Embed Size (px)
Citation preview

Latest information on Sterkfontein’sAustralopithecus skeleton and a newlook at Australopithecus
R.J. Clarke*
AFTER A DECADE OF CAREFUL EXCAVATION,it is now possible to explain how theskeleton came to be in that isolated
position in the cavern. Furthermore, it isapparent that the fossil does not belong toeither Australopithecus africanus or to A. afa-rensis, but to an individual belonging to, orclosely affiliated to, the second Australopithecusspecies that is represented in SterkfonteinMember 4 and Makapansgat.
IntroductionIt was 10 years ago in this journal that an
announcement was made of the discoveryin the Silberberg Grotto of the SterkfonteinCaves, northwest of Johannesburg, of thefirst-ever associated skull and skeleton ofan ape-man.1 At that time, all we haduncovered were the lower legs with partsof the feet, a complete skull, and a leftradius that was later shown to belong to amonkey. Although it was not possible toplace the fossil into a species, it was seento be that of an Australopithecus differingfrom Australopithecus africanus and wasthought to date to older than three mil-lion years ago (Myr), possibly 3.5 Myr. Itwas subsequently dated to 3.3 Myr bypalaeomagnetism.2 Taphonomically, itwas observed that the skeleton lay on atalus slope and that fossils of other ani-mals were virtually absent, in contrastwith the eastern end of the cave infill inthe Silberberg Grotto, which had a denseconcentration of bones including partialskeletons of monkeys and carnivores. Itwas noted that this raised the question ofhow such an intact skeleton came to be inthat isolated spot in the cavern.
Ten years of slow, careful excavation inthe concrete-like breccia have revealedthe answer to that question. The generalsequence of events that formed theSterkfontein infills has been described byClarke,3 but there is one big mysteryconcerning this skeleton (classified asStW 573) that needs to be explained. Of allthe fossils, animal and hominid, recoveredfrom Sterkfontein since 1936, it is onlyStW 573 that represents a near-completeskeleton, and if the feet had not beenblasted off by lime miners, it wouldundoubtedly have been complete.
At the eastern end of the Silberberg
Grotto, bones were heavily concentratedapparently by water. Although there aresome articulated parts of skeletons ofcarnivores, monkeys and bovids, nothingis anywhere near complete. What thenwere the specific conditions that allowedfor the preservation of StW 573 as a com-plete skeleton? We have so far uncoveredthe skull, left arm and hand,32 right armand hand, right scapula, right clavicle,several ribs and vertebrae, sacrum andpelvis, and both legs with some of the footbones. The rest of the foot bones, blastedaway by mining, have not been found(Figs 1–4).
The length of time taken to extract thisskeleton from the Silberberg Grotto hasbeen governed by several factors. The in-dividual bones comprising the skeleton,whilst in general anatomical relationshipto one another, have in some parts beendisplaced and broken up by ancient envi-ronmental factors, including talus collapseand water. Because of this, it is not possibleto predict exactly where in the concrete-like breccia any skeletal element may
occur. The bones themselves are soft andcan be easily damaged by tools if excava-tion is too rapid. The aim has been to lo-cate the position of all elements of theskeleton within the breccia with minimalrisk of damage prior to removing blocks tothe laboratory for final cleaning. Anothermajor consideration has been the need touncover clues as to what happened tothat individual.
There are several mysteries that wewould like to unravel. How did thatindividual come to be in the cave? Why isthe skeleton so unique in being near-complete? Why are parts of it broken upand at different levels from other parts?Why is much of the left pelvis disinte-grated? Why is this the only hominidfossil so far discovered in the SilberbergGrotto? Why were there virtually noother animal bones with it when it wasfirst uncovered?
Because of the methodical and slowexcavation, we can now provide possibleand probable answers to these questions.The completeness of the individual andlack of any carnivore damage to the bonesindicate that it was not preyed upon bycarnivores but that it fell into a shaftleading into the cavern. The reasons forthe fall are unknown. We do know that atthe other (eastern) end of the SilberbergGrotto is a concentration of bones, mainlyof carnivores and monkeys, from animalsthat fell in to a natural death trap.4 Theprobability is that StW 573 also fell into a
Research in Action South African Journal of Science 104, November/December 2008 443
*Institute for Human Evolution & School of AnatomicalSciences, University of the Witwatersrand, Private Bag 3,WITS 2050, South Africa.E-mail: [email protected] Fig. 1. The skull and left humerus of StW 573.

shaft, below which was a talus slope thatfed into the western end of the SilberbergGrotto. Although there are some articu-lated parts of primate, bovid and carni-vore at the eastern end, there are nocomplete or near-complete skeletons.This is probably due to the individualbones and partially articulated elementshaving been transported by water fromtheir original point of deposition. Thereare certainly signs that the bones wereconcentrated and loosely stratified bywater.3
Situation of the Australopithecusskeleton
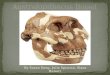
When the StW 573 skeleton was firstbeing uncovered, there was a noticeablenear-absence of other fossils in its vicinity.Only a monkey left radius and a leopardhalf-mandible were present. As we havecontinued to dig deeper in order toundercut the skeleton for lifting, we haveuncovered an increasing number ofanimal remains, including two monkeycrania and a monkey ulna. It is notewor-thy that these bones are isolated finds,and there is no sign of any articulatedskeleton of an animal. Why then shouldthe Australopithecus be so complete? Theanswer seems to be in its position on thetalus slope. It is on top of the talus slope,not far below the roof, and was appar-ently one of the last bodies to fall onto thatslope before the point of ingress wasblocked and the talus was sealed over by athick flowstone (Fig. 5). Such pure flow-stone can develop only when there is nofurther entry of debris. Thus there was noopportunity for the skeleton to be brokenup and scattered by such further entry ofrocks and sediment.
The breakage that has occurred to theskeleton has been occasioned by waterflowing through the talus beneath it,removing a softer reddish deposit, andforming a cavity into which the centralpart of the skeleton and its surroundingmatrix collapsed.5 The extensive cavity isstill clearly visible where the lime minershave sectioned it by blasting away thebreccia downslope of the skeleton. Fur-thermore, our excavation to undercutthe skeleton has revealed some smallercavities lined with calcium carbonatewithin the talus deposit and suggestive ofwater flow through the talus. It is thisancient water flow that was probablyresponsible for the disintegration of theleft side of the pelvis. And it is the collapseof the central part of the skeleton andslight movements of the surroundingmatrix that caused some of the bones tobreak.
