Embed Size (px)
Citation preview

LIM Kinase 1 Accumulates in PresynapticTerminals During Synapse Maturation
JAY Y. WANG,1 DONALD J. WIGSTON,2 HOWARD D. REES,3 ALLAN I. LEVEY,3
AND DOUGLAS L. FALLS1*1Department of Biology, Emory University, Atlanta, Georgia 30322
2Department of Physiology, Emory University, Atlanta, Georgia 303223Department of Neurology, Emory University, Atlanta, Georgia 30322
ABSTRACTLIM kinase 1 (LIMK1) is a cytoplasmic protein kinase that is highly expressed in
neurons. In transfected cells, LIMK1 binds to the cytoplasmic tail of neuregulins andregulates the breakdown of actin filaments. To identify potential functions of LIMK1 in vivo,we have determined the subcellular distribution of LIMK1 protein within neurons of the ratby using immunomicroscopy. At neuromuscular synapses in the adult hindlimb, LIMK1 wasconcentrated in the presynaptic terminal. However, little LIMK1 immunoreactivity wasdetected at neuromuscular synapses before the 2nd week after birth, and most motoneuronterminals were not strongly LIMK1 immunoreactive until the 3rd week after birth. Thus,LIMK1 accumulation at neuromuscular synapses coincided with their maturation. Incontrast, SV2, like many other presynaptic terminal proteins, can be readily detected atneuromuscular synapses in the embryo. Similar to its late accumulation at developingsynapses, LIMK1 accumulation at regenerating neuromuscular synapses occurred long afterthese synapses first formed. In the adult ventral spinal cord, LIMK1 was concentrated in asubset of presynaptic terminals. LIMK1 gradually accumulated at spinal cord synapsespostnatally, reaching adult levels only after P14. This study is the first to implicate LIMK1 inthe function of presynaptic terminals. The concentration of LIMK1 in adult, but not nascent,presynaptic terminals suggests a role for this kinase in regulating the structural or functionalcharacteristics of mature synapses. J. Comp. Neurol. 416:319–334, 2000. r 2000 Wiley-Liss, Inc.
Indexing terms: synaptogenesis; nonreceptor protein kinase; neuromuscular junction; spinal
motoneurons; synaptic plasticity
Protein kinases play a major role in regulating thedevelopment and function of the nervous system (forreview, see Levitan, 1994; Valtorta et al., 1996; Tokuda andHatase, 1998). Several cytoplasmic kinases, such as CAM-kinase and src, and receptor kinases, have been found atsynapses. Some of these synapse-associated kinases arelocalized to the presynaptic terminal, others to the postsyn-aptic membrane or subsynaptic cytoplasm, and some arefound both presynaptically and postsynaptically. Theirfunctions include regulation of neurotransmitter receptorsynthesis and anchoring, neurotransmitter receptor func-tion, neurotransmitter release, and synaptic morphology.An important step in elucidating the functions of suchsynapse-associated kinases is defining their location withinsynapses and the temporal pattern of their accumulationat synapses during development.
LIM Kinase 1 (LIMK1) is a cytoplasmic protein kinaseidentified in several screens for novel kinases expressed inthe nervous system (Bernard et al., 1994; Mizuno et al.,1994; Cheng and Robertson, 1995; Proschel et al., 1995).
The name LIM Kinase 1 derives from the fact that LIMK1is the first protein found to contain both LIM domains,which are zinc-finger motifs that mediate protein-proteininteractions (Dawid et al., 1998) and a kinase domain. Onthe basis of genetic evidence, LIMK1 hemizygosity wasproposed as the cause of a cognitive defect seen in patientswith Williams syndrome, but this association was recentlyquestioned (Tassabehji et al., 1999). However, studies ofcells transfected with LIMK1 have revealed several pos-sible roles for this kinase. Evidence from our laboratorydemonstrates that LIMK1 can interact with the cytoplas-mic tail of neuregulins (Wang et al., 1998), suggesting thatLIMK1 may regulate signaling by neuregulins (NRGs).
Grant sponsor: National Institutes of Health; Grant number: GM 56337.*Correspondence to: Douglas L. Falls, Department of Biology, Rollins
Research Building, 1510 Clifton Road, Emory University, Atlanta, GA30322. E-mail: [email protected]
Received 18 May 1999; Revised 22 September 1999; Accepted 23 Septem-ber 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 416:319–334 (2000)
r 2000 WILEY-LISS, INC.

LIMK1 can also phosphorylate and inhibit the actin-binding protein cofilin (Arber et al., 1998; Yang et al.,1998). Cofilin severs actin filaments; thus, inhibition ofcofilin by LIMK1 could increase filamentous actin and/orslow remodeling of the actin cytoskeleton.
In a study of the interaction between LIMK1 and theNRG cytoplasmic tail, we discovered that LIMK1, likeNRG, is present at adult neuromuscular synapses (Wanget al., 1998). Previously, LIMK1 immunoreactivity hadbeen noted in the cytoplasm and nuclei of some neurons ofthe adult mouse brain (Bernard et al., 1994), but synapticlocalization of LIMK1 has not been reported.
Our finding that LIMK1 is present at adult neuromuscu-lar synapses raised a number of new questions, includingwhether LIMK1 is located in the presynaptic or postsynap-tic component of synapses, whether LIMK1 is also found atcentral synapses, and when during development it accumu-lates at synapses. Here we have used immunohistochemi-cal methods to answer these questions. We report thatLIMK1 is concentrated in the presynaptic terminals ofadult neuromuscular synapses and the presynaptic termi-nals of some synapses in the adult spinal cord. In contrastto the synaptic vesicle protein SV2, which reaches its adultlevels in terminals before birth, LIMK1 immunoreactivityat embryonic and early postnatal synapses is near thebackground level. Similarly, LIMK1 accumulation at regen-erating synapses lags behind SV2. These findings suggestthat LIMK1 regulates the structure and/or function ofadult presynaptic terminals.
MATERIALS AND METHODS
Reagents
SV2 and 2H3 hybridoma supernatants were obtainedfrom the Developmental Studies Hybridoma Bank main-tained by the University of Iowa (Iowa City, IA); rabbitanti-S100 antibody from Dako (Carpinteria, CA); fluores-cein- and rhodamine-conjugated secondary antibodies fromJackson ImmunoResearch Laboratories, Inc. (West Grove,PA); fluorescein and Texas red a-bungarotoxin (a-BTX)from Molecular Probes, Inc. (Eugene, OR); horseradishperoxidase (HRP)-conjugated goat anti-rabbit antibody fromPierce (Rockford, IL); and biotinylated goat anti-rabbitand the ABC Elite kit from Vector (Burlingame, CA).
Constructs and calculation ofLIMK1-predicted molecular weight
A LIMK1 expression construct was created by append-ing the myc-epitope tag onto the N-terminal end of themouse LIMK1 complementary deoxyribonucleic acid(cDNA) sequence as described in Wang et al. (1998). Themyc-LIMK1 cDNA encodes a protein of 661 amino acidswith a predicted molecular weight of 73 kDa.
Conceptual translation of the rat LIMK1 cDNA se-quence that was first isolated predicts that the rat LIMK1protein is 647 amino acids in length (Nunoue et al., 1995).The predicted mass of this 647 amino acid polypeptide is71 kDa, and in discussing our results, we explicitly con-sider only this mass. Somewhat smaller LIMK1 peptidesmight also be produced as a consequence of alternativesplicing or translation initiation downstream of the 58-most AUG (Bernard et al., 1994; Cheng and Robertson,1995; Proschel et al., 1995). Recently, a cDNA encoding aLIMK1 form entirely lacking the kinase domain wasisolated (Edwards and Gill, 1999). All of these possible
variants, with the exception of the kinase-deleted form,contain the peptide sequence used to generate our LIMK1antibody.
Production of the LIMK1 antibody
The LIMK1 antiserum LK1-EU1 was produced by immu-nizing a rabbit with the synthetic peptide a-acetyl-KETYRRGESSLPAHPEVPD conjugated to horseshoe crabhemocyanin (H1757; Sigma, St. Louis, MO) using glutaral-dehyde. The underlined amino acids correspond to the 18C-terminal amino acids of the rat LIMK1 protein (aminoacids 630–647 of Genbank accession D31873; Nunoue etal., 1995). Immunization and harvesting of serum wereperformed by Covance Research Products (Denver, PA).Serum was affinity purified by using the immunizingpeptide conjugated to Affigel 15 (BioRad, Hercules, CA)according to the manufacturer’s instructions. The affinity-purified antibody preparation had an immunoglobulinconcentration of 1.5 mg/ml, as estimated from Coomassieblue-stained sodium dodecyl sulfate (SDS) gels. This affin-ity-purified preparation was used for all of the experi-ments reported in this study. A preliminary characteriza-tion of the LK1-EU1 antibody has been reported (Wang etal., 1998).
Animals and surgery
Wistar Sprague-Dawley rats (Harlan, Indianapolis, IN)were used for all experiments. Ages of animals rangedfrom embryonic day 15 (E15 5 15 days postcoitus) toapproximately P90 (postnatal day 90). Rats are born atE21–E22, become weanlings at approximately P21, andbecome adults (sexually mature) around P50–60. Forsurgery, 2- or 3-month-old male rats, weighing approxi-mately 400 g, were anesthetized with ketamine (8 mg/kg)and xylazine (12 mg/kg). For nerve transection, the sciaticnerve was exposed in the upper thigh, and a 0.5-cmsegment of the nerve was removed. The gastrocnemiusmuscle was harvested 5 days later. For regenerationstudies, the sciatic nerve was crushed at midthigh levelwith no. 5 forceps. At various times after the crush, thesoleus muscle was harvested. For transection of ventraland dorsal roots, the left L4 roots were exposed by using adorsal approach. The ventral and dorsal roots were cut,and the left L4 dorsal root ganglion was removed. Thespinal cord was harvested 5 or 10 days later, and sectionswere prepared from the L4 level of the cord. The unoper-ated leg (muscle denervation experiments) or unoperatedside of the spinal cord (transection of roots) from the samerats was used as controls. All surgical procedures andanimal handling were performed in accordance with EmoryUniversity’s guidelines for animal care.
Tissue immunoblots
Adult rats were killed by CO2 inhalation. Tissues ofinterest were immediately dissected, rinsed with 13 phos-phate-buffered saline (PBS: 0.1 M phosphate buffer, pH7.4, 150 mM NaCl), and frozen on dry ice. The tissues werethen immersed in HNTE buffer (50 mM HEPES, pH 7.4,150 mM NaCl, 1% vol/vol Triton X-100, 5 mM ethylenedi-aminetetraacetic acid [EDTA], 10% vol/vol glycerol and‘‘Complete’’ protease inhibitor cocktail [1 tablet/10 mlbuffer; Boehringer Mannheim]). The tissues were homog-enized by using a polytron (model PT3000; Brinkmann).The homogenates were subjected to centrifugation at15,000 g for 20 minutes at 4°C to remove insoluble
320 J.Y. WANG ET AL.

