHs of Plasmodium and Toxoplasma and the location of this group on a branch uniting Bifidobacterium LDH and all other bacterial LDHs. The node uniting Synechocystis with Plasmodi- urn and Toxoplasma is very strongly supported (100% bootstrap support). This result, along with the location of the midpoint root, suggested to us that these three LDHs might not be orthologous to the other LDHs in the tree. A search of GenBank using BLASTP revealed that the closest matches to these sequences were to the malate dehydrogenases (MDH, E.C. 1.1.1.37) of three eubacteria (Bacillus subtilis [Jin, de Jesus-Berries, and Sonenshein 19961, Bacillus sp. [Wynne et al. 19961, and Chlorojlexus aurantiacus [Synstad, Emmerhoff, and Sir- evag 19961). The cytoplasmic and mitochondrial MDHs of eukaryotes and eubacterial MDHs have been shown to be related to L-LDHs in both sequence and, especial- ly, three-dimensional structure (Birktoft et al. 1982). Se- quence comparisons suggested that LDHs and MDHs represent two distinct groups whose members are more closely related among themselves than to members of the other group (Iwabe et al. 1989). Recently, an MDH from the archaebacterium HuZoarcuZa marismortui was cloned that exhibits greater sequence similarity to LDHs than to other MDHs (Cendrin et al. 1993). The MDHs of the three eubacteria mentioned above appear to ex- hibit a similar relationship to LDHs (Jin, de Jesus-Ber- rios and Sonenshein 1996; Synstad, Emmerhoff, and
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019
Tunicate LDH 1277
10 20 I I
Hsa A _-____---_ -MATLK---- Hsa C .......... ..S.V ..... Fhe A .......... .-MSTQ .... Sac A .................... Pma Hsa B :::::::::: 1::
ST ............
Fhe B .......... ..SSVL .... Fhe C ........... ..SVL .... Spl ........... ..D..GFD I Cel .......... MAS.I ..... Hvu A MHKASSLSEL GFDAGGASSG Blo .......... .----- .... Taq .......... .----- .... Mhy .......... .----- .... Bst .......... .----- .... Pfa .......... .----- .... ssp .......... .----- ....
Hsa A Hsa C Fhe A Sac A Pma Hsa B Fhe B Fhe C SPl Cel Hvu A Blo Taq MhY Bst Pfa Ssp
Hsa A Hsa C Fhe A Sac A Pma Hsa B Fhe B Fhe C SPl Cel Hvu A Blo Taq MhY Bst Pfa Ssp
Hsa A Hsa C Fhe A Sac A Pma Hsa B Fhe B Fhe C SPl Cel Hvu A Blo Taq MhY Bst Pfa SSP
110 120
30 40 I I
-DQLIYNLLK EEQTP----- .E.. .EK.IE DDENS..... .EK..SHVM. ..PVG.C... ..K..GH.AT SQEPRS.... .GK..HEMVP SKERDPP... .EK..APVAE ..A.VP.... .QK..TP.AS SSAE.P.... .HK..TP.AC SSPE.P.... MSE.FTEVCP DIPKSG.... .-EVFAEIAA PVENS..... FFRPVADGCP ATP.SSAVPH .------MAE TTVK...... .--------- -----..... .MKKFNKKGE IKLMK.... . .------MNA MKNNG..... .__--____- __m.****. .-MN.LEYAP IACQS.....
130 140 I I I I
KIVS-GKDYN VTANSKLVII TAGARQQEG- ----ESRLNL .VT......S .S...RI..V . . . . . . . . . . . . . ..T..A. ... A.D . ..S .... ..V .VV .................... ......... S .S.G....V. .................... ... A.D . ..S . ..G.R ..W ....................
.A.D :;I.A.D ..
..S ........
..I .VV ..V ..A .R ..VV .. ..V ................................
.A.D :~NG.D ..
..S ..S..RI .VV . ..V ......... ..R ....
. .S .S...RI ..V ..... ..P ........ ..S. TVKA.DT ..S I..G...C W .... ..R ...... ..T..S. R.S-..T .AA ..K..D . ..V .... ..IP ..... ..T .... S.DG.SD.PE ICRDADM.V. . ..P..KP .... ..Q...E. VW.R.SGW.E DLEGARV ..V A..VA.RP ..... ..T..Q. IS .. .RYE.K DLKDADFIV. . ..RP.KP ..... ..T..E. VDIW.HG ..D DCRDAD..V. C...N.KP ..... ..T..D. CK..GSNT .D DL.GADV ..V . ..FTKAP .K SDKEWN.DD. SKIIGTNE.E A..G.DV.V. . ..LPRRP .... ..M..DD.
70 80 I I I I I I
--QNKITWG VGAVGMACAI SILMKDLADE LALVDVI--E DKLKGEMMDL QHGSLFLRTP ... C ... I .. T .......... ..L .......... ..A ..L .............. ..FS .S ..R..V .... ..M....S .. .V.L...C ...... ..M ....... ..A .... ..A...K .H Y
::Hh:;:: ::b::::i:: V .M
:+:iR::::: .::::.v ...... .H.A
:i:::::::: L:::::.K.A .. N ......... ..R..V ...... E::::::; ::
.G.S ........ ..L .................. ..Q ..
.LR..C ...... ..M ... .R ......... ..L...K .S ..R..V .... ..Q .... ..V T..LRE ........ ..V ... ..V ............ ..K .. ..PS.V .... ..M.....G M .VVL.G.CTD .V....V ..Q . ..Q..V ........ ..JZN I ..HG.V .... ..Q .... ..Y . ..QQN..N. .C....V ..A ........... ..LA.T.H C RRLT..S.I. A.N....I .Q T..TQN .... I....AL ..P . ..R..AL .. ..AAA..PR V
-T.LA.I. A . . ::-V.VGI . .
..STL .F AAAQRGI.R. IV.E.IA ..K ERVEA.VL.M .. ..S.YP .V S.F..S.T .Y ALVLQGV.R. VV .. .LD ..R KLAQAHAE.I L.ATP.AH-.
..PI..ALI. A.N..NSFL Y AAMNQG..S. YGII.IN ..P .FAD.NAF.F EDA.AS.PF.
..GARVV.I. A.F..ASYV F ALMNQGI ... IV.I.AN ... S.AI.DA ..F N..KV.APK.
. . KA..VL . . S.MI.GVM.T L.VQ.N.G-D VV.F.IV ..K NMPH.KAL.T S.TNVMAYSN
.. -WQV .... A.N..RTL .Q RLVQQNV.-N VV.L.IV ..P GLPQ.IAL .. MAAQSVEEYD
150 160 170 180 190 200 I I I I I I
VQRNVNIFKF IIPNVVKYSP NCKLLIVSNP VDILTYVAWK ISGFPKNRVI GSGCNLDSAR ..... A.M.1 . ..AI.H ... D..I.V ........ ..IV ... ..L.VT ............. .............. I ..... ..I..V .............. L....RH .... ..T ...... ............. DI..H .. D.1I.V .... ..V ....... L..L.MH.I. .......... .............. I ..... ..I..V .............. L..L..H ............. ...... V ......
. ..g:::: D.IIIV ......... ..T .. L..L..H .............