The question of why the StW 573 skeletonis the only hominid so far found in the
Silberberg Grotto remains a mystery. Thecavern has been extensively mined andall the quarried breccia, in addition tothe breccia that we excavated, has beenprocessed. Although it is full of animalbones, not a single tooth of a hominid has
ever been recovered, apart from those inthe skull of StW 573. This near-absence ofhominid fossils contrasts greatly with therichness of Australopithecus remains inMember 4. The answer to this puzzlecould be either that early in the history of
444 South African Journal of Science 104, November/December 2008 Research in Action
Fig. 2. The left forearm and hand of StW 573.
Fig. 3. The left hand of StW 573. Fingers clenched and thumb across palm.
Fig. 4. The right scapula of StW 573 (centre right) in ventral view. Back of skull at top left.
the cave infilling at Sterkfontein Australo-pithecus was not common in the vicinity ofthe caves, or that unlike the monkeys andcarnivores, it was not in the habit of fallingdown the shafts. By Member 4 times,perhaps 700 000 years later, there aredefinite signs that Australopithecus wasbeing preyed upon and its bones andteeth were being washed into a largercave opening.6,36
To which species does StW 573belong?
When the Australopithecus skull andskeleton was first announced in 1998, itwas not possible to determine the speciesto which it belongs. It could be stated onlythat it is different from A. africanus. As theuncovering of the bones progressed, itappeared that the skull had some similari-ties to A. afarensis in its muscular occiput,sagittal crest, and robust zygomatic arch.There were, however, also some differ-ences. It is now apparent that StW 573does not belong to either A. africanus orA. afarensis, but that it represents an indi-vidual either belonging to, or very closelyaffiliated to, the second Australopithecusspecies that is well represented in Sterk-fontein Member 4 and Makapansgat.7,8
The continued removal of breccia fromthe cranium has revealed that the lowerpart of the StW 573 face has been pushedupward and backward, which gave it afalse appearance of having a relativelyshort, squat face. If allowance is made forthis, then the combination of longer face,robust zygomatic arch, muscular occiput,small sagittal crest at the back of thecranium, and lack of supraorbital thicken-ing distinguishes this skull from Austra-lopithecus africanus and aligns it closelywith specimens such as StW 505 andStW 252, which belong to the secondAustralopithecus species as discussed below.
Australopithecus africanusThe type specimen of this species is the
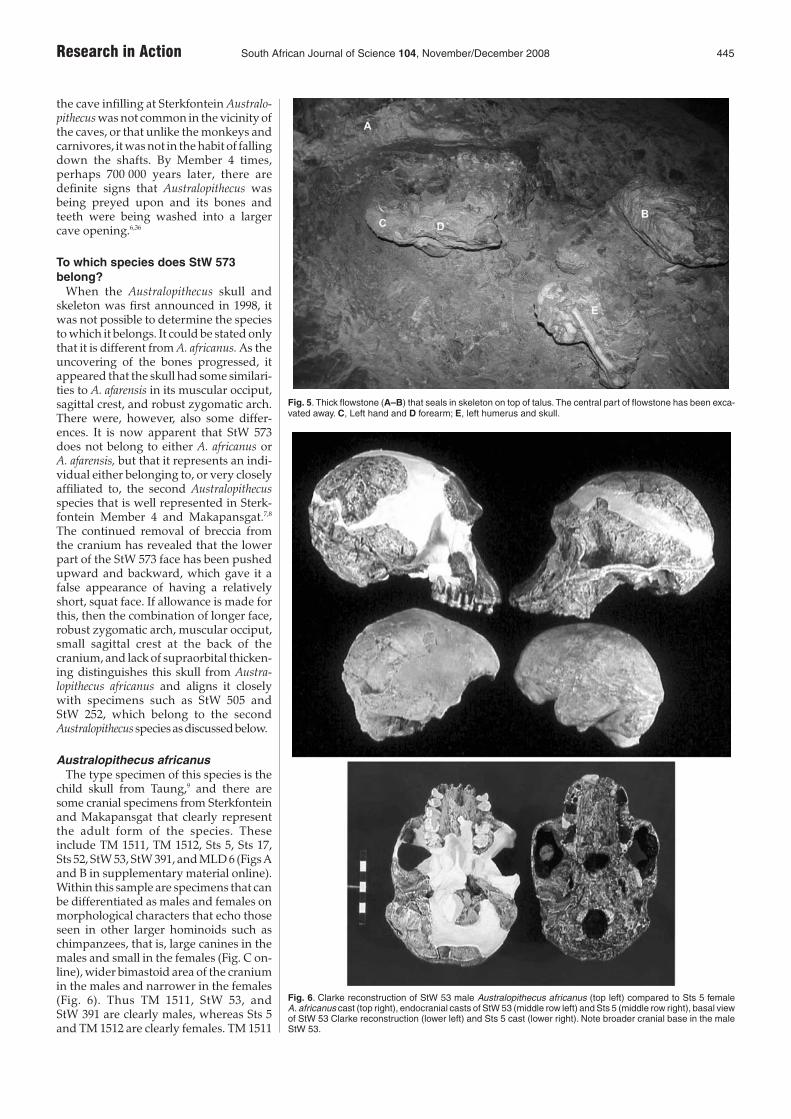
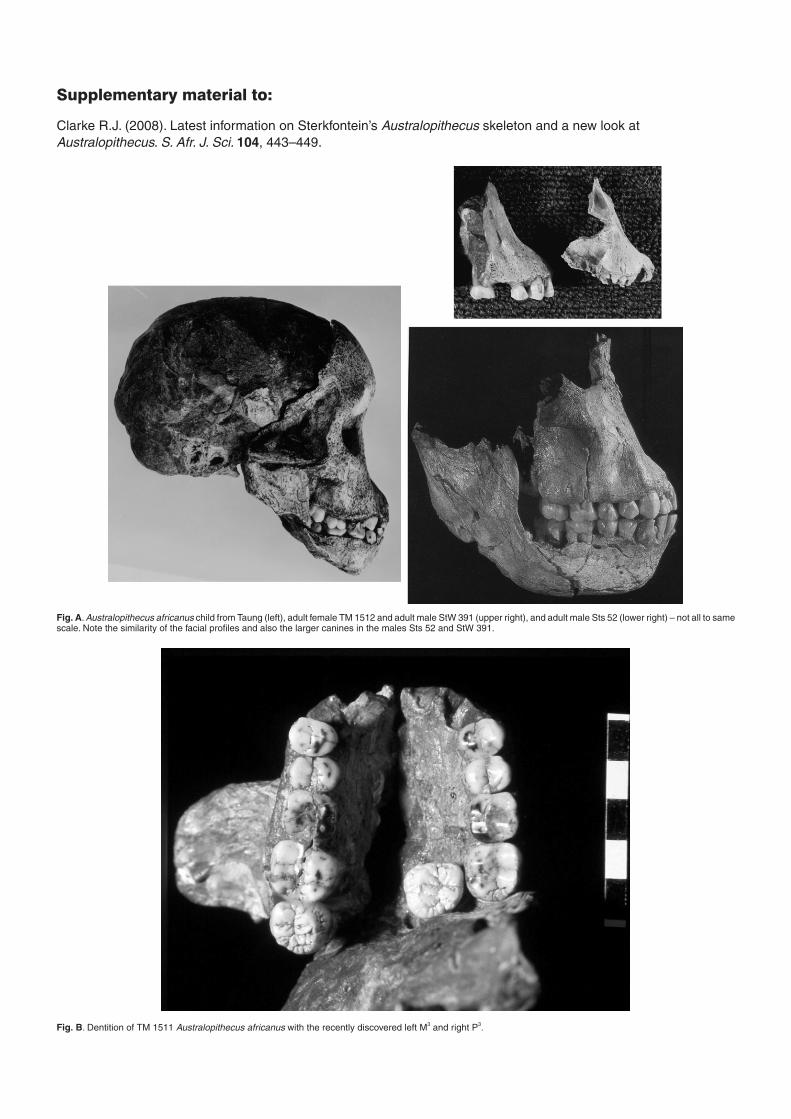
child skull from Taung,9 and there aresome cranial specimens from Sterkfonteinand Makapansgat that clearly representthe adult form of the species. Theseinclude TM 1511, TM 1512, Sts 5, Sts 17,Sts 52, StW 53, StW 391, and MLD 6 (Figs Aand B in supplementary material online).Within this sample are specimens that canbe differentiated as males and females onmorphological characters that echo thoseseen in other larger hominoids such aschimpanzees, that is, large canines in themales and small in the females (Fig. C on-line), wider bimastoid area of the craniumin the males and narrower in the females(Fig. 6). Thus TM 1511, StW 53, andStW 391 are clearly males, whereas Sts 5and TM 1512 are clearly females. TM 1511
Research in Action South African Journal of Science 104, November/December 2008 445