material, and the supernatants (hereafter referred to astissue lysates) were transferred to fresh tubes. The proteinconcentration of the tissue lysates was analyzed by usingthe BioRad DC protein assay kit. For Western blotting, analiquot of each tissue lysate was mixed with an equal volumeof 23 SDS-1,4-dithiothreitol (DTT) sample buffer (23 5 6%wt/vol SDS, 20% vol/vol glycerol, 0.01% wt/vol bromophenolblue, 125 mM Tris-HCl, pH 6.8, 100 mM DTT), the sample washeated to 95°C for 5 minutes, and 40 µg of total protein fromeach tissue lysate was loaded on a 10% SDS-polyacrylamidegel. Immunoblotting was performed as previously described(Wang et al., 1998). The LK1-EU1 antibody was used at a finalconcentration of 0.75 µg/ml. For the peptide competitioncontrol, 3.75 µg of LK1-EU1 was incubated with 15 µg ofimmunizing peptide in a volume of 20 µl for 2 hours atroom temperature, and this mixture was used in place ofthe LK1-EU1 antibody in the immunoblot procedure (finalantibody concentration 5 0.75 µg/ml).
Immunoprecipitation
Tissue lysates were prepared in a nondenaturing lysisbuffer as described above. Two hundred micrograms ofbrain lysate protein or 1 mg of spinal cord, kidney, ormuscle lysate protein in a volume of 1 ml of HNTE bufferwas used for each immunoprecipitation. Lysates wereprecleared by incubation with 10 µg of rabbit IgG (Sigma)and 20 µl of protein A-agarose (Sigma) for 1 hour at 4°C.The protein-A beads were then spun down at 700 g, andthe supernatants were transferred to fresh tubes contain-ing 5 µg of LK1-EU1. After incubation for 1 hour at 4°C, 20µl of protein-A agarose was added to the tissue lysate/antibody mixture and incubated on a rotator for 4 hours at4°C. The beads were washed with HNTE buffer at roomtemperature (4 washes, 3 minutes each), and bound pro-tein was eluted by incubation in 50 µl of 23 SDS-DTTsample buffer for 30 minutes at room temperature. Theprotein-A agarose beads were sedimented by centrifuga-tion, and the eluate was transferred to a clean tube. Beforeloading on the gel, the eluate was heated for 5 minutes at95°C. Fifteen microliters of the eluate (representing theprecipitate from approximately 30% of the starting amountof tissue lysate) was analyzed by immunoblot as previously
described (Wang et al., 1998). For the peptide competitioncontrol, 5 µg of LK1-EU1 was incubated with 15 µg of theimmunizing peptide in a volume of 20 µl for 2 hours atroom temperature. This mixture was then added to theprecleared tissue lysates, and immunoprecipitation wasperformed as described above (final antibody concentra-tion 5 5 µg/ml).
Fluorescence immunohistochemistry
Rats were anesthetized with 4% wt/vol chloral hydrate(P21 and adult) or ether (P0–P14) and perfused throughthe heart with 4% wt/vol paraformaldehyde in 0.1 Msodium phosphate, pH 7.4. Hindlimb muscles and spinal
Fig. 1. The LK1-EU1 antibody specifically identifies LIMK1 inWestern blot and immunoprecipitation assays. A: Western blot analy-sis of lysates prepared from adult rat brain or from COS-7 cellstransfected with myc-LIMK1. The antibody labeled a single band ineach lysate (left two lanes). No bands were labeled when the antibodywas preincubated with the immunizing peptide (right two lanes). Thelabeled protein in the COS-7 cell lysate has a relative molecularweight of 73 kDa, the size expected for the 661 amino acid myc-LIMK1protein. The protein labeled in the brain lysate has a relativemolecular weight of 83 kDa. B: Western blot analysis of adult rattissues. For each tissue lysate, 40 µg of total protein was loaded. Asingle band at 83 kDa was recognized in brain, but not in the othertissues tested. C: Immunoprecipitation of LIMK1 from tissue lysates.Both the immunoprecipitation and the Western blot analysis of theprecipitates were performed with the LK1-EU1 antibody. An 83-kDaband was detected in brain and spinal cord but not in skeletal muscleor kidney. When the antibody was preincubated with the immunizingpeptide before immunoprecipitation, no LIMK1 immunoreactivity wasfound in the precipitate (left lane). One milligram of spinal cord,kidney, and muscle lysate protein was used for the immunoprecipita-tions, but only 200 µg of brain protein was used. Thus, the relativeintensity of the observed bands indicates that the level of LIMK1 inspinal cord is more than in kidney or muscle, but somewhat less thanin brain.
LIMK1 IN PRESYNAPTIC TERMINALS 321

