Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 210, No. 1, August, pp. 90-97, 1981
Leucine Aminopeptidase (Bovine Lens): Synthesis and Kinetic Properties of Ortho-, Meta-, and Para-Substituted Leucyl-Anilides’
ALLEN TAYLOR,2 FRANCIS E. TISDELL, AND FREDERICK H. CARPENTER3
Department of Biochemistry, University of California, Berkeley, Calqwnia 9&?20
Received December 2,198O
The synthesis of a number of leucyl derivatives of substituted anilides and their prop- erties as substrates and inhibitors of Zn2+-Mgz+ leucine aminopeptidase (EC 3.4.11.1) at pH 8.5 and 30°C are described. The compounds include leucyl-X where X is o-, m-, or p aminobenzenesulfonic acid, o-, m-, or panisidine, and m- or p-aminobenzenesulfonyl fluoride. The latter two sulfonyl fluorides, designed to be active site-directed irreversible inhibitors, turned out to be good substrates for leucine aminopeptidase. The K, and V values of the above compounds as substrates for leucine aminopeptidase are reported. N-Leucyl-m-aminobenzenesulfonate exhibits desirable properties (solubility much greater than K,, h at 295 nm of 2000 M-' cm-‘, and V of 306 rmol min-’ mg-‘) as a substrate for a spectrophotometric assay of leucine aminopeptidase. With the exception of N-leucyl- paminobenzenesulfonate, all of the above compounds are inhibitors of the hydrolysis of leucyl-pnitroanilide by leucine aminopeptidase with Ki values approximately their K, values when they are used as substrates. Despite wide variability in steric bulk, chemical composition, and electrical charge of the substituted anilides, the K, values of the above compounds vary over a narrow range (0.5 to 4.8 mM), which indicates that the leucyl moiety plays the predominant role in the determination of K,,, values. Although the K, values of m- substituents are similar to those of o- substituents, the V values for m- substituents are much greater than those for o- substituents, which suggests that o- substituents interfere with the catalytic process. N-Leucyl-p-aminobenzenesulfonate and N-alanyl-p-aminobenzenesulfonate as well as the nonsubstrate paminobenzenesulfonate stimulate rather than inhibit the proteolysis of leucyl-pnitroanilide. The stimulation has no effect on V but lowers the K, for the hydrolysis of leucyl-pnitroanilide, which is compatible with these compounds’ serving as nonessential activators.
While the mechanism of action and the quarternary structure of carboxypepti- dases and endopeptidases are known in substantial detail, information concerning the aminopeptidases is less well developed (3-5).
Leucine aminopeptidase activity has been detected in many tissues and organs
’ This work was supported in part by grants from the National Institutes of Health to F.H.C. (EY 00813 and AM 00618) and to A.T. (EY 02342). Preliminary reports of this work have appeared (1, 2).
’ Recipient of a National Institutes of Health post- doctoral fellowship, 5-FO2-54141. Current address, Thompson Chemistry Laboratories, Williams Col- lege, Williamstown Mass. 81267.
3 To whom correspondence should be addressed.
of animals, plants, and bacteria, and has been associated with several pathological conditions (3, 6-10). The present study concerns leucine aminopeptidase from bo- vine lens (EC 3.4.11.1). The native enzyme contains six identical subunits per mole- cule (11-14). Each subunit consists of a single polypeptide of 54,000 daltons (12) which contains one Zn2+ in a structural site and another Zn2+, or other divalent metal, in an activation site (15-1’7). Ex- change of Mn2+ or M@;2+ for the activation site Zn2+ is associated with a 24- or ll-fold change, respectively, in the rate of hydro- lysis, with little effect on the Km values of amide substrates (15).
The present work was initiated in a
0003-9861/81/090090-08$02.00/O Copyright 0 1981 by Academic Press. Inc. All rights of reproduction in any form reserved.
90

ACTION OF BOVINE LEUCINE AMINOPEPTIDASE ON SUBSTITUTED ANILIDES 91
search for an active site-directed irre- versible inhibitor for leucine aminopepti- dase. The putative inhibitors were mod- eled on the work of Baker and Hurlbut (18) who made use of sulfonyl fluorides to form covalent bonds at the active site of chymotrypsin. The proposed inhibitors, N- leucyl-m- or p-aminobenzenesulfonyl flu- oride, proved to be good substrates and led to the investigation of similar compounds for use in the assay and elucidation of the mechanism of action of leucine aminopep- tidase.