...... v ..c ..TI.V .... ..V....T .. L..L..H .... ..T ...... D ........ H ... LI.RH .. D.IIIV .... ..V....T .. L..L.MH .... ..T ...... ......... H . ..QIA .... SA1.V ..... ..LM ...... L.N..R ..... ..T ...... ..... E ... G . ..QL ..... DTCI.V .... ..V....T .. L..L.RE ..F . ..T ...... L ..ALYR K .V.P.AEH .iT
.. DAL..V ..V .......
....... L....AS ..T....S. .. .L.A .M..L..VA. .AIYMLIT ..... ..A.H..Q. LT.L.E.QIF ..T ......
LD..AQV.A D VV.KIL.AA. EAV...AT .. ..VM.Q..Y R L..L.PE ..V . ..TI..T .. .AD.IR.IRE .ALK.KESGF SGISI..A ... ..I.RAYR D A...SDQK ... ..TV..T .. .DK.IA..R S .VES.MASGF QGLF.VAT ...... ..AT .. F..L.HE .... ..TI..T .. LPL.NK.MIE .GGHIK.NC. .AFIIV.T .. ..VMVQLLH Q H..V...KI. .L.GV..TS. LGK.A..VA Q GAREALR ... .AI.IV.T .. L.VM..L ... VT.L.SQ ..M .MAGV .....
210 220 230 240 250 260 270 280 290 300 I I I I I I I I I I
FRYLMGERLG VHPLSCHGWV LGEHGDSSVP VWSGMNVAGV SLKTLHPDLG TDKDKEQWK- EVHKQVVESA YNIKLK--G YTSWAIGLSV ADLAESIMKN . . . . I..K.. . ..T.....I I......... L...V..... A....D.K.. ..S...H... NI....IQ.. ..I....... . . . . . . . . . . M..VG..L.. H . . . . . . KFH L..S.....I V........A . . ..V.I... ..Q..N.NM. A.G.S.N... .L..K..DG. . . . . . . . . . . . . . . . ..M.. . ..V...V..
. . . . . . . . . . . . SS...... I......... . . . . . . . . . . . ..E...E.. . . . . ..N... KL..D..D.. . . . . . . . . . . . . . . . . . . . . . . . ..T.... S . . . . . . . . . .NSA.....I I......... . . ..V..... G.QS.N..I. .P..G.D... S......D.. . . . . . . . . . . . . . . . . . . . . . . . ..T.L..
. . . . . A.K.. I..S.....I . . . . . . . ..A . . ..V..... ..QE.N.EM. ..N.S.N... . . ..M..... . . . . . . . . . . ..N....... . ..I..ML.. .M.A.... I.AS.FN... . . . . ..T... . . ..A..... ..QK.N.EI. ..G....... AT..A..D.. . . . . . . . . . . ..N....F.. . ..T...V..
::F..ADK.. I.SS.FN..I . . . . ..T... T N.Q..N.NI. ..F.E.N... .T..M..D.. . . . . . . . . . . ..N....... . ..T..L.R. ..H.IA.K.N LS.V.V...I I......... il:::V::&i C.NSI..RI. YPDGP.G.D. KI.....DG. .D..R..... ..N......C .E.LAT.LHH ..F.LS.K.N IA.S.....I I........A . . ..V..... T.HEIK..I. EKT.N.H.EA .I..K..D.. ..I....... . . . . . . . . . . .KI.QG.FS. ..F.IA.H.D .NAQDVQAYM V....... .A I..SIS.G.M PAFKSLR.SH RSF.EAALE. GIRRA..GG. . . ..G..... . . . . . ..Y.. .S..A.LLRD L.F.IAQQT. .NVKNV.AYI A......E.. L.ESATIG.. PMCDWT.LP. H.PLDADKRE .I.QE.KNA. .KI.NG.... A.NY...M.G V.II.AVLHD ..A.LAQH.L .A.Q.V.AY. V......E.L . ..SAQ.G.. D.EAFAQAR- GRALTPDDRL RIDEG.RRA. .RI.EG.... A.YYG..AGL .R.TRA.LTD LQFAIAK.AK .S.N.VQAY. M...... .FV AY.NIKI..E CFCAYSKLT- GIDSS-NYEK .LEYP.SRR. ..I.NR...R A.FYG..AAI .KIVSN.I.D ..F.L..YFS .A.QNV.AYI I.....TEL. . ..QAYIGVM PIRK.VESK- GEEAQ-KDLE RIFVN.RDA. .QI.EK.... A.YYG.AMGL .RVTRA.LH. LK.YISQK.N .C.RDVNAHI V.A..NKM.L LKRYIT.G.1 P.QEFINN-- KL-ISDAELE AIFDRT.NT. L.IVN.H..A SPYV.PAAAI IEM...YL.D LKAFIAMK.. AC.SDINTL. ..G...LML. LPRYCT.S.. PITE.I.P-- ------.TIE .LVERTRNGG A.IAA.LQT. TAYY.PAS.A .VMV...LR.
310 320 330 340 350 360 370 I I I I I I I
LRRVHPVSTM IKGLYGIKD- -DVFLSVPCI LGQNGISDLV KVTLTSEEEA RLKKSADTLW GIQKELQ-F- V .E. .EL..
:iIi::::::i V~:iiI:i.. .E.. .I..V ..R..V..V. .IN.N..... LF....E... N...D.I...
..I..V ..NS.LT.VI HM..KP...K Q.V...E... .V....T.L. .C........ V.DF....N. . . . . ..L..V .DNH...NI. .MK.KPD..Q Q.Q...T... D...D.K... . . . . . . . . . H C..QH.VH.. . . . . ..L..V ..SE..T.II NQ..KK.... QVQ...E... NV....T... .S.I...... V..M...EN. .E....L... .NAR.LTSVI NQK.KDD.V. Q......... D...D.KDL. .S........ V.DMF..GE. .E....L..V .NGS.VGSV. NM...AA.V. Q......... . . ..D.KDL. MN.I...... A..M...G.. .E.Y..L..V .NSG.VGSV. NM...D..V. Q.QG..S... D...D.RDI. RH.1.. .TCF V..R...T.. . . .C..L..V .NC..VNSI. N.D..A.... MI....M.IA DV..G.K.W. S.N.FAL..N V..FH..N.. . ..Y..L.W ..SA.LTHV. .QQ..EA.VQ K.HN..KA.L N.NG1V.M. Q .VL AS.FH..S.G HE....L.AR Tiii::.
..RG..LGVA EMD..EA.A. Q.RR..K... ENCQL.D.L. S. L.DFH..S.. .-ICM.. .TL .NRQ.VNNTI NTPVSDK.L. A..R..E..K ETAAQFG...
EKG.FT..LF TPNE.VEE. .-.A..L.R. ..AR.VEATL YPR.NE..RQ A.RR..EI.K .AASA.G... TKNIMIAGAN LR.E..FHG. .-.NIG..W ..A.. .EKII EIS.NDK.KE KFA..VAIID K.YQDAIKNI ENAILT..AY LD....ER.. .-.YIG..AV INR...RNI EIE.NDD.KN .FHH..A..K SVLARAFTR. .KK.LIC..L LE.Q..HS.. .-1.GGT.W ..A..VEQVI ELQ.N...-- --.AKF.EiAI AET.RMKALA QS.IL.AA.Y LD.A..L... .-I..G...R ..CR.VE.IL E.Q..P..K. A.HL..FiAVR LNIDVALAMV
_--
. . .