Fig. 5. Thick flowstone (A–B) that seals in skeleton on top of talus. The central part of flowstone has been exca-vated away. C, Left hand and D forearm; E, left humerus and skull.
Fig. 6. Clarke reconstruction of StW 53 male Australopithecus africanus (top left) compared to Sts 5 femaleA. africanus cast (top right), endocranial casts of StW 53 (middle row left) and Sts 5 (middle row right), basal viewof StW 53 Clarke reconstruction (lower left) and Sts 5 cast (lower right). Note broader cranial base in the maleStW 53.

and StW 53 closely resemble each otherand both have large canine sockets com-pared to Sts 5 and TM 1512. Furthermore,StW 53 is wider across the mastoid regionthan is Sts 5 (Fig. 9). Of these specimens,StW 53 needs to be discussed in detail,because it is commonly but incorrectlyreferred to in text books and researchpublications as Homo habilis.
In 1976, at the southwestern end of hisSterkfontein excavation, Alun Hughesrecovered hominid teeth and cranialfragments of one individual (StW 53)from the decalcified breccia within asolution pocket. The right posteriorportion of the braincase was embedded insolid breccia in the wall of the solutionpocket, thus indicating the exact locationand breccia type from which the otherfragments had been decalcified andscattered. The breccia at that western endwas, at the time, thought to be allMember 5 with an overlying exposure ofMember 6 in a small area on the northernside. As Member 5 had yielded abundantearly stone tools, and as no stone toolswhatsoever had occurred in Member 4,10
believed that StW 53 must be a cranium ofearly Homo. Subsequently, it was frequentlyreferred to as Homo habilis because of itssimilarity to the OH 24 Homo habilis fromOlduvai Gorge Bed 1.11,12 Later stratigraphicinvestigation by Clarke13 showed thatStW 53 did not in fact come fromtool-bearing Member 5, but from a hang-ing remnant of Member 4.14 Furthermore,at a conference in Orce, Spain, in 1995,15
Clarke observed that StW 53 and supposedHomo habilis specimens OH 13, OH 24,and KNM ER 1813, had small brains andflat noses like those of Australopithecus,and in these respects differed greatlyfrom the much larger-brained OH 7 typespecimen of Homo habilis, as well as fromKNM ER 1470 Homo habilis, which hasa prominent nasal skeleton. He alsoreferred to the work of Spoor16 and Spooret al.,17 who found that the bony labyrinthof the StW 53 temporal was uniqueamong hominid fossils in dimensions ofsemicircular canals, which were betweenthose of great apes and australopithecinesand those of cercopithecoids. Nothing inits anatomy suggested that StW 53 wasanything but an Australopithecus and itcertainly came from the non-tool-bearingMember 4, just like the other Australo-pithecus fossils from Sterkfontein.14
Clarke12 made a reconstruction of theStW 53 cranium ‘based solely on anatomi-cal considerations and symmetry’. Hetermed it a ‘preliminary’ reconstructionbecause there were several other isolatedfragments of vault and orbit, some ofwhich might possibly be incorporated inany future reconstruction. Furthermore,
there was no contact between the facialportion and the frontal and temporals.Thus, very minor adjustments might beconsidered in the future to the angulationand vertical position of the face relative tothe braincase. This did not mean thatClarke did not have full confidence in theform and size of the braincase as he hadrestored it. There were some good contactsand some near contacts between fragmentsof the posterior vault and this, plus thesmall size of the undistorted frontal(smaller than Sts 5), provided a soundbasis for the reconstruction.
