Embed Size (px)
Citation preview

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 1/42
Protein Structure-Function #6
Determination of Protein Structure
and Protein Engineering

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 2/42
X-Ray Crystallography

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 3/42
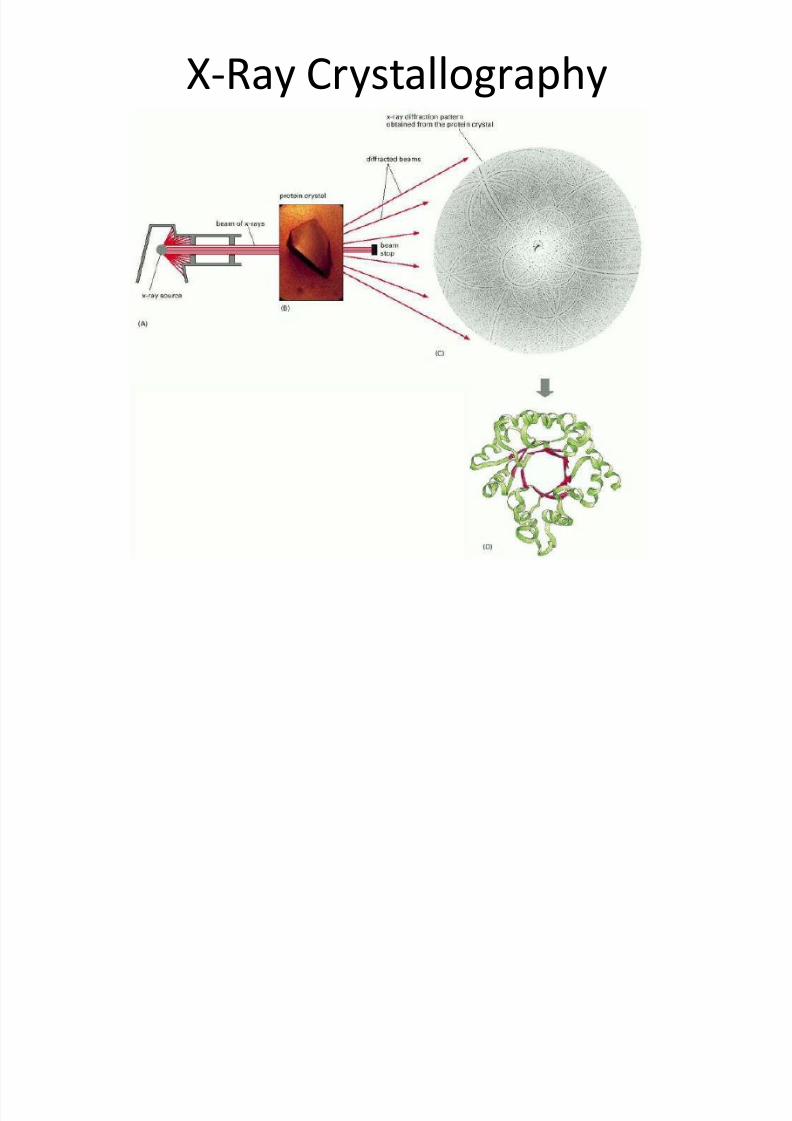
• X-rays are a form ofelectromagnetic radiation with awavelength typically around 0.1nm(the diameter of a hydrogen atom).
• If a narrow parallel beam of x-raysis directed at a sample of pure
protein, most of the x-rays passstraight through it.
• A small fraction will however bescattered by the atoms in the
sample. If the sample is a wellordered crystal, the scatteredwaves reinforce one another atcertain points and appear asdiffraction spots when the x-rays
are recorded by a suitable detector.
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 4/42
X-Ray Crystallography