Most kinetic investigations of leucine aminopeptidase activity have utilized spectrophotometric observation of the hy- drolysis of leucyl-pnitroanilide or leucyl- amide. There are problems with both as- say systems. Leucyl-p-nitroanilide is sol- uble only up to one-half K,,, of the Zn2+- Zn2+ or Zn’+-Mp enzyme (1, 16, l?), so it is impossible to investigate the kinetics under conditions approaching saturation of the enzyme. Although leucyl-amide is soluble to >lO X Km, it has been reported (19) that a transpeptidation reaction oc- curs at high substrate concentrations. Since transpeptidation results in no net peptide bond cleavage, it interferes with those assays designed to detect such cleav- age. Several of the compounds reported here fulfill two criteria for useful sub- strates, i.e., high solubility and significant change in extinction coefficient upon hy- drolysis.
A detailed kinetic analysis of the Km and V values of the variously substituted leu- cyl-anilides revealed pertinent informa- tion about the mechanism of action of the enzyme.
EXPERIMENTAL PROCEDURES
Materials. Leucine aminopeptidase in the Zn*+- Zn2+ form was isolated from bovine lens tissue by a modification (20) of the procedure of Hanson et al. (21). Polyacrylamide gel electrophoresis was used to check the homogeneity of the native (11) and dena- tured enzyme (12). For kinetic studies the Znz+-Zn*+ enzyme was converted to the Zn*+-Mgz+ form by in- cubation for 12 h at 3’7’C of a combination of 10 ~1 leucine aminopeptidase solution (10 to 15 mg/ml in 0.1 M Tris .HCl, pH 8.0) with 1.14 ml 2.2 mM NaHC03/ Na&03, pH 9.5, 10.8 mM MgClz. The Zn2+-Mgz+ en-
zyme had a specific activity of >600 pmol min-’ mg-’ in the leucyl-amide assay (15).
Leucyl-amide. HCl, alanyl-amide, leucyl-p-ni- troanilide, Boc-Leu,” Boc-Leu-ONp, Leu-Ala, Leu- Gly-Gly, and Ala-Gly-Gly were purchased from Vega-Fox. Sulfanilic, metanilic, and orthanilic acids, the o-, m-, and panisidines, and the m- and pami- nobenzenesulfonyl fluorides were obtained from Eastman. N,N’-Dicyclohexylcarbodiimide and tri- fluoroacetic acid were obtained from Aldrich, and Matheson, Coleman and Bell, respectively. Silica gel for column chromatography was 40-140 mesh from J, T. Baker. All other chemicals were purchased from Fisher Scientific, Eastman, Schwartz-Mann, Sigma, or MCB, and were of the highest purity available.
Syntheses. The syntheses of all the leucyl-anisi- dides and the leucyl-sulfonylfluorides were identical to that of leucyl-m-anisidide and are an adaptation of the procedure of Tarenghi and Bergonzi.’
Synthesis of leucyl-m-anisidide. m-Anisidine (0.99 g, 8.0 mmol) was added to a solution of Boc-leucine. Hz0 (2.2 g, 8.8 mmol) in 25 ml CHzClz. A solution of N,W-dicyclohexylcarbodiimide (2.0 g, 9.7 mmol) in 10 ml CHzClz was dripped into the flask and the mix- ture was stirred at room temperature for ‘72 h. The precipitate of dicyclohexylurea was removed and washed with CHzClz. The volume of the combined supernatants was reduced to 5 ml and the concen- trate was applied to a 2 X 100-cm silica gel column. Elution was performed with CHCls:EtOAc (9:l). Al- iquots of the column fractions were subjected to thin- layer chromatography on silica gel plates (Analtech) in the same solvent system. Those fractions contain- ing Boc-Leu-m-anisidide were pooled and the sol- vents were removed and replaced with CHCls. After extraction of the CHCls solution with 10 mM HCl, the organic phase was dried over anhydrous MgSO,. Dry HCl gas (Matheson) was bubbled into the CHCls so- lution for 20 min to effect the removal of the Boc group. A crystalline product was obtained several hours after Ccl, was added to the CHC& solution.
