Embed Size (px)
Citation preview

ORIGINAL RESEARCH PAPER
Life history traits of Blaptostethus pallescens (Hemiptera:Anthocoridae), a candidate for use in augmentative biologicalcontrol in Egypt
Islam S. Sobhy • Amany M. Abdul-Hamid •
Awad A. Sarhan • Ahmed A. Shoukry •
Nasser S. Mandour • Stuart R. Reitz
Received: 24 December 2013 / Accepted: 12 February 2014 / Published online: 5 March 2014
� The Japanese Society of Applied Entomology and Zoology 2014
Abstract Blaptostethus pallescens Poppius (Hemiptera:
Anthocoridae) is an abundant native predator in mango
orchards and other cropping systems in Egypt. To deter-
mine suitable mass-rearing conditions for this little-studied
species, we assessed some of its biological characteristics.
Testing its thermal response at three constant temperatures
(20, 25, 30 �C), showed that immature development time
and adult longevity decreased with increasing temperature.
Reproductive success of individual females was greatest
when reared at 25 �C (84.3 ± 3.1 eggs) rather than at
20 �C (46.6 ± 2.0 eggs) or 30 �C (65.2 ± 2.5 eggs).
Although B. pallescens reared at 25 �C had a significantly
higher net reproductive rate (R0), which may be attributed
to their relatively rapid development and high fecundity,
we argue that 30 �C seems to be more convenient for
rearing B. pallescens, as mean generation time (T) and
doubling time (DT) are clearly shorter, thus more indi-
viduals could be reared per unit of time at 30 �C. Mating
significantly reduced male and female longevity, as
unmated adults lived 25–45 % longer than mated individ-
uals did. Unmated females did not lay eggs, suggesting that
mating is a prerequisite for egg maturation. Adult males
and females performed best, in terms of longevity, when
fed Ephestia kuehniella Zeller (Lepidoptera: Pyralidae)
eggs instead of non-prey diets. However, diets of plant sap
or pollen could sustain adults in times of limited egg
availability. Because its biology is similar to that of other
subtropical anthocorids already reared for augmentative
releases, B. pallescens may be amenable to mass-rearing
using already established techniques. Therefore, B. pal-
lescens could be used to improve augmentative biological
control in crops such as mango or maize in Egypt where it
already naturally occurs, and therefore would not engender
concerns over non-target effects that an exotic, generalist
biological control agent would.
Keywords Temperature � Mating � Non-prey foods � Life
table � Anthocorids mass rearing
Introduction
Because of emerging concerns regarding the potential
negative effects on non-target species and ecosystems (De
Clercq 2002), the use of exotic biological control agents
has been subjected to greater regulation and more intense
risk assessments, which have restricted new uses of such
agents (van Lenteren et al. 2006).
One alternative to the use of exotic natural enemies is to
enhance the use of indigenous natural enemies in biological
control. Indigenous natural enemies provide certain
advantages because of their ability to readily exploit native
or invasive pests as their prey, but also to persist on
alternate prey when target pests are rare or absent (Sy-
mondson et al. 2002). Nevertheless, to improve the use of
indigenous natural enemies in biological control will
require additional information on their basic biology and
I. S. Sobhy (&) � A. M. Abdul-Hamid �A. A. Sarhan � A. A. Shoukry � N. S. Mandour
Department of Plant Protection, Public Service Center for
Biological Control (PSCBC), Faculty of Agriculture, Suez Canal
University, Ismailia 41522, Egypt
e-mail: [email protected]
I. S. Sobhy
Plant-Insect Interactions Group, Institute of Plant Science and
Resources, Okayama University, Kurashiki 710-0046, Japan
S. R. Reitz
Department of Crop and Soil Sciences, Oregon State University,
710 SW 5th Ave, Ontario, OR 97914, USA
123
Appl Entomol Zool (2014) 49:315–324
DOI 10.1007/s13355-014-0252-4

life history traits so that their mass rearing may be opti-
mized (Oida and Kadono 2012).
In the Mediterranean Basin and sub-Saharan Africa,
several anthocorid species have been reported as important
natural enemies in various cropping systems (Hernandez
and Stonedahl 1999). One of these species of interest is
Blaptostethus pallescens Poppius, a sub-tropical species
that can be abundant in grain warehouses in Egypt where
mites are common (Tawfik and El-Husseini 1971). Blapt-
ostethus pallescens may also suppress populations of cer-
tain pests of maize. There, it preys on eggs and young
larvae of lepidopteran pests, including Chilo agamemnon
Bleszynski, Ostrinia nubilalis (Hubner) and Spodoptera
littoralis (Boisduval), and it is also known to prey on
certain sucking insect pests, such as Rhopalosiphum maidis
Fitch, and some species of Tetranychus spider mites
(Tawfik and El-Husseini 1971). Blaptostethus pallescens can
also be abundant in mango (Mangifera indica L.) orchards
where it preys on larvae and eggs of the honey dew moth,
Cryptoblabes gnidiella (Milliere), which is a key pest of this
extremely valuable cash crop in Egypt (Sarhan, unpublished
data). Ballal et al. (2012) found that B. pallescens could feed
on two species of mealybug. Its wide range of prey renders B.
pallescens as an ideal candidate for mass rearing and aug-
mentative releases in a subtropical area, yet very little is
known of its basic biological traits.
Comprehensive knowledge of anthocorid predators
actually extends to only a few species of a few genera (e.g.,
Anthocoris, Orius and Xylocoris) that have been used
successfully in biological control programs (Lattin 1999).
Given that basic biological characteristics have been crit-
ical for the development of mass-rearing protocols for
these species (Grenier and De Clercq 2003; van Lenteren
2012), it is therefore important to investigate other antho-
corid species that may be amenable to mass rearing and
successful use in biological control programs.
We therefore conducted this study to investigate some
basic biological characteristics of B. pallescens. We
determined the effects of temperature on the development
and reproduction of B. pallescens. In addition, we inves-
tigated the impact of mating on adult longevity under
various temperatures, as well as the influence of non-prey
food on adult longevity. These findings will be useful for
developing mass-rearing programs for this promising an-
thocorid biological control agent.
Materials and methods
Stock culture of B. pallescens
A colony of B. pallescens was established from nymphs
and adults collected from mango inflorescences at the
Experimental Farm, Faculty of Agriculture, Suez Canal
University in the Ismailia Governorate, Egypt (30�360Nlatitude and 32�240E longitude). Adults and nymphs were
maintained in 1 l plastic transparent jars (10 cm diame-
ter 9 20 cm height), which were covered with muslin held
in place by rubber bands. Each jar was provided with
sufficient quantities of fresh, loose Ephestia kuehniella
Zeller (Lepidoptera: Pyralidae) eggs as a food supply,
which is a commonly used diet for anthocorid predators
(Schmidt et al. 1995). Ephestia kuehniella eggs were pro-
vided from the mass-rearing line at the Public Service
Centre for Biological Control (PSCBC), Faculty of Agri-
culture, Suez Canal University. Ephestia kuehniella were
reared on a wheat germ-based diet. A green bean pod
(Phaseolus vulgaris L.) was provided in each jar as an
oviposition substrate (Isenhour and Yeargan 1981). Bean
pods with newly deposited eggs were removed and
replaced daily and kept in the previously described plastic
jars. Jars were examined daily for emergence of B. pal-
lescens nymphs. Soon after hatching, nymphs were care-
fully transferred to new plastic jars and were provisioned
with E. kuehniella eggs, and small styrofoam balls to offer
hiding places and reduce cannibalism (Sobhy et al. 2010).