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 5/42
• The position and intensity of eachspot contains information about thelocations of the atoms within thecrystal that gave rise to it.
• Deducing the three dimensionalstructure of a large molecule formthe diffraction pattern is acomplicated task and was not
achieved for a protein moleculeuntil the 1960s.
• An electron density map is producedfrom the overall diffraction patternusing a mathematical techniquecalled a Fourier transform.
• A model for the structure is thenbuilt that is consistent with theelectron-density map.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 6/42
Problems•
In recent years X-ray diffraction has become increasinglyautomated, however many problems still remain withthe technique.
• The slowest step is often the generation of suitable
crystals.
• This requires large amounts of very pure protein andoften involves many years of trial and error, searching for
the proper crystallization conditions.
• There are still many proteins, in particular membraneproteins, which have so far resisted all attempts tocrystallize them.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 7/42
• The physical environment of the crystal is notidentical with that within a living cell.
• A crystal imposes a space and time average onthe structure deduced from the analysis and
thus provides little information about molecularmotion within the protein (e.g. calmodulin).
• The conformation of the molecules within thecrystal can also be affected by nonphsysiologicalfactors such as incidental protein-proteininteractions.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 8/42
•
Structures derived through this methodshould always be confirmed by comparingstructural data acquired by other means.
• Also mobile portions of the protein aredifficult to determine as they may have adifferent confirmation in different proteinmolecules within the crystal.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 9/42
Nuclear Magnetic Resonance (NMR)Spectroscopy
• This technique depends on the fact that certain atomicnuclei are intrinsically magnetic.
• The simplest example is the hydrogen nucleus (1H), which
is a proton.