Synthesis of N-&ucyl)-m-aminobenzenesu&bnic acid. Boc-Leu-ONp (4.4 g, 12.9 mmol) was dissolved in 30 ml dry pyridine at 42°C. Metanilic acid (2.0 g, 11.5 mmol) was added and the reaction mixture was stirred for 16 h at 80°C. Solution of the reactants occurred after 2 h. The pyridine was removed under vacuum, leaving a thick resin. The resin was dis- solved in 40 ml of water and the pH was adjusted to 3.2 with 6 M HOAc. The white precipitate which formed was removed by gravity filtration and dis-
4 Abbreviations used: Boc-, t-butyloxycarbonyl; EtOAc, ethyl acetate; HOAc, acetic acid; ONp, pni- trophenyl ester; THF, tetrahydrofuran.
’ Personal communication from G. Tarenghi, Miles Italiana S.P.A., Divisione Armes, Laboratorie di Ri- cerche, 2205 Garlate, (CO), Italy.

92 TAYLOR, TISDELL,
TABLE I
EXTINCTION COEFFICIENT DIFFERENCES BETWEEN LEUCYL-X AND X IN 0.1 M Tris.HCl/O.Ol M HCOa
pH 8.5,30°C AT WAVELENGTHS USED IN KINETICS ASSAYS
La-x
l,eu- -0 0 sag Leu-
-4 0 S02T
La”-Nd@O;
Leu-NH 0 Q so; I.eu-NE 0
P 3 Le”-NH -u 0 OCHj Leu-NH 0
-$. a1
I,eu-NH 0
-P O "3
.I used x
AE~ between for assay Leu-X and X
nm H-1 cm-’
290 10,300
295 300
295 830 300 550
295 2.000
300 2,500
300 1,850
295 360
300 170
carded. Nitrophenol was extracted from the aqueous phase with three 30-ml portions of EtOAc. The water was removed by rotary evaporation, leaving a residue which was redissolved in ethanol. Upon removal of the solvent, there remained a friable resin which was crushed to a powder. Trifluoroacetic acid (5 ml) was added to the dry powder, causing immediate evolu- tion of carbon dioxide. After the reaction was stirred for 20 min. the solution was poured into cold diethyl ether with rapid stirring. The resulting precipitate was maintained at -10°C for 6 h, then collected and dried over Pz05. The syntheses of N-(leucyl)-pami- nobenzenesulfonic acid, N-(leucyl)-o-aminobenzene- sulfonic acid, and N-(alanyl)-paminobenzenesul- fonic acid were performed in the same fashion.
Methods. Ultraviolet and visible absorption spectra and enzyme kinetics were performed in a buffer con- sisting of 0.1 M Tris.HCl, 0.01 M NaHCOs, pH 8.5, on either a Cary 118 or a Cary 15 recording spectro- photometer equipped with thermostatted cells set at 30°C. Actual wavelengths used to monitor the hy- drolysis of the peptide bond of the substituted leucyl- anilides are recorded in Table I.
The usual assay involved the addition of 100 ~1 of the Zn’+-Mg2f enzyme in the activation buffer to a solution of the substrate(s) in the Tris buffer indi- cated above. Total volume was 2.5 ml. In experiments with N-(leucyl)-m-aminobenzenesulfonyl fluoride, there existed the possibility of aqueous hydrolysis of
AND CARPENTER
the fluoride and corresponding changes in extinction even before enzyme was added. In experiments in- volving these substrates, this situation was avoided by adding a stock solution of these compounds in 25 al of dimethyl sulfoxide to the appropriate buffer- enzyme solution. This amount of dimethyl sulfoxide had no effect on activity under these conditions. For those derivatives lacking a substituted anilide (i.e., Leu-Ala, Leu-Gly-Gly, Ala-Gly-Gly), and for those derivatives which had very slow rates of hydrolysis (N-[leucyll-o-aminobenzenesulfonic acid, N-[leucyl]- o-anisidide), aliquots were removed from the hydro- lysis mixture at various time intervals and diluted with 0.2 Mcitrate buffer at pH 2.2 to stop the reaction. These aliquots were subjected to amino acid analysis on a modified Beckman 120B amino acid analyzer using a 0.9 X 57-cm column packed with A-15 resin which was eluted with 0.2 M citrate buffer (15). In all kinetic experiments the concentration of substrate was varied over at least a lo-fold range. Initial ve- locities were determined from reactions which in- volved less than 5% depletion of the initial substrate concentration. Values of K,,, and V were calculated from the computer program HYPERB (22).
