Embed Size (px)
DESCRIPTION
Lessons Learned from the Microbial Analysis ofthe Herschel Spacecraft during Assembly,Integration, and Test Operations
Citation preview

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/259209683
LessonsLearnedfromtheMicrobialAnalysisoftheHerschelSpacecraftduringAssembly,Integration,andTestOperations
ARTICLEinASTROBIOLOGY·DECEMBER2013
ImpactFactor:2.59·DOI:10.1089/ast.2013.1024·Source:PubMed
CITATIONS
7
READS
48
9AUTHORS,INCLUDING:
ChristineMoissl-Eichinger
MedicalUniversityofGraz
68PUBLICATIONS956CITATIONS
SEEPROFILE
AlexanderJosefProbst
UniversityofCalifornia,Berkeley
39PUBLICATIONS1,313CITATIONS
SEEPROFILE
MichaelaStieglmeier
Ludwig-Maximilians-UniversityofMunich
15PUBLICATIONS499CITATIONS
SEEPROFILE
PetraRettberg
GermanAerospaceCenter(DLR)
320PUBLICATIONS2,472CITATIONS
SEEPROFILE
Availablefrom:AlexanderJosefProbst
Retrievedon:21March2016

Research Article
Lessons Learned from the Microbial Analysisof the Herschel Spacecraft during Assembly,
Integration, and Test Operations
Christine Moissl-Eichinger,1 Rudiger Pukall,2 Alexander J. Probst,1 Michaela Stieglmeier,1,*
Petra Schwendner,1,3 Maximilian Mora,1 Simon Barczyk,3 Maria Bohmeier,3 and Petra Rettberg3
Abstract
Understanding microbial diversity in spacecraft assembly clean rooms is of major interest with respect to planetaryprotection considerations. A coordinated screening of different clean rooms in Europe and South America bythree German institutes [Deutsches Zentrum fur Luft- und Raumfahrt (DLR), Leibniz-Institut DSMZ-DeutscheSammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), and the Institute of Microbiology and ArchaeaCenter, University of Regensburg] took place during the assembly, test, and launch operations of the Herschelspacecraft in 2006–2009. Through this campaign, we retrieved critical information regarding the microbiome withinthese clean rooms and on the Herschel spacecraft, which served as a model for upcoming ESA mission prepara-tions. This ‘‘lessons learned’’ document summarizes and discusses the data we obtained during this samplingcampaign. Additionally, we have taken the opportunity to create a database that includes all 16S rRNA genesequences ever retrieved from molecular and cultivable diversity studies of spacecraft assembly clean rooms tocompare the microbiomes of US, European, and South American facilities. Key Words: Planetary protection—Spacecraft assembly facility—Microbial diversity—Herschel. Astrobiology 13, xxx–xxx.
1. Introduction
Assessing the microbial diversity in clean room facilitiesis of importance in many disciplines. In pharmaceuti-
cal environments, the presence of potential pathogens mustbe monitored, whereas space agencies are concerned aboutpossible contamination of spacecraft. Such contamina-tion could result in the transfer of Earth microbes to other(potentially habitable) planets and interference in future life-detection missions through microbial contamination (plan-etary protection; for a review see Crawford, 2005).
A great deal of time and effort on the part of spaceagencies is, and has been, necessary to analyze the micro-biome of spacecraft and associated clean rooms with regardto planetary protection. These efforts have recently provokeddebate over the possibility of overprotecting Mars (Fairenand Schulze-Makuch, 2013). The authors claim that Mars hasalready been contaminated with Earth life in the past (byeither meteorites or spacecraft); thus, current planetary pro-tection efforts could be significantly scaled back for financialand scientific reasons. However, the authors of the afore-
mentioned article do not address the problem of false-positive or false-negative life-detection results that couldpossibly be obtained by using life-detection instrumentscontaminated by Earth life signatures. Such a contaminationcould therefore tremendously affect the ongoing life-detec-tion mission and possibly lead to misinterpretations con-cerning subsequent science-driven missions or even tomission failure (Conley and Rummel, 2013).
Thus, analyzing and interpreting the microbiome ofspacecraft-associated environments is crucial for life-detection mission success.
Microbial contamination in and around spacecraft hasbeen assessed with a variety of cultivation-based (La Ducet al., 2007; Stieglmeier et al., 2009, 2012; Moissl-Eichingeret al., 2012) and cultivation-independent methods (Venkat-eswaran et al., 2001; La Duc et al., 2009; Cooper et al., 2011;Vaishampayan et al., 2013a). Although the results from mi-crobial analyses are strongly dependent on time, location,method, and other factors (clean room maintenance, missionstatus, etc.), these studies have revealed a vast diversity ofmicroorganisms in clean rooms. These organisms are mostly
1Institute of Microbiology and Archaea Center, University of Regensburg, Regensburg, Germany.2Leibniz-Institut DSMZ—Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany.3German Aerospace Center (DLR), Institute of Aerospace Medicine, Cologne, Germany.*Present address: Department of Genetics in Ecology, Vienna, Austria.
ASTROBIOLOGYVolume 13, Number 12, 2013ª Mary Ann Liebert, Inc.DOI: 10.1089/ast.2013.1024
1

human-associated, but hardy (spore-forming) bacteria arefrequently detected in close proximity to and on spacecraft.Spacecraft and clean room contamination by these organismsposes a potential threat to planetary protection.
NASA has a long tradition of implementing planetary pro-tection (Puleo et al., 1977), and NASA’s endeavors in bioburdenassessment have followed established protocols [NASA stan-dard procedures for the microbial examination of space hard-ware (NASA-HBBK-6022, 2007)] that focus on heat-shock-surviving (spore-forming) microbes. The number of colony-forming units that appear after such a heat shock (e.g., 80�C for15 min) offers an indication as to the overall cleanliness of aspacecraft and represents the microbial load (‘‘bioburden’’).ESA has adopted many of these protocols but has also estab-lished other methods to improve detection of possible contam-inants (ECSS-Q-ST-70-55 Working Group, 2008; Probst et al.,2010b).
Currently, ESA is preparing the ExoMars mission, which isscheduled for launch in two stages in 2016 and 2018. Duringthe preparation phase of this mission, optimal procedures arebeing chosen to analyze bioburden and biodiversity in facili-ties and on spacecraft. Therefore, ESA announced three pro-jects that aimed at the understanding of microbial diversity inclose vicinity of a spacecraft: Determination of the MicrobialDiversity of Spacecraft Assembly and Testing Facilities (Bio-Div, main contractor: DLR, Cologne; subcontractor: DSMZ,Braunschweig), Archaeal and Specific Bacterial Communitiesin Spacecraft Associated Clean Rooms (main contractor: Uni-versity of Regensburg), and Characterization, ControlledStorage, and Publication of Planetary Protection AssociatedBacterial Isolates (main contractor: DSMZ). Together, theseprojects will help to elucidate the microbial bioburden andbiodiversity in European and South American facilities beforethe ExoMars mission is implemented. During these surveys,isolation strategies led to the identification of numerous mi-crobial strains that are representative of the clean room di-versity. These isolates were collected, archived, and released tothe public through the creation of a microbial culture collec-tion, maintained by the Leibniz Institute DSMZ in Braunsch-weig, Germany (Moissl-Eichinger et al., 2012).
As a model spacecraft for future missions, the Herschelspace telescope was followed through different locations inEurope and South America and sampled under assembly,integration, and testing activities in each facility until thelaunch from Kourou, French Guiana. Although Herschel wasnot subject to biocleanliness control (it represents planetaryprotection category 1 according to COSPAR; COSPAR,2011), the housing clean rooms were operated as ISO 8 andISO 5 (ISO 146441, www.iest.org), which ensured a low levelof particulate and microbial contamination that simulatedcomparable conditions during the assembly, test, and launchoperations for ExoMars. Herschel is a large ESA space tele-scope that covers the spectral range from far-infrared tosubmillimeter wavelengths during its observations of distantobjects in the Universe. The Herschel telescope was laun-ched, together with the Planck spacecraft, from Kourou,French Guiana, in 2009. It reached its point of destination(Lagrange Point L2) and was employed to deliver spectraldata that cannot be observed from Earth (Lis et al., 2012).
Some data collected through this project (‘‘Herschel mis-sion’’) have previously been described (Stieglmeier et al.,2009, 2012), as has the above-mentioned culture collection
(Moissl-Eichinger et al., 2012). Another publication with respectto the microbial diversity detected in the spacecraft assemblyfacilities in Kourou can be found in this issue (Schwendner et al.,2013). The article summarizes and discusses the data that wereobtained in order to provide a ‘‘lessons learned’’ document,which could serve as an information basis for the design offuture sampling campaigns as well as a source for the com-parison with data from other clean room facilities. In addition,we also took this opportunity to create a database that includesall (molecular and isolates’) near full-length 16S rRNA genesequences retrieved and published from any spacecraft as-sembly facility. This will provide a valuable resource for futuresurveys that include comparative analyses of previous culti-vation and molecular data.
2. Materials and Methods
2.1. Description of the sampling campaign
The microbial analysis of the Herschel space telescope andits housing facilities took place from 2006 to 2009 at differentlocations in Europe and South America. During this timeframe, five sampling events were performed, and the micro-bial community was analyzed within the framework of thethree aforementioned projects. The BioDiv project aimed tofollow the European Cooperation for Space Standardiza-tion (ECSS) standard of ESA for the cultivation of heat-shock-resistant microbes (bioburden determination; ECSS-Q-ST-70-55 Working Group, 2008) from surfaces, as it isrecommended by space agencies for the estimation of con-tamination by predominantly spore-forming microbes. Thisincluded the establishment of the ‘‘milliflex-assay’’ as a rapidmethod to detect colonies within 7 h. Additionally, the BioDivproject focused on the overall diversity that was cultivableby applying R2A medium (to obtain vegetative organisms,‘‘vegetatives’’). Air sampling was also performed to under-stand the distribution of microbes within the clean room.
The project designated Archaeal and Specific BacterialCommunities in Spacecraft Associated Clean Rooms applied 32different cultivation conditions in addition to the standardprocedure (‘‘alternative’’ cultivation methods) to investigate abroader spectrum of extremotolerant microbes. Extreme con-ditions with respect to temperature, pH, salt content, oxygenavailability, and nutrient content were used to cultivate tolerantmicrobes and understand the requirements of a clean roomcommunity. The conditions applied were chosen with respect topossible survivability in space, so that media were also usedthat provided conditions for autotrophic or diazotrophic mi-crobes (primary producers). Additionally, molecular analyseswere applied to facilitate understanding of the entire microbialcommunity (uncultivable majority plus cultivable minority),which included quantitative PCR (qPCR) analysis as well ascloning of 16S rRNA gene sequences from Bacteria and Ar-chaea. The resulting isolates were organized and archived at theDSMZ in Braunschweig (Moissl-Eichinger et al., 2012).
2.2. Sampling and sample extraction
Five samplings were performed in three different space-craft assembly clean room facilities, one in Germany, a sec-ond in the Netherlands, and a third in French Guiana. Thefacilities in Germany (Friedrichshafen) were maintained byEADS Astrium (European Aeronautic Defense and Space
2 MOISSL-EICHINGER ET AL.

