Embed Size (px)
Citation preview

Lecture 6Antibody genes II
•Mechanism of class switching•Regulation of class switching•Somatic mutation•T cell dependent antibody response•Affinity maturation•T independent antibody responses
Next time: Mid term test
Review session Saturday Jan 22
4:00-6:50PM YORK 2722
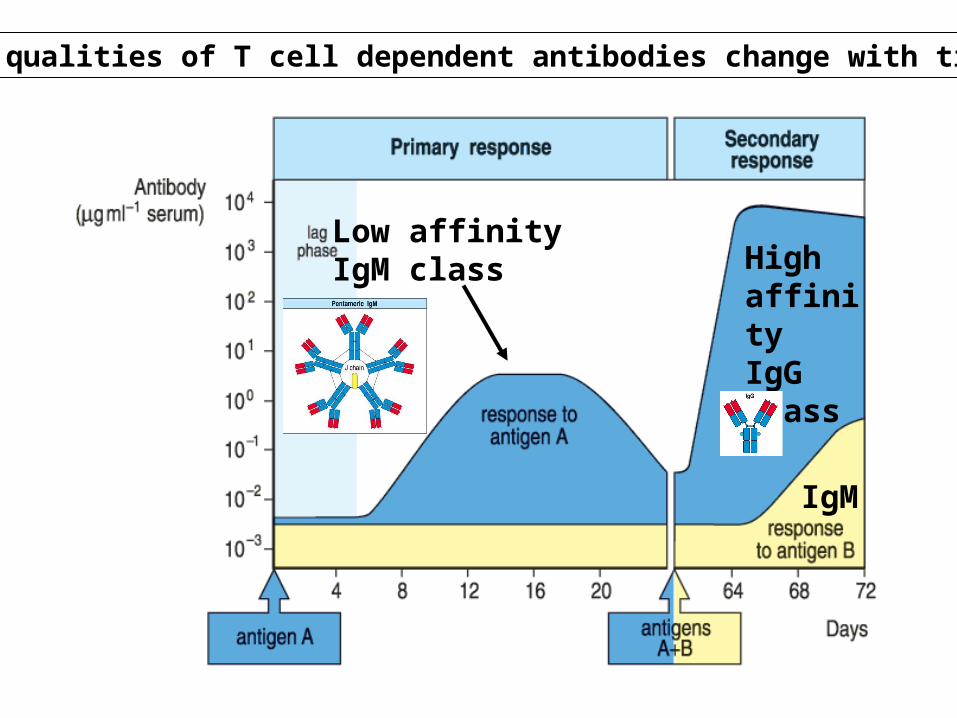
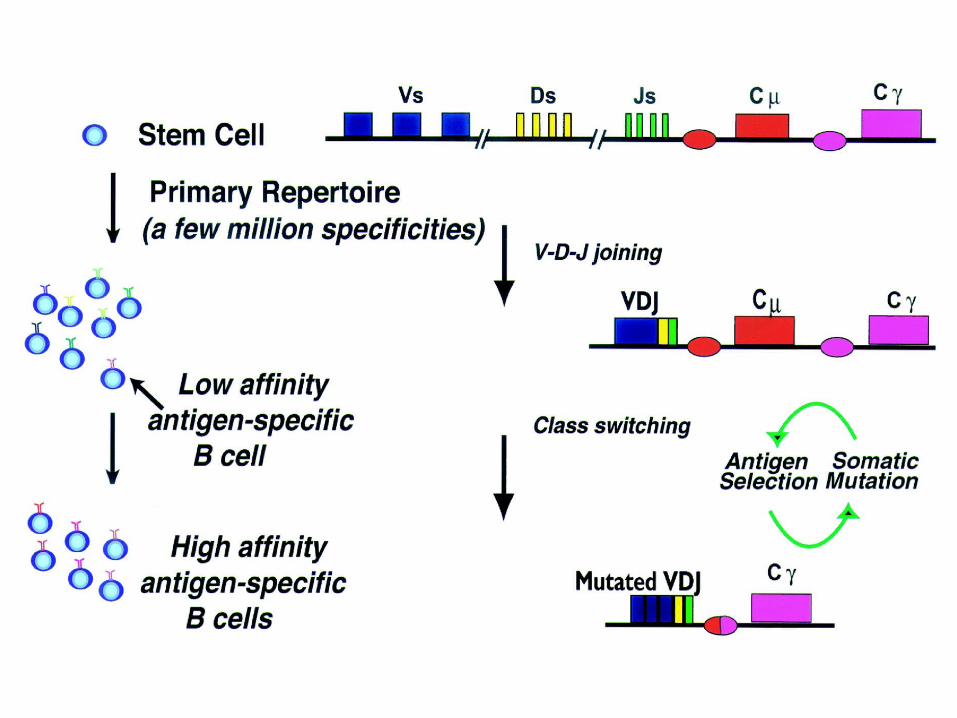
Low affinityIgM class High
affinityIgG class
IgM
The qualities of T cell dependent antibodies change with time
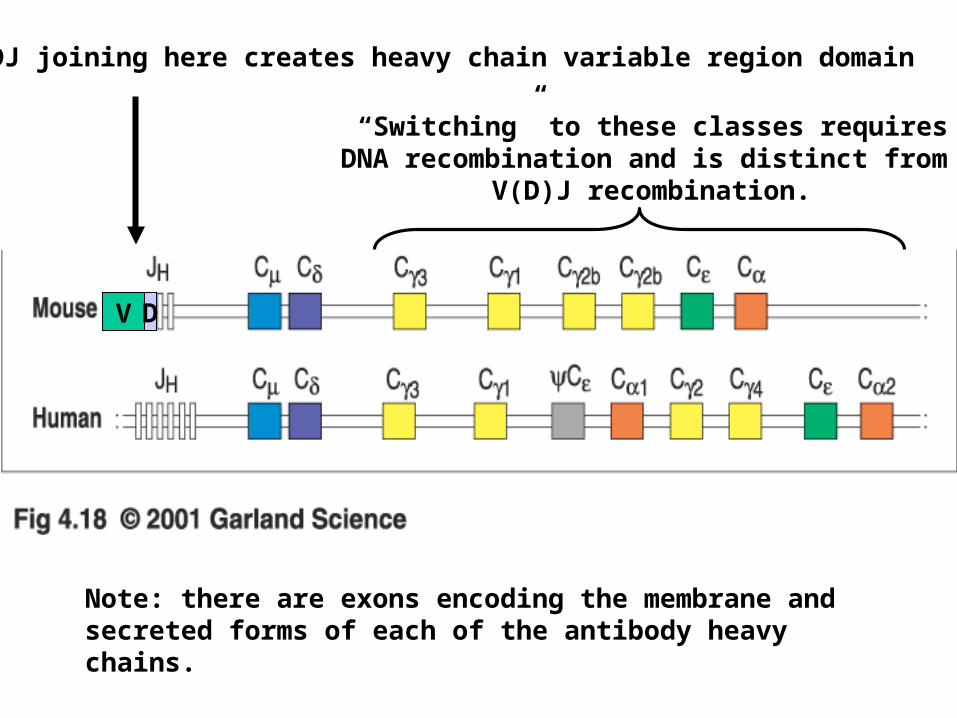
VDJ joining here creates heavy chain variable region domain
V D
Note: there are exons encoding the membrane and secreted forms of each of the antibody heavy chains.
“Switching” to these classes requiresDNA recombination and is distinct from
V(D)J recombination.
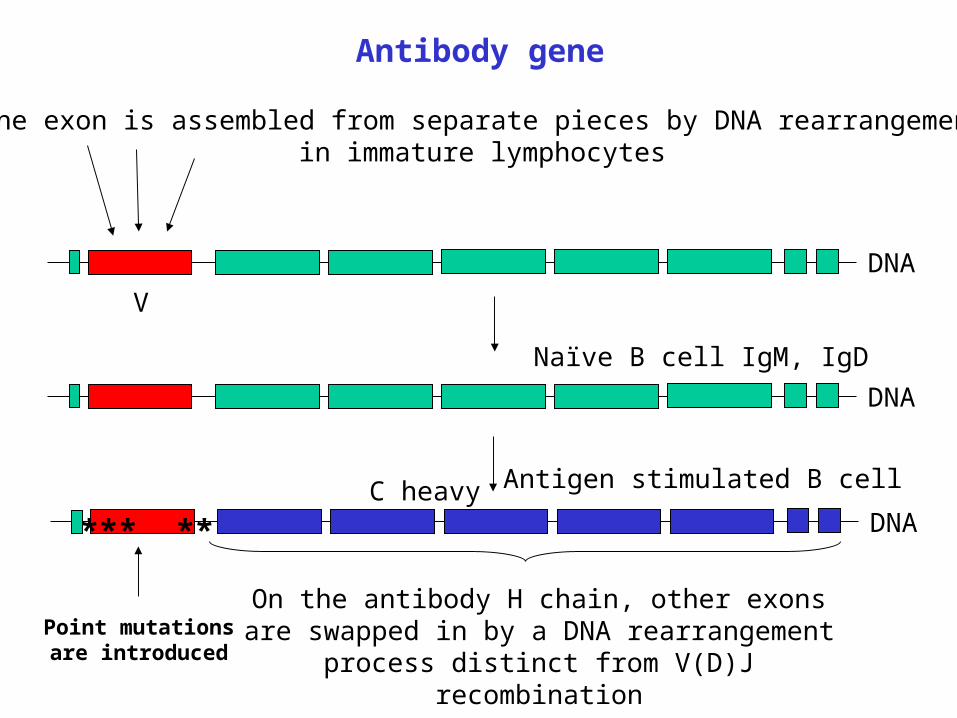
Antibody gene
DNA
One exon is assembled from separate pieces by DNA rearrangementin immature lymphocytes
DNA
V
Naïve B cell IgM, IgD
DNA
On the antibody H chain, other exons are swapped in by a DNA rearrangement process distinct from
V(D)J recombination
C heavy Antigen stimulated B cell
*** **
Point mutationsare introduced
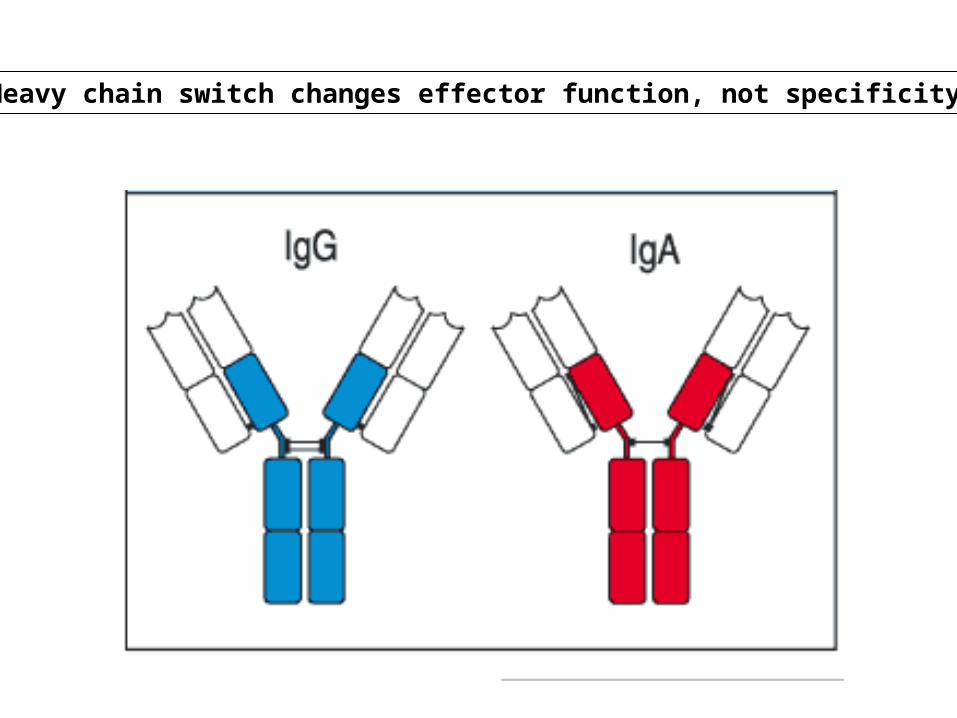
Heavy chain switch changes effector function, not specificity