Curnoe and Tobias18 stated that theycompared ‘the cranium, as initially assem-bled by Hughes and Clarke, with the laterreconstruction by Clarke (1985)12’ and that‘the anatomical placement of a number ofcritical bones differed between the two.’They then went on to state, ‘Thus, twodifferent interpretations of the morphol-ogy of StW 53 have existed until now (i.e.studies of original remains, as assembledby Alun Hughes, and alterations to thisrestoration made on cast pieces by Clarkeduring his reconstruction).’ This is untrue.There was no earlier restoration of thecranium, either by Hughes and Clarke orby Hughes alone. In the initial publicationby Hughes and Tobias,10 the separateportions of the cranium are not assem-bled. In an earlier announcement,19 thereis a photograph of the separate face andfrontal, left parieto-occipital fragmentand left occipital condyle placed in profileon a photographic stand in the darkroomby Hughes and Clarke to indicate, as thecaption says, ‘some of the discoveredfragments reassembled in an attempt togive an idea of the outline of the cranium.’The same photograph is published byTobias20 with the caption stating that it is‘a tentative alignment of some parts of thespecimen, effected by A.R. Hughes andR.J. Clarke.’ The plasticene supportingthe parieto-occipital fragment can be seenin this photograph.
There never was a three-dimensionalrestoration of the cranium prior to theClarke reconstruction of 1985, and therewere no differences between the 1976photograph and the Clarke 1985 recon-struction in the anatomical placement ofthose few fragments. Nevertheless, Curnoeand Tobias used these non-existent differ-ences to justify their making a ‘new recon-struction’. To achieve this, they dissembledthe anatomically sound near-contact be-tween the left and right occipital portionsand separated them by about 8 mm. Now,in their ‘reconstruction’ the nuchal line isabout 8 mm longer on the right of theinion than it is on the left (see theirfigures 8c and 14). They have thus artifi-cially and unjustifiably widened the
braincase and made it asymmetrical.They also internally opened the suturalcontact between the left posterior parietalfragment and occipital and flexed theparietal upwards such that the parietalfragment, in their words, ‘is now moreelevated at its anterior end’. This hasmade the vault higher and they haveelongated the vault greatly by extendingthe gap abnormally between the posteriorbraincase and the fronto-facial portion toproduce an oddly elongated but relativelynarrow braincase. The result of these andother modifications is that the maxilla isgreatly twisted relative to the mid sagittalplane (their figure 9). They comment onthis as follows: ‘thus, the whole facialskeleton of StW 53 is twisted to the right ofthe median sagittal plane. We have notcorrected for this in our reconstruction, asClarke (1985) did in his, preferring insteadto recognize more fully the extent ofdamage the specimen has suffered.’ Thisis untrue. Clarke12 did not mention thecorrection of any distortion and had noneed to, because there was no distortionof the cranium to correct!
The maxilla, frontal bone, left parieto-occipital portions, left temporal, and leftoccipital condyle, as well as several otherisolated fragments of braincase and aright ascending ramus of the mandible,were recovered from the decalcifiedbreccia and none of these pieces wasdeformed in any way. The right posteriorportion of the braincase was excavatedfrom the hard breccia wall of the solutionpocket where it was exposed in the formof the manganese-stained broken sectionthrough the right parietal, occipital andtemporal. A photograph of this is pub-lished in Tobias,20 as well as a drawingreconstructing the postulated sequenceof events leading up to breakage of theskull. The manganese staining indicatedthe decalcified edge of the right braincase.Some of the loose fragments from thedecalcified breccia would originally havebeen in contact with this manganese-stained exposed margin. When thisbraincase portion was removed andcleaned of breccia, it could be seen thatthe parietal had been pushed downwardsslightly inside the temporal and occipital,and there was some fracturing and slightdisplacement of parts of the parietal andoccipital near asterion. There was, how-ever, no serious distortion of the individ-ual bone fragments comprising thespecimen. Once the bones were separatedand put in their correct position byClarke,12 this portion of the braincaseexhibited good landmarks and surfacecontours that, together with the otherisolated fragments, provided an excellentbasis for the reconstruction of an undis-
446 South African Journal of Science 104, November/December 2008 Research in Action

torted cranium. Curnoe and Tobias,18
however, stated that there is ‘deforma-tion of, and damage to, the right parieto-occipito-temporal fragment’ and said that‘transverse curvature of the vault wasmore difficult to reconstruct because ofextensive distortion and damage, especiallyon the right parietal and temporal bonesand on the right part of the occipital.’
The Clarke12 reconstruction of this region,mirror imaging the available data providedby the left parieto-occipital and temporaland occipital condyle fragments, was notdeformed. Curnoe and Tobias, however,cut up the cast of the right braincaseportion as reconstructed by Clarke (theirfigure 10) and bent contacts betweenfragments in an unnatural manner. Theyalso separated the good contact betweentwo parts of the right occipital base androtated the upper portion clockwise rela-tive to the lower portion, leaving a gap of9 mm at the medial end. All of this hasproduced a deformity that they havemirror imaged in their reconstruction ofthe left side, producing a bizarrely shapedprofile when viewed from the rear thatdoes not resemble any known hominidcranium (see their figures 14 and 8c).
Reconstruction, in the fossil context,means building up to its original formsomething that is fragmented. This isdone by utilizing the preserved anatomicallandmarks and contours, and this is howClarke12 made the reconstructed StW 53cranium. Curnoe and Tobias18 have notdone this, but instead have widely sepa-rated a near-contact, unrealistically flexedsuture and bone contacts and unnaturallyextended the gap between front and backto produce a braincase that is wider,higher and longer than it should be.Furthermore, it is now deformed. Thustheirs is not a valid reconstruction becauseit has not followed the anatomical guide-lines and contours and therefore does notreflect the original form of the cranium. Itis a construct of how they think it shouldbe, rather than a reconstruction of how itactually was. In their own words, Curnoeand Tobias18 admit that ‘our reconstructiondiffers in important respects from theearlier one, especially in terms of neuro-cranial length, breadth, and height. How-ever, given that StW 53 exhibits extensivedamage, these dimensions are most likelyprone to much error in reconstruction.’Despite this, they have still maintainedthat the maximal cranial length of their‘reconstruction’ is virtually identical tothe Homo habilis cranium KNM ER 1470(their figure 18) and their overall conclusionbased on their artificially enlarged anddeformed StW 53 braincase, as well astheir interpretation of the anatomy, is thatit is a representative of Homo habilis.