• The spinning of a proton generates a magnetic moment.This moment can take either of two orientations, or spinstates (called α and β), when an external magnetic field is
applied.
• The energy difference between these states is proportionalto the strength of the imposed magnetic field.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 10/42
• The α state has a slightly lower energy and henceis slightly more populated (by a factor of the orderof 1.00001 in a typical experiment) because it isaligned with the field.
• A spinning proton in an α state can be raised to anexcited state (β state) by applying a pulse of
electromagnetic radiation (a radio-frequency, orRF, pulse), provided the frequency corresponds tothe energy difference between the α and the βstates.
• In these circumstances, the spin will change fromα to β; in other words, resonance will be obtained.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 11/42
• A resonance spectrum for a molecule can beobtained by varying the magnetic field at aconstant frequency of electromagneticradiation or by keeping the magnetic fieldconstant and varying electromagnetic radiation.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 12/42
• These properties can be used to examine the chemicalsurroundings of the hydrogen nucleus.
• The flow of electrons around a magnetic nucleusgenerates a small local magnetic field that opposes theapplied field.
• The degree of such shielding depends on the surroundingelectron density. Consequently, nuclei in differentenvironments will change states, or resonate, at slightlydifferent field strengths or radiation frequencies.
• The nuclei of the perturbed sample absorbelectromagnetic radiation at a frequency that can bemeasured.
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 13/42
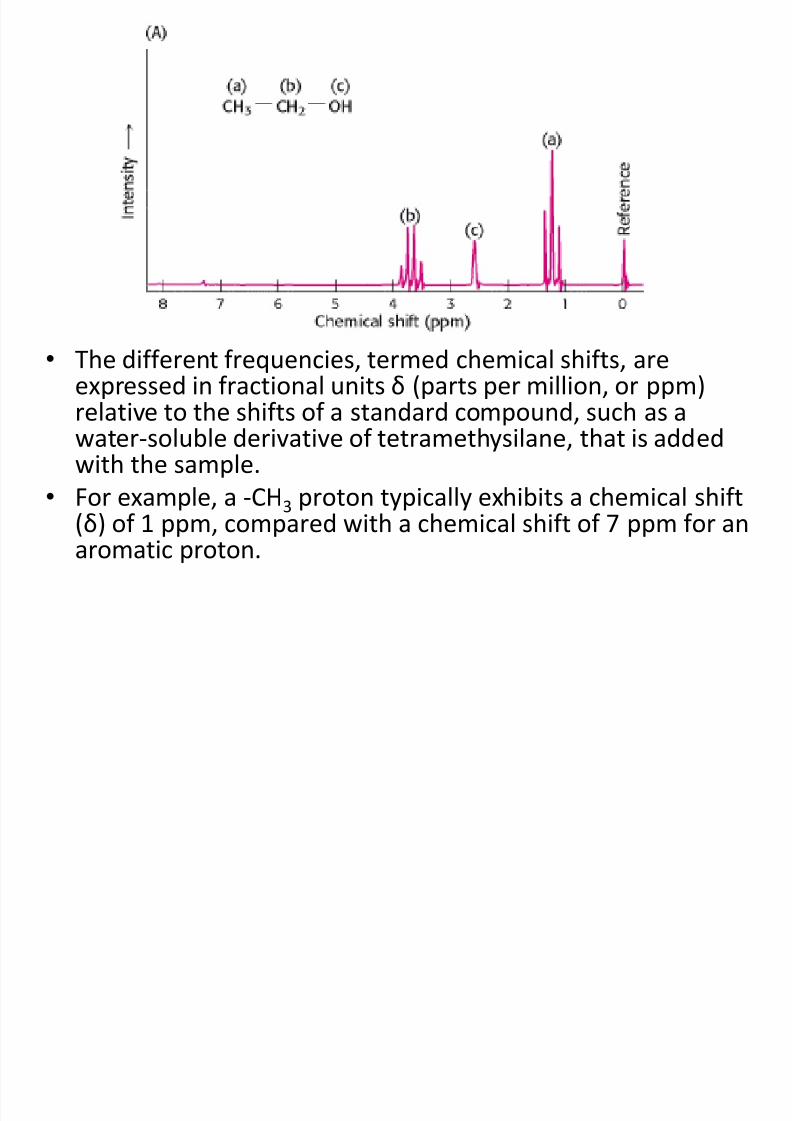
• The different frequencies, termed chemical shifts, areexpressed in fractional units δ (parts per million, or ppm)
relative to the shifts of a standard compound, such as awater-soluble derivative of tetramethysilane, that is addedwith the sample.
• For example, a -CH3 proton typically exhibits a chemical shift(δ) of 1 ppm, compared with a chemical shift of 7 ppm for an
aromatic proton.
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 14/42
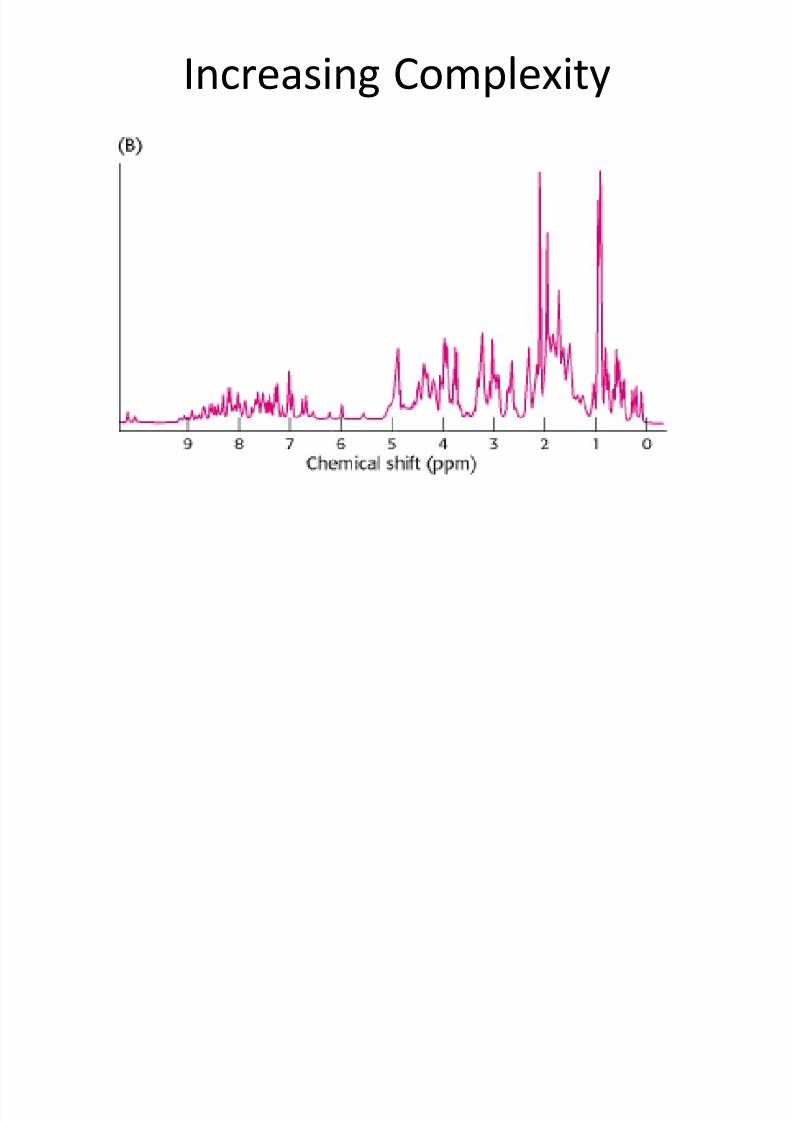
Increasing Complexity