Field-collected adults and nymphs were added on a regular
basis to refresh the colony and to increase its genetic var-
iation (Leon-Beck and Coll 2009). Upon eclosion, adults
were sexed and placed in new plastic jars, supplied with the
same type of prey and oviposition substrates. Colonies
were kept in climatic chambers maintained at 25 ± 1 �C,
70 ± 10 % relative humidity, and an L16:D8 photoperiod.
Experiment 1: effect of temperature on preimaginal
development
Three different constant temperature regimes were tested:
20, 25, and 30 �C. Climatic chambers for this experiment
were maintained within ±1 �C of the test temperature, and
at 70 ± 10 % relative humidity and an L16:D8
photoperiod.
Sections of bean pod with newly deposited B. pallescens
eggs (0–12 h) were kept individually in small petri dishes
(9 cm diameter 9 1.5 cm height) and maintained under
each of the tested temperatures. Each container was
inspected daily to determine when nymphs emerged, and
subsequently, the incubation period for eggs. The incuba-
tion period was considered as the time from oviposition
until the nymphs were enclosed.
Upon eclosion, nymphs (0–6 h old) were separated
individually into the above-described petri dishes with the
use of a small, fine-hair brush. According to the availability
of newly hatched nymphs, there were 41–49 replicates for
each temperature treatment. Each nymph was provided
initially with 100 (&0.250 mg) loose, fresh eggs of E.
316 Appl Entomol Zool (2014) 49:315–324
123

kuehniella. This amount was increased by an additional 50
E. kuehniella eggs after each molt. These increases in the
amount of prey were based on preliminary tests to ensure
that sufficient prey were available to predators. A piece of
paper towel was placed on the bottom of the test arenas to
facilitate locomotion by the nymphs (Yanik and Unlu
2011). Every 12 h, the containers were inspected to
determine survivorship and timing of molting by B. pal-
lescens nymphs until adulthood and the subsequent sex
ratio of adults.
Experiment 2: effect of temperature on female
oviposition and longevity
Fecundity and longevity were determined for females
emerging from the immature development tests. Adults
were tested at the same temperature at which they were
reared as nymphs (20, 25, and 30 ± 1 �C; 70 ± 10 %
relative humidity; L16:D8 photoperiod). Newly emerged
adults were paired (one female with one male), and indi-
vidual pairs were placed separately in petri dishes (9 cm
diameter 9 1.5 cm height) for copulation. Six hours after
introduction, males were removed so that the responses of
females could be determined without interference from
males. There were 24–34 replicates per temperature
treatment.
Females were supplied daily with bean pods as ovipo-
sition sites and an excess of fresh E. kuehniella eggs as
prey, until death. Bean pods containing deposited eggs
were replaced daily. The numbers of eggs laid daily were
counted under a stereoscope (209). These bean pods were
then held under the same environmental conditions as the
females and were inspected daily under a stereoscope to
determine the number of hatched eggs. Hatched eggs were
identified by the opened visible operculum. Female lon-
gevity was recorded.
Experiment 3: effect of temperature and mating
on adult longevity
To ensure that individuals did not mate prior to the
experiment, 5th instars were placed individually in petri
dishes, as described above, and these nymphs were provi-
sioned with bean pods and an excess of fresh E. kuehniella
eggs until they reached adulthood. Upon adult eclosion,
males and females were kept individually isolated for the
unmated cohorts. To obtain mated B. pallescens, individual
males and females were placed together for mating pairs.
After six hours of pairing males and females, the males
were removed and separated to other petri dishes so that the
longevity of individual mated predators could be deter-
mined as was done for the unmated ones. Excess fresh E.
kuehniella eggs were provided as prey; fresh bean pods
were added to all arenas, as both a moisture source and
oviposition substrate. Bean pods were inspected under a
stereoscope (209) to determine the fecundity of mated and
virgin females. Containers were examined daily for survi-
vorship and for calculating the length of the oviposition
periods. Experiments were conducted under three constant
temperatures (20, 25, and 30 ± 1 �C; 70 ± 10 % relative
humidity; L16:D8 photoperiod). There were 36–38 repli-
cates per temperature treatment.
Experiment 4: effect of alternative foods on adult
longevity
To assess potential adult food sources for use in mass-
rearing programs, we investigated the effects of plant sap,
pollen, honey, water, and E. kuehniella eggs on the lon-
gevity of both males and females. A starvation treatment
without food or water was added as a control. Blaptostethus
pallescens used in the experiments were provisioned with
an excess of E. kuehniella eggs during their nymphal
development. Newly emerged adults were paired for 6 h
for copulation. Pairs (n = 20) were then separated to
individual petri dishes to determine longevity for each sex.
Adults were assigned to different food sources at ran-
dom. In this experiment, no oviposition substrates were
provided, except in the plant sap treatment where bean
pods were added to provide plant sap (Bonte et al. 2012).
For both water and honey treatments, saturated cotton
wicks were placed into the adult containers and replaced
daily. For the pollen treatment, 1.5 mg of dried pollen from
Egyptian clover (Trifolium alexandrinum L.) was sprinkled
into petri dishes on a regular basis. Pollen was supplied
from the Plant Protection Research Institute (PPRI), Agri-
cultural Research Center (ARC), El-Dokki, Egypt. Fresh E.
kuehniella eggs were added daily in excess, as previously
described. Experiments were maintained under 25 ± 1 �C,
70 ± 10 % relative humidity, and an L16:D8 photoperiod.
Statistical analyses
In the first experiment, the development time data for each
nymphal instar were analyzed using the non-parametric
Kruskal–Wallis analysis of variance (ANOVA) on ranks,
and Dunn’s method was subsequently used to compare
treatment means. Data for overall nymphal stage devel-
opment time and total preimaginal development time pas-
sed normality tests (Shapiro–Wilk); therefore, one-way
ANOVA was used for these analyses, and the Holm–Sidak
method was used for all pairwise multiple comparisons.
Data on female fecundity were analyzed by one-way
ANOVA after passing normality tests (Shapiro–Wilk), and
treatment means were compared using the Holm–Sidak
method.
Appl Entomol Zool (2014) 49:315–324 317
123

Two-way ANOVAs were conducted to evaluate the
effects of mating and temperature levels on male and
female longevity. Where no interaction was found, means
were separated using a Tukey’s test. When interactions
were significant, pairwise multiple comparison procedures
(Holm–Sidak method) were used.