The Clarke12 reconstruction, which wasbased on anatomy and contours of thebones (which contra Curnoe and Tobiasdo not ‘exhibit extensive damage’), showsthat the StW 53 cranial size and shapedoes not differ significantly from Austra-lopithecus africanus in the form of Sts 5(Fig. 6). The facial structure of the two isvery similar with small, narrow muzzle,small nasal aperture and flat nasal skeleton,and both have a narrow rectangularpalate. The frontal bones of both resembleeach other in size and form with a promi-nent metopic ridge. The wider inter-mastoid breadth in StW 53 is undoubtedlybecause it is a male A. africanus, whereasSts 5 is a female. This is indicated also inthe dentition. Sts 5 has smaller canine andcheek teeth sockets than does StW 53,which more resembles Sts 52 and StW 391.In its small canine and premolar sockets,Sts 5 is similar to TM 1512, which is alsoundoubtedly a female. The larger caninesocket in the apparent males has causedthe socket to bulge more anteriorly, whichcan contribute to a less pronouncedappearance of the premaxillary region. Itshould also be noted that StW 53 has thepremaxillary bone visible on both sides ofthe upper nasal margin. This is an ape-likeand Australopithecus character seen inTaung and MLD 6 and does not occur inHomo. Hughes and Tobias10 noted thecoincidence of StW 53 fragments beingfound on the days representing the 40thanniversary of Broom’s first visit toSterkfontein on 9 August 1936, and hisdiscovery of the first adult cranium on 17August 1936. The coincidence was actuallymore striking than they realized, becauseStW 53 is remarkably similar to Broom’sfirst adult cranium, TM 1511. The twocrania resemble each other in the form oftheir face and teeth, even though theteeth of StW 53 are heavily worn, and thefrontal bone of StW 53 fits extremely wellwith the size and curvature of the frontallobe of the endocast (Sts 60) of TM 1511.
There is in fact nothing in the anatomyof StW 53 to align it with Homo habilis (inthe form of OH 7, KNM ER 1470 andOH 65) rather than with Australopithecus.The type specimen of Homo habilis isOlduvai Hominid 7 (OH 7), consisting ofthe two parietals and mandible of a juve-nile.21 The parietals alone indicate a muchlarger-brained hominid than any Austra-lopithecus and these parietals match in sizeand shape those of the near-completecranium KNM ER 1470.22 Hence, the 1470cranium cannot be distinguished fromHomo habilis and there is no justificationfor its having been placed in a separatespecies, H. rudolfensis (e.g. refs 23, 24). Thediscovery of a complete dentition inmaxilla of OH 65 from Olduvai Gorge
Bed 1 (ref. 25) supports the placement ofKNM ER 1470 in Homo habilis. The maxillaof OH 65 matches in shape and size that of1470 and the cheek tooth crowns, thoughsmall by comparison with Australopithecus,have widely-flaring roots like those ofKNM ER 1470. The palates of both fossilsare broad and horseshoe-shaped, thusdiffering radically from the narrow, rect-angular palates of Australopithecus.
It is rather the smaller-brained, moreAustralopithecus-like fossils OH 13, KNMER 1813, OH 24 and OH 62 which shouldbe removed from the taxon Homo habilis asthey have no similarity to Homo habilis asrepresented by the type specimen OH 7,together with KNM ER 1470 and OH 65.The question of whether the smaller-brained forms should be classed asAustralopithecus rather than Homo hasbeen raised before.26,27 As far as the small-brained StW 53 is concerned, there seemsno doubt that it should be classed asA. africanus, apparently male, with strongsimilarities to Broom’s original 1936cranium TM 1511, as well as to Sts 5.
A second Australopithecus speciesin South Africa
Ever since the discovery by Broom ofthe first adult Australopithecus fossil atSterkfontein caves in 1936, all otherAustralopithecus fossils from that site havegenerally been regarded as belonging toone species. Initially, Broom28 named thefirst cranium, TM 1511, as Australopithecustransvaalensis, a species distinct from thechild of Taung that had been namedAustralopithecus africanus.9 Two years later,following the discovery at Sterkfontein ofa child mandibular symphysis with a verylarge canine, Broom29 decided that it wasso distinct from the Taung child that heshould make it a new genus, Plesianthropus.He considered that the larger-toothed fos-sils from Sterkfontein were males and thesmaller-toothed fossils were females ofone species.
Dart30 named the first Australopithecusfossil from Makapansgat (the occipitalMLD 1) as a new species, Australopithecusprometheus, and all subsequent fossilsfrom that site were placed in the samespecies. Thus, although each of the threesites, Taung, Sterkfontein and Makapans-gat, were considered to have differentspecies of ape-man, it did not occur toeither Broom or Dart that they may havemore than one species represented withinthe sites of Sterkfontein and Makapansgat.
