Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Mar. 1985, p. 599-6070099-2240/85/030599-09$02.00/0Copyright C 1985, American Society for Microbiology
Leucine Incorporation and Its Potential as a Measure of ProteinSynthesis by Bacteria in Natural Aquatic Systemst
DAVID KIRCHMAN,+* ELIZABETH K'NEES, AND ROBERT HODSONDepartment of Microbiology and Institute of Ecology, University of Georgia, Athens, Georgia 30602
Received 23 July 1984/Accepted 21 December 1984
Leucine incorporation was examined as a method for estimating rates of protein synthesis by bacterialassemblages in natural aquatic systems. The proportion of the total bacterial population that took up leucinein three marine environments was high (>50%). Most of the leucine (>90%) taken up was incorporated intoprotein, and little (<20%) was degraded to other amino acids, except in two oligotrophic marine environments.In samples from these two environments, ca. 50% of the leucine incorporated had been degraded to otheramino acids, which were subsequently incorporated into protein. The degree of leucine degradation appears todepend on the organic carbon supply, as the proportion of 3H-radioactivity incorporated into protein that wasrecovered as [3H]leucine after acid hydrolysis increased with the addition of pyruvate to oligotrophic watersamples. The addition of extracellular leucine inhibited total incorporation of [14C]pyruvate (a precursor forleucine biosynthesis) into protein. Furthermore, the proportion of [14C]pyruvate incorporation into proteinthat was recovered as [14C]leucine decreased with the addition of extracellular leucine. These results show thatthe addition of extracellular leucine inhibits leucine biosynthesis by marine bacterial assemblages. The molarfraction of leucine in a wide variety of proteins is constant, indicating that changes in leucine incorporationrates reflect changes in rates of protein synthesis rather than changes in the leucine content of proteins. Theresults demonstrate that the incorporation rate of [3H]leucine into a hot trichloroacetic acid-insoluble cellfraction can serve as an index of protein synthesis by bacterial assemblages in aquatic systems.
The rate of protein synthesis could be a valuable param-eter for evaluating the role of bacteria in trophic dynamicsand biogeochemical cycles of aquatic systems. Protein is a
major macromolecule in bacterial cells and constitutes overhalf of the dry weight of bacteria in pure culture (14) and inmixed marine assemblages (8). Thus, protein synthesis perse must be a significant fraction of total biomass production.Since protein synthesis depends on the utilization of nitrog-enous compounds taken up from the environment, the rateof protein synthesis may be a more direct measure thanestimates derived from total bacterial production of nitrogenutilization by heterotrophic bacteria in aquatic systems. Therate of protein synthesis would also account for a largefraction of the rate of utilization of compounds for energyproduction, since 96% of the total energy expended forsynthesizing macromolecules by Escherichia coli, for exam-ple, is used to synthesize proteins (14). Furthermore, be-cause protein is a valuable food source for heterotrophicorganisms (31), rates of net bacterial protein synthesis mayreflect more accurately than estimates of total bacterialproduction the contribution of bacteria to the energeticneeds of organisms that graze on bacteria.
Rates of protein synthesis in total microbial assemblageshave been estimated from the incorporation of 35SO4 (4).However, there are two problems with the application of thistechnique to bacterial assemblages, especially in marineenvironments. First, phytoplankton are responsible for a
large proportion of total SO4 uptake (84% in a freshwaterlake [15]), and thus changes in rates of 15 O4 incorporationcould be due to either the phytoplankton or the bacteria.
* Corresponding author.t Contribution no. 532 of the Marine Institute, University of
Georgia, Sapelo Island, GA 31327.t Present address: Department of Molecular Genetics and Cell
Biology, The University of Chicago, Chicago, IL 60637.
Second, the incorporation of 35SO4 is difficult to measure inmarine environments because the high concentration ofsulfate in seawater isotopically dilutes the added 35SO4. Forthese reasons we chose to examine the incorporation ofleucine to complement the existing method based on 35 o4
incorporation.Leucine incorporation is often used in laboratory studies
as an index of protein synthesis (for examples, see refer-ences 22, 32, and 37). In its simplest form, the methodconsists of adding radiolabeled leucine and measuring theaccumulation of radioactivity in the hot trichloroacetic acid(TCA)-insoluble cell fraction collected on membrane filters.The hot TCA extract contains protein, peptidoglycan, andperhaps some complex polysaccharides (10, 29). The use ofleucine incorporation rates to estimate the rate of proteinsynthesis by bacterial assemblages in aquatic environmentshas not been examined.
This paper reports several experiments on leucine uptakeand biosynthesis by marine bacterial assemblages. Althoughthe experiments were designed to explore aspects of leucineincorporation as a method to measure protein synthesis bynatural bacterial assemblages, the work described here is thecontinuation of our efforts (17) to examine the regulation oftransport and incorporation of amino acids by bacterialassemblages. The regulation of amino acid uptake per se isimportant to understand because of the role of these com-pounds in the dissolved nitrogen cycle and in supplyingnitrogen for bacterial growth. Kirchman and Hodson (17)demonstrated the tight coupling between transport of aminoacids such as leucine and their incorporation into protein. Inthis report we examine the biosynthesis of leucine and theutilization of leucine to synthesize other amino acids.
MATERIALS AND METHODS
Sampling sites. We conducted experiments at several lo-cations to minimize the risk that our results would be appli-
599
Vol. 49, No. 3
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.

600 KIRCHMAN, K'NEES, AND HODSON
cable to only one type of aquatic environment. Our initialexperiments were carried out during May 1983 on the R. V.Cape Hatteras in continental shelf and Gulf Stream watersoff the southeast coast of the United States. The precisegeographical locations are given with the results of eachexperiment. Water was collected with Nisken bottles from5 m below the surface, unless otherwise indicated. Subse-quent experiments were completed during October 1983 onthe R. V. Cape Florida, which was anchored off the coast ofAndros Island, Bahamas. Subsurface samples were collectedin polycarbonate bottles from a mangrove estuary (FreshCreek) on Andros Island and brought back to the CapeFlorida for experimentation. In addition, Nisken bottles wereused to collect samples from S m in waters ca. 100 m deep ata distance of 2 km offshore. We refer to these samples asoriginating from the oligotrophic Tongue of the Ocean, sinceleucine turnover rates and bacterial abundance were low.Several experiments were conducted at Sapelo Island, Ga.Samples were collected in polycarbonate bottles from DoboySound (Marsh Landing) and brought back to the laboratoryat Sapelo Island for experimentation. At all sites the exper-iments began within an hour after collection of the samples.The uptake studies were conducted at the in situ temperatureand in the dark.
Proportion of bacteria incorporating leucine. The propor-tion of the total bacterial assemblage that took up leucinewas estimated by a microautoradiographic technique devel-oped by Tabor and Neihof (30) and Meyer-Reil (21). Samples(10 ml) from the Bahamian estuary and the Tongue of theOcean were incubated with [3H]leucine for 4 h. The specificactivity of the added leucine was 50 ,uCi/nmol (New EnglandNuclear Corp.), and the final added concentration was 0.5nM. After incubation, Formalin was added such that thefinal concentration was 2%. To check for abiotic adsorptionof [3H]leucine, Formalin was added before the addition ofthe radiolabel, and the sample was processed with theothers. The samples were stored at 4°C until analyzed. Amore extensive study was conducted with samples fromSapelo Island. The concentrations used and incubation timesare given below.The microautoradiographic procedure followed was the
MARG-E method described by Tabor and Neihof (30).Briefly, appropriate sample volumes were filtered throughNuclepore filters (pore size, 0.2 ,um) and rinsed once with 5ml of filter-sterilized seawater. With the help of a safetylight, the filters were transferred to acid-cleaned microscopeslides that had been coated with NTB-2 autoradiographicemulsion (Eastman Kodak Co.). The side of the filter withthe sample was placed face down into the emulsion. Theemulsion-coated slide with the filter was then placed onto acold surface to solidify in total darkness. After solidifying,the slides were incubated in the dark over dessicant for 3days at 15°C.The emulsion-coated slides with filters were developed
and then stained with acridine orange as described by Taborand Neihof (30). After being destained with a series of citratebuffers (30), the filters were removed by first soaking theslides in 1% glycerol for 1 min and then gently peeling thefilters away from the emulsion once they were dried. Theemulsion-coated slides were then examined under epifluor-escence microscopy to count the acridine orange-stainedbacteria and under bright-field microscopy to check foraccumulation of silver grains associated with the bacteria.To be scored as a bacterium that took up leucine, at leastthree silver grains had to be within a distance from thestained cell that was less than the length of the bacterium.