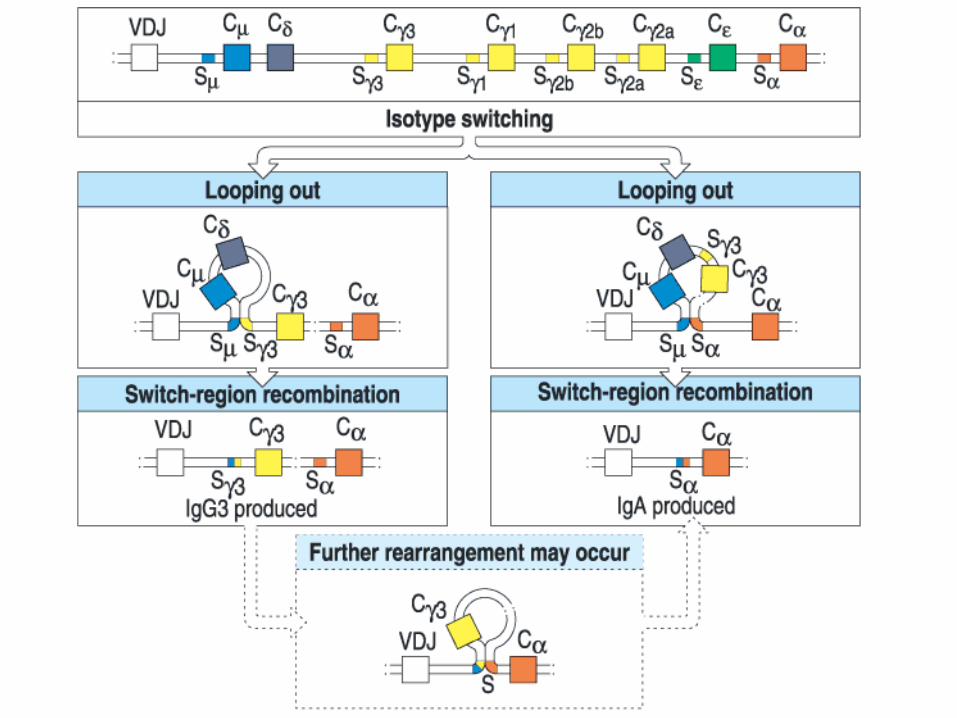

VDJ CCS S
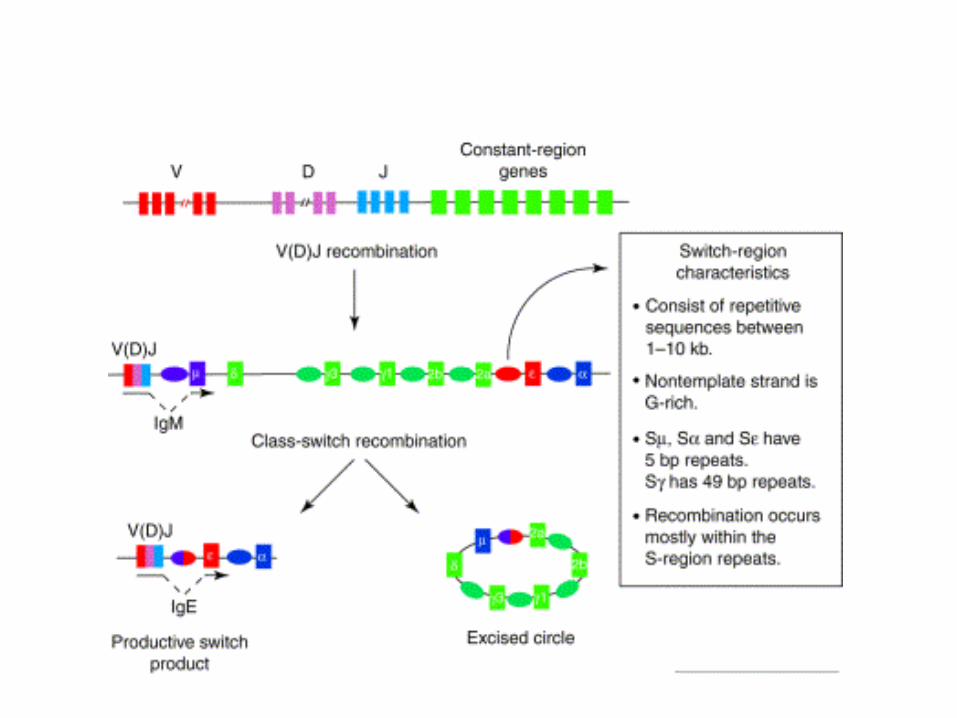
•Joining can occur anywhere within the large, repetitive S-regions. There is no obvious conserved sequence motif.•Unlike V(D)J recombination, this gene splicing occurs between, rather than within, coding sequences.•Like V(D)J recombination, targeting of elements for rearrangement is correlated with prior “sterile transcription”
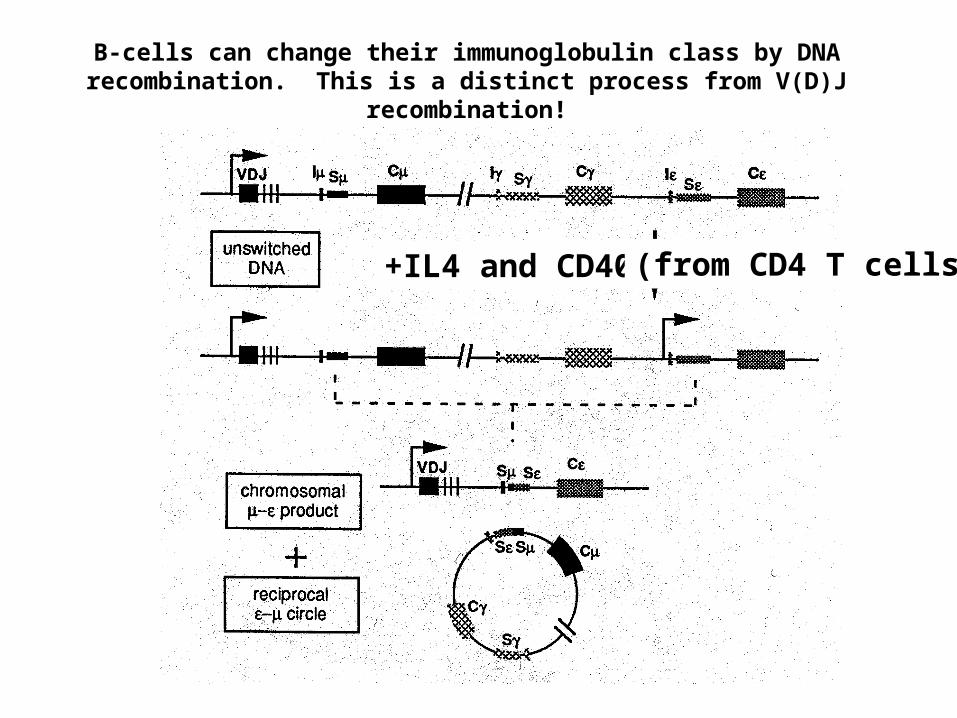
B-cells can change their immunoglobulin class by DNA recombination. This is a distinct process from V(D)J recombination!
+IL4 and CD40L (from CD4 T cells)
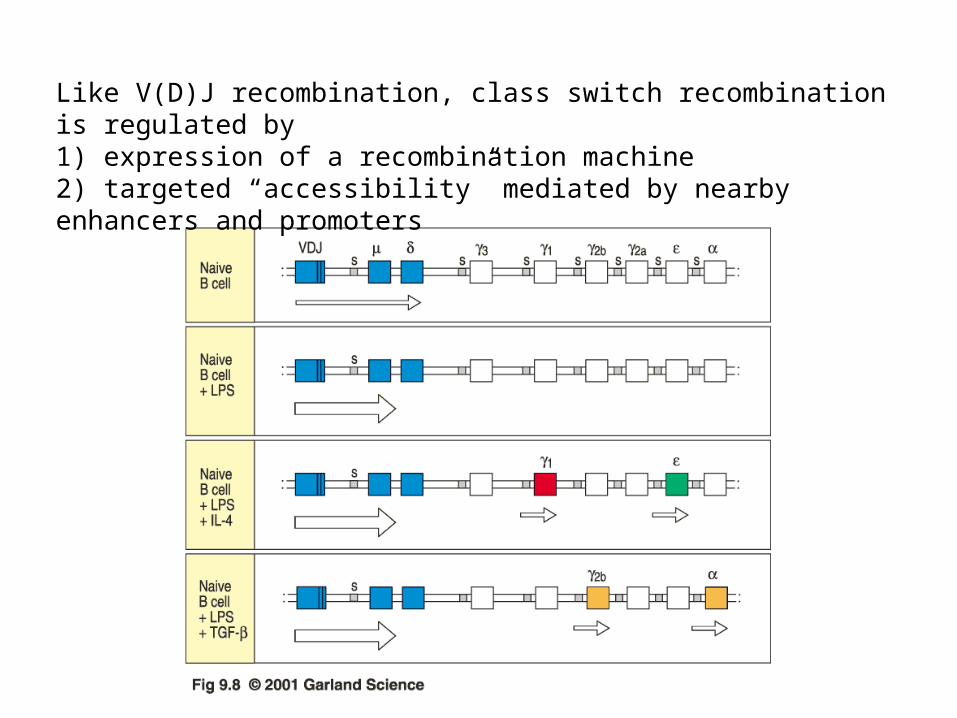
Like V(D)J recombination, class switch recombination is regulated by 1) expression of a recombination machine2) targeted “accessibility” mediated by nearby enhancers and promoters
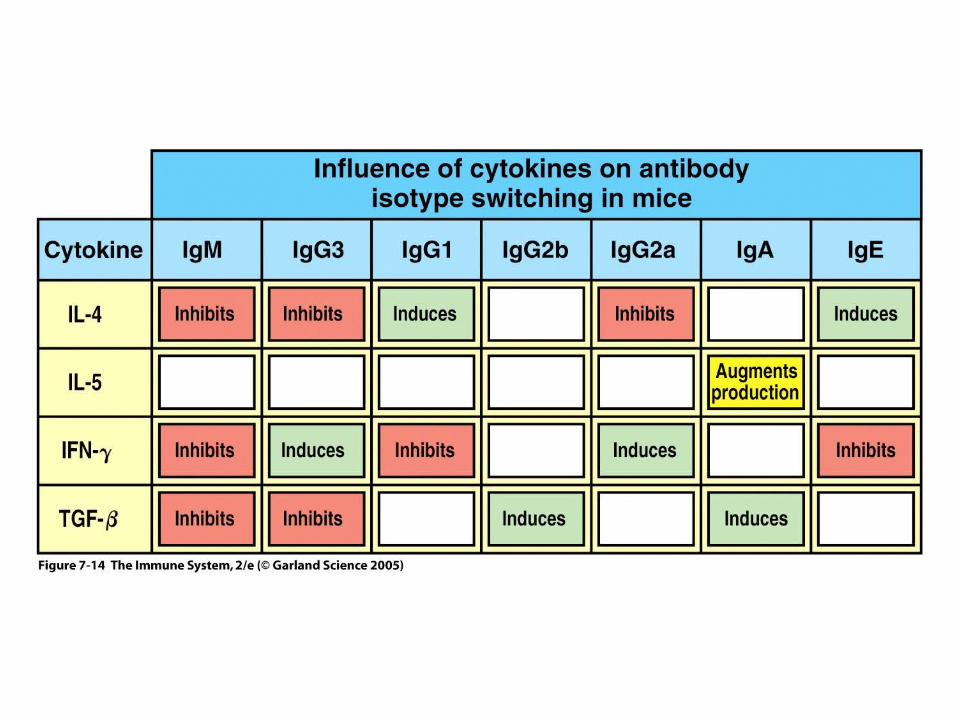
Figure 7-14
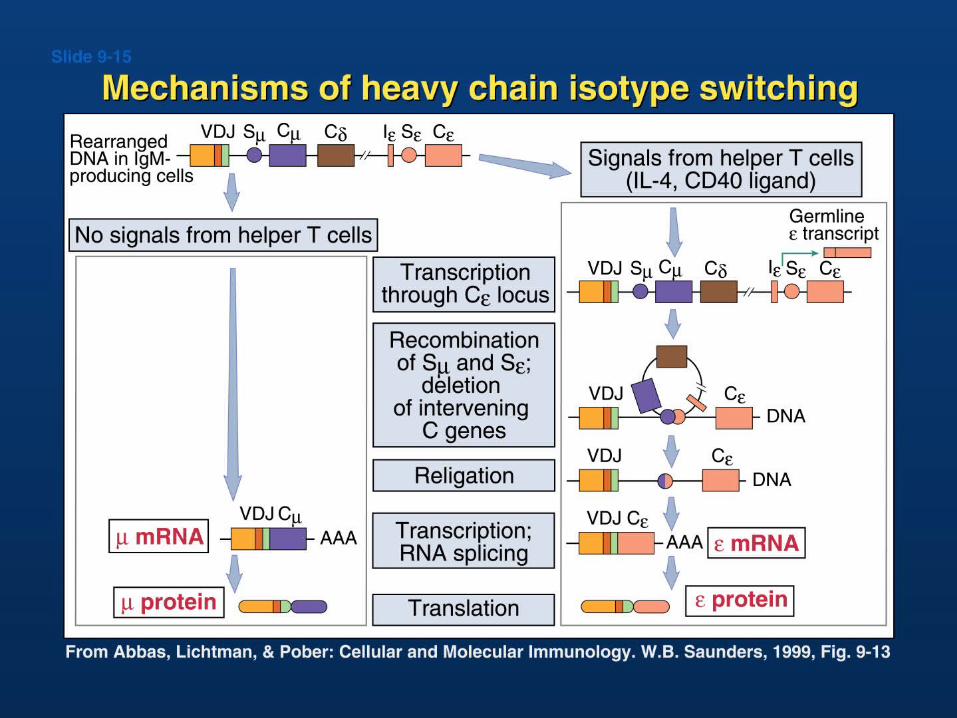
Ig isotype class switching

Concepts in class switching
•Switching to H chain classes IgG, IgA, and IgE involves irreversible DNA recombination.•During this process the H chain variable region retains the same antigen specificity.•Recombination occurs within S (switch) regions, which are several kilobases in length and highly repetitive (i.e, nothing like the recognition sequences for V(D)J recombination).