Embed Size (px)
Citation preview

Leaf Growth in Dactylis glomerata following Defoliation
BY
J. L. DAVIDSON' AND F. L. MILTHORPE
Unwertity of Nottingham School of Agriculture, Sutton Bonmgton, Loughborovgh
A B S T R A C T
Defoliation to a height of 2-5 cm considerably reduced the increase in leaf areain young Dactylis glomerata (cocksfoot) plants compared with that of intact plants,the reduction in the rate of appearance of new leaves being relatively greaterthan the reduction in expansion of existing leaves. The growth of those expandingleaves which were cut during defoliation accounted for 94 per cent of the totalincrease in leaf area during the first four days after defoliation. In such a leaf,expanding cells were confined to a basal section which was well below theligule of the enclosing fully expanded leaf.
There was a positive relationship between rate of leaf expansion and totalsoluble carbohydrate content of the stubble when the carbohydrate content wasvaried by placing the plants in the dark, but not when it was varied by de-foliation and subsequent growth. These and other results suggest that theconcentration of soluble carbohydrate in the bases of expanding leaves was afactor controlling leaf expansion following defoliation, and that the concentra-tion in any one leaf depended on the photosynthetic contribution from itsexposed portion.
When the external nutrient supply was high, removal of the laminae of fullyexpanded leaves, which comprised about two-thirds of the total leaf area, did notreduce leaf expansion. When the nutrient status was low, these leaves were ofprimary importance, presumably because of their role as a source of labilenutrients.
I N T R O D U C T I O N
ONE of the more controversial topics in grassland agronomy is the ques-tion of the significance of soluble carbohydrate 'reserves' in regrowth
which follows defoliation. Field studies have shown that growth after de-foliation is generally associated with a decline in the soluble carbohydratecontent of both roots and stubble (e.g. Graber et al., 1927; Weinmann, 1948).These changes led to the belief that after defoliation carbohydrate reservesare converted to structural components of new and expanding cells and maycontrol the rate of regrowth. Weinmann (1952) argued from these conceptsthat the aim of pasture management must be to maintain an adequate levelof reserves in the desirable species of the sward.
The significance of the soluble carbohydrates in growth generally was ques-tioned when Archbold (1945) showed that accumulated sugars and fructosansin the stem of barley contributed little to subsequent grain development. Morerecently, the arguments have been reviewed by May (i960) who concluded
1 C.S.I.R.O. Division of Plant Industry, Deniliquin, N.S.W., Australia.
[Annals of Botany, N.S. Vol. 30, No. 118, 1966.]

174 Davidson and Milthorpe—Leaf Growth in Dactylis
that the role of accumulated soluble carbohydrates in regrowth of grass hasyet to be established.
Growth following defoliation depends first of all on the development of aphotosynthetic surface. It seems reasonable to expect that the factors mostimportant in determining rates of regrowth will be those upon which leafgrowth after defoliation is primarily dependent. The series of experimentsdescribed here was an attempt to examine the influence of some of the mostlikely factors to be concerned during the period immediately followingdefoliation.
DEVELOPMENT OF THE LEAF SURFACE
Measurements were made of the relative contributions of leaves cut by thedefoliation treatment and of younger leaves to the developing leaf surface.
Experimental Procedures
In this and subsequent experiments reported here, the cultural techniquesemployed to grow plants of S37 cocksfoot {Dactylis glomerata L.) were thesame as previously reported (Davidson and Milthorpe, 1965). Plants weregrown in sand culture with standard Long Ashton nutrient solution (Hewitt,1952) at 220 C with a 16-hour day of 2-90 cal visible radiation/cm2/hr. Whenthe plants were five weeks old all exposed laminae were removed and eachtiller was cut to a height of 2-5 cm. The areas of cut and uncut leaves weremeasured 2, 4, 8, and 16 days later, five plants being taken at each harvest.
Results
Growth measurements on uncut plants of the same age and size grownunder the same conditions were reported previously (Davidson and Mil-thorpe, 1965). Comparison with these show that defoliation to 2-5 cm causedmarked reductions in the rate of leaf appearance and leaf-area increase(Table 1).
