Embed Size (px)
Citation preview

Copyright # 2006 John Wiley & Sons, Ltd. Received 10 November 2004Accepted 18 April 2005
AQUATIC CONSERVATION: MARINE AND FRESHWATER ECOSYSTEMS
Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
Published online in Wiley InterScience(www.interscience.wiley.com). DOI: 10.1002/aqc.724
Linking geomorphic character, behaviour and condition tofluvial biodiversity: implications for river management
BRUCE C. CHESSMANa,*, KIRSTIE A. FRYIRSb and GARY J. BRIERLEYc
aNSW Department of Infrastructure, Planning and Natural Resources, Parramatta, NSW 2150, AustraliabDepartment of Physical Geography, Division of Environmental and Life Sciences, Macquarie University,
North Ryde, NSW 2109, AustraliacSchool of Geography and Environmental Science, University of Auckland, Private Bag 92019,
Auckland, New Zealand
ABSTRACT
1. The River Styles1 framework is a geomorphic approach to the classification of river types,assessment of the physical condition of rivers, and planning of physical rehabilitation. However, thelinkages between River Styles and aquatic biodiversity conservation are still only weakly developed.2. In this study, 41 sites in the Bega River basin in New South Wales, Australia, were classified
according to River Style and geomorphic condition, and surveyed for four biological assemblages:diatoms, aquatic and semi-aquatic macrophytes, aquatic macroinvertebrates and fish.3. Each assemblage differed significantly among River Styles. However, in the case of diatoms and
fish, these differences could be accounted for by geographic clustering of sites in the same RiverStyle, and a tendency for River Styles to occupy particular altitudinal zones and sizes of streams.This result was attributed to the overriding influences of water quality on diatoms and of altitude-related variation in water temperature and distance from the ocean on fish. For macrophytes andmacroinvertebrates, geomorphic river type appeared to exert a direct influence, probably viavariation in physical habitat characteristics.4. Geomorphic condition, judged as good, moderate or poor by reference to the inferred natural
condition of each River Style, was also significantly associated with differences in biologicalassemblages other than fish. Twice as many taxa appeared to favour sites in good geomorphiccondition as favoured sites in poor condition. Many of the taxa associated with sites in poorcondition are alien taxa introduced to Australia since European settlement.5. These findings imply that protection of reaches that are in good geomorphic condition is likely
to be critical for the maintenance of indigenous biodiversity, and that rehabilitation of geomorphiccondition can assist in the rehabilitation of native riverine biota.Copyright # 2006 John Wiley & Sons, Ltd.
KEY WORDS: diatom; fish; geomorphology; macroinvertebrate; macrophyte; river condition; river rehabilitation;
River Style
*Correspondence to: Dr B.C. Chessman, NSW Department of Infrastructure, Planning and Natural Resources, PO Box 3720,Parramatta, NSW 2124, Australia. E-mail: [email protected]

INTRODUCTION
The management of biodiversity in rivers and riparian zones sometimes involves the direct addition orremoval of biota, such as the restocking of fish populations or the control of alien pest species. More often,however, goals for fluvial biodiversity are pursued indirectly through the manipulation of environmentaldrivers such as the flow regime (Stanford et al., 1996; Poff et al., 1997; Robinson and Uehlinger, 2003) andphysical structure (Frissell et al., 1986; Rhoads et al., 1999). Success in this approach relies on anunderstanding of ecosystem patterns, dynamics and interrelationships at a range of spatial and temporalscales (Hildrew, 1996; Petts, 2000; Ward et al., 2001; Frothingham et al., 2002). A failure to establish such afoundation can result in locally inappropriate actions, which do not achieve the desired outcomes (Rosgen,1994; Ebersole et al., 1997; Lockwood and Pimm, 1999).
Fluvial geomorphology provides a logical base from which to manipulate aquatic biodiversity throughthe medium of physical habitat (Brussock et al., 1985; Sear, 1994; Harper and Everard, 1998; Newson,2002), and via geomorphological influences on other driving variables such as temperature (Hawkins et al.,1997). Rehabilitation of riverine habitats through the reinstatement of important fluvial processes isreceiving increasing impetus from developments in public policy such as the European Union’s WaterFramework Directive and Habitats Directive (Clarke et al., 2003). A geomorphic template approach isbased on an understanding of the natural physical character and behaviour of a river in its landscape andvalley settings, how these properties have changed under the influence of human disturbance, and theirlikely future trajectory. Consequently, rehabilitation works can be tailored to suit individual river reachesand their contemporary hydrological and sedimentological drivers. This perspective underlies the RiverStyles1 framework of Brierley and Fryirs (2000, 2005).
River Styles are lengths of river with a characteristic degree of valley confinement, a certain channelplanform and a particular suite of geomorphic units and bed materials (Brierley and Fryirs, 2005). TheRiver Styles framework has four stages (Brierley et al., 2002). Stage 1 involves the classification of riverreaches throughout a drainage basin according to valley setting (ranging from fully confined to laterallyunconfined), channel planform (channel sinuosity, lateral stability, continuity and number), geomorphicunit assemblage and dominant bed materials. Geomorphic units include features such as bedrock steps,point bars, sand sheets, pools, riffles and cascades. Bed materials range from bedrock and large boulders tosand and mud. In Stage 2, the geomorphic condition of each reach is assessed in relation to a ‘natural’reference condition for the relevant style (Fryirs, 2003). This reference condition is inferred from reaches ofthe style with little artificial modification and from historical analysis of reach evolution. In Stage 3, thereach is placed on an evolutionary pathway of progressive deterioration or recovery in condition, andwhere applicable the potential for recovery is assessed (Fryirs and Brierley, 2000). Together, these stagesprovide the basis for setting the priority of reaches for conservation or rehabilitation, as well as informingthe design of structural works where needed to assist the latter. This forms Stage 4 of the framework(Brierley et al., 2002).
Each River Style operates under a specific set of controls dictated by topography (slope and lateralconfinement) and hydrology (stream power). Certain flow characteristics, such as base flows, bankfull flowsand over-bank events, mould different assemblages of geomorphic units, and dictate the way in which bedmaterials are stored and reworked along a reach. These processes are modified by human disturbances ofcatchments and riparian zones, such as the removal of vegetation. Potential linkages between the RiverStyles framework and aquatic biodiversity arise from the expectation that differences in the assemblage ofgeomorphic units and bed materials among reaches of varying geomorphic style and condition willengender predictable differences in biological communities through the medium of physical habitat(Thomson et al., 2001, 2004; Brierley and Fryirs, 2005). If this expectation is realized, rehabilitation ofgeomorphic condition across the suite of River Styles present in a basin should have consequent benefits forthe conservation of natural aquatic biodiversity. In other words, unless other factors such as poor water
B.C. CHESSMAN ET AL.268
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

quality are limiting, natural biodiversity should be maximized where diverse River Styles exist in goodgeomorphic condition.
This paper evaluates relationships among River Styles, their geomorphic condition, and four fluvialbiological assemblages. These are diatoms, aquatic and semi-aquatic macrophytes, aquatic macroinverte-brates and fish. Biological assemblages are shown to differ among both River Styles and reaches in differinggeomorphic condition. Some consequent issues are explored, which concern the further development oflinkages between fluvial geomorphology and aquatic ecology. Implications are presented for riverrehabilitation that is based on improving geomorphic condition.