Company), while the other clean rooms were located at theESA European Space Research and Technology Centre (ES-TEC) in Noordwijk and at the Centre Spatial Guyanais (CSG)in Kourou, maintained by the French space agency CentreNational d’Etudes Spatiales (CNES). More details about thesampling schedule (dates, clean room classes, samplingtypes, etc.) are provided in Table 1. Table 1 also depicts theabbreviations for each sampling event, which are used asfollows: ESTEC samplings: ES1, ES2; Friedrichshafen sam-plings: FR1, FR2; Kourou sampling: KO. The clean roomswere operated as ISO 8 and ISO 5 (ISO 146441, www.iest.org), which corresponds to an upper level of 3,520,000 (max.particle diameter: 0.5 lm), 832,000 (1 lm), 29,300 (5 lm)particles per cubic meter of air (ISO 8) or 100,000 (0.1 lm),23,700 (0.2 lm), 10,200 (0.3 lm), 3,520 (0.5 lm), 832 (1 lm), 29(5 lm) particles per cubic meter of air (ISO 5).
Samples were taken from clean room surfaces that in-cluded mainly floor and ground support equipment (GSE),spacecraft, and clean room air. Throughout the campaign,different sampling tools were used. These included nylon-flocked swabs (552C regular swab, microRheologics, Brescia,Italy), wipes (Spec-Wipe 4, VWR International GmbH,Darmstadt, Germany; 15 cm · 15 cm), and SpongeSicles(Biotrace [3M], St. Paul, MN, USA). In the first sampling(Table 1), witness plates (V2A 1.4301, no. 4 finish, 25 mm ·5 mm · 1 mm) were exposed for 3 days during clean roomoperations. BiSKits (biological sampling kits, QuicksilverAnalytics, Abingdon, MD, USA) were used for the collectionof samples for molecular analyses. Air samples were col-lected with the AirPort MD8 air sampler (Sartorius, Ger-many), which was supplemented with sterile gelatinmembrane filters. Comprehensive descriptions of samplingprocedures and extractions are described elsewhere (Stiegl-meier et al., 2012) and are summarized together with subse-quent cultivation strategies in Table 2.
2.3. Sample processing for bioburden measurementsaccording to ECSS standard documentECSS-Q-ST-70-55C
The sampling and sample processing procedure is out-lined in document ECSS-Q-ST-70-55C (ECSS-Q-ST-70-55
Working Group, 2008) and was also described in detailpreviously (Stieglmeier et al., 2012). In brief, premoistenedwipes were used for sampling and afterward extracted in atotal volume of 40 mL PBST [phosphate-buffered saline so-lution including 0.02% (v/v) Tween 80] by vortexing andsonication. Aliquots were either heat-shocked (15 min, 80�C;‘‘wipe assay E.1 for mesophilic aerobic spores and heat-tolerant bacteria,’’ determination of microbial bioburden) ordirectly processed (‘‘wipe assay E.2 for aerobic mesophilicbacteria,’’ determination of vegetatives) by pour-plating intrypticase soy agar (TSA) and R2A (Becton Dickinson, Hei-delberg, Germany), respectively. All plates were incubatedfor 72 h at 32�C. Colony numbers were monitored every 24 h;colonies representative in size, color, abundance, and shapewere randomly picked for further characterization, purifiedby two subsequent streak-outs, and subjected to 16S rRNAgene sequencing.
2.4. Sample processing for the cultivationof specialized microorganisms (alternative assays)
SpongeSicle samples were extracted either under an-aerobic or aerobic conditions as previously described(Stieglmeier et al., 2009). For the cultivation of specializedmicroorganisms, more than 30 different media and con-ditions were applied as follows. Anaerobes and facultativeanaerobes: anaerobic thioglycollate liquid medium (TG),anaerobic thioglycollate agar (TGA; this medium was usedfor colony counts of anaerobes), anaerobic trypticase soyliquid medium (TS), anaerobic trypticase soy agar (TSA;all media described in Stieglmeier et al., 2009). All anaer-obes were grown under nitrogen atmosphere or mixtureswith hydrogen and carbon dioxide as described previously(Stieglmeier et al., 2009). Diazotrophs were grown inmodified Hino and Wilson N-free medium under nitrogenatmosphere or under a nitrogen atmosphere supplementedwith 1% oxygen. Autotrophs were isolated on autotrophichomoacetogen liquid medium (AHM) and autotrophic all-rounder liquid medium (AAM; Stieglmeier et al., 2009). Formedia focusing on the isolation of archaea, refer to Moissl-Eichinger (2011a). Sulfate-reducing microbes were grownon modified DSMZ 63 agar (Moissl-Eichinger et al., 2012).
Table 1. Sampling Locations, Dates, Clean Room Classes, Number of Samples Taken,
and Number of Isolates Obtained
Sampling Cultivation Sample type Isolates
Date SitesClean roomclass (ISO)1
Cultivationapproach Swabs
Wipes/SpongeSicles2 Air
Witnessplates BiSKits Blanks
No. ofcoloniesobtained
No. ofisolates
processed
No. ofbacterialspecies
12.12.2006 ES1:ESTEC
8 Standard 42 5 15 54 - 6 166 74 20
17.04.2007 FR1:EADS
5 Standard 36 12 6 - - 20 66 4 3Alternative 9 8 - - - 7 1181 158 26
20.11.2007 FR2:EADS
8 Standard 62 18 - - - 15 54 52 12Alternative 9 9 - - 5 12 562 127 18
11.03.2008 ES2:ESTEC
8 Standard 82 16 16 - - 14 91 76 40Alternative 9 4 - - 4 9 1816 216 34
15.04.2009 KO:CSG
8 Standard 5 21 14 - - 5 42 22 11Alternative 8 5 - - 4 5 > 3000 141 49
1Clean room classification according to ISO 14644 (for further details please see Materials and Methods).2Wipes were used for bioburden measurements following the ESA standard; SpongeSicles were used for alternative cultivation analyses.
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 3