Infrared spectra were obtained on Perkin-Elmer Models 237B and 257 spectrophotometers. Varian Associates T-60, A-60, and a Perkin-Elmer R-12B were used to record NMR spectra. Melting points were determined in open capillary tubes using either a Thomas-Hoover melting point apparatus or a Lab- oratory Devices Melt-Temp and are uncorrected. El- emental analyses were performed by the Micro- analytical Laboratory, Department of Chemistry, University of California, Berkeley.
RESULTS
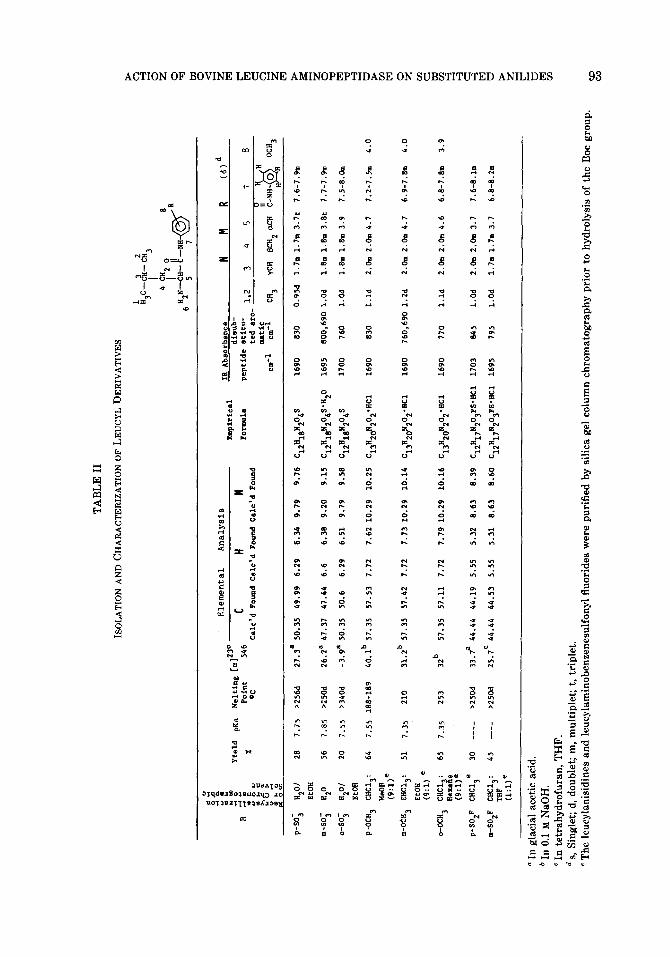
Table II shows the yields obtained in the synthesis of the various leucyl-anilide de- rivatives along with their pK, values, melting points, optical rotations, elemen- tal analyses, and their characterization by infrared absorption and nuclear magnetic resonance. Utilizing differences in absorp- tion spectrum between these peptide an- alogs and the products of their hydrolysis (Table I), kinetic constants were deter- mined for these compounds in assays in which their hydrolysis was monitored alone or in the presence of an alternate substrate (Table III). Each of the com- pounds exhibited a solubility in the assay buffer that was severalfold larger than their Km values. Consequently, unlike leu- cyl-pnitroanilide, which is soluble only to about one-half its Km value, the measure-
TABL
E II
ISOL
ATIO
N AN
DCHA
RACT
ERIZA
TIONO
FLEU
CYLD
ERIV
ATIV
ES
&L
A3
4 CH
2 0
H Nd
"-U-N
$gS
6 2
” 4;
rz
Eleme
ntal
Analy
sis
Yield
p"a
Keltln
S kx
~230
Em
pirica
l IR
Ab
sorba
nce
N M
R
(6)
d :t
R z5
PO
iW
546
C H
N dis
ub-
Jbw
x FO
rOlU
la pe
ptide
~lt
itu-
QC
Cnlc'd
Po
und
Calc'd
Fo
und
Calc'd
Fo
und
ted
(1=0-
Is2
3 '
' z&
g m
atie
'g
; cm
-1
cm-1
CH
3
p-50
; H2
0/ 20
7.7
5 >Z
SCd
EfOH
m-S
O;
iI20
56
7.85
X506
o-so;
H20/
20
7.55
>340
d Et
OH
P-OC
H3
CHCl
3 :
64
7.55
188-1
89
HeOH
c9
:1je
m-OC
H3
cHc1
3: 51
7.3
5 21
0 no
n (9
:l) e
o-OCH
3 CH
C13:
65
7.35
253
Hcxe
ne
(9:l)e
p-S
02P
CHC1
3e
30
----
>250
d
m-S
O21
C"C1
3: 45
---
_ >2
50d
THP
(l:lJe
27.3a
50
.35
49.99
6.2
9 6.3
4 9.7
9 9.7
6
26.2=
47
.37
47.44
6.6
6.3
8 9.2
0 9.1
5
-3.9'
50.35
50
.6 6.2
9 6.5
1 9.7
9 9.5
8
40.1b
57
.35
57.53
7.7
2 7.6
2 10
.29
10.25
31.2b
51
.35
57.42
7.7
2 7.1
3 10
.29
10.14
3Zb
57.35
57
.11
7.12
7.79
10.29
10
.16
33.P
44
.44
44.19
5.5
5 5.3
2 8.6
3 6.3
9
25.7’
44
.44
44.53
5.5
5 5.3
1 8.6
3 8.6
0
C12H
xlN20
4S
C12"1
8N2%
S."20
ClZHlB
N204
S
1690
1695
1700
1690
1690
77
0 l.ld
2.