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 15/42
• It is possible to resolve most protons in manyproteins by using this technique of one-dimensional NMR.
•
With this information, we can then deducechanges to a particular chemical group underdifferent conditions, such as theconformational change of a protein from adisordered structure to an α helix in response
to a change in pH.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 16/42
NOESY
• We can garner even more information byexamining how the spins on different protonsaffect their neighbors.
• By inducing a transient magnetization in asample through the application a radio-
frequency pulse, it is possible to alter the spinon one nucleus and examine the effect on thespin of a neighbouring nucleus.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 17/42
NOESY
• Especially revealing is a two-dimensionalspectrum obtained by Nuclear OverhauserEnhancement Spectroscopy (NOESY), which
graphically displays pairs of protons that are inclose proximity, even if they are not closetogether in the primary structure

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 18/42
• The basis for this technique is the nuclear Overhausereffect (NOE), an interaction between nuclei that isproportional to the inverse sixth power of the distance
between them.
• Magnetization is transferred from an excited nucleus toan unexcited one if they are less than about 5 Å apart.
• In other words, the effect provides a means of detectingthe location of atoms relative to one another in thethree-dimensional structure of the protein. The diagonalof a NOESY spectrum corresponds to a one-dimensionalspectrum. The off-diagonal peaks provide crucial newinformation: they identify pairs of protons that are lessthan 5 Å apart
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 19/42
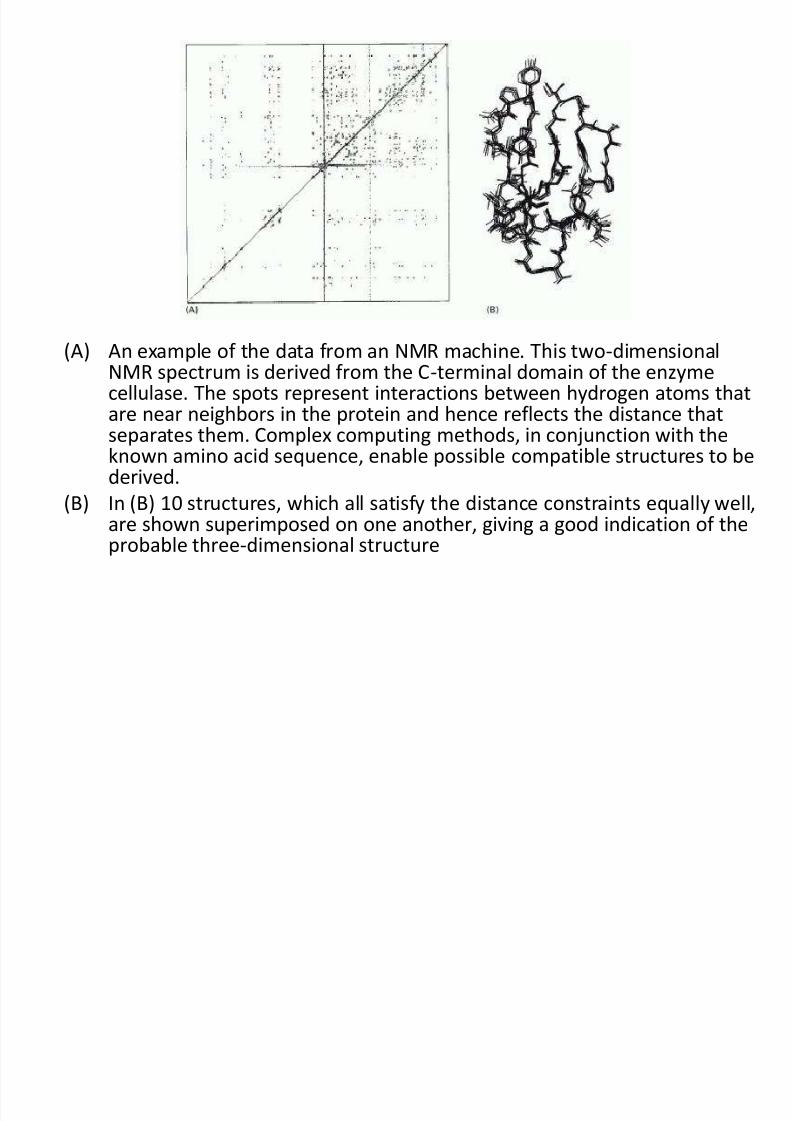
(A) An example of the data from an NMR machine. This two-dimensionalNMR spectrum is derived from the C-terminal domain of the enzymecellulase. The spots represent interactions between hydrogen atoms that
are near neighbors in the protein and hence reflects the distance thatseparates them. Complex computing methods, in conjunction with theknown amino acid sequence, enable possible compatible structures to bederived.
(B) In (B) 10 structures, which all satisfy the distance constraints equally well,are shown superimposed on one another, giving a good indication of the
probable three-dimensional structure

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 20/42
Why So Many Structures?
• A family of related structures is generated for three reasons.
• First, not enough constraints may be experimentallyaccessible to fully specify the structure.
• Second, the distances obtained from analysis of the NOESYspectrum are only approximate.
•
Finally, the experimental observations are made not on singlemolecules but on a large number of molecules in solutionthat may have slightly different structures at any givenmoment. Thus, the family of structures generated from NMRstructure analysis indicates the range of conformations forthe protein in solution.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 21/42
Problems?
• At present, NMR spectroscopy can determinethe structures of only relatively small proteins(<40 kd), but its resolving power is certain to
increase.
• The power of NMR has been greatly enhanced
by the ability to produce proteins labeleduniformly or at specific sites with 13C, 15N, and2H with the use of recombinant DNAtechnology