cord were dissected and immersed in 15% wt/vol sucrose in0.1 M sodium phosphate, pH 7.4, until they sank, trans-ferred to 30% sucrose in 0.1 M sodium phosphate, pH 7.4,for 2–3 days, and then frozen in OCT compound by usingliquid nitrogen. To harvest embryonic tissue, a pregnantfemale rat was killed by CO2 inhalation, and embryos wereremoved to a 100-mm Petri dish containing PBS. Thehindlimbs were dissected and immersed in 4% paraformal-dehyde in 0.1 M of sodium phosphate, pH 7.4, overnight at4°C. The tissues were cryoprotected with sucrose andfrozen in OCT compound as described above. Ten-microme-ter thick sections were cut in a cryostat and processed forimmunohistochemistry as previously described (Wang etal., 1998).
Antibody concentration or dilutions used for immunohis-tochemistry were as follows: LK1-EU1, 1.5 µg/ml; anti-SV2, 1:100; 2H3, 1:1,000; S100, 1:200; lissamine rhoda-mine sulfonyl chloride (LRSC)-conjugated donkey anti-rabbit, 1:200; fluorescein isothiocyanate conjugate (FITC)-conjugated donkey anti-mouse 1:200; and LRSC-conjugateddonkey anti-mouse, 1:200. Fluorescein a-BTX (see Fig. 2)was used at 5 µg/ml. Texas-red a-BTX (see Fig. 3) was usedat 1 µg/ml. When used, a-BTX was applied with thesecondary antibody. Note that in Figure 3, for the SV2/BTXdouble-label panels, Texas red a-BTX is displayed in greeninstead of the usual red, and FITC-conjugated donkeyanti-mouse is displayed in red. The specificity of LK1-EU1label was tested by preincubating the antibody with theimmunizing peptide for 2 hours at room temperature (10µg peptide per 1 µg antibody in a volume of 20 µl;antibody/peptide mixture diluted with PBS). This mixturewas then applied to tissue sections as described above(final antibody concentration 5 1.5 µg/ml).
Sections were scanned by using a BioRad 1024 laserscanning confocal microscope (BioRad) coupled to a ZeissAxioskop.A203 Plan-Neofluar (NA0.5), a 403 Plan-Neofluar(NA0.75), and a 633 Plan-Apochromat oil immersion lens (NA1.4) were used. A Z-series of images was collected for eachpreparation at a step of 0.5 µm. The images shown in thefigures were compiled from a stack of 10–20 individual Z-series images by using the BioRad Lasersharp software,except that in Figure 7A2 and B each panel shows a singleZ-series image. Images were further processed by usingAdobePhotoshop. All images within a single figure were collectedidentically, and all images within a single figure showing the
same fluorophore were processed identically. For each figure,images in the same row show the same section, and superim-position of red and green fluorescence is seen as yellow.Control experiments were performed in which sections weredeveloped with only one primary antibody but with bothsecondary antibodies used for double labeling. Each primaryantibody used was tested in this type of control experiment.These experiments demonstrated the absence of bleedthroughand cross-labeling. They also showed that, in the absence ofappropriate primary antibody, no signal was produced by anyof the secondary antibodies used.
Subjective rating of LIMK1 and SV2 accumulation atneuromuscular synapses during postnatal developmentwas performed as follows: sections were double labeledwith fluorescein-a-BTX and either LK1-EU1 or anti-SV2antibody visualized by a rhodamine-conjugated secondary.At postnatal time points, essentially all AChR clusters inthe normal rat hindlimb are at synapses (see Results).Therefore, for purposes of this assessment, a synapse wasdefined as a cluster of acetylcholine receptors (AChRs)identified by a-BTX label. Each field was first examinedwith fluorescein optics to identify AChR clusters. The fieldwas then examined with rhodamine optics, and the inten-sity of the LIMK1 (or SV2) label that was colocalized withthe clustered AChRs was scored. At each developmentalage studied, at least 300 synapses were scored for colocal-ized LIMK1 label intensity and at least 95 synapses werescored for colocalized SV2 label intensity. The score as-signed was based on a subjective assessment of the aver-age brightness of the LIMK1 (or SV2) label colocalizedwith the BTX label. Intensities were scored as ‘‘Strong,’’‘‘Moderate,’’ ‘‘Weak,’’ or ‘‘Not detected.’’ The observer scor-ing these sections was not informed of the age of theanimal from which each section was harvested.
Immunoelectronmicroscopy
Immunoelectronmicroscopy was performed as describedelsewhere (Gilmor et al., 1996). The LK1-EU1 was used at1.5 µg/ml. Biotinylated goat anti-rabbit secondary wasused at 1:200. The avidin-biotin-complex method was usedto visualize immunoreactivity. The peroxidase reactionproduct was developed in 0.05% diaminobenzidine (DAB)and 0.01% H2O2. The immunostained tissue was postfixedwith 1% osmium tetroxide and embedded in Eponate 12(Ted Pella, Redding, CA). The embedded tissue was thin
Fig. 2. LIMK1 is concentrated in presynaptic terminals at theneuromuscular synapse. Adult rat hindlimb muscle was cut longitudi-nally, and sections were processed for immunohistochemistry andanalyzed by confocal microscopy. A: The distribution of LIMK1 isnearly identical to the distribution of the presynaptic terminal markerSV2 at the adult neuromuscular synapse. B: In both en face andcross-sectional (inset in B3) views, the distribution of LIMK1 isdistinct from the distribution of the postsynaptic membrane AChRs.The cross-sectional profile, which represents the LIMK1 and AChRdistribution along the dotted line of Figure B3, was digitally recon-structed from the confocal Z-stack data. In B4, the fluorescent image ofB3 is superimposed on a low-power differential interference contrast(DIC) view of the muscle tissue in which this synapse was observed.Four muscle fibers are seen. The long axis of these fibers runs left toright in this image. Striations of the fibers are seen oriented perpen-dicular to the long axis. The synapse shown was the only synapseobserved in this field. As illustrated in this panel, most of the synapsesobserved in the longitudinal muscle sections used in this study areviewed en face. Insets in panels B1 and B2 demonstrate that preincu-bation of the LK1-EU1 antibody with the immunizing peptide abol-ishes synaptic labeling. C: The distributions of SV2 and the Schwanncell marker S100 are distinct. Because the available LIMK1 and S100
antibodies were both raised in rabbit, we inferred the distribution ofLIMK1 relative to S100 from the SV2/S100 double-label images. In thesuperimposed SV2/S100 image (C3), a large proportion of the coloredpixels are red or green, demonstrating that the confocal images resolvethe different spatial localization of SV2 and S100. Because thedistribution of LIMK1 appears nearly identical to that of SV2 (A1–3),the SV2/S100 double-label result indicates that LIMK1 is not concen-trated in the Schwann cells capping the neuromuscular synapse.Arrow points to the soma of a Schwann cell. D: Five days aftertransection of the sciatic nerve, LIMK1 label is dramatically reducedat synaptic sites in denervated muscle, but the distribution of AChRsappears no different than that at innervated adult synapses. The rapiddisappearance of LIMK1 label from the synaptic site provides furtherevidence that LIMK1 is a presynaptic protein. E: LIMK1 is lessabundant in motoneuron axons than at neuromuscular synapses ofthe adult rat. Neurofilament-M (NF-M) is known to be excluded fromthe nerve terminal, but it is found throughout the rest of the axon. TheLIMK1 label seen along the axon is weak compared with the LIMK1label in the nerve terminal. We do not yet know if the faint label alongthe axon is within the axon or in Schwann cells or other cells borderingthe axon. Scale bars 5 20 µm.
322 J.Y. WANG ET AL.

sectioned at 50 nm on an ultramicrotome and examined byusing a Hitachi H7500 electron microscope.
RESULTS
Characterization of the LIMK1 antibody
A LIMK1 antiserum, LK1-EU1, was raised by immuniz-ing a rabbit with a peptide corresponding to the carboxy-
terminal amino acid sequence of rat and mouse LIMK1.The LK1-EU1 antibody preparation used in this study wasaffinity purified from this serum.ABLAST search (Altschulet al., 1997) revealed no proteins other than LIMK1 withclose sequence similarity to the peptide used for immuniza-tion. In particular, there is no sequence similarity betweenthe immunizing peptide and the carboxy-terminal regionof the LIMK1 homologue LIMK2 (Nunoue et al., 1995).
Figure 2
LIMK1 IN PRESYNAPTIC TERMINALS 323