Oligotrophic microorganisms were grown on 1:10 and 1:100diluted R2A agar (Moissl-Eichinger et al., 2012). Alkaliphiles/alkalitolerants and acidophiles/acidotolerants were isolated onR2A agar with adjusted pH (pH 9, pH 11, and pH 3.5, re-spectively; Moissl-Eichinger et al., 2012). Halophiles and halo-tolerants were grown on R2A agar containing 3.5% and 10%NaCl (w/v), whereas psychrophilic/psychrotolerant andthermophilic/thermotolerant microbes were isolated on R2Aafter incubation at 4�C, 10�C, 50�C, and 60�C, respectively(Moissl-Eichinger et al., 2012). Microaerophilic/micro-aerotolerant microorganisms were isolated on R2A under areduced oxygen atmosphere [98% nitrogen, 2% O2 (v/v),Moissl-Eichinger et al., 2012]. All incubations were performedat 32�C unless otherwise stated. Colony counts were obtainedfrom agar plates only. Enriched microbes in liquid media weretransferred to agar plates and purified thereon.
2.5. Sample processing of air samples
Air samples, representing 500 L of filtered air, were pro-cessed as described by Moissl-Eichinger et al. (2012).
2.6. Sample processing of witness plates
The witness plates were transferred into sterile tubescontaining 50 mL of rinse solution. The tubes were sonicatedfor 120 s at 35 kHz. R2A plates were inoculated with 5 mL ofthe suspension by the pour-plate technique for mesophilicvegetative bacteria. Platings were performed in triplicate,and plates were incubated for 72 h at 25�C. The colonies werecounted after 24, 48, and 72 h. The remaining suspensionfrom each sample was heat-shocked at 80�C for 15 min, and5 mL was plated on TSA (in triplicate) for heat-tolerantbacteria and spores. These plates were incubated at 32�C,and the colonies were counted after 24, 48, and 72 h.
2.7. Culture collection and taxonomic analyses
Representative isolates were freeze-dried for long-termstorage under ambient conditions. In parallel, cryopreserva-tion of bacterial strains above liquid nitrogen was performedas described (Hippe, 1991). Currently, publicly available bac-terial strains as well as their DSM numbers have been pub-lished by Moissl-Eichinger et al. (2012). The strains can beordered from the DSMZ and will be supplied as freeze-driedcultures in glass ampoules or as actively growing culture onrequest. Detailed information about each strain is accessible atwww.dsmz.de/research/microorganisms/projects/european-space-agency-microbial-strain-collection.html. A list of strains is
also provided at http://www.dsmz.de/catalogues/catalogue-microorganisms/specific-catalogues/esa-strains.html, whichoffers the possibility for selection of specific strains. Thesestrains are also included in the general online catalogue ofthe DSMZ and can easily be found by their DSM number. Allstrains were identified based on 16S rRNA gene sequenceanalysis in which partial or near full-length sequences wereused, as described by Moissl-Eichinger et al. (2012). TheEzTaxon Server containing the 16S rRNA gene sequences oftype strains of prokaryotes was used for similarity-basedidentification (Kim et al., 2012).
2.8. Sample processing of samplesfor molecular analyses
Molecular analysis of samples, including DNA extraction,16S rRNA gene PCR and cloning, sequencing, phylogeneticanalyses, and quantitative 16S rRNA gene PCR, was performedaccording to the instructions of Stieglmeier et al. (2012). Resultshave been published by Stieglmeier et al. (2012) and Schwendneret al. (2013) and are revisited in a comparative manner.
2.9. Database construction and statistical analyses
Bacterial 16S rRNA gene sequences that were generatedin previous publications by cloning (Moissl et al., 2007; LaDuc et al., 2009; Probst et al., 2010a; Vaishampayan et al.,2010; Schwendner et al., 2013; Stieglmeier et al., 2012) andcould be found in public databases (NCBI, Greengenes,5431 sequences, median length 1424 nucleotides; acces-sion numbers: DQ532126–DQ532150; FJ191310–FJ194034;GQ129843–GQ130128; EU704699–EU706281; FJ957429–FJ957855; EU373541–EU888578; HQ434559–HQ434621) wereretrieved and used to generate a database that includedessential metadata. After SINA-alignment (Pruesse et al.,2012), the multiple sequence alignment was fed into Dna-dist (Felsenstein, 1989, 1993) to compute a distance matrixwith Jukes-Cantor correction. The distance matrix was usedfor operational taxonomic unit (OTU) grouping in mothur(Schloss et al., 2009; 2.5% dissimilarity cutoff, furthestneighbor algorithm). In case 16S rRNA gene sequences werenon-overlapping, these were grouped in separate OTUs bychoosing 5% dissimilarity between them. OTU abundanceswere either retrieved directly during this method (giventhat all sequences of a clone library were submitted to apublic database) or were manually added from individualpublications (e.g., Moissl et al., 2007). The abundance ofdetected OTUs was normalized to an artificial total sum to
Table 2. Summary of Sampling and Cultivation/Analysis Strategies
Strategy and procedure Air Bioburden Vegetatives Alternative Molecular analyses
Sampling of: Air Surfaces1 Surfaces1 Surfaces1 Surfaces1
Sampling tool: AirPort air sampler Swabs, wipes Swabs, wipes SpongeSicles BiSKitsSample extraction in: n.a. PBST2 PBST2 PBST2 n.a.Heat shock: - 80�C, 15 min - - -Cultivation: TSA3, 32�C TSA, 32�C R2A, 32�C Various n.a.
Processing of witness plates is not included, since this type of sampling was performed only once.1Mainly floor and ground support equipment.2Phosphate-buffered saline solution including 0.02% (v/v) Tween 80.3Gelatin filter was placed on TSA.n.a., not applicable.
4 MOISSL-EICHINGER ET AL.

ensure their biological/statistical comparability. Hier-archical clustering (based on average-linkage) and non-metric multidimensional scaling (NMDS), both based onBray-Curtis distance measure, were generated in the Rprogramming environment (R Development Core Team,2011) with the aid of the Vegan Package. For comparison ofthe cultivable and molecular diversity, all publicly available16S rRNA gene sequences from spacecraft-associated cleanroom facilities were added to the existing database. The entireset of 16S rRNA gene sequences was then classified with theBayesian method (Wang et al., 2007; Schloss et al., 2009)against a manually updated taxonomy database, whose se-quences were grouped at 98% level (available at http://www.secondgenome.com/go/2011-greengenes-taxonomy). The cul-tivable and molecular diversity was then compared on genuslevel. The 16S rRNA gene database created for this study,which includes all 16S rRNA genes from molecular studiesplus all 16S rRNA genes from isolates, can be provided bythe corresponding author upon request.
3. Results
3.1. Cultivable microbial diversity
As shown in Table 1, a large number of colonies wereobtained during the sampling campaign, and almost 900isolates have been processed (i.e., purified and sequenced).These isolates were obtained via different enrichmentconditions (bioburden, vegetative assay, alternative as-
says). A combination of all these three cultivation ap-proaches resulted in isolates belonging to 130 differentbacterial species, whereas the largest number of differenttaxa was obtained from the Kourou samples. The isolatesspanned 53 genera and 4 bacterial phyla [Actinobacteria,Bacteroidetes/Chlorobi, Firmicutes, Proteobacteria (a, b,c)]., Most of them were deposited at the DSMZ and areavailable for the public and further research (see Moissl-Eichinger et al., 2012; Schwendner et al., 2013). A summary ofall isolated bacterial genera obtained from all assays per-formed in samplings FR1, FR2, ES2, and KO is given inFigure 1, which also indicates the differences in microbialdiversity of each single sampling event.
Bacteria that were isolated from each of the facilities includedthe spore-forming microbes Bacillus and Paenibacillus but alsonon-spore-forming Gram-positives (Micrococcus, Staphylococcus)and Gram-negative Stenotrophomonas. The success of the alter-native enrichment strategies is demonstrated by the isolation of,for example, strictly anaerobic propionibacteria, retrieved fromthree samplings (FR1, FR2, ES2).
Further details with respect to the overall cultivable mi-crobial diversity have been given in other publications(Stieglmeier et al., 2009, 2012; Schwendner et al., 2013).
3.2. Cultivable, heat-shock-resistant microbialdiversity (‘‘bioburden’’)
The heat-shock-resistant microbial diversity obtained fol-lowing the ECSS standard for bioburden determination
FIG. 1. Venn diagram of the microbial diversity (genera) detected by cultivation. Microbial genera that were detected bycultivation and molecular detection methods are highlighted in bold. Results are shown for FR1, FR2, ES2, and KO only, sinceES1 sampling and analyses were performed without alternative cultivation methods.
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 5

spanned three phyla (Table 3). The overwhelming majorityof isolates were assigned to the phylum Firmicutes, withgenus Bacillus dominating. Bacillus pumilus was found ineach of the locations, as were representatives of the B. subtilisgroup.
Besides Gram-positive, spore-forming bacteria, a broadervariety of non-spore-formers and Gram-negative microor-ganisms were obtained. In particular, Staphylococcus strainsappeared frequently on agar plates, apparently resisting theapplication of heat shock (80�C for 15 min; Table 3).
3.3. Microbial diversity with respect to collectionand cultivation strategy
The cultivation success of microorganisms is strongly de-pendent on the enrichment medium used and cultivationstrategy applied. To visualize these differences, the fourdifferent sampling and enrichment methods were displayedaccording to their success in covering the bacterial diversity(Fig. 2).
Bacillus, Micrococcus, and Staphylococcus were detected byat least three different isolation strategies, whereas a broaddiversity of microbes (in, e.g., Kourou samples) was obtainedonly by applying a greater number of alternative cultivationconditions.
3.4. Diversity of cultivable, strictly and facultativelyanaerobic bacteria
The procedures and the successful isolation of strictlyand facultatively anaerobic bacteria from clean room sam-ples have been previously described (Stieglmeier et al., 2009;see also Probst et al., 2010a). During the Herschel campaign,most of the isolates recovered under anaerobic cultivationconditions were facultative anaerobes. Also, some strictlyanaerobic microbes were retrieved from each location,
adding up to 12% of all isolates obtained from alternativeassays (Table 4). Different strains of Propionibacterium wereisolated from FR1, FR2, and ES2. Spore-forming clostridiawere isolated from the ESTEC (ES2) and South Americanfacilities.
3.5. Diversity of extremotolerant bacteria
The use of alternative media for the enrichment of mi-croorganisms from spacecraft assembly facilities increasedthe overall isolate diversity by a factor of 2.6 (18 and 47genera for standard and alternative cultivation, respectively).In total, 29 microbial genera were exclusively obtained byproviding alternative cultivation conditions. The cultivableclean room microorganisms showed a clear preference foralkaline media, as well as media with reduced organiccompounds (Table 5).
Two Bacillus strains, one Paenibacillus strain, and Micro-coccus flavus were exclusively isolated on R2A medium, pH11. Dermacoccus sp. was enriched on plates with pH 5, as wasMycobacterium chubuense, Angustibacter sp., Terrabacter sp.,and Hymenobacter rigui, the only representative of the Bac-teroidetes phylum.
A broad diversity of oligotrophic and oligotolerant bac-teria was obtained on 1:100 diluted R2A: Acinetobacter, Bal-neimonas, Brevundimonas, Citrobacter, Kocuria, Microbacterium,Micrococcus, Moraxella, Paenibacillus, Sanguibacter, Staphylo-coccus, Stenotrophomonas, Streptomyces (Moissl-Eichinger,2011b). Colony counts obtained from media for oligotrophswere often comparable to or exceeded counts from classicalR2A medium (Table 5).
Sporosarcina globispora (FR2 sampling) was only isolatedunder low-temperature conditions (10�C) and was the soleisolate that was unable to grow at 32�C in subsequent cul-tivations. All other microbes isolated at 4�C and 10�C werecapable of growing at the standard incubation temperature,
Table 3. Cultivable, Heat-Shock-Resistant Microbial Diversity (‘‘Bioburden’’)
Phylum Genus Gram Spores1 ES1 ES2 FR2 KO
Actinobacteria Micrococcus + - M. luteus, M. lylaeRothia + - R. mucilaginosa
Firmicutes Bacillus + + B. pumilus,B. licheniformis,B. mojavensis,B. subtilis2
B. pumilus, B. circulans,B. firmus, B. flexus,B. licheniformis,B. megaterium,B. mojavensis,B. simplex, B. sp.,B. subtilis2
B. pumilus,B. subtilis2
B. pumilus,B. cereus,B. subtilis2
Paenibacillus + + P. humicus,P. favisporus
P. sp.
Staphylococcus + - S. caprae,S. epidermidis,S. haemolyticus,S. hominis
S. epidermidis S. sp.
Solibacillus + + S. silvestris(a-) Proteobacteria Paracoccus - - P. yeeii(c-) Proteobacteria Acinetobacter - - A. lwoffii A. sp.
Stenotrophomonas - - S. maltophilia
Heat-shock-resistant isolates from FR1 samplings were not characterized phylogenetically and are therefore not included in this table.1Isolates capable of forming spores.2B. subtilis refers to representatives of the B. subtilis group due to an insufficient resolution of the 16S rRNA gene sequence analysis; no
further characterization was performed.
6 MOISSL-EICHINGER ET AL.

