Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Nov. 2010, p. 4839–4849 Vol. 78, No. 110019-9567/10/$12.00 doi:10.1128/IAI.00248-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Limited Role of Secreted Aspartyl Proteinases Sap1 to Sap6 inCandida albicans Virulence and Host Immune Response in
Murine Hematogenously Disseminated Candidiasis�
Alexandra Correia,1 Ulrich Lermann,2 Luzia Teixeira,3,4 Filipe Cerca,4,5 Sofia Botelho,4Rui M. Gil da Costa,4 Paula Sampaio,1 Fatima Gartner,4 Joachim Morschhauser,2
Manuel Vilanova,4,5 and Celia Pais1*Centre of Molecular and Environmental Biology (CBMA), Department of Biology, University of Minho, 4710-057 Braga, Portugal1;
Institut fur Molekulare Infektionsbiologie, Universitat Wurzburg, Josef Schneider Str. 2, D-97080 Wurzburg, Germany2;Unidade Multidisciplinar de Investigacao Biomedica (UMIB), Universidade do Porto, Largo Prof. Abel Salazar 2, 4099-003 Porto,
Portugal3; Instituto de Ciencias Biomedicas de Abel Salazar (ICBAS), Universidade do Porto, Largo Prof. Abel Salazar 2,4099-003 Porto, Portugal4; and Instituto de Biologia Molecular e Celular (IBMC), Rua Campo Alegre,
4099-003 Porto, Portugal5
Received 12 March 2010/Returned for modification 23 March 2010/Accepted 25 July 2010
Candida albicans secreted aspartyl proteinases (Saps) are considered virulence-associated factors. Severalmembers of the Sap family were claimed to play a significant role in the progression of candidiasis establishedby the hematogenous route. This assumption was based on the observed attenuated virulence of sap-nullmutant strains. However, the exclusive contribution of SAP genes to their attenuated phenotype was notunequivocally confirmed, as the Ura status of these mutant strains could also have contributed to theattenuation. In this study, we have reassessed the importance of SAP1 to SAP6 in a murine model ofhematogenously disseminated candidiasis using sap-null mutant strains not affected in their URA3 geneexpression and compared their virulence phenotypes with those of Ura-blaster sap mutants. The mediansurvival time of BALB/c mice intravenously infected with a mutant strain lacking SAP1 to SAP3 was equivalentto that of mice infected with wild-type strain SC5314, while those infected with mutant strains lacking SAP5showed slightly extended survival times. Nevertheless, no differences could be observed between the wild typeand a �sap456 mutant in their abilities to invade mouse kidneys. Likewise, a deficiency in SAP4 to SAP6 hadno noticeable impact on the immune response elicited in the spleens and kidneys of C. albicans-infected mice.These results contrast with the behavior of equivalent Ura-blaster mutants, which presented a significantreduction in virulence. Our results suggest that Sap1 to Sap6 do not play a significant role in C. albicansvirulence in a murine model of hematogenously disseminated candidiasis and that, in this model, Sap1 to Sap3are not necessary for successful C. albicans infection.
The polymorphic yeast Candida albicans is an importantopportunistic human pathogen causing infections that rangefrom superficial mucosal lesions to life-threatening systemicdisease. It is by far the most common cause of fungal invasiveinfections, which could be attributed to the little immunosup-pression required to predispose an individual to invasive Can-dida infections (39). Host physical barriers and immune systemintegrity are crucial factors in controlling the establishment ofinfection. However, the high adaptability of C. albicans todifferent host niches, by the expression of appropriate sets ofvirulence-related genes, is also a determinant (19, 51). Severalof these virulence attributes may participate in and influencethe infective process, depending on the site and stage of inva-sion and on the nature of the host response (37). The secretionof hydrolytic enzymes during infection is required as a viru-lence attribute to aid adhesion, invasion, and the destruction ofhost immune factors, in addition to nutrient acquisition (21).
Among these enzymes, secreted aspartyl proteinases (Sap),encoded by a 10-member gene family (SAP1 to SAP10) havebeen the most extensively studied (35). The 10 SAP genes thatcompose this family can be divided into subfamilies based onamino acid sequence homology alignments (SAP1 to SAP3,SAP4 to SAP6, SAP9, and SAP10). These genes exhibit differ-ential expression profiles at different stages and sites of infec-tion (33, 35, 46, 49) and have been linked with the virulence ofthe fungus since their discovery (10, 27, 48).
The contribution of the SAP1 to SAP3, SAP4 to SAP6, SAP7,and SAP9 and SAP10 genes to virulence in different models ofinfection has been studied by using sap-null mutant strains (1,13, 16, 20, 23, 25, 26, 29, 34, 43, 54). The subfamily consistingof the SAP4 to SAP6 genes, in particular, was shown to con-tribute significantly to C. albicans virulence in models of acutesystemic candidiasis, murine peritonitis, and Candida gastro-intestinal infection (16, 25, 43). These genes are expressedmainly during hypha formation (22, 34), and SAP5 in particularwas found to be upregulated at all time points after eitherintravenous (i.v.) or intraperitoneal (i.p.) infection of mice (44,50, 57).
Hube et al. (20) previously reported that �sap1, �sap2, and�sap3 null mutants displayed attenuated virulence in models of
* Corresponding author. Mailing address: Centro de Biologia Mo-lecular e Ambiental da Universidade do Minho (CMBA), Departa-mento de Biologia, Campus de Gualtar, 4710-057 Braga, Portugal.Phone: 351 253604312. Fax: 351 253678980. E-mail: [email protected].
� Published ahead of print on 2 August 2010.
4839
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

acute systemic candidiasis. The triple deletion of SAP4 to SAP6resulted in a more marked impact on C. albicans virulence insimilar experimental settings, suggesting an important role forthese hypha-related genes in the establishment of disseminatedcandidiasis (43). These mutant strains were generated fromauxotrophic laboratory strain CAI4 with the most commonmethod used for disrupting genes in C. albicans, the Ura-blaster technique (17). The use of the URA3 marker for mutantconstruction in C. albicans can lead to a misinterpretation ofthe results in mutant virulence studies (5, 8, 11, 28). Althoughthis can now be overcome by the integration of URA3 at theENO1 (52) or RPS10 (8) locus, the mutant strains used inearlier studies did not share a site of URA3 integration. There-fore, it is conceivable that the Ura status could have influencedthe results, and thus, the attenuated nature of these mutantsduring acute systemic candidiasis remains to be confirmed un-equivocally (38). Therefore, in this study, we have used a set of�sap null mutants constructed, as described previously byLermann and Morschhauser (29), from prototrophic wild-type(WT) strain SC5314 using the SAT1-flipping strategy (41) toreaddress the importance of the SAP1 to SAP6 genes in amurine model of hematogenously disseminated candidiasis. Inaddition, we analyzed the histopathology of several organs andaspects of the immune response elicited in the spleens andkidneys of BALB/c mice infected with the wild-type strainand a sap-null mutant lacking SAP4 to SAP6.
MATERIALS AND METHODS
Mice. Male BALB/c mice, 8 to 10 weeks old, were purchased from CharlesRiver (Barcelona, Spain) and kept under specific-pathogen-free conditions at theAnimal Facility of the Instituto de Ciencias Biomedicas Abel Salazar, Porto,Portugal. All procedures involving mice were performed according to the Euro-pean Convention for the Protection of Vertebrate Animals Used for Experimen-tal and Other Scientific Purposes (ETS 123), the 86/609/EEC directive, andPortuguese rules (DL 129/92).
Candida albicans and culture conditions. The C. albicans strains used in thisstudy are listed in Table 1. All strains were maintained as frozen stocks in 30%glycerol at �80°C. Yeast growth was tested in synthetic glucose minimal (2%
glucose, 0.67% Bacto yeast nitrogen base without amino acids) and complexyeast-peptone-dextrose (YPD) (2% glucose, 1% yeast extract, 2% Bacto pep-tone) liquid media at 30°C and 37°C in a shaking incubator for 24 h. Growth wasmeasured at 60-min intervals, and the generation times were calculated. Hyphaformation was induced by the addition of calf serum (10%) (Invitrogen, Carls-bad, CA) or N-acetylglucosamine (2.5 mM) (Sigma, St. Louis, MO) at 37°C. Toprepare the inocula for infection, C. albicans strains were grown in a shakingincubator for 14 h at 30°C in Winge medium (0.2% glucose, 0.3% yeast extract).Yeast cells were harvested, washed twice with sterile, nonpyrogenic phosphate-buffered saline (PBS), counted in a hemocytometer, and resuspended at theappropriate concentrations. Inocula were confirmed by CFU counts on Sab-ouraud dextrose agar (Difco, Detroit, MI) for up to 48 h at 37°C.
C. albicans hematogenously disseminated infections. Mice were injected in-travenously (i.v.) in the lateral tail vein with 1 � 105 or 5 � 105 C. albicans yeastcells in 0.2 ml PBS. To evaluate the progress of hematogenously disseminatedcandidiasis, mice were weighed and monitored twice daily. Moribund mice werehumanely terminated, and their deaths were recorded as occurring on the fol-lowing day.
Other groups of mice were infected with 5 � 104 yeast cells and sacrificed 3and 7 days postinfection to determine organ fungal burden and/or immunologicalparameters. Control mice were injected i.v. with PBS. Kidneys were asepticallyremoved, weighed, homogenized, and quantitatively cultured on Sabouraud dex-trose agar (Difco) at 37°C. Values are expressed as log CFU per gram of tissue.Alternatively, kidneys, liver, lungs, and brain were fixed in 10% phosphate-buffered formaldehyde, followed by periodic acid-Schiff (PAS) reagent stainingand counterstaining of the paraffin-embedded tissues with hematoxylin in orderto evaluate both fungal morphology and the composition and distribution ofinflammatory infiltrates.