Later it became general practice toregard all the Australopithecus fossils fromthe three sites as belonging to the onespecies, A. africanus, and that has been theprevailing view up to the present day. Adissenting view has been expressed by
Research in Action South African Journal of Science 104, November/December 2008 447
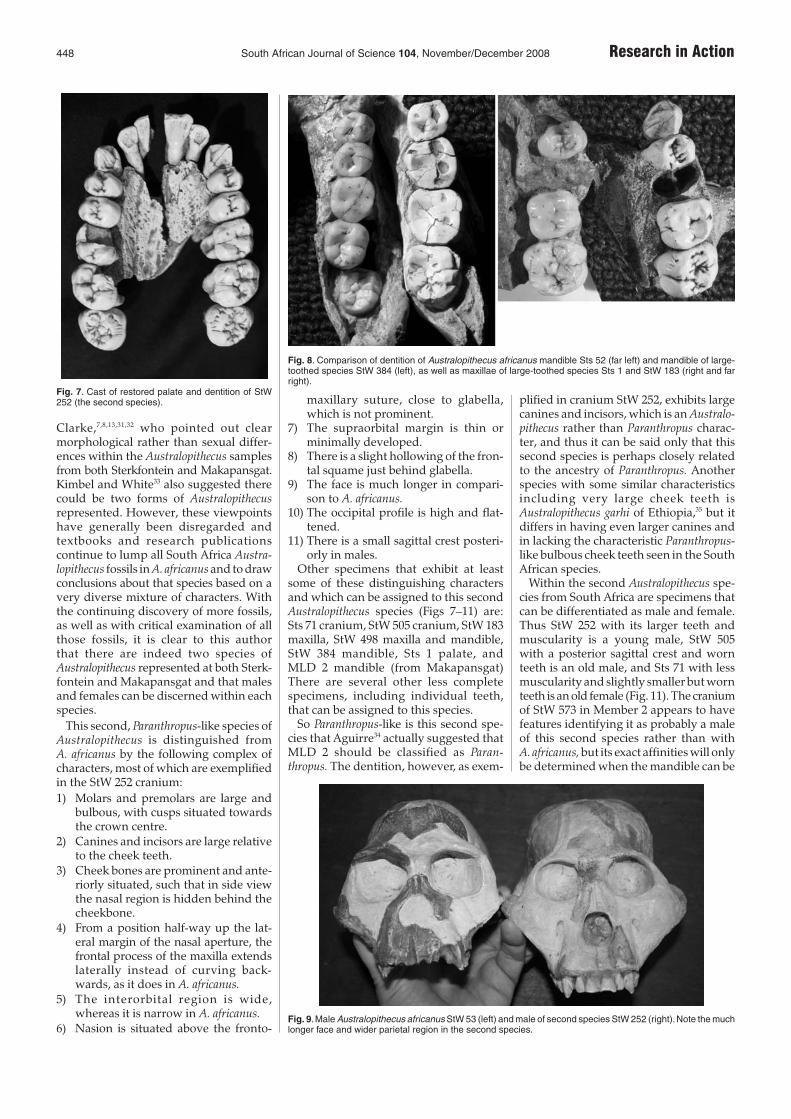
Clarke,7,8,13,31,32 who pointed out clearmorphological rather than sexual differ-ences within the Australopithecus samplesfrom both Sterkfontein and Makapansgat.Kimbel and White33 also suggested therecould be two forms of Australopithecusrepresented. However, these viewpointshave generally been disregarded andtextbooks and research publicationscontinue to lump all South Africa Austra-lopithecus fossils in A. africanus and to drawconclusions about that species based on avery diverse mixture of characters. Withthe continuing discovery of more fossils,as well as with critical examination of allthose fossils, it is clear to this authorthat there are indeed two species ofAustralopithecus represented at both Sterk-fontein and Makapansgat and that malesand females can be discerned within eachspecies.
This second, Paranthropus-like species ofAustralopithecus is distinguished fromA. africanus by the following complex ofcharacters, most of which are exemplifiedin the StW 252 cranium:1) Molars and premolars are large and
bulbous, with cusps situated towardsthe crown centre.
2) Canines and incisors are large relativeto the cheek teeth.
3) Cheek bones are prominent and ante-riorly situated, such that in side viewthe nasal region is hidden behind thecheekbone.
4) From a position half-way up the lat-eral margin of the nasal aperture, thefrontal process of the maxilla extendslaterally instead of curving back-wards, as it does in A. africanus.
5) The interorbital region is wide,whereas it is narrow in A. africanus.
6) Nasion is situated above the fronto-
maxillary suture, close to glabella,which is not prominent.
7) The supraorbital margin is thin orminimally developed.
8) There is a slight hollowing of the fron-tal squame just behind glabella.
9) The face is much longer in compari-son to A. africanus.
10) The occipital profile is high and flat-tened.
11) There is a small sagittal crest posteri-orly in males.
Other specimens that exhibit at leastsome of these distinguishing charactersand which can be assigned to this secondAustralopithecus species (Figs 7–11) are:Sts 71 cranium, StW 505 cranium, StW 183maxilla, StW 498 maxilla and mandible,StW 384 mandible, Sts 1 palate, andMLD 2 mandible (from Makapansgat)There are several other less completespecimens, including individual teeth,that can be assigned to this species.
So Paranthropus-like is this second spe-cies that Aguirre34 actually suggested thatMLD 2 should be classified as Paran-thropus. The dentition, however, as exem-
plified in cranium StW 252, exhibits largecanines and incisors, which is an Australo-pithecus rather than Paranthropus charac-ter, and thus it can be said only that thissecond species is perhaps closely relatedto the ancestry of Paranthropus. Anotherspecies with some similar characteristicsincluding very large cheek teeth isAustralopithecus garhi of Ethiopia,35 but itdiffers in having even larger canines andin lacking the characteristic Paranthropus-like bulbous cheek teeth seen in the SouthAfrican species.
Within the second Australopithecus spe-cies from South Africa are specimens thatcan be differentiated as male and female.Thus StW 252 with its larger teeth andmuscularity is a young male, StW 505with a posterior sagittal crest and wornteeth is an old male, and Sts 71 with lessmuscularity and slightly smaller but wornteeth is an old female (Fig. 11). The craniumof StW 573 in Member 2 appears to havefeatures identifying it as probably a maleof this second species rather than withA. africanus, but its exact affinities will onlybe determined when the mandible can be
448 South African Journal of Science 104, November/December 2008 Research in Action
Fig. 8. Comparison of dentition of Australopithecus africanus mandible Sts 52 (far left) and mandible of large-toothed species StW 384 (left), as well as maxillae of large-toothed species Sts 1 and StW 183 (right and farright).
Fig. 7. Cast of restored palate and dentition of StW252 (the second species).
Fig. 9.Male Australopithecus africanus StW 53 (left) and male of second species StW 252 (right).Note the muchlonger face and wider parietal region in the second species.