TABLE I
Mean increases in leaf area and leaf number of intact and defoliatedplants under otherwise similar conditions
TreatmentIntact plantsDefoliated to a
a height of2-5 cm .
Onginalleaf area
(cm1)215
2 1 2(before
defoliation)
Increase inleaf area
(cm1)2 3 0
116
Increase inleaf number
35
8-6
Tuneinterval(days)
7
8
Increase in leaf number was reduced relatively more than increase in leafarea. Indeed, immediately following defoliation, leaf expansion was almostentirely dependent on the expansion of the older expanding leaves which
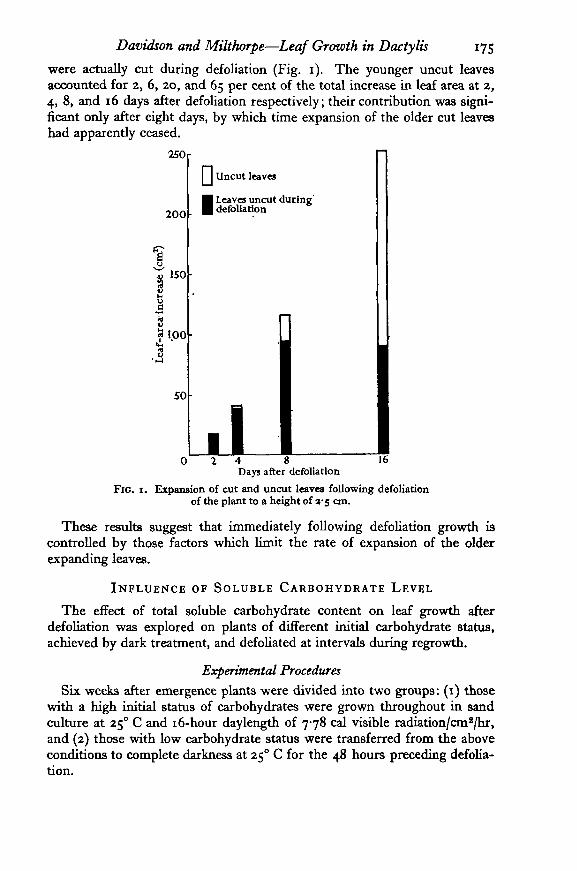
Davidson and Milthorpe—Leaf Growth in Dactylis 175
were actually cut during defoliation (Fig. 1). The younger uncut leavesaccounted for 2, 6, 20, and 65 per cent of the total increase in leaf area at 2,4, 8, and 16 days after defoliation respectively; their contribution was signi-ficant only after eight days, by which time expansion of the older cut leaveshad apparently ceased.
050
200
ISO
50
Uncut leavesD•
Leaves uncut during'defoliation
A4 8Days after defoliation
16
FIG. 1. Expansion of cut and uncut leaves following defoliationof the plant to a height of 3-5 an.
These results suggest that immediately following defoliation growth iscontrolled by those factors which limit the rate of expansion of the olderexpanding leaves.
INFLUENCE OF SOLUBLE CARBOHYDRATE LEVEL
The effect of total soluble carbohydrate content on leaf growth afterdefoliation was explored on plants of different initial carbohydrate status,achieved by dark treatment, and defoliated at intervals during regrowth.
Experimental Procedures
Six weeks after emergence plants were divided into two groups: (1) thosewith a high initial status of carbohydrates were grown throughout in sandculture at 250 C and 16-hour daylength of 778 cal visible radiation/cm2/hr,and (2) those with low carbohydrate status were transferred from the aboveconditions to complete darkness at 250 C for the 48 hours preceding defolia-tion.
176 Davidson and Milthorpe—Leaf Growth in Dactylis
The plants were defoliated by removing all exposed laminae and were thenreturned to the lighted growth room. Defoliation was repeated at two-dayintervals for eight days. One sub-group of six plants of initially high carbo-hydrate status was kept in complete darkness for these eight days. At eachdefoliation three replicates of the treatments other than that in continuousdarkness were preserved for carbohydrate determinations.