•Recombination is targeted by DNA "accessibility" that is controlled by sterile transcription starting at the I-region. •I region transcription is regulated by short and long range cytokines provided mainly by T cells.•The specificity of the cytokines determines the H-chain class to be used, and hence, the effector function.
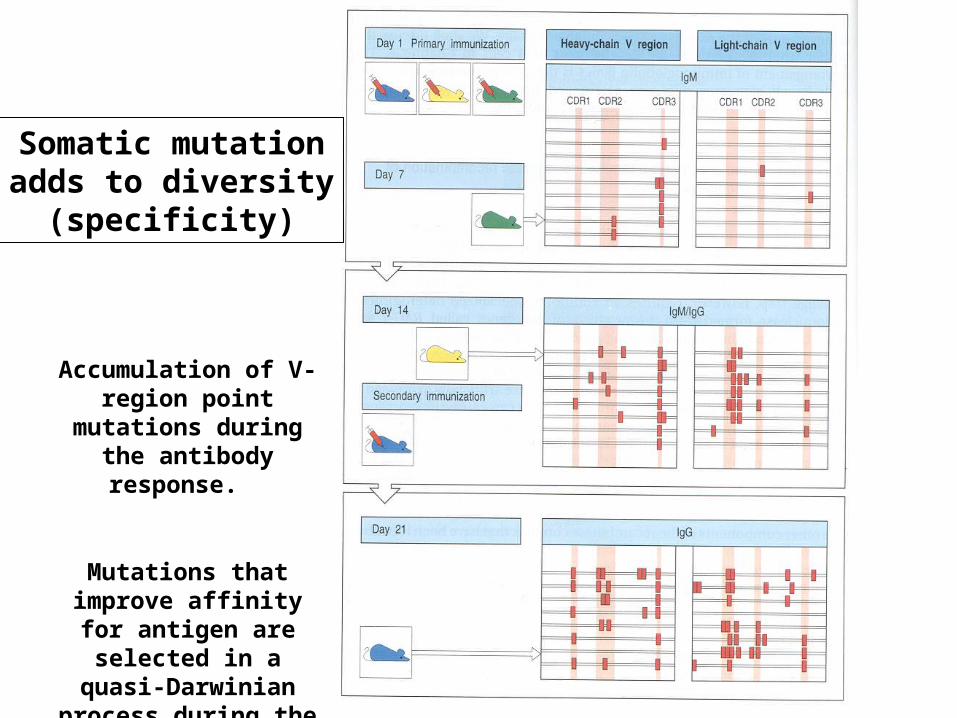
Accumulation of V-region point mutations
during the antibody response.
Mutations that improve affinity for antigen are
selected in a quasi-Darwinian process during the immune
response.
Somatic mutationadds to diversity
(specificity)

From Gearhart and Bogenhagen PNAS 80:3439, 1983
Somatic mutations are focussed on the variable region exon
Promoter Enhancer elements
Different cells carrying the same V gene were compared for sequence changes
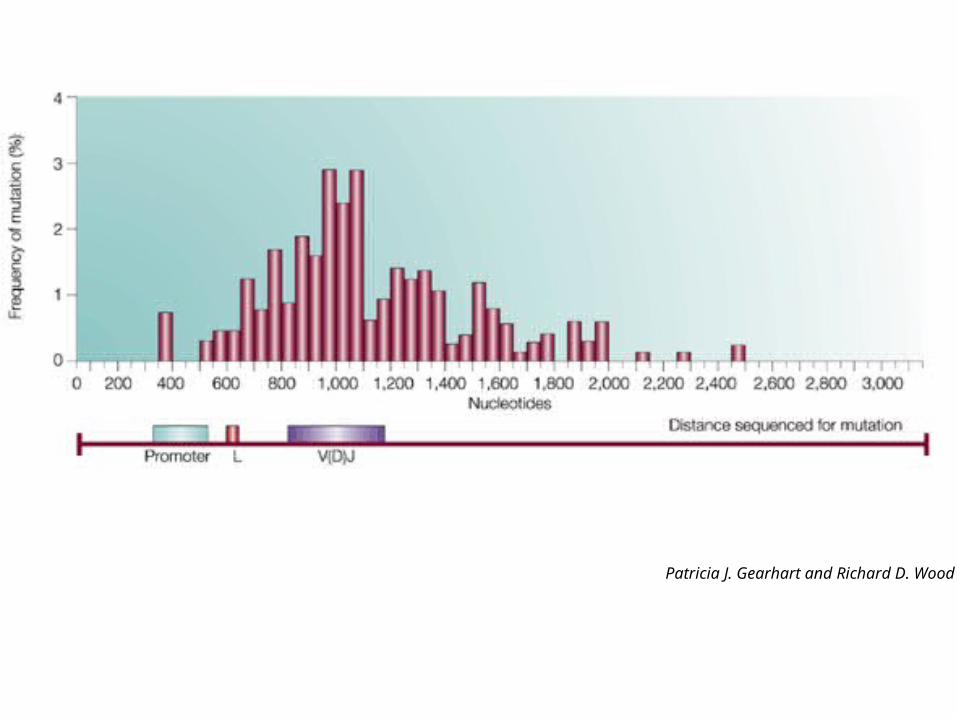
Patricia J. Gearhart and Richard D. Wood
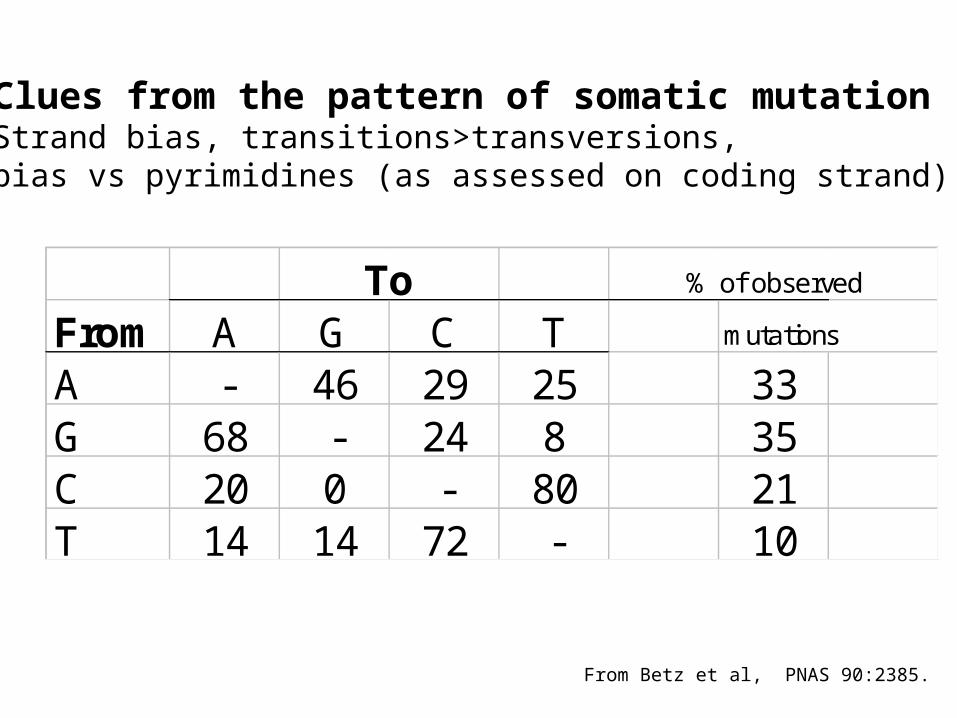
To % of observed
From A G C T mutations
A - 46 29 25 33G 68 - 24 8 35C 20 0 - 80 21T 14 14 72 - 10
Clues from the pattern of somatic mutationStrand bias, transitions>transversions, bias vs pyrimidines (as assessed on coding strand).
From Betz et al, PNAS 90:2385.