Om
2.01,~
4.6
6.8
-7.8m
3.9
1695
830
0.95d
1.7
m
1.7m
3.
7t
800,6
90
l.Od
l.Sm
1.8
1 3.8
t
760
l.Od
1.8m
1.8
m
3.9
830
l.ld
2.Om
2.o
m
4.7
760,6
90
1.2d
2.Qm
2.
Om
4.7
6.9-
7.8m
4.
0
a15
l.od
2.Om
2.
Qm
3.7
795
l.Od
1.7m
1.7
m
3.7
7.6-
7.9m
7.7-
7.9m
7.5~8
.Om
7.2-7
.5m
4.0
7.6~8
.1m
6.84.2
m
a In
gl
acia
l ac
etic
acid
. *
In
0.1
M
NaOH
. ’
In
tetra
hydr
ofura
n, TH
F.
d s,
Si
nglet
; d,
do
ublet
; m
, m
ultip
let;
t, tri
plet.
e Th
e le
ucyla
nisid
ines
an
d leu
cylam
inobe
nzen
esulf
onyl
fluor
ides
were
pu
rified
by
si
lica
gel
colum
n ch
rom
atogr
aphy
pr
ior
to
hydr
olysis
of
th
e Bo
c gr
oup.

94 TAYLOR, TISDELL, AND CARPENTER
TABLE III
KINETIC CONSTANTS FOR L-LEUCYL, L-ALANYC DERIVATIVES AND PEPTIDES T = 3O”C, pH 8.5,0.1 M Tris.HCl, 0.01 M HCO;, LAP’ in Zn*+-Mg2f FORM
Compound % vm in Competition with Leu-Yb InM
* 5 Y . HIM
3.2
3 2.4
OCH \3
Ieu-NH- 0 0 4.8
2.0
S02F
2.3
La-Ala 0.7 770 = 0.6
Leu-Gly-Gly 1.0 804 =
Ala-Gly-Gly 50 640 =
600 6.8
3.9
300
co.1 c 2.2
7.8 2.5
5.0 2.2
0.7 =
2.0
1000 sob -NH
0.9
,100 -NH~-NO,
8.7
2-0; -NH- 0 c
CH3
1.6
6.2
-NK 0 0
NO2
-N
3.5 -NH u-
0 NO2
-
’ LAP, Leucine aminopeptidase. * All compounds except LmSF exhibited competitive inhibition when included with the substrate shown. ’ Checked by amino acid analysis.