and most of them were also detected under other growthconditions provided. Growth at lower temperatures wasdelayed compared to elevated temperatures. The only ther-motolerant microorganism isolated was Geobacillus sp. (ES2sampling).
Some isolates revealed multitolerance with regard to dif-ferent cultivation approaches. For instance, Micrococcus[isolation conditions: R2A 1:100, pH 11, and salt concentra-tions of 10% (w/v)] and Paenibacillus (isolation conditions:R2A 1:100, pH 11, anoxic, and 50�C) were also grown onmedium for CO2- and N2-fixing microbes. CO2-fixing activitywas found for Staphylococcus, but isolates of this genus alsoshowed the ability to grow under reduced organic concen-trations, alkaline pH, elevated salt concentration, and an-aerobic conditions. Moreover, isolates classified as Bacillusturned out to be physiologically versatile, being enrichedeven at pH 11, without oxygen, 50�C or elevated salt con-centrations.
3.6. Molecular microbial diversity basedon 16S rRNA gene cloning
Details with respect to molecular analyses based on bac-terial 16S rRNA gene pool cloning and sequencing from lo-cations FR1, FR2, ES2, and KO have been published earlier(Stieglmeier et al., 2012; Schwendner et al., 2013). Overall,the molecular analyses revealed the presence of signaturesof up to 10 bacterial phyla within the spacecraft assemblyclean rooms [Acidobacteria, Actinobacteria, Bacteroidetes,Chloroflexi, Cyanobacteria, Deinococcus/Thermus, Firmi-cutes, Gemmatimonadetes, Planctomycetes, and Proteo-bacteria (a, b, c)], whereas the highest diversity was obtainedfrom KO samples and the lowest diversity from FR1.
Propionibacterium and Staphylococcus were the only micro-bial genera that were detected in all facilities (Fig. 3). Inter-estingly, the molecular analyses revealed a relatively goodoverlap with cultivation results, and 18 microbial genera
FIG. 2. Number of bacterial genera obtained by differentcultivation strategies (isolates obtained after heat-shocktreatment and enrichment on TSA (‘‘Bioburden’’), isolatesgrown on R2A without prior heat shock (‘‘Vegetatives’’), andisolates obtained under alternative cultivation conditions(‘‘Alternative’’). ES1 and FR1 are not shown (alternative as-say not performed/low number of analyzed isolates). Airsampling was not performed at the FR2 sampling event.
Table 4. Number of Isolates That Were Obtained
under Strictly Anaerobic Conditions Proportional
to All Isolates from Alternative Cultivation
Procedures (in %)
No of isolates FR1 FR2 ES KO
.isolated underanaerobic conditions
7 (27%) 8 (44%) 12 (35%) 15 (31%)
.that were strictlyanaerobic
1 (4%) 1 (6%) 4 (12%) 1 (2%)
Table 5. Average Colony-Forming Units (CFU)
Obtained from Floor Samples from Each
Facility, Sorted by the Alternative Enrichment
Method Applied
Enrichment type Conditions FR1 FR2 ES2 KO
Oligotrophs R2A 1:10 > 2 · 104 2,600 12,000 > 2 · 104
Oligotrophs R2A 1:100 > 2 · 104 1,500 2,300 > 2 · 104
Alkaliphiles pH 9 1,300 2,900 14,000 > 2 · 104
Alkaliphiles pH 11 4,000 460 8,400 > 2 · 104
Acidophiles pH 5 940 38 0 1,700Acidophiles pH 3 0 0 0 80Psychrotolerants 4�C 560 970 1,900 > 2 · 104
Psychrotolerants 10�C 0 38 190 2,400Microaerophiles 3% O2 3,800 1,900 1,200 2,900Desiccation 190 0 0 n.d.Thermophiles 50�C 190 0 190 20Thermophiles 60�C 0 0 160 0Salt tolerants 3.5% NaCl 3,200 590 8,100 880Salt tolerants 10% NaCl 190 77 1,400 140Anaerobes 0% O2 0 170 380 580Bioburden* Heat shock n.a. 940 170 710Vegetatives* R2A n.a. n.a. 2,800 3,400
Only colony counts from solid media are shown. For comparison,CFU obtained from bioburden measurements as well as from thevegetative assay are given in the last two rows. ES1 is not shownsince no enrichment on alternative media was performed.
*According to ECSS standardn.a., not applicable due to overgrowth of the plates (swarming
colonies); n.d., not done.
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 7