Honjo's bombshell:Knocking out a single gene blocks both somatic
mutation and class switching
• A gene specifically expressed in activated B cells undergoing class switching was knocked out.
• Mutant has hyper IgM syndrome.• IgM is only antibody class present in blood.• After immunization antibody V-regions have no
sign of somatic mutation.
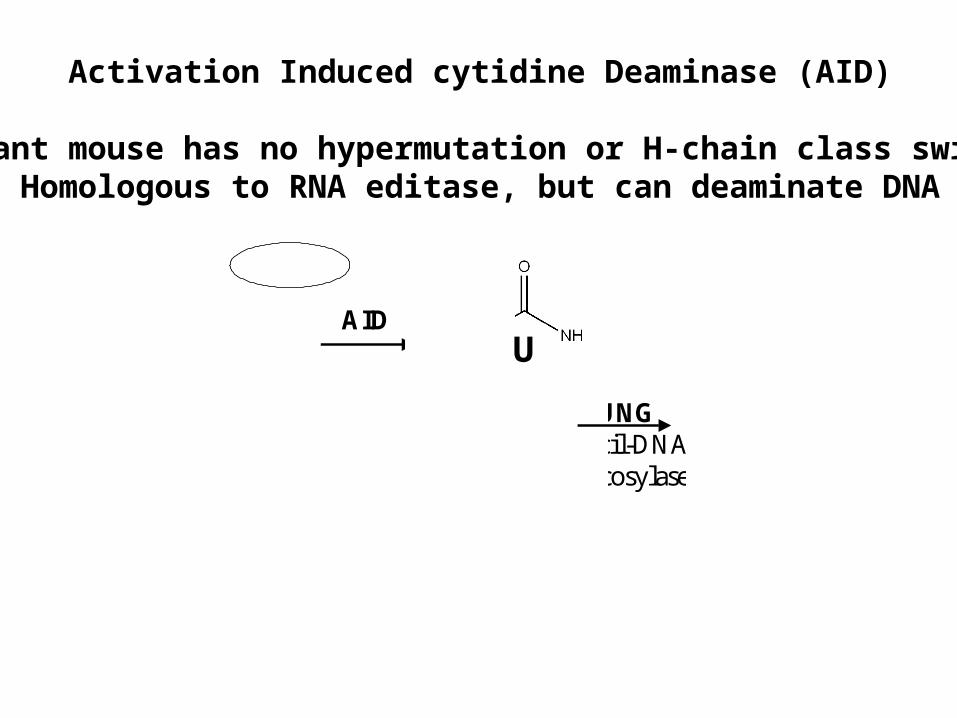
Activation Induced cytidine Deaminase (AID)
Mutant mouse has no hypermutation or H-chain class switchHomologous to RNA editase, but can deaminate DNA
UNGuracil-DNAglycosylase
UAID
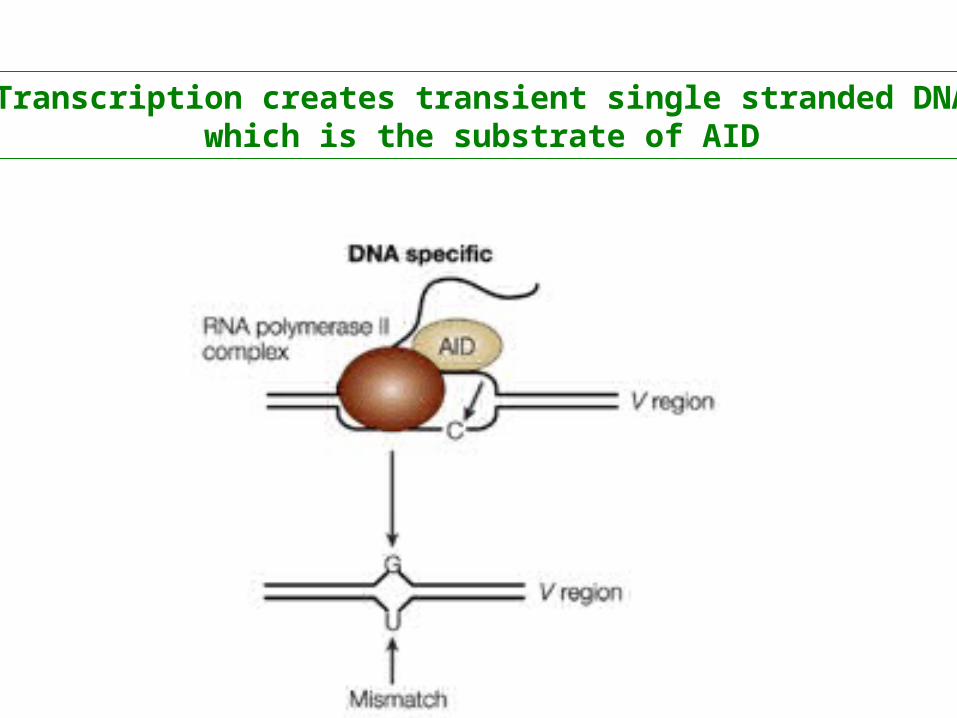
Transcription creates transient single stranded DNAwhich is the substrate of AID
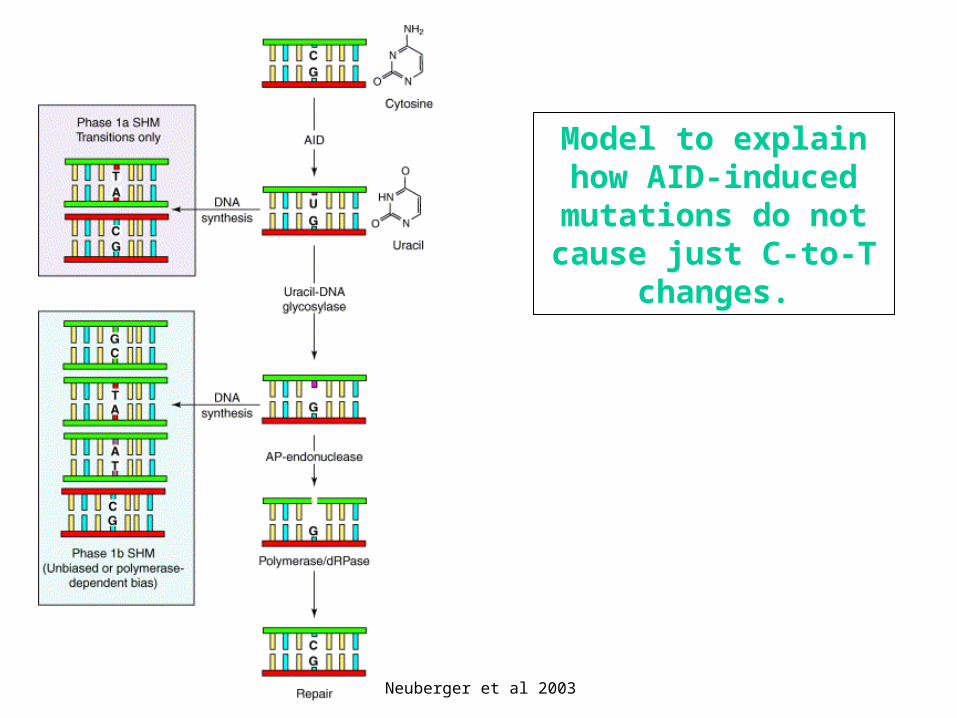
Neuberger et al 2003
Model to explainhow AID-induced mutations do not cause just C-to-T
changes.
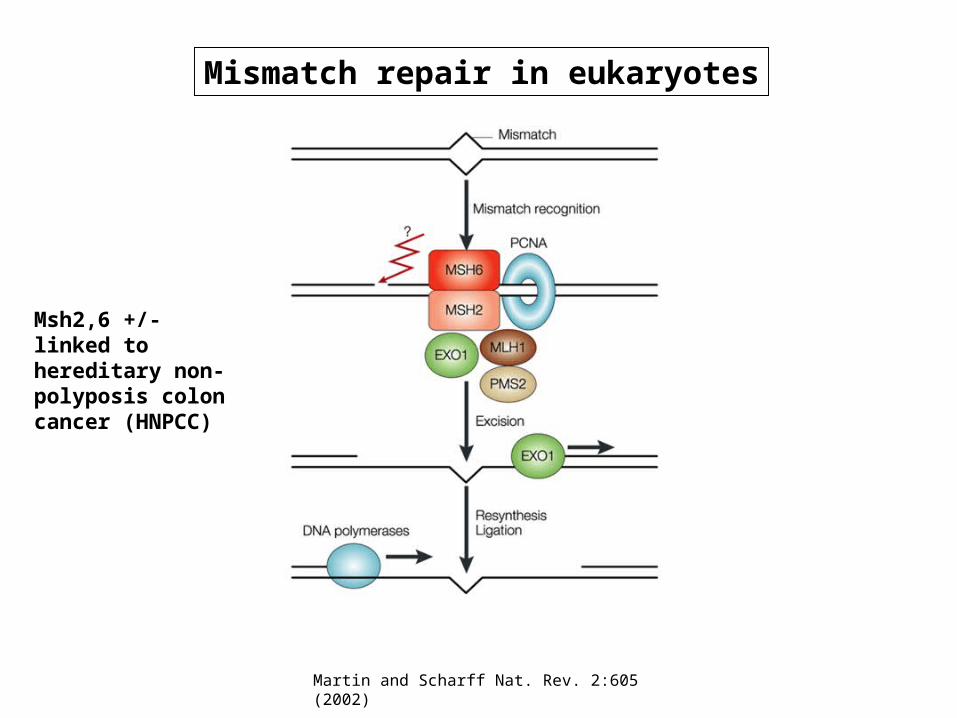
Martin and Scharff Nat. Rev. 2:605 (2002)
Mismatch repair in eukaryotes
Msh2,6 +/- linked to hereditary non-polyposis colon cancer (HNPCC)

-If mutations are routinely removed from replicating DNA, a process that prevents repair locally would target mutation. If so, knockouts of DNA repair genes would have little or no effect.
-If mutations are introduced by massive local DNA damage, possibly needed to overwhelm the normal repair mechanisms, then repair mutants would have increased mutation rates in the targeted regions (near assembled VDJs).
-Alternatively, DNA repair enzymes may be needed to generate mutations.
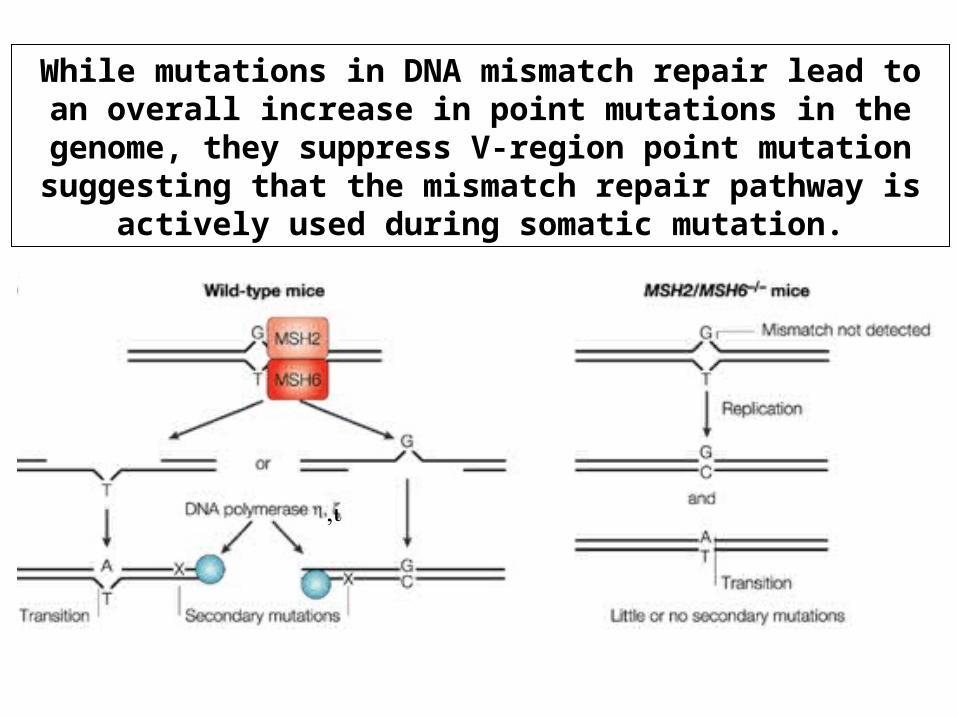
While mutations in DNA mismatch repair lead to an overall increase in point mutations in the genome, they suppress V-region point mutation suggesting that the mismatch repair
pathway is actively used during somatic mutation.