Quantitative real-time PCR (qRT-PCR). BALB/c mice were i.v. infected with1 � 105 C. albicans SC5314, SAP123MS4C, and SAP456MS4A yeast cells. TotalRNA was isolated from the kidneys of three mice per group 3 and 7 days afterinfection. Briefly, the kidneys were removed, homogenized in PBS, and centri-fuged at 1,500 � g at 4°C for 10 min. Pellets were washed twice with ice-coldRNase-free water and frozen in liquid nitrogen until RNA extraction, which wasperformed by using the hot acidic phenol method (4). Total RNA was incubatedwith DNase I, amplification grade (Invitrogen), for 15 min at room temperatureto eliminate genomic DNA contamination. DNase I was inactivated according tothe manufacturer’s instructions.
The Superscript III Platinum two-step qRT-PCR kit with SYBR green (In-vitrogen) was used to generate first-strand cDNA from each DNase I-treatedRNA sample, as follows: 10 min at 65°C, 60 min at 37°C, and 10 min at 65°C.Quantitative PCR (qPCR) was performed with Platinum SYBR green qPCRSuperMix-UDG (Invitrogen). Three microliters of each cDNA sample wasadded to a 25-�l PCR mixture containing 12.5 �l of Platinum SYBR green qPCRSuperMix-UDG, 0.5 �l of 10 �M specific forward and reverse primers (Table 2),and 8.5 �l of RNase-free water. Each reaction was performed with a CorbettRotor-Gene 6000 instrument (Qiagen). Thermocycling conditions for SAP andACT1 quantification were 2 min at 50°C (UDG incubation) and 5 min at 95°C,followed by 40 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s. Thespecificity of each primer pair was verified by the presence of a single melting-temperature peak. The calibration and efficiency of SAP and ACT1 primers wereassessed in titration experiments using C. albicans SC5314 genomic DNA (500 ngto 5 pg) in serial dilutions. A negative (water) control and a four-point curve ofSC5314 genomic DNA were included in each run. DNase I-treated RNA wasused to exclude genomic DNA contamination. SAP1 to SAP10 gene expressionwas normalized to the housekeeping gene ACT1 and analyzed by using both thestandard curve and the comparative threshold cycle (CT) (��CT) methods. Dataare presented as the fold difference in expression relative to wild-type (WT) geneexpression from infected mice. Each experimental condition was performed intriplicate, and reactions were done in duplicate on different days for reproduc-ibility purposes.
Flow cytometric analysis. The assessment of cell surface and cytoplasmiclineage or activation markers on different splenic leukocyte populations wasperformed by flow cytometric analysis (fluorescence-activated cell sorter [FACS]analysis). Spleens were aseptically removed and homogenized to single-cell sus-pensions in Hanks’ balanced salt solution (HBSS) (Sigma). A total number of1 � 106 leukocytes were stained per sample.
The following monoclonal antibodies (MAbs), along with the respective iso-type controls, were used (at previously determined optimal dilutions) for immu-nofluorescence cytometric analysis with a FACScan apparatus (Becton Dickin-son, San Jose, CA) using CELLQUEST software (Becton Dickinson):phycoerythrin (PE) anti-mouse CD40 (1C10) and biotin anti-mouse major his-tocompatibility complex (MHC) class II (NIMR-4) (Southern Biotechnology
TABLE 1. Candida albicans strains used in this study
Strain(s) Genotype Reference
SC5314 Prototrophic wild-type strain 18
SAP123MS4C andSAP123MS4D
sap1�::FRT/sap1�::FRT 29sap2�::FRT/sap2�::FRTsap3�::FRT/sap3�::FRT
SAP456MS4A andSAP456MS4B
sap4�::FRT/sap4�::FRT 29sap5�::FRT/sap5�::FRTsap6�::FRT/sap6�::FRT
SAP4MS4A sap4�::FRT/sap4�::FRT 29SAP5MS4A sap5�::FRT/sap5�::FRT 29SAP6MS4A sap6�::FRT/sap6�::FRT 29CAF2-1 URA3/ura3�::imm434 17
M119 sap1�::hisG/sap1�::hisG 25sap2�::hisG/sap2�::hisGsap3�::hisG/sap3�::hisG::URA3::hisG
DSY459 sap6�::hisG/sap6�::hisG 43sap4�::hisG/sap4�::hisGsap5�::hisG/sap5�::hisG::URA3::hisG
4840 CORREIA ET AL. INFECT. IMMUN.
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Associates, Birmingham, AL); fluorescein isothiocyanate (FITC) anti-mouse/ratFoxp3 (FJK-16S), PE-Cy5 anti-mouse CD4 (L3T4) (RM4-5), and PE anti-mouseF4/80 antigen (BM8) (eBioscience, San Diego, CA); and FITC anti-mouseCD11c (HL3), FITC anti-mouse Ly-6G and Ly-6C (Gr-1) (RB6-8C5), PE anti-mouse CD25 (PC61), FITC anti-mouse CD45R/B220 (RA3-6B2), PE anti-mouse CD80 (B7-1) (16-10A1), PE anti-mouse CD86 (B7-2) (GL1), PE ratanti-mouse interleukin-4 (IL-4) (BVD4-1D11), FITC anti-mouse gamma inter-feron (IFN-�) (XMG1.2), and PE anti-mouse IL-10 (JES5-2A5) (BD Pharmin-gen, San Diego, CA). Biotin-conjugated MAbs were revealed with streptavidin-PE-Cy5 (BD Pharmingen). Cells were preincubated for 15 min with anti-Fc�R (akind gift of Jocelyne Demengeot, Gulbenkian Institute of Science, Oeiras, Por-tugal) before CD11c and Foxp3 staining. The Foxp3 Staining Buffer set (eBio-science) was used for the fixation and permeabilization of splenocytes previouslysurface stained with CD4 and CD25 MAbs.
The intracellular expression of the cytokines IFN-�, IL-4, and IL-10 wasdetected in splenic CD4� T lymphocytes. The intracellular expression of thecytokines IFN-� and IL-4 was also detected in renal CD4� T lymphocytes.Splenocytes were obtained as described above. Red blood cell lysis was per-formed by incubation with 0.15 M ammonium chloride. Cells were washed andresuspended in complete RPMI medium (Sigma) (RPMI 1640 medium supple-mented with 50 U of penicillin/ml, 50 �g of streptomycin/ml, 1% HEPES buffer[Sigma], 10% fetal calf serum [FCS] [Invitrogen], and 5 �M 2-mercaptoethanol).The kidneys were minced with a razor blade and incubated for 30 min at 37°C inRPMI 1640 complete medium containing collagenase D (Sigma-Aldrich) at 2�g/ml. Cells were homogenized to single-cell suspensions, washed, and resus-pended in RPMI 1640 complete medium. Mononuclear cells were separatedfrom the above-described suspensions by layering 5 ml onto 2.5 ml of a polysu-crose-sodium ditrizoate solution (Histopaque 1083; Sigma) and centrifuged at800 � g for 20 min at room temperature. Mononuclear cells collected from themedium-Histopaque interface were washed and resuspended in RPMI 1640complete medium. Spleen and renal cells (1 � 106 cells) were transferred into96-well tissue culture plates (Nunc, Roskilde, Denmark) and stimulated for 4.5 hwith 20 ng/ml phorbol myristate acetate (Sigma) and 200 ng/ml ionomycin(Sigma) in the presence of 10 �g/ml of brefeldin A (Roche, Penzberg, Germany).
The staining of the cell surface marker CD4 was performed as describedabove, after a preincubation step of 15 min with anti-Fc�R, followed by fixationwith 2% formaldehyde. Cells were permeabilized with 0.5% saponin in flowcytometric buffer (PBS containing 1% bovine serum albumin [BSA] and 10 mMsodium azide), and subsequently, cells were incubated for 15 min with anti-Fc�Rand stained for 30 min at room temperature with the appropriate antibody.Intracellular staining with the isotypic controls was performed to confirm thespecificity of antibody binding.
Th1 cells were defined as being CD4� IFN-�� IL-4�, and Th2 cells weredefined as being CD4�IFN-�� IL-4�. Ratios of Th1/Th2 cells were generated todetermine the presence of a polarized immune response. The immune responsein noninfected mice was defined as being unpolarized.
Serum IFN-�, IL-4, and IL-10 measurements. The concentration of IL-4 in thesera of C. albicans-infected mice and noninfected controls was quantified withthe Quantikine M murine IL-4 enzyme-linked immunosorbent assay (ELISA)kit, and serum IFN-� and IL-10 concentrations were quantified with Duo-SetELISA kits (all from R&D Systems, Minneapolis, MN) according to the manu-facturer’s instructions.
Statistical analysis. Unless otherwise stated, results shown are from one ex-periment representative of three independent experiments. The statistical sig-
nificance of results was determined by the unpaired Student t test, and survivaldata were analyzed with the log-rank test by using GraphPad Prism 4 software(GraphPad Software, Inc., La Jolla, CA). Results were considered statisticallysignificant with P values of less than 0.05.
RESULTS
Virulence of C. albicans in a murine model of hematog-enously disseminated infection. Prior to the virulence studies,we determined the generation time for each strain in syntheticdefined and complex media at 30° and 37°C and the ability toform hyphae in serum- or N-acetylglucosamine-containing me-dia. No yeast growth defects were observed under the condi-tions tested (similar generation times), and filamentous growthwas similar to that of wild-type (WT) strain SC5314 under thehypha-inducing conditions analyzed (data not shown).