Total bacterial abundance in each sample was enumeratedby the standard acridine orange direct-count method (12,34). This estimate of the total bacterial abundance wascompared with the number of bacteria counted in the mi-croautoradiographic preparations to determine whether thebacteria were completely transferred from the Nucleporefilter to the NTB-2 emulsion. All the bacteria were trans-ferred, as total bacterial abundance estimated by the regulardirect-count procedure and from the microautoradiographicpreparation differed by less than 10%. Subsamples of theFormalin-fixed seawater used for the microautoradiographicmeasurements were also filtered onto membrane filters (poresize, 2 ,um) and rinsed. The filters, after base hydrolysis (seebelow), were radioassayed directly to determine the turn-over rate of leucine (assimilated radioactivity divided byadded radioactivity) in each sample.
Possible uptake of [3H]leucine by cyanobacteria was alsoexamined. The same microautoradiographic procedure wasused as described above, except that the filters were notstained with acridine orange. The concentration of the addedleucine was 0.5 nM, and the incubation time was 4 h. Insteadof staining the bacteria, we looked with epifluorescent andbright-field microscopy for accumulation of silver grainsassociated with autofluorescing particles. This experimentwas conducted during May 1984 with water samples from100 m, which was the depth of the chlorophyll maximum, inthe Gulf Stream (30°10.10' N; 79029.23' W).
Recovery of [3HJleucine from protein hydrolysates. Thebasic procedure we propose to estimate protein synthesis isto measure the incorporation of [3H]leucine into hot TCA-insoluble material that is collected on membrane filters. Thedetails of our procedure is as follows. [4,5-3H]leucine and, insome experiments, unlabeled leucine or pyruvate were addedto water samples. Acid-killed controls were also analyzed tocorrect for abiotic uptake. The specific activity of the addedleucine ranged from 50 to 56.5 ,uCi/nmol, depending on thebatches obtained from New England Nuclear or AmershamCorp. The final concentration of the added [3H]leucine was0.5 nM. At appropriate times (the precise times are givenwith each experiment) subsamples were withdrawn, andTCA was added such that the final concentration was 5%.The subsamples were extracted in a boiling water bath for 30min. After the subsamples cooled, the TCA-insoluble mate-rial was collected on either Nuclepore or Gelman filters(pore size, 0.2 ,um) and rinsed twice with ca. 3 ml of5% coldTCA. The filters were either treated for liquid scintillationcounting to measure total incorporation into the hot TCA-insoluble cell fraction or frozen at -20°C for later analysis.One series of experiments examined total uptake of leucine.In that experiment the subsamples were not extracted inTCA but were filtered through membrane filters and rinsedtwice with 3.0 ml of filtered seawater. The filters used tomeasure total uptake or incorporation into protein were firsttreated with 1.0 ml of Protosol (New England Nuclear) for atleast 1 h at 50°C. This treatment was effective in minimizingself-absorption of the low-energy beta particles, since thesame results were obtained by combusting the filters in aBiological Oxidizer (R. J. Harvey Co.), which results in ahomogeneous sample in the cocktail. After the treatmentwith the alkaline regeant, the samples were neutralized with0.5 ml of 0.5 N HCl, and 10 ml of Scintiverse I (FisherScientific Co.) was added. Quench correction was by thechannels ratio method calculated by a Beckman LS-9000scintillation counter.We determined to what extent [3H]leucine was trans-
formed to other compounds, particularly other amino acids,
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.

LEUCINE INCORPORATION AS AN INDEX OF PROTEIN SYNTHESIS
that could be subsequently incorporated into the hot TCA-insoluble cell fraction. The following procedure was usedwith the Nuclepore filters. The additional steps necessaryfor Gelman filters are described below. The frozen Nu-cleopore filters were thawed and added to 5 ml ampulesalong with 2 ml of 6 N HCl. After being flushed with argon orN2, the ampules were sealed and placed in a 110°C oven for20 h. The HCl, containing amino acids and other monomersresulting from the acid hydrolysis, was transferred to a glassscintillation vial (20 ml) and evaporated on a hot plate (50°C)under a stream of N2The dried acid hydrolysate was dissolved in 5 ml of
distilled water and desalted. A cation-exchange column wasprepared with Dowex 50W-X4 (200-400 mesh; hydrogenform), which had been rinsed several times with distilledwater. After the acid hydrolysate was added to the column,the anionic and neutral compounds were eluted with 5 ml ofdeionized water. The aqueous elution was saved and radio-assayed. The amino acids were then eluted with 4 ml of 1 MNH40H.The ammonium solution, which contained the amino acids
resulting from the acid hydrolysis, was dried down on a hotplate (50°C) under a stream of N2. The evaporated samplesthen were dissolved in 50 xl of 2 mM leucine. Leucine wasseparated from the other amino acids and monomers by amethod of thin-layer chromatography (TLC) designed toseparate leucine from isoleucine, valine, and phenylalanine(20). The amino acids were applied in 10-,u aliquots to silicagel TLC plates. The same volume of 2 mM leucine wasspotted in an adjacent lane for every plate. We found theWhatman Linear-K plates convenient to use because of theirpreabsorbent areas and channeled lanes. The plates weredeveloped in a methyl ethyl ketone-pyridine-water-glacialacetic acid (73:5:20:2) solution. After chromatography, thelane containing the standard was sprayed with ninhydrin(0.1% [wt/vol] in acetone) to visualize the leucine. Thesample lanes were covered to prevent contact with theninhydrin.Based on the Rf of the leucine standard developed on the
same plate, the leucine spots in the adjacent sample laneswere scraped and placed in 1.0 ml of water in a scintillationvial. The remaining sections of the lane, including thepreabsorbent area, were added to a different scintillation vialcontaining 1.0 ml of water. After the compounds wereallowed to elute off the TLC scrapings for 30 min, 10 ml ofScintiverse I was added. Recovery of amino acids from thesample lanes was 100%. For some experiments we scrapeddifferent areas of the sample lanes corresponding to theamino acids less polar (above the leucine spot on thechromatograph) and more polar (below the leucine spot)than leucine. Selected amino acids, including isoleucine,valine, alanine, and phenylalanine, were chromatographedas standards for these experiments.We used Gelman filters in some of our experiments. When
these cellulose acetate filters were hydrolyzed directly in 6 NHCl, the resulting caramel material could not be chromato-graphed. Instead, the filters were treated with the proteolyticenzyme, pepsin. First, the filters were cut in half. One halfwas radioassayed without pepsin treatment. The other halfwas placed in 1.0 ml of the pepsin (1.0 mg/ml of 0.1 N HCl)for 6 h at 37°C. We found that more radioactivity wassolubilized from the filter with increasing pepsin concentra-tions, but that concentrations higher than 1.0 mg ml-1resulted in a caramel material that could not be chromato-graphed. The removal of radioactivity from the filter did notincrease when the filters were incubated for longer than 6 h.