METHODS
Study area
The Bega River basin covers 2850 km2 in the State of New South Wales, Australia (Plate 1). Most of thebasin is drained by the Bemboka and Brogo rivers, which rise at altitudes around 1200m and unite to formthe Bega River near the town of Bega. The basin also includes several discrete stream systems to the northof the Bega River, the largest of which is the Murrah River system. All stream systems drain eastward to theTasman Sea within the Pacific Ocean. The geology of the basin is mostly Devonian granite andgranodiorite, and its soils are typically deep, sandy and well-drained. About half of the basin is coveredwith indigenous forest, whereas the remainder is cleared for grazing by dairy and beef cattle. This clearing,which followed the arrival of European settlers in the mid-nineteenth century, has resulted in profoundgeomorphic changes along many rivers. These include massive channel incision and widening, sometimesresulting in the creation of continuous stream channels where discontinuous channels previously existed,and associated mobilization and deposition of vast quantities of sediment (Brooks and Brierley, 1997, 2000;Brierley and Fryirs, 1998, 1999, 2000, 2005; Fryirs and Brierley, 1998, 1999, 2000, 2001; Brierley et al.,1999). The Brogo and Bemboka rivers are both impounded in their upper reaches, and river flows areabstracted extensively for irrigation, and to a much lesser extent for urban water supply.
Geomorphic classification and assessment
The classification of reaches within the Bega River basin into River Styles, and the assessment of theirgeomorphic condition, have been described by Brierley and Fryirs (2000), Brierley et al. (2002) and Fryirs(2001, 2003). River reaches throughout the basin that are greater than third order were classified into nineRiver Styles (Plate 1; Table 1). The condition of each reach (Figure 1) was rated as either ‘good’ (close tothe inferred natural reference condition), ‘moderate’ (somewhat modified by human impact) or ‘poor’(severely modified) on the basis of variables that are considered relevant for each River Style (Fryirs, 2001,2003). One of the key variables is the assemblage of geomorphic units. In a particular River Style, differentgeomorphic units are present at various stages in the evolutionary sequence of human impact and recovery.For example, a reach in good condition belonging to the River Style known as ‘partly confined valley withbedrock-controlled discontinuous floodplain’ is expected to have pools induced by bedrock that areseparated by riffles, islands and runs. If a sand sheet is present, filling pools and smothering riffles and runs,the reach is considered to be in poor geomorphic condition for that River Style (see Table 2 for furtherdetail).
Biological surveys
Surveys of diatoms, macrophytes and macroinvertebrates took place in February–March 2000, with fishsampled in May–July of the same year. Forty-one sites were surveyed, representing all nine River Styles
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 269
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
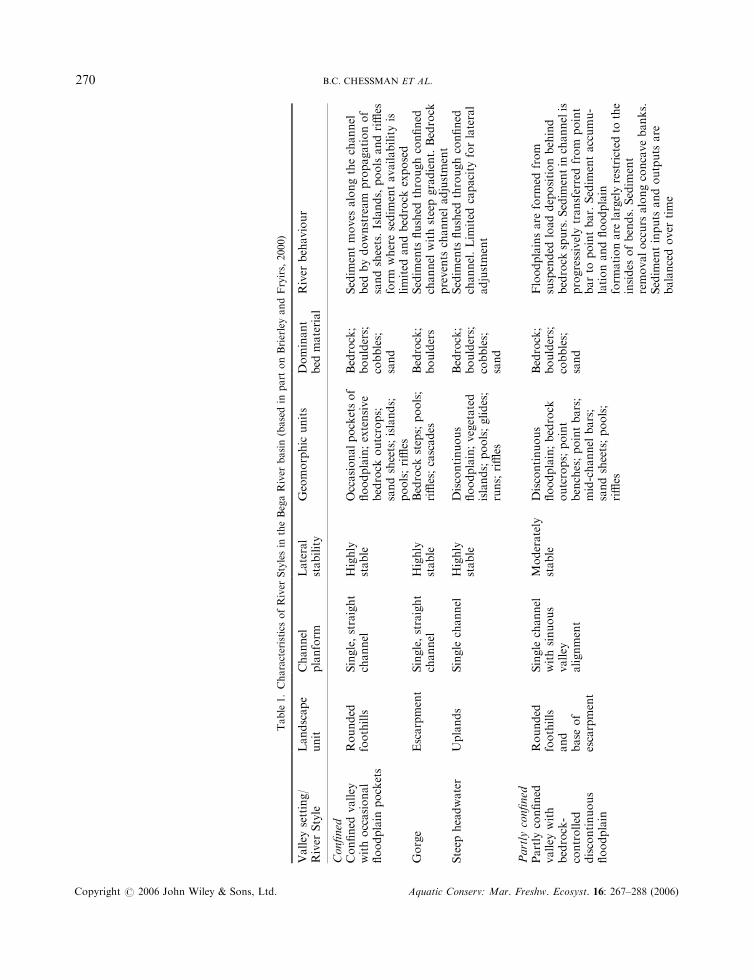
Table1.Characteristics
ofRiver
Stylesin
theBegaRiver
basin(basedin
part
onBrierleyandFryirs,2000)
Valley
setting/
River
Style
Landscape
unit
Channel
planform
Lateral
stability
Geomorphic
units
Dominant
bed
material
River
behaviour
Confined
Confined
valley
withoccasional
floodplain
pockets
Rounded
foothills
Single,straight
channel
Highly
stable
Occasionalpocketsof
floodplain;extensive
bedrock
outcrops;
sandsheets;islands;
pools;riffles
Bedrock;
boulders;
cobbles;
sand
Sedim
entmoves
alongthechannel
bed
bydownstream
propagationof
sandsheets.Islands,poolsandriffles
form
wheresedim
entavailabilityis
limited
andbedrock
exposed
Gorge
Escarpment
Single,straight
channel
Highly
stable
Bedrock
steps;pools;
riffles;cascades
Bedrock;
boulders
Sedim
ents
flushed
throughconfined
channel
withsteepgradient.Bedrock
prevents
channel
adjustment
Steep
headwater
Uplands
Single
channel
Highly
stable
Discontinuous
floodplain;vegetated
islands;pools;glides;
runs;riffles
Bedrock;
boulders;
cobbles;
sand
Sedim
ents
flushed
throughconfined
channel.Lim
ited
capacity
forlateral
adjustment
Partly
confined
Partly
confined
valley
with
bedrock-
controlled
discontinuous
floodplain
Rounded
foothills
and
base
of
escarpment
Single
channel
withsinuous
valley
alignment
Moderately
stable
Discontinuous
floodplain;bedrock
outcrops;point
benches;pointbars;
mid-channel
bars;
sandsheets;pools;
riffles
Bedrock;
boulders;
cobbles;
sand
Floodplainsare
form
edfrom
suspended
loaddepositionbehind
bedrock
spurs.Sedim
entin
channelis
progressivelytransferredfrom
point
barto
pointbar.Sedim
entaccumu-
lationandfloodplain
form
ationare
largelyrestricted
tothe
insides
ofbends.Sedim
ent
removaloccurs
alongconcavebanks.