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 22/42
Circular Dichroism Spectroscopy
Protein dynamics
andsecondary structuredetermination
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 23/42
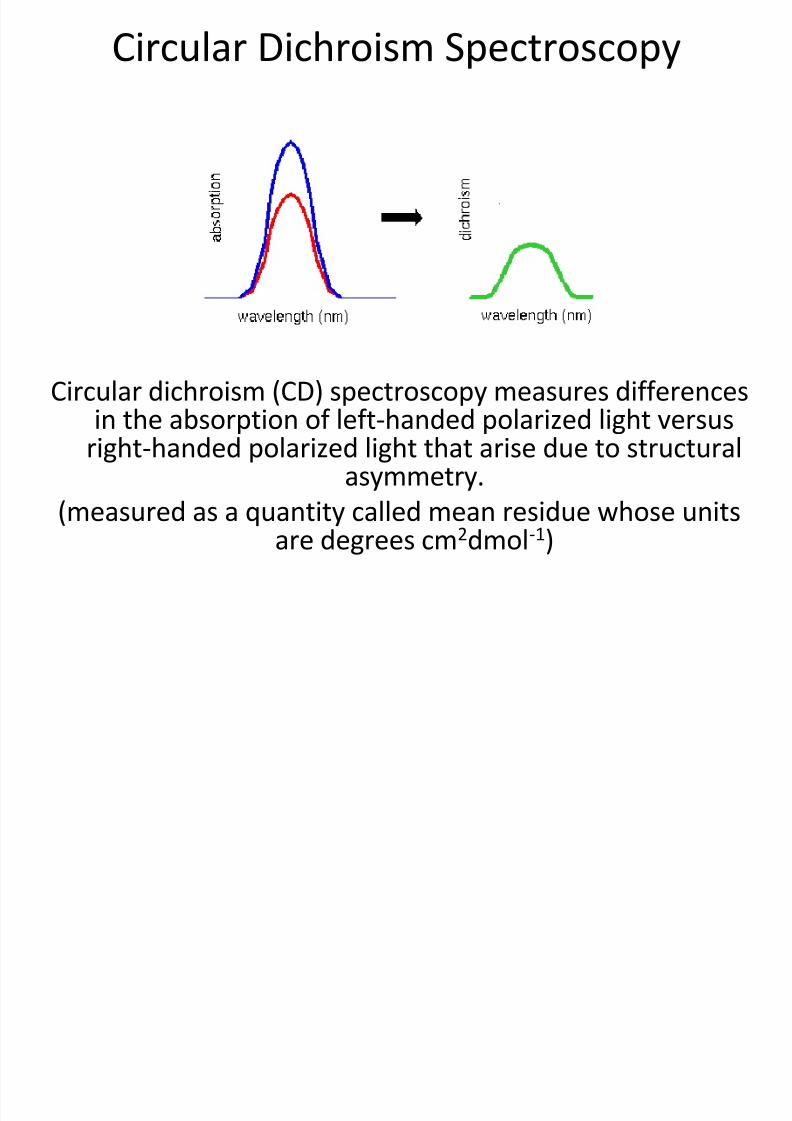
Circular Dichroism Spectroscopy
Circular dichroism (CD) spectroscopy measures differences
in the absorption of left-handed polarized light versusright-handed polarized light that arise due to structuralasymmetry.
(measured as a quantity called mean residue whose unitsare degrees cm2dmol-1)

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 24/42
CD Spectroscopy•
The absence of regular structure results in zero CDintensity, while an ordered structure results in a spectrumwhich can contain both positive and negative signals.
• Chiral or asymmetric molecules produce a CD spectrumbecause they absorb left and right handed polarized lightto different extents and are thus considered to beoptically active.
• Biological macromolecules such as proteins and DNA arecomposed of optically active elements and because theycan adopt different types of 3D structures, each type ofmolecule produces a distinct CD spectrum.
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 25/42
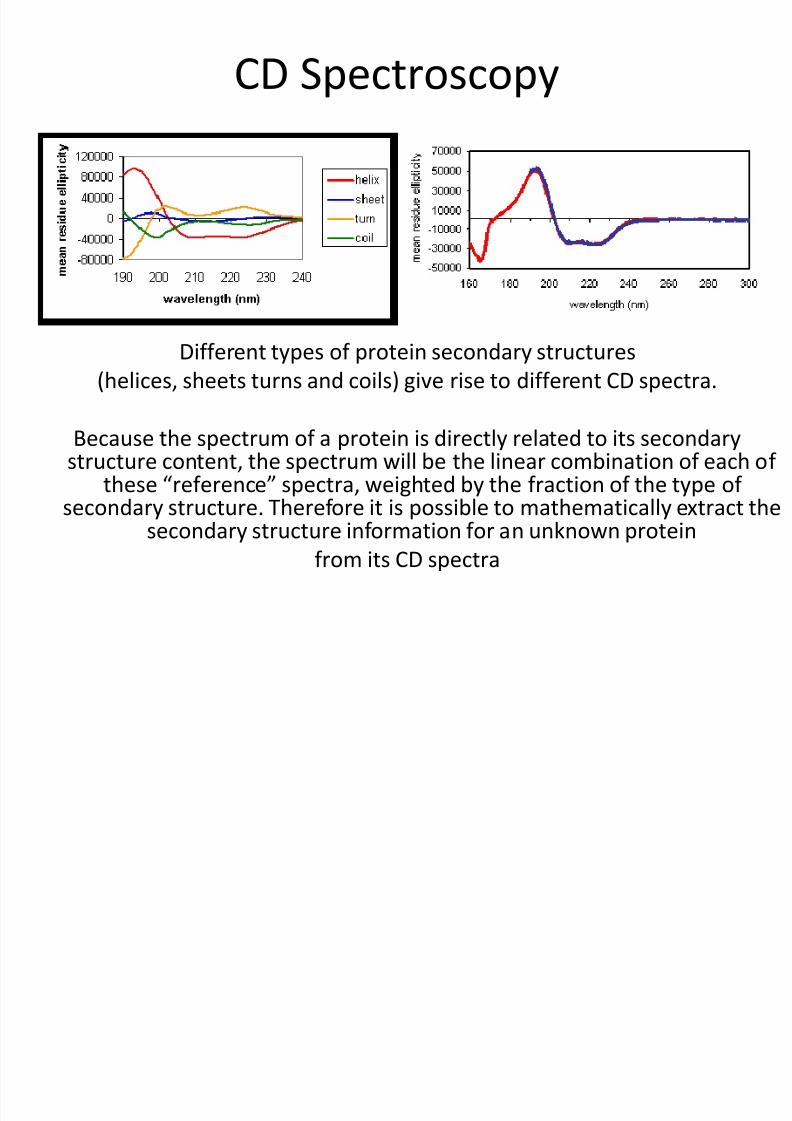
CD Spectroscopy
Different types of protein secondary structures(helices, sheets turns and coils) give rise to different CD spectra.
Because the spectrum of a protein is directly related to its secondarystructure content, the spectrum will be the linear combination of each of
these “reference” spectra, weighted by the fraction of the type ofsecondary structure. Therefore it is possible to mathematically extract the
secondary structure information for an unknown protein
from its CD spectra