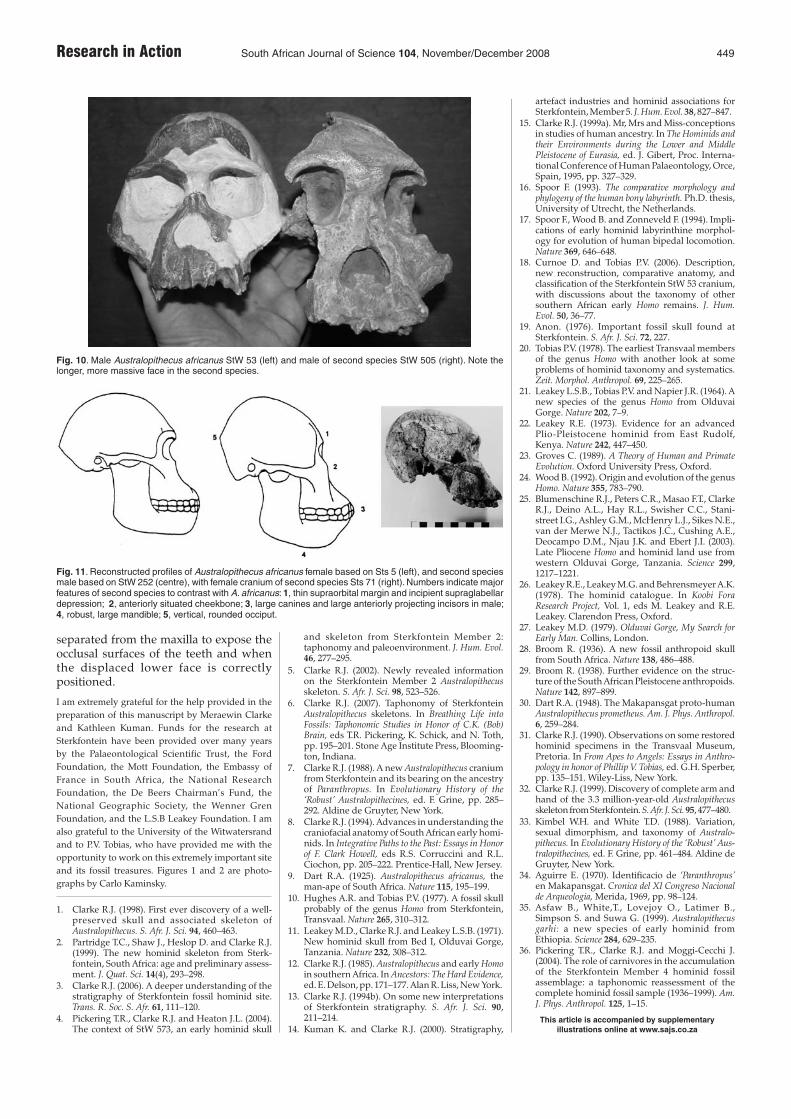
separated from the maxilla to expose theocclusal surfaces of the teeth and whenthe displaced lower face is correctlypositioned.
I am extremely grateful for the help provided in thepreparation of this manuscript by Meraewin Clarkeand Kathleen Kuman. Funds for the research atSterkfontein have been provided over many yearsby the Palaeontological Scientific Trust, the FordFoundation, the Mott Foundation, the Embassy ofFrance in South Africa, the National ResearchFoundation, the De Beers Chairman’s Fund, theNational Geographic Society, the Wenner GrenFoundation, and the L.S.B Leakey Foundation. I amalso grateful to the University of the Witwatersrandand to P.V. Tobias, who have provided me with theopportunity to work on this extremely important siteand its fossil treasures. Figures 1 and 2 are photo-graphs by Carlo Kaminsky.
1. Clarke R.J. (1998). First ever discovery of a well-preserved skull and associated skeleton ofAustralopithecus. S. Afr. J. Sci. 94, 460–463.
2. Partridge T.C., Shaw J., Heslop D. and Clarke R.J.(1999). The new hominid skeleton from Sterk-fontein, South Africa: age and preliminary assess-ment. J. Quat. Sci. 14(4), 293–298.
3. Clarke R.J. (2006). A deeper understanding of thestratigraphy of Sterkfontein fossil hominid site.Trans. R. Soc. S. Afr. 61, 111–120.
4. Pickering T.R., Clarke R.J. and Heaton J.L. (2004).The context of StW 573, an early hominid skull
and skeleton from Sterkfontein Member 2:taphonomy and paleoenvironment. J. Hum. Evol.46, 277–295.
5. Clarke R.J. (2002). Newly revealed informationon the Sterkfontein Member 2 Australopithecusskeleton. S. Afr. J. Sci. 98, 523–526.
6. Clarke R.J. (2007). Taphonomy of SterkfonteinAustralopithecus skeletons. In Breathing Life intoFossils: Taphonomic Studies in Honor of C.K. (Bob)Brain, eds T.R. Pickering, K. Schick, and N. Toth,pp. 195–201. Stone Age Institute Press, Blooming-ton, Indiana.
7. Clarke R.J. (1988). A new Australopithecus craniumfrom Sterkfontein and its bearing on the ancestryof Paranthropus. In Evolutionary History of the‘Robust’ Australopithecines, ed. F. Grine, pp. 285–292. Aldine de Gruyter, New York.
8. Clarke R.J. (1994). Advances in understanding thecraniofacial anatomy of South African early homi-nids. In Integrative Paths to the Past: Essays in Honorof F. Clark Howell, eds R.S. Corruccini and R.L.Ciochon, pp. 205–222. Prentice-Hall, New Jersey.
9. Dart R.A. (1925). Australopithecus africanus, theman-ape of South Africa. Nature 115, 195–199.
10. Hughes A.R. and Tobias P.V. (1977). A fossil skullprobably of the genus Homo from Sterkfontein,Transvaal. Nature 265, 310–312.
11. Leakey M.D., Clarke R.J. and Leakey L.S.B. (1971).New hominid skull from Bed I, Olduvai Gorge,Tanzania. Nature 232, 308–312.
12. Clarke R.J. (1985). Australopithecus and early Homoin southern Africa. In Ancestors: The Hard Evidence,ed. E. Delson, pp. 171–177. Alan R. Liss, New York.
13. Clarke R.J. (1994b). On some new interpretationsof Sterkfontein stratigraphy. S. Afr. J. Sci. 90,211–214.
14. Kuman K. and Clarke R.J. (2000). Stratigraphy,
artefact industries and hominid associations forSterkfontein, Member 5. J. Hum. Evol. 38, 827–847.