were detected by both methods. Staphylococcus, for instance,was identified by cultivation and cultivation-independentmethods in all sampled facilities. Propionibacterial molecularsignatures were found in each clean room, and isolates werecultivated from three samplings (FR1, FR2, ES2).
Signatures of putative spore-forming microbes were notdetected with the molecular approach, verifying our previ-ous assumption that spore formers seem to be presentmainly as spores, which reduces the chance of being recog-nized by DNA extraction methods without bead-beating.
3.7. Molecular microbial diversity comparison
A database was generated with all molecular and culti-vable 16S rRNA gene sequences from clean room environ-ments that were publicly available at NCBI (National Centerfor Biotechnology Information, March 2012). Using classifi-cation of each sequence with the Bayesian algorithm againsta manually curated and updated Greengenes database (seeMaterials and Methods), we identified 230 different micro-bial genera present in clean room environments. One hun-dred forty-nine of these 230 microbial taxa were identifiedvia cloning only, and 53 were detected via molecular andcultivation tools. Twenty-eight genera were retrieved incultivation only and completely escaped the molecular assay.The amount of cultivable bacteria in clean room environ-ments was estimated to be 35% based on genus level, which
is extraordinarily high and can mainly be attributed to theapplication of alternate cultivation assays (see above).
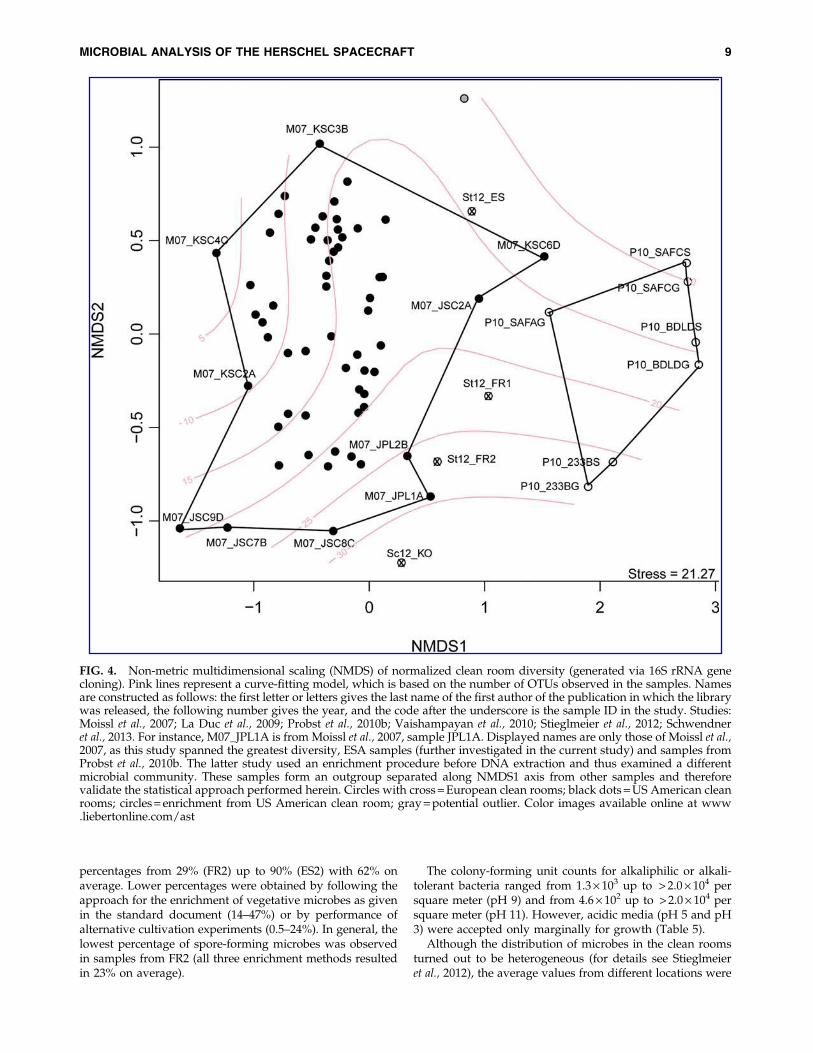
With regard to the molecular diversity only, a statisticalcomparison of all data generated with 16S rRNA genecloning from American and European clean room facilitieswas performed. Non-metric multidimensional scaling (Fig. 4)showed an outgroup of anaerobic enrichment cultures pub-lished by Probst et al. (2010a) and therefore confirmed ourstatistical approach. Microbial communities detected byMoissl et al. (2007) spanned the major part of the ordinationanalysis, which suggests that the study covered a greatproportion of the microbial diversity of clean room facilities.Samples from ESA spacecraft assembly facilities were not asclosely related to samples from NASA clean rooms as ex-pected, which indicates that the geographic relatedness of aclean room may have influence on the microbial communitystructure.
3.8. Microbial abundance based on cultivation,bioburden measurement, and molecular analysis
Spore-forming isolates were cultured from each of thefacilities. The heat-shock application as well as various en-richment conditions influenced the percentage of spore-forming organisms obtained compared to the total numberof isolates (Fig. 5). The bioburden protocol clearly supportsthe selective enrichment of spore formers and yields
FIG. 3. Venn diagram of the microbial diversity (genera) in the four sampling events FR1, FR2, ES2, and KO, detected bymolecular methods based on 16S rRNA gene analyses of the gene pool. Microbial genera that were detected by cultivationand molecular detection methods are highlighted in bold. Sequences that could not be phylogenetically classified on genuslevel are not shown.
8 MOISSL-EICHINGER ET AL.
percentages from 29% (FR2) up to 90% (ES2) with 62% onaverage. Lower percentages were obtained by following theapproach for the enrichment of vegetative microbes as givenin the standard document (14–47%) or by performance ofalternative cultivation experiments (0.5–24%). In general, thelowest percentage of spore-forming microbes was observedin samples from FR2 (all three enrichment methods resultedin 23% on average).
The colony-forming unit counts for alkaliphilic or alkali-tolerant bacteria ranged from 1.3 · 103 up to > 2.0 · 104 persquare meter (pH 9) and from 4.6 · 102 up to > 2.0 · 104 persquare meter (pH 11). However, acidic media (pH 5 and pH3) were accepted only marginally for growth (Table 5).
Although the distribution of microbes in the clean roomsturned out to be heterogeneous (for details see Stieglmeieret al., 2012), the average values from different locations were
FIG. 4. Non-metric multidimensional scaling (NMDS) of normalized clean room diversity (generated via 16S rRNA genecloning). Pink lines represent a curve-fitting model, which is based on the number of OTUs observed in the samples. Namesare constructed as follows: the first letter or letters gives the last name of the first author of the publication in which the librarywas released, the following number gives the year, and the code after the underscore is the sample ID in the study. Studies:Moissl et al., 2007; La Duc et al., 2009; Probst et al., 2010b; Vaishampayan et al., 2010; Stieglmeier et al., 2012; Schwendneret al., 2013. For instance, M07_JPL1A is from Moissl et al., 2007, sample JPL1A. Displayed names are only those of Moissl et al.,2007, as this study spanned the greatest diversity, ESA samples (further investigated in the current study) and samples fromProbst et al., 2010b. The latter study used an enrichment procedure before DNA extraction and thus examined a differentmicrobial community. These samples form an outgroup separated along NMDS1 axis from other samples and thereforevalidate the statistical approach performed herein. Circles with cross = European clean rooms; black dots = US American cleanrooms; circles = enrichment from US American clean room; gray = potential outlier. Color images available online at www.liebertonline.com/ast
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 9

quite consistent. Interestingly, the bioburden measurementsrevealed, independently from the clean room class, a con-tamination in the range of 102 heat-shock-resistant cultiva-bles per square meter of clean room floor and GSE, whereasthe cultivation approach without heat shock (vegetatives)resulted in one order of magnitude higher colony counts(Table 6). Nevertheless, compared to the data derived fromwipe sampling, the swab samples delivered inconsistentdata, most likely due to the small area sampled, which re-sulted in a higher variation (25 cm2, compared to up to 1 m2
sampling area for wipes; Stieglmeier et al., 2012).
3.9. Archaea
Archaea were detected in samples from FR2, ES2, andKO (other sampling campaigns were not used for the de-tection of archaea). ES2 samples even revealed archaealfluorescence in situ hybridization (FISH) signals of rod-shaped cells. The average, estimated cell number of archaeaper square meter of clean room floor did not exceed 1.2 · 104
(Moissl-Eichinger, 2011a; based on qPCR results), althoughtheir constant presence suggests a larger role within the cleanroom environment (further details, see Moissl-Eichinger,2011a). The detected archaeal diversity spans two differentarchaeal phyla: Thaumarchaeota and Euryarchaeota (me-thanogens and halophiles).
3.10. Fluorescence in situ hybridization (FISH)analyses
Fluorescence in situ hybridization was used to visualizemicrobes in original samples and to assign a phylogeneticaffiliation to certain morphologies. Although FISH is notfeasible for use in low-biomass samples, we were able tovisualize microbes in each sample. Different morphologieswere detected (cocci, rods, diplococci), revealing all goodsignals with bacteria-targeted probes. Examples for positiveFISH were given by Schwendner et al. (2013) and Moissl-Eichinger (2011a). Paenibacilli were visualized with specifi-cally designed probes in some of the Kourou samples
(Schwendner et al., 2013). Strong fluorescence signals suggesta high content of ribosomal RNA and are an indicator formicrobial activity. Interestingly, many microbes were foundto form aggregates with particles, which made their detec-tion quite difficult. Domain-specific FISH revealed also thepresence of archaea (Moissl-Eichinger, 2011a).
4. Discussion and Lessons Learned
In preparation for the upcoming ExoMars mission, ESAhas increased its efforts to analyze and characterize the mi-crobial diversity and abundance on and around spacecraftduring assembly, test, and launch operations. Although notsubject to planetary protection requirements, the Herschelspacecraft was used as a model for establishing and prac-ticing microbiological methods to analyze bioburden andbiodiversity. The microbiome of built environments and inparticular of confined clean rooms is indeed special. Due to arestricted exchange with the environment, microbes aretransferred via items or humans only and subsequentlysuppressed by harsh environmental conditions within theclean room. The confinement and applied cleaning proce-dures in classified clean rooms (ISO 146441) reduce the mi-crobial load significantly and select hardy microorganisms,such as spore formers. The Herschel project covered differentmethodologies to analyze spacecraft-associated microorgan-isms. We used a cultivation approach with more than 30different media to isolate bacteria and to feed the novel ESAculture collection (Moissl-Eichinger et al., 2012), and usedmolecular methods that allowed for insight into the uncul-tivable part of the microbiome. In the following section, wesummarize the major findings of the project and make rec-ommendations for changes in procedures or to improveprotocols.
Throughout this campaign, surface sampling was per-formed with different sampling tools, such as wipes, swabs,witness plates, BiSKits, and SpongeSicles. Each of these toolswas functional and returned useful data. However, futureESA studies will mainly use wipes and swabs for a number
FIG. 5. Cultivated number of spore formersproportional to all isolates (in %), dependenton the cultivation approach used: isolatesobtained after heat-shock treatment and en-richment on TSA (‘‘Bioburden’’), isolatesgrown on R2A without prior heat shock(‘‘Vegetatives’’), and isolates obtained underalternative cultivation conditions (‘‘Alter-native’’). +: The alternative approach wasnot performed for ES1. ‘‘Bioburden’’ and‘‘Vegetatives’’ counts from FR1 were notconsidered since only a very low number ofisolates were phylogenetically analyzed.
10 MOISSL-EICHINGER ET AL.