Pepsin had no effect on free leucine, since we were able torecover >95% of [3H]leucine by TLC when the free aminoacid was pepsin treated. After treatment the filter wasradioassayed and compared with the untreated filter todetermine how much radioactivity was solubilized by thepepsin treatment. The pepsin solution, containing the par-tially digested TCA-insoluble material, was added to anampule along with 1.06 ml of concentrated HCl to give a 6 NHCl solution. After flushing, the ampule was sealed andincubated in a 110°C oven for 20 h. The acid hydrolysate wasthen desalted and chromatographed as described above.Gelman filters and the pepsin treatment were used for
experiments whose results are shown below (see Fig. 3, 4,and 7). We repeated these experiments when possible andobtained the same results. We compared Gelman and Nu-cleopore filters directly and found that the measured propor-tion of total 3H-radioactivity recovered as [3H]leucine wasthe same for both filters. Furthermore, both filters areequally effective in trapping radioactivity in the hot TCA-in-soluble fraction when used for samples from natural aquaticenvironments. Nuclepore filters were not as effective asGelman filters in trapping hot TCA precipitates from highcell densities of E. coli.
Incorporation of [14C]pyruvate into protein. To investigatethe biosynthesis of leucine, we examined the incorporationof [3-_4C]pyruvate (New England Nuclear) into the hotTCA-insoluble fraction. The specific activity of the addedpyruvate was 15 mCi/mmol. The experimental protocol wassimilar to that described above for [3H]leucine incorpora-tion. After incubation of the water sample with [14C]pyruvate,the samples were extracted in hot 5% TCA for 30 min. Thehot TCA-insoluble material was collected on filters, whichwere either radioassayed directly or frozen for later TLCanalysis. The transformation of [14C]pyruvate to [14C]leucinewas examined by hydrolyzing the filters in 6 N HCl andseparating the amino acids by TLC as described for[3H]leucine incorporation. The proportion of [14C]pyruvateused for leucine biosynthesis was estimated from the amountof [14C]leucine recovered compared with the radioactivitysampled from the remaining sections of the TLC plate.We tested whether the presence of extracellular leucine
inhibited the biosynthesis of leucine. The proportion of[14C]pyruvate recovered as [14C]leucine from the hydrolyzedTCA-insoluble material was measured after incubation withdifferent concentrations of nonradioactive leucine along withthe [14C]pyruvate. The nonradioactive leucine was addedjust before addition of the [14C]pyruvate. The incubationtimes and concentration of [14C]pyruvate are given for eachexperiment below. Acid-killed controls were also analyzedand used to correct for abiotic uptake. One experimenttested for the effect of leucine on total uptake of[14C]pyruvate. In that experiment the subsamples werefiltered, rinsed twice with 3 ml of filter-sterilized seawater,and radioassayed.Data analysis. Several experiments measured the incorpo-
ration into protein of [3H]leucine in the presence of differentconcentrations of nonradioactive leucine. Since this is anal-ogous to the approach of Wright and Hobbie (36), wecalculated the maximum incorporation rate (Vmax) by usingstandard nonlinear parameter estimation techniques (18),which fit the equation of Wright and Hobbie directly to thedata without linear transformation. The lines drawn were fitby eye. RESULTS
Several parameters must be examined to demonstrate thata hot TCA extraction after incubation of a water sample with
VOL. 49, 1985 601
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.
602 KIRCHMAN, K'NEES, AND HODSON
TABLE 1. Proportion of bacterial assemblages that incorporatedleucine in three marine environments'
No. of %Environment [3H]leucine Turnover bacteria dpm Leucine-
concn (nM) (% h-1) ml-' cell-' active(x 105) bacteria
Tongue of the 0.5 5.9 1.1 0.0850 61Ocean
Bahamian 0.5 17 6.1 0.0200 53estuary
Sapelo Island 0.5 2.8 10 0.0066 3410 2.1 10 0.0740 63
"The incubation time was 2 h for samples from the Tongue of the Oceanand the Bahamian estuary. The incubation time for samples from SapeloIsland was 4 h. The interpolated values for the percentage of leucine-activebacteria in 2-h samples are 23% for 0.5 nM leucine and 53% for 10 nM leucine.This interpolation is based on data shown in Fig. 1.
[3H]leucine is an adequate method for measuring the rate ofprotein synthesis by bacteria in natural microbial assem-blages. Here we examined the following parameters: (i) theproportion of the total bacterial assemblage that assimilatesleucine, (ii) the degree of leucine degradation to othercompounds incorporated into the hot TCA-insoluble frac-tion, and (iii) de novo synthesis of leucine.
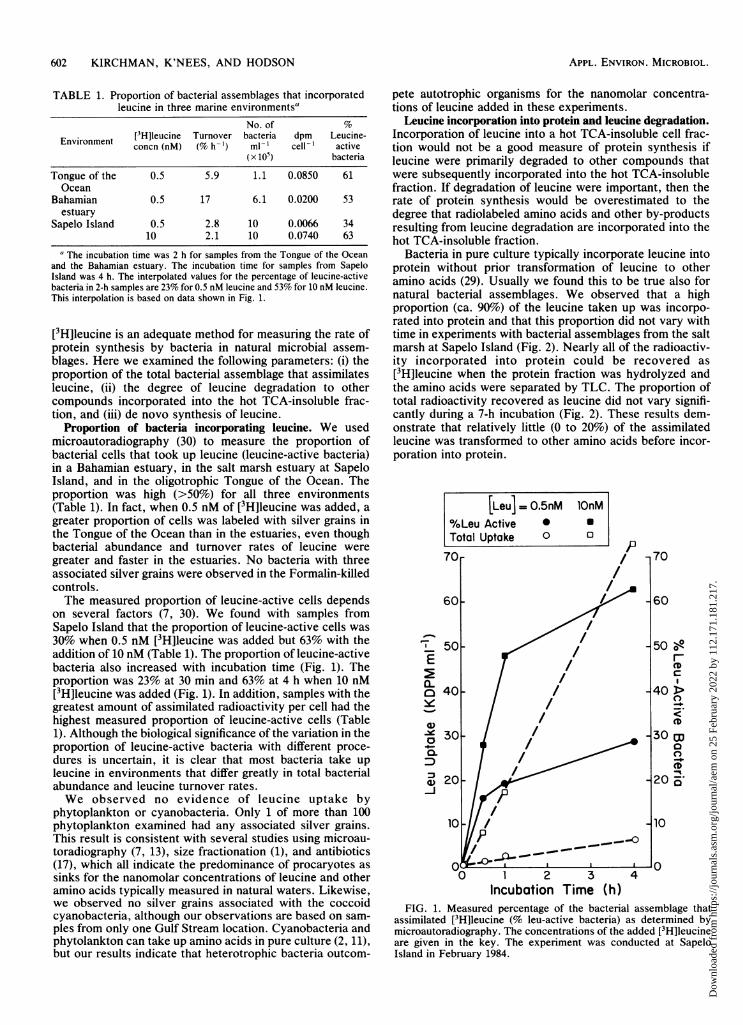
Proportion of bacteria incorporating leucine. We usedmicroautoradiography (30) to measure the proportion ofbacterial cells that took up leucine (leucine-active bacteria)in a Bahamian estuary, in the salt marsh estuary at SapeloIsland, and in the oligotrophic Tongue of the Ocean. Theproportion was high (>50%) for all three environments(Table 1). In fact, when 0.5 nM of [3H]leucine was added, agreater proportion of cells was labeled with silver grains inthe Tongue of the Ocean than in the estuaries, even thoughbacterial abundance and turnover rates of leucine weregreater and faster in the estuaries. No bacteria with threeassociated silver grains were observed in the Formalin-killedcontrols.The measured proportion of leucine-active cells depends
on several factors (7, 30). We found with samples fromSapelo Island that the proportion of leucine-active cells was30% when 0.5 nM [3H]leucine was added but 63% with theaddition of 10 nM (Table 1). The proportion of leucine-activebacteria also increased with incubation time (Fig. 1). Theproportion was 23% at 30 min and 63% at 4 h when 10 nM[3H]leucine was added (Fig. 1). In addition, samples with thegreatest amount of assimilated radioactivity per cell had thehighest measured proportion of leucine-active cells (Table1). Although the biological significance of the variation in theproportion of leucine-active bacteria with different proce-dures is uncertain, it is clear that most bacteria take upleucine in environments that differ greatly in total bacterialabundance and leucine turnover rates.We observed no evidence of leucine uptake by
phytoplankton or cyanobacteria. Only 1 of more than 100phytoplankton examined had any associated silver grains.This result is consistent with several studies using microau-toradiography (7, 13), size fractionation (1), and antibiotics(17), which all indicate the predominance of procaryotes assinks for the nanomolar concentrations of leucine and otheramino acids typically measured in natural waters. Likewise,we observed no silver grains associated with the coccoidcyanobacteria, although our observations are based on sam-ples from only one Gulf Stream location. Cyanobacteria andphytolankton can take up amino acids in pure culture (2, 11),but our results indicate that heterotrophic bacteria outcom-
pete autotrophic organisms for the nanomolar concentra-tions of leucine added in these experiments.