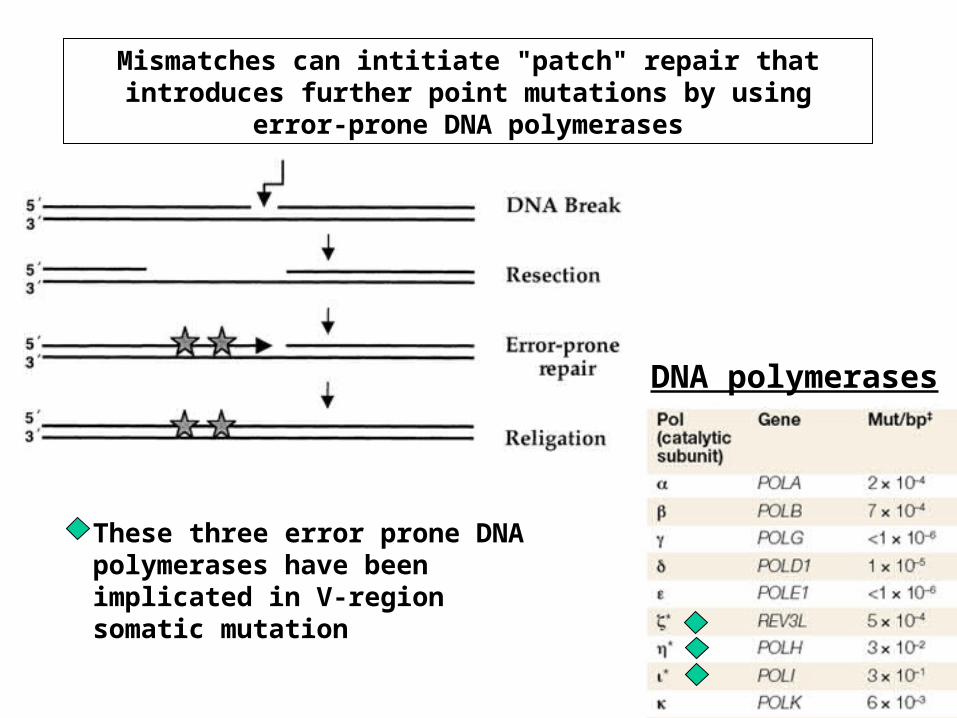
Mismatches can intitiate "patch" repair that introduces further point mutations by using error-prone DNA polymerases
These three error prone DNA polymerases have been implicated in V-region somatic mutation
DNA polymerases
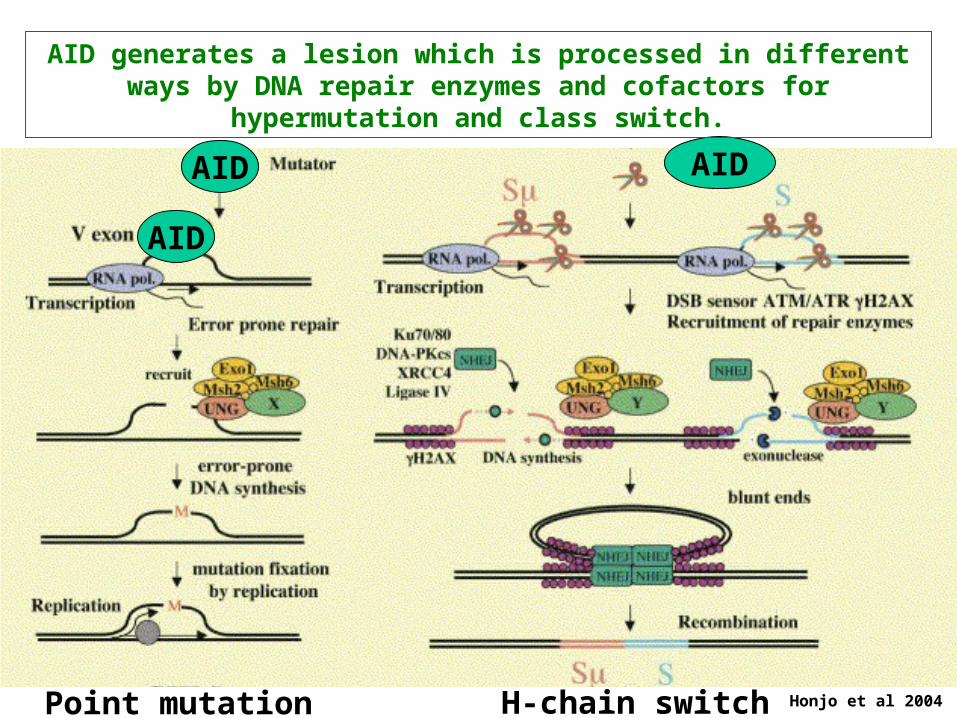
Honjo et al 2004Point mutation H-chain switch
AID AID
AID
AID generates a lesion which is processed in different ways by DNA repair enzymes and cofactors for hypermutation and class switch.

Somatic Mutation Concepts
• Surprisingly, the diversity generated by V(D)J recombination is insufficient, and is supplemented by somatic point mutation!
• Somatic mutation is induced upon antigen activation among mature B cells.
• CD4 T cell help is required.• Mutation is localized to antibody V region.• Mutation is initiated by AID at cytidines by deamination
(C>U); other repair enzymes extend the mutation.• In class switching, AID appears to generate staggered
nicks in S-regions, leading to the recruitment of the non-homologous end joining enzymes (only this part of the mechanism is reminiscent of V(D)J recombination).
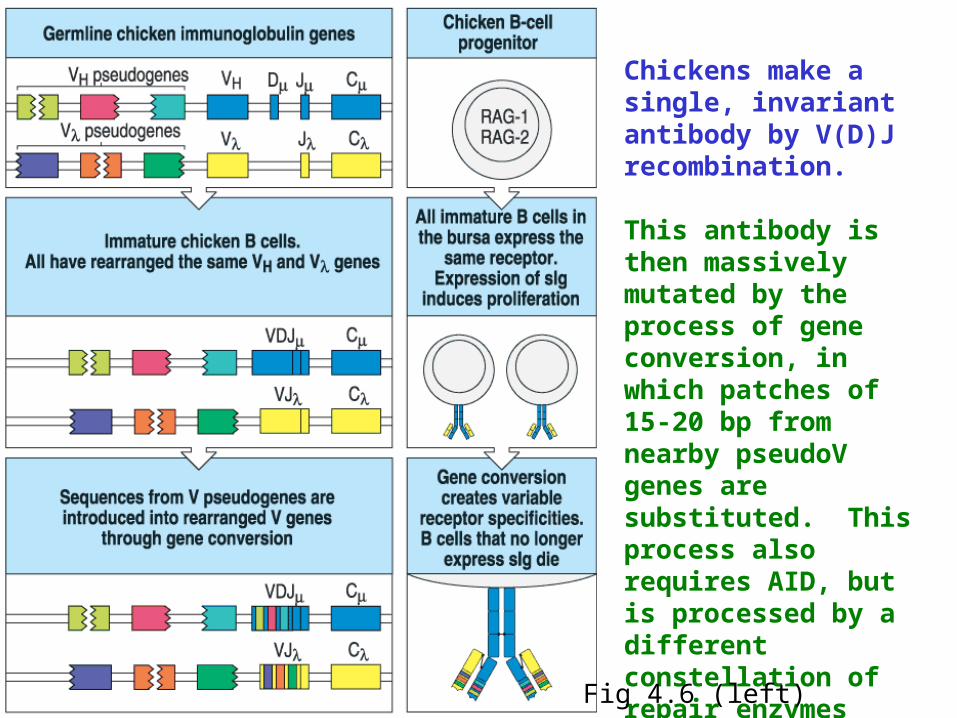
Chickens make a single, invariant antibody by V(D)J recombination.
This antibody is then massively mutated by the process of gene conversion, in which patches of 15-20 bp from nearby pseudoV genes are substituted. This process also requires AID, but is processed by a different constellation of repair enzymes that point mutation or class switch.