The specificity of the LK1-EU1 antibody was tested inWestern blot and immunoprecipitation assays. In Westernblots, the antibody labeled a single band in lysates ofCOS-7 cells transfected with myc-LIMK1 (Fig. 1A) butlabeled no bands in lysates of untransfected COS-7 cells(Wang et al., 1998). The antibody also labeled a single bandin lysates of brain, but no proteins were labeled in lysatesof skeletal muscle, heart, kidney, or liver (Fig. 1B). Proteinimmunoprecipitated by the LK1-EU1 antibody from tissuelysates was also analyzed by Western blot. In immunopre-cipitates of brain and spinal cord, the antibody labeled asingle band, but no bands were labeled in immunoprecipi-tates of skeletal muscle or kidney (Fig. 1C). These resultsare consistent with previous reports that LIMK1 mRNA ismore abundant in nervous system tissue than in othertissues (Bernard et al., 1994; Mizuno et al., 1994; Chengand Robertson, 1995; Okano et al., 1995; Proschel et al.,1995). Preincubation of the antibody with the peptide usedfor immunization abolished labeling on immunoblots (Fig.1A), blocked immunoprecipitation (Fig. 1C), and abolishedor markedly reduced immunohistochemical labeling (insetin Fig. 2B2). Taken together, these data strongly supportthe hypothesis that LK1-EU1 specifically recognizes theLIMK1 protein.
It may seem surprising that LIMK1 was not detected inmuscle tissue, as we have observed LIMK1 immunoreactiv-ity at neuromuscular synapses (Wang et al., 1998; see alsoFig. 2). However, because proteins in axons and at syn-apses constitute only a tiny percentage of the total proteinin muscle, proteins present in relatively high concentra-tions at synapses may have a low concentration in muscletissue lysates. Thus, proteins concentrated at the neuro-muscular synapse might be readily observed in immunohis-tochemical assays but still not detected in biochemicalassays of muscle lysates.
The apparent molecular weight of myc-LIMK1 in COScell lysates is 73 kDa, the size predicted from the knownrat LIMK1 cDNA sequence (see Materials and Methods fordiscussion of predicted LIMK1 size). However, the appar-ent molecular weight of LIMK1 in rat brain and spinalcord lysates is 83 kDa (Fig. 1). No Western analysis ofLIMK1 in brain or other tissues has previously beenpublished. However, the identification of the 83-kDa bandas LIMK1 has been confirmed by using an antibody raisedagainst the N-terminal region of LIMK1 (Falls et al.,unpublished data). Possible reasons for the difference inmobility of COS cell and rat brain LIMK1 include posttrans-lational modifications, such as phosphorylation, or analternatively spliced LIMK1 messenger ribonucleic acid(mRNA) encoding a larger protein.
LIMK1 is concentrated in the presynapticterminal at mature neuromuscular synapses
We previously showed that LIMK1 is present at adultneuromuscular synapses (Wang et al., 1998). This findingraised the question of whether LIMK1 is contained withinmotoneuron terminals, postsynaptic muscle fibers, or thesynapse-associated Schwann cells (see Hall and Sanes,1993 and references therein for discussion and illustrationof neuromuscular synapse structure and development). Toanswer this question we used confocal immunomicroscopyto compare the distribution of LIMK1 at adult neuromus-cular synapses to the presynaptic terminal protein SV2(Buckley and Kelly, 1985; Lupa and Hall, 1989), thepostsynaptic membrane AChRs (Dennis, 1981; Anderson
et al., 1984), and the Schwann cell protein S100 (Woolf etal., 1992; Son and Thompson, 1995a; Son and Thompson,1995b; Hirata et al., 1997).
The distribution of LIMK1 and SV2 at neuromuscularsynapses appeared nearly identical (Fig. 2A). In contrast,the distribution of LIMK1 was distinct from that of AChRs(Fig. 2B) and S100 (compare Fig. 2A with Fig. 2C). Thesedata indicate that most LIMK1 at neuromuscular syn-apses is in the presynaptic terminals, not in the postsynap-tic region of the muscle or in Schwann cells. Moreover, theprecise alignment of the LIMK1 and SV2 immunoreactiv-ity suggests that LIMK1 is especially abundant nearactive zones, because synaptic vesicles within the nerveterminal are most abundant near the active zones (forreview, see Hall and Sanes, 1993; Burns and Augustine,1995).
Denervation of hindlimb muscle was performed to fur-ther test the hypothesis that synaptic LIMK1 is princi-pally located in the presynaptic terminal. Labeling forLIMK1 and SV2 was dramatically reduced at synapticsites within 5 days after denervation, but the AChRpattern at this time appeared similar to that of innervatedsynapses (Fig. 2D; SV2 result not shown). The nerveterminal is known to degenerate soon after denervation.Thus, the rapid disappearance of LIMK1 from the synapticsite is consistent with presynaptic localization of LIMK1,although it must be noted that this result alone does notexclude the possibility of postsynaptic localization (Astrowet al., 1992).
To determine if LIMK1 is similarly abundant in axonsand nerve terminals, sections of muscle were doublelabeled for LIMK1 and neurofilament-M (NF-M), a cyto-skeletal component found along the length of motor axons,but not in their terminals (Lupa and Hall, 1989; Lee andCleveland, 1996). LIMK1 label appeared much more in-tense in the presynaptic terminals than along the axons(Fig. 2E), suggesting that LIMK1 is specifically traffickedto the presynaptic terminal of motoneurons.
LIMK1 accumulates at neuromuscularsynapses during the postnatal period
of synaptic maturation
To assess the potential for LIMK1 involvement in neuro-muscular synapse formation and maturation, we exam-ined the accumulation of LIMK1 at developing neuromus-cular synapses and along intramuscular axons. In ourstudies, we compared the embryonic and postnatal distri-bution of LIMK1 to both AChRs (Figs. 3 and 4) and SV2(Fig. 5).
Synapse formation, defined as colocalization of synapticvesicle proteins and acetylcholine receptors, begins in therat hindlimb around E18 (Rubinstein and Kelly, 1981;Condon et al., 1990). At E18, occasional small areas ofoverlap between SV2 and AChR label were observed, andmost AChR clusters were near SV2-labeled axons (Fig.3A). At E20, strong SV2-label colocalized with most AChRclusters (Fig. 3C), indicating that by E20 most AChRclusters are located at nascent synapses. Postnatally,strong SV2 label was colocalized with all clusters of AChRsexamined (Fig. 4).
The relationship of AChR clusters to LIMK1 immunore-activity was quite different. At E18, LIMK1 was evidentalong portions of the intramuscular nerves, but few AChRclusters were found near LIMK1-labeled regions of thesenerves (Fig. 3B; see also Fig. 5 E18). At E20, only faint
324 J.Y. WANG ET AL.

Fig. 3. Little LIMK1 is observed at embryonic neuromuscularsynapses. Longitudinal sections from rat hindlimb muscle were doublelabeled with a-BTX and the SV2 antibody (A and C) or LK1-EU1 (Band D). In each set of panels, the positions of two AChR clusters areindicated with arrows or arrowheads. A: At E18, the anti-SV2antibody labels intramuscular axon bundles and small spots borderingthese bundles. On the basis of their proximity to the axon bundles,these spots likely correspond to axon terminals or the shafts of smallaxonal branches. Most AChR clusters are associated with SV2-labeledaxons, but at this early stage of synapse development many AChRclusters do not overlap, or only partially overlap, SV2-labeled nerves.
B: At E18, LIMK1 label is present along some segments of theintramuscular nerves (compare Fig. 5), but AChR clusters are gener-ally not associated with these LIMK1-labeled nerve segments. C: AtE20, SV2 label is still found along the shafts of axons. Most brightAChR clusters are colocalized with SV2 label. Many of these AChRclusters are overlain by a bright, spade-shaped patch of SV2 label thatlikely corresponds to a nerve terminal. D: At E20, only faint patches ofLIMK1 immunoreactivity are observed in muscle. In contrast to SV2,the LIMK1 immunoreactivity rarely overlaps AChR label at this stageof development. Scale bars 5 20 µm in E18; 30 µm in E20.

patches of LIMK1 immunoreactivity were observed, andthere continued to be little overlap of the LIMK1 label withAChR label (Fig. 3D and Fig. 4). Postnatally, the intensityof LIMK1 label that overlapped AChR clusters increasedonly gradually. At P0 and P2 ,5% of the synapses exam-ined were even weakly LIMK1 immunoreactive. At P6approximately 50% were weakly labeled, and a few per-cent were moderately labeled. The amount of labeling wasgreater at P14, but it was not until P21 that <50% of thesynapses were strongly LIMK1 immunoreactive. Only inadults were most of the synapses examined stronglyLIMK1 immunoreactive.
In the experiments just described (Figs. 3 and 4), thedistribution of LIMK1 relative to AChRs was assessed. Tofurther define the accumulation of LIMK1 in the develop-ing neuromuscular system, we compared the distributionof LIMK1 to SV2. For these experiments, we first evalu-ated the changes that occur in SV2 distribution duringhindlimb neuromuscular development. Before the forma-tion of neuromuscular synapses, and also during the earlystages of synaptogenesis, SV2 was observed along thelength of intramuscular axons (Fig. 3A and C; Fig. 5 E15and E18). Subsequently, SV2 immunoreactivity becameprogressively more restricted, until by P21, SV2 was foundonly within the presynaptic terminal and the immediatelyadjacent preterminal axon (not shown). Similar observa-tions for the progressive restriction of SV2 immunoreactiv-ity to the synapse have previously been reported in a studyof neuromuscular synapse formation in the mouse dia-phragm (Lupa and Hall, 1989).
Comparison of the distribution of LIMK1 with SV2 atdifferent developmental stages revealed expression ofLIMK1 both along axons and at synapses. These two
aspects of LIMK1 neuromuscular expression had differenttime courses. The expression along axons was most promi-nent at E15, the first age we examined (Fig. 5 E15). AfterE15, LIMK1 immunoreactivity along intramuscular axonsrapidly declined (Fig. 5 E18) and remained low at all laterstages examined (see Fig. 2E). It is not yet clear if theLIMK1 that is distributed along embryonic axons is withinthe axoplasm, or if instead it is expressed in Schwann cellsor other cells of the peripheral nerve.
The expression of LIMK1 in presynaptic terminals wasfirst observed much later in development. At E18, whensynapses are initially forming, presynaptic terminals can-not unequivocally be identified by the pattern of SV2 label(see Fig. 3A). However, little or no LIMK1 immunoreactiv-ity was observed in the distal portions of the SV2-labeledaxonal arbors, suggesting that levels of LIMK1 are low innerve terminals at this stage (Fig. 5 E18). By E20, and atsubsequent stages, nerve terminals can be clearly identi-fied by the pattern of SV2 label (see Fig. 3C). At E20 andalso at P0, LIMK1 immunoreactivity was not readilydetected at nerve terminals (Fig. 5 E20 and P0). However,by P6 small spots of faint LIMK1 label were observed atsome nerve terminals (Fig. 5 P6). LIMK1 label at nerveterminals gradually increased over the first postnatalweeks, but adult levels were not reached until after P14(Fig. 5 P14, P21, and adult).
These data (Figs. 3–5) demonstrate that the principalaccumulation of LIMK1 at neuromuscular synapses occursover the first several weeks after birth and that the levelsof LIMK1 are low when synapses are first formed, relativeto the levels in the adult.
LIMK1 at regenerating neuromuscularsynapses
To determine if the delayed appearance of LIMK1 atdeveloping neuromuscular synapses was related to the
Fig. 4. SV2 appears at developing neuromuscular synapses beforeLIMK1. Scoring of LIMK1 (main graph) and of SV2 immunoreactivity(inset) at neuromuscular synapses was performed as described inMaterials and Methods. The bars for the ‘‘not detected’’ category arenot shown on the histograms. Percentages in the ‘‘not detected’’category were as follows: E20, 98%; P0, 98%; P2, 97%; P6, 42%; P10,17%; P14, 7%; P21, 0%; Adult, 0%. For each age analyzed, the sum of‘‘not detected,’’ weak, moderate, and strong labeling is 100%. Note thatalthough 100% of the synapses scored showed strong SV2 label at allages examined (inset), the percentage of synapses strongly labeled bythe LIMK1 antibody slowly increased over the first few weeks afterbirth (main graph).
Fig. 5. Comparison of LIMK1 and SV2 distribution during neuro-muscular development. Longitudinal sections from rat hindlimb musclewere double labeled with the LK1-EU1 and SV2 antibodies. E15: AtE15, a time before neuromuscular synapse formation in the hindlimb,SV2 and LIMK1 immunoreactivity are seen along intramuscular axonbundles. Although the distributions of SV2 and LIMK1 overlap, thereis not the precise correspondence that is observed at the adultneuromuscular synapse. E18: The distribution of LIMK1 and SV2immunoreactivity along the intramuscular axon shafts is similar toE15, but the intensity of the LIMK1 immunoreactivity is diminished.LIMK1 immunoreactivity is not colocalized with most small spots ofSV2 label (arrows). On the basis of their proximity to the axonbundles, these spots of SV2 label likely correspond to axon terminalsor small axonal branches (compare Fig. 3A). E20: Strong SV2 immuno-reactivity is found in distal axons and nerve terminals. Presynapticterminals can usually be recognized by the pattern of SV2 label(arrows; compare Fig. 3C). Weak LIMK1 immunoreactivity overlapssome SV2-labeled structures (open arrowhead), which, based on theirlinear appearance, are likely to be axon shafts. However, little or noLIMK1 immunoreactivity colocalizes with the SV2-labeled nerveterminals. P0: SV2 label is found only in nerve terminals (arrows) andadjacent axonal segments. The nerve terminal is easily identified atthis and all subsequent stages. Little LIMK1 is observed in thevicinity of nerve terminals. P6: Weak LIMK1 label (arrows) colocalizeswith some SV2-labeled nerve terminals. P14: LIMK1 label in nerveterminals is greater than at P6. The variability in the intensity ofLIMK1 label within nerve terminals is larger than observed in theadult. P21: LIMK1 label in nerve terminals is brighter than at P14.The intensity of the LIMK1 label within nerve terminals is also lessvariable than at P14, but still more variable than in the adult. Adult:The pattern of LIMK1 label and SV2 label are nearly identical (seealso Fig. 2A). Scale bar 5 50 µm.
326 J.Y. WANG ET AL.