Results
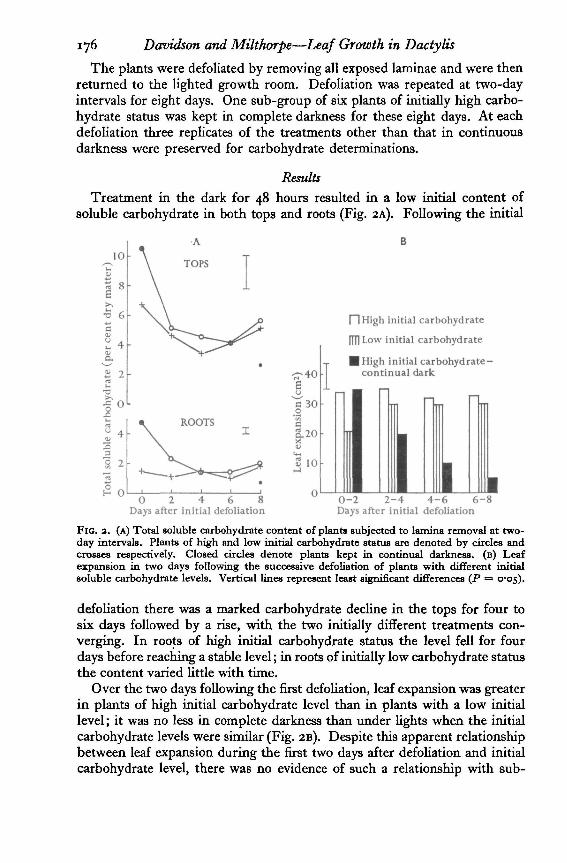
Treatment in the dark for 48 hours resulted in a low initial content ofsoluble carbohydrate in both tops and roots (Fig. 2A). Following the initial
10
J
\ 8
£ 4
£ 2
v 4
28 2
l o
ROOTSI 3 0
L20
s 10I—I
initial carbohydrate
Iffl Low initial carbohydrate
• High initial carbohydrate-continual dark
0 2 4 6 8Days after initial defoliation
0-2 2-4 4 -6 6-8Days after initial defoliation
FIG. 3. (A) Total soluble carbohydrate content of plants subjected to lamina removal at two-day intervals. Plants of high and low initial carbohydrate status are denoted by circles andcrosses respectively. Closed circles denote plants kept in continual darkness, (B) Leafexpansion in two days following the successive defoliation of plants with different initialsoluble carbohydrate levels. Vertical lines represent least significant differences (P =• 0-05).
defoliation there was a marked carbohydrate decline in the tops for four tosix days followed by a rise, with the two initially different treatments con-verging. In roots of high initial carbohydrate status the level fell for fourdays before reaching a stable level; in roots of initially low carbohydrate statusthe content varied little with time.
Over the two days following the first defoliation, leaf expansion was greaterin plants of high initial carbohydrate level than in plants with a low initiallevel; it was no less in complete darkness than under lights when the initialcarbohydrate levels were similar (Fig. 2B). Despite this apparent relationshipbetween leaf expansion during the first two days after defoliation and initialcarbohydrate level, there was no evidence of such a relationship with sub-

Davidson and Milthorpe—Leaf Growth in Dactylis 177
sequent defoliation. For instance, in plants initially of high carbohydratestatus, although the initial total soluble carbohydrate content in both tops androots was approximately double that at subsequent defoliations, leaf ex-pansion was scarcely altered. Also, over the two days following the secondand subsequent defoliations, leaf expansion in plants of low initial carbo-hydrate status was greater than it was after the first defoliation even thoughthe soluble carbohydrate content was then lower.
A reduction in soluble carbohydrate content brought about by growthfollowing defoliation did not lead to reduced leaf expansion; a similar reduc-tion resulting from dark treatment was associated with a significant reductionin leaf expansion following defoliation.
After eight days in complete darkness, when the rate of leaf expansion wasvery low compared with other treatments, the soluble carbohydrate contentof the tops was approximately half that of the other treatments. This supportsthe conclusion of Albert (1927) that growth in complete darkness stops whenconsiderable carbohydrate remains, and suggests that not all of the totalsoluble carbohydrate is available for leaf expansion or can be used in it.