Leucine incorporation into protein and leucine degradation.Incorporation of leucine into a hot TCA-insoluble cell frac-tion would not be a good measure of protein synthesis ifleucine were primarily degraded to other compounds thatwere subsequently incorporated into the hot TCA-insolublefraction. If degradation of leucine were important, then therate of protein synthesis would be overestimated to thedegree that radiolabeled amino acids and other by-productsresulting from leucine degradation are incorporated into thehot TCA-insoluble fraction.
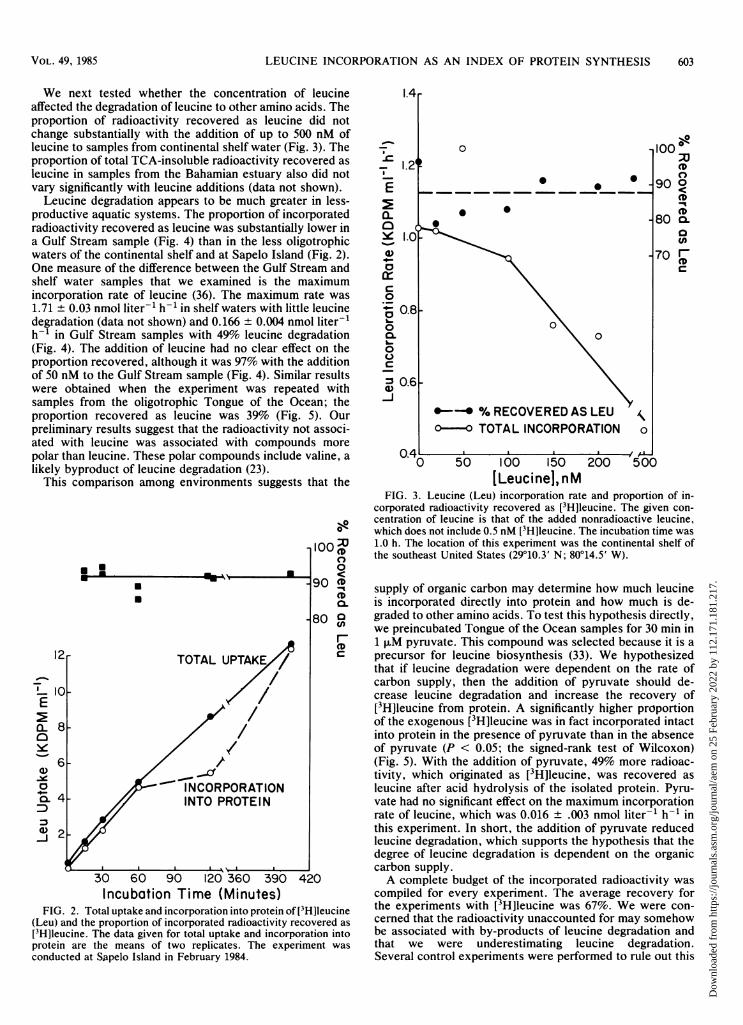
Bacteria in pure culture typically incorporate leucine intoprotein without prior transformation of leucine to otheramino acids (29). Usually we found this to be true also fornatural bacterial assemblages. We observed that a highproportion (ca. 90%) of the leucine taken up was incorpo-rated into protein and that this proportion did not vary withtime in experiments with bacterial assemblages from the saltmarsh at Sapelo Island (Fig. 2). Nearly all of the radioactiv-ity incorporated into protein could be recovered as[3H]leucine when the protein fraction was hydrolyzed andthe amino acids were separated by TLC. The proportion oftotal radioactivity recovered as leucine did not vary signifi-cantly during a 7-h incubation (Fig. 2). These results dem-onstrate that relatively little (0 to 20%) of the assimilatedleucine was transformed to other amino acids before incor-poration into protein.
[Leu] = 0.5nM lOnM%/OLeu Active 0*Total Uptake ° °
0-
E
L
0)0IC
0)1-J-
1 2 3Incubation Time (h)
70
60
50 orC
40 >0
(D30 C
00O.
20 a.
10
FIG. 1. Measured percentage of the bacterial assemblage thatassimilated [3H]leucine (% leu-active bacteria) as determined bymicroautoradiography. The concentrations of the added [3H]leucineare given in the key. The experiment was conducted at SapeloIsland in February 1984.
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.
LEUCINE INCORPORATION AS AN INDEX OF PROTEIN SYNTHESIS
We next tested whether the concentration of leucineaffected the degradation of leucine to other amino acids. Theproportion of radioactivity recovered as leucine did notchange substantially with the addition of up to 500 nM ofleucine to samples from continental shelf water (Fig. 3). Theproportion of total TCA-insoluble radioactivity recovered asleucine in samples from the Bahamian estuary also did notvary significantly with leucine additions (data not shown).