of reasons. First, different sampling tools have different re-covery efficiencies (Probst et al., 2011) and can, therefore,considerably affect comparability of results across multiplestudies, which can only be overcome by standardizing pro-tocols for sampling and for sample processing. Although thesponge-based sampling tools (BiSKits, SpongeSicles) deliv-ered good results, the possibility of sponge particle loss orbuffer residues on a spacecraft or in its close vicinity was aconcern and resulted in their exclusion (G. Kminek, personalcommunication).
In addition, analyses reported severe problems with theBiSKit tool with respect to DNA contamination of the in-cluded buffer (Kwan et al., 2011). Recent efforts have focusedon selection of wipes and swabs for sampling of clean roomand spacecraft surfaces, which can be used for both culti-vation and molecular assays. The nylon-flocked swab hasbeen evaluated previously for the efficient recovery of sporesfrom various surfaces (Probst et al., 2010b), but a lower effi-ciency compared to cotton swabs in recovering free DNA hasbeen found (Kwan et al., 2011). However, cotton swabs aremade of natural fibers, which are, as mentioned for sponges,a problem with respect to particle control. In addition, in-terference with subsequent DNA-based detection methodscould become a problem. A similar problem occurs withwipes, which cannot currently be delivered (certified) DNA-free. This has necessitated the inclusion of a step to removeany potential DNA contamination (dry-heat treatment;170�C, 24 h; A. Auerbach, personal communication). Basedon these arguments and the current status of knowledge,polyester wipes (for surfaces up to 1 m2) and nylon-flockedswabs (for surfaces up to 25 cm2) remain the sampling toolsof choice. Future studies should be based on the two selectedsampling tools for spacecraft and clean room surfaces. Suchan approach for bioburden and biodiversity studies willminimize bias potentially introduced by differing samplingtechniques and subsequent extraction protocols.
Since only 1% of all microorganisms can be cultivatedunder defined laboratory conditions (Amann et al., 1995),molecular techniques are generally useful to detect micro-organisms that cannot be found by cultivation. Molecularanalyses performed in this study were based on DNA ex-traction without bead-beating, qPCR analyses, and classical16S rRNA gene cloning (Stieglmeier et al., 2012; Schwendneret al., 2013). Molecular technologies that have been used tocharacterize clean room microbial diversity include 16SrRNA gene cloning (Venkateswaran et al., 2001; La Duc et al.,2004; Moissl et al., 2007, 2008; Probst et al., 2010a; Moissl-Eichinger, 2011a; Stieglmeier et al., 2012; Schwendner et al.,2013), PhyloChip DNA microarray of the generation 2 and 3(La Duc et al., 2009; Probst et al., 2010a; Cooper et al., 2011;Vaishampayan et al., 2013a), and 16S rRNA gene pyro-sequencing (La Duc et al., 2012; Vaishampayan et al., 2013a).PhyloChip and pyrosequencing were reported to detect agreater level of diversity than classical technologies but haveonly been conducted on a restricted number of samples.Although these two state-of-the-art technologies allowgreater insight into the biodiversity present, the majority ofclean room samples have only been analyzed with classical16S rRNA gene cloning.
Our comparative analysis demonstrated the Herschelclean room microbial diversity to somewhat differ from thatreported in NASA clean rooms. Furthermore, the Herschel
Ta
bl
e6.
Ab
un
da
nc
eo
fM
ic
ro
bia
lC
el
ls
pe
rS
qu
ar
eM
et
er
Cl
ea
nR
oo
mF
lo
or
,G
ro
un
dS
up
po
rt
Eq
uip
me
nt
,a
nd
Sp
ac
ec
ra
ft
,E
st
im
at
ed
by
Cu
lt
iv
at
io
n
(B
io
bu
rd
en
:S
ta
nd
ar
dA
ssa
yIn
cl
ud
in
gH
ea
tS
ho
ck
;V
eg
et
at
iv
es:
Cu
lt
iv
ab
le
Mic
ro
or
ga
nism
so
nR
2Aw
it
ho
ut
He
at
Sh
oc
k)
an
dq
PC
R
An
aly
sis
Sam
pli
ng
loca
tion
Too
lE
S1
ES
2F
R1
FR
2K
O1
Cu
ltiv
atio
n:
Bio
bu
rden
Flo
or,
GS
EW
ipe
n.a
.(4
0–n
.a)
170
(0–9
40)
n.a
.94
0(2
20–1
500)
710
(190
–160
0)S
wab
n.a
.(4
0–n
.a.)
1400
(0–1
1,00
0)82
00(0
–1.5
·10
4)
0n
.d.
Sp
acec
raft
Sw
ab11
.3(0
–40)
500
(0–1
000)
420
(0–1
.0·
103)
027
(0–9
7)C
ult
ivat
ion
:V
eget
ativ
esF
loo
r,G
SE
Wip
en
.a.
(190
–n.a
.)28
00(3
30–6
400)
n.a
.n
.a.
3400
(210
0–47
00)
Sw
ab12
.5(0
–20)
1.2
·10
4(0
–7.4
·10
4)
n.a
.30
0(0
–150
0)n
.d.
Sp
acec
raft
Sw
ab23
00(0
–2.5
·10
5)
6500
(0–1
.8·
104)
50(0
–150
0)50
8(0
–100
0)32
(0–4
2)q
PC
R2
Flo
or,
GS
EB
iSK
itn
.d.
2.6
·10
7(1
.1·
106
to7.
6·
107)
8.2
·10
6(3
.3·
106
to1.
3·
107)
3.9
·10
6(3
.0·
105
to1.
1·
107)
n.d
.S
pac
ecra
ftS
wab
n.d
.6.
1·
105
n.d
.1.
5·
108
n.d
.
Th
era
ng
eo
fco
lon
yco
un
tso
rce
llco
un
tsd
etec
ted
isg
iven
inp
aren
thes
is.
n.a
.,n
ot
app
lica
ble
du
eto
ov
erg
row
tho
fth
ep
late
s(s
war
min
gco
lon
ies)
;n
.d.,
no
td
on
e.1S
pac
ecra
ftsa
mp
les
wer
eta
ken
fro
mA
rian
e5
fair
ing
(Sch
wen
dn
eret
al.,
2013
).2N
um
ber
of
bac
teri
aw
ases
tim
ated
bas
edo
n16
SrR
NA
gen
eco
pie
s,co
rrec
ted
by
fact
or
4.1
[eac
hb
acte
rial
cell
con
tain
sse
ver
alco
pie
so
f16
SrR
NA
gen
es;
ple
ase
refe
rto
Lee
etal
.(2
009)
for
furt
her
det
ails
].
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 11

clean room contained organisms not previously reported inNASA clean room samples. The reasons for this observationare currently unclear but could be assigned to the geo-graphical location of the clean rooms (see also Moissl et al.,2007) or to method biases (DNA extraction technique, se-lected primers, PCR conditions, etc.; Von Wintzingerodeet al., 1997).
Nevertheless, this analysis demonstrated that synchro-nizing the protocols would also allow more comprehensivecomparisons, since this would reduce methodological biases.In addition, it is necessary to further characterize individualclean rooms to create a more complete picture of the mi-crobial signatures that might vary temporarily and spatially.Stieglmeier et al. (2012) clearly showed microbial contami-nation within a clean room to be extremely variable andhighly heterogeneous. As a consequence, frequent samplingand sampling of multiple locations within a clean room isimperative in establishing an accurate picture of the cleanroom microbiome.
Another suggested change to processing methods is theinclusion of a bead-beating step in the DNA extraction pro-tocol; although appearing frequently in cultivation assays,spore-forming bacteria were not at all detectable via 16SrRNA gene cloning in the Herschel campaign. This suggeststhat the DNA extraction method employed by these studieswas unable to open spores but nevertheless showed thepresence of those bacteria as spores and not as vegetativecells.
Answering the question of whether microorganisms arealive or dead in clean room facilities can be a crucial factorwhen assessing the potential risk of a forward contamina-tion. RNA-targeted studies may provide better evidence formicrobial viability than DNA-based studies as RNA de-grades faster; it is, however, also more difficult to extract(Vaishampayan et al., 2013a). As an alternative, studies haverecently used propidium monoazide (PMA) as a viabilitymarker (Probst et al., 2012; Vaishampayan et al., 2013a). PMAhas proven useful for the analysis of intact cells in variousmicrobial communities (Nocker et al., 2007). Masking DNAof disrupted cells for PCR downstream processing revealedthe biodiversity, which was alive or at least undisrupted atthe time point of sampling. The application of this technol-ogy to US American clean rooms demonstrated a lowermicrobial diversity and abundance than previously assumedby standard qPCR, pyrosequencing, and PhyloChip G3(Vaishampayan et al., 2013a). It has also been shown thatmission-critical clean rooms exhibited a lower microbialrichness and abundance than non-mission clean rooms withrespect to the living microbial proportion (Vaishampayanet al., 2013a). This indicates that cleaning protocols werehighly efficient. However, conservative planetary protectionconsiderations must assess on an individual basis the risk ofeach possible biological contaminant—alive or dead. PMA-combined technologies may prove useful in future cleanroom analyses. These molecular methods will not, however,replace time-consuming cultivation efforts. Only cultivationcan conclusively demonstrate the viability of organisms re-covered from clean rooms. More important, this approachallows microbial isolates to be collected, which can serve astest objects for cleaning and sterilization efforts. The analysisof resistances and adaptations of those microbial isolates bythe scientific community can add critical information to
planetary protection considerations (Venkateswaran et al.,2003; Link et al., 2004).
Samples from the South American clean room were foundby this study to have the broadest diversity of cultivablebacterial genera. The external environment has been pro-posed to exhibit a major influence on the clean room mi-crobiome (Moissl et al., 2007), and this seems to be true in thiscase, as the humid and warm climate supports biodiversity(Fierer et al., 2012; Schwendner et al., 2013). Cultivable bio-diversity is also influenced by the maintenance proceduresand is significantly reduced under higher particulate control,as observed for the FR1 sampling event. However, micro-organisms that were cultivated in samples from all four cleanrooms were the typical clean room contaminants: Bacillus,Micrococcus, Paenibacillus, and Staphylococcus. Interestingly,Stenotrophomonas, a contaminant not previously considered atypical clean room inhabitant, was cultivated from all loca-tions on standard media. Stenotrophomonas species werefound in air filters in two shopping malls in Singapore(Tringe et al., 2008) and have attracted interest with respect tomulti-drug-resistance and causing nosocomial infections inimmunocompromised people (Denton et al., 1998). In theliterature, representatives of these microbes were reported tobe resistant to a number of toxic metals, including cobalt,silver, mercury, and cadmium. This may be important forplanetary protection considerations (Pages et al., 2008).
Although heat shock was applied, a relatively low per-centage of spore formers were recovered on culture media.Micrococcus, Staphylococcus, Rothia, Acinetobacter, Stenotropho-monas, Paracoccus were among the heat-shock-surviving, non-spore-forming bacteria found in this study. Micrococcus andStaphylococcus are typical clean room contaminants and couldtherefore significantly influence colony counts retrieved inbioburden measurements. For a detailed discussion pleaserefer to Stieglmeier et al. (2012).
A large number of isolates have not been assigned toa species due to larger ( > 2.5%) phylogenetic distance ofthe 16S rRNA gene compared to closest related type spe-cies, or an unclear phylogenetic situation. One isolate(Tersicoccus phoenicis gen. nov, sp. nov.) was found torepresent a novel genus and was recently described(KO_PS43; Vaishampayan et al., 2013b). Interestingly,this species was isolated in parallel in a US clean room(Kennedy Space Center) independent of this study. An-other novel isolate was described 2 years earlier (Behrendtet al., 2010). A spore-forming organism, Paenibacilluspurispatii sp. nov., was isolated from a sample taken at theISO 8 clean room in ESTEC (ES2 sampling). The enrichmentof both bacteria was achieved in alternative media underanaerobic conditions (Hino and Wilson NH3-free mediumand TSA, respectively).
The application of alternative media has increased thecultivable microbial diversity immensely. For instance, theappearance of Propionibacterium is worth mentioning. Thismicroorganism was isolated from three European cleanrooms, and its cultivation was only possible by the use ofalternative cultivation media (anaerobic conditions). In ac-cordance with previous studies (e.g., La Duc et al., 2007),many isolates were found to tolerate ‘‘extreme’’ cultivationconditions. Media at pH 9 or 11 and low-nutrient plates, suchas R2A, revealed similar or higher colony counts. This couldindicate an adaptation of the microorganisms to (alkaline)
12 MOISSL-EICHINGER ET AL.