. . .
. . .
. . .
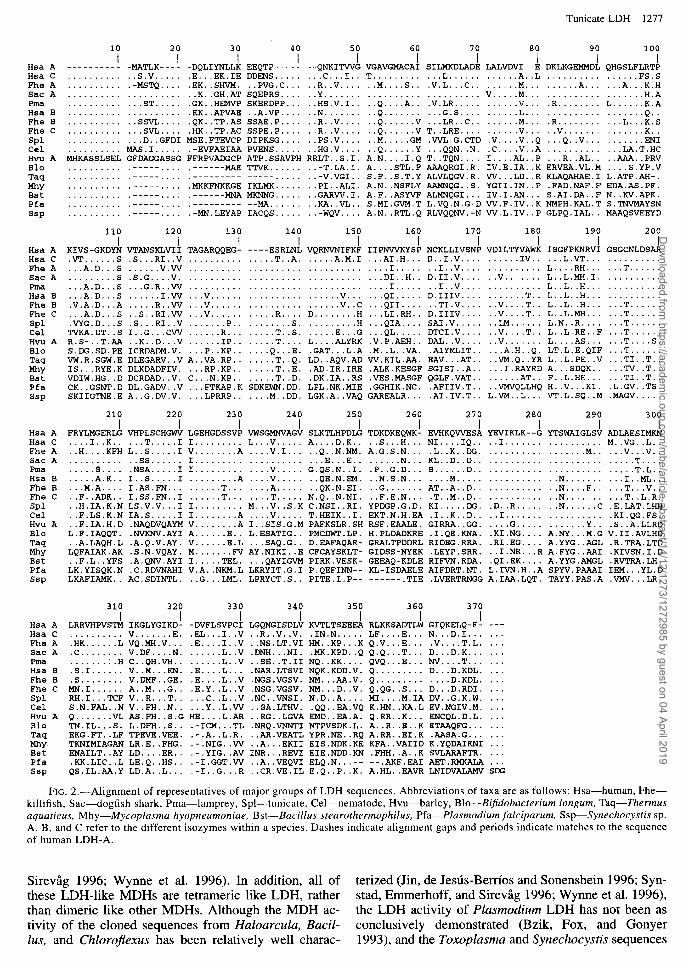
FIG. 2.-Alignment of representatives of major groups of LDH sequences. Abbreviations of taxa are as follows: Hsa-human, Fhe- killifish, Sac-dogfish shark, Pma-lamprey, Spl-tunicate, Cel-nematode, Hvu-barley, Blo--B$dobacterium Zongum, Taq-Z’hermus aquaticus, Mhy-Mycoplasma hyopneumoniae, Bst-Bacillus stearothermophilus, Pfa-Plasmodium falciparum, Ssp-Synechocystis sp. A, B, and C refer to the different isozymes within a species. Dashes indicate alignment gaps and periods indicate matches to the sequence of human LDH-A.
Sirevgg 1996; Wynne et al. 1996). In addition, all of terized (Jin, de Jesus-Berries and Sonenshein 1996; Syn- these LDH-like MDHs are tetrameric like LDH, rather stad, Emmerhoff, and Sirevag 1996; Wynne et al. 1996), than dimeric like other MDHs. Although the MDH ac- the LDH activity of Plasmodium LDH has not been as tivity of the cloned sequences from Haloarculu, Bacil- conclusively demonstrated (Bzik, Fox, and Gonyer Zus, and Chlorojkxus has been relatively well charac- 1993), and the Toxoplasma and Synechocystis sequences
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1278 Stock et al.
Table 1 Summary of Pairwise Differences Among LDH Sequences
Pairwise Comparisons Range of Distance@
Bacteria to vertebrate .................. 0.59-0.73 Apicomplexa to vertebrate. ............. 0.68-0.73 Plant to vertebrate. .................... 0.46-0.54 Nematode to vertebrate ................ 0.35-0.46 Tunicate to vertebrate. ................. 0.36-0.44 Within vertebrates. .................... SO.40 Tunicate to vertebrate LDH-A. .......... 0.35-0.39 Tunicate to lamprey ................... 0.36 Tunicate to vertebrate LDH-B ........... 0.37-0.40 Tunicate to mammal LDH-C ............ 0.39-0.44 Tunicate to killifish LDH-C ............. 0.38
a Distances were calculated using the alignment in figure 2, with gaps in the pairwise comparisons deleted and all other positions considered. Distances are proportions of nonidentical residues.
have been identified as LDHs only by sequence simi- larity to Plasmodium LDH (Kaneko et al. 1995; Yang and Parmley 1995). Regardless of whether the “LDHs” of Plasmodium, Toxoplasma, and Synechocystis actually function as LDHs or MDHs, we suggest that they are not orthologous to the other LDHs in figure 3. Strong support for this hypothesis is provided by the presence in B. subtilis of a conventional LDH (Fig. 3) in addition to the LDH-like MDH described above.
Phylogeny of Eukaryotic LDHs
The midpoint rooting of figure 3, which supports a monophyletic group of nonprotistan eukaryotic LDHs distinct from bacterial LDHs, is consistent with the re- sults of simultaneous analysis of LDH and MDH (Iwabe et al. 1989). This monophyletic group of plant and an- imal LDHs was found in 100% of the bootstrap NJ trees. Plant LDHs also form a monophyletic group with 100% bootstrap support, although this is not surprising given that all of the species are members of the family Po- aceae. The monophyly of animal LDHs was found in 99% of the bootstrap NJ tree. All of the above clusters were also found in parsimony analyses (not shown).
Phylogenetic Position of Styela LDH
The phylogeny of the LDHs of the three animal phyla matches that of the organisms. In the tree in figure 3, nematode LDH clusters outside of urochordate (Stye- la) and chordate LDHs, but with only moderate boot- strap support (60%). Figure 3 shows significant support (97% of the bootstrap trees) for Styelu LDH being out- side of a monophyletic group of all of the vertebrate LDHs. This result indicates that all of the gene dupli- cations within the vertebrates examined occurred after the divergence of tunicates and vertebrates. In an at- tempt to provide increased resolution of the relation- ships of tunicate and vertebrate LDHs, we conducted a neighbor-joining analysis without the more distantly re- lated and less reliably alignable bacterial and protistan LDHs and have rooted the resulting tree between plant and animal LDHs (fig. 4). Such an analysis shows higher support for the theory that the tunicate LDH is more closely related to vertebrate LDH than is nematode LDH (92% bootstrap support), but lower support for the
monophyly of vertebrate LDHs (87% of the bootstrap trees).
We used maximum-parsimony analyses to evaluate the relative support for alternative hypotheses on the phylogenetic relationships of tunicate LDH (fig. 4 and table 2). It has been suggested based on immunochem- ical comparisons that tunicate LDH is most closely re- lated to LDH-C and lamprey LDH (Baldwin, Mortimer, and Patak 1988). These authors assumed that mammal and fish LDH-Cs were orthologous, although this rela- tionship was strongly contradicted by a subsequent phy- logenetic analysis of a teleost LDH-C sequence (Quat- tro, Woods and Powers 1993). Not surprisingly, given this latter result, the forced grouping of Styela LDH, lamprey LDH, and mammal and fish LDH-Cs required a large number of extra steps (31 for equal weighting, 46 for PROTPARS weighting) relative to the most par- simonious trees and in the case of PROTPARS weight- ing was significantly worse as judged by the method of Templeton (1983). Relaxing the requirement that mam- mal LDH-C be a part of this cluster (Baldwin, Mortimer, and Patak’s [1988] immunochemical data were based on fish LDH-C) still results in trees significantly worse than unconstrained trees. Clustering of tunicate LDH with mammalian LDH-Cs could also be rejected, but a spe- cific relationship with fish LDH-C could not. Additional phylogenetic positions of the tunicate LDH inside the vertebrate LDH clade (not necessarily suggested by any authors) were also evaluated with respect to the number of extra steps. The clustering of the tunicate LDH with gnathostome LDH-B and teleost LDH-C required the fewest additional steps. Hypotheses clustering Styela LDH with LDH-C remained among the least likely.