ments of K,,, values were not hampered by alternate substrate, the alternate sub- limitations in substrate solubilities. Each strate exhibits the properties of a com- of the kinetic parameters reported for ex- petitive inhibitor. The measured Ki value periments which involve monitoring the of the alternate substrate should be equal proteolysis of leucyl-pnitroanilide (Table to the K,,, value measured directly on the III) is the average of at least 4, and in some alternate substrate. Inspection of Table cases more than 10 determinations. All III reveals that, in accordance with theory, other experiments were performed a min- the Ki value of most of the alternate sub- imum of two times. strates closely approximated their Km
According to Dixon and Webb (23), values when measured alone. One excep- when the action of an enzyme on one sub- tion was N-leucyl-o-aminobenzenesulfo- strate is measured in the presence of an nate which was hydrolyzed too slowly to
ACTION OF BOVINE LEUCINE AMINOPEPTIDASE ON SUBSTITUTED ANILIDES 95
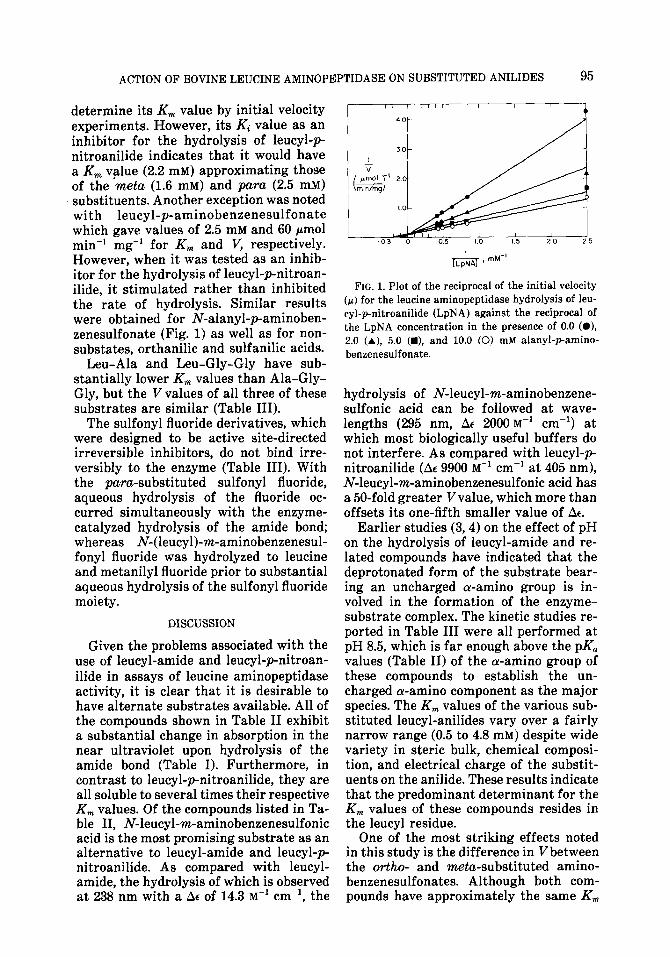
determine its Km value by initial velocity experiments. However, its Ki value as an inhibitor for the hydrolysis of leucyl-p- nitroanilide indicates that it would have a Km value (2.2 mM) approximating those of the meta (1.6 mM) and purct (2.5 mM) substituents. Another exception was noted with leucyl-p-aminobenzenesulfonate which gave values of 2.5 mM and 60 pmol min-’ mg-’ for Km and V, respectively. However, when it was tested as an inhib- itor for the hydrolysis of leucyl-pnitroan- ilide, it stimulated rather than inhibited the rate of hydrolysis. Similar results were obtained for N-alanyl-paminoben- zenesulfonate (Fig. 1) as well as for non- substates, orthanilic and sulfanilic acids.
Leu-Ala and Leu-Gly-Gly have sub- stantially lower K,,, values than Ala-Gly- Gly, but the Vvalues of all three of these substrates are similar (Table III).
The sulfonyl fluoride derivatives, which were designed to be active site-directed irreversible inhibitors, do not bind irre- versibly to the enzyme (Table III). With the paru-substituted sulfonyl fluoride, aqueous hydrolysis of the fluoride oc- curred simultaneously with the enzyme- catalyzed hydrolysis of the amide bond; whereas N-(leucyl)-m-aminobenzenesul- fonyl fluoride was hydrolyzed to leucine and metanilyl fluoride prior to substantial aqueous hydrolysis of the sulfonyl fluoride moiety.
DISCUSSION
Given the problems associated with the use of leucyl-amide and leucyl-p-nitroan- ilide in assays of leucine aminopeptidase activity, it is clear that it is desirable to have alternate substrates available. All of the compounds shown in Table II exhibit a substantial change in absorption in the near ultraviolet upon hydrolysis of the amide bond (Table I). Furthermore, in contrast to leucyl-p-nitroanilide, they are all soluble to several times their respective Km values. Of the compounds listed in Ta- ble II, N-leucyl-m-aminobenzenesulfonic acid is the most promising substrate as an alternative to leucyl-amide and leucyl-p- nitroanilide. As compared with leucyl- amide, the hydrolysis of which is observed at 238 nm with a AC of 14.3 M-’ cm-‘, the
FIG. 1. Plot of the reciprocal of the initial velocity (F) for the leucine aminopeptidase hydrolysis of leu- cyl-p-nitroanilide (LpNA) against the reciprocal of the LpNA concentration in the presence of 0.0 (O), 2.0 (A), 5.0 (m), and 10.0 (0) mM alanyl-pamino- benzenesulfonate.