15. Clarke R.J. (1999a). Mr, Mrs and Miss-conceptionsin studies of human ancestry. In The Hominids andtheir Environments during the Lower and MiddlePleistocene of Eurasia, ed. J. Gibert, Proc. Interna-tional Conference of Human Palaeontology, Orce,Spain, 1995, pp. 327–329.
16. Spoor F. (1993). The comparative morphology andphylogeny of the human bony labyrinth. Ph.D. thesis,University of Utrecht, the Netherlands.
17. Spoor F., Wood B. and Zonneveld F. (1994). Impli-cations of early hominid labyrinthine morphol-ogy for evolution of human bipedal locomotion.Nature 369, 646–648.
18. Curnoe D. and Tobias P.V. (2006). Description,new reconstruction, comparative anatomy, andclassification of the Sterkfontein StW 53 cranium,with discussions about the taxonomy of othersouthern African early Homo remains. J. Hum.Evol. 50, 36–77.
19. Anon. (1976). Important fossil skull found atSterkfontein. S. Afr. J. Sci. 72, 227.
20. Tobias P.V. (1978). The earliest Transvaal membersof the genus Homo with another look at someproblems of hominid taxonomy and systematics.Zeit. Morphol. Anthropol. 69, 225–265.
21. Leakey L.S.B., Tobias P.V. and Napier J.R. (1964). Anew species of the genus Homo from OlduvaiGorge. Nature 202, 7–9.
22. Leakey R.E. (1973). Evidence for an advancedPlio-Pleistocene hominid from East Rudolf,Kenya. Nature 242, 447–450.
23. Groves C. (1989). A Theory of Human and PrimateEvolution. Oxford University Press, Oxford.
24. Wood B. (1992). Origin and evolution of the genusHomo. Nature 355, 783–790.
25. Blumenschine R.J., Peters C.R., Masao F.T., ClarkeR.J., Deino A.L., Hay R.L., Swisher C.C., Stani-street I.G., Ashley G.M., McHenry L.J., Sikes N.E.,van der Merwe N.J., Tactikos J.C., Cushing A.E.,Deocampo D.M., Njau J.K. and Ebert J.I. (2003).Late Pliocene Homo and hominid land use fromwestern Olduvai Gorge, Tanzania. Science 299,1217–1221.
26. Leakey R.E., Leakey M.G. and Behrensmeyer A.K.(1978). The hominid catalogue. In Koobi ForaResearch Project, Vol. 1, eds M. Leakey and R.E.Leakey. Clarendon Press, Oxford.
27. Leakey M.D. (1979). Olduvai Gorge, My Search forEarly Man. Collins, London.
28. Broom R. (1936). A new fossil anthropoid skullfrom South Africa. Nature 138, 486–488.
29. Broom R. (1938). Further evidence on the struc-ture of the South African Pleistocene anthropoids.Nature 142, 897–899.
30. Dart R.A. (1948). The Makapansgat proto-humanAustralopithecus prometheus. Am. J. Phys. Anthropol.6, 259–284.
31. Clarke R.J. (1990). Observations on some restoredhominid specimens in the Transvaal Museum,Pretoria. In From Apes to Angels: Essays in Anthro-pology in honor of Phillip V. Tobias, ed. G.H. Sperber,pp. 135–151. Wiley-Liss, New York.
32. Clarke R.J. (1999). Discovery of complete arm andhand of the 3.3 million-year-old Australopithecusskeleton from Sterkfontein. S. Afr. J. Sci. 95, 477–480.
33. Kimbel W.H. and White T.D. (1988). Variation,sexual dimorphism, and taxonomy of Australo-pithecus. In Evolutionary History of the ‘Robust’ Aus-tralopithecines, ed. F. Grine, pp. 461–484. Aldine deGruyter, New York.
34. Aguirre E. (1970). Identificacio de ‘Paranthropus’en Makapansgat. Cronica del XI Congreso Nacionalde Arqueologia, Merida, 1969, pp. 98–124.
35. Asfaw B., White,T., Lovejoy O., Latimer B.,Simpson S. and Suwa G. (1999). Australopithecusgarhi: a new species of early hominid fromEthiopia. Science 284, 629–235.
36. Pickering T.R., Clarke R.J. and Moggi-Cecchi J.(2004). The role of carnivores in the accumulationof the Sterkfontein Member 4 hominid fossilassemblage: a taphonomic reassessment of thecomplete hominid fossil sample (1936–1999). Am.J. Phys. Anthropol. 125, 1–15.
This article is accompanied by supplementaryillustrations online at www.sajs.co.za
Research in Action South African Journal of Science 104, November/December 2008 449
Fig. 10. Male Australopithecus africanus StW 53 (left) and male of second species StW 505 (right). Note thelonger, more massive face in the second species.
Fig. 11. Reconstructed profiles of Australopithecus africanus female based on Sts 5 (left), and second speciesmale based on StW 252 (centre), with female cranium of second species Sts 71 (right). Numbers indicate majorfeatures of second species to contrast with A. africanus: 1, thin supraorbital margin and incipient supraglabellardepression; 2, anteriorly situated cheekbone; 3, large canines and large anteriorly projecting incisors in male;4, robust, large mandible; 5, vertical, rounded occiput.
Supplementary material to:
Clarke R.J. (2008). Latest information on Sterkfontein’s Australopithecus skeleton and a new look atAustralopithecus. S. Afr. J. Sci. 104, 443–449.
Fig. A. Australopithecus africanus child from Taung (left), adult female TM 1512 and adult male StW 391 (upper right), and adult male Sts 52 (lower right) – not all to samescale. Note the similarity of the facial profiles and also the larger canines in the males Sts 52 and StW 391.
Fig. B. Dentition of TM 1511 Australopithecus africanus with the recently discovered left M3 and right P3.

Fig. C. Palatal view of TM 1512 female Australopithecus africanus (left) and StW 391 male A. africanus (right). Note the larger canine socket and wider premolars in themale.



















![Australopithecus Sediba[1]](https://img.pdfslide.net/doc/110x75/577ccfad1a28ab9e78904834/australopithecus-sediba1.jpg)