In the non-prey food effect experiment, data were het-
eroscedastic (according to the Levene test); therefore, the
data were analyzed using the Kruskal–Wallis ANOVA
(H test), and means were compared pairwise with Dunn’s
method. These analyses were performed with a SigmaPlot
12.3 (SYSTAT Inc., Chicago, IL, USA).
Percentage data (nymphal survival, sex ratio, and egg
hatchability) were examined with generalized linear mod-
els (GLM) fitted by maximum quasi-binomial estimation in
the software package R (R Development Core Team 2009).
Life-table parameters were estimated for B. pallescens
at each temperature regime, using the data obtained for
survivorship and the age-specific fecundity of adults along
with the survivorship and development of all immature
stages of B. pallescens. Parameter estimates for the net
reproductive rate (R0), mean generation time (T), intrinsic
rate of increase (rm), and the finite rate of increase (k) were
calculated according to the description of Birch (1948),
whereas doubling time (DT) was calculated according to
Kairo and Murphy (1995) as follows:
R0 ¼X
lxmx;
T ¼X
xlxmx=X
lxmx;
rm ¼ ln R0=T ;
k ¼ exp (rmÞ;
DT ¼ ln2=rm;
where x is the age of female (days), lx is the survivorship at
the corresponding time, and mx is age-specific fecundity.
The life-table parameters were estimated on the basis of
female sex ratio.
Results
Effect of temperature on preimaginal development
Generally, the developmental time of B. pallescens was
significantly faster as temperatures increased. Development
was over 1.5-fold faster at 25 �C than at 20 �C, and over
1.5-fold faster at 30 �C than at 25 �C (Table 1). Significant
differences were found in developmental time for all B. Ta
ble
1D
evel
op
men
tti
mes
for
imm
atu
rest
ages
of
Bla
pto
stet
hu
sp
all
esce
ns
rear
edo
nE
ph
esti
aku
ehn
iell
aeg
gs
atth
ree
con
stan
tte
mp
erat
ure
sw
ith
a1
6L
:8D
ph
oto
per
iod
Tem
per
atu
re
(�C
)
Incu
bat
ion
per
iod
(day
s)
nL
eng
tho
fn
ym
ph
alst
ages
(day
s)A
To
tal
ny
mp
hal
per
iod
To
tal
pre
imag
inal
per
iod
(day
s)
Sex
rati
o
(fem
ale,
%)
1st
2n
d3
rd4
th5
th
20
9.2
6±
0.4
9a
41
8.4
2±
0.5
4a
6.7
5±
0.4
3a
4.1
9±
0.4
1a
5.9
4±
0.5
6a
7.4
3±
0.7
1a
34
.00
±1
.24
a4
3.3
4±
1.1
8a
45
.45
±0
.43
a
25
5.5
3±
0.5
3b
49
5.1
8±
0.3
9b
3.8
1±
0.3
1b
3.2
9±
0.2
3a
4.5
7±
0.3
9a
5.2
6±
0.4
1b
22
.32
±0
.74
b2
7.7
4±
0.8
7b
64
.1±
0.5
5a
30
3.9
0±
0.3
1c
46
3.3
5±
0.2
6c
2.5
6±
0.2
3c
2.0
8±
0.1
8b
2.9
4±
0.2
3b
3.5
2±
0.3
1c
14
.59
±0
.51
c1
8.5
0±
0.6
2c
58
.82
±0
.56
a
*V
alu
esar
ep
rese
nte
das
mea
n±
SE
AM
ean
sw
ith
inea
chco
lum
nfo
llo
wed
by
the
sam
ele
tter
are
no
tsi
gn
ifica
ntl
yd
iffe
ren
tac
cord
ing
toD
un
n’s
met
ho
dfo
rin
cub
atio
np
erio
dan
dd
iffe
ren
tn
ym
ph
alin
star
s,o
rb
ased
on
the
Ho
lm–
Sid
akte
stfo
rto
tal
ny
mp
hal
and
pre
imag
inal
per
iod
s(p
[0
.05
)
318 Appl Entomol Zool (2014) 49:315–324
123
pallescens instars among the tested temperature regimes
(first instar: H2 = 53.78, p \ 0.001; second: H2 = 45.49,
p \ 0.001; third: H2 = 26.13, p \ 0.001; fourth: H2 = 22.43,
p \ 0.001, fifth: H2 = 25.59, p \ 0.001, and total nymphal
stage: F2,94 = 133.54, p \ 0.001). The total preimaginal
period was drastically affected by temperature (F2,92 = 167.6,
p = 0.001; Table 1). Sex ratios did not differ from unity, and
the sex ratios of adults did not differ significantly according to
temperature (F2,92 = 0.9748, p = 0.3811; Table 1).
Blaptostethus pallescens nymphs were able to complete
their development at all tested temperature regimes, but there
were significant differences in nymphal survivorship to
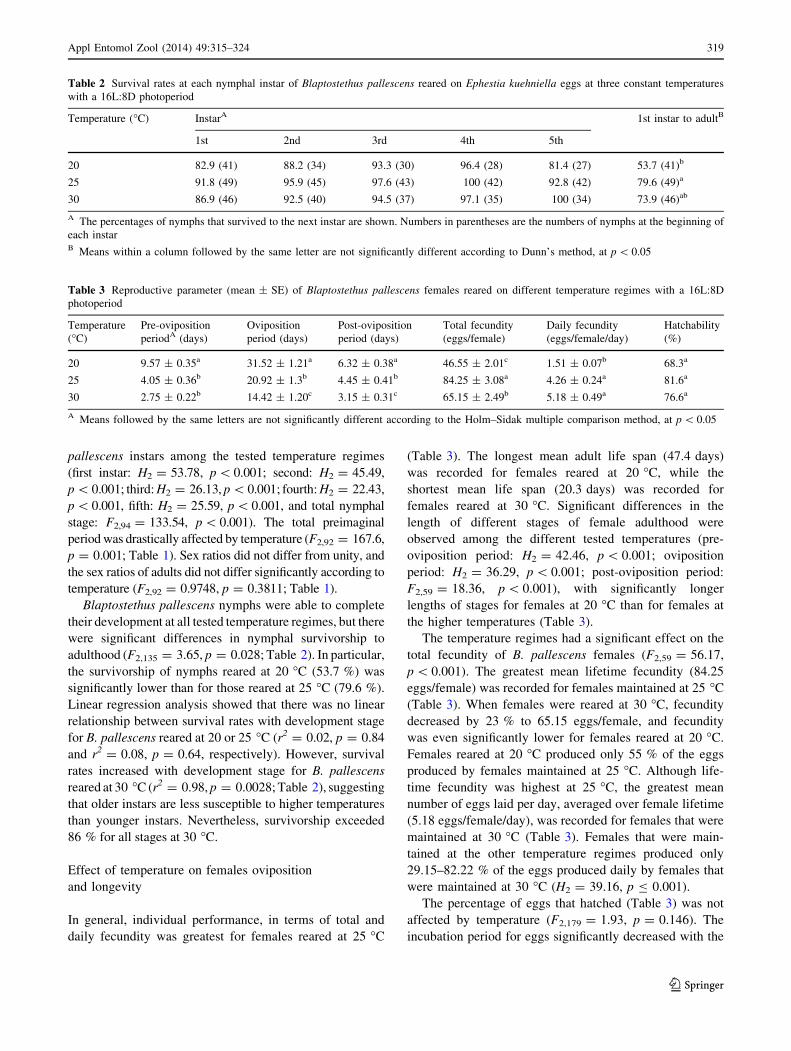
adulthood (F2,135 = 3.65, p = 0.028; Table 2). In particular,
the survivorship of nymphs reared at 20 �C (53.7 %) was
significantly lower than for those reared at 25 �C (79.6 %).