Phylogeny of the Vertebrate LDHs
The phylogenetic relationships among vertebrate LDHs were investigated with plant and invertebrate out- groups as shown in figure 4. Parsimony and neighbor- joining analyses gave similar topologies, although the bootstrap values were generally higher with neighbor- joining. The values discussed below are from this latter analysis. A branch uniting all of the jawed vertebrate (gnathostome) LDH-Bs as well as teleost fish (killifish) LDH-C, Xenopus LDH-A, and Xenopus LDH-C was found in 90% of the bootstrap trees. The tree topology supports a number of gene duplications within this clade. Ninety-three percent of the bootstrap trees support a derivation of teleost LDH-C from teleost LDH-B after this group diverged from tetrapods, as initially proposed on immunochemical grounds (Whitt 1969; Marker-t, Shaklee, and Whitt 1975) and subsequently supported at the protein sequence level (Quattro, Woods, and Pow- ers 1993). Two duplications are also indicated (with 100% bootstrap support) in the Xenopus lineage after it diverged from amniotes. One is likely to be the result of tetraploidy (Kobel and Du Pasquier 1986), while the other may be a gene duplication postulated by Wolff and Kobel (1982). Stock and Powers ( 1995) concluded from similar results that LDH-A and LDH-C have been in- correctly designated in Xenopus, with LDH-A probably remaining to be cloned. Aside from these likely dupli-
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

Tunicate LDH 1279
_c Cattle LDH-A Pig LDH-A
loo_ i Rabbit LDH-A
Mouse LDH-A 77 _ ' Rat LDH-A
- Human LDH-A 81 - Chicken LDH-A
Fox LDH-C Human LDH-C
45 ‘M-“i;wL;FC
51 - Dogfish shark LDH-A Lamprey LDH
Kiilifish LDH-A Killifish LDH-B
Killifish LDH-C
100 , Tunicate LDH Nematode LDH
58 100
100 8. longum LDH
( T. aquaticus LDH T. caldophilus LDH
D. radiodurans LDH T. maritima LDH
M. hyopneumoniae LDH M. genitalium LDH
87
P. acidilactici LDH
95 S. mutans LDH
FIG. 3.-Neighbor-joining tree of 54 LDH sequences. The tree is rooted at the midpoint of the longest path length. Amino acid distances were calculated with a gamma correction (a = 2). Alignment positions l-44 were omitted from consideration, and the pairwise deletion option of MEGA was used to treat alignment gaps. Numbers at nodes indicate the results of a bootstrap analysis with 1,000 replications. Bootstrap support within mammal LDH isozymes has been omitted for clarity. Numbers in a larger font indicate the support for the interrelationships among the major groups of eukaryotic LDHs.
cations, the phylogeny of the LDH-B genes matches the well-accepted organismal phylogeny of (teleost fish, (frogs, (birds, (mammals)))).
Although LDH-Bs and their derivatives form a well-supported clade in figure 4, the sister group of this clade (lamprey LDH + gnathostome LDH-A + mam- mal LDH-C) is found in only 47% of the bootstrap trees. Within this latter group, there is strong support, how- ever, for the theory that mammalian LDH-Cs are derived from within LDH-A. The strongest contradiction of the proposals that LDH-C is the earliest divergence among vertebrate LDHs (e.g., Li et al. 1983; Baldwin, Morti- mer, and Patak 1988; Crawford, Constantino, and Pow- ers 1989; Baldwin, Patak, and Mortimer 1992; Tsoi and Li 1994; Tsuji et al. 1994; Mannen et al. 1996) is pro-
vided by the node uniting dogfish shark LDH-A with mammal LDH-C and tetrapod LDH-A (96% bootstrap support). Further support for a derivation of mammalian LDH-Cs from LDH-A is provided by the node uniting them with chicken and mammal LDH-As (80% of the bootstrap trees). Mammalian LDH-Cs are specifically placed as the sister group of bird and mammal LDH-As (77% bootstrap support), a position that requires gene losses in a number of amniote groups, as discussed by Matson (1989). This grouping may be due to a rapid rate of evolution of LDH-C, which is obvious from in- spection of the within-mammal divergences relative to those of LDH-A and LDH-B (fig. 4; Whitt 1984). A clustering of mammal LDH-C and mammal LDH-A, which does not require additional gene losses, is found
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1280 Stock et al.
77 27
8C
Cattle LDH-A
Pig LDH-A
IL Rabbit LDH-A 99 60 rt Mouse LDH-A
Rat LDH-A
Human LDH-A
) 1 - Chicken LDH-A I I I- Fox LDH-C ,
- Human LDH-C 97
I Mouse LDH-C
Rat LDH-C
Dogfish shark LDH-A
I- Killifish LDH-A
88 60 ~
96 91
Xenopus LDH-A
Duck LDH-B
Chicken LDH-B
L Mouse LDH-B
Tunicate LDH
Nematode LDH
FIG. 4.-Neighbor-joining and parsimony analyses of 24 verte- brate LDH sequences with plant and invertebrate outgroups. The to- pology indicated was obtained from a neighbor-joining analysis con- ducted as described in figure 3. Numbers above the nodes indicate the results of a bootstrap neighbor-joining analysis with 1,000 replications. Numbers below the branches indicate the support obtained in a boot- strap parsimony analysis. Some sequences with close relatives were deleted from the parsimony analyses to reduce the amount of time required; these were cattle LDH-A, pig LDH-A, rabbit LDH-A, rat LDH-A, Xenopus LDH-B, duck LDH-B, pig LDH-B, and barley LDH- B. Three most-parsimonious trees were found with 100 replications of a random addition sequence and TBR branch swapping (Swofford 1991). These differed from the topology shown by placing mammalian LDH-Cs within or as the sister group of mammalian LDH-As (the only aspect of the three trees in which they differed among each other) and by grouping killifish LDH-A with lamprey LDH. The bootstrap par- simony results indicated were obtained with 100 bootstrap replications, each with 10 replications of a random addition sequence and TBR branch swapping.
in the most parsimonious trees and in 48% of the boot- strap trees from a maximum-parsimony analysis (not shown).
The phylogeny of gnathostome LDH-A and lam- prey LDH (suggested to be orthologous to LDH-A by Stock and Whitt [1992]) in figure 4 does not match the generally accepted phylogeny of the organisms; moving the LDH-A of the killifish to a sister group relationship with tetrapod LDH-A would bring the two phylogenies into agreement. Such a position of the killifish LDH-A is contradicted by two weakly supported nodes (47%
and 48% bootstrap support) and one strongly supported one (96% bootstrap support). The additional gene du- plications and losses required by the tree in figure 4 over a tree matching the organismic phylogeny have been discussed by Tsuji et al. (1994), Quattro et al. (1993, and Stock and Powers (1995).