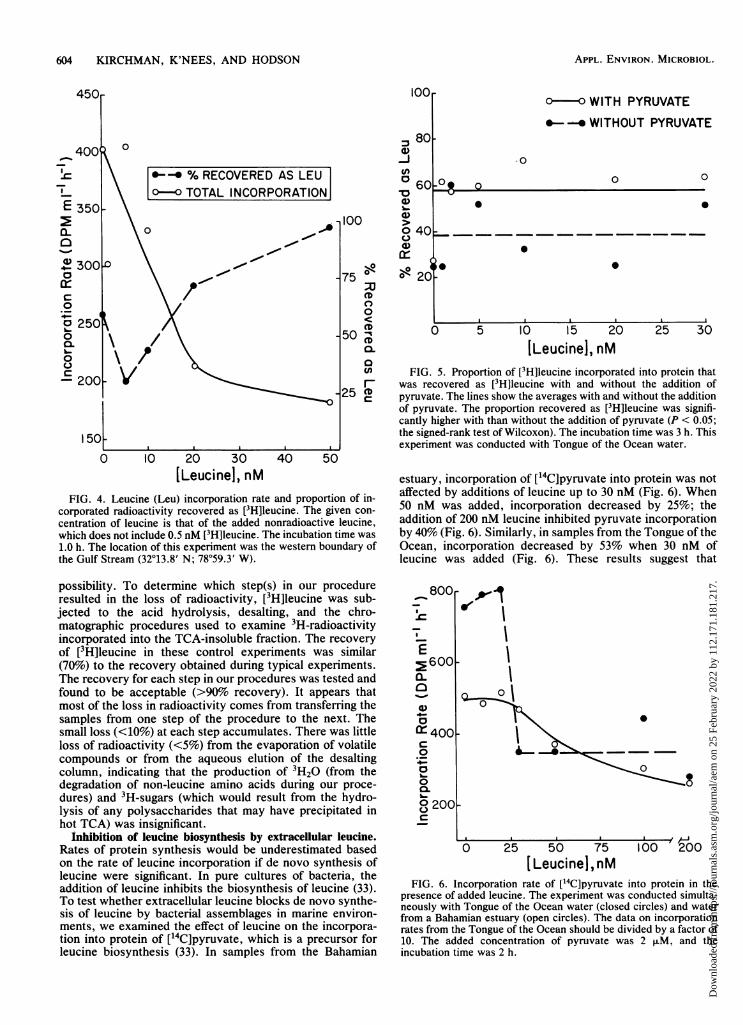
Leucine degradation appears to be much greater in less-productive aquatic systems. The proportion of incorporatedradioactivity recovered as leucine was substantially lower ina Gulf Stream sample (Fig. 4) than in the less oligotrophicwaters of the continental shelf and at Sapelo Island (Fig. 2).One measure of the difference between the Gulf Stream andshelf water samples that we examined is the maximumincorporation rate of leucine (36). The maximum rate was1.71 + 0.03 nmol liter-1 h-' in shelf waters with little leucinedegradation (data not shown) and 0.166 ± 0.004 nmol liter-'h-1 in Gulf Stream samples with 49% leucine degradation(Fig. 4). The addition of leucine had no clear effect on theproportion recovered, although it was 97% with the additionof 50 nM to the Gulf Stream sample (Fig. 4). Similar resultswere obtained when the experiment was repeated withsamples from the oligotrophic Tongue of the Ocean; theproportion recovered as leucine was 39% (Fig. 5). Ourpreliminary results suggest that the radioactivity not associ-ated with leucine was associated with compounds morepolar than leucine. These polar compounds include valine, alikely byproduct of leucine degradation (23).
This comparison among environments suggests that the
I14
E
Q-I
C
y
CD
-J
I100
1.4r
_ 1.2E
0j0-a:-
0._
O 0.80
0.Q..o0
3 0.6,1
OA
0
-* _* 90
0 0
0
*--@% RECOVERED AS L0-o TOTAL INCORPORATION 0
0s) 50 100 150 200 50)[Leucine] nM
100
80
00
CD0
70 rc
FIG. 3. Leucine (Leu) incorporation rate and proportion of in-corporated radioactivity recovered as [3H]leucine. The given con-centration of leucine is that of the added nonradioactive leucine,
°-e which does not include 0.5 nM [3H]leucine. The incubation time was; 1.0 h. The location of this experiment was the continental shelf ofm the southeast United States (29°10.3' N; 80014.5' W).v J0
-90 M
80 0rCDa
30 60 90 120 360 390 420Incubation Time (Minutes)
FIG. 2. Total uptake and incorporation into protein of [3H]leucine(Leu) and the proportion of incorporated radioactivity recovered as[3H]leucine. The data given for total uptake and incorporation intoprotein are the means of two replicates. The experiment wasconducted at Sapelo Island in February 1984.
supply of organic carbon may determine how much leucineis incorporated directly into protein and how much is de-graded to other amino acids. To test this hypothesis directly,we preincubated Tongue of the Ocean samples for 30 min in1 ,uM pyruvate. This compound was selected because it is aprecursor for leucine biosynthesis (33). We hypothesizedthat if leucine degradation were dependent on the rate ofcarbon supply, then the addition of pyruvate should de-crease leucine degradation and increase the recovery of[3H]leucine from protein. A significantly higher proportionof the exogenous [3H]leucine was in fact incorporated intactinto protein in the presence of pyruvate than in the absenceof pyruvate (P < 0.05; the signed-rank test of Wilcoxon)(Fig. 5). With the addition of pyruvate, 49% more radioac-tivity, which originated as [3H]leucine, was recovered asleucine after acid hydrolysis of the isolated protein. Pyru-vate had no significant effect on the maximum incorporationrate of leucine, which was 0.016 ± .003 nmol liter-' h-V inthis experiment. In short, the addition of pyruvate reducedleucine degradation, which supports the hypothesis that thedegree of leucine degradation is dependent on the organiccarbon supply.A complete budget of the incorporated radioactivity was
compiled for every experiment. The average recovery forthe experiments with [3HJleucine was 67%. We were con-cerned that the radioactivity unaccounted for may somehowbe associated with by-products of leucine degradation andthat we were underestimating leucine degradation.Several control experiments were performed to rule out this
VOL. 49, 1985 603
.f
Li I a I I , i-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.
604 KIRCHMAN, K'NEES, AND HODSON
0
0--e* % RECOVERED AS LEUo---o TOTAL INCORPORATION
\ 01,.A
/
V
10 20 30 40 50
IOC
75
50
25
100
= 80-J= 0 aIna 60 0-a0
0 40 _0
0- 820
0 20o
CD00
FIG. 5. Er was recoveC pc pyruvate. TI
of pyruvate,cantly highethe signed-r;experiment
o-o WITH PYRUVATE
0_- -WITHOUT PYRUVATE
0
.
0
0
0
0
5 10 15 20 25 30[Leucinel, nM
Proportion of [3H]leucine incorporated into protein thatbred as [3H]leucine with and without the addition ofhe lines show the averages with and without the additionThe proportion recovered as [3H]leucine was signifi-
zr with than without the addition of pyruvate (P < 0.05;*ank test of Wilcoxon). The incubation time was 3 h. Thiswas conducted with Tongue of the Ocean water.
[Leucinel, nMFIG. 4. Leucine (Leu) incorporation rate and proportion of in-
corporated radioactivity recovered as [3H]leucine. The given con-centration of leucine is that of the added nonradioactive leucine,which does not include 0.5 nM [3H]leucine. The incubation time was1.0 h. The location of this experiment was the westem boundary ofthe Gulf Stream (32°13.8' N; 78°59.3' W).
possibility. To determine which step(s) in our procedureresulted in the loss of radioactivity, [3H]leucine was sub-jected to the acid hydrolysis, desalting, and the chto-matographic procedures used to examine 3H-radioactivityincorporated into the TCA-insoluble fraction. The recoveryof [3H]leucine in these control experiments was similar(70%) to the recovery obtained during typical experiments.The recovery for each step in our procedures was tested andfound to be acceptable (>90% recovery). It appears thatmost of the loss in radioactivity comes from transferring thesamples from one step of the procedure to the next. Thesmall loss (<10%) at each step accumulates. There was littleloss of radioactivity (<5%) from the evaporation of volatilecompounds or from the aqueous elution of the desaltingcolumn, indicating that the production of 3H20 (from thedegradation of non-leucine amino acids during our proce-dures) and 3H-sugars (which would result from the hydro-lysis of any polysaccharides that may have precipitated inhot TCA) was insignificant.