Linear regression analysis showed that there was no linear
relationship between survival rates with development stage
for B. pallescens reared at 20 or 25 �C (r2 = 0.02, p = 0.84
and r2 = 0.08, p = 0.64, respectively). However, survival
rates increased with development stage for B. pallescens
reared at 30 �C (r2 = 0.98, p = 0.0028; Table 2), suggesting
that older instars are less susceptible to higher temperatures
than younger instars. Nevertheless, survivorship exceeded
86 % for all stages at 30 �C.
Effect of temperature on females oviposition
and longevity
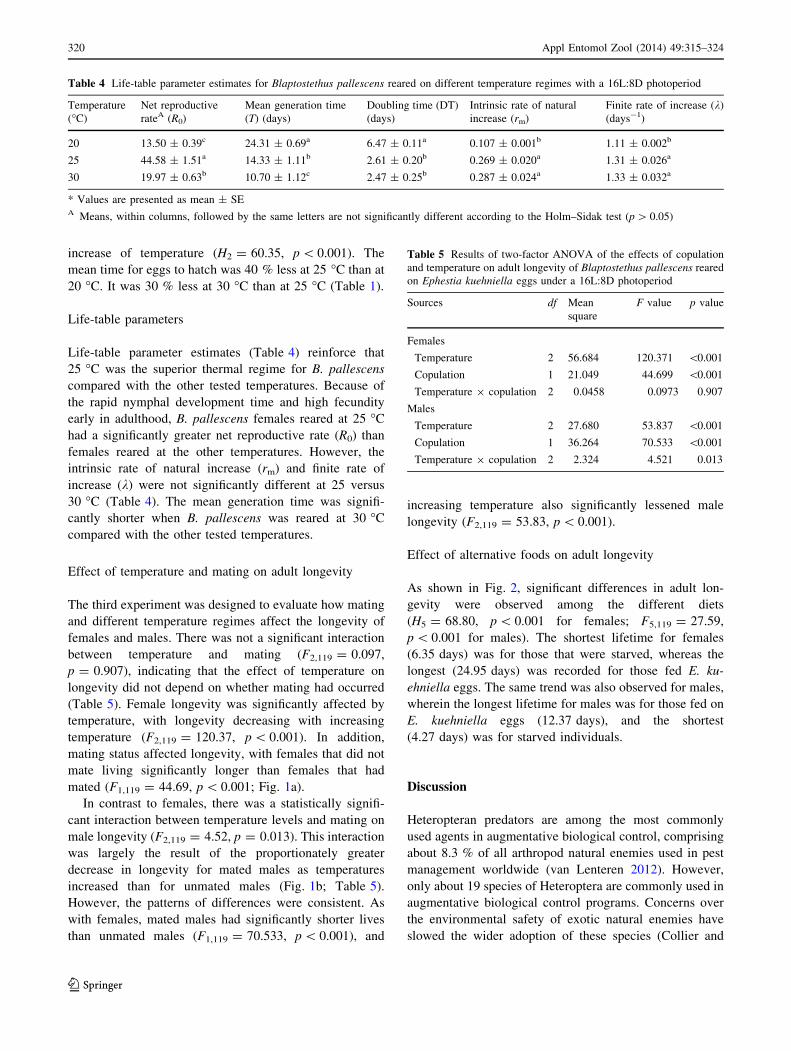
In general, individual performance, in terms of total and
daily fecundity was greatest for females reared at 25 �C
(Table 3). The longest mean adult life span (47.4 days)
was recorded for females reared at 20 �C, while the
shortest mean life span (20.3 days) was recorded for
females reared at 30 �C. Significant differences in the
length of different stages of female adulthood were
observed among the different tested temperatures (pre-
oviposition period: H2 = 42.46, p \ 0.001; oviposition
period: H2 = 36.29, p \ 0.001; post-oviposition period:
F2,59 = 18.36, p \ 0.001), with significantly longer
lengths of stages for females at 20 �C than for females at
the higher temperatures (Table 3).
The temperature regimes had a significant effect on the
total fecundity of B. pallescens females (F2,59 = 56.17,
p \ 0.001). The greatest mean lifetime fecundity (84.25
eggs/female) was recorded for females maintained at 25 �C
(Table 3). When females were reared at 30 �C, fecundity
decreased by 23 % to 65.15 eggs/female, and fecundity
was even significantly lower for females reared at 20 �C.
Females reared at 20 �C produced only 55 % of the eggs
produced by females maintained at 25 �C. Although life-
time fecundity was highest at 25 �C, the greatest mean
number of eggs laid per day, averaged over female lifetime
(5.18 eggs/female/day), was recorded for females that were
maintained at 30 �C (Table 3). Females that were main-
tained at the other temperature regimes produced only
29.15–82.22 % of the eggs produced daily by females that
were maintained at 30 �C (H2 = 39.16, p B 0.001).
The percentage of eggs that hatched (Table 3) was not
affected by temperature (F2,179 = 1.93, p = 0.146). The
incubation period for eggs significantly decreased with the
Table 2 Survival rates at each nymphal instar of Blaptostethus pallescens reared on Ephestia kuehniella eggs at three constant temperatures
with a 16L:8D photoperiod
Temperature (�C) InstarA 1st instar to adultB
1st 2nd 3rd 4th 5th
20 82.9 (41) 88.2 (34) 93.3 (30) 96.4 (28) 81.4 (27) 53.7 (41)b
25 91.8 (49) 95.9 (45) 97.6 (43) 100 (42) 92.8 (42) 79.6 (49)a
30 86.9 (46) 92.5 (40) 94.5 (37) 97.1 (35) 100 (34) 73.9 (46)ab
A The percentages of nymphs that survived to the next instar are shown. Numbers in parentheses are the numbers of nymphs at the beginning of
each instarB Means within a column followed by the same letter are not significantly different according to Dunn’s method, at p \ 0.05
Table 3 Reproductive parameter (mean ± SE) of Blaptostethus pallescens females reared on different temperature regimes with a 16L:8D
photoperiod
Temperature
(�C)
Pre-oviposition
periodA (days)
Oviposition
period (days)
Post-oviposition
period (days)
Total fecundity
(eggs/female)
Daily fecundity
(eggs/female/day)
Hatchability
(%)
20 9.57 ± 0.35a 31.52 ± 1.21a 6.32 ± 0.38a 46.55 ± 2.01c 1.51 ± 0.07b 68.3a
25 4.05 ± 0.36b 20.92 ± 1.3b 4.45 ± 0.41b 84.25 ± 3.08a 4.26 ± 0.24a 81.6a
30 2.75 ± 0.22b 14.42 ± 1.20c 3.15 ± 0.31c 65.15 ± 2.49b 5.18 ± 0.49a 76.6a
A Means followed by the same letters are not significantly different according to the Holm–Sidak multiple comparison method, at p \ 0.05
Appl Entomol Zool (2014) 49:315–324 319
123
increase of temperature (H2 = 60.35, p \ 0.001). The
mean time for eggs to hatch was 40 % less at 25 �C than at
20 �C. It was 30 % less at 30 �C than at 25 �C (Table 1).