Discussion Gene Duplications Leading to Multiple Vertebrate LDHs Occurred After the Divergence of Tunicates and Vertebrates
Our analyses strongly support a sister group rela- tionship between the LDH of Styela plicata and a clade consisting of all of the vertebrate LDHs that have been sequenced to date (those of lamprey and jawed verte- brates). If it is assumed that the locus we sequenced is the only one encoding LDH in the genome of Styela (see below), then this phylogenetic result suggests that all of the LDH duplications represented by the present collection of vertebrate sequences occurred after the di- vergence of vertebrates and tunicates. This pattern of duplication is consistent with the hypotheses of Markert, Shaklee, and Whitt (1975) and Fisher et al. (1980), which were formulated largely in the absence of se- quence data. Our phylogenetic results strongly contra- dict the hypothesis of Baldwin, Mortimer, and Patak (1988) that tunicate LDH is more closely related to the LDH of lampreys and even more so to the LDH-C of mammals and teleost fish than it is to the LDH-A and LDH-B of jawed vertebrates. This hypothesis was based on immunochemical comparisons of the LDH of the tu- nicate Pyura stolonifera (distantly related to S. plicata) with the LDH-A, LDH-B, and LDH-C of a teleost fish, as well as on comparisons of amino acid compositions of tunicate and other vertebrate LDH isozymes. The lack of agreement of our phylogenetic conclusions based on sequence comparisons with earlier conclusions based on cruder immunochemical and amino acid composition analyses suggests that hypotheses on the relationships of echinoderm LDH (Baldwin, Patak, and Mortimer 1992) and hagfish LDHs (Baldwin, Mortimer, and Patak 1988 and included references) to the LDHs of lamprey and jawed vertebrates that are based on similar evidence are no longer persuasive.
LDH Locus Number in Tunicates and Other Invertebrate Groups
Our PCR amplifications of Styela L.dh did not pro- vide any evidence for the existence of more than one locus in this species. A number of the primers we used are coniplementary to regions with amino acid sequenc- es conserved among multiple vertebrate paralogs. This suggests that if the Styela locus we amplified were or- thologous to a specific type of vertebrate LDH, then the primers would be capable of amplifying Styela genes orthologous to other vertebrate genes. However, ampli- fication of additional loci may have failed for a number of reasons, including restricted expression or unique substitutions along the Styela lineage. A more system- atic determination of the number of loci present is pos-
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

Tunicate LDH 128 1
Table 2 Lengths of Constrained Trees in Parsimony Analyses
EQUAL WEIGHTS~ PROTPARSC
CONSTRAINTS Length Extra Steps Length Extra Stepsd
Nonee.................................................. 928 0 1,103 0 (tunicate, killifish C, mammal C, lamprey) . . . . . . . . . . . . . . . . . . . 959 31 1,149 46” (tunicate, killifish C, lamprey). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 948 20 1,128 25” (tunicate, killifish C) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 936 8 1,119 16 (tunicate, mammal C). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 935 7 1,120 17” (tunicate, all A except Xenopus, lamprey). . . . . . . . . . . . . . . . . . . . 936 8 1,118 15 (tunicate, all A except Xenopus, mammal C, lamprey) . . . . . . . . . 933 5 1,113 10 (tunicate, all B, all Xenopus). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 939 11 1,122 19 (tunicate, all B, all Xenopus, killifish C) . . . . . . . . . . . . . . . . . . . . . 931 3 1,111 8
a Constraints consisted of requiring the indicated group of sequences to be monophyletic with no other relationships specified inside or outside this group. See figure 4 for the sequences included in the analyses.
b Lengths of trees calculated with all amino acid transformations weighted equally. c Lengths of trees calculated with amino acid transformations weighted according to the number of nonsynonymous nucleotide substitutions required. d Asterisks indicate topologies judged to be significantly worse than the most parsimonious tree by the test of Templeton (1983) as implemented in PROTPARS
(Felsenstein 1993). ’ e Unconstrained trees placed the tunicate outside of all vertebrate LDHs.
sible with the characterization of LDH enzymatic activ- ity; such investigations should detect additional loci that have diverged extensively in sequence but not in func- tion. The most thorough study of this type in tunicates is that of Baldwin, Mortimer, and Patak (1988) on P. stoloniferu. Nondenaturing electrophoresis and histo- chemical staining of crude extracts of whole animals and a number of tissues revealed evidence for a single locus, as did analyses of purified LDH activity. Similar elec- trophoretic investigations of LDH activity suggest that echinoderms (Baldwin, Patak, and Mortimer 1992) and amphioxus (Fisher et al. 1980) also possess a single LDH locus. Assuming the generally accepted phylogeny (Peterson 1995) of (echinoderms, (tunicates, (amphiox- us, (vertebrates)))), the simplest explanation of these lo- cus numbers and our phylogenetic results is that the an- cestral state of deuterostomes (a single LDH locus) has been retained by echinoderms, tunicates, and amphiox- us, while LDH duplications have occurred within ver- tebrates after their divergence from amphioxus. This hy- pothesis was proposed previously by Fisher et al. (1980). ’
If, in spite of evidence to the contrary, an additional tunicate LDH locus did exist, the implications for LDH evolution in vertebrates would depend on the phyloge- netic position of the new locus in the LDH tree. Clus- tering with the existing tunicate locus would imply a duplication in tunicates after their divergence from ver- tebrates and would not alter hypotheses of the timing of vertebrate gene duplications. Because of the phyloge- netic position of the known tunicate locus, any other position of the new locus would require that one of the two types of loci was lost in all vertebrates. Placement of the new locus within the clade consisting of verte- brate LDHs would require the loss of at least one type of locus in the tunicate as well. In other words, even if an additional tunicate locus is discovered, it will not be possible to postulate that particular vertebrate duplica- tions occurred earlier than the divergence of tunicates and vertebrates without also postulating a more complex pattern of gains and losses.
The Location of the Root in the Tree of LDHs
We described above the evidence that the LDHs of Synechocystis, Plasmodium, and Toxoplasma are not or- thologous to the other LDHs in the tree and may in fact be more closely related to tetrameric MDHs. With the exception of a weakly supported node placing Bijido- bacterium LDH closer to plant and animal LDHs than are other bacterial LDHs, the tree in figure 3 (rooted at the midpoint) depicts a phylogeny of the remaining LDHs corresponding to the organismal phylogeny of (bacteria, (plants, (nematodes, (tunicates, (verte- brates))))). This provides support for a root separating monophyletic groups of bacterial and eukaryotic LDHs (joining an internal branch with 100% bootstrap support in fig. 3), as any other root position would require a greater number of gene duplication and loss events. The clustering of bacterial LDH outside of vertebrate LDHs has also been obtained in trees of LDHs rooted with MDH sequences (Iwabe et al. 1989; Cendrin et al. 1993; Wynne et al. 1996).