Inhibition of leucine biosynthesis by extracellular leucine.Rates of protein synthesis would be underestimated basedon the rate of leucine incorporation if de novo synthesis ofleucine were significant. In pure cultures of bacteria, theaddition of leucine inhibits the biosynthesis of leucine (33).To test whether extracellular leucine blocks de novo synthe-sis of leucine by bacterial assemblages in marine environ-ments, we examined the effect of leucine on the incorpora-tion into protein of [14C]pyruvate, which is a precursor forleucine biosynthesis (33). In samples from the Bahamian
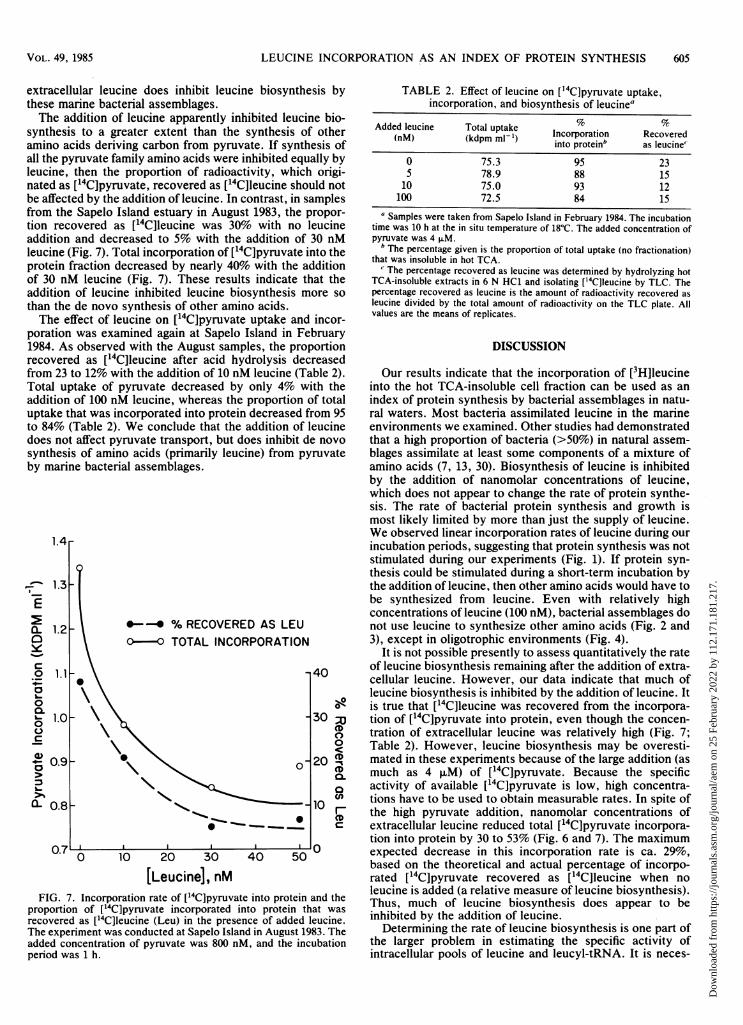
estuary, incorporation of ['4C]pyruvate into protein was notaffected by additions of leucine up to 30 nM (Fig. 6). When50 nM was added, incorporation decreased by 25%; theaddition of 200 nM leucine inhibited pyruvate incorporationby 40% (Fig. 6). Similarly, in samples from the Tongue of theOcean, incorporation decreased by 53% when 30 nM ofleucine was added (Fig. 6). These results suggest that
800r
-c
Em6000L0
0
CZ-0
W-400c0.
8 200
*--1
I
I
I
0
I
0 25 50 75 100 ''200
| Leucinel, nMFIG. 6. Incorporation rate of [14C]pyruvate into protein in the
presence of added leucine. The experiment was conducted simulta-neously with Tongue of the Ocean water (closed circles) and waterfrom a Bahamian estuary (open circles). The data on incorporationrates from the Tongue of the Ocean should be divided by a factor of10. The added concentration of pyruvate was 2 FM, and theincubation time was 2 h.
4001I
7
E 35001-
.? 3000
c03 250o
000
-E2001
I50L0
APPL. ENVIRON. MICROBIOL.
450r
I
I
/._l
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.
LEUCINE INCORPORATION AS AN INDEX OF PROTEIN SYNTHESIS
extracellular leucine does inhibit leucine biosynthesis bythese marine bacterial assemblages.The addition of leucine apparently inhibited leucine bio-
synthesis to a greater extent than the synthesis of otheramino acids deriving carbon from pyruvate. If synthesis ofall the pyruvate family amino acids were inhibited equally byleucine, then the proportion of radioactivity, which origi-nated as [14C]pyruvate, recovered as [14C]leucine should notbe affected by the addition of leucine. In contrast, in samplesfrom the Sapelo Island estuary in August 1983, the propor-tion recovered as ['4C]leucine was 30% with no leucineaddition and decreased to 5% with the addition of 30 nMleucine (Fig. 7). Total incorporation of ["4C]pyruvate into theprotein fraction decreased by nearly 40% with the additionof 30 nM leucine (Fig. 7). These results indicate that theaddition of leucine inhibited leucine biosynthesis more sothan the de novo synthesis of other amino acids.The effect of leucine on [14C]pyruvate uptake and incor-
poration was examined again at Sapelo Island in February1984. As observed with the August samples, the proportionrecovered as [14C]leucine after acid hydrolysis decreasedfrom 23 to 12% with the addition of 10 nM leucine (Table 2).Total uptake of pyruvate decreased by only 4% with theaddition of 100 nM leucine, whereas the proportion of totaluptake that was incorporated into protein decreased from 95to 84% (Table 2). We conclude that the addition of leucinedoes not affect pyruvate transport, but does inhibit de novosynthesis of amino acids (primarily leucine) from pyruvateby marine bacterial assemblages.
1.4r
1.3kI-
E
0-
0._
c
0CL-a)
0
1-
0~
1.2F _--4 % RECOVERED AS LEU0-O TOTAL INCORPORATION
1.11
1.0k
0.91
0.8k N%.
-.-%0~~~
0.7 0 10 20 30 40 50
[Leucine], nM
od2O
FIG. 7. Incorporation rate of ['4C]pyruvate into protein and theproportion of ['4C]pyruvate incorporated into protein that wasrecovered as [14C]leucine (Leu) in the presence of added leucine.The experiment was conducted at Sapelo Island in August 1983. Theadded concentration of pyruvate was 800 nM, and the incubationperiod was 1 h.
TABLE 2. Effect of leucine on ["4C]pyruvate uptake,incorporation, and biosynthesis of leucinea
Added leucine Total uptake % %(nM) (kdpm ml-') Incorporation Recoveredinto protein' as leucine'
0 75.3 95 235 78.9 88 1510 75.0 93 12
100 72.5 84 15
a Samples were taken from Sapelo Island in February 1984. The incubationtime was 10 h at the in situ temperature of 18°C. The added concentration ofpyruvate was 4 ,uM.
b The percentage given is the proportion of total uptake (no fractionation)that was insoluble in hot TCA.
' The percentage recovered as leucine was determined by hydrolyzing hotTCA-insoluble extracts in 6 N HC1 and isolating [14Cjleucine by TLC. Thepercentage recovered as leucine is the amount of radioactivity recovered asleucine divided by the total amount of radioactivity on the TLC plate. Allvalues are the means of replicates.
DISCUSSION
Our results indicate that the incorporation of [3H]leucineinto the hot TCA-insoluble cell fraction can be used as anindex of protein synthesis by bacterial assemblages in natu-ral waters. Most bacteria assimilated leucine in the marineenvironments we examined. Other studies had demonstratedthat a high proportion of bacteria (>50%) in natural assem-blages assimilate at least some components of a mixture ofamino acids (7, 13, 30). Biosynthesis of leucine is inhibitedby the addition of nanomolar concentrations of leucine,which does not appear to change the rate of protein synthe-sis. The rate of bacterial protein synthesis and growth ismost likely limited by more than just the supply of leucine.We observed linear incorporation rates of leucine during ourincubation periods, suggesting that protein synthesis was notstimulated during our experiments (Fig. 1). If protein syn-thesis could be stimulated during a short-term incubation bythe addition of leucine, then other amino acids would have tobe synthesized from leucine. Even with relatively highconcentrations of leucine (100 nM), bacterial assemblages donot use leucine to synthesize other amino acids (Fig. 2 and3), except in oligotrophic environments (Fig. 4).