To explore the role of the aspartyl proteinases Sap1 to Sap6as virulence factors in the course of hematogenously dissemi-nated candidiasis, BALB/c mice were initially infected i.v. with5 � 105 and 1 � 105 cells of C. albicans WT strain SC5314and two independent series of homozygous-deletion triple-mutant strains lacking the SAP1 to SAP3 genes (SAP123MS4Cand SAP123MS4D) or lacking the SAP4 to SAP6 genes(SAP456MS4A and SAP456MS4B) (29). The independentmutant strains behaved indistinctively (data not shown), andtherefore, the results presented below correspond to thoseobtained with strains SAP123MS4C and SAP456MS4A.
Mice infected with the highest inoculum showed 100% mor-tality for every group by day 6 after infection, and no differ-ences could be observed (data not shown).
No differences in survival time were observed after i.v. in-fection of BALB/c mice with 1 � 105 CFU of the WT or�sap123 mutant strain SAP123MS4C (P � 0.5698 by log-ranktest) (Fig. 1A). Mice infected with �sap456 mutant strainSAP456MS4A had an extended overall survival time comparedto that of their WT-infected counterparts (Fig. 1). The mediansurvival times of SAP456MS4A-infected mice were 13 and 16versus 10 and 12 days for WT-infected mice, as determined forthe independent experiments shown in Fig. 1A and B, respec-tively. All mice injected with either the WT or the triple-mutant strain SAP123MS4C succumbed to candidal infectionwithin 17 days. In contrast, at the end of the experimentalperiod (30 days), 15 to 20% of the mice infected with the�sap456 mutant survived infection. Nevertheless, the kidneysof the surviving mice had Candida microabscesses and gran-
TABLE 2. SAP and ACT1 primers and expected fragment lengths
GenePrimer (5�–3�) Amplicon
length (bp)Forward Reverse
ACT1 TGCTGAACGTATGCAAAAGG TGAACAATGGATGGACCAGA 186SAP1 TGAGGCTGCTGGTGATTATG TGCCAACAGCTTTGAGAGAA 224SAP2 ATCAGCTGGTTTCGTTGCTT GGGACAGCTTGTCTTTTGGA 105SAP3 TGTTACTGGTCCCCAAGGTGAA CTTGTCCTTGACCAGCTTGACAT 209SAP4 AATGATGTGGGCAAAAGAGG ACGGCATTTGAATCTGGAAC 155SAP5 ATTTCCCGTCGATGAGACTG ACCACGCCATTTTGGAATAC 205SAP6 GTCAACGCTGGTGTCCTCTT GCAGGAACGGAGATCTTGAG 197SAP7 TTCTCGTGATGCTGTCCAAG CCAGCAGGAAGACCATAAGC 183SAP8 TTTGGTGGGGTTGATAATGC GGCAGCAGCCAATTTATCAG 198SAP9 ACCGGGTCTTCAGATTTGTG TTCCTCGTCGGTTTCTATGG 180SAP10 AACGGAAATGTTGCTTCTGG TGAATCGCCTATCGAAAACC 192
VOL. 78, 2010 ROLE OF Saps IN MURINE DISSEMINATED CANDIDIASIS 4841
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
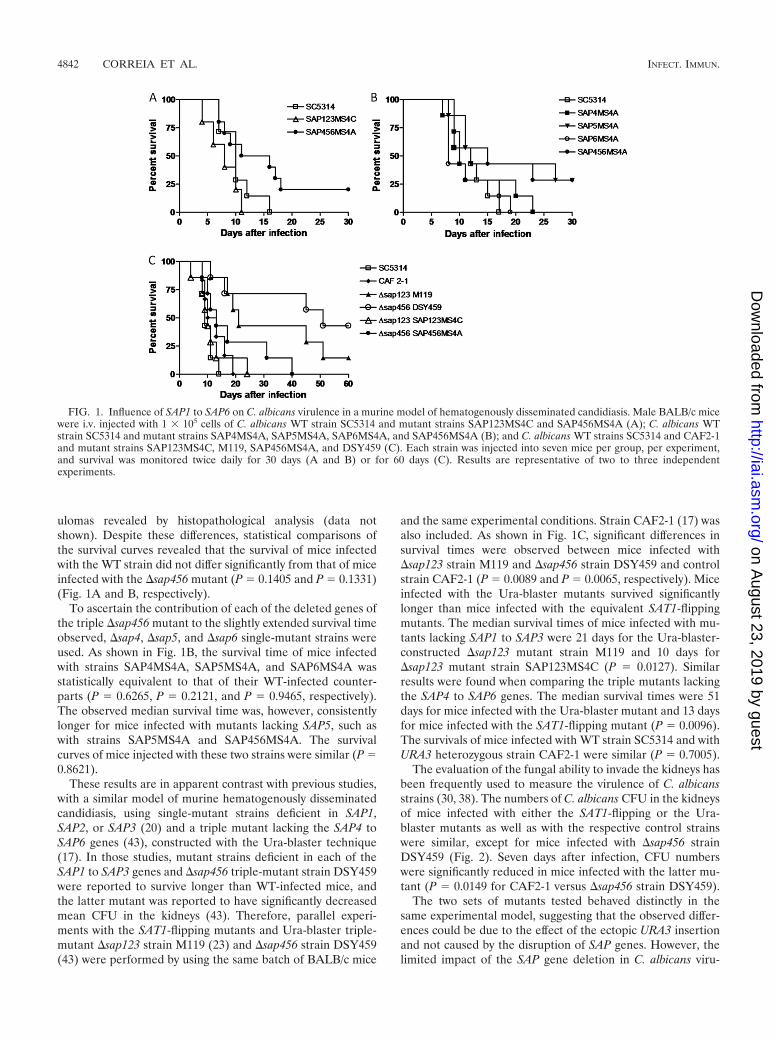
ulomas revealed by histopathological analysis (data notshown). Despite these differences, statistical comparisons ofthe survival curves revealed that the survival of mice infectedwith the WT strain did not differ significantly from that of miceinfected with the �sap456 mutant (P � 0.1405 and P � 0.1331)(Fig. 1A and B, respectively).
To ascertain the contribution of each of the deleted genes ofthe triple �sap456 mutant to the slightly extended survival timeobserved, �sap4, �sap5, and �sap6 single-mutant strains wereused. As shown in Fig. 1B, the survival time of mice infectedwith strains SAP4MS4A, SAP5MS4A, and SAP6MS4A wasstatistically equivalent to that of their WT-infected counter-parts (P � 0.6265, P � 0.2121, and P � 0.9465, respectively).The observed median survival time was, however, consistentlylonger for mice infected with mutants lacking SAP5, such aswith strains SAP5MS4A and SAP456MS4A. The survivalcurves of mice injected with these two strains were similar (P �0.8621).
These results are in apparent contrast with previous studies,with a similar model of murine hematogenously disseminatedcandidiasis, using single-mutant strains deficient in SAP1,SAP2, or SAP3 (20) and a triple mutant lacking the SAP4 toSAP6 genes (43), constructed with the Ura-blaster technique(17). In those studies, mutant strains deficient in each of theSAP1 to SAP3 genes and �sap456 triple-mutant strain DSY459were reported to survive longer than WT-infected mice, andthe latter mutant was reported to have significantly decreasedmean CFU in the kidneys (43). Therefore, parallel experi-ments with the SAT1-flipping mutants and Ura-blaster triple-mutant �sap123 strain M119 (23) and �sap456 strain DSY459(43) were performed by using the same batch of BALB/c mice
and the same experimental conditions. Strain CAF2-1 (17) wasalso included. As shown in Fig. 1C, significant differences insurvival times were observed between mice infected with�sap123 strain M119 and �sap456 strain DSY459 and controlstrain CAF2-1 (P � 0.0089 and P � 0.0065, respectively). Miceinfected with the Ura-blaster mutants survived significantlylonger than mice infected with the equivalent SAT1-flippingmutants. The median survival times of mice infected with mu-tants lacking SAP1 to SAP3 were 21 days for the Ura-blaster-constructed �sap123 mutant strain M119 and 10 days for�sap123 mutant strain SAP123MS4C (P � 0.0127). Similarresults were found when comparing the triple mutants lackingthe SAP4 to SAP6 genes. The median survival times were 51days for mice infected with the Ura-blaster mutant and 13 daysfor mice infected with the SAT1-flipping mutant (P � 0.0096).The survivals of mice infected with WT strain SC5314 and withURA3 heterozygous strain CAF2-1 were similar (P � 0.7005).
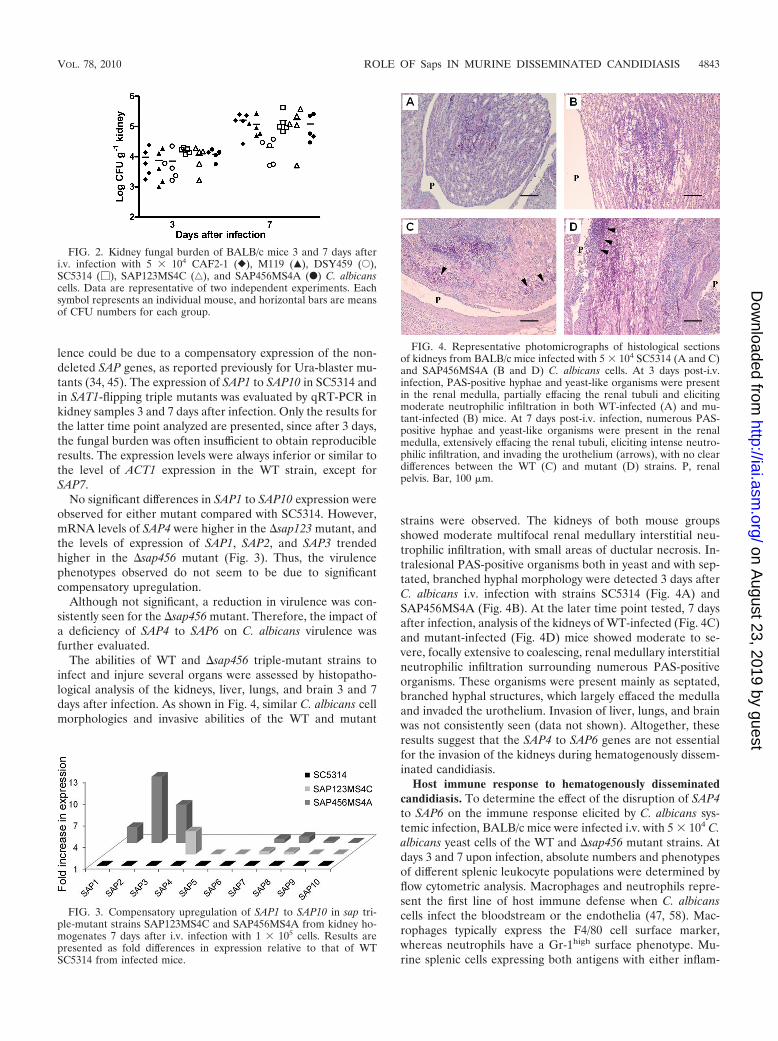
The evaluation of the fungal ability to invade the kidneys hasbeen frequently used to measure the virulence of C. albicansstrains (30, 38). The numbers of C. albicans CFU in the kidneysof mice infected with either the SAT1-flipping or the Ura-blaster mutants as well as with the respective control strainswere similar, except for mice infected with �sap456 strainDSY459 (Fig. 2). Seven days after infection, CFU numberswere significantly reduced in mice infected with the latter mu-tant (P � 0.0149 for CAF2-1 versus �sap456 strain DSY459).