Sedim
entinputs
andoutputs
are
balancedover
time
B.C. CHESSMAN ET AL.270
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
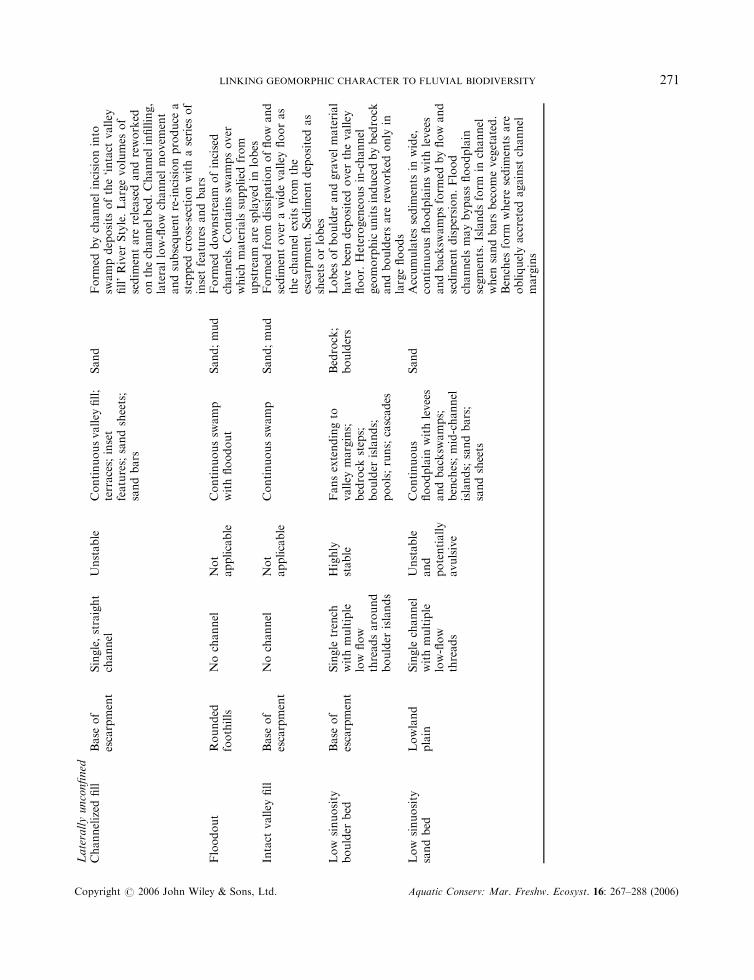
Laterallyunconfined
Channelized
fill
Base
of
escarpment
Single,straight
channel
Unstable
Continuousvalley
fill;
terraces;inset
features;sandsheets;
sandbars
Sand
Form
edbychannel
incisioninto
swampdeposits
ofthe‘intact
valley
fill’River
Style.Largevolumes
of
sedim
entare
releasedandreworked
onthechannelbed.Channelinfilling,
laterallow-flow
channel
movem
ent
andsubsequentre-incisionproduce
astepped
cross-sectionwithaseries
of
insetfeaturesandbars
Floodout
Rounded
foothills
Nochannel
Not
applicable
Continuoussw
amp
withfloodout
Sand;mud
Form
eddownstream
ofincised
channels.Containssw
ampsover
whichmaterialssupplied
from
upstream
are
splayed
inlobes
Intact
valley
fill
Base
of
escarpment
Nochannel
Not
applicable
Continuoussw
amp
Sand;mud
Form
edfrom
dissipationofflow
and
sedim
entover
awidevalley
flooras
thechannel
exitsfrom
the
escarpment.Sedim
entdepositedas
sheets
orlobes
Low
sinuosity
boulder
bed
Base
of
escarpment
Single
trench
withmultiple
low
flow
threadsaround
boulder
islands
Highly
stable
Fansextendingto
valley
margins;
bedrock
steps;
boulder
islands;
pools;runs;cascades
Bedrock;
boulders
Lobes
ofboulder
andgravel
material
havebeendepositedover
thevalley
floor.Heterogeneousin-channel
geomorphicunitsinducedbybedrock
andbouldersare
reworked
only
inlargefloods
Low
sinuosity
sandbed
Lowland
plain
Single
channel
withmultiple
low-flow
threads
Unstable
and
potentially
avulsive
Continuous
floodplain
withlevees
andbacksw
amps;
benches;mid-channel
islands;sandbars;
sandsheets
Sand
Accumulatessedim
ents
inwide,
continuousfloodplainswithlevees
andbacksw
ampsform
edbyflow
and
sedim
entdispersion.Flood
channelsmaybypass
floodplain
segments.Islandsform
inchannel
when
sandbars
becomevegetated.
Benches
form
wheresedim
ents
are
obliquelyaccretedagainst
channel
margins
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 271
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
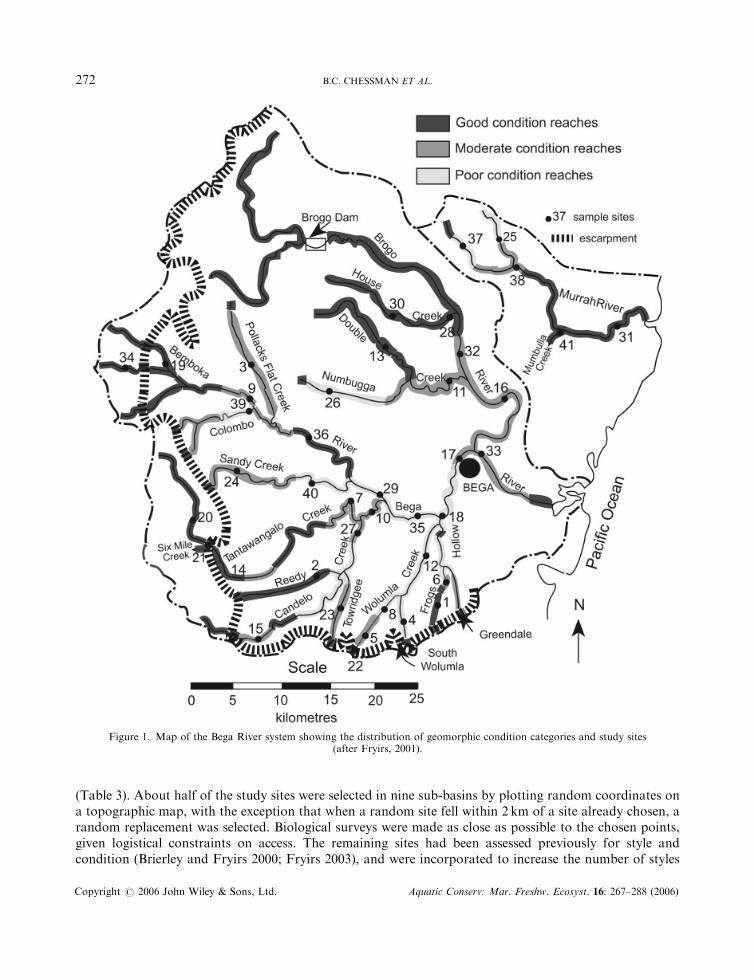
(Table 3). About half of the study sites were selected in nine sub-basins by plotting random coordinates ona topographic map, with the exception that when a random site fell within 2 km of a site already chosen, arandom replacement was selected. Biological surveys were made as close as possible to the chosen points,given logistical constraints on access. The remaining sites had been assessed previously for style andcondition (Brierley and Fryirs 2000; Fryirs 2003), and were incorporated to increase the number of styles
Figure 1. Map of the Bega River system showing the distribution of geomorphic condition categories and study sites(after Fryirs, 2001).