cleaning detergents and to low nutrient availability in cleanroom environments.
The application of these two conditions and the cultiva-tion under strictly anaerobic conditions has added crucialinsights with respect to the cultivable microbiome fromclean rooms. As a consequence, ESA has proposed to addthese three cultivation strategies as an optional procedure tothe standard document in order to analyze not only theheat-shock-resistant bioburden but also possibly adaptedmicroorganisms.
Archaea were detectable in low, but constant, abundancein all clean rooms analyzed to date; furthermore, in one casethese organisms were even shown to be intact and physio-logically active (Moissl et al., 2008; Moissl-Eichinger, 2011a).The role of Eury- and Thaumarchaeota in such built envi-ronments is currently unclear. Similarly, it is also unclearwhether these organisms pose a risk to planetary protectionefforts. Recent experiments have proven the association ofThaumarchaeota with human skin (Probst et al., 2013), whichis in accordance with the observation that most microbialcontaminants in clean rooms are human-associated.
Fluorescence in situ hybridization has proven useful forthe visualization of (active) archaea and bacteria in cleanroom samples. Although the overall number of cells wasrather low, detection of coccoid and rod-shaped microbeswas possible, which were mainly attached to particles. Thisobservation supported the decision to sonicate the wipesduring extraction steps in order to loosen the attachment ofmicrobial cells to particle or wipe surfaces.
Although a presumably large portion of the microbialcommunity present on and around spacecraft can be de-tected, analyzed, and visualized, the microbiome in cleanrooms is still mysterious in many ways. However, eachanalysis performed will contribute important data for futuresampling campaigns as did the Herschel study. The spaceagencies’ efforts are substantial to reduce microbial con-tamination and to confine the risk for extraterrestrial envi-ronments and life-detection missions as much as possible.This is an effort that will require constant adaptation andimprovement of protocols in order to deliver a better un-derstanding of the microbial diversity and biocontaminationin space travel.
Acknowledgments
Part of the research described in this paper was carried outby DLR under contract with ESA, ESTEC contract no. 20234/06/NL/EK. The work performed in Regensburg was fundedby ESA, ESTEC contract no. 20508/07/NL/EK. We thankthe Herschel Project Team for support during sampling. Wethank Gerhard Kminek for critical discussion. Alexander J.Probst was supported by the National German AcademicFoundation (Studienstiftung des deutschen Volkes). We aregrateful to Anna Auerbach and Melanie Duckstein for pro-viding information, lab management, and technical assis-tance. We thank Gabriel Milinovich for critically reading themanuscript.
Abbreviations
DLR, Deutsches Zentrum fur Luft- und Raumfahrt; DSMZ,Deutsche Sammlung von Mikroorganismen und Zellkulturen
GmbH; ECSS, European Cooperation for Space Standardiza-tion; ESTEC, European Space Research and TechnologyCentre; FISH, fluorescence in situ hybridization; GSE, groundsupport equipment; OTU, operational taxonomic unit; PMA,propidium monoazide; qPCR, quantitative PCR; TG, thio-glycollate liquid medium; TGA, thioglycollate agar; TS, tryp-ticase soy liquid medium; TSA, trypticase soy agar.
References
Amann, R.I., Ludwig, W., and Schleifer, K.H. (1995) Phyloge-netic identification and in situ detection of individual micro-bial cells without cultivation. Microbiol Rev 59:143–169.
Behrendt, U., Schumann, P., Stieglmeier, M., Pukall, R., Au-gustin, J., Sproer, C., Schwendner, P., Moissl-Eichinger, C.,and Ulrich, A. (2010) Characterization of heterotrophic nitri-fying bacteria with respiratory ammonification and denitrifi-cation activity—description of Paenibacillus uliginis sp. nov., aninhabitant of fen peat soil and Paenibacillus purispatii sp. nov.,isolated from a spacecraft assembly clean room. Syst ApplMicrobiol 33:328–336.
Conley, C.A. and Rummel, J.D. (2013) Appropriate protection ofMars. Nat Geosci 6:587–588.
Cooper, M., La Duc, M.T., Probst, A., Vaishampayan, P., Stam,C., Benardini, J.N., Piceno, Y.M., Andersen, G.L., and Ven-kateswaran, K. (2011) Comparison of innovative molecularapproaches and standard spore assays for assessment of sur-face cleanliness. Appl Environ Microbiol 77:5438–5444.
COSPAR. (2011) COSPAR Planetary Protection Policy (20 October2002, as amended to 24 March 2011), COSPAR, Paris. Avail-able online at https://cosparhq.cnes.fr/sites/default/files/pppolicy.pdf.
Crawford, R.L. (2005) Microbial diversity and its relationship toplanetary protection. Appl Environ Microbiol 71:4163–4168.
Denton, M., Todd, N.J., Kerr, K.G., Hawkey, P.M., and Little-wood, J.M. (1998) Molecular epidemiology of Steno-trophomonas maltophilia isolated from clinical specimens frompatients with cystic fibrosis and associated environmentalsamples. J Clin Microbiol 36:1953–1958.
ECSS-Q-ST-70-55 Working Group. (2008) Space product assur-ance: microbial examination of flight hardware and cleanrooms. ECSS-Q-ST-70-55C, European Cooperation for SpaceStandardization, ESA Requirements and Standards Division,European Space Agency, ESTEC, Noordwijk, the Netherlands.
Fairen, A.G. and Schulze-Makuch, D. (2013) The overprotectionof Mars. Nat Geosci 6:510–511.
Felsenstein, J. (1989) PHYLIP—Phylogeny Inference Package(version 3.2). Cladistics 5:164–166.
Felsenstein, J. (1993) PHYLIP (Phylogeny Inference Package)version 3.5c. Distributed by the author. Department of Ge-netics, University of Washington, Seattle.
Fierer, N., Leff, J.W., Adams, B.J., Nielsen, U.N., Bates, S.T.,Lauber, C.L., Owens, S., Gilbert, J.A., Wall, D.H., and Ca-poraso, J.G. (2012) Cross-biome metagenomic analyses of soilmicrobial communities and their functional attributes. ProcNatl Acad Sci USA 109:21390–21395.
Hippe, H.H. (1991) Maintenance of methanogenic bacteria. InMaintenance of Microorganisms and Cell Cultures, edited by B.E.Kirsop and A. Doyle, Academic Press, London, pp 101–114.
Kim, O.S., Cho, Y.J., Lee, K., Yoon, S.H., Kim, M., Na, H., Park,S.C., Jeon, Y.S., Lee, J.H., Yi, H., Won, S., and Chun, J. (2012).Introducing EzTaxon-e: a prokaryotic 16S rRNA gene se-quence database with phylotypes that represent unculturedspecies. Int J Syst Evol Microbiol 62:716–721.
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 13