Life-table parameters
Life-table parameter estimates (Table 4) reinforce that
25 �C was the superior thermal regime for B. pallescens
compared with the other tested temperatures. Because of
the rapid nymphal development time and high fecundity
early in adulthood, B. pallescens females reared at 25 �C
had a significantly greater net reproductive rate (R0) than
females reared at the other temperatures. However, the
intrinsic rate of natural increase (rm) and finite rate of
increase (k) were not significantly different at 25 versus
30 �C (Table 4). The mean generation time was signifi-
cantly shorter when B. pallescens was reared at 30 �C
compared with the other tested temperatures.
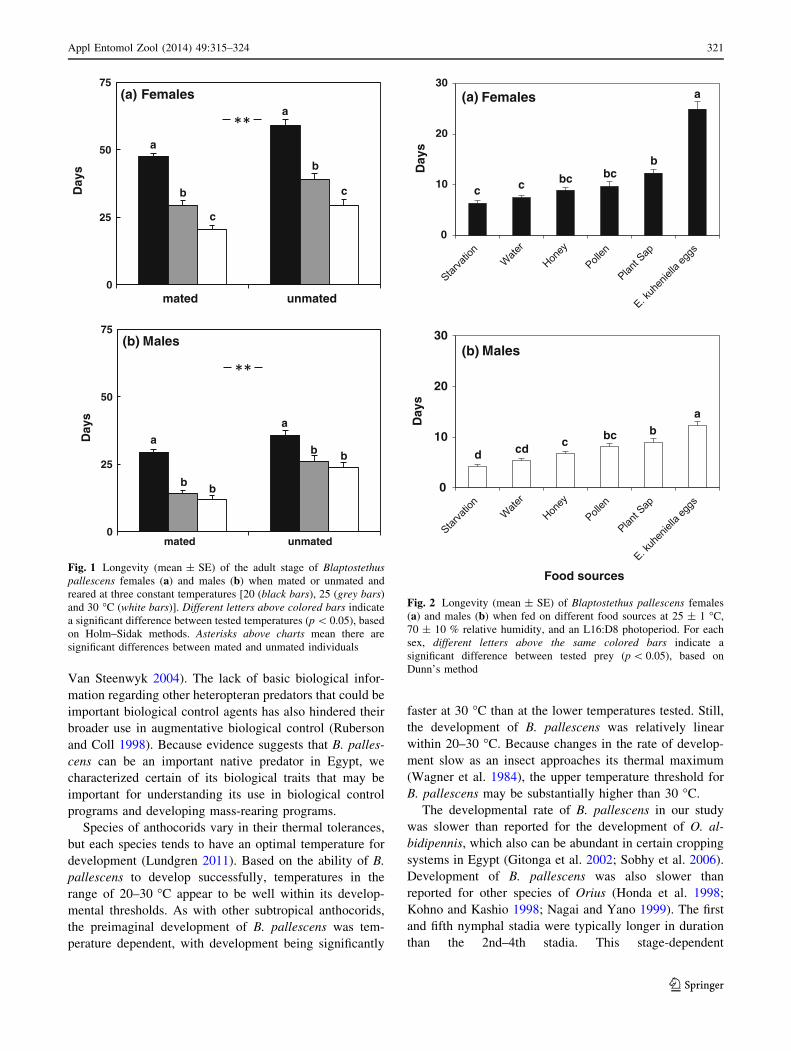
Effect of temperature and mating on adult longevity
The third experiment was designed to evaluate how mating
and different temperature regimes affect the longevity of
females and males. There was not a significant interaction
between temperature and mating (F2,119 = 0.097,
p = 0.907), indicating that the effect of temperature on
longevity did not depend on whether mating had occurred
(Table 5). Female longevity was significantly affected by
temperature, with longevity decreasing with increasing
temperature (F2,119 = 120.37, p \ 0.001). In addition,
mating status affected longevity, with females that did not
mate living significantly longer than females that had
mated (F1,119 = 44.69, p \ 0.001; Fig. 1a).
In contrast to females, there was a statistically signifi-
cant interaction between temperature levels and mating on
male longevity (F2,119 = 4.52, p = 0.013). This interaction
was largely the result of the proportionately greater
decrease in longevity for mated males as temperatures
increased than for unmated males (Fig. 1b; Table 5).
However, the patterns of differences were consistent. As
with females, mated males had significantly shorter lives
than unmated males (F1,119 = 70.533, p \ 0.001), and
increasing temperature also significantly lessened male
longevity (F2,119 = 53.83, p \ 0.001).
Effect of alternative foods on adult longevity
As shown in Fig. 2, significant differences in adult lon-
gevity were observed among the different diets
(H5 = 68.80, p \ 0.001 for females; F5,119 = 27.59,
p \ 0.001 for males). The shortest lifetime for females
(6.35 days) was for those that were starved, whereas the
longest (24.95 days) was recorded for those fed E. ku-
ehniella eggs. The same trend was also observed for males,
wherein the longest lifetime for males was for those fed on
E. kuehniella eggs (12.37 days), and the shortest
(4.27 days) was for starved individuals.