The two sets of mutants tested behaved distinctly in thesame experimental model, suggesting that the observed differ-ences could be due to the effect of the ectopic URA3 insertionand not caused by the disruption of SAP genes. However, thelimited impact of the SAP gene deletion in C. albicans viru-
FIG. 1. Influence of SAP1 to SAP6 on C. albicans virulence in a murine model of hematogenously disseminated candidiasis. Male BALB/c micewere i.v. injected with 1 � 105 cells of C. albicans WT strain SC5314 and mutant strains SAP123MS4C and SAP456MS4A (A); C. albicans WTstrain SC5314 and mutant strains SAP4MS4A, SAP5MS4A, SAP6MS4A, and SAP456MS4A (B); and C. albicans WT strains SC5314 and CAF2-1and mutant strains SAP123MS4C, M119, SAP456MS4A, and DSY459 (C). Each strain was injected into seven mice per group, per experiment,and survival was monitored twice daily for 30 days (A and B) or for 60 days (C). Results are representative of two to three independentexperiments.
4842 CORREIA ET AL. INFECT. IMMUN.
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
lence could be due to a compensatory expression of the non-deleted SAP genes, as reported previously for Ura-blaster mu-tants (34, 45). The expression of SAP1 to SAP10 in SC5314 andin SAT1-flipping triple mutants was evaluated by qRT-PCR inkidney samples 3 and 7 days after infection. Only the results forthe latter time point analyzed are presented, since after 3 days,the fungal burden was often insufficient to obtain reproducibleresults. The expression levels were always inferior or similar tothe level of ACT1 expression in the WT strain, except forSAP7.
No significant differences in SAP1 to SAP10 expression wereobserved for either mutant compared with SC5314. However,mRNA levels of SAP4 were higher in the �sap123 mutant, andthe levels of expression of SAP1, SAP2, and SAP3 trendedhigher in the �sap456 mutant (Fig. 3). Thus, the virulencephenotypes observed do not seem to be due to significantcompensatory upregulation.
Although not significant, a reduction in virulence was con-sistently seen for the �sap456 mutant. Therefore, the impact ofa deficiency of SAP4 to SAP6 on C. albicans virulence wasfurther evaluated.
The abilities of WT and �sap456 triple-mutant strains toinfect and injure several organs were assessed by histopatho-logical analysis of the kidneys, liver, lungs, and brain 3 and 7days after infection. As shown in Fig. 4, similar C. albicans cellmorphologies and invasive abilities of the WT and mutant
strains were observed. The kidneys of both mouse groupsshowed moderate multifocal renal medullary interstitial neu-trophilic infiltration, with small areas of ductular necrosis. In-tralesional PAS-positive organisms both in yeast and with sep-tated, branched hyphal morphology were detected 3 days afterC. albicans i.v. infection with strains SC5314 (Fig. 4A) andSAP456MS4A (Fig. 4B). At the later time point tested, 7 daysafter infection, analysis of the kidneys of WT-infected (Fig. 4C)and mutant-infected (Fig. 4D) mice showed moderate to se-vere, focally extensive to coalescing, renal medullary interstitialneutrophilic infiltration surrounding numerous PAS-positiveorganisms. These organisms were present mainly as septated,branched hyphal structures, which largely effaced the medullaand invaded the urothelium. Invasion of liver, lungs, and brainwas not consistently seen (data not shown). Altogether, theseresults suggest that the SAP4 to SAP6 genes are not essentialfor the invasion of the kidneys during hematogenously dissem-inated candidiasis.
Host immune response to hematogenously disseminatedcandidiasis. To determine the effect of the disruption of SAP4to SAP6 on the immune response elicited by C. albicans sys-temic infection, BALB/c mice were infected i.v. with 5 � 104 C.albicans yeast cells of the WT and �sap456 mutant strains. Atdays 3 and 7 upon infection, absolute numbers and phenotypesof different splenic leukocyte populations were determined byflow cytometric analysis. Macrophages and neutrophils repre-sent the first line of host immune defense when C. albicanscells infect the bloodstream or the endothelia (47, 58). Mac-rophages typically express the F4/80 cell surface marker,whereas neutrophils have a Gr-1high surface phenotype. Mu-rine splenic cells expressing both antigens with either inflam-
FIG. 2. Kidney fungal burden of BALB/c mice 3 and 7 days afteri.v. infection with 5 � 104 CAF2-1 (}), M119 (Œ), DSY459 (E),SC5314 (�), SAP123MS4C (‚), and SAP456MS4A (F) C. albicanscells. Data are representative of two independent experiments. Eachsymbol represents an individual mouse, and horizontal bars are meansof CFU numbers for each group.
FIG. 3. Compensatory upregulation of SAP1 to SAP10 in sap tri-ple-mutant strains SAP123MS4C and SAP456MS4A from kidney ho-mogenates 7 days after i.v. infection with 1 � 105 cells. Results arepresented as fold differences in expression relative to that of WTSC5314 from infected mice.
FIG. 4. Representative photomicrographs of histological sectionsof kidneys from BALB/c mice infected with 5 � 104 SC5314 (A and C)and SAP456MS4A (B and D) C. albicans cells. At 3 days post-i.v.infection, PAS-positive hyphae and yeast-like organisms were presentin the renal medulla, partially effacing the renal tubuli and elicitingmoderate neutrophilic infiltration in both WT-infected (A) and mu-tant-infected (B) mice. At 7 days post-i.v. infection, numerous PAS-positive hyphae and yeast-like organisms were present in the renalmedulla, extensively effacing the renal tubuli, eliciting intense neutro-philic infiltration, and invading the urothelium (arrows), with no cleardifferences between the WT (C) and mutant (D) strains. P, renalpelvis. Bar, 100 �m.
VOL. 78, 2010 ROLE OF Saps IN MURINE DISSEMINATED CANDIDIASIS 4843
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
matory or immunosuppressive function have also been de-scribed in the context of C. albicans infections (31, 56).According to the expression of these two surface markers,three cell populations were analyzed in this study: F4/80high
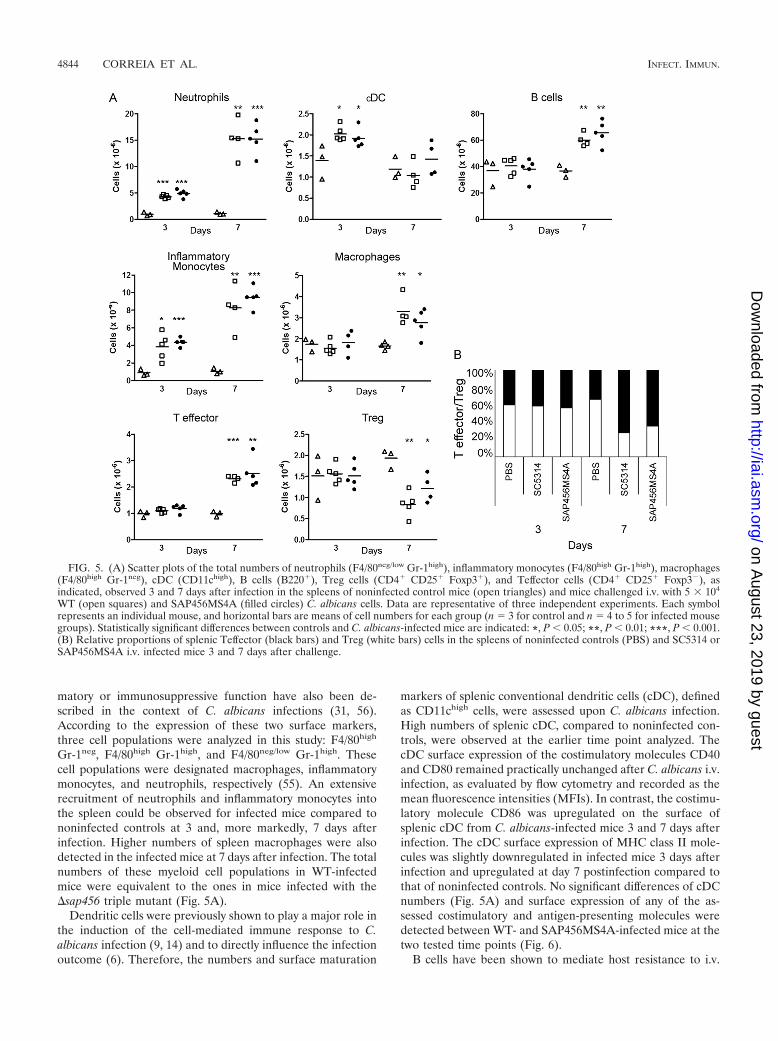
Gr-1neg, F4/80high Gr-1high, and F4/80neg/low Gr-1high. Thesecell populations were designated macrophages, inflammatorymonocytes, and neutrophils, respectively (55). An extensiverecruitment of neutrophils and inflammatory monocytes intothe spleen could be observed for infected mice compared tononinfected controls at 3 and, more markedly, 7 days afterinfection. Higher numbers of spleen macrophages were alsodetected in the infected mice at 7 days after infection. The totalnumbers of these myeloid cell populations in WT-infectedmice were equivalent to the ones in mice infected with the�sap456 triple mutant (Fig. 5A).