LDH Evolution from Bacteria to Vertebrates
Determining whether large numbers of LDH gene duplications and losses have occurred outside of chor- dates requires more extensive collection of LDH se- quences. Our phylogenetic results obtained with LDHs from a relatively small number of nonchordate taxa im- ply an evolutionary pattern of a single locus persisting in most lineages with occasional (and usually recent) gene duplications. Examples of gene duplications in nonvertebrate taxa include the appearance of LDH-A and LDH-B of Bacillus psychrosaccharolyticus after its divergence from the other Bacillus species in the tree and the LDH-A and LDH-B of barley, which resulted from a duplication after the divergence of barley from the other two members of the family Poaceae (fig. 3). Although the number of LDH loci in invertebrates has not been well characterized, Drosophila melanogaster is believed to have a single locus for L-LDH (Alahiotis et al. 1983), and the evidence for a single locus in an echi- noderm (Baldwin, Patak, and Mortimer 1992) has been
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1282 Stock et al.
discussed above. A number of insects and molluscs are believed to lack LDH activity altogether, which would be loss events according to our phylogenetic trees (e.g., fig. 3). However, the biggest remaining question in the evolution of invertebrate LDHs relates to the orthology of the loci encoding the proteins or enzyme activities that have been described (Long 1976; Siebenaller et al. 1983). All of the LDH sequences that we have analyzed and all known chordate LDHs are specific for the L- stereoisomer of lactate (Whitt 1984). A number of groups of invertebrates have D-LDHs, however, and the presence of D- or L-LDH appears to be mutually exclu- sive for any given taxon (Long 1976). Sequence anal- yses have shown that bacterial D-LDHs form a separate family from L-LDHs that is much more distantly related to the latter than are MDHs (Kochhar et al. 1992). No invertebrate D-LDH sequences are available to allow a determination of their relationship to L-LDH, although Siebenaller et al. (1983) suggested that the two types of LDH were not orthologous based on the sequence of a few short peptide fragments from a D-LDH of a horse- shoe crab. If this hypothesis is correct, then L-LDH would have to have been lost and replaced by D-LDH at least four times in invertebrates (Long 1976).
Relationships Within Vertebrate LDHs
The relationships of vertebrate LDHs that we ob- tained in our analyses are generally consistent with those of Stock and Whitt (1992), Quattro, Woods, and Powers (1993), Quattro et al. (1995), and Stock and Powers (1995). There was strong support for the derivation of teleost LDH-C from LDH-B after the divergence of tel- eosts and tetrapods and a derivation of mammalian LDH-C from LDH-A. This contradicts other sequence analyses that suggested that LDH-C represents the ear- liest divergence among vertebrate LDHs (Li et al. 1983; Crawford, Constantino, and Powers 1989; Tsoi and Li 1994; Tsuji et al. 1994; Mannen et al. 1996). The results of these analyses differed from ours largely in the lo- cation of the root of the vertebrate LDH tree. We con- ducted a series of phylogenetic analyses with varying subsets of outgroup sequences to the vertebrate LDHs (not shown) and obtained a root in or between mam- malian LDH-Cs and other LDHs only with highly di- vergent sequences as outgroups. The analyses of Tsoi and Li (1994) and Mannen et al. (1996), in which the position of LDH-C differed from ours with respect to topological relationships as well, were conducted with the unweighted pair-group with arithmetic mean (UPGMA) method, one that has been shown to be high- ly sensitive to unequal rates of evolution (Huelsenbeck and Hillis 1993).
Our analyses also agreed with previous studies in providing strong support for recent duplications of LDH-B in Xenopus and the monophyly of a group con- sisting of LDH-B and its derivatives in teleosts and Xen- opus. Although the lamprey LDH clusters with LDH-As as in the analyses of Stock and Whitt (1992), this result was not strongly supported, and at least one node strong- ly contradicted the orthology of killifish LDH-A with tetrapod and shark LDH-As. Such a result was also ob-
tained in other studies that included teleost LDH-A se- quences (Tsoi and Li 1994; Tsuji et al. 1994; Quattro et al 1995). This phylogenetic result cannot be brought in line with the orthology of LDH-As by rerooting the tree. Whether or not it is artifactual may be addressed by analyses of the LDH-As of other groups of primitive bony fishes.
The interrelationships of four major lineages of LDHs (shark LDH-A + tetrapod LDH-A + mammal LDH-C; teleost LDH-A, lamprey LDH, and jawed ver- tebrate LDH-Bs and derivatives) were not well resolved in our analyses, as indicated by low bootstrap support. The persistence of at least some of these low bootstrap values in the absence of an outgroup (analysis not shown) suggests that the sequence of more closely re- lated outgroups (amphioxus is the only candidate inver- tebrate) will not by itself resolve all aspects of the phy- logeny of vertebrate LDHs. The collection of additional vertebrate sequences may decrease or eliminate support for potentially artifactual groupings but is unlikely to result in their replacement with strongly supported al- ternatives. The resolution of the phylogeny of vertebrate LDHs may require analyses more sophisticated than the rather simple methods we have used, such as the as- signment of different expected rate categories to differ- ent sites and the consideration of structural or functional information in weighting changes.
Gene Duplications in Early Vertebrate Evolution
Regardless of the exact pattern of gene duplication leading to multiple LDH loci in vertebrates, our analyses suggest that all duplications occurred after the diver- gence of tunicates and vertebrates and that at least some of them occurred early in the diversification of verte- brates. This evolutionary pattern is the same as that sug- gested by Markert, Shaklee, and Whitt (1975) and Fisher et al. (1980). In the latter study, it was specifically pro- posed that the earliest LDH duplications occurred in jawless vertebrates after the divergence of vertebrates from cephalochordates (amphioxus).
Ohno (1970) proposed that extensive gene duplica- tions had occurred at two or more points in chordate phy- logeny, based largely on evidence from genome size es- timates. The first of these episodes of large-scale gene duplication was proposed to have occurred in the lineage leading to amphioxus and vertebrates after the divergence of tunicates, and the second in the fish or amphibian an- cestors of amniotes. Ohno suggested that the latter du- plication was due to polyploidy but that the first may have been due either to polyploidization or extensive tan- dem duplication. Fisher et al. (1980) interpreted the evo- lution of four isozyme systems (including LDH) as in- dicating one or more rounds of polyplodization in the early vertebrate or prevertebrate lineage. Holland et al. (1994) and Sharman and Holland (1996) surveyed the phylogenetic relationships and number of loci encoding genes involved in the regulation of development and also concluded that widespread gene duplications occurred at two periods in chordate evolution. These authors pro- posed a slightly different timing for the duplication epi- sodes than Ohno (1970), however. The first phase was
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

Tunicate LDH 1283
proposed to have occurred close to the origin of verte- brates after the divergence of this lineage from amphi- oxus, and the second near the origin of gnathostomes. Our results indicating that the duplication of LDH loci in vertebrates occurred after the divergence of tunicates is consistent with both the hypotheses of Ohno (1970) and of Holland et al. (1994) and Sharman and Holland (1996). A more precise understanding of the timing of the du- plications leading to multiple LDH loci and their rela- tionship to the duplications of other genes awaits the se- quencing of the LDHs of amphioxus and hagfish.