It is not possible presently to assess quantitatively the rateof leucine biosynthesis remaining after the addition of extra-cellular leucine. However, our data indicate that much ofleucine biosynthesis is inhibited by the addition of leucine. Itis true that [14C]leucine was recovered from the incorpora-tion of [14C]pyruvate into protein, even though the concen-tration of extracellular leucine was relatively high (Fig. 7;Table 2). However, leucine biosynthesis may be overesti-mated in these experiments because of the large addition (asmuch as 4 p.M) of [14C]pyruvate. Because the specificactivity of available [14Clpyruvate is low, high concentra-tions have to be used to obtain measurable rates. In spite ofthe high pyruvate addition, nanomolar concentrations ofextracellular leucine reduced total [14C]pyruvate incorpora-tion into protein by 30 to 53% (Fig. 6 and 7). The maximumexpected decrease in this incorporation rate is ca. 29%,based on the theoretical and actual percentage of incorpo-rated [14C]pyruvate recovered as [14C]leucine when noleucine is added (a relative measure of leucine biosynthesis).Thus, much of leucine biosynthesis does appear to beinhibited by the addition of leucine.
Determining the rate of leucine biosynthesis is one part ofthe larger problem in estimating the specific activity ofintracellular pools of leucine and leucyl-tRNA. It is neces-
VOL. 49, 1985 605
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.

606 KIRCHMAN, K'NEES, AND HODSON
sary to know the specific activity of the leucyl-tRNA pool todirectly calculate the rate at which [3H]leucine is incorpo-
rated into protein (19). For a number of reasons this specificactivity would be difficult, if not impossible, to estimate forbacterial assemblages occurring in most aquatic systems. Itis doubtful that the pools in those microorganisms incorpo-rating leucine (primarily bacteria) could be distinguishedfrom the pools in microorganisms not incorporating leucine.Phytoplankton, which do not take up significant amounts ofamino acids (1, 7, 17) but which have amino acids pools (35),undoubtedly have larger internal leucine and leucyl-tRNApools than the bacteria, as the phytoplankton biomass ismuch larger than the bacterial biomass in the euphotic zone
of aquatic systems.To circumvent the specific activity problem, we have been
using an approach based on the suggestions of Moriarty andPollard (24) and Kirchman et al. (16) to measure a conver-
sion factor, which relates the incorporation rate of leucine tothe rate of protein synthesis. The results will be reported inmore detail elsewhere (D. Kirchman and R. Hodson, man-
uscript in preparation). Briefly, grazing activity by bacte-rivores, which usually prevents bacterial abundance fromincreasing significantly, is reduced by adding eucaryoticinhibitors (25) or by diluting the microbial assemblage withfiltered water (6, 16). Bacterial abundance and biomassincrease over time after the grazing activity has been re-
duced, if not eliminated. At various times subsamples are
withdrawn, and the incorporation rate of leucine is measuredas described by Moriarty and Pollard (24); the incorporationrate of [3H]leucine is measured in the presence of differentconcentrations of nonradioactive leucine. A conversion fac-tor is then calculated by using the equations described byKirchman et al. (16).
In addition to variation in the specific activity of leucyl-tRNA, another potential problem in using the rate of leucineincorporation to calculate rates of protein synthesis is pos-
sible variation in the amount of leucine in bacterial protein.However, the molar fraction of leucine in a wide variety ofproteins is relatively constant. Reeck and Fisher (28) com-
piled the reported values for the amino acid content of 207proteins. The leucine molar fraction of prokaryotic enzymes
is 8.8% (standard deviation [SD] = 2.1%); the leucinecontent of eucaryotic enzymes is 8.2% (SD = 2.0%). Theaverage leucine molar fraction given in a more recentcompilation of 61 bacterial proteins is 8.8% (SD = 2.0%)(27). These averages agree well with the best estimate of theleucine content in natural bacterial assemblages. The data ofHagstrom et al. (8) indicate that the leucine molar fraction ofa marine bacterial assemblage in continuous culture withunamended seawater was 9.1 and 8.3% at dilution rates of0.018 h-1 and 0.11 h-1, respectively. The important point isthat, based on the extensive data set on leucine content ofproteins studied in the laboratory, 95% of all proteins willhave a leucine content that varies on the average less than0.3% (a confidence interval based on n = 207 and SD =
2.1%) from the mean of 8.8%. Thus, variation in leucinecontent is very unlikely to cause significant variation in ratesof leucine incorporation and hence in the calculated rate ofprotein synthesis by natural bacterial assemblages.Because protein is a large proportion of bacterial biomass
(ca. 50%), the rate of protein synthesis should be a largefraction of the rate of total bacterial production. Rates ofleucine incorporation can be used to estimate bacterialbiomass production; the rate of leucine incorporation isdivided by the proportion of leucine in protein (8.8%) andthe amount of protein in bacterial cells (ca. 50%). However,
estimates of bacterial production based on protein synthesis(leucine incorporation) will equal estimates of productionbased on DNA synthesis (thymidine incorporation; seereferences 6 and 7) only during balanced growth. Rates ofprotein and DNA synthesis, normalized to the amount ofeach macromolecule per cell, are equal during balancedgrowth (14, 26). Whether the bacterial assemblage is inbalanced or unbalanced growth is an important question toaddress. The proportion of primary production processed bybacteria has been estimated from rates of DNA synthesisand by assuming that the uptake of organic compounds isclosely coupled to the incorporation of [3H]thymidine (5, 6),which is the case during balanced growth. Given that theconditions for bacterial growth change on small temporaland spatial scales, it is unclear what percentage of the timebacterial assemblages are in balanced growth. Althoughsome investigators have concluded that bacterial assem-blages in aquatic systems are in balanced growth (3, 9, 17),we believe that comparing rates of thymidine and leucineincorporation is a direct approach for examining the extentof balanced versus unbalanced growth. We have observedsignificant temporal and spatial differences between rates of[3H]thymidine incorporation into DNA and rates of[3H]leucine incorporation into protein (D. Kirchman, R.Murray, L. Pomeroy, and R. Hodson, manuscript in prepa-ration).
In addition to elucidating how well leucine incorporationmeasures bacterial protein synthesis, the results discussedhere increase our understanding of the relationships amongleucine uptake, leucine biosynthesis, and incorporation intoprotein. Our results demonstrate that bacterial assemblagescan quickly regulate the relative rates of leucine degradationand biosynthesis in response to the addition of organiccompounds. When pyruvate was added, the amount ofleucine degraded to other amino acids decreased on a timescale shorter than the presumptive growth rate of the bacte-ria. Similarly, leucine biosynthesis decreased quickly (<1 h)when leucine was added. In both cases, the bacteria appar-ently maximize their utilization of extracellular leucine forprotein synthesis, rather than synthesizing leucine de novoor utilizing leucine for the synthesis of other amino acids.Experiments such as those described here give insight intothe regulation of bacterial metabolism in response to changesin the dissolved organic carbon pool.
ACKNOWLEDGMENTSWe thank D. Monticello for his advice on methods to examine
leucine metabolism. We also thank L. Pomeroy for allowing us toparticipate on cruises of the Cape Hatteras.
This work was supported in part by National Science Foundationgrants BSR 8215587 and OCE 8117834 and by National Oceano-graphic and Atmospheric Administration sea grant NA 80AA-D-00091.
LITERATURE CITED1. Azam, F., and R. E. Hodson. 1977. Size distribution and activity
of marine microheterotrophs. Limnol. Oceanogr. 22:492-501.2. Chapman, J. S., and J. C. Meeks. 1983. Glutamine and gluta-
mate transport by Anabaena variabilis. J. Bacteriol. 156:122-129.
3. Cuhel, R. L., H. W. Jannasch, and C. D. Taylor. 1983. Microbialgrowth and macromolecular synthesis in the northwestern At-lantic ocean. Limnol. Oceanogr. 28:1-18.