Dendritic cells were previously shown to play a major role inthe induction of the cell-mediated immune response to C.albicans infection (9, 14) and to directly influence the infectionoutcome (6). Therefore, the numbers and surface maturation
markers of splenic conventional dendritic cells (cDC), definedas CD11chigh cells, were assessed upon C. albicans infection.High numbers of splenic cDC, compared to noninfected con-trols, were observed at the earlier time point analyzed. ThecDC surface expression of the costimulatory molecules CD40and CD80 remained practically unchanged after C. albicans i.v.infection, as evaluated by flow cytometry and recorded as themean fluorescence intensities (MFIs). In contrast, the costimu-latory molecule CD86 was upregulated on the surface ofsplenic cDC from C. albicans-infected mice 3 and 7 days afterinfection. The cDC surface expression of MHC class II mole-cules was slightly downregulated in infected mice 3 days afterinfection and upregulated at day 7 postinfection compared tothat of noninfected controls. No significant differences of cDCnumbers (Fig. 5A) and surface expression of any of the as-sessed costimulatory and antigen-presenting molecules weredetected between WT- and SAP456MS4A-infected mice at thetwo tested time points (Fig. 6).
B cells have been shown to mediate host resistance to i.v.
FIG. 5. (A) Scatter plots of the total numbers of neutrophils (F4/80neg/low Gr-1high), inflammatory monocytes (F4/80high Gr-1high), macrophages(F4/80high Gr-1neg), cDC (CD11chigh), B cells (B220�), Treg cells (CD4� CD25� Foxp3�), and Teffector cells (CD4� CD25� Foxp3�), asindicated, observed 3 and 7 days after infection in the spleens of noninfected control mice (open triangles) and mice challenged i.v. with 5 � 104
WT (open squares) and SAP456MS4A (filled circles) C. albicans cells. Data are representative of three independent experiments. Each symbolrepresents an individual mouse, and horizontal bars are means of cell numbers for each group (n � 3 for control and n � 4 to 5 for infected mousegroups). Statistically significant differences between controls and C. albicans-infected mice are indicated: *, P 0.05; **, P 0.01; ***, P 0.001.(B) Relative proportions of splenic Teffector (black bars) and Treg (white bars) cells in the spleens of noninfected controls (PBS) and SC5314 orSAP456MS4A i.v. infected mice 3 and 7 days after challenge.
4844 CORREIA ET AL. INFECT. IMMUN.
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
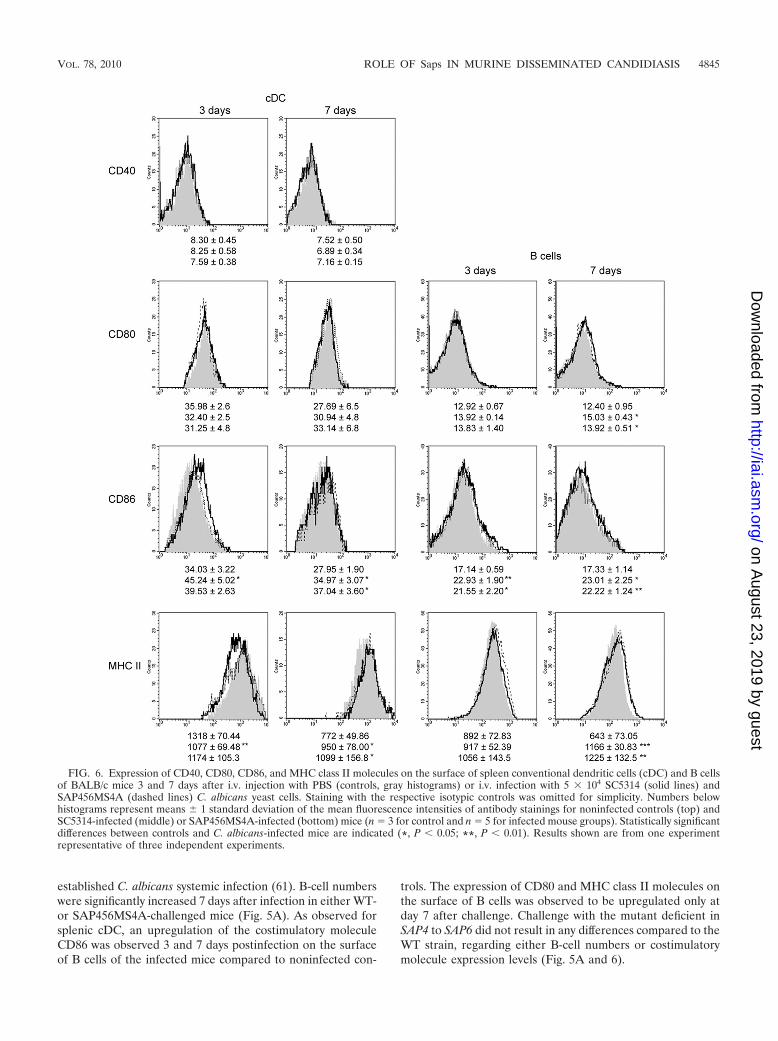
established C. albicans systemic infection (61). B-cell numberswere significantly increased 7 days after infection in either WT-or SAP456MS4A-challenged mice (Fig. 5A). As observed forsplenic cDC, an upregulation of the costimulatory moleculeCD86 was observed 3 and 7 days postinfection on the surfaceof B cells of the infected mice compared to noninfected con-
trols. The expression of CD80 and MHC class II molecules onthe surface of B cells was observed to be upregulated only atday 7 after challenge. Challenge with the mutant deficient inSAP4 to SAP6 did not result in any differences compared to theWT strain, regarding either B-cell numbers or costimulatorymolecule expression levels (Fig. 5A and 6).
FIG. 6. Expression of CD40, CD80, CD86, and MHC class II molecules on the surface of spleen conventional dendritic cells (cDC) and B cellsof BALB/c mice 3 and 7 days after i.v. injection with PBS (controls, gray histograms) or i.v. infection with 5 � 104 SC5314 (solid lines) andSAP456MS4A (dashed lines) C. albicans yeast cells. Staining with the respective isotypic controls was omitted for simplicity. Numbers belowhistograms represent means 1 standard deviation of the mean fluorescence intensities of antibody stainings for noninfected controls (top) andSC5314-infected (middle) or SAP456MS4A-infected (bottom) mice (n � 3 for control and n � 5 for infected mouse groups). Statistically significantdifferences between controls and C. albicans-infected mice are indicated (*, P 0.05; **, P 0.01). Results shown are from one experimentrepresentative of three independent experiments.
VOL. 78, 2010 ROLE OF Saps IN MURINE DISSEMINATED CANDIDIASIS 4845
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
Although CD4� T cells have been reported to have littleinfluence on survival and on fungal burden during acute sys-temic candidiasis (3, 24), the CD4� T-cell subset of naturallyoccurring regulatory T (Treg) cells was shown to promote hostsusceptibility to C. albicans (36, 53) and to limit tissue damageand/or enhance healing but not to directly augment the clear-ance of the organism from infected tissues (32). To ascertainthe impact of a deficiency of SAP4 to SAP6 on the immuneresponse mediated by CD4� T-lymphocyte cell populations,the numbers of CD4�, CD4� CD25�, and CD4� CD25�
Foxp3� (Treg) cells and also of CD4� CD25� Foxp3� (Tef-fector) cells in the spleens of the infected BALB/c mice wereassessed. The numbers of splenic CD4� and CD4� CD25� Tcells of infected mice were not significantly different fromthose of noninfected controls (data not shown). However, asassessed by Foxp3 expression within the CD4� CD25� T-cellpopulation, reduced percentages and numbers of Treg cellswere observed 7 days after C. albicans infection. This corre-lated with higher splenic percentages and numbers of Teffector
cells in these mice (Fig. 5A), resulting in higher Teffector/Tregratios than those for noninfected controls (Fig. 5B). The per-centages and numbers of both Teffector and Treg cells werenot significantly different between the two C. albicans-infectedgroups.