Fig 4.6 (left)
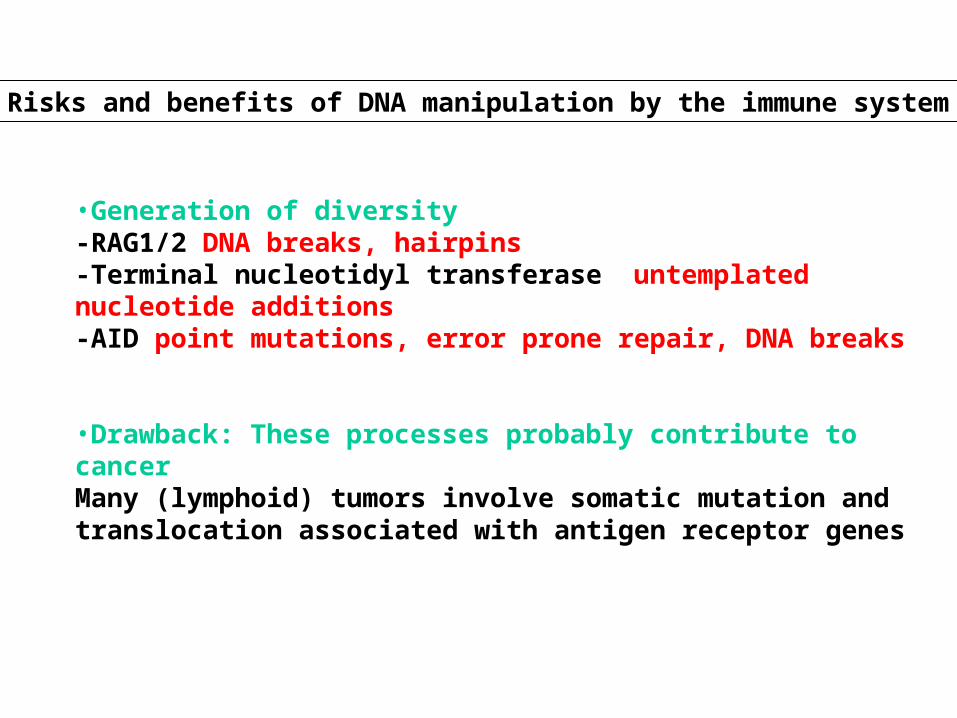
Risks and benefits of DNA manipulation by the immune system
•Generation of diversity-RAG1/2 DNA breaks, hairpins-Terminal nucleotidyl transferase untemplated nucleotide additions-AID point mutations, error prone repair, DNA breaks
•Drawback: These processes probably contribute to cancerMany (lymphoid) tumors involve somatic mutation and translocation associated with antigen receptor genes
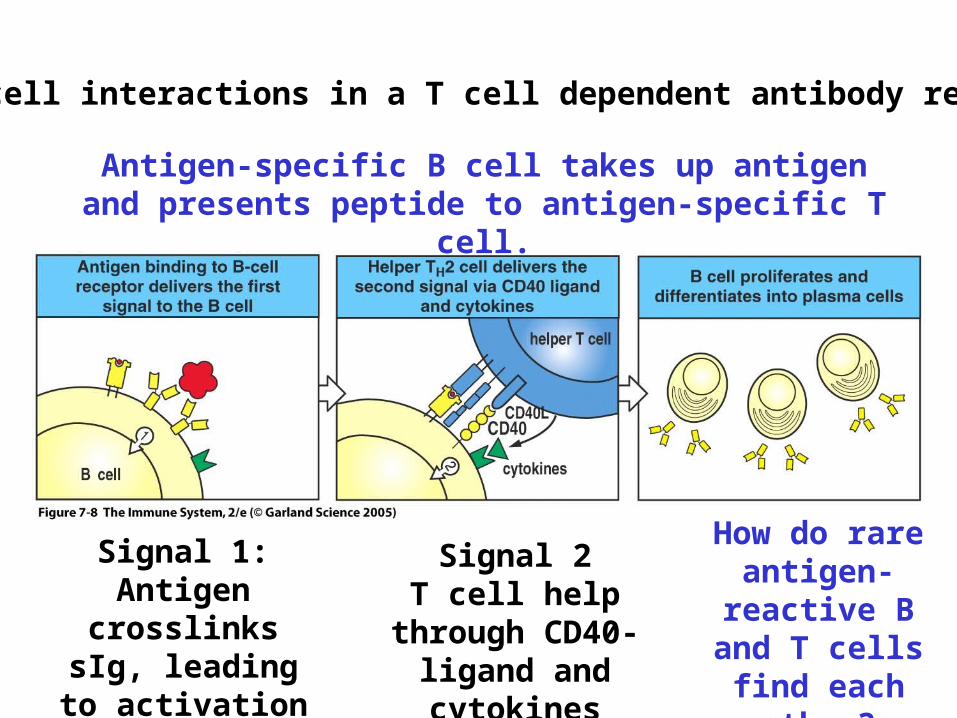
Cell:cell interactions in a T cell dependent antibody response
Antigen-specific B cell takes up antigen and presents peptide to antigen-specific T cell.
Signal 1:Antigen crosslinks
sIg, leading to activation
Signal 2T cell help through CD40-ligand and
cytokines
How do rare antigen-reactive
B and T cells find each
other?
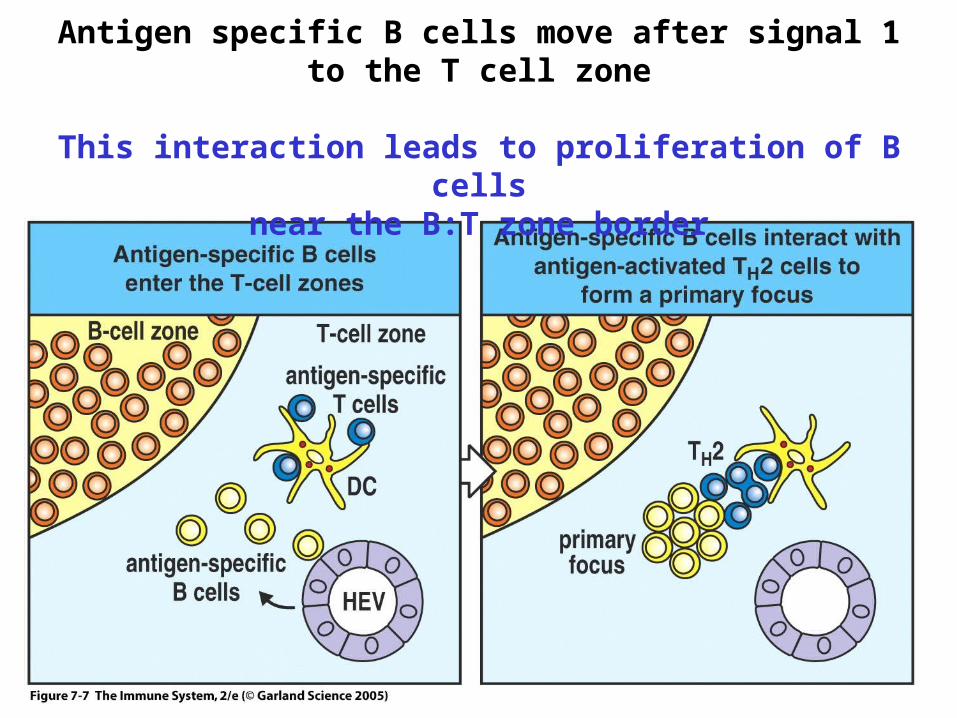
Antigen specific B cells move after signal 1 to the T cell zone
This interaction leads to proliferation of B cellsnear the B:T zone border