T H E SITE OF LEAF EXPANSION
Experimental Procedures
Measurements to determine the location of expanding cells within laminaeand leaf sheaths were confined to leaves on the main tiller. After having beengrown for five weeks at 22° C and 3 -2 cal visible radiation/cm*/hr with a 16-hourdaylength, plants were selected for uniformity of length of main tiller.
1. The lamina. All visible laminae (sections above ligules) on the maintiller were marked with Indian ink at four or five equal intervals. The lengthsand median widths of each section were measured at the time of marking andagain after 24 hours.
2. The leaf base. When the young plants are defoliated by the removal ofall laminae, the remaining stubble consists of the sheaths of the older leaveswith ligules exposed (which will be referred to as 'outer' leaves) and all parts ofyounger leaves enclosed by these sheaths (referred to as 'inner' leaves). Thisstubble, with laminae still attached, was marked to denote four measured sec-tions by length—the basal 1 cm (Section d), and the remainder divided intothree equal parts (Sections c, b, and a, respectively, reading from the base).At the top of each section a fine needle was pushed horizontally through thecentre of the tiller. After 24 hours the length of each section was measuredon all marked leaves. Using the method of Wright (1961) cell numbers wereestimated in each section of expanding and fully expanded leaves.
Results
There was no measurable change in the dimensions of exposed laminaeover the 24-hour period. This confirms the conclusions of Sharman (1942),Esau (1943), and Begg and Wright (1962) that the exposed parts of grass leaves

178 Davidson and Milthorpe—Leaf Growth in Dactylis
are fully expanded, as are those leaves with exposed ligules. Measurementsof lamina expansion can therefore be confined to those leaves whose ligulesare not yet exposed.
TABLE 2
The extension in 24 hours, initial cell number, and carbohydrate content ofsuccessive sections of inner and outer leaf bases
Distance Initial total solublefrom tiller Extension Cell number carbohydrate content
Section base (cm) (cm) (io'/g fresh weight) (per cent dry weight)Inner Outer Inner Outer Inner Outer
a 6-0-8-4 ° ° 6-o 1-7 8-9 4-8b 3-5-6-0 o o 51 12 236 5-4c 1-0-3-5 I-S ° io-6 1-2 32-0 9-2d o-i-o i-o o 24-7 2-5 14-9 14-9
S.E. . . ±0-05 . . ±1-42 ±0-08
The piercing technique employed on the stubble did result in reduced leafextension—3-2^0-14 and 2-5^0-14 cm/day in normal and pierced plants,respectively—perhaps by damaging vascular tissue and thereby reducing sub-strate flow to expanding cells. However, the technique served to indicate thatcell extension was confined to the basal Sections c and d which consisted ofsmall densely packed cells (Table 2). The zone of extension did not extendinto Section b in any replicate and, therefore, was probably limited to thelowest 3 cm of the stubble.
T H E EXTENT OF CARBOHYDRATE REDUCTION INDIFFERENT LEAVES
Measurements were made of the reduction in the soluble carbohydratecontent of expanding and fully expanded leaves brought about by treatmentin the dark and growth following defoliation.
Experimental Procedures
Plants were grown at 250 C and 7-78 cal visible radiation/cm2/hr for 16 hoursper day for six weeks after emergence. One group was then defoliated byremoving all laminae and kept under the same conditions (defoliation treat-ment); another was put into continuous darkness at a similar temperature(dark treatment), and a third group was grown without disturbance (normaltreatment). After two days in these conditions the plants were stripped of allexposed laminae, and the bases of expanding and fully expanded leavesseparated. These portions were then analysed for soluble carbohydrates.
Results
In each of the treatments 74 per cent of the dry matter of the stubble wasaccounted for by the sheaths of outer fully expanded leaves and 36 per cent bythe younger expanding leaves. Total soluble carbohydrates comprised 10-3,6-5, and 4-8 per cent of the dry matter in the normal, defoliation, and dark

Davidson and Milthorpe—Leaf Growth in Dactylis 179
treatments, respectively. In the younger expanding leaves the total solublecarbohydrate concentration was almost double that of the outer, older, fullyexpanded leaves (Table 3).