Discussion
Heteropteran predators are among the most commonly
used agents in augmentative biological control, comprising
about 8.3 % of all arthropod natural enemies used in pest
management worldwide (van Lenteren 2012). However,
only about 19 species of Heteroptera are commonly used in
augmentative biological control programs. Concerns over
the environmental safety of exotic natural enemies have
slowed the wider adoption of these species (Collier and
Table 4 Life-table parameter estimates for Blaptostethus pallescens reared on different temperature regimes with a 16L:8D photoperiod
Temperature
(�C)
Net reproductive
rateA (R0)
Mean generation time
(T) (days)
Doubling time (DT)
(days)
Intrinsic rate of natural
increase (rm)
Finite rate of increase (k)
(days-1)
20 13.50 ± 0.39c 24.31 ± 0.69a 6.47 ± 0.11a 0.107 ± 0.001b 1.11 ± 0.002b
25 44.58 ± 1.51a 14.33 ± 1.11b 2.61 ± 0.20b 0.269 ± 0.020a 1.31 ± 0.026a
30 19.97 ± 0.63b 10.70 ± 1.12c 2.47 ± 0.25b 0.287 ± 0.024a 1.33 ± 0.032a
* Values are presented as mean ± SEA Means, within columns, followed by the same letters are not significantly different according to the Holm–Sidak test (p [ 0.05)
Table 5 Results of two-factor ANOVA of the effects of copulation
and temperature on adult longevity of Blaptostethus pallescens reared
on Ephestia kuehniella eggs under a 16L:8D photoperiod
Sources df Mean
square
F value p value
Females
Temperature 2 56.684 120.371 \0.001
Copulation 1 21.049 44.699 \0.001
Temperature 9 copulation 2 0.0458 0.0973 0.907
Males
Temperature 2 27.680 53.837 \0.001
Copulation 1 36.264 70.533 \0.001
Temperature 9 copulation 2 2.324 4.521 0.013
320 Appl Entomol Zool (2014) 49:315–324
123
Van Steenwyk 2004). The lack of basic biological infor-
mation regarding other heteropteran predators that could be
important biological control agents has also hindered their
broader use in augmentative biological control (Ruberson
and Coll 1998). Because evidence suggests that B. palles-
cens can be an important native predator in Egypt, we
characterized certain of its biological traits that may be
important for understanding its use in biological control
programs and developing mass-rearing programs.
Species of anthocorids vary in their thermal tolerances,
but each species tends to have an optimal temperature for
development (Lundgren 2011). Based on the ability of B.
pallescens to develop successfully, temperatures in the
range of 20–30 �C appear to be well within its develop-
mental thresholds. As with other subtropical anthocorids,
the preimaginal development of B. pallescens was tem-
perature dependent, with development being significantly
faster at 30 �C than at the lower temperatures tested. Still,
the development of B. pallescens was relatively linear
within 20–30 �C. Because changes in the rate of develop-
ment slow as an insect approaches its thermal maximum
(Wagner et al. 1984), the upper temperature threshold for
B. pallescens may be substantially higher than 30 �C.
The developmental rate of B. pallescens in our study
was slower than reported for the development of O. al-
bidipennis, which also can be abundant in certain cropping
systems in Egypt (Gitonga et al. 2002; Sobhy et al. 2006).
Development of B. pallescens was also slower than
reported for other species of Orius (Honda et al. 1998;
Kohno and Kashio 1998; Nagai and Yano 1999). The first
and fifth nymphal stadia were typically longer in duration
than the 2nd–4th stadia. This stage-dependent
0
25
50
75
mated unmated
Femalesa
b
c
a
b
c
0
25
50
75
mated unmated
Males
a
b ba
b b
Day
sD
ays
(a)
(b)
Fig. 1 Longevity (mean ± SE) of the adult stage of Blaptostethus
pallescens females (a) and males (b) when mated or unmated and
reared at three constant temperatures [20 (black bars), 25 (grey bars)
and 30 �C (white bars)]. Different letters above colored bars indicate
a significant difference between tested temperatures (p \ 0.05), based
on Holm–Sidak methods. Asterisks above charts mean there are
significant differences between mated and unmated individuals
0
10
20
30
ab
cd cd
bc
Males
0
10
20
30
b
a
c bc bcc
Females
Food sources
Day
sD
ays
(a)
(b)
Fig. 2 Longevity (mean ± SE) of Blaptostethus pallescens females
(a) and males (b) when fed on different food sources at 25 ± 1 �C,
70 ± 10 % relative humidity, and an L16:D8 photoperiod. For each
sex, different letters above the same colored bars indicate a
significant difference between tested prey (p \ 0.05), based on
Dunn’s method
Appl Entomol Zool (2014) 49:315–324 321
123

developmental pattern is typical of other anthocorids (Is-
enhour and Yeargan 1981; Sanchez and Lacasa 2002).
Despite the slower development of B. pallescens, its
survivorship to adulthood was far superior to that of O.
albidipennis or other species of Orius (Cocuzza et al. 1997;
Gitonga et al. 2002; Nagai and Yano 1999). Survivorship
was [80 % for all nymphal stages at all tested tempera-
tures. Often, anthocorids experience high mortality in the
first stadium (Cocuzza et al. 1997; Honda et al. 1998;
Nagai and Yano 1999).
Females had the greatest individual reproductive success
at 25 �C. The preoviposition period for females reared at
this temperature was not significantly longer than the pre-
oviposition period of females reared at 30 �C. Therefore,
females reared at either temperature could begin laying
eggs at similar times and lay similar numbers of eggs each
day. However, the greater longevity of females reared at
25 �C would enable these to produce more offspring. In
fact, lifetime fecundity was 1.3–1.8 times higher at 25 �C
than at 30 or 20 �C.
However, the selection of temperatures for mass-rearing
programs must also consider the effect of temperature on
the overall reproductive output of a colony rather than that
of individual insects. The longevity of B. pallescens
females decreased with increasing temperature from 20 to
30 �C, which is in accordance with that recorded for O.
albidipennis adults fed on highly nutritious prey such as
Frankliniella occidentalis (Pergande) (Cocuzza et al. 1997)
or E. kuehniella eggs (Sobhy et al. 2006). This pattern of
decreasing longevity with increasing temperature has also
been found for other species of Orius (Carvalho et al. 2005;
Gitonga et al. 2002). Despite the shorter adult longevity, a
substantially greater number of predators could be pro-
duced per unit of time in mass-rearing programs at 30 �C
than at the lower tested temperatures. Based on our results,
each female would produce 17.1 adult progeny at 20 �C,
54.7 at 25 �C, and 36.9 at 30 �C. Yet, approximately 17
generations of B. pallescens could develop in 1 year at
30 �C, compared with approximately 11 generations at
25 �C and only seven generations at 20 �C. Therefore, the
population growth rate would be substantially greater at
30 �C than at the lower temperatures.
Mating status had significant effects on the longevity of
B. pallescens females and males. Unmated males lived for
6–12 days longer than mated males, depending on tem-
perature. Honda et al. (1998) found that unmated females
of Orius minutus (L.) and O. sauteri (Poppius) lived over
40 % longer than their mated counterparts. Male Hemip-
tera often transfer a significant amount of their body mass
in the form of nuptial gifts, i.e., sperm, associated body
fluids, as well as some nutrients in their spermatophore to
females during mating, and this loss of body resources may
shorten male longevity (Krupke et al. 2008). The mean life
span of unmated females was 9–12 days longer than the
life span of mated ones at the three tested temperatures,
which represents a 25–45 % increase in longevity.
Based on our observations, B. pallescens females that
had mated once would vigorously resist subsequent mating
attempts, which is consistent with previous (Tawfik and El-
Husseini 1971) findings. Although this resistance may be
explained by the presence of mating inhibitors in the male
ejaculate (Arnqvist and Nilsson 2000; Yamane et al. 2011),
females of B. pallescens may benefit by avoiding multiple
matings. Anthocorids are known to mate by traumatic
insemination (Schuh and Stys 1991). This form of mating
may pose risks to females from wounds or injury (Stutt and
Siva-Jothy 2001), which may reduce female lifespan.