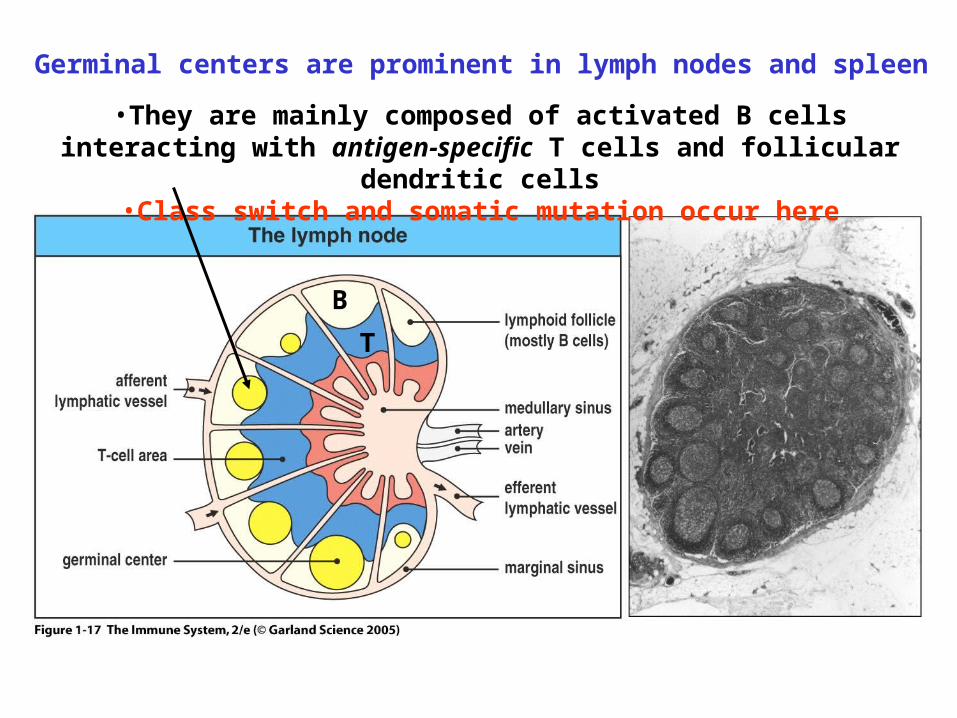
•They are mainly composed of activated B cells interacting with antigen-specific T cells and follicular dendritic cells
•Class switch and somatic mutation occur here
B
T
Germinal centers are prominent in lymph nodes and spleen
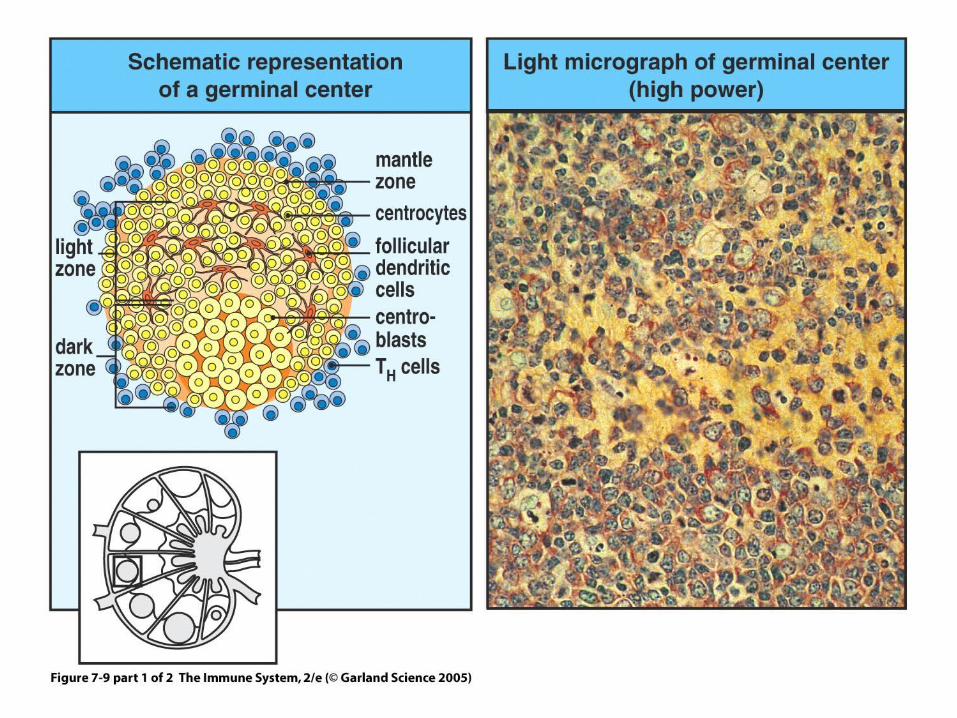
Figure 7-9 part 1 of 2
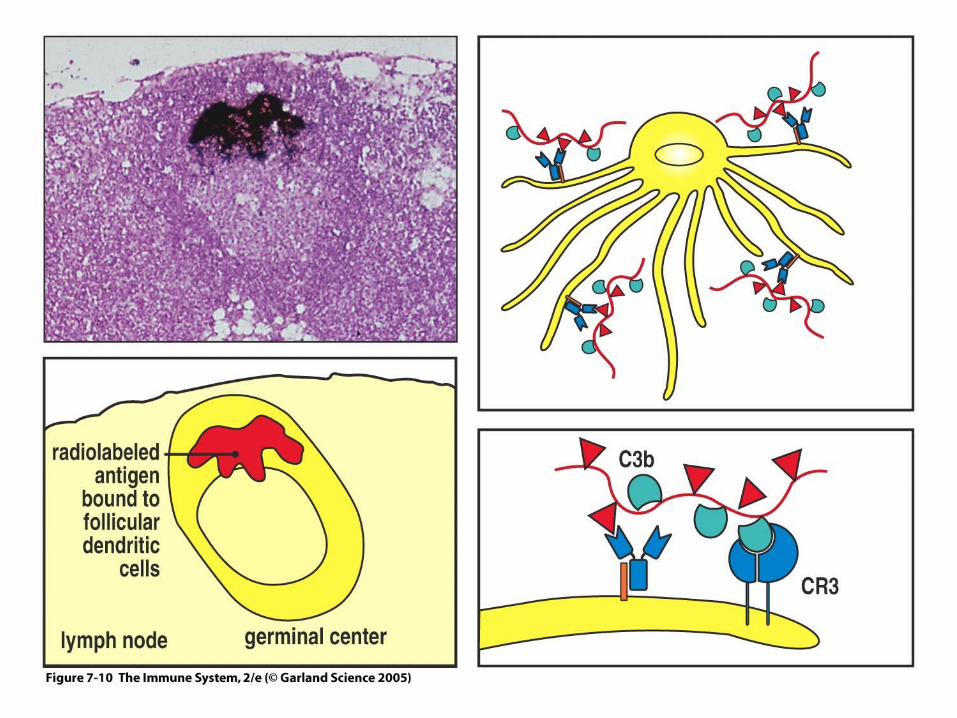
Figure 7-10
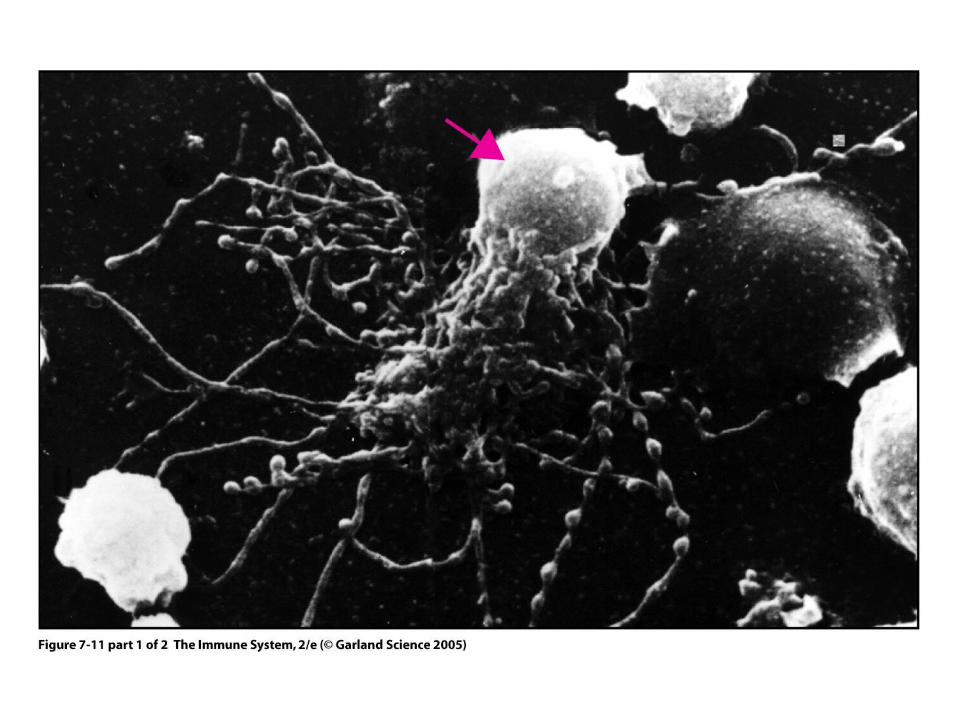
Figure 7-11 part 1 of 2
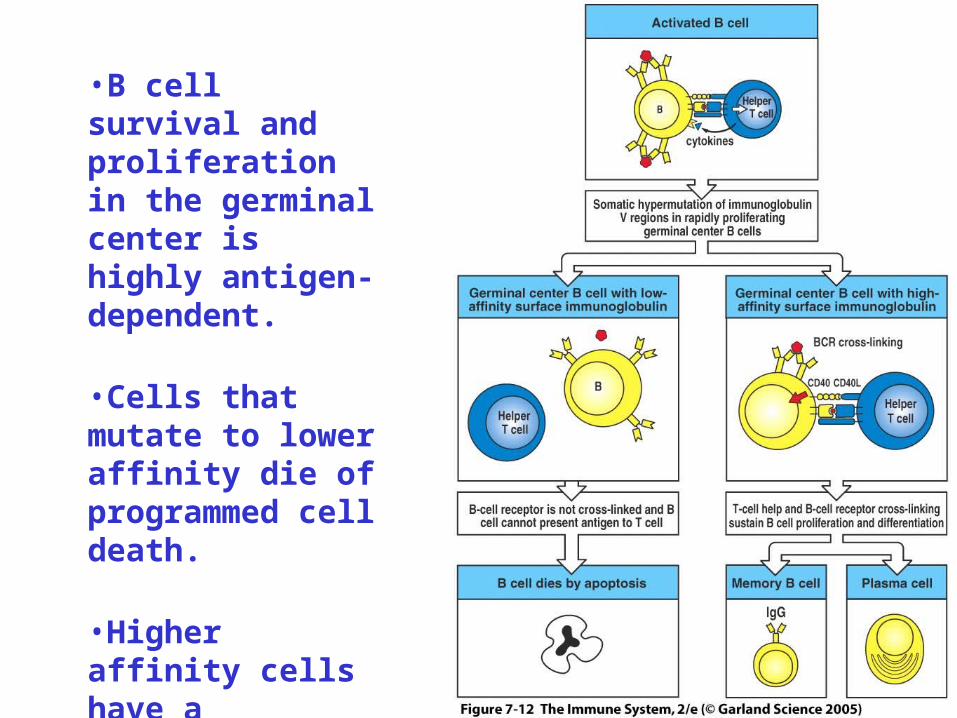
•B cell survival and proliferation in the germinal center is highly antigen- dependent.
•Cells that mutate to lower affinity die of programmed cell death.
•Higher affinity cells have a competitive advantage.
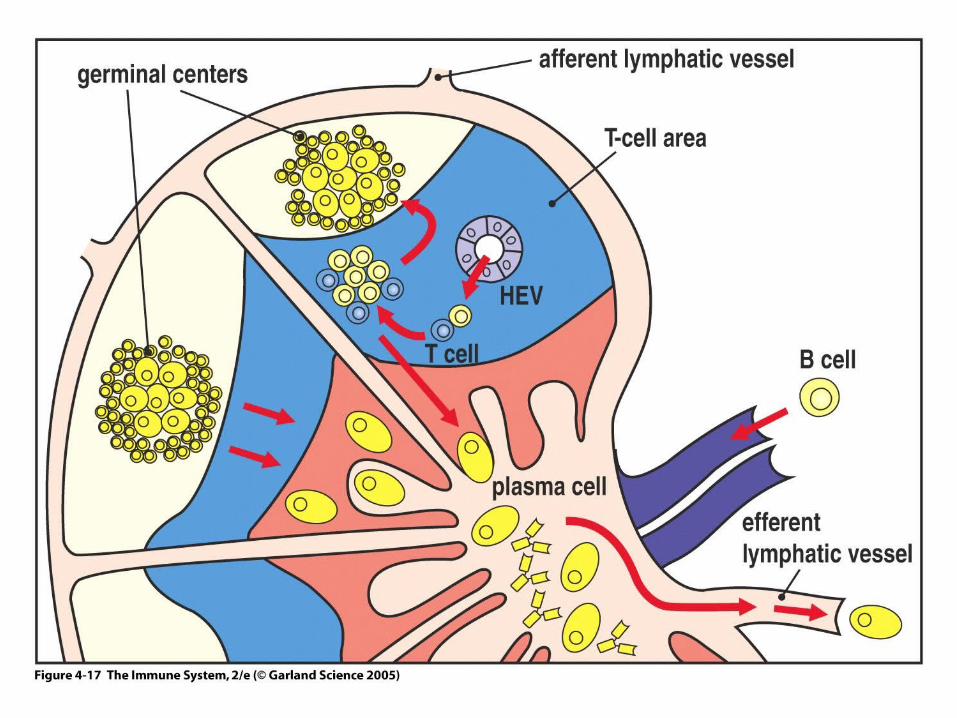
Figure 4-17
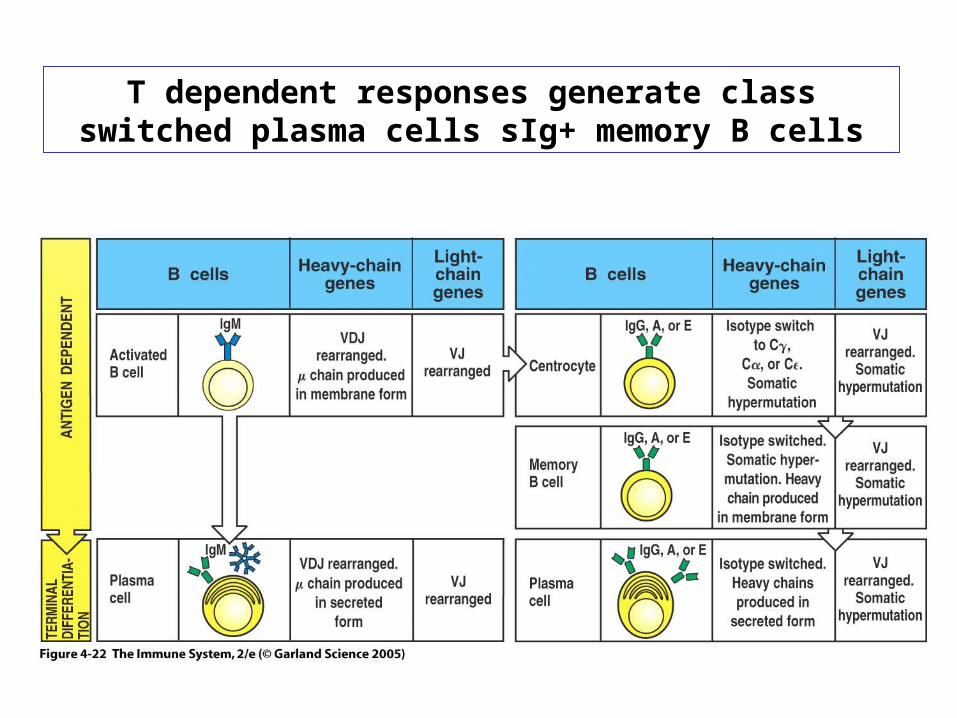
T dependent responses generate class switched plasma cells sIg+ memory B cells
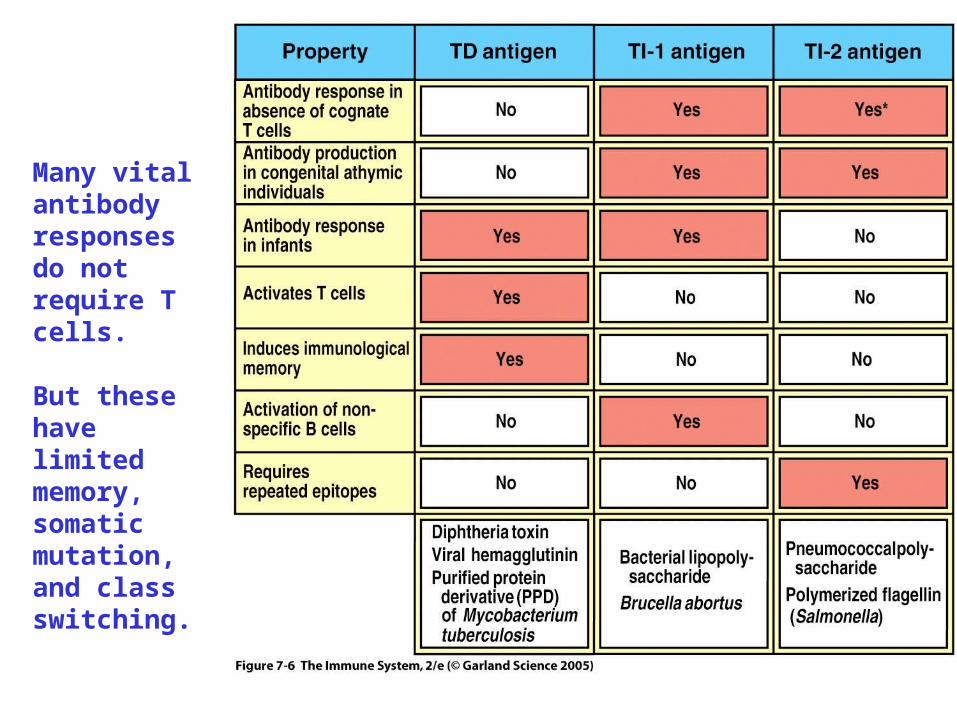
Figure 7-6Many vital antibody responses do not require T cells.