That the duplications leading to LDH-A and LDH- B involved large genomic regions is suggested by the linkage relationship of these two genes in mammals; mouse and human Ldh-Bs are located on chromsomes 6 and 12p, respectively, and mouse and human Ldh-As are located on chromosomes 7 and llp (Debry and Seldin 1996). In both species, the chromosome segments con- taining Ldh-A and Ldh-B contain additional loci with paralogous relationships between the two regions, and it has been proposed that these chromosomal segments are ancestrally related, having arisen by a process such as polyploidization (Lundin 1993). A relatively recent origin of mammalian Ldh-C from Ldh-A, as we have suggested based on phylogenetic analyses of LDH genes, is consis- tent with the close linkage of these two loci (perhaps within 100 kb-Stubbs et al. 1994). Ldh-C and Ldh-A have also been shown to be linked in some fish species (Morizot 1990) despite evidence from sequence analyses that the former gene is closely related to Ldh-B. This latter result suggests that linkage arrangements in modern species may bear a complex relationship to gene dupli- cation histories. Nevertheless, it would be interesting to compare phylogenies of chordate gene families contain- ing paralogs linked to mammalian or fish LDH loci with the duplication history that we have proposed for the LDH gene family. Phylogenetic congruence between LDH and other linked gene families would provide ad- ditional evidence for our hypothesis.
Acknowledgments
We wish to thank Patricia Schulte for the gift of competent bacterial cells. This study was supported by NSF grants BSR-87-17417 (to G.S.W.), BSR-87-18425 (to D.A.P.), BSR-88-15362 (to G.S.W. and D.W.S.), BSR-90-22648 (to D.A.P.), and DEB 96-29572 (to D.A.P.), an NSF Predoctoral Fellowship in Genetics (to D.W.S.), and an Alfred F? Sloan Postdoctoral Fellowship in Studies of Molecular Evolution (to J.M.Q.).
LITERATURE CITED
ABAD-ZAPATERO, C., J. I? GRIFFITH, J. L. SUSSMANN, and M. G. ROSSMANN. 1987. Refined crystal structure of dogfish M4 apo-lactate dehydrogenase. J. Mol. Biol. 198:445-467.
ALAHIOTIS, S. N., A. ONOUFRIOU, M. FOTAKI, and M. PELE- CANOS. 1983. Drosophila lactate dehydrogenase: develop- mental aspects. Biochem. Genet. 21:199-211.
BALDWIN, J., K. MORTIMER, and A. PATAK. 1988. Do ascidians possess the ancestral subunit type of vertebrate lactate de- hydrogenase? J. Exp. Zool. 246: 109-l 14.
BALDWIN, J., A. PATAK, and K. MORTIMER. 1992. Echinoderm (Holothuria atra) lactate dehydrogenase: affinities with the putative ancestral chordate enzyme. Biochem. Syst. Ecol. 20535-539.
BIRKTOFT, J. J., R. T FERNLEY, R. A. BRADSHAW, and L. J. BANASZAK. 1982. Amino acid sequence homology among the 2-hydroxy acid dehydrogenases: mitochondrial and cy- toplasmic malate dehydrogenases form a homologous sys- tem with lactate dehydrogenase. Proc. Natl. Acad. Sci. USA 79:6166-6170.
BJOURSON, A. J., and J. E. COOPER. 1992. Band-stab PCR: a simple technique for the purification of individual PCR products. Nucleic Acids Res. 20:4675.
BZIK, D. J., B. A. Fox, and K. GONYER. 1993. Expression of Plasmodium falciparum lactate dehydrogenase in Esche- richia coli. Mol. Biochem. Parasitol. 59: 155-166.
CENDRIN, E, J. CHROBOCZEK, G. ZACCAI, H. EISENBERG, and M. MEVARECH. 1993. Cloning, sequencing, and expression in Escherichia coli of the gene coding for malate dehydro- genase of the extremely halophilic archaebacterium HaZ- oarcula marismortui. Biochemistry 32:4308-43 13.
CRAWFORD, D. L., H. R. CONSTANTINO, and D. A. POWERS. 1989. Lactate dehydrogenase-B cDNA from the teleost Fundulus heteroclitus: evolutionary implications. Mol. Biol. Evol. 6:369-383.
DEBRY, R. W., and M. E SELDIN. 1996. Human/mouse homol- ogy relationships. Genomics 33:337-35 1.
FELSENSTEIN, J. 1993. PHYLIP phylogeny inference package. Version 3.5~. University of Washington, Seattle.
FISHER, S. E., J. B. SHAKLEE, S. D. FERRIS, and G. S. WHIT-T. 1980. Evolution of five multilocus isozyme systems in the chordates. Genetica 52/53:73-85.
FROHMAN, M. A. 1990. RACE: rapid amplification of comple- mentary DNA ends. Pp. 28-38 in M. A. INNIS, D. H. GEL- FAND, J. J. SNINSKY, and T J. WHITE, eds. PCR protocols: a guide to methods and applications. Academic Press, New York.
HENDRIKS, W., J. W. M. MULDERS, M. A. BIBBY, C. SLINGSBY, H. BLOEMENDAL, and W. W. DE JONG. 1988. Duck lens E- crystallin and lactate dehydrogenase B4 are identical: a sin- gle-copy gene product with two distinct functions. Proc. Natl. Acad. Sci. USA 85:7114-7118.
HOLLAND, I? W. H. 1992. Homeobox genes in vertebrate evo- lution. BioEssays 14:267-273.
HOLLAND, I? W. H., J. GARCIA-FERNANDEZ, N. A. WILLIAMS, and A. SIDOW. 1994. Gene duplications and the origins of vertebrate development. Development (Suppl.): 125-l 33.
HUELSENBECK, J. P, and D. M. HILLIS. 1993. Success of phy- logenetic methods in the four-taxon case. Syst. Biol. 42: 247-264
IWABE, N., K.-I. KUMA, M. HASEGAWA, S. OSAWA, and T. MIYATA. 1989. Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proc. Natl. Acad. Sci. USA 86:9355- 9359.
JIN, S., M. DE JESCJS-BERR~OS, and A. L. SONENSHEIN. 1996. A Bacillus subtilis malate dehydrogenase gene. J. Bacterial. 178:560-563.
KANEKO, T., A. TANAKA, S. SATO, H. KOTANI, T. SAZUKA, N. MIYAJIMA, M. SUGIURA, and S. TABATA. 1995. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. I. Sequence features in the 1 Mb region from map positions 64% to 92% of the genome. DNA Res. 2:153-166.
KOBEL, H. R. and L. Du PASQUIER. 1986. Genetics of poly- ploid Xenopus. Trends Genet. 2: 3 10-3 15.
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019

1284 Stock et al.
KOCHHAR, S., I? E. HUNZIKER, I? LEONG-MORGENTHALER, and H. HJOTTINGER. 1992. Evolutionary relationship of NAD+- dependent D-lactate dehydrogenase: comparison of primary structure of 2-hydroxy acid dehydrogenases. Biochem. Bio- phys. Res. Commun. 184:60-66.
KOZAK, M. 1996. Interpreting cDNA sequences: some insights from studies on translation. Mamm. Genome 7563-574.
KUMAR, S., K. TAMURA, and M. NEI. 1993. MEGA: molecular evolutionary genetics analysis. Version 1 .Ol . Pennsylvania State University, University Park.
LI, S. S.-L., W. M. FITCH, Y.-C. E. PAN, and E S. SHARIEF. 1983. Evolutionary relationships of vertebrate lactate de- hydrogenase isozymes A4 (muscle), B, (heart), and C4 (tes- tis). J. Biol. Chem. 258:7029-7032.