4. Cuhel, R. L., C. D. Taylor, and H. W. Jannasch. 1982. Assim-ilatory sulfur metabolism in marine microorganisms: consider-ations for the application of sulfate incorporation into protein asa measurement of natural population protein synthesis. Appl.Environ. Microbiol. 43:160-168.
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.

LEUCINE INCORPORATION AS AN INDEX OF PROTEIN SYNTHESIS
5. Ducklow, H. W., and D. Kirchman. 1983. Bacterial dynamicsand distribution during a spring diatom bloom in the HudsonRiver Plume. J. Plankton Res. 5:333-555.
6. Fuhrman, J. A., and F. Azam. 1980. Bacterioplankton second-ary production estimates for coastal waters of British Columbia,Antarctica, and California. Appl. Environ. Microbiol. 39:1085-1095.
7. Fuhrman, J. A., and F. Azam. 1982. Thymidine incorporation asa measure of heterotrophic bacterioplankton production inmarine surface waters: evaluation and field results. Mar. Biol.66:109-120.
8. Hagstrom, A., J. A. Ammerman, S. Henrichs, and F. Azam.Bacterioplankton growth in seawater. II. Organic matter utili-zation during steady-state growth. Mar. Ecol. Prog. Ser.18:41-48.
9. Hanson, R. B., and H. K. Lowery. 1983. Nucleic acid synthesisin oceanic microplankton from the Drake Passage, Antarctica:evaluation of steady-state growth. Mar. Biol. 73:79-89.
10. Hanson, R. S., and J. A. Phillips. 1981. Chemical composition,p. 328-364. In P. Gerhardt, R. G. E. Murray, R. N. Costilow,E. W. Nester, W. A. Wood, N. R. Kreig, and G. B. Phillips(ed.), Manual of methods for general bacteriology. AmericanSociety for Microbiology, Washington, D.C.
11. Hellebust, J. A., and J. Lewin. 1977. Heterotrophic nutrition, p.169-197. In D. Werner (ed.), The biology of diatoms. Universityof California Press, Berkeley.
12. Hobbie, J. E., R. J. Daley, and S. Jasper. 1977. Use ofNuclepore filters for counting bacteria by fluorescence micros-copy. Appl. Environ. Microbiol. 33:1225-1228.
13. Hoppe, H. G. 1976. Determination and properties of activelymetabolizing heterotrophic bacteria in the sea, investigated bymeans of microautoradiography. Mar. Biol. 36:291-302.
14. Ingraham, J. L., 0. Maal$e, and F. C. Neidhardt. 1983. Growthof the bacterial cell. Sinauer Association, Sunderland, Mass.
15. Jordan, M. J., and G. E. Likens. 1980. Measurement of plank-tonic bacterial production in an oligotrophic lake. Limnol.Oceanogr. 25:719-732.
16. Kirchman, D., H. W. Ducklow, and R. Mitchell. 1982. Estimatesof bacterial growth from changes in uptake rates and biomass.Appl. Environ. Microbiol. 44:1296-1307.
17. Kirchman, D., and R. Hodson. 1984. Inhibition by peptides ofamino acid uptake by bacterial populations in natural waters:implications for the regulation of amino acid transport andincorporation. Appl. Environ. Microbiol. 47:624-631.
18. Li, W. K. W. 1983. Consideration of errors in estimating kineticparameters based on Michaelis-Menten formalism in microbialecology. Limnol. Oceanogr. 28:185-190.
19. Matin, A. F., and M. Rabinowitz. 1977. Measurement of half-lifeof rat cardiac myosin heavy chain with leucyl-tRNA used asprecursor pool. J. Biol. Chem. 252:3422-3429.
20. McBride, R. W., D. W. Jolly, B. M. Kadis, and T. E. Nelson.1979. Rapid thin-layer chromatographic separation of isoleu-cine, leucine, and phenylalanine. J. Chromatogr. 168:290-291.
21. Meyer-Reil, L. A. 1978. Autoradiography and epifluorescencemicroscopy combined for the determination of number andspectrum of actively metabolizing bacteria in natural waters.
Appl. Environ. Microbiol. 36:506-512.22. Mitra, R. S. 1984. Protein synthesis in Escherichia coli during
recovery from exposure to low levels of Cd2+. Appl. Environ.Microbiol. 47:1012-1016.
23. Monticello, D. J., and R. N. Costilow. Interconversion of valineand leucine by Clostridium sporogenes. J. Bacteriol.152:946-949.
24. Moriarty, D. J. W., and P. C. Pollard. 1981. DNA synthesis asa measure of bacterial productivity in seagrass sediments. Mar.Ecol. Prog. Ser. 5:151-156.
25. Newell, S. Y., B. F. Sherr, E. B. Sherr, and R. D. Fallon. 1983.Bacterial response to presence of eukaryote inhibitors in waterfrom a coastal marine environment. Mar. Environ. Res.10:147-157.
26. Pritchard, R. H., and D. W. Tempest. 1982. Growth: cells andpopulations, p. 99-123. In J. Mandelstam, K. Mcquillen, and I.Dawes (ed.), Biochemistry of bacterial growth. John Wiley &Sons, Inc., New York.
27. Reeck, G. R. 1983. Amino acid compositions of selected pro-teins, p. 15-29. In A. I. Laskin and H. A. Lechevalier (ed.),Handbook of microbiology, vol. 2. Microbial composition. CRCPress, Inc., Boca Raton, Fla.
28. Reeck, G. R., and L. Fisher. 1973. A statistical analysis of theamino acid compositions of proteins. Int. J. Peptide ProteinRes. 5:109-117.
29. Roberts, R. B., D. B. Cowie, R. H. Abelson, E. T. Bolton, andR. J. Britten. 1963. Studies of biosynthesis in Escherichia coli.Carnegie Institution of Washington publication no. 607. Car-negie Institute of Washington, Washington, D.C.
30. Tabor, P. S., and R. A. Neihof. 1982. Improved microautoradi-ographic method to determine individual microorganisms activein substrate uptake in natural waters. Appl. Environ. Microbiol.44:945-953.
31. Tenore, K. R., R. B. Hanson, B. E. Dornseif, and C. N.Wiederhold. 1979. The effect of organic nitrogen supplement onthe utilization of different sources of detritus. Limnol. Oce-anogr. 24:350-355.
32. Turnbough, C. L. 1983. Regulation of Escherichia coli aspartatetranscarboamylase synthesis by guanosine tetraphosphate andpyrimidine ribonucleoside triphosphates. J. Bacteriol. 153:998-1007.
33. Umbarger, H. E. 1978. Amino acid biosynthesis and its regula-tion. Annu. Rev. Biochem. 47:533-606.
34. Watson, S. W., T. J. Novitsky, H. L. Quinby, and F. W. Valois.1977. Determination of bacterial number and biomass in themarine environment. Appl. Environ. Microbiol. 33:940-946.
35. Wheeler, P. W. 1983. Phytoplankton nitrogen metabolism, p.309-346. In E. Carpenter and D. Capone (ed.), Nitrogen in themarine environment. Academic Press, Inc., New York.
36. Wright, R. T., and J. E. Hobbie. 1965. Uptake of organic solutesin lake water. Limnol. Oceanogr. 10:22-28.
37. Yamamori, T., K. Ito, Y. Nakamura, and T. Yura. 1978.Transient regulation of protein synthesis in Escherichia coliupon shift-up of growth temperature. J. Bacteriol. 134:1133-1140.
VOL. 49, 1985 607
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 25
Feb
ruar
y 20
22 b
y 11
2.17
1.18
1.21
7.





