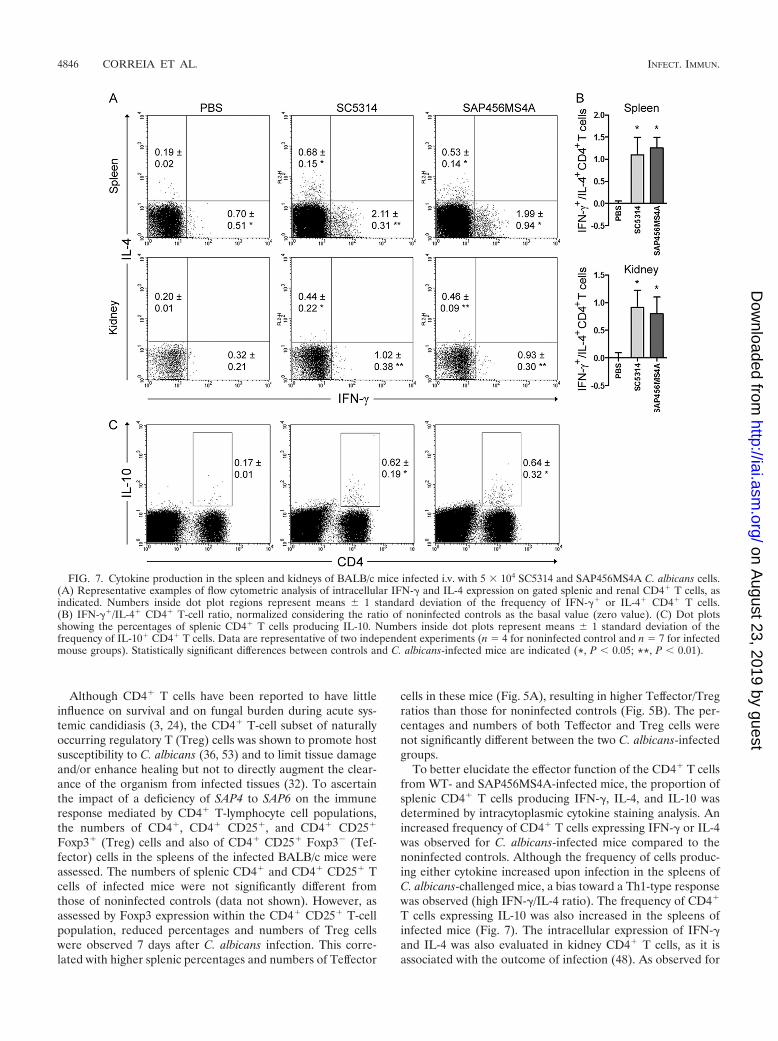
To better elucidate the effector function of the CD4� T cellsfrom WT- and SAP456MS4A-infected mice, the proportion ofsplenic CD4� T cells producing IFN-�, IL-4, and IL-10 wasdetermined by intracytoplasmic cytokine staining analysis. Anincreased frequency of CD4� T cells expressing IFN-� or IL-4was observed for C. albicans-infected mice compared to thenoninfected controls. Although the frequency of cells produc-ing either cytokine increased upon infection in the spleens ofC. albicans-challenged mice, a bias toward a Th1-type responsewas observed (high IFN-�/IL-4 ratio). The frequency of CD4�
T cells expressing IL-10 was also increased in the spleens ofinfected mice (Fig. 7). The intracellular expression of IFN-�and IL-4 was also evaluated in kidney CD4� T cells, as it isassociated with the outcome of infection (48). As observed for
FIG. 7. Cytokine production in the spleen and kidneys of BALB/c mice infected i.v. with 5 � 104 SC5314 and SAP456MS4A C. albicans cells.(A) Representative examples of flow cytometric analysis of intracellular IFN-� and IL-4 expression on gated splenic and renal CD4� T cells, asindicated. Numbers inside dot plot regions represent means 1 standard deviation of the frequency of IFN-�� or IL-4� CD4� T cells.(B) IFN-��/IL-4� CD4� T-cell ratio, normalized considering the ratio of noninfected controls as the basal value (zero value). (C) Dot plotsshowing the percentages of splenic CD4� T cells producing IL-10. Numbers inside dot plots represent means 1 standard deviation of thefrequency of IL-10� CD4� T cells. Data are representative of two independent experiments (n � 4 for noninfected control and n � 7 for infectedmouse groups). Statistically significant differences between controls and C. albicans-infected mice are indicated (*, P 0.05; **, P 0.01).
4846 CORREIA ET AL. INFECT. IMMUN.
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

the spleen, the frequency of CD4� T cells expressing the cy-tokines IFN-� and IL-4 increased in infected mice. Although atrend toward extended survival was observed for mice infectedwith mutant strain SAP456MS4A, the percentages of CD4� Tcells producing the cytokine IFN-� or IL-4 in the kidneys ofeither group of infected mice were similar, resulting in equiv-alent Th1/Th2 cell ratios (Fig. 7). No serum IFN-�, IL-4, andIL-10 was detected by ELISA 3 and 7 days after infection ofeither infected or uninfected mice (data not shown).
Overall, these results indicate that a deficiency of SAP4 toSAP6 does not have a significant impact on the immune re-sponse elicited in the spleen and kidneys of BALB/c micehematogenously challenged with C. albicans.
DISCUSSION
The secretion of aspartyl proteinases has long been recog-nized as a virulence-associated trait of Candida albicans (10,27, 49). The importance of specific Sap isoenzymes for thepathogenicity of this fungus has been investigated with differ-ent infection models by comparing the virulence of mutantsdeficient in individual or multiple SAP genes with that of a WTcontrol strain. In this study, the importance of SAP1 to SAP6gene expression for C. albicans virulence was evaluated byusing sap-null mutants derived from WT strain SC5314. Thevirulence of mutant strains lacking SAP1 to SAP3 was indis-tinguishable from that of WT strain SC5314, while the deletionof SAP4 to SAP6 caused a slight attenuation in virulence.Previous reports have shown that a deficiency in SAP4 to SAP6attenuated virulence to a higher extent than did a deficiency inSAP1, SAP2, or SAP3 (20, 43). Here, an increased mediansurvival time was consistently observed for mice infected withmutants lacking the SAP5 gene, such as the single-deletionmutant strain SAP5MS4A or the triple-deletion mutant strainSAP456MS4A, compared to that of animals infected with WTstrain SC5314. However, the differences found were small andnot always significant. Moreover, histopathology analysis didnot indicate a reduced ability of the SAP456MS4A mutant toinvade the kidneys, although sap-null mutant strains lackingthe SAP6 gene were previously shown to have reduced inva-siveness in a model of experimental peritonitis (16). The de-letion of SAP4 to SAP6 did not result in clear differences inhypha formation, and similar morphotypes were observed,both in vitro and in vivo, for WT and mutant strains. This is notunexpected, as SAP4 to SAP6 expression is associated with, butnot required for, hyphal morphology (16). Additionally, ex-pression levels of SAP4 to SAP6 may not be directly linked toorgan invasion, since a previously reported C. albicans strainexpressing high levels of SAP4 to SAP6 was noninvasive (57).
The results obtained with SAT1-flipping mutants contrastwith those obtained when using the Ura-blaster sap-null mu-tants, which survived much longer. When another parameterassociated with C. albicans virulence was analyzed, such askidneys CFU, no differences were observed among mousegroups, except for the ones infected with strain DSY459, whichpresented a lower fungal burden. Discrepancies between dif-ferent methods of evaluating virulence have already been re-ported for mice intravenously infected with C. albicans mutantstrains, including sap-null mutants, where differences in mousesurvival were not associated with differences in organ fungal
burden (20, 54, 62). The differences found between the two setsof mutants are most likely due to the ectopic insertion ofURA3, which must have contributed to the reduced virulenceof the Ura-blaster-constructed mutant strains. It is widelyknown that the Ura status of C. albicans strains influencesadherence (5) and virulence (28, 52). Although this can beovercome by the integration of URA3 at the ENO1 (52) orRPS10 (8) locus, the strains used in this study and in previouslyreported studies (20, 43) did not share a site of URA3 integra-tion.
The disruption of SAP1 to SAP3 and SAP4 to SAP6 led to anincreased level of expression of SAP4 and SAP1 to SAP3,respectively, suggesting that C. albicans attempts to compen-sate the functional loss of these subfamilies by upregulatingalternative SAP genes during hematogenously disseminatedcandidiasis. Therefore, the compensatory upregulation ob-served could be, to some extent, contributing to the lack of aphenotype seen for these mutants. However, the equivalentUra-blaster mutants, despite the compensatory upregulationreported previously (34, 45), showed markedly reduced viru-lence in this model.
Recently, Lermann and Morschhauser (29) and Naglik et al.(34) reevaluated the role of SAP1 to SAP6 in a model ofreconstituted human epithelia (RHE) and reported that SAP1to SAP6 were not essential for successful C. albicans RHEinfection, in contrast to previous reports (45, 46). The presentstudy thus reports an additional model in which the SAP genesubfamilies SAP1 to SAP3 and SAP4 to SAP6 seem to havelittle influence on the outcome of infection.
As mice infected with the �sap456 triple mutant displayed aslightly extended survival time, it could be expected that itmight result from a more effective host immune response. Thiswould be in agreement with a previous report suggesting animmunomodulatory role of Sap4 to Sap6 upon macrophagephagocytosis (7). However, the analysis of diverse features ofthe innate and acquired immune response elicited inBALB/c mice upon infection with either the WT or the�sap456 triple mutant did not show any significant differ-ences between these two yeast strains. The similar abilitiesof both strains to recruit inflammatory cells are in accor-dance with their similar observed virulences, taking intoaccount the prominent role of innate immunity and of neu-trophils in particular, in host protection against dissemi-nated candidiasis (2, 60).
The proportions of splenic Teffector and Treg cells in thespleens of mice infected with either the WT or the �sap456mutant were highly similar. Likewise, the frequencies of CD4�
T cells expressing IFN-�, IL-4, and IL-10, cytokines previouslyshown to be relevant for resistance or susceptibility to systemiccandidiasis (48), were similar in the two infected mousegroups. In vivo models indicate that regulatory T cells attenu-ate Th1-type antifungal responses and induce tolerance to thefungus (32, 36). As higher IFN-�/IL-4 ratios were observed forsplenic and renal CD4� T cells of infected mice than those ofnoninfected controls, it can be assumed that even though thekidneys of infected mice presented a high fungal burden 7 daysafter challenge, a protective Th1-type response of an equiva-lent magnitude might be occurring in both WT- and �sap456mutant-infected mice.