hydrolysis of N-leucyl-m-aminobenzene- sulfonic acid can be followed at wave- lengths (295 nm, At 2000 M-’ cm-‘) at which most biologically useful buffers do not interfere. As compared with leucyl-p- nitroanilide (Ae 9900 M-’ cm-’ at 405 nm), N-leucyl-m-aminobenzenesulfonic acid has a 50-fold greater Vvalue, which more than offsets its one-fifth smaller value of At.
Earlier studies (3,4) on the effect of pH on the hydrolysis of leucyl-amide and re- lated compounds have indicated that the deprotonated form of the substrate bear- ing an uncharged a-amino group is in- volved in the formation of the enzyme- substrate complex. The kinetic studies re- ported in Table III were all performed at pH 8.5, which is far enough above the pK, values (Table II) of the a-amino group of these compounds to establish the un- charged a-amino component as the major species. The Km values of the various sub- stituted leucyl-anilides vary over a fairly narrow range (0.5 to 4.8 mM) despite wide variety in steric bulk, chemical composi- tion, and electrical charge of the substit- uents on the anilide. These results indicate that the predominant determinant for the Km values of these compounds resides in the leucyl residue.
One of the most striking effects noted in this study is the difference in Vbetween the o&ho- and m&a-substituted amino- benzenesulfonates. Although both com- pounds have approximately the same Km

96 TAYLOR, TISDELL, AND CARPENTER
values, the meta substituent is hydrolyzed at a rate several orders of magnitude greater than that of the ortho compound. A similar, although not as dramatic, dif- ference is noted between the o- and m-an- isidides. It is unlikely that these differ- ences can be attributed to inductive effects propagated through the benzene ring to the peptide bond. A more likely explana- tion involves an interference with the cat- alytic process, either through steric per- turbation of the active site and/or (in the case of the N-leucyl-o-aminobenzenesul- fonate) the proximity of a negative charge. These observations yield suggestive, al- though not conclusive, evidence on the mechanism of action of leucine aminopep- tidase. The fact that the ortho- and meta- substituted anilides have approximately the same K, value while differing sub- stantially in their V values suggests that in the minimal kinetic expression
E+S ~ES$E+P,
K, is equal to KS and that the rate-lim- iting step involves some term in k3. Fur- ther evidence for this suggestion is af- forded by a comparison of Leu-Gly-Gly with Ala-Gly-Gly. Although their K, val- ues differ by more than 50-fold, their V values are essentially the same. If K, is equal to K,, one might expect the same rate of release of the dipeptide (Gly-Gly) from the saturated enzyme.
One set of observations obtained in these studies is left without resolution. iV-Leucyl-paminobenzenesulfonate gives values of 2.5 mM and 60 Nmol min-’ mg-’ for K, and V, respectively, when used as a substrate for leucine aminopeptidase. However, when this compound is tested as an inhibitor in the hydrolysis of leucyl-p nitroanilide, it stimulates rather than in- hibits hydrolysis. Similarly, the non- substrates paminobenzenesulfonate and o-aminobenzenesulfonate (but not m- aminobenzenesulfonate) and the very poor substrate N-alanyl-p-aminobenzenesul- fonate stimulate the proteolysis of leucyl- p-nitroanilide. As shown by Fig. 1, in- creasing concentrations of the activator N-alanyl-paminobenzenesulfonate do not
change the Vfor the hydrolysis of leucyl- pnitroanilide, but do lower the apparent K, value. Such an effect is compatible with this compound’s serving as a nones- sential activator, but further work is needed in order to clarify its role’in the catalytic mechanism.
The N-leucyl-aminobenzenesulfonyl fluorides were designed to be active site- directed affinity labels by analogy to sim- ilar compounds which inactivate chymo- trypsin (18). These compounds not only did not inactivate leucine aminopeptidase, but proved to be quite good substrates. These results complement other attempts to la- bel a nucleophilic site at the active center by use of chloromethyl ketones (24-26) or by serine (4) or sulfhydryl reagents (15). All have failed to implicate a specific crit- ical nucleophile in the mechanism of ac- tion of leucine aminopeptidase.