Figure 5
LIMK1 IN PRESYNAPTIC TERMINALS 327

developmental state of the motor axon terminal or of themuscle, we examined the expression of LIMK1 in musclesat various times after crushing the sciatic nerve (Fig. 6).After a crush injury to the nerve, the injured axonsdegenerate distal to the crush. Subsequently, synapses areregenerated at the former synaptic sites by axons growingthrough the crush and down the remaining nerve sheath(see Rich and Lichtman, 1989, and references therein fordiscussion of reinnervation of neuromuscular synapses).
Five days after nerve crush, little LIMK1 immunoreac-tivity remained at former synaptic sites (Fig. 6), and SV2label could not be detected (data not shown). This loss ofSV2 immunoreactivity indicates the loss of the presynap-tic terminal after the crush. By 9 days after nerve crush,SV2-positive terminal arborizations could be found, butonly rare, faint spots of LIMK1 immunoreactivity weredetected at these newly regenerated synapses. IncreasingLIMK1 expression at regenerating synapses was evidentby 14 days after nerve crush, and by 23 days LIMK1expression had returned to its normal adult levels. Thepattern of LIMK1 label in presynaptic terminals 9 and 14days after the crush is reminiscent of the pattern seen atdeveloping synapses 6 and 14 days after birth, respectively(see Fig. 5). No increase in axonal LIMK1 immunoreactiv-ity accompanied the reaccumulation of synaptic LIMK1 atany of the time points examined (Fig. 6). Thus, as duringpostnatal development, the expression of LIMK1 in regen-erating motor nerve terminals lagged 1–2 weeks behindSV2.
LIMK1 is concentrated in presynapticterminals of the adult spinal cord
Our finding that LIMK1 is concentrated at the neuromus-cular synapse encouraged us to investigate whether LIMK1is also concentrated at central synapses. We have focusedour analysis on the synapses around large cells in theventral horn of the spinal cord. On the basis of their sizeand position, many of these cells are likely to be a-motoneu-rons (see Burke, 1998; Walmsley et al., 1998, and refer-ences therein for discussion and illustration of ventralhorn cellular and synaptic anatomy). By using electronand confocal immunomicroscopy, we found that LIMK1immunoreactivity is concentrated in some, but not all, ofthe presynaptic terminals forming synapses in the adultventral spinal cord (Figs. 7 and Fig. 8 Adult). LIMK1-immunoreactive terminals were observed to form syn-apses both on the soma of neurons (Fig. 7B and C) and ondendrites (Fig. 7D and E). Some dendrites were observedto receive inputs from both labeled and unlabeled termi-nals (Fig. 7D and E).
Analysis of spinal cord segments that had been injuredby transection of the ventral and dorsal roots furthersupported the hypothesis that LIMK1 is concentrated inpresynaptic terminals of central synapses. In the intactadult spinal cord, a dense, nearly continuous collar ofSV2-labeled synaptic boutons outlined the large neuronsof the ventral horn (Fig. 7B and Fig. 8 Adult). Some, butnot all, of these boutons also showed strong LIMK1 label.After transection of ventral and dorsal roots, the numberof LIMK1- and SV2-immunoreactive boutons outlining theperimeter of large neurons in the affected segments wassubstantially reduced (Fig. 8 Lesioned). This result isconsistent with the well-documented withdrawal (or ‘‘strip-ping’’) of presynaptic terminals from the surface of axoto-mized motoneurons (Watson, 1974; Matthews and Nelson,
1975; Mendell et al., 1976; Eugene and Taxi, 1991) andfurther supports the idea that the punctate LIMK1 labelalong the surface of uninjured motoneurons represents theconcentration of LIMK1 in presynaptic terminals.
Having established that LIMK1 is concentrated in pre-synaptic terminals of the adult ventral spinal cord, weinvestigated the time course of LIMK1 accumulation inthese presynaptic terminals. In the first days after birth,few SV2-labeled boutons in the ventral spinal cord showeddetectable LIMK1 immunoreactivity (Fig. 8, P0 and P6).The intensity of LIMK1 label that was colocalized withSV2-labeled boutons increased only gradually, similar tothe gradual postnatal increase at developing neuromuscu-lar synapses. By P14, many boutons showed weak LIMK1immunoreactivity, and some boutons were strongly immu-noreactive. However, even at P14 (Fig. 8) and P21 (notshown), LIMK1 label in boutons appeared less intensethan in the adult.
An additional finding from these studies was that thecytoplasm and nucleus of motoneurons in the adult areLIMK1 immunoreactive (Figs. 7 and 8). Identification ofLIMK1 protein within the soma of motoneurons was notentirely unexpected, because in situ hybridization studieshad previously demonstrated relatively high amounts ofLIMK1 mRNA in large neurons of the ventral spinal cord(Proschel et al., 1995; Mori et al., 1997), and we had foundLIMK1 at motoneuron terminals (Fig. 2). However, not allproteins synthesized by motoneurons are easily detectedin the soma. For example, we observed little SV2 immuno-reactivity in the motoneuron cell body (Figs. 7 and 8).Thus, although SV2 and LIMK1 are trafficked to a similarlocation within the motoneuron terminal, their relativeabundance in the terminal and soma is different.
LIMK1 accumulated in motoneuron nuclei with a timecourse that roughly paralleled its accumulation in motoneu-ron terminals and synapses of the ventral horn. Althoughat P0, the cytoplasm of motoneurons was strongly LIMK1immunoreactive, LIMK1 label was not seen in motoneuronnuclei at this stage (Fig. 8). The concentration of LIMK1 inthe nucleus gradually increased over the first few weeksafter birth, until by P14 the intensity of LIMK1 immunore-activity was similar in the nucleus and cytoplasm.
DISCUSSION
This study is the first to provide a detailed characteriza-tion of the distribution of LIMK1 protein within neurons ofadult and developing animals. Our major findings are thatLIMK1 is concentrated in presynaptic terminals of adultneuromuscular and spinal cord synapses and that LIMK1accumulates in developing and regenerating nerve termi-nals long after these terminals establish contact with theirtargets. Our experiments also revealed developmentalchanges in the amount of LIMK1 along axons and inneuronal nuclei.
LIMK1 protein-binding domainsand kinase activity
LIMK1, like a number of other synapse-associated pro-teins, has several domains that are likely to function inprotein-protein binding. These include two LIM domains(Sanchez-Garcia and Rabbitts, 1994; Dawid et al., 1995;Dawid et al., 1998), a PDZ-like domain (Bredt, 1998; Hataet al., 1998), and a serine-proline-rich region. Several ofthe proteins already known to bind to LIMK1 are present
328 J.Y. WANG ET AL.