Although our results suggest that B cells may have a role in
VOL. 78, 2010 ROLE OF Saps IN MURINE DISSEMINATED CANDIDIASIS 4847
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

the activation of T cells during experimental disseminated can-didiasis, in accordance with the increased susceptibility ob-served for B-cell-deficient mice (61), they also indicate that thedeficiency of SAP4 to SAP6 does not affect such a role of Bcells.
Differences in C. albicans morphology have been frequentlyshown to influence both the type and magnitude of the hostimmune response in the course of candidiasis. Dendritic cells,and also neutrophils, modulate adaptive responses to the fun-gus, depending on the Candida morphotype encountered (14,42, 47). As indistinguishable morphotypes were found for theWT and the �sap456 mutant strain both in vitro and in vivo,this is also in agreement with the lack of significant differencesobserved in the immune responses elicited by these strains.
A relative independence on aspartyl proteinase activity forthe establishment of hematogenously disseminated candidiasiswas previously reported (15), as treatment with pepstatin A, apotent protease inhibitor, did not protect mice against intra-venous infection with C. albicans. As previously suggested, anexplanation for these observations may be the requirement forSap only where anatomical barriers had to be crossed prior todissemination (15, 26). When C. albicans cells are delivereddirectly into the bloodstream, low-molecular-weight peptidesare available, and yeast growth may be protease independent.
The relative importance of specific SAP genes for C. albicanspathogenicity is greatly determined by the type of infection andits dependence on protease activity for the successful invasionand colonization of various host niches. Treatment with pep-statin A resulted in reduced virulence in intranasal (15) andintraperitoneal (26) models but had no protective effect in theintravenous model. The subfamily of the SAP genes SAP1 toSAP3, and SAP2 in particular, was proven to be important in amodel of rat vaginal infection, while SAP4 to SAP6 had littleimpact on this infection model (12). In contrast, only �sap456mutants, and �sap6 in particular, showed reduced virulence ina murine model of Candida peritonitis and keratitis, while thedeletion of the SAP1, SAP2, or SAP3 gene had no significanteffect on these infection models (16, 23). Moreover, immuno-logical neutralization of Sap2 was shown to have a protectiveeffect on C. albicans-infected hosts during vaginal and oralinfection (12, 40) and also in experimental peritonitis (59).
Although individual processes resulting from the action of asingle gene or a small group of genes may be important inspecific stages of infection, cooperative gene functions are es-sential for the multiple processes of C. albicans infection (38).Thus, although the proteinase family as a whole may contrib-ute to C. albicans virulence in the course of acute systemiccandidiasis, other factors must be the major contributors toinvasion and cell damage in this model.
ACKNOWLEDGMENTS
We are indebted to Bernhard Hube from Hans Knoell Institute,Jena, Germany, for providing C. albicans CAF2-1 and the Ura-blastersap mutant strains.
This work was supported by Fundacao para a Ciencia e Tecnologiagrant POCI/SAU-IMI/58014/2004 and FEDER. Alexandra Correiaand Filipe Cerca were supported by FCT fellowships SFRH/BD/31354/2006 and SFRH/BD/27638/2006, respectively. Luzia Teixeira was sup-ported by FSE and MCTES through POPH-QREN-Tipologia 4.2.
REFERENCES
1. Albrecht, A., A. Felk, I. Pichova, J. R. Naglik, M. Schaller, P. de Groot, D.MacCallum, F. C. Odds, W. Schafer, F. Klis, M. Monod, and B. Hube. 2006.Glycosylphosphatidylinositol-anchored proteases of Candida albicans targetproteins necessary for both cellular processes and host-pathogen interac-tions. J. Biol. Chem. 281:688–694.
2. Ashman, R. B. 2008. Protective and pathologic immune responses againstCandida albicans infection. Front. Biosci. 13:3334–3351.
3. Ashman, R., C. S. Farah, S. Wanasaengsakul, Y. Hu, G. Pang, and R. L.Clancy. 2004. Innate versus adaptive immunity in Candida albicans infection.Immunol. Cell Biol. 82:196–204.
4. Ausubel, F. M. (ed.). 1993. Current protocols in molecular biology, 2nd ed.Wiley, New York, NY.
5. Bain, J. M., C. Stubberfield, and N. A. Gow. 2001. Ura-status-dependentadhesion of Candida albicans mutants. FEMS Microbiol. Lett. 204:323–328.
6. Bonifazi, P., T. Zelante, C. D’Angelo, A. De Luca, S. Moretti, S. Bozza, K.Perruccio, R. G. Iannitti, G. Giovannini, C. Volpi, F. Fallarino, P. Puccetti,and L. Romani. 2009. Balancing inflammation and tolerance in vivo throughdendritic cells by the commensal Candida albicans. Mucosal Immunol.2:362–374.
7. Borg-von Zepelin, M., S. Beggah, K. Boggian, D. Sanglard, and M. Monod.1998. The expression of the secreted aspartyl proteinases Sap4 to Sap6 fromCandida albicans in murine macrophages. Mol. Microbiol. 28:543–554.
8. Brand, A., D. M. MacCallum, A. J. P. Brown, N. A. Gow, and F. C. Odds.2004. Ectopic expression of URA3 can influence the virulence phenotypesand proteome of Candida albicans but can be overcome by targeted reinte-gration of URA3 at the RPS10 locus. Eukaryot. Cell 3:900–909.
9. Buentke, E., and A. Sheynius. 2003. Dendritic cells and fungi. APMIS 111:789–796.
10. Cassone, A., F. de Bernardis, F. Mondello, T. Ceddia, and L. Agatensi. 1987.Evidence for a correlation between proteinase secretion and vulvovaginalcandidosis. J. Infect. Dis. 156:777–783.
11. Cheng, S., M. H. Nguyen, Z. Zhang, H. Jia, M. Handfield, and C. J. Clancy.2003. Evaluation of the roles of four Candida albicans genes in virulence byusing gene disruption strains that express URA3 from the native locus. Infect.Immun. 71:6101–6103.
12. De Bernardis, F., M. Boccanera, D. Adriani, A. Girolamo, and A. Cassone.2002. Intravaginal and intranasal immunizations are equally effective ininducing vaginal antibodies and conferring protection against vaginal candi-diasis. Infect. Immun. 70:2725–2729.
13. De Bernardis, F., S. Arancia, L. Morelli, B. Hube, D. Sanglard, W. Schafer,and A. Cassone. 1999. Evidence that members of the secretory aspartylproteinase gene family, in particular SAP2, are virulence factors for Candidavaginitis. J. Infect. Dis. 179:201–208.
14. d’Ostiani, C. F., G. Del Sero, A. Bacci, C. Montagnoli, A. Spreca, A. Men-cacci, P. Ricciardi-Castagnoli, and L. Romani. 2000. Dendritic cells discrim-inate between yeasts and hyphae of the fungus Candida albicans: implica-tions for initiation of T helper cell immunity in vitro and in vivo. J. Exp. Med.191:1661–1673.
15. Fallon, K., K. Bausch, J. Noonan, E. Huguenel, and P. Tamburini. 1997.Role of aspartic proteases in disseminated Candida albicans infection inmice. Infect. Immun. 65:551–556.
16. Felk, A., M. Kretschmar, A. Albrecht, M. Schaller, S. Beinhauer, T. Nich-terlein, D. Sanglard, H. C. Korting, W. Schafer, and B. Hube. 2002. Candidaalbicans hyphal formation and the expression of the Efg1-regulated protein-ases Sap4 to Sap6 are required for the invasion of parenchymal organs.Infect. Immun. 70:3689–3700.
17. Fonzi, W. A., and M. Y. Irwin. 1993. Isogenic strain construction and genemapping in Candida albicans. Genetics 134:717–728.
18. Gillum, A. M., E. Y. Tsay, and D. R. Kirsch. 1984. Isolation of the Candidaalbicans gene for orotidine-5�-phosphate decarboxylase by complementationof S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 198:179–182.
19. Hostetter, M. K. 1994. Adhesins and ligands involved in the interaction ofCandida spp. with epithelial and endothelial surfaces. Clin. Microbiol. Rev.7:29–42.
20. Hube, B., D. Sanglard, F. C. Odds, D. Hess, M. Monod, W. Schafer, A. J.Brown, and N. A. Gow. 1997. Disruption of each of the secreted aspartylproteinase genes SAP1, SAP2, and SAP3 of Candida albicans attenuatesvirulence. Infect. Immun. 65:3529–3538.
21. Hube, B., and J. Naglik. 2001. Candida albicans proteinases: resolving themystery of a gene family. Microbiology 147:1997–2005.
22. Hube, B., M. Monod, D. A. Schofield, A. J. Brown, and N. A. Gow. 1994.Expression of seven members of the gene family encoding secretory asparticproteinases in Candida albicans. Mol. Microbiol. 14:87–99.
23. Jackson, B. E., K. R. Wilhelmus, and B. Hube. 2007. The role of secretedaspartyl proteinases in Candida albicans keratitis. Invest. Ophthalmol. Vis.Sci. 48:3559–3565.
24. Jones-Carson, J., A. Vazquez-Torres, T. Warner, and E. Balish. 2000. Dis-parate requirement for T cells in resistance to mucosal and acute systemiccandidiasis. Infect. Immun. 68:2363–2365.
4848 CORREIA ET AL. INFECT. IMMUN.
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

25. Kretschmar, M., A. Felk, P. Staib, M. Schaller, D. Hess, M. Callapina, J.Morschhauser, W. Schafer, H. C. Korting, H. Hof, B. Hube, and T. Nich-terlein. 2002. Individual acid aspartic proteinases (Saps) 1-6 of Candidaalbicans are not essential for invasion and colonization of the gastrointestinaltract in mice. Microb. Pathog. 32:61–70.