TABLE 3
The soluble carbohydrate contents of the bases of expanding and fullyexpanded leaves in plants subjected to dark and defoliation treatments
for 2 daysCarbohydrate content
TreatmentNormal
Defoliation
Dark
S.E.
LeavesExpandingFully
expandedExpandingFully
expandedExpandingFully
expanded
(per cent
Alcohol-solubleIO-I
5 98-5
2-73 7
2'I
±049
dry matter)
FructosanIO-I
1 61 8
i ' i
2-5
1-2
±O-I9
Totalsoluble
1 3 2
7-51 0 3
3 86 2
3 4±0-52
Reduction oftowi as per
cent of normalvalues
2 2
49S3
55
The dark treatment led to the same relative reduction of carbohydrate con-centration in expanding as in fully expanded leaves, whereas, with the de-foliation treatment, the relative reduction in the expanding leaves was onlyhalf of that of the older fully expanded leaves which comprised three-quartersof the total dry weight. The results suggest that only that soluble carbohydrateheld in the base of an expanding leaf is used for subsequent expansion of thatleaf; the much greater quantity of soluble carbohydrate in the bases of olderfully expanded leaves may have little direct influence on leaf expansion butmay well be used by roots and tiller buds.
INFLUENCE OF LEVEL OF DEFOLIATION ON LEAF GROWTH
The effect of varying the height of cutting on the subsequent rate of leafexpansion was investigated.
Experimental ProceduresPlants were grown under the conditions described in the section on leaf
expansion above for five weeks and the tillers were then marked to delineateSections a, b, c, and d, as described on p. 177. All laminae were removed andthe stubble was cut off at the top of Sections a, b, c, and d. Leaf expansion wasmeasured after 24 hours.
ResultsThe removal of all laminae led to a 44 per cent reduction in leaf extension
(Table 4). Further removal of Section a, the top one-third of the stubble

180 Davidson and Milthorpe—Leaf Growth in Dactylis
length, caused no further reduction, but removal of Sections b and c led toreductions of 62 and 91 per cent respectively in leaf extension compared withuncut plants. The effects on leaf-area increase were essentially the same.
TABLE 4
Leaf growth in the first 24 hours after defoliation from maintillers cut to different heights
Level of defoliation—to the top of section
Intact plantsabcdS.E.
Stubble length(cm)
8-88 95 93-5i -o
±0-73
Leaf extension(cm)
3-2i-81 9I '2o-3
±0-15
Leaf areaincrease
(cm1)
3 32-12-1I ' lO-2
±O-2I
Reduced leaf expansion which follows the removal of all tissue above thelimit of cell expansion is presumably the result of depleted substrate (andpossibly hormone) supply. The drastic effect of removing all but 1 cm ofstubble involves the added consequence of removing part of the reservoirof cells upon which leaf expansion depends.
INFLUENCE OF NUTRIENT SUPPLY ON LEAF GROWTH AFTERDEFOLIATION
The effects of leaf removal were examined under conditions of continuousand restricted nutrient supply.
Experimental Procedures
Plants were grown at 220 C under a 16-hour day of 3-2 cal visible radiation/cm2/hr for six weeks after emergence with the normal nutrient supply.Half the number of pots were then leached with standard and half with one-tenth standard Long Ashton nutrient solution. Two days later, plants in eachgroup were subjected to one of the following defoliation treatments:
(1) undefoliated; (2) laminae of fully expanded leaves removed; (3) visiblelaminae of expanding leaves removed; (4) all visible laminae removed.Leaf expansion of the main tiller was measured after 29 hours.