Fig. 6. LIMK1 reappears slowly at regenerating neuromuscularsynapses. The sciatic nerve of adult rats was crushed at midthighlevel. Soleus muscle was harvested 5, 9, 14, and 23 days after crush.The right panels show either a-BTX (5 day) or both SV2 and NF-Mlabel (9, 14, and 23-day). 5 day: Former synaptic sites were identifiedby labeling AChRs with a-BTX. LIMK1 is dramatically decreased atthe former synaptic sites (arrow) compared with LIMK1 at innervatedadult synapses (see Figs. 2 and 5). The faint speckles of LIMK1 label inthe end-plate area may represent residual nerve terminal LIMK1 thathas not yet been degraded or LIMK1 in the Schwann cell or musclecomponent of the injured synapse. At 5 days after crush, no SV2 labelis detected at the former synaptic site (data not shown), consistent
with the expected degeneration of the nerve terminal and the portionof the axon distal to the site of injury. 9 day: Some NF-M-labeled axonsreappeared in the soleus muscle, often ending in SV2-labeled termi-nals characteristic of neuromuscular synapses. Faint LIMK1 labelwas observed at the synapses and along the course of the axons. 14day: Although most areas of the regenerating synapse display little orno LIMK1 immunoreactivity, small areas within some synapsesdisplay brighter label. This variability is similar to that seen atdeveloping synapses (compare Fig. 5 P6, P14, and P21). 23 day:LIMK1 expression has returned to adult levels and is preciselycolocalized with SV2 at reinnervated synaptic sites. Scale bar 520 µm.
in nerve terminals, including neuregulin (Sandrock et al.,1995; Wang et al., 1998), PKC isoforms (Kuroda et al.,1996; Majewski et al., 1997), and actin (Hirokawa et al.,1989; Yang et al., 1998).
The kinase domain of LIMK1 can phosphorylate cofilinon serine 3 (Arber et al., 1998; Rosenblatt and Mitchison,1998; Yang et al., 1998) and thereby inhibit degradation of
F-actin. When LIMK1 is expressed in differentiated PC12cells, the actin cytoskeleton is dramatically altered. In PC12cells that overexpress LIMK1, the growth cones on neuritesare enlarged, and their F-actin content is increased. Con-versely, in PC12 cells transfected with a mutated form ofLIMK1-lacking kinase activity, the growth cones are reducedin size (Arber et al., 1998). However, whether cofilin or related
Figure 7
330 J.Y. WANG ET AL.

proteins such as actin depolymerizing factor (ADF) could serveas a substrate of LIMK1 in nerve terminals is unclear, becausemembers of theADF/cofilin family of actin-regulating proteinshave not yet been reported in adult nerve terminals.
LIMK1 binds to neuregulins through interaction of theLIM domain region of LIMK1 with the NRG cytoplasmictail (Wang et al., 1998). NRGs have now been found bothpresynaptically and postsynaptically at the neuromuscu-lar synapse (Burden, 1998). Our finding that LIMK1 is apresynaptic terminal protein focuses further investigationof the NRG-LIMK1 interaction on the presynaptic side ofsynapses. If LIMK1 regulates actin in the nerve terminal,then alteration of the neuronal cytoskeleton may be adownstream effect of intercellular signaling initiatedthrough the NRG cytoplasmic tail.
Accumulation of LIMK1at maturing synapses
Although functional synapses first form in the rathindlimb and spinal cord during embryonic development,these synapses gradually acquire their adult characteris-tics over the first few weeks after birth (Conradi andRonnevi, 1977; Dennis et al., 1981; Grinnell, 1995; Nguyenand Lichtman, 1996; Seebach and Mendell, 1996). We havefound that LIMK1 accumulates in the nerve terminals ofthese synapses roughly in parallel with this synapticmaturation.
Many of the proteins involved in the functions of adultpresynaptic terminals are already abundant in terminalswithin the first few days after they form synapses (Lupaand Hall, 1989; Li and Dahlstrom, 1993), suggesting thatthese proteins perform functions required by the imma-ture and the adult synapse. However, a few proteinspresent in adult nerve terminals accumulate later, as thesynapse matures to its adult phenotype. Two such proteinsare the guanosine triphosphate (GTP)-binding proteinrab3a (Stettler et al., 1994) and the SNAP-receptor bind-ing protein complexin (Ono et al., 1998). Perturbations inboth rab3a and complexin function can modulate neuro-transmitter release, and each has been proposed to regu-late maturation of the transmitter release process (Ober-hauser et al., 1992; Lledo et al., 1993; McMahon et al.,1995; Ono et al., 1998). To our knowledge LIMK1 is the
first kinase to join this small group of proteins known tobecome concentrated at synapses during their maturation.In contrast to the maturational increase in LIMK1, twogrowth-associated proteins, GAP-43 and CAP-23, havebeen shown to gradually decrease at neuromuscular syn-apses over the first few weeks after birth (Caroni andBecker, 1992; Caroni et al., 1997), and it has been sug-gested that this decline contributes to the reduction inmorphological plasticity seen at maturing motoneuronterminals.
Our findings suggest that LIMK1 plays a role in adultnerve terminals. We predict that the activities controlledby LIMK1 will appear or mature in presynaptic terminalsduring the period of LIMK1 accumulation. On the basis ofthese considerations and the finding that LIMK1 canregulate cofilin, two candidate functions of nerve terminalLIMK1 are regulation of the morphological plasticity ofthe nerve terminal and regulation of neurotransmitterrelease. Both of these aspects of nerve terminal physiologyare known to change dramatically during the period overwhich LIMK1 accumulates in nerve terminals (Balice-Gordon and Lichtman, 1990; Bazzy, 1994; Seebach andMendell, 1996, and references therein). Furthermore, ac-tin filament turnover in synaptosomes and cultured neu-rons is regulated by depolarization, and the stabilization ofsynaptosomal actin filaments can affect neurotransmitterrelease (Bernstein and Bamburg, 1985; Bernstein andBamburg, 1989; Bernstein et al., 1998). This raises thepossibility that LIMK1-mediated regulation of the actincytoskeleton may govern some aspects of the nerve termi-nal’s response to electrical activity.
In summary, LIMK1 accumulates in presynaptic termi-nals during synapse maturation, suggesting LIMK1 regu-lates activities of the nerve terminal that change duringthis maturational process. The time course of LIMK1’saccumulation in presynaptic terminals and its previouslydemonstrated effects on actin suggest a role for LIMK1 inthe regulation of nerve terminal morphological stabilityand/or neurotransmitter release. Now that LIMK1 hasbeen identified as a nerve terminal protein, it will beimportant to determine the effects of perturbing LIMK1function on nerve terminal biology.
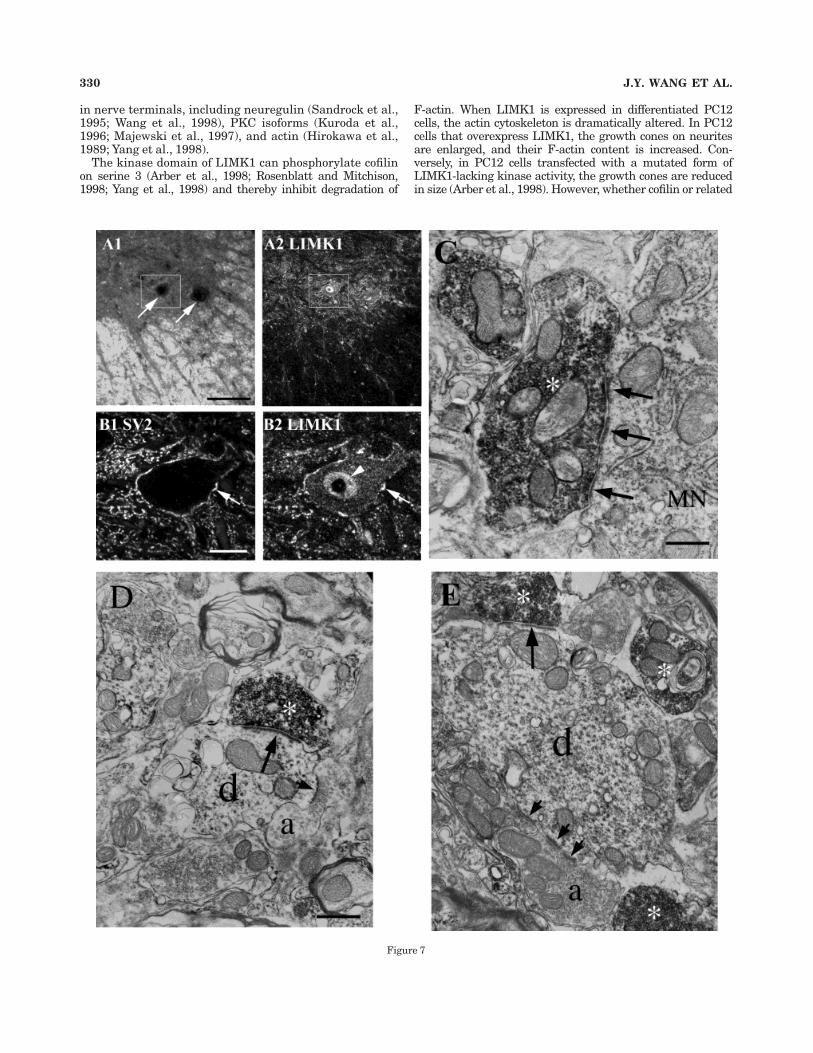
Fig. 7. LIMK1 is concentrated in the presynaptic terminals ofsome synapses in the adult ventral spinal cord. Confocal fluorescencemicroscopy (A2, B) and immunoperoxidase electron microscopy (C–E)were used to visualize LIMK1 immunoreactivity. A: Low magnifica-tion images of the ventral horn region from an adult spinal cordvisualized by using illumination with a darkfield condenser (A1) orfluorescence microscopy to reveal LIMK1 immunoreactivity in thesame field (A2). In these images, the gray matter of the ventral horn isbordered at the bottom (ventral) and very right (lateral) by thesurrounding white matter (lightest areas in A1). Two large ventralhorn cells, which appear darker than the surrounding portions of thegray matter in A1, are indicated by arrows. On the basis of their sizeand position, these cells are likely to be a-motoneurons. Many of thelarger spots of LIMK1 label in the white matter colocalize withHoechst dye 33342 labeling of nuclei (not shown). Because LIMK1label is present in many presynaptic terminals within the gray matter(see below), the smaller puncta of LIMK1 label in the white mattermay correspond to LIMK1 label at synapses along dendrites thatextend out into the white matter. B: Higher magnification fluorescenceimages of the area boxed in panel A. Bright puncta of SV2 label (B1),marking presynaptic terminals, and LIMK1 label (B2) outline thelarge neuron in this field. The arrow points to one of the puncta at
which LIMK1 and SV2 label are colocalized along the edge of theneuron. There is also strong LIMK1 label in the nucleus (arrowhead),but not the nucleolus (unlabeled spot within the nucleus), and lessintense LIMK1 label diffusely through the cytoplasm. In contrast, thecytoplasm and nucleus of this neuron are unlabeled in the SV2 image.In the neuropil surrounding this cell, there are prominent puncta ofLIMK1 label. The pattern of SV2 label in the neuropil appears similar,but not identical, to the pattern of LIMK1 label. Electron microscopydemonstrates that many of the LIMK1-labeled puncta along the edgesof neurons and in the neuropil represent presynaptic terminals (seebelow). C: Electron micrograph demonstrating LIMK1 label in anaxon terminal (asterisk) forming synapses (arrows) on the soma of amotoneuron (MN). D,E: Electron micrographs demonstrating LIMK1label in axon terminals (asterisks) forming synapses (long arrows) ondendrites (d) in the ventral horn of the spinal cord. Note that thesesame dendrites also form synapses (short arrows) with unlabeledterminals (a). In C–E, both long and short arrows point to densematerial underlying the postsynaptic membrane, and clear synapticvesicles can be observed in each of the LIMK1-labeled presynapticterminals. Scale bars 5 100 µm in A; 20 µm in B; 0.5 µm in C; 0.67 µmin D,E.
LIMK1 IN PRESYNAPTIC TERMINALS 331