26. Kretschmar, M., B. Hube, T. Bertsch, D. Sanglard, R. Merker, M. Schroder,H. Hof, and T. Nichterlein. 1999. Germ tubes and proteinase activity con-tribute to virulence of Candida albicans in murine peritonitis. Infect. Immun.67:6637–6642.
27. Kwon-Chung, K. J. K., D. Lehman, C. Good, and P. T. Magee. 1985. Geneticevidence for role of extracellular proteinase in virulence of Candida albicans.Infect. Immun. 49:571–575.
28. Lay, J., K. Henry, J. Clifford, Y. Koltin, C. E. Bulawa, and J. M. Becker.1998. Altered expression of selectable marker URA3 in gene-disrupted Can-dida albicans strains complicates interpretation of virulence studies. Infect.Immun. 66:5301–5306.
29. Lermann, U., and J. Morschhauser. 2008. Secreted aspartic proteases arenot required for invasion of reconstituted human epithelia by Candida albi-cans. Microbiology 154:3281–3295.
30. MacCallum, D. M., and F. C. Odds. 2004. Temporal events in the intrave-nous challenge model for experimental Candida albicans infections in femalemice. Mycoses 48:151–161.
31. Mencacci, A., C. Montagnoli, A. Bacci, E. Cenci, L. Pitzurra, A. Spreca, M.Kopf, A. H. Sharpe, and L. Romani. 2002. CD80�Gr-1� myeloid cells inhibitdevelopment of antifungal Th1 immunity in mice with candidiasis. J. Immu-nol. 169:3180–3190.
32. Montagnoli, C., A. Bacci, S. Bozza, R. Gaziano, P. Mosci, A. H. Sharpe, andL. Romani. 2002. B7/CD28-dependent CD4�CD25� regulatory T cells areessential components of the memory-protective immunity to Candida albi-cans. J. Immunol. 169:6298–6308.
33. Naglik, J., A. Albrecht, O. Bader, and B. Hube. 2004. Candida albicansproteinases and host/pathogen interactions. Cell. Microbiol. 6:915–926.
34. Naglik, J. R., D. Moyes, J. Makwana, P. Kanzaria, E. Tsichlaki, G. Weindl,A. R. Tappuni, C. A. Rodgers, A. J. Woodman, S. J. Challacombe, M.Schaller, and B. Hube. 2008. Quantitative expression of the Candida albicanssecreted aspartyl proteinase gene family in human oral and vaginal candidi-asis. Microbiology 154:3266–3280.
35. Naglik, J. R., S. J. Challacombe, and B. Hube. 2003. Candida albicanssecreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol.Biol. Rev. 67:400–428.
36. Netea, M. G., R. Sutmuller, C. Hermann, C. A. A. Van der Graaf, J. W. M.Van der Meer, J. H. van Krieken, T. Hartung, G. Adema, and B. J. Kullberg.2004. Toll-like receptor 2 suppresses immunity against Candida albicansthrough induction of IL-10 and regulatory T cells. J. Immunol. 172:3712–3718.
37. Odds, F. C. 1994. Candida species and virulence. ASM News 60:313–318.38. Odds, F. C., N. A. R. Gow, and A. J. Brown. 2006. Toward a molecular
understanding of Candida albicans virulence, p. 305–319. In J. Heitman et al.(ed.), Molecular principles of fungal pathogenesis. ASM Press, Washington,DC.
39. Pappas, P. G. 2006. Invasive candidiasis. Infect. Dis. Clin. North Am. 20:485–506.
40. Rahman, D., M. Mistry, S. Thavarai, S. J. Challacombe, and J. R. Naglik.2007. Murine model of concurrent oral and vaginal Candida albicans colo-nization to study epithelial host-pathogen interactions. Microbes Infect.9:615–622.
41. Reuß, O., Å. Vik, R. Kolter, and J. Morschhauser. 2004. The SAT1 flipper,an optimized tool for gene disruption in Candida albicans. Gene 341:119–127.
42. Romani, L., C. Montagnoli, S. Bozza, K. Perruccio, A. Spreca, P. Allavena,S. Verbeek, R. A. Calderone, F. Bistoni, and P. Puccetti. 2004. The exploi-tation of distinct recognition receptors in dendritic cells determines the fullrange of host immune relationships with Candida albicans. Int. Immunol.16:149–161.
43. Sanglard, D., B. Hube, M. Monod, F. C. Odds, and N. A. Gow. 1997. A triple
deletion of the secreted aspartyl proteinase genes SAP4, SAP5, and SAP6 ofCandida albicans causes attenuated virulence. Infect. Immun. 65:3539–3546.
44. Saville, S. P., A. L. Lazzell, C. Monteagudo, and J. L. Lopez-Ribot. 2003.Engineered control of cell morphology in vivo reveals distinct roles for yeastand filamentous forms of Candida albicans during infection. Eukaryot. Cell2:1053–1060.
45. Schaller, M., H. C. Korting, W. Schafer, J. Bastert, W. Chen, and B. Hube.1999. Secreted aspartic proteinase (Sap) activity contributes to tissue dam-age in a model of human oral candidosis. Mol. Microbiol. 34:169–180.
46. Schaller, M., M. Bein, H. C. Korting, S. Baur, G. Hamm, M. Monod, S.Beinhauer, and B. Hube. 2003. The secreted aspartyl proteinases Sap1 andSap2 cause tissue damage in an in vitro model of vaginal candidiasis based onreconstituted human vaginal epithelium. Infect. Immun. 71:3227–3234.
47. Shoham, S., and S. M. Levitz. 2005. The immune response to fungal infec-tions. Br. J. Haematol. 129:569–582.
48. Spellberg, B., D. Johnston, Q. T. Phan, J. E. Edwards, Jr., S. W. French, A. S.Ibrahim, and S. G. Filler. 2003. Parenchymal organ, and not splenic, immu-nity correlates with host survival during disseminated candidiasis. Infect.Immun. 71:5756–5764.
49. Staib, F. 1969. Proteolysis and pathogenicity of Candida albicans strains.Mycopathol. Mycol. Appl. 37:345–348.
50. Staib, P., M. Kretschmar, T. Nichterlein, H. Hof, and J. Morschhauser.2000. Differential activation of a Candida albicans virulence gene familyduring infection. Proc. Natl. Acad. Sci. U. S. A. 97:6102–6107.
51. Sundstrom, P. 1999. Adhesins in Candida albicans. Curr. Opin. Microbiol.2:353–357.
52. Sundstrom, P., J. E. Cutler, and J. F. Staab. 2002. Reevaluation of the roleof HWP1 in systemic candidiasis by use of Candida albicans strains withselectable marker URA3 targeted to the ENO1 locus. Infect. Immun. 70:3281–3283.
53. Sutmuller, R. P., M. H. den Brok, M. Kramer, E. J. Bennink, L. W. Toonen,B. J. Kullberg, L. A. Joosten, S. Akira, M. G. Netea, and G. J. Adema. 2006.Toll-like receptor 2 controls expansion and function of regulatory T cells.J. Clin. Invest. 116:485–494.
54. Taylor, B. N., H. Hannemann, M. Sehnal, A. Biesemeier, A. Schweizer, M.Rollinghoff, and K. Schroppel. 2005. Induction of SAP7 correlates withvirulence in an intravenous infection model of candidiasis but not in a vaginalinfection model in mice. Infect. Immun. 73:7061–7063.
55. Taylor, P. R., G. D. Brown, A. B. Geldhof, L. Martinez-Pomares, and S.Gordon. 2003. Pattern recognition receptors and differentiation antigensdefine murine myeloid cell heterogeneity ex vivo. Eur. J. Immunol. 33:2090–2097.
56. Taylor, P. R., S. V. Tsoni, J. A. Willment, K. M. Dennehy, M. Rosas, H.Findon, K. Haynes, C. Steele, M. Botto, S. Gordon, and G. D. Brown. 2007.Dectin-1 is required for b-glucan recognition and control of fungal infection.Nat. Immunol. 8:31–38.
57. Thewes, S., M. Kretschmar, H. Park, M. Schaller, S. G. Filler, and B. Hube.2007. In vivo and ex vivo comparative transcriptional profiling of invasive andnon-invasive Candida albicans isolates identifies genes associated with tissueinvasion. Mol. Microbiol. 63:1606–1628.
58. Vazquez-Torres, A., and E. Balish. 1997. Macrophages in resistance to can-didiasis. Microbiol. Mol. Biol. Rev. 61:170–192.
59. Vilanova, M., L. Teixeira, I. Caramalho, E. Torrado, A. Marques, P. Ma-dureira, A. Ribeiro, P. Ferreira, M. Gama, and J. Demengeot. 2004. Protec-tion against systemic candidiasis in mice immunized with secreted asparticproteinase 2. Immunology 111:334–342.
60. Vonk, A. G., M. G. Netea, J. W. M. van der Meer, and B. J. Kullberg. 2006.Host defence against disseminated Candida albicans infection and implica-tions for antifungal immunotherapy. Expert Opin. Biol. Ther. 6:891–903.
61. Wagner, R. D., A. Vazquez-Torres, J. Jones-Carson, T. Warner, and E.Balish. 1996. B cell knockout mice are resistant to mucosal and systemiccandidiasis of endogenous origin but susceptible to experimental systemiccandidiasis. J. Infect. Dis. 174:589–597.
62. Warenda, A. J., S. Kauffman, T. P. Sherrill, J. M. Becker, and J. B. Konopka.2003. Candida albicans septin mutants are defective for invasive growth andvirulence. Infect. Immun. 71:4045–4051.
Editor: G. S. Deepe, Jr.
VOL. 78, 2010 ROLE OF Saps IN MURINE DISSEMINATED CANDIDIASIS 4849
on August 23, 2019 by guest
http://iai.asm.org/
Dow
nloaded from