Alternatively, energy invested in egg maturation and ovi-
position may reduce female longevity (Simmons and
Kotiaho 2007).
Mating status not only affected the longevity of B.
pallescens females, but we also found that virgin females
did not lay eggs. This finding is consistent to what has been
reported for other anthocorids (Honda et al. 1998; Ito and
Nakata 1998; Leon-Beck and Coll 2009). Shapiro and
Shirk (2010) observed that no fully mature eggs were
present in the ovaries of unmated females of Orius pumilio
(Champion). Horton et al. (2005) found that delays in
mating for three species of Anthocoris (Anthocoridae) led
to delays in oocyte development and caused a prolongation
of the preoviposition period. However, this delay in oocyte
development would not be beneficial for mass production.
Therefore, a near-unity sex ratio would help to assure that
females mate soon after eclosion.
Mass-rearing techniques of natural enemies can be fur-
ther optimized by reducing costs and labor inputs during
the rearing process (van Lenteren and Tommasini 2003).
Ephestia kuehniella eggs are routinely used in rearing an-
thocorids for augmentative release because of their high
nutritional value (Bonte and De Clercq 2011; Ferkovich
et al. 2007). However, E. kuehniella eggs are expensive
because of the cost of maintaining colonies of this insect.
Hence, using alternative foods such as artificial diets or
non-prey nutrition instead of E. kuehniella eggs, which are
routinely used in the commercial production, might reduce
the cost of the rearing process, as the market price for E.
kuehniella eggs still remains high (De Clercq et al. 2005).
In our study, we focused on the influence of different
diets on adult life span. Starved females died after only
6.35 days, which would be only about 2 days after the end
of their preoviposition period. However, starved females
are not likely to be able to produce any eggs (Shapiro and
Shirk 2010). Given that females provisioned with water did
not survive longer than starved females, it appears the lack
of nutrients rather than simply desiccation was responsible
for the short adult longevity. Honey, pollen, and plant sap
322 Appl Entomol Zool (2014) 49:315–324
123

may have provided limited amounts of necessary nutrients,
but none were nearly as nutritious as E. kuehniella eggs.
The high quality of E. kuehniella eggs is likely related to
their relatively high nitrogen content (Ferkovich et al.
2007). Plant sap, honey, and pollen can supplement
arthropod prey to achieve better rearing quality, but are not
sufficient alone for the propagation of colonies of B. palles-
cens (Chin-Ling et al. 1999; Kiman and Yeargan 1985).
However, these materials are readily available and less
expensive than E. kuehniella eggs, and thus they may be used
to provisionally sustain populations of B. pallescens in the
event that there is a temporary shortage of E. kuehniella eggs.
Our findings provide data on the basic biology of B.
pallescens and potential mass-rearing conditions for this
predator. We found that E. kuehniella eggs are a suitable
food for rearing nymphs and adults of B. pallescens.
Temperature levels substantially impact the reproductive
performance and survivorship of B. pallescens. Given our
results, a temperature of approximately 30 �C would
maximize the production of colonies for mass-rearing
purposes. Unquestionably, more studies should be under-
taken in terms of predation rates and population growth,
which are important for forecasting the role of B. palles-
cens as a biological control agent. However, the successful
rearing of this predator may provide an opportunity for it to
be used in augmentative biological control programs in its
native range, crops such as mango and maize.
Acknowledgments The authors would like to deeply thank Prof.
M.F.S. Tawfik, Faculty of Agriculture, Cairo University, Egypt, for
his help in the identification of Blaptostethus pallescens individuals
that were used to build up our stock colony. We acknowledge Lesley
Smart, Rothamsted Research, for her comments on the earlier version
of the manuscript. This work was supported by the Public Service
Centre for Biological Control (PSCBC), Faculty of Agriculture, Suez
Canal University, Ismailia, Egypt.
References
Arnqvist G, Nilsson T (2000) The evolution of polyandry: multiple
mating and female fitness in insects. Anim Behav 60:145–164
Ballal CR, Gupta T, Joshi S (2012) Predatory potential of two
indigenous anthocorid predators on Phenacoccus solenopsis
Tinsley and Paracoccus marginatus Williams and Granara de
Willink. J Biol Control 26:18–22
Birch LC (1948) The intrinsic rate of natural increase of an insect
population. J Anim Ecol 17:15–26
Bonte M, De Clercq P (2011) Influence of predator density, diet and
living substrate on developmental fitness of Orius laevigatus.
J Appl Entomol 135:343–350
Bonte J, Vangansbeke D, Maes S, Bonte M, Conlong D, De Clercq P
(2012) Moisture source and diet affect development and
reproduction of Orius thripoborus and Orius naivashae, two
predatory anthocorids from southern Africa. J Insect Sci 12:1
(available online: insectscience.org/12.1)
Carvalho LM, Bueno VHP, Mendes SM (2005) Influence of
temperature on the reproduction and longevity of the predator
Orius thyestes Herring (Hemiptera, Anthocoridae). Rev Bras
Entomol 49:409–414
Chin-Ling W, Yen-Jong W, Mon-Yu H, Ching-Tien Y, Yin-Fu C
(1999) Selection of proper food materials for rearing Orius
strigicollis (Poppius) (Hemiptera: Anthocoridae). Chin J Ento-
mol 19:319–329
Cocuzza GE, De Clercq P, Lizzio S, Van De Veire M, Tirry L,
Degheele D, Vacante V (1997) Life tables and predation activity
of Orius laevigatus and O. albidipennis at three constant
temperatures. Entomol Exp Appl 85:189–198
Collier T, Van Steenwyk R (2004) A critical evaluation of augmen-
tative biological control. Biol Control 31:245–256
De Clercq P (2002) Dark clouds and their silver linings: exotic
generalist predators in augmentative biological control. Neotrop
Entomol 31:169–176
De Clercq P, Arijs Y, Van Meir T, Van Stappen G, Sorgeloos P,
Dewettinck K, Rey M, Grenier S, Febvay G (2005) Nutritional
value of brine shrimp cysts as a factitious food for Orius
laevigatus (Heteroptera: Anthocoridae). Biocontrol Sci Technol
15:467–479
Ferkovich SM, Venkatesan T, Shapiro JP, Carpenter JE (2007)
Presentation of artificial diet: effects of composition and size of
prey and diet domes on egg production by Orius insidiosus
(Heteroptera: Anthocoridae). Fla Entomol 90:502–508
Gitonga LM, Lohr B, Overholt WA, Magambo JK, Mueke JM (2002)
Effect of temperature on the development of Orius albidipennis
Reuter, a predator of the African legume flower thrips, Megalu-
rothrips sjostedti trybom. Insect Sci Appl 22:215–220
Grenier S, De Clercq P (2003) Comparison of artificially vs. naturally
reared natural enemies and their potential for use in biological
control. In: van Lenteren JC (ed) Quality control and production
of biological control agent. CAB International, Wallingford,
pp 115–131
Hernandez LM, Stonedahl GM (1999) A review of the economically
important species of the genus Orius (Heteroptera: Anthocori-
dae) in East Africa. J Nat His 33:543–568
Honda JY, Nakashima Y, Hirose Y (1998) Development, reproduc-
tion and longevity of Orius minutus and Orius sauteri (Het-
eroptera: Anthocoridae) when reared on Ephestia kuehniella
eggs. Appl Entomol Zool 33:449–453
Horton DR, Lewis TM, Neven LG (2005) Ovarian development and
lipid reserves are affected by mating delays in three species of
Anthocoris (Hemiptera: Anthocoridae). Can Entomol
137:328–336
Isenhour DJ, Yeargan KV (1981) Effect of temperature on the
development of Orius insidiosus, with notes on laboratory
rearing. Ann Entomol Soc Am 74:114–116
Ito K, Nakata T (1998) Effect of photoperiod on reproductive
diapause in the predatory bugs, Orius sauteri (Poppius) and O.