Results
At each nutrient level 63 per cent of the total leaf area was removed withthe fully expanded leaves and 37 per cent with the younger expanding leaves.The effect of reducing the nutrient supply on leaf growth was rapid. Withintwo days the low nutrient supply led to a significant reduction in leaf growth(Table 5). At both nutrient levels complete lamina removal reduced leafextension to the same value. At the high (normal) level of nutrient supplythe removal of all fully expanded leaves, i.e. two-thirds of the total leaf area,

NoneFully expandedExpandingAll
4 34 63-42-6
Davidson and MiUhorpe—Leaf Growth in Dactylis 181
did not reduce the rate of leaf extension. However, this was reduced signi-ficantly by removing the exposed laminae of the expanding leaves. This sup-ports the concept already developed that under the conditions of continuousnutrient supply maintained in these experiments, leaf expansion depended
TABLE 5
Leaf extension from the main tiller of plants sub-jected to different defoliation treatments at two
levels of the external nutrient supply
Leaf extension (cm)
High nutrient Low nutrientLaminae removed supply supply
3'32-8362-s
S.E. . . ±026 ±026
primarily on the photosynthesis of the expanding leaves themselves. Whenthese exposed laminae were removed, photosynthate from the older, fullyexpanded leaves was presumably drawn upon since their removal led toa further reduction in extension.
At low nutrient status the results were not conclusive. Following theremoval of all fully expanded leaves, leaf extension was similar to that whenall leaves were removed and significantly less than when only expanding leaveswere removed; yet the removal of either category of leaf resulted in extensionnot significantly different from that of the uncut plants. In order to test theseresponses further, plants were grown as before except that no nutrients wereadded from two weeks prior to defoliation in order to deplete further thenutrient status of the plants. Leaf extension was then measured four daysafter defoliation and was I I - I , io-o, and 8-6^0-^2 cm from the three treat-ments, uncut, expanding leaves removed, and fully expanded leaves removed,respectively. These further results indicate that, under conditions of limitingnutrient status, the removal of fully expanded leaves caused a significantreduction in leaf extension, whereas the removal of the younger, expandingleaves had no significant effect.
The results suggest that there may be an interaction of nutrient and carbo-hydrate limitations on leaf growth following defoliation. The fully expandedleaves were presumably a reservoir of labile nutrients, and their removal,when the nutrient status was low, led to reduced leaf extension; further, theseolder leaves possibly supply to the roots carbohydrate required for continueduptake of nutrients. When the nutrient supply was high, leaf extension wasapparently controlled by the carbohydrate concentration in the base of theexpanding leaves, and was influenced primarily by the photosynthate pro-duced by the exposed laminae of these leaves.

182 Davidson and Milthorpe—Leaf Growth in Dactytis
DISCUSSION
In cocksfoot, as in other grasses, the leaves which are still expanding arethose emerging vertically from the enclosing sheaths; leaves which are fullyexpanded are those with fully exposed ligules from which the laminae bendback. In the experiments, these visible expanding leaves were mainly respon-sible for the initial increase in leaf area after defoliation. In the first four daysafter cutting to a height of 2-5 cm, only 6 per cent of the increase in leaf areacould be attributed to expanding leaves too small to be cut by the treatmentimposed.
Some confusion is evident in publications concerning the zone of leafexpansion. Sampson (1952) and Stoddart and Smith (1955) stated that thegrowing region of grasses is concentrated at the base of the blade and the baseof sheath. Sharman (1942) concluded from measurements of epidermal cellsof maize leaves that cell extension stopped when the tissue emerged from thesheath. Begg and Wright (1962), working with Phalaris anmdmacea, agreedwith this view and suggested that the sudden cessation of cell expansion uponemergence from the sheath was associated with phytochrome; however, theyfound that their measurements of epidermal cell lengths were too variable todetermine when cell expansion stopped. In the present study measurementsof leaf extension indicate that cell expansion was restricted to regions wellwithin the surrounding sheaths and had ceased before the tissue emerged fromthem.
The observation of a higher concentration of soluble carbohydrates inexpanding compared with fully expanded leaves and the apparent increase inconcentration towards the leaf base is compatible with the account of leafdevelopment presented by Sharman (1942) from a detailed morphologicalexamination. He showed that, in fully expanded leaves, differentiated vascularelements are linked directly with those of the roots. He also found thatvascular differentiation began at the leaf tip and extended down towards theleaf base, so that the lowest region of expanding leaves is without differentiatedvascular tissue. Presumably, for carbohydrates to move out of an expandingleaf, they must traverse by diffusion this basal densely packed region ofdividing cells; this could result in a build-up of carbohydrate above this zone.If this is the explanation of carbohydrate build-up in the base of an expandingleaf, then the carbohydrate there would be derived from photosynthesis ofthe exposed blade of the expanding leaf itself. In this respect, autoradiographicstudies by Williams (1964) showed very little export of carbohydrate fromyoung leaves; export only commenced when laminae were almost fullyexpanded.