B.C. CHESSMAN ET AL.272
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

Table 2. Example of typical attributes of reaches in good, moderate and poor geomorphic condition for one River Style in theBega River basin (partly confined valley with bedrock-controlled discontinuous floodplain) (based in part on Fryirs (2001) and
Brierley and Fryirs (2005))
Attribute Good Moderate Poor
Channel Not expanded. Low width-depthratio, with significant localvariability induced by bedrockoutcrops, vegetation and woodydebris. Cross-sections areasymmetrical at bend apices andirregular at inflection points.Islands and bars are vegetatedwith hardy shrubs and aquaticgrasses. Natural or low rate oferosion of concave banks.
Expanded, including at inflectionpoints. High width-depth ratio.Cross-sections are asymmetrical atbend apices and symmetrical atinflection points. Point bars andbenches are largely unvegetated ordominated by alien species. Nowoody debris. Localized erosion ofconcave banks.
Overwidened, including atinflection points. High width–depthratio. Cross-sections areasymmetrical at bend apices andsymmetrical at inflection points.No within-channel vegetation orwoody debris. Accelerated rates ofconcave bank erosion and channelexpansion along the entire reach.
Channelplanform andvegetation
Low-sinuosity, single channelwithin a meandering valley.Moderate lateral stability.Occasional bedrock outcrops,bank-attached bars and sandsheets cause division of flow.Bar-island-riffle complexes areseparated by pools. Point barsand point benches occur onbends. Discontinuous pockets offloodplain may be scouredaround large trees and shrubs.Continuous or scattered riparianwoody vegetation consists mainlyof indigenous species. Banks arelined with Lomandra sp. Pointbars are colonized by tussocks.Some hardy shrubs on benches.
Low-sinuosity, single channel withina meandering valley. Laterallyunstable at concave banks.Point benches, point bars, concavebenches, localized sand sheetsand well-defined low-flow channelare present. Occasional bedrockoutcrops. Discontinuous pockets offloodplain are either scoured orstripped. Sparse cover of riparianwoody vegetation. Floodplaindominated by pasture.
Low-sinuosity, single channelwithin a meandering valley.Laterally unstable at concavebanks and inflection points,reflecting channel expansion. Pointbars, sand sheets, localized bedrockoutcrops and multi-stringedlow-flow channel are present.Discontinuous pockets offloodplain are either stripped orcharacterized by short-cuttingflood-channels or extensivesand sheets. No riparian woodyvegetation. Floodplain dominatedby pasture.
Bedcharacter
Well-segregated bedload, withdiscrete pockets of material ofdifferent textures. Some graveldeposits in riffles. Bars andbenches comprise sands. Highin-stream roughness (vegetationand woody debris) promotelocalized deposition of fine-grainedmaterials and organics. Highhydraulic diversity. Bed is stablewith no incision or aggradation.Balance maintained betweensediment input and output alongthe reach. Acts as a sedimenttransport zone.
Poorly sorted bed material. Sandsheets create local homogeneity,reducing roughness and the range ofhydraulic diversity. Bed isunstable with local aggradationand incision. However, a balance ismaintained between sediment inputand output along the reach. Acts as asediment transport zone.
Near-homogeneous (unsorted)in-stream and floodplain sandsheets. Hydraulically homogeneous.Bed is unstable with majoraggradation and local incision.Sediment transport is limited. Actsas a sediment accumulation zone.
Examplephotograph
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 273
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
and condition categories included, and to generate replicates. Macrophytes were surveyed at all sites. Theother groups of biota were surveyed only where there was sufficient water for sampling. A duplicatemacrophyte survey was performed at one site.
Two diatom samples were taken at each site where surface water was present. Where possible, sampleswere taken from stone surfaces at depths of about 150mm } one in fast-flowing water and one in slow-flowing water. However, flowing water and stony substrata were absent or rare at several sites, and in thesecases samples were taken in still water and from substrata such as submerged wood and macrophytes.Environmental variation was such that consistency in sampled habitats and flow conditions could seldombe maintained even within a style. Each substratum was shaken underwater for 3 s to remove loose silt anddebris, and then scraped with a sharpened wooden spatula. Scrapings were preserved in Lugol’s iodinesolution and forwarded to a specialist laboratory, where the samples were cleared and mounted andapproximately 300 valves per sample were identified to species level.
Aquatic and semi-aquatic macrophyte species (excluding trees and shrubs) were recorded by a carefulsearch of four large quadrats at each site. Regardless of channel size, each quadrat measured 20m in thedownstream direction and extended laterally across the streambed and out to 20m from the edge of the bedon each bank. The quadrats were separated by 40-m gaps. Each species observed was scored for eachquadrat as absent (0), isolated (1), scattered (3), forming beds or stands (5) or overgrowing or filling thechannel (7). In about 10% of cases, a decision between adjacent categories was difficult, and anintermediate score was used (e.g. isolated–scattered scored 2).
Three macroinvertebrate samples were taken at each site, generally one from a riffle or similargeomorphic unit with fast current, one out of the current at the edge of a pool, and one away from the edgein the dominant aquatic geomorphic unit (usually a glide or pool). At sites where little or no flowing waterwas present, the riffle sample was substituted with a sample from the dominant or subdominant geomorphicunit, so that total sampling effort was consistent. Each sample was collected with a pond net (0.25-mmmesh) by kick or sweep techniques for a period of 3min. Samples were placed in white sorting trays andpicked by eye for 30min with the aim of retrieving the maximum number of invertebrate types (Chessman,1995). Specimens were preserved with 70% ethanol and sent to a specialist laboratory for identification,generally to family level. This level was chosen to constrain costs, and because current taxonomicknowledge enables only about 40% of river macroinvertebrates collected in routine surveys in New SouthWales to be identified to species level.
Fish were surveyed by the New South Wales Fisheries Department with a backpack electrofisher,operated in a single pass for about 30min at most sites. At some sites with little water, less time (down to16min) was expended. Stunned fish were collected in a dip net and placed in containers of water to recover.Fish were identified to species, counted and subsequently released. Fish observed but not captured werealso recorded.
Table 3. Number of sites sampled in each combination of River Style and geomorphic condition in the Bega River basin. Style symbolsare used in Table 4 and Figure 2
River Style Symbol Good Moderate Poor
Channelized fill A 1 3 4Confined valley with occasional floodplain pockets B 4 4 3Floodout C } 1 }Gorge D 2 } }Intact valley fill E 1 1 }Low sinuosity boulder bed F 1 } }Low sinuosity sand bed G 1 4 1Partly confined valley with bedrock-controlled discontinuous floodplain H 3 3 2Steep headwater I 2 } }
B.C. CHESSMAN ET AL.274
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

Data analysis
Diatom data for each site were expressed as the proportion of valves represented by each species, averagedfor the two samples. Macrophyte data were expressed as the average score per species per quadrat,macroinvertebrate data as the average number of specimens per family per sample and fish data as thenumber of specimens per species per 30min of electrofishing. Diatom, macroinvertebrate and fish data werethen transformed for analysis as logarithm (xþ 1). Macrophyte data were not log-transformed because thescheme used for rating abundance in the field already approximated a logarithmic scale.