ACKNOWLEDGMENTS
The authors wish to thank Sharon Glasser Fischer for aid with the kinetic studies, Dr. Charles Sawyer for help with computer programs, Dr. Steve Rosen- berg for consultations on spectrophotometric mea- surements, and Jackson Lee and David Beardsley for aid with optical rotation measurements.
1.
2.
3.
4.
5.
REFERENCES
TAYLOR, A., AND CARPENTER, F. H. (1977) Pacific Slope Biochemistry Conference Abstracts, June 27-29, p. 31, Oregon State University, Corvalis.
TAYLOR, A., AND CARPENTER, F. H. (1978) Fed. Proc. 37,1696.
SMITH, E. L., AND HILL, R. L. (1960) in The En- zymes (Boyer, P. D., Lardy, H., and Myrback, K., eds.), 2nd ed., Vol. 4, Part A, pp. 37-62, Academic Press, New York.
DELANGE, R. J., AND SMITH, E. L. (1972) in The Enzymes (Boyer, P. D., ed.), 3rd ed., Vol. 3, pp. 81-102, Academic Press, New York.
HANSON, H., AND FROHNE, M. (1976) in Methods in Enzymolology (Lorand, L., ed.), Vol. 45, pp. 564-521, Academic Press, New York.
DEVI, A. (1963) B&hem. Biophys. Acta 73,155- 158.
UETE, T., SHMANO, N., AND SHIMIZU, S. (1974) Clin. Chem. 20,834~837.
SWANSON, A. A., AND TRUESDALE, A. W. (1974) Ophthalmic Res. 6,235-W.
HAHN, U., SWANSON, H. A., AND HOCKWIN, 0. (1976) Albrect Graefes Arch. Klin. Exp. Ophthalmol. 199.197296.

ACTION OF BOVINE LEUCINE AMINOPEPTIDASE ON SUBSTITUTED ANILIDES 97
lo. VAN HEYNINGEN, R., AND TRAYHURN, P. (1976) Exp. Eye Res. 22,625-637.
11. MELBYE, S. W., AND CARPENTER, F. H. (1971) J. Biol. Chem. 246,2459-2463.
12. CARPENTER, F. H., AND HARRINGTON, K. T. (1972) J. Biol. Chem. 247, 5580-5586.
13. JURNAK, F., RICH, A., VAN LOON-KLAASSEN, L., BLOEMENDAL, H., TAYLOR, A., AND CARPEN- TER, F. H. (1977) J. Mol. Biol. 112,149-153.
14. TAYLOR, A., WLODAWER, A., AND CARPENTER, F. H. (1979) J. Ultra&%&. Res. 68.92-196.
15. CARPENTER, F. H., AND VAHL, J. M. (1973) J. BioL Chem. 248, 294-304.
16. THOMPSON, G. A., AND CARPENTER, F. H. (1976) J. Biol. Chem. 251,53-60.
17. THOMPSON, G. A., ANDCARPENTER, F. H.(1976) J. Biol. Chem. 251.1618-1624.
18. BAKER,B. R., ANDHURLBUT, J. A. (1969) J. Med. Chem. 12. 221-224.
19. HANSON, H., AND LASCH, J. (1967) Hqvpe Seyler’s 2. PhysioL Chem. 348,1525-1539.
20. MELBYE, S. W. (1970) Ph.D. thesis, University of California, Berkeley.
21. HANSON, H., GLASSER, D., AND KIRSCHKE, H. (1965) Hoppe Seykr’s 2. PhysioL Chem. 340, 107-125.
22. HANSON, K. R., LING, R., AND HAVIR, E. (1967) B&hem. Biophys, Res. Commun. 29,194-197.
23. DIXON, M., AND WEBB, E. C. (1958) Enzymes, p. 91, Academic Press, New York.
24. VAHL, J. M. (1970) Ph.D. thesis, University of California, Berkeley.
25. BIRCH, P. L., EL OBIED, H. A., AND AKHTAR, M. (1972) Arch. Biochim. Biophys. 148,447-448.
26. FI~KAU, S., FORSTER, U., PASCUAL, C., AND SCHUNCK, W. H. (1974) Eur. J. Biochem. 44, 523-528.





