minutus (Linnaeus) (Heteroptera: Anthocoridae). Appl Entomol
Zool 33:115–120
Kairo MTK, Murphy ST (1995) The life history of Rodolia iceryae
Janson (Coleoptera: Coccinellidae) and the potential for use in
innoculative release against Icerya pattersoni Newstead (Ho-
moptera: Margarodidae) on coffee. J Appl Entomol 119:487–491
Kiman ZB, Yeargan KV (1985) Development and reproduction of the
predator Orius insidiosus (Hemiptera, Anthocoridae) reared on
diets of selected plant material and arthropod prey. Ann Entomol
Soc Am 78:464–467
Kohno K, Kashio T (1998) Development and prey consumption of
Orius sauteri (Poppius) and O. minutus (L.) (Heteroptera:
Anthocoridae) fed on Frankliniella occidentalis (Pergande)
(Thysanoptera: Thripidae). Appl Entomol Zool 33:227–230
Krupke CH, Brunner JF, Jones VP (2008) Factors influencing mate
choice in Euschistus conspersus Uhler (Heteroptera: Pentatom-
idae). Environ Entomol 37:192–197
Appl Entomol Zool (2014) 49:315–324 323
123

Lattin JD (1999) Bionomics of the Anthocoridae. Annu Rev Entomol
44:207–231
Leon-Beck M, Coll M (2009) The mating system of the flower bug
Orius laevigatus. Biol Control 50:199–203
Lundgren JG (2011) Reproductive ecology of predaceous Heterop-
tera. Biol Control 59:37–52
Nagai K, Yano E (1999) Effects of temperature on the development
and reproduction of Orius sauteri (Poppius) (Heteroptera:
Anthocoridae), a predator of Thrips palmi Karny (Thysanoptera:
Thripidae). Appl Entomol Zool 34:223–230
Oida H, Kadono F (2012) Development of Geocoris varius and G.
proteus (Hemiptera: Geocoridae) provided with Ephestia ku-
ehniella (Lepidoptera: Pyralidae) eggs. Appl Entomol Zool
47:365–372
R Development Core Team (2009) R: a language and environment for
statistical computing, 2.9.1 edn. R Foundation for Statistical
Computing, Vienna. http://www.R-project.org
Ruberson JR, Coll M (1998) Research needs for predatory Heterop-
tera. In: Coll M, Ruberson JR (eds) Predatory Heteroptera: their
ecology and use in biological control. Entomological Society of
America, Lanham, pp 225–233
Sanchez JA, Lacasa A (2002) Modelling population dynamics of
Orius laevigatus and O. albidipennis (Hemiptera: Anthocoridae)
to optimize their use as biological control agents of Frankliniella
occidentalis (Thysanoptera: Thripidae). Bull Entomol Res
92:77–88
Schmidt JM, Richards PC, Nadel H, Ferguson G (1995) A rearing
method for the production of large numbers of the insidious
flower bug, Orius insidiosus (Say) (Hemiptera: Anthocoridae).
Can Entomol 127:445–447
Schuh RT, Stys P (1991) Phylogenetic analysis of cimicomorphan
family relationships (Heteroptera). J N Y Entomol Soc
99:298–350
Shapiro JP, Shirk PD (2010) Ovarian development in predacious
Orius pumilio: relationship to diet, mating, and juvenile
hormone. Ann Entomol Soc Am 103:971–978
Simmons LW, Kotiaho JS (2007) The effects of reproduction on
courtship, fertility and longevity within and between alternative
male mating tactics of the horned beetle, Onthophagus binodis.
J Evol Biol 20:488–495
Sobhy I, Sarhan A, Shoukry A, El-Kady G, Mandour N (2006) Effects
of different temperature levels on the biological attributes of
Orius albidipennis (Reuter) (Hemiptera: Anthocoridae). J Agric
Res Suez Canal Univ 6:213–216
Sobhy I, Sarhan A, Shoukry A, El-Kady G, Mandour N, Reitz S
(2010) Development, consumption rates and reproductive biol-
ogy of Orius albidipennis reared on various prey. Biocontrol
55:753–765
Stutt AD, Siva-Jothy MT (2001) Traumatic insemination and sexual
conflict in the bed bug Cimex lectularius. Proc Natl Acad Sci
98:5683–5687
Symondson WOC, Sunderland KD, Greenstone MH (2002) Can
generalist predators be effective biocontrol agents? Annu Rev
Entomol 47:561–594
Tawfik MFS, El-Husseini MM (1971) The life history of the
Blaptostethus pallescens Fieber, Poppius (Hemiptera: Anthoco-
ridae). Bull Soc Entomol Egypt 55:239–352
van Lenteren JC (2012) The state of commercial augmentative
biological control: plenty of natural enemies, but a frustrating
lack of uptake. Biocontrol 57:1–20
van Lenteren JC, Tommasini MG (2003) Mass production, storage,
shipment and release of natural enemies. In: van Lenteren JC
(ed) Quality control and production of biological control agents:
theory and testing procedures. CABI Publishing, Wallingford,
pp 181–189
van Lenteren JC, Bale J, Bigler F, Hokkanen HMT, Loomans AJM
(2006) Assessing risks of releasing exotic biological control
agents of arthropod pests. Annu Rev Entomol 51:609–634
Wagner TL, Wu H-I, Sharpe PJH, Schoolfield RM, Coulson RN
(1984) Modeling insect development rates: a literature review
and application of a biophysical model. Ann Entomol Soc Am
77:208–225
Yamane T, Higuchi H, Takahashi A, Ishimoto M (2011) Reduction of
mating receptivity and changes in longevity of rice leaf bug,Trigonotylus caelestialium (Heteroptera: Miridae), females after
mating. Appl Entomol Zool 46:559–564
Yanik E, Unlu L (2011) Influences of temperature and humidity on
the life history parameters and prey consumption of Anthocoris
minki Dohrn (Heteroptera: Anthocoridae). Appl Entomol Zool
46:177–184
324 Appl Entomol Zool (2014) 49:315–324
123





