For each biotic assemblage, all surveyed sites were ordinated by two-dimensional non-metricmultidimensional scaling (NMS) with the PC-ORD software package (McCune and Mefford, 1999). Thestatistical significance of differences among River Styles and geomorphic condition categories was tested foreach assemblage by the MRPP (multi-response permutation procedure) in PC-ORD. This is anonparametric procedure that tests the null hypothesis of no difference among predefined groups ofentities (in this case, sites grouped by River Style or condition category). Because of limitations in theMRPP algorithm, River Styles represented by only one site for a given assemblage had to be excluded.Indicator species analysis in PC-ORD (Dufrene and Legendre, 1997) was used to identify individual taxathat were characteristic of particular River Styles or condition categories. This analysis generates anindicator value, which ranges from 0 if the taxon shows no discrimination among the predefined groups to100 if the taxon is exclusively associated with a single group. For all of these analyses, biological differencesamong sites were expressed by the S�rensen (Bray–Curtis) dissimilarity measure.
Since sites of the same River Style and condition category tended to be geographically clustered (see Plate1 and Figure 1), it is possible that biological similarities among sites of the same style or condition couldarise simply from the physical proximity of these sites, rather than from their geomorphic affinity. Thiscould occur, for example, because organisms can disperse more readily among sites that are geographicallyclose than among distant sites. It could also occur through confounding by environmental factors otherthan style and condition. For example, water temperature regime (related to altitude) and flow regime(related to catchment area above the site) are also likely to be more similar at nearby than at distant sites.
In an effort to disentangle the effects of these potentially confounding factors, every possible pairing ofsites was characterized as same-style or different-style. The biological difference between members of eachpair was expressed by the S�rensen dissimilarity measure applied to transformed data for each biologicalassemblage, as described above. The straight-line geographic distance (in kilometres) between the membersof each pair was calculated, together with the absolute difference in altitude and in drainage area. Theabsolute difference in geomorphic condition was also calculated for each pair, after scoring good conditionas 3, moderate condition as 2 and poor condition as 1. Analysis of covariance (ANCOVA) was then used totest the contribution of style, condition, geographic distance, altitude and catchment area to the biologicaldifferences. A separate analysis was run for each assemblage, with biological difference as the dependentvariable, style as a fixed factor (either same-style or different-style) and difference in condition, geographicdistance, difference in altitude and difference in catchment area as covariates. The altitudinal andcatchment-area differences were logarithmically transformed as the raw values had skewed distributions.
RESULTS
Biological differences among River Styles and geomorphic condition categories
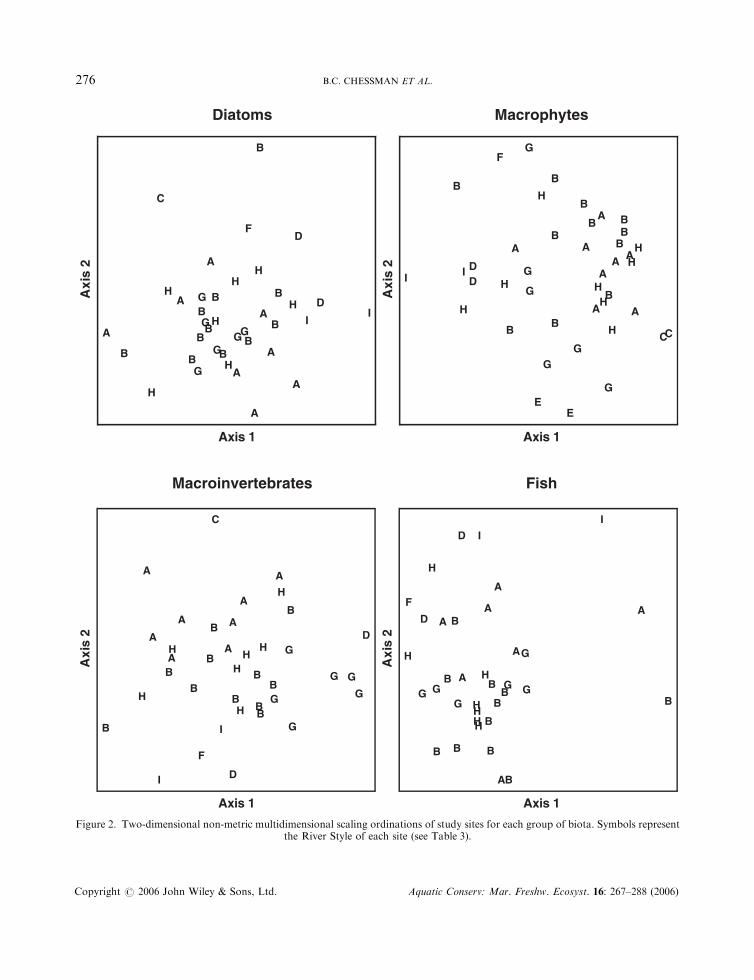
Two-dimensional NMS ordinations of all sites surveyed for each biotic assemblage had medium stressvalues (0.20 for diatoms, 0.21 for macrophytes, 0.18 for macroinvertebrates and 0.19 for fish). Althoughsites of the same style were often clustered in ordination space, the styles overlapped one anotherconsiderably (Figure 2). MRPP revealed highly significant differences among River Styles for each
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 275
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
Diatoms Macrophytes
A
A
AA
C
H B
B
B
B
HH
HG
GG
F
I
D
DA
A
A
H
BB
H
G
G
G
IB
B
A
BB
H
B
Axis 1
Axi
s 2
E
A
A
AA
CC
H
H
BB
B
BH
H
H
G
GG
F
IDD
E
A
A A
H
B
B
H
G
G
GI
B
B
A
B
B
H
B
Axis 1
Axi
s 2
Macroinvertebrates Fish
A
A A
A
C
H
B
BB
B
H
H
H G
G
G
F
I
D
DA
A
A
H
BBH
G
G
G
I
B
B
A
B
B
H
B
Axis 1
Axi
s 2
A
A
A
H
B
B
B
BH
H
H
GG G
F
ID
D A
A
A
H
BB
H
G
GG
I
B B
A
B
B
H
B
Axis 1
Axi
s 2
Figure 2. Two-dimensional non-metric multidimensional scaling ordinations of study sites for each group of biota. Symbols representthe River Style of each site (see Table 3).