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 26/42
CD Spectroscopy Has Been Used toMonitor
• Secondary structure
• Conformational changes
• Environmental effects
• Protein folding and denaturation
• Dynamics
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 27/42
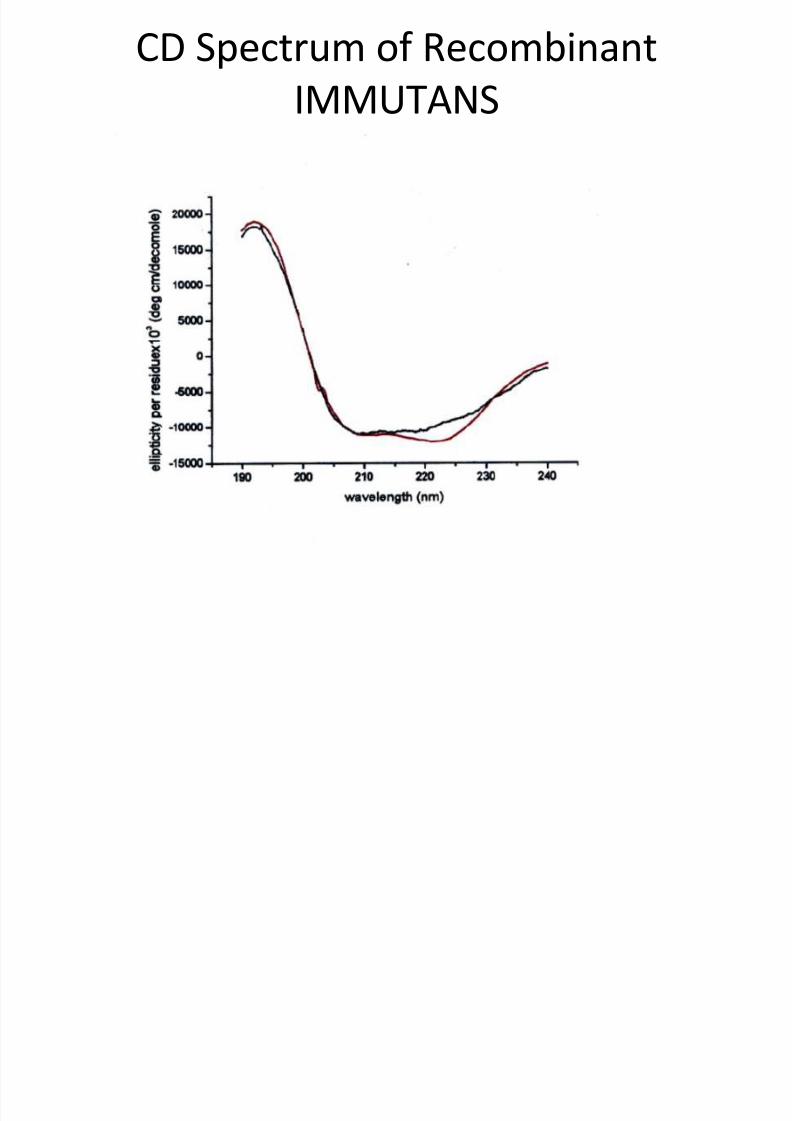
CD Spectrum of RecombinantIMMUTANS