Figure 8
332 J.Y. WANG ET AL.

ACKNOWLEDGMENTS
We thank Heather Black-Brewster for technical assis-tance; Elizabeth Robertson (Harvard University) for pro-viding the mouse LIMK1 cDNA; Steve L’Hernault (EmoryUniversity), Sho Ono (Emory University), Mark Rich(Emory University), Marla Luskin (Emory University andLee Hilborne (UCLA School of Medicine) for criticallyreading the manuscript; and Arthur English (Emory Uni-versity) for help with denervations.
LITERATURE CITED
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, LipmanDJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of proteindatabase search programs. Nucleic Acids Res 25:3389–3402.
Anderson MJ, Klier FG, Tanguay KE. 1984. Acetylcholine receptor aggrega-tion parallels the deposition of a basal lamina proteoglycan duringdevelopment of the neuromuscular junction. J Cell Biol 99:1769–1784.
Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, Bernard O,Caroni P. 1998. Regulation of actin dynamics through phosphorylationof cofilin by LIM-kinase. Nature 393:805–809.
Astrow SH, Sutton LA, Thompson WJ. 1992. Developmental and neuralregulation of a subsarcolemmal component of the rat neuromuscularjunction. J Neurosci 12:1602–1615.
Balice-Gordon RJ, Lichtman JW. 1990. In vivo visualization of the growthof pre- and postsynaptic elements of neuromuscular junctions in themouse. J Neurosci 10:894–908.
Bazzy AR. 1994. Developmental changes in rat diaphragm endplateresponse to repetitive stimulation. Brain Res Dev Brain Res 81:314–317.
Bernard O, Ganiatsas S, Kannourakis G, Dringen R. 1994. Kiz-1, a proteinwith LIM zinc finger and kinase domains, is expressed mainly inneurons. Cell Growth Differ 5:1159–1171.
Bernstein BW, Bamburg JR. 1985. Reorganization of actin in depolarizedsynaptosomes. J Neurosci 5:2565–2569.
Bernstein BW, Bamburg JR. 1989. Cycling of actin assembly in synapto-somes and neurotransmitter release. Neuron 3:257–265.
Bernstein BW, DeWit M, Bamburg JR. 1998. Actin disassembles reversiblyduring electrically induced recycling of synaptic vesicles in culturedneurons. Brain Res Mol Brain Res 53:236–251.
Bredt DS. 1998. Sorting out genes that regulate epithelial and neuronalpolarity. Cell 94:691–694.
Buckley K, Kelly RB. 1985. Identification of a transmembrane glycoproteinspecific for secretory vesicles of neural and endocrine cells. J Cell Biol100:1284–1294.
Burden SJ. 1998. The formation of neuromuscular synapses. Genes Dev12:133–148.
Burke RE. 1998. Spinal cord: ventral horn. In: Shepherd GM, editor. Thesynaptic organization of the brain. New York: Oxford University Press.p 77–120.
Burns ME, Augustine GJ. 1995. Synaptic structure and function: dynamicorganization yields architectural precision. Cell 83:187–194.
Caroni P, Aigner L, Schneider C. 1997. Intrinsic neuronal determinantslocally regulate extrasynaptic and synaptic growth at the adult neuro-muscular junction. J Cell Biol 136:679–692.
Caroni P, Becker M. 1992. The downregulation of growth-associatedproteins in motoneurons at the onset of synapse elimination is con-trolled by muscle activity and IGF1. J Neurosci 12:3849–3861.
Cheng AK, Robertson EJ. 1995. The murine lim-kinase gene (limk) encodesa novel serine threonine kinase expressed predominantly in trophoblastgiant cells and the developing nervous system. Mech Dev 52:187–197.
Condon K, Silberstein L, Blau HM, Thompson WJ. 1990. Development ofmuscle fiber types in the prenatal rat hindlimb. Dev Biol 138:256–274.
Conradi S, Ronnevi LO. 1977. Ultrastructure and synaptology of the initialaxon segment of cat spinal motoneurons during early postnatal develop-ment. J Neurocytol 6:195–210.
Dawid IB, Breen JJ, Toyama R. 1998. LIM domains: multiple roles asadapters and functional modifiers in protein interactions. Trends Genet14:156–162.
Dawid IB, Toyama R, Taira M. 1995. LIM domain proteins [review]. C RAcad Sci III 318:295–306.
Dennis MJ. 1981. Development of the neuromuscular junction: inductiveinteractions between cells. Annu Rev Neurosci 4:43–68.
Dennis MJ, Ziskind-Conhaim L, HarrisAJ. 1981. Development of neuromus-cular junctions in rat embryos. Dev Biol 81:266–279.
Edwards DC, Gill GN. 1999. Structural features of LIM kinase that controleffects on the actin cytoskeleton. J Biol Chem 274:11352–11361.
Eugene D, Taxi J. 1991. Effects of axotomy on synaptic transmission andstructure in frog sympathetic ganglia. J Neurocytol 20:404–419.
Gilmor ML, Nash NR, Roghani A, Edwards RH, Yi H, Hersch SM, Levey AI.1996. Expression of the putative vesicular acetylcholine transporter inrat brain and localization in cholinergic synaptic vesicles. J Neurosci16:2179–2190.
Grinnell AD. 1995. Dynamics of nerve-muscle interaction in developing andmature neuromuscular junctions. Physiol Rev 75:789–834.
Hall ZW, Sanes JR. 1993. Synaptic structure and development: theneuromuscular junction. Cell 72:99–121.
Hata Y, Nakanishi H, Takai Y. 1998. Synaptic PDZ domain-containingproteins. Neurosci Res 32:1–7.
Hirata K, Zhou C, Nakamura K, Kawabuchi M. 1997. Postnatal develop-ment of Schwann cells at neuromuscular junctions, with special refer-ence to synapse elimination. J Neurocytol 26:799–809.
Hirokawa N, Sobue K, Kanda K, Harada A, Yorifuji H. 1989. The cytoskel-etal architecture of the presynaptic terminal and molecular structure ofsynapsin 1. J Cell Biol 108:111–126.
Kuroda S, Tokunaga C, Kiyohara Y, Higuchi O, Konishi H, Mizuno K, GillGN, Kikkawa U. 1996. Protein-protein interaction of zinc finger limdomains with protein kinase c. J Biol Chem 271:31029–31032.
Lee MK, Cleveland DW. 1996. Neuronal intermediate filaments. Annu RevNeurosci 19:187–217.
Levitan IB. 1994. Modulation of ion channels by protein phosphorylationand dephosphorylation. Annu Rev Physiol 56:193–212.
Li JY, Dahlstrom AB. 1993. Distribution of GAP-43 in relation to CGRP andsynaptic vesicle markers in rat skeletal muscles during development.Brain Res Dev Brain Res 74:269–282.
Lledo PM, Vernier P, Vincent JD, Mason WT, Zorec R. 1993. Inhibition ofRab3B expression attenuates Ca(21)-dependent exocytosis in rat ante-rior pituitary cells. Nature 364:540–544.
Lupa MT, Hall ZW. 1989. Progressive restriction of synaptic vesicle proteinto the nerve terminal during development of the neuromuscularjunction. J Neurosci 9:3937–3945.
Fig. 8. LIMK1 accumulates postnatally at synapses of the ventralspinal cord. Cross sections of rat spinal cord from different developmen-tal stages were double labeled with the SV2 and LK1-EU1 antibodies.Presynaptic terminals appear as small spots (puncta) of strong SV2immunoreactivity. P0: Few SV2-labeled puncta are LIMK1 immunore-active, indicating that LIMK1 is not concentrated at most P0 synapsesof the ventral horn. At this stage, the cytoplasm of motoneurons, butnot their nuclei, is LIMK1 immunoreactive. P6: The density ofsynaptic boutons, as assessed by SV2 label, is greater at P6 than at P0.As at P0, few synaptic boutons are LIMK1 immunoreactive. P14: Asmall proportion of the synaptic boutons are strongly LIMK1 immuno-reactive (arrows). Many others are weakly to moderately immunoreac-tive. Nuclei of motoneurons now display some LIMK1 label. Adult: AtP60 the proportion of LIMK1-labeled synaptic boutons is larger thanat P14, and more of the synapses are strongly labeled (arrows). Notethe prominent ‘‘collar’’ of SV2-labeled boutons outlining the soma oflarge neurons. Many of these boutons are strongly LIMK1 immunore-active (arrows). Lesioned: Ten days after unilateral transection ofventral and dorsal roots, the near continuous collar of SV2 immunore-activity observed around uninjured motoneurons is no longer evident.Prominent puncta of colocalized LIMK1/SV2 immunoreactivity alongthe perimeter of the motoneurons are rarely observed (arrow marksone example). This finding suggests that the puncta of SV2 andLIMK1 label at the edge of uninjured motoneurons represent nerveterminals, which withdraw from the surface of the injured neurons.LIMK1 and SV2 label of other synapses (i.e., synapses not on the somaof large neurons) was not evidently different from normal adultventral horn. There also were no changes noted in cytoplasmic ornuclear LIMK1 immunoreactivity. The images labeled ‘‘Adult’’ and‘‘Lesioned’’ are from the intact and operated sides, respectively, of thesame section. Scale bars 5 30 µm in P0–P14 and 45 µm (Adult 1Lesioned). For each panel, ventral is down and lateral is to the right.
LIMK1 IN PRESYNAPTIC TERMINALS 333