A positive relationship between the rate of leaf expansion and totalsoluble carbohydrate content of the stubble existed when the carbohydratecontent was varied by dark treatment, but not when it was varied by growthfollowing defoliation. The results indicate that the carbohydrate of primaryimportance in developing the leaf surface is that present above the zone of
Davidson and Milthorpe—Leaf Growth in Dactytis 183
expansion of expanding leaves (and possibly in roots of those grasses in whichcarbohydrates accumulate in roots). An originally small but increasing supplyto these zones following defoliation will normally result from photosynthesisof the upper sections as the leaves grow. There is no such source in darknessor for older leaves which are fully expanded when cut; the soluble carbo-hydrate concentration in fully expanded leaves, which comprise the greater
- 3
- 4
- 50 10 15 20 25
Days after defoliation30 3S
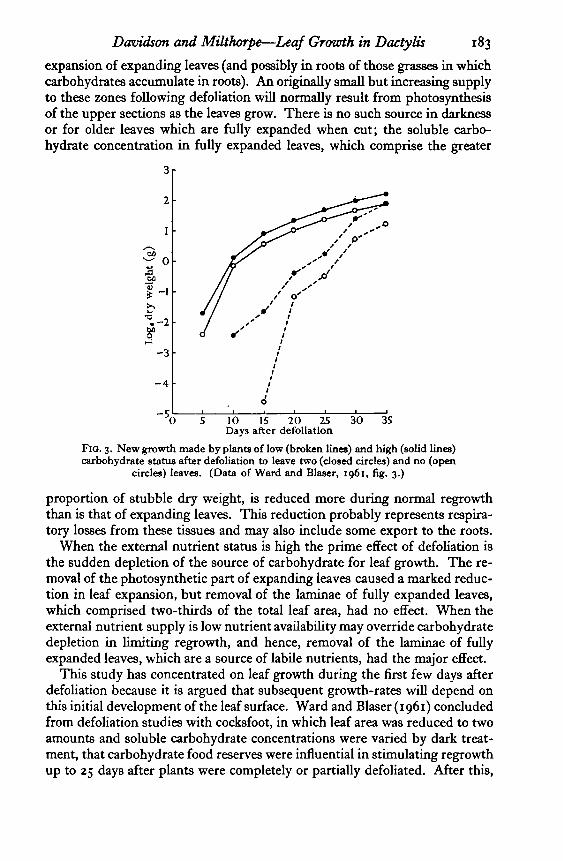
FIG. 3. New growth made by plants of low (broken lines) and high (solid lines)carbohydrate status after defoliation to leave two (closed circles) and no (open
circles) leaves. (Data of Ward and Blaser, 1061, fig. 3.)
proportion of stubble dry weight, is reduced more during normal regrowththan is that of expanding leaves. This reduction probably represents respira-tory losses from these tissues and may also include some export to the roots.
When the external nutrient status is high the prime effect of defoliation isthe sudden depletion of the source of carbohydrate for leaf growth. The re-moval of the photosynthetic part of expanding leaves caused a marked reduc-tion in leaf expansion, but removal of the laminae of fully expanded leaves,which comprised two-thirds of the total leaf area, had no effect. When theexternal nutrient supply is low nutrient availability may override carbohydratedepletion in limiting regrowth, and hence, removal of the laminae of fullyexpanded leaves, which are a source of labile nutrients, had the major effect.