B.C. CHESSMAN ET AL.276
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

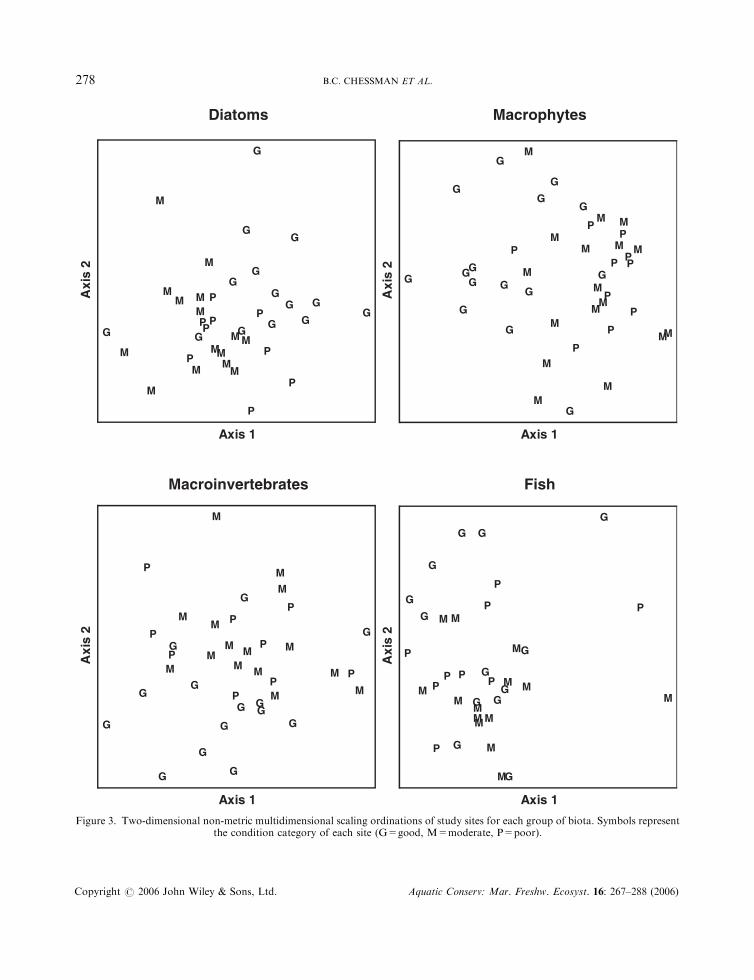
assemblage (P ¼ 0:002 for diatoms; P50:001 for macrophytes and macroinvertebrates; P ¼ 0:001 for fish).Differences according to condition category were significant for diatoms (P ¼ 0:002), macrophytes(P50:001) and macroinvertebrates (P ¼ 0:002), but not for fish (P > 0:05). Except in the case of fish, sitesin good condition tended to segregate in ordination space, whereas sites in moderate or poor conditionoverlapped considerably (Figure 3).
Indicator species analysis identified 15 diatom species, nine macrophyte species, 18 macroinvertebratefamilies and three fish species (from study totals of 173 species, 83 species, 123 families and 17 species,respectively) that differed significantly (P50:05) among styles in absolute or relative abundance (Table 4).These numbers are about 2–3 times as great as expected by chance at a ¼ 0:05 (about 9, 4, 6 and 1,respectively). Individual indicator taxa reached their greatest abundance in five of the nine styles,particularly those symbolized in Table 4 as D (gorge), G (low sinuosity sand bed) and I (steep headwater).No indicator taxon was most abundant in styles B (confined valley with occasional floodplain pockets), E(intact valley fill), F (low sinuosity boulder bed) or H (partly confined valley with bedrock-controlleddiscontinuous floodplain).
Most of the macrophyte species that differed significantly among styles had their strongest associationwith the ‘floodout’ style (style C in Table 4). This style is characterized particularly by the presence ofcontinuous swampy areas and substrata of sand and mud (Table 1) – conditions that would clearly favourproliferation of large emergent macrophytes such as Eleocharis sp., Phragmites australis, Rumex brownii,Schoenoplectus validus and Typha orientalis. In contrast the ‘gorge’ style (style D in Table 4), with a steepgradient and rocky substrata, was strongly associated with smaller, shallow-rooted plants (Ranunculus sp.and Rorippa nasturtium-aquaticum). Alternanthera denticulata, which typically grows on unshaded sand andgravel bars, was most strongly associated with the style known as ‘low sinuosity sand bed’ (style G inTable 4).
Of the individual macroinvertebrate families that differed significantly among styles (Table 4), the groupassociated with the ‘channelized fill’ style (style A in Table 4) comprised Aeshnidae, Ceinidae,Lumbriculidae and Naucoridae. This style occurs at the base of the escarpment and is characterized byextensive sand sheets and sand bars (Table 1). In contrast, the ‘gorge’ style (style D in Table 4), with itsdominance of bedrock steps, boulders, riffles and cascades, was associated with the Calamoceratidae,Cordulephyidae, Ecnomidae, Hydryphantidae, Limnesiidae and Telephlebiidae. Several macroinvertebratefamilies } the Corydalidae, Dixidae, Helicopsychidae, Hydrophilidae, Mideopsidae and Ptilodactylidae }
were most strongly associated with the ‘steep headwater’ style (style I in Table 4). Characteristics of thisstyle include a range of flow environments and stony substrata (Table 1). Differences in substratumrequirements probably explain some of these associations } for example, extensive sand would suitLumbriculidae, which are sediment-burrowing worms, whereas Helicopsychidae, which graze algae fromrock surfaces (Gooderham and Tsyrlin, 2002), require stony substrata.
Thirteen diatom species, 19 macrophyte species, 19 macroinvertebrate families and one fish speciesdiffered significantly (P50:05) in absolute or relative abundance among geomorphic condition categories(Table 5). With the exception of fish, these numbers are again substantially greater than expected bychance. Thirty-four of these taxa reached their greatest abundance in the ‘good’ category, compared withonly one in the ‘moderate’ category and 17 in the ‘poor’ category. Five or six of the taxa with their greatestabundance in the ‘poor’ category are alien species introduced by humans to Australia. (The native or alienstatus of one plant species, the water pepper Persicaria hydropiper, is uncertain.) In contrast, no alienspecies were identified among those most associated with good geomorphic condition.
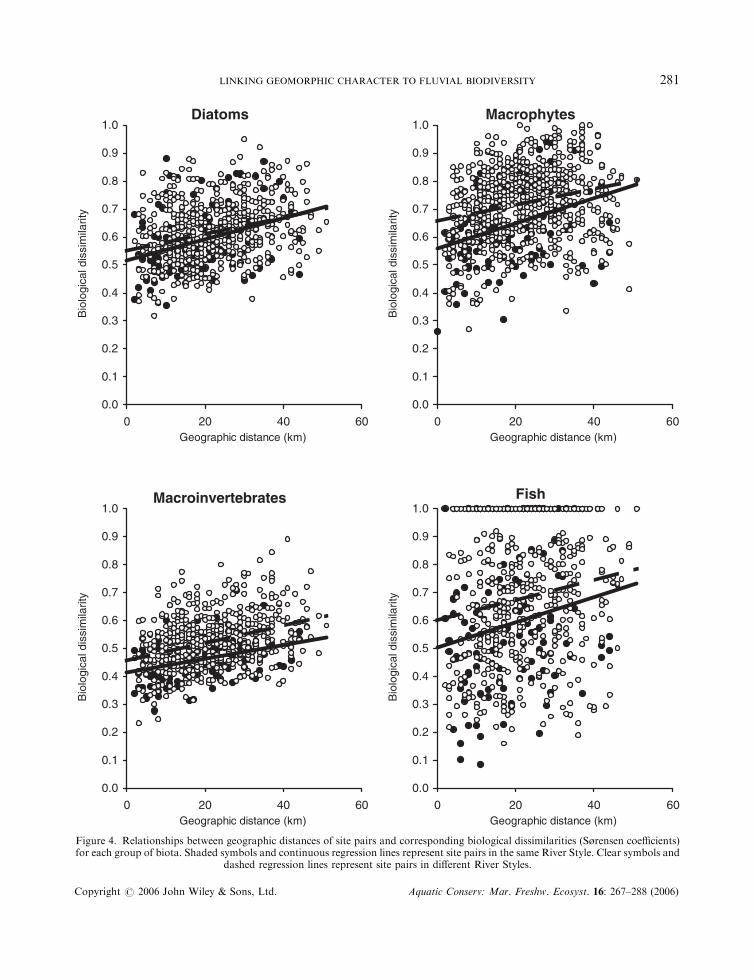
Effects of geographic separation, altitude and catchment area
For macrophytes, macroinvertebrates and fish, but not for diatoms, two sites with a given geographicseparation tended to be more biologically dissimilar if they were in different River Styles than if they were
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 277
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
Diatoms Macrophytes
G
M
PM
M
M G
M
M
P
GG
MM
MP
G
G
G
GM
P
P
M
GP
G
G
M
M
GP
G
P
MM
P
G
Axis 1
Axi
s 2
G
G
M
PM
MM
M
P
GM
M
PG
G
M
M
MP
G
GGG
M
M
P P
M
G
P
G
G
M
MG
P
G
P
M
M
P
G
Axis 1
Axi
s 2
Macroinvertebrates Fish
G
M P
M
M
M
G
MM
P
G
G
M M
M
P
G
G
G
GM
P
P
M
GPG
G
M
M
G
P
G
P
M
M
P
G
Axis 1
Axi
s 2
M
P
M
M
G
M
M
PG
G
M
MM P
G
GG
G M
P
P
M
GP
G
G
MM
G
P G
P
M
M
P
G
Axis 1
Axi
s 2
Figure 3. Two-dimensional non-metric multidimensional scaling ordinations of study sites for each group of biota. Symbols representthe condition category of each site (G=good, M=moderate, P=poor).