Majewski H, Kotsonis P, Iannazzo L, Murphy TV, Musgrave IF. 1997.Protein kinase C and transmitter release. Clin Exp Pharmacol Physiol24:619–623.
Matthews MR, Nelson VH. 1975. Detachment of structurally intact nerveendings from chromatolytic neurones of rat superior cervical ganglionduring the depression of synaptic transmission induced by post-ganglionic axotomy. J Physiol (Lond) 245:91–135.
McMahon HT, Missler M, Li C, Sudhof TC. 1995. Complexins: cytosolicproteins that regulate SNAP receptor function. Cell 83:111–119.
Mendell LM, Munson JB, Scott JG. 1976. Alterations of synapses onaxotomized motoneurones. J Physiol (Lond) 255:67–79.
Mizuno K, Okano I, Ohashi K, Nunoue K, Kuma K, Miyata T, Nakamura T.1994. Identification of a human cDNA encoding a novel protein kinasewith two repeats of the LIM/double zinc finger motif. Oncogene9:1605–1612.
Mori T, Okano I, Mizuno K, Tohyama M, Wanaka A. 1997. Comparison oftissue distribution of two novel serine/threonine kinase genes contain-ing the lim motif (limk-1 and limk-2) in the developing rat. Brain ResMol Brain Res 45:247–254.
Nguyen QT, Lichtman JW. 1996. Mechanism of synapse disassembly at thedeveloping neuromuscular junction. Curr Opin Neurobiol 6:104–112.
Nunoue K, Ohashi K, Okano I, Mizuno K. 1995. LIMK-1 and LIMK-2, twomembers of a LIM motif-containing protein kinase family. Oncogene11:701–710.
Oberhauser AF, Monck JR, Balch WE, Fernandez JM. 1992. Exocytoticfusion is activated by Rab3a peptides. Nature 360:270–273.
Okano I, Hiraoka J, Otera H, Nunoue K, Ohashi K, Iwashita S, Hirai M,Mizuno K. 1995. Identification and characterization of a novel family ofserine threonine kinases containing two n-terminal lim motifs. J BiolChem 270:31321–31330.
Ono S, Baux G, Sekiguchi M, Fossier P, Morel NF, Nihonmatsu I, Hirata K,Awaji T, Takahashi S, Takahashi M. 1998. Regulatory roles of complex-ins in neurotransmitter release from mature presynaptic nerve termi-nals. Eur J Neurosci 10:2143–2152.
Proschel C, Blouin MJ, Gutowski NJ, Ludwig R, Noble M. 1995. Limk1 ispredominantly expressed in neural tissues and phosphorylates serine,threonine and tyrosine residues in vitro. Oncogene 11:1271–1281.
Rich MM, Lichtman JW. 1989. In vivo visualization of pre- and postsynapticchanges during synapse elimination in reinnervated mouse muscle. JNeurosci 9:1781–1805.
Rosenblatt J, Mitchison TJ. 1998. Actin, cofilin and cognition. Nature393:739–740.
Rubinstein NA, Kelly AM. 1981. Development of muscle fiber specializationin the rat hindlimb. J Cell Biol 90:128–144.
Sanchez-Garcia I, Rabbitts TH. 1994. The LIM domain: a new structuralmotif found in zinc-finger-like proteins. Trends Genet 10:315–320.
Sandrock AW, Goodearl ADJ, Yin QW, Chang D, Fischbach GD. 1995. Aria isconcentrated in nerve terminals at neuromuscular junctions and atother synapses. J Neurosci 15:6124–6136.
Seebach BS, Mendell LM. 1996. Maturation in properties of motoneuronsand their segmental input in the neonatal rat. J Neurophysiol 76:3875–3885.
Son YJ, Thompson WJ. 1995a. Nerve sprouting in muscle is induced andguided by processes extended by Schwann cells. Neuron 14:133–141.
Son YJ, Thompson WJ. 1995b. Schwann cell processes guide regenerationof peripheral axons. Neuron 14:125–132.
Stettler O, Moya KL, Zahraoui A, Tavitian B. 1994. Developmental changesin the localization of the synaptic vesicle protein rab3A in rat brain.Neuroscience 62:587–600.
Tassabehji M, Metcalfe K, Karmiloff-Smith A, Carette MJ, Grant J, DennisN, Reardon W, Splitt M, Read AP, Donnai D. 1999. Williams syndrome:use of chromosomal microdeletions as a tool to dissect cognitive andphysical phenotypes. Am J Hum Genet 64:118–125.
Tokuda M, Hatase O. 1998. Regulation of neuronal plasticity in the centralnervous system by phosphorylation and dephosphorylation. Mol Neuro-biol 17:137–156.
Valtorta E, Benfenati F, Basudev H. 1996. Role of protein kinases in nerveterminal maturation and function. Biochem Soc Trans 24:645–653.
Walmsley B, Alvarez FJ, Fyffe RE. 1998. Diversity of structure and functionat mammalian central synapses. Trends Neurosci 21:81–88.
Wang JY, Frenzel KE, Wen D, Falls DL. 1998. Transmembrane neuregulinsinteract with LIM kinase 1, a cytoplasmic protein kinase implicated indevelopment of visuospatial cognition. J Biol Chem 273:20525–20534.
Watson WE. 1974. Cellular responses to axotomy and to related procedures.Br Med Bull 30:112–115.
Woolf CJ, Reynolds ML, Chong MS, Emson P, Irwin N, Benowitz LI. 1992.Denervation of the motor endplate results in the rapid expression byterminal Schwann cells of the growth-associated protein GAP-43. JNeurosci 12:3999–4010.
Yang N, Higuchi O, Ohashi K, Nagata K, Wada A, Kangawa K, Nishida E,Mizuno K. 1998. Cofilin phosphorylation by LIM-kinase 1 and its role inRac-mediated actin reorganization. Nature 393:809–812.
334 J.Y. WANG ET AL.