This study has concentrated on leaf growth during the first few days afterdefoliation because it is argued that subsequent growth-rates will depend onthis initial development of the leaf surface. Ward and Blaser (1961) concludedfrom defoliation studies with cocksfoot, in which leaf area was reduced to twoamounts and soluble carbohydrate concentrations were varied by dark treat-ment, that carbohydrate food reserves were influential in stimulating regrowthup to 25 days after plants were completely or partially defoliated. After this,

184 Davidson and Milthorpe—Leaf Growth in Dactylis
regrowth rates were dependent on leaf area (cf. their fig. 3). If, however,these results are plotted on a logarithmic scale, so that the slope between twopoints is the measure of relative growth-rate (Fig. 3), it is clear that theadvantage attributed to a higher leaf area existed from the first harvest, andthe influence associated with carbohydrate status was confined to the earlygrowth phase. Their results in fact suggest that some factor associated withcarbohydrate status influences leaf expansion immediately after defoliation,with early differences in leaf area resulting in subsequent differences in abso-lute growth-rates. Results from the study reported here indicate that thisfactor is the soluble carbohydrate level within the bases of expanding leaves.
The first effects of defoliation on growth may be explained largely in termsof effects on the supply of carbohydrate to leaves capable of expansion.Growth substances may also have an important influence, but their role at thisstage must remain a matter for speculation.
ACKNOWLEDGEMENTS
An Overseas Studentship awarded by the Commonwealth Scientific andIndustrial Research Organization enabled one of us (J. L. D.) to take partin this work which was included in a thesis submitted to the University ofNottingham for the degree of Doctor of Philosophy.
LITERATURE CITED
ALBERT, W. B., 1927. Studies on the growth of alfalfa and some perennial grasses. J. Am.Soc. Agron. 19, 624-54.
ARCHBOLD, H. K., 1945. Some factors concerned in the process of starch storage in thebarley grain. Nature, Lond. 156, 70-73.
BEGG, J. E., and WRIGHT, M. J., 1962. Growth and development of leaves from intercalarymeristems in Phalarii arundinacea L. Ibid. 194, 1097—8.
DAVIDSON, J. L., and MILTHORPE, F. L., 1965. The effect of temperature on the growth ofcocksfoot (Dactylis glomerata L.). Ann. Bot. 29, 407-18.
ESAU, K., 1943. Ontogeny of the vascular bundle in Zea mays. Hilgardia, 15, 327—56.GRABER, L. F., NELSON, N. T., LEUKBL, W. A., and ALBERT, W. B., 1927. Organic food re-
serves in relation to the growth of alfalfa and other perennial herbaceous plants. Bull.Wis. agric. Exp. Stn. 80, 3-128.
HEWITT, E. J., 1952. Sand and water culture methods used in the study of plant nutrition.Tech. Commun. Commonw. Bur. Hort. Plantn Crops No. 22.
MAY, L. H., i960. The utilization of carbohydrate reserves in pasture plants after defoliation.Herb. Abstr. 30, 239—45.
SAMPSON, A. W., 1952. Range Management. Principles and Practices. Wiley, New York.SHARMAN, B. C , 1942. Developmental anatomy of the shoot of Zea mays L. Ann. Bot. 6,
245-82.STODDART, L. A , and SMITH, A. W., 1955. Range Management. McGraw-Hill, New York.WARD, C. Y., and BLASER, R. E., 1961. Carbohydrate food reserves and leaf area in regrowth
of orchard grass. Crop Set. 1, 366-70.WETNMANN, H., 1948. Underground development and reserves of grasses. J. Br. Grassld
Soc. 3, 115-40.1952. Carbohydrate reserves in grasses. Proc. 6th Int. Grassld Congr. 1, 655-60.
WILLIAMS, R. D., 1964. Assimilation and translocation in perennial grasses. Ann. Bot. 28,419-26.
WRIGHT, S. T. C , 1961. Growth and cellular differentiation in the wheat coleoptile (Triticumvulgare L.). I. Estimation of cell number, cell volume and certain nitrogenous consti-tuents. J. exp. Bot. 12, 303-18.






![SISTEMATIKA IN HOROLOGIJA SKUPINE NAVADNE PASJE … · SISTEMATIKA IN HOROLOGIJA SKUPINE NAVADNE PASJE TRAVE (Dactylis glomerata agg.) V SLOVENIJI [DOKTORSKA DISERTACIJA] SYSTEMATICS](https://img.pdfslide.net/doc/110x75/5e4bca03ca227d5d900d47ce/sistematika-in-horologija-skupine-navadne-pasje-sistematika-in-horologija-skupine.jpg)






