LONG, G. L. 1976. The stereospecific distribution and evolu- tionary significance of invertebrate lactate dehydrogenases. Comp. Biochem. Physiol. 55B:77-83.
LUNDIN, L. G. 1993. Evolution of the vertebrate genome as reflected in paralogous chromosomal regions in man and the house mouse. Genomics 16:1-19.
MANNEN, H., S. C.-M. TSOI, D. B. PICKFORD, J. A. DONALD, L. J. GUILLETTE, and S. S.-L. LI. 1996. Sequences of the lizard cDNAs encoding lactate dehydrogenase (LDH) iso- zymes A (muscle) and B (heart). Gene 171:303-304.
MARCHUK, D., M. DRUMM, A. SAULINO, and E S. COLLINS. 199 1. Construction of t-vectors: a rapid and general system for direct cloning of unmodified PCR products. Nucleic Ac- ids Res. 19: 1154.
MARKERT, C. L. 1984. Biochemistry and function of lactate dehydrogenase. Cell Biochem. Funct. 2:131-134.
MARKERT, C. L., J. B. SHAKLEE, and G. S. WHITT. 1975. Evo- lution of a gene. Science 189: 102-l 14.
MATSON, R. H. 1989. Distribution of the testis-specific LDH- X among avian taxa with comments on the evolution of the LDH gene family. Syst. Zool. 38:106-l 15.
MORIZOT, D. C. 1990. Use of fish gene maps to predict ancestral vertebrate genome organization. Pp. 207-234 in Z.-I. OGITA and C. L. MARKERT, eds. Isozymes: structure, function, and use in biology and medicine. Wiley-Liss, New York.
OHNO, S. 1970. Evolution by gene duplication. Springer, New York.
PETERSON, K. J. 1995. A phylogenetic test of the calcichordate scenario. Lethaia 28:25-38.
QUATTRO, J. M., D. D. POLLOCK, M. POWELL, H. A. WOODS, and D. A. POWERS. 1995. Evolutionary relations among vertebrate muscle-type lactate dehydrogenases. Mol. Mar. Biol. Biotechnol. 4:224-23 1.
QUATTRO, J. M., H. A. WOODS, and D. A. POWERS. 1993. Sequence analysis of teleost retina-specific lactate dehydro- genase C: evolutionary implications for the vertebrate lac- tate dehydrogenase gene family. Proc. Natl. Acad. Sci. USA 90: 242-246.
SAMBROOK, J., E. E FRITSCH, and T. MANIATIS. 1989. Molec- ular cloning: a laboratory manual. 2nd edition. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
SHARMAN, A. C., and I? W. H. HOLLAND. 1996. Conservation, duplication, and divergence of developmental genes during chordate evolution. Neth. J. Zool. 46:47-67.
SHEETS, M. D., S. C. OGG, and M. I? WICKENS. 1990. Point mutations in AAUAAA and the poly (A) addition site: ef- fects on the accuracy and efficiency of cleavage and poly- adenylation in vitro. Nucleic Acids Res. 18:5799-5805.
SIEBENALLER, J. E, T. L. ORR, B. B. OLWIN, and S. S. TAYLOR. 1983. Comparison of the D-lactate stereospecific dehydro- genase of kmulus polyphemus with active-site regions of L-lactate dehydrogenases. Biochim. Biophys. Acta 749: 153-162.
STOCK, D. W., K. D. MOBERG, L. R. MAXSON, and G. S. WHI’IT. 199 1. A phylogenetic analysis of the 18s ribosomal RNA sequence of the coelacanth Latimeria chalumnae. En- viron. Biol. Fish. 32:99-l 17.
STOCK, D. W., and D. A. POWERS. 1995. The cDNA sequence of the lactate dehydrogenase-A of the spiny dogfish (Squa- Zus acanthias): corrections to the amino acid sequence and an anlysis of the phylogeny of vertebrate lactate dehydro- genases. Mol. Mar. Biol. Biotechnol. 4:284-294.
STOCK, D. W., and G. S. WHITT. 1992. Evolutionary implica- tions of the cDNA sequence of the single lactate dehydro- genase of a lamprey. Proc. Natl. Acad. Sci. USA 89:1799- 1803.
STUBBS, L., E. M. RINCHIK, E. GOLDBERG, B. RUDY, M. A. HANDEL, and D. JOHNSON. 1994. Clustering of six human 11~15 gene homologs within a 500-kb interval of proximal mouse chromosome 7. Genomics 24:324-332.
SWOFFORD, D. L. 1991. PAUP phylogenetic analysis using par- simony. Version 3.0s. Illinois Natural History Survey, Champaign.
SYNSTAD, B., 0. EMMERHOFF, and R. SIREVAG. 1996. Malate dehydrogenase from the green gliding bacterium Chloro- jlexus uuruntiucus is phylogenetically related to lactic de- hydrogenases. Arch. Microbial. 165:346-353.
TEMPLETON, A. R. 1983. Phylogenetic inference from restric- tion endonuclease cleavage site maps with particular refer- ence to the evolution of humans and the apes. Evolution 371221-244.
THOMPSON, J. D., D. HIGGINS, and T. J. GIBSON. 1994. CLUS- TAL W improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position- specific gap penalties and weight matrix choice. Nucleic Acids Res. 22:4673-4680.
TSOI, S. C.-M., and S. S.-L. LI. 1994. The nucleotide and de- duced amino-acid sequences of a cDNA encoding lactate dehydrogenase from Cuenorhubditis eleguns: the evolution- ary relationships of lactate dehydrogenases from mammals, birds, amphibian, fish, nematode, plants, bacteria, myco- plasma, and plasmodium. Biochem. Biophys. Res. Com- mun. 205:558-564.
TSUJI, S., M. A. QURESHI, E. W. Hou, W. M. FITCH, and S. S.- L. LI. 1994. Evolutionary relationships of lactate dehydro- genases (LDHs) from mammals, birds, an amphibian, fish, barley, and bacteria: LDH cDNA sequences from Xenopus, pig and rat. Proc. Natl. Acad. Sci. USA 91:9392-9396.
WHITT, G. S. 1969. Homology of lactate dehydrogenase genes: E gene function in the teleost nervous system. Science 166: 1156-l 158.
- 1984. Genetic, developmental and evolutionary as- pects of the lactate dehydrogenase isozyme system. Cell Biochem. Funct. 2: 134-139.
WOLFF, J., and H. R. KOBEL. 1982. Lactate dehydrogenase of Xenopus luevis luevis and Xenopus borealis depends on a multiple gene system. J. Exp. Zool. 223:203-210.
WYNNE, S. A., D. J. NICHOLLS, M. D. SCAWEN, and T. K. SUNDARAM. 1996. Tetrameric malate dehydrogenase from a thermophilic Bacillus: cloning, sequence and overexpres- sion of the gene encoding the enzyme and isolation and characterization of the recombinant enzyme. Biochem. J. 3171235-245.
YANG, S., and S. E PARMLEY. 1995. A bradyzoite stage-spe- cifically expressed gene of Toxoplasma gondii encodes a polypeptide homologous to lactate dehydrogenase. Mol. Biochem. Parasitol. 73:291-294.
SHOZO YOKOYAMA, reviewing editor
Accepted August 29, 1997
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/14/12/1273/1272985 by guest on 04 April 2019





