But these have limited memory, somatic mutation, and class switching.
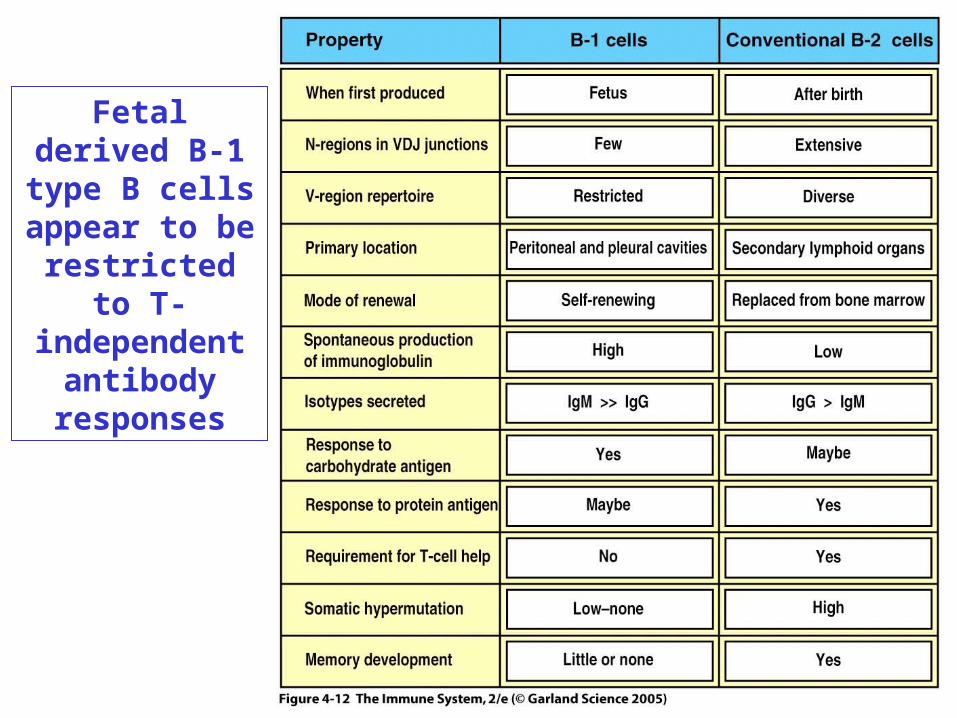
Figure 4-12Fetal derived B-1 type B
cells appear to be restricted to T-independent
antibody responses
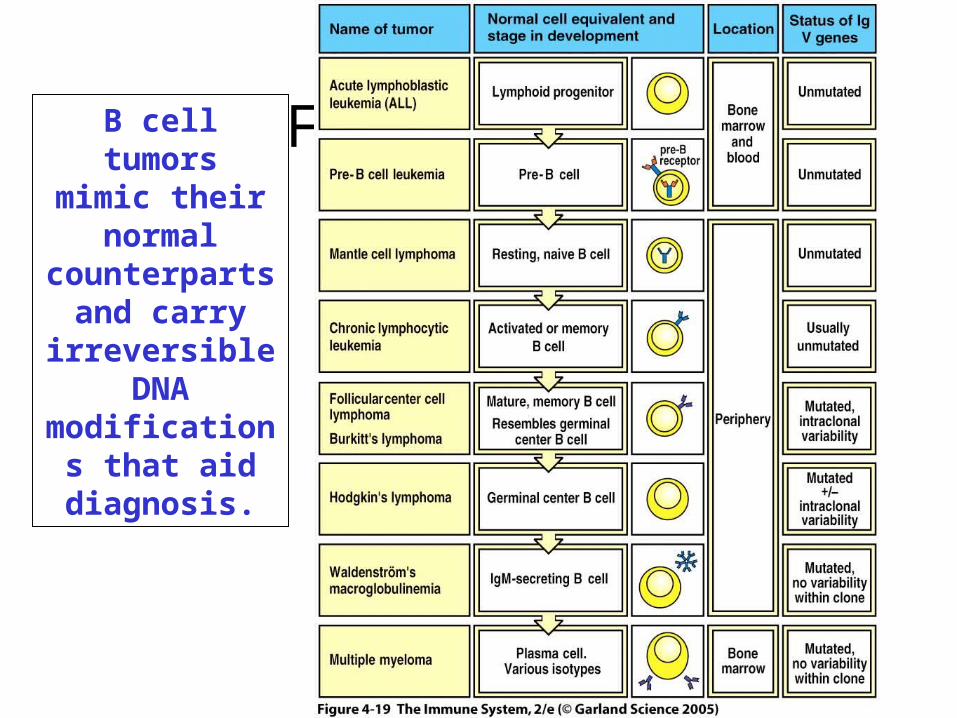
Figure 4-19B cell tumorsmimic their
normal counterparts
and carry irreversible
DNA modifications
that aid diagnosis.
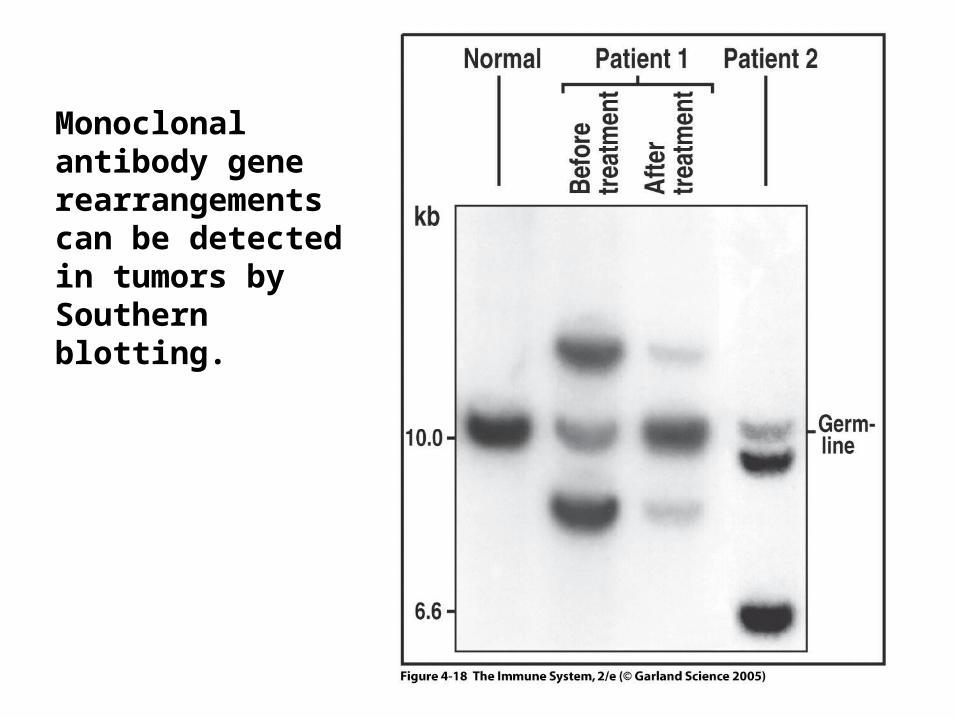
Monoclonal antibody gene rearrangements can be detected in tumors by Southern blotting.
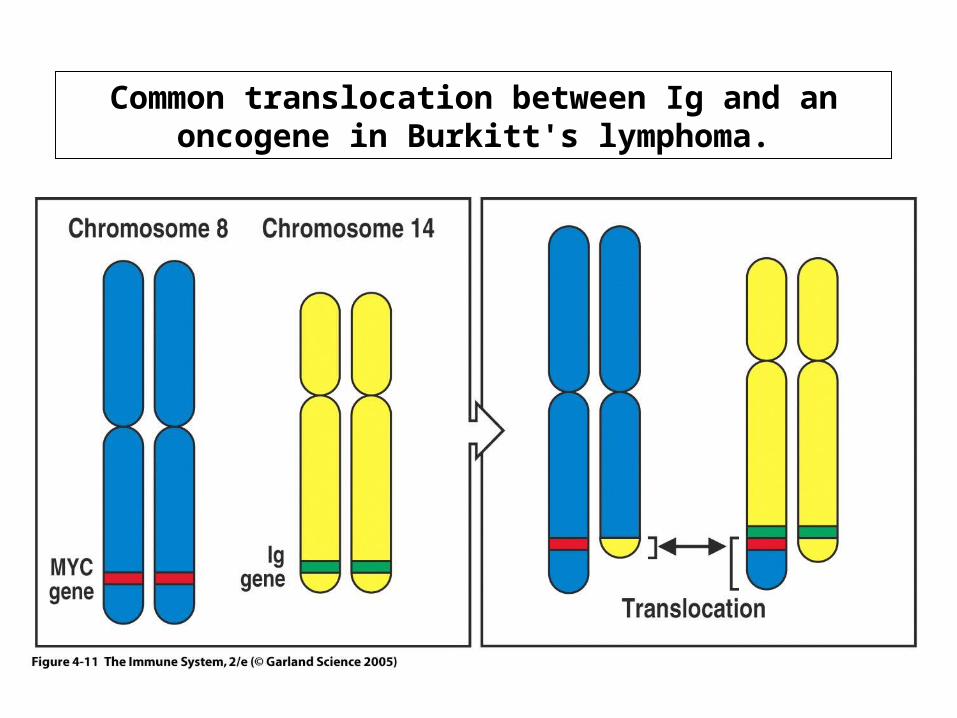
Common translocation between Ig and an oncogene in Burkitt's lymphoma.

Modern drug research and development (duration ~10 years):•random screening of millions of compounds in bioassay
to find initial candidates•refining candidates based on minor substitutions and selection
for improved affinity and specificity•toxicity and efficacy trials
The antibody response (duration ~14 days):•select a few thousand (or fewer) cells among millions of B cells•point mutate and select to develop highest affinities*
*Important for recognition of microbes that mutate rapidly•class switch to appropriate effector class
End result: a long lived, high affinity antibody in circulationAntibody molecules now behave like preexisting innate immune
molecules, allowing an instantaneous response to the specific microbe.
Note: a high affinity antibody is effective at lower dose, increasing the lifespan of memory

Summary of lecture 6
• Molecular mechanisms of class switching and somatic hypermutation. AID and DNA repair enzymes.
• Mismatch repair system is involved in point mutation.• Double strand break repair is involved in class switch.• CD4 T cells are important for class switch regulation
and the initiation of somatic mutation.• The T cell dependent antibody response requires cell
migration to bring T and B cells together.• Somatic mutation happens in the germinal center.• Selection in the germinal center favors high affinity
cells.• Plasma cells and memory B cells are formed during the
T dependent antibody response.
Next time: Mid term test
Review session Saturday Jan 22
4:00-6:50PM YORK 2722





