B.C. CHESSMAN ET AL.278
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

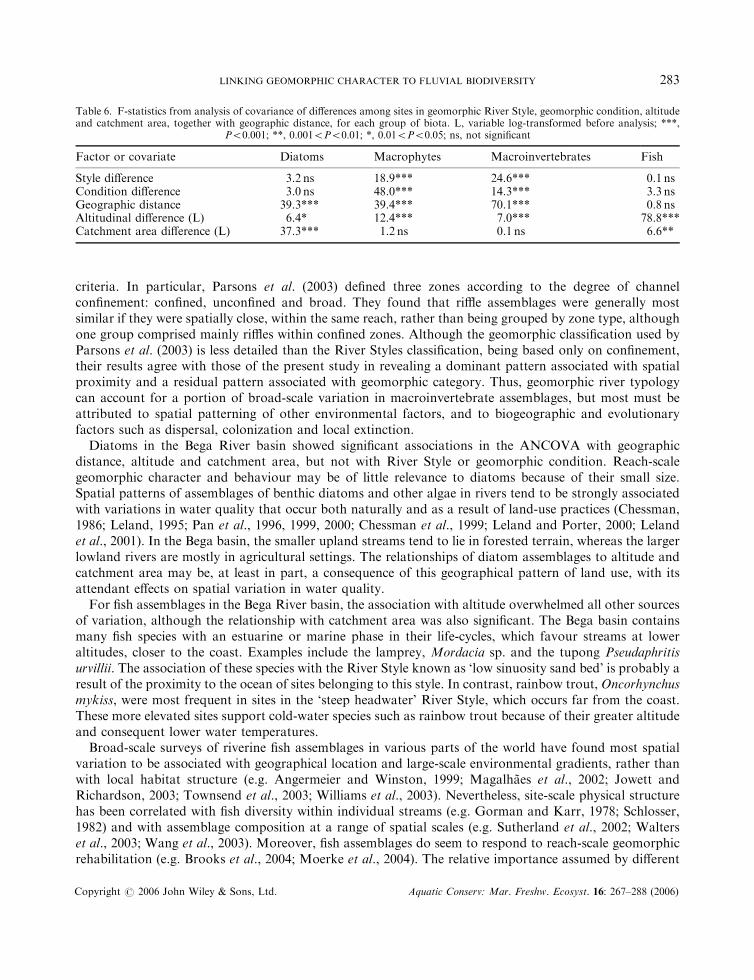
both in the same style (Figure 4). ANCOVA indicated that biological dissimilarity between members of sitepairs was significantly related to River Style and geomorphic condition for macrophytes andmacroinvertebrates, but not for diatoms and fish (Table 6). The effect of altitude was significant for all
Table 4. Taxa that differed significantly in abundance among River Styles in indicator species analysis (P50:05), listed according tothe style where each was most common. Asterisks indicate alien taxa. Refer to Table 3 for explanation of style symbols
Style Group Taxon Indicator value Probability
A Macrophytes Juncus articulatus* 33.7 0.002A Macroinvertebrates Aeshnidae 47.2 0.021A Macroinvertebrates Ceinidae 40.9 0.006A Macroinvertebrates Lumbriculidae* 35.1 0.001A Macroinvertebrates Naucoridae 57.3 0.038C Macrophytes Eleocharis sp. 64.3 0.031C Macrophytes Phragmites australis 47.3 0.048C Macrophytes Rumex brownii 39.9 0.005C Macrophytes Schoenoplectus validus 43.3 0.009C Macrophytes Typha orientalis 64.2 0.002D Diatoms Diatomella balfouriana 42.5 0.040D Diatoms Frustulia rhomboides 56.4 0.030D Diatoms Gomphonema micropus 58.5 0.024D Diatoms Nitzschia recta 68.8 0.012D Diatoms Rhopalodia gibba 45.7 0.029D Macrophytes Ranunculus sp. 72.7 0.012D Macrophytes Rorippa nasturtium-aquaticum* 37.0 0.001D Macroinvertebrates Calamoceratidae 46.1 0.041D Macroinvertebrates Cordulephyidae 58.8 0.015D Macroinvertebrates Ecnomidae 46.4 0.014D Macroinvertebrates Hydryphantidae 59.7 0.025D Macroinvertebrates Limnesiidae 68.0 0.015D Macroinvertebrates Telephlebiidae 53.5 0.022G Diatoms Anorthoneis excentrica 37.8 0.046G Diatoms Diploneis elliptica 53.1 0.026G Diatoms Navicula cryptocephala 27.8 0.009G Diatoms Navicula incertata 69.0 0.013G Diatoms Reimeria sinuata 71.6 0.006G Macrophytes Alternanthera denticulata 66.5 0.031G Macroinvertebrates Hygrobatidae 47.0 0.033G Macroinvertebrates Tipulidae 38.8 0.035G Fish Mordacia sp. 93.8 0.001G Fish Pseudaphritis urvillii 54.8 0.029I Diatoms Cocconeis placentula 28.1 0.017I Diatoms Gomphonema angustum 37.0 0.006I Diatoms Gomphonema minutum 70.6 0.005I Diatoms Navicula cryptotenella 63.6 0.019I Diatoms Nitzschia dissipata 36.5 0.050I Macroinvertebrates Corydalidae 39.0 0.032I Macroinvertebrates Dixidae 56.5 0.029I Macroinvertebrates Helicopsychidae 80.4 0.009I Macroinvertebrates Hydrophilidae 27.4 0.022I Macroinvertebrates Mideopsidae 40.7 0.025I Macroinvertebrates Ptilodactylidae 66.7 0.012I Fish Oncorhynchus mykiss* 100.0 0.009
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 279
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
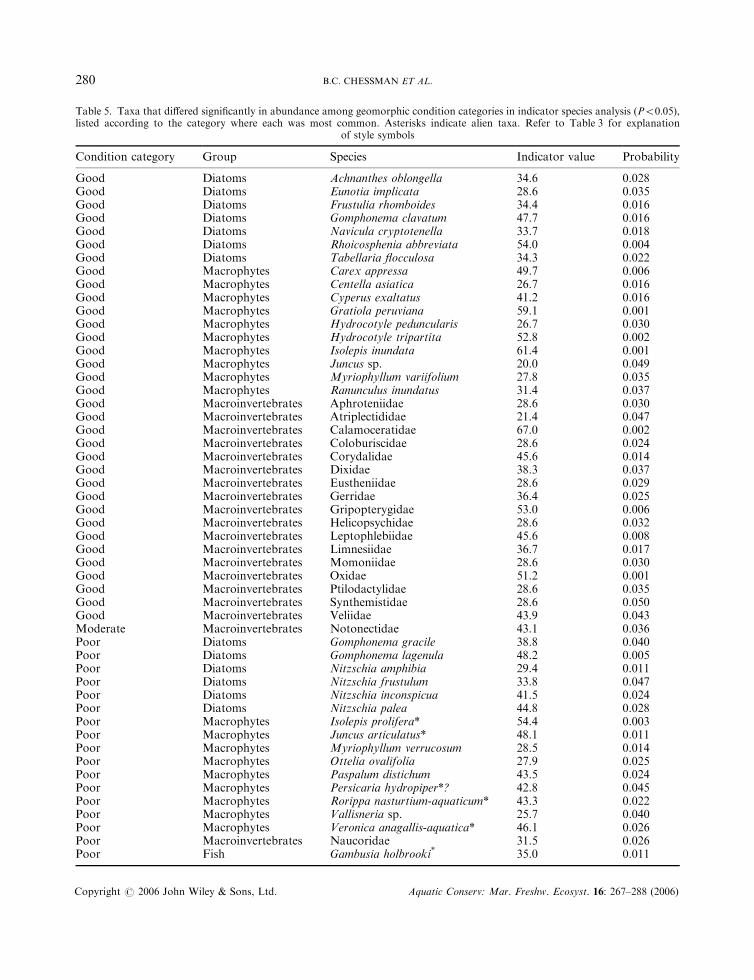
Table 5. Taxa that differed significantly in abundance among geomorphic condition categories in indicator species analysis (P50:05),listed according to the category where each was most common. Asterisks indicate alien taxa. Refer to Table 3 for explanation
of style symbols
Condition category Group Species Indicator value Probability
Good Diatoms Achnanthes oblongella 34.6 0.028Good Diatoms Eunotia implicata 28.6 0.035Good Diatoms Frustulia rhomboides 34.4 0.016Good Diatoms Gomphonema clavatum 47.7 0.016Good Diatoms Navicula cryptotenella 33.7 0.018Good Diatoms Rhoicosphenia abbreviata 54.0 0.004Good Diatoms Tabellaria flocculosa 34.3 0.022Good Macrophytes Carex appressa 49.7 0.006Good Macrophytes Centella asiatica 26.7 0.016Good Macrophytes Cyperus exaltatus 41.2 0.016Good Macrophytes Gratiola peruviana 59.1 0.001Good Macrophytes Hydrocotyle peduncularis 26.7 0.030Good Macrophytes Hydrocotyle tripartita 52.8 0.002Good Macrophytes Isolepis inundata 61.4 0.001Good Macrophytes Juncus sp. 20.0 0.049Good Macrophytes