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 28/42
• Some of the advantages of CD spectroscopyare that it requires only very small amounts ofprotein (100mg or less)
• and measurement can be done very quickly(30 mins or less).
• But this technique will not allow the 3Dstructure to be determined.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 29/42
• Single Particle Cryo- EM
• Visualizing biological
complexes such as viruses,small organelles, andmacromolecular biologicalcomplexes of 200 kDa or
larger preserved in non-crystaline ice

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 30/42
Cryo-EM analysis 31 Å resolution

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 31/42

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 32/42
GroEL-GroES Complex (Cpn60)
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 33/42
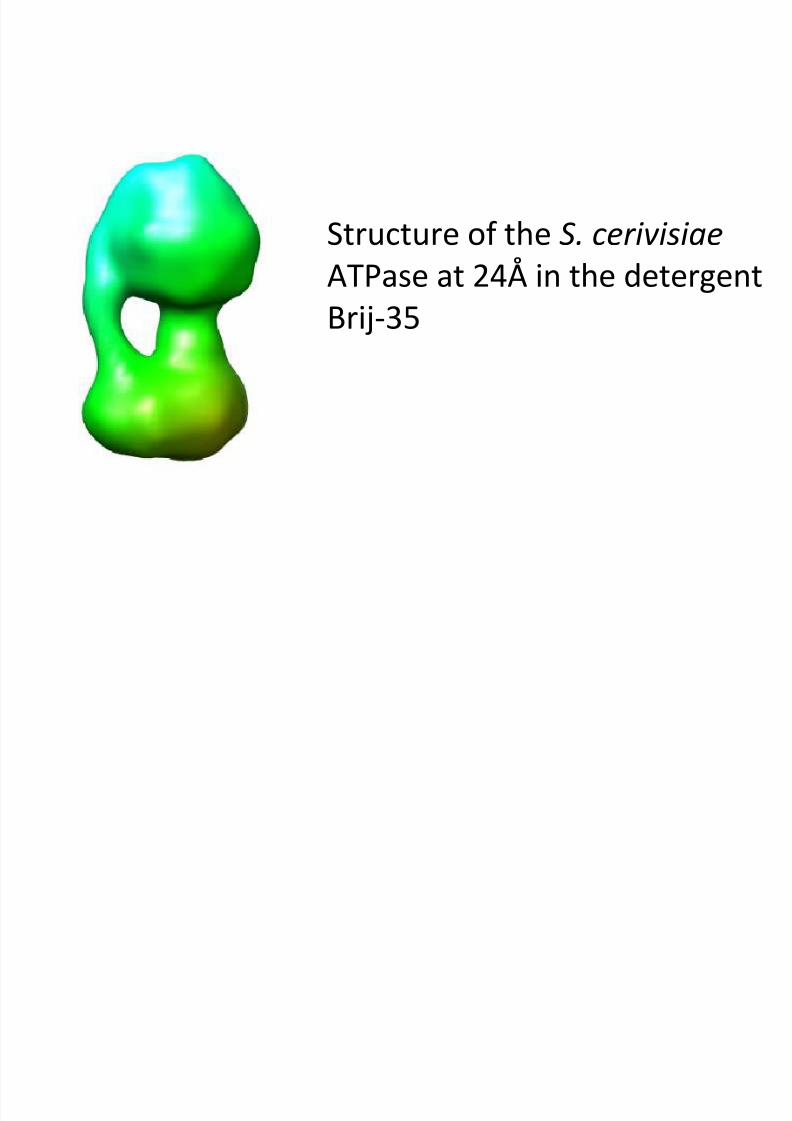
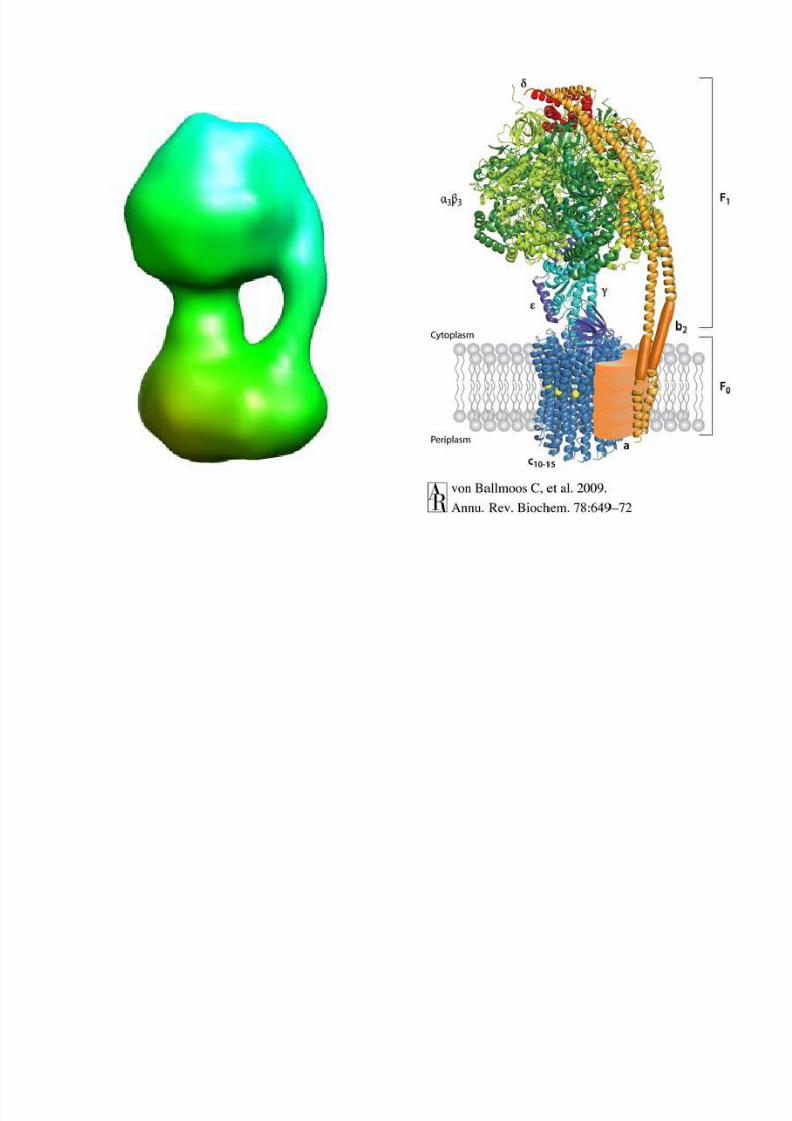
Structure of the S. cerivisiae
ATPase at 24Å in the detergentBrij-35
8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 34/42

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 35/42
Protein Engineering
1. Dissection of the structure and activity of existing proteinsby making systematic alterations of their structures andexamining the changes in their properties.
2. Production of novel proteins for use in medicine andindustry.
3. We do not know how to design functional protein de novo,but we can make minor alterations of existing proteins to
give useful changes in activity. We can also cut and pastesegments of one protein to another. A good example is thehumanization of monoclonal antibodies by grafting themouse antigen-binding loops onto human frame workallowing use in cancer treatments.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 36/42
• The prerequisite for protein engineeringstudies is that the enzyme has been cloned anexpressed.
• Further, unless only relatively crudeinformation is required, it is essential that the
structure has been solved at high resolution

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 37/42
Choice of Mutation
• Choose a mutation that deletes part of a side chain or leads toan isosteric change. Deletions are preferred to mutations thatincrease the size of the side chain, especially in the interior of aprotein or at an enzyme-substrate interface. Any increase involume of the side chain is liable to distort the structure of the
enzyme whereas small changes in the enzyme cavity can betolerated.
• Avoid creating buried unpaired charges. Removal of a groupthat solvates a buried charge is dangerous. Solvation energies
of ions are so high that charged groups must be solvated.Unless there is open access of solvent to buried chargeresidues, the structure of mutant proteins will rearrange sothat there is solvation by water or other groups on the enzyme.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 38/42
Choice of Mutation
• Delete the minimal number of interactions. It isdifficult enough to analyze the change in justone interaction. Avoid the deletion of multiple
interactions where possible.
• Do not add new functional groups to sidechains. The addition of a new group on a sidechain can cause local reorganization ofstructure if a new group can make novelinteractions

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 39/42
All previous rules may be
disobeyed when appropriate!

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 40/42
Choice of Mutation
• The ideal mutation is a nondisruptive deletion,which is one which simply removes aninteraction without causing a disruption or
reorganization of a structure.
• When in doubt mutate to alanine. This isgenerally a deletion mutation and alanine hasfew quirks. Glycine, because of it widerfreedom of conformation can have odd effectson structure

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 41/42
Random Mutagenesis
• The process of random mutagenesis is alsouseful in the study of protein structurefunction.
• Random mutations may be induced by PCR orby chemical mutagenesis. This process can beuseful in identifying important residues whendetailed structural information is notavailable.

8/13/2019 Lecture 6 - Protein Structure-Function 6
http://slidepdf.com/reader/full/lecture-6-protein-structure-function-6 42/42
Mutagenesis may also be performed atthe gross level
• Instead of changing a single amino acid residue,whole sections of the protein may be removed byexcising a stretch of the gene. This is termeddeletion mutagenesis.
• Domains of proteins may be spliced together togive novel proteins.
• Groups can also be added to proteins to act astags or as aids in purification process e.g. His tagsor thioredoxin.





