Kwan, K., Cooper, M., La Duc, M.T., Vaishampayan, P., Stam,C., Benardini, J.N., Scalzi, G., Moissl-Eichinger, C., andVenkateswaran, K. (2011) Evaluation of procedures forthe collection, processing, and analysis of biomolecules fromlow-biomass surfaces. Appl Environ Microbiol 77:2943–2953.
La Duc, M.T., Kern, R., and Venkateswaran, K. (2004) Microbialmonitoring of spacecraft and associated environments. MicrobEcol 47:150–158.
La Duc, M.T., Dekas, A., Osman, S., Moissl, C., Newcombe, D.,and Venkateswaran, K. (2007) Isolation and characterization ofbacteria capable of tolerating the extreme conditions of cleanroom environments. Appl Environ Microbiol 73:2600–2611.
La Duc, M.T., Osman, S., Vaishampayan, P., Piceno, Y., Ander-sen, G., Spry, J.A., and Venkateswaran, K. (2009) Compre-hensive census of bacteria in clean rooms by using DNAmicroarray and cloning methods. Appl Environ Microbiol75:6559–6567.
La Duc, M.T., Vaishampayan, P., Nilsson, H.R., Torok, T.,and Venkateswaran, K. (2012) Pyrosequencing-derivedbacterial, archaeal, and fungal diversity of spacecraft hardwaredestined for Mars. Appl Environ Microbiol 78:5912–5922.
Lee, Z.M.-P., Bussema, C., and Schmidt, T.M. (2009) RrnDB:documenting the number of rRNA and tRNA genes in bac-teria and archaea. Nucleic Acids Res 37:D489–D493.
Link, L., Sawyer, J., Venkateswaran, K., and Nicholson, W.(2004) Extreme spore UV resistance of Bacillus pumilus isolatesobtained from an ultraclean spacecraft assembly facility. Mi-crob Ecol 47:159–163.
Lis, D.C., Schilke, P., Bergin, E.A., Emprechtinger, M., andHEXOS Team. (2012) Hot, metastable hydronium ion in thegalactic centre: formation pumping in X-ray-irradiated gas?Philos Trans A Math Phys Eng Sci 370:5162–5173.
Moissl, C., Osman, S., La Duc, M.T., Dekas, A., Brodie, E., De-Santis, T., and Venkateswaran, K. (2007) Molecular bacterialcommunity analysis of clean rooms where spacecraft are as-sembled. FEMS Microbiol Ecol 61:509–521.
Moissl, C., Bruckner, J.C., and Venkateswaran, K. (2008) Ar-chaeal diversity analysis of spacecraft assembly clean rooms.ISME J 2:115–119.
Moissl-Eichinger, C. (2011a) Archaea in artificial environments:their presence in global spacecraft clean rooms and impact onplanetary protection. ISME J 5:209–219.
Moissl-Eichinger, C. (2011b) Extremophiles in spacecraft assemblyclean rooms. In Adaption of Microbial Life to Environmental Ex-tremes: Novel Research Results and Application, edited by H. Stan-Lotter and S. Fendrihan, Springer, Wien, pp 231–261.
Moissl-Eichinger, C., Rettberg, P., and Pukall, R. (2012) The firstcollection of spacecraft-associated microorganisms: a publicsource for extremotolerant microorganisms from spacecraftassembly clean rooms. Astrobiology 12:1024–1034.
NASA-HBBK-6022. (2007) NASA handbook for the microbial ex-amination of space hardware. NASA, Washington, DC. Avail-able online at http://planetaryprotection.nasa.gov/documents.
Nocker, A., Sossa-Fernandez, P., Burr, M.D., and Camper,A.K. (2007) Use of propidium monoazide for live/deaddistinction in microbial ecology. Appl Environ Microbiol73:5111–5117.
Pages, D., Rose, J., Conrod, S., Cuine, S., Carrier, P., Heulin, T.,and Achouak, W. (2008) Heavy metal tolerance in Steno-trophomonas maltophilia. PLoS One 3:e1539.
Probst, A., Vaishampayan, P., Osman, S., Moissl-Eichinger, C.,Andersen, G.L., and Venkateswaran, K. (2010a) Diversity ofanaerobic microbes in spacecraft assembly clean rooms. ApplEnviron Microbiol 76:2837–2845.
Probst, A., Facius, R., Wirth, R., and Moissl-Eichinger, C. (2010b)Validation of a nylon-flocked-swab protocol for efficient re-covery of bacterial spores from smooth and rough surfaces.Appl Environ Microbiol 76:5148–5158.
Probst, A., Facius, R., Wirth, R., Wolf, M., and Moissl-Eichinger,C. (2011) Recovery of Bacillus spore contaminants from roughsurfaces: a challenge to space mission cleanliness control. ApplEnviron Microbiol 77:1628–1637.
Probst, A., Mahnert, A., Weber, C., Haberer, K., and Moissl-Eichinger, C. (2012) Detecting inactivated endospores influorescence microscopy using propidium monoazide. Inter-national Journal of Astrobiology 11:117–123.
Probst, A., Auerbach, A.K., and Moissl-Eichinger, C. (2013) Ar-chaea on human skin. PLoS One 8:e65388.
Pruesse, E., Peplies, J., and Glockner, F.O. (2012) SINA: accuratehigh-throughput multiple sequence alignment of ribosomalRNA genes. Bioinformatics 28:1823–1829.
Puleo, J.R., Fields, N.D., Bergstrom, L., Oxborrow, G.S., Stabekis,P.D., and Koukol, R. (1977) Microbiological profiles of theViking spacecraft. Appl Environ Microbiol 33:379–384.
Schloss, P.D., Westcott, L., Ryabin, T., Hall, J.R., Hartmann, M.,Hollister, E.B., Lesniewski, R.A., Oakley, B.B., Parks, D.H.,Robinson, C.J., Sahl, J.W., Stres, B., Thallinger, G.G., VanHorn, D.J., and Weber, C.F. (2009) Introducing mothur: open-source, platform-independent, community-supported soft-ware for describing and comparing microbial communities.Appl Environ Microbiol 75:7537–7541.
Schwendner, P., Moissl-Eichinger, C., Barczyk, S., Bohmeier, M.,Pukall, R., and Rettberg, P. (2013) Insights into the microbialdiversity and bioburden in a South American spacecraftassembly clean room. Astrobiology 5, doi:10.1089/ast.2013.1023.
Stieglmeier, M., Wirth, R., Kminek, G., and Moissl-Eichinger, C.(2009) Cultivation of anaerobic and facultatively anaerobicbacteria from spacecraft-associated clean rooms. Appl EnvironMicrobiol 75:3484–3491.
Stieglmeier, M., Rettberg, P., Barczyk, S., Bohmeier, M., Pukall,R., Wirth, R., and Moissl-Eichinger, C. (2012) Abundance anddiversity of microbial inhabitants in European spacecraft-associated clean rooms. Astrobiology 12:572–585.
R Development Core Team (2011) R: a language and environ-ment for statistical computing. R Foundation for StatisticalComputing, Vienna, Austria. http://www.R-project.org
Tringe, S.G., Zhang, T., Liu, X., Yu, Y., Lee, W.H., Yap, J., Yao, F.,Suan, S.T., Ing, S.K., Haynes, M., Rohwer, F., Wei, C.L., Tan, P.,Bristow, J., Rubin, E.M., and Ruan, Y. (2008) The airborne me-tagenome in an indoor urban environment. PLoS One 3:e1862.
Vaishampayan, P., Osman, S., Andersen, G., and Venkateswaran,K. (2010) High-density 16S microarray and clone library–basedmicrobial community composition of the Phoenix spacecraftassembly clean room. Astrobiology 10:499–508.
Vaishampayan, P., Probst, A.J., La Duc, M.T., Bargoma, E.,Benardini, J.N., Andersen, G.L., and Venkateswaran, K.(2013a) New perspectives on viable microbial communities inlow-biomass cleanroom environments. ISME J 7:312–324.
Vaishampayan, P., Moissl-Eichinger, C., Pukall, R., Schumann,P., Sproer, C., Augustus, A., Hayden Roberts, A., Namba, G.,Cisneros, J., Salmassi, T., and Venkateswaran, K. (2013b) De-scription of Tersicoccus phoenicis gen. nov., sp. nov. isolatedfrom spacecraft assembly clean room environments. Int J SystEvol Microbiol 63:2463–2471.
Venkateswaran, K., Satomi, M., Chung, S., Kern, R., Koukol, R.,Basic, C., and White, D. (2001) Molecular microbial diversityof a spacecraft assembly facility. Syst Appl Microbiol 24:311–320.
14 MOISSL-EICHINGER ET AL.

Venkateswaran, K., Kempf, M., Chen, F., Satomi, M., Nicholson,W., and Kern, R. (2003) Bacillus nealsonii sp. nov., isolated froma spacecraft-assembly facility, whose spores are gamma-ra-diation resistant. Int J Syst Evol Microbiol 53:165–172.
Von Wintzingerode, F., Gobel, U.B., and Stackebrandt, E. (1997)Determination of microbial diversity in environmental sam-ples: pitfalls of PCR-based rRNA analysis. FEMS Microbiol Rev21:213–229.
Wang, Q., Garrity, G.M., Tiedje, J.M., and Cole, J.R. (2007) NaıveBayesian classifier for rapid assignment of rRNA sequencesinto the new bacterial taxonomy. Appl Environ Microbiol73:5261–5267.
Address correspondence to:Christine Moissl-Eichinger
Institute of Microbiology and Archaea CenterUniversity of Regensburg
Universitaetsstrasse 3193053 Regensburg
Germany
E-mail: [email protected]
Submitted 19 April 2013Accepted 26 October 2013
MICROBIAL ANALYSIS OF THE HERSCHEL SPACECRAFT 15





