Myriophyllum variifolium 27.8 0.035Good Macrophytes Ranunculus inundatus 31.4 0.037Good Macroinvertebrates Aphroteniidae 28.6 0.030Good Macroinvertebrates Atriplectididae 21.4 0.047Good Macroinvertebrates Calamoceratidae 67.0 0.002Good Macroinvertebrates Coloburiscidae 28.6 0.024Good Macroinvertebrates Corydalidae 45.6 0.014Good Macroinvertebrates Dixidae 38.3 0.037Good Macroinvertebrates Eustheniidae 28.6 0.029Good Macroinvertebrates Gerridae 36.4 0.025Good Macroinvertebrates Gripopterygidae 53.0 0.006Good Macroinvertebrates Helicopsychidae 28.6 0.032Good Macroinvertebrates Leptophlebiidae 45.6 0.008Good Macroinvertebrates Limnesiidae 36.7 0.017Good Macroinvertebrates Momoniidae 28.6 0.030Good Macroinvertebrates Oxidae 51.2 0.001Good Macroinvertebrates Ptilodactylidae 28.6 0.035Good Macroinvertebrates Synthemistidae 28.6 0.050Good Macroinvertebrates Veliidae 43.9 0.043Moderate Macroinvertebrates Notonectidae 43.1 0.036Poor Diatoms Gomphonema gracile 38.8 0.040Poor Diatoms Gomphonema lagenula 48.2 0.005Poor Diatoms Nitzschia amphibia 29.4 0.011Poor Diatoms Nitzschia frustulum 33.8 0.047Poor Diatoms Nitzschia inconspicua 41.5 0.024Poor Diatoms Nitzschia palea 44.8 0.028Poor Macrophytes Isolepis prolifera* 54.4 0.003Poor Macrophytes Juncus articulatus* 48.1 0.011Poor Macrophytes Myriophyllum verrucosum 28.5 0.014Poor Macrophytes Ottelia ovalifolia 27.9 0.025Poor Macrophytes Paspalum distichum 43.5 0.024Poor Macrophytes Persicaria hydropiper*? 42.8 0.045Poor Macrophytes Rorippa nasturtium-aquaticum* 43.3 0.022Poor Macrophytes Vallisneria sp. 25.7 0.040Poor Macrophytes Veronica anagallis-aquatica* 46.1 0.026Poor Macroinvertebrates Naucoridae 31.5 0.026Poor Fish Gambusia holbrooki* 35.0 0.011
B.C. CHESSMAN ET AL.280
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
Diatoms
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
0 20 40 60Geographic distance (km)
Bio
logi
cal d
issi
mila
rity
Macrophytes
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
0 20 40 60Geographic distance (km)
Bio
logi
cal d
issi
mila
rity
Macroinvertebrates
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
0 20 40 60Geographic distance (km)
Bio
logi
cal d
issi
mila
rity
Fish
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
0 20 40 60Geographic distance (km)
Bio
logi
cal d
issi
mila
rity
Figure 4. Relationships between geographic distances of site pairs and corresponding biological dissimilarities (S�rensen coefficients)for each group of biota. Shaded symbols and continuous regression lines represent site pairs in the same River Style. Clear symbols and
dashed regression lines represent site pairs in different River Styles.
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 281
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

four biological assemblages and geographic distance was significant for all but fish. Catchment area wassignificant for diatoms and fish only. The significance levels in Table 6 should be interpreted with somecaution because the data, covering all possible site pairs, are not all statistically independent. For example,the differences between sites 1 and 2, between sites 1 and 3, and between sites 2 and 3 are not independent ofone another. However, the significance levels for the relationships of style and condition with macrophytesand macroinvertebrates were very high (P50:001).
DISCUSSION
The MRPP analysis showed that all four biological assemblages differed significantly among River Styles inthe Bega River basin, and all assemblages except fish differed according to geomorphic condition. Thus,sites of the same geomorphic style and condition tended to be more biologically similar than sites ofdifferent styles or sites in differing condition. This could have arisen through direct biological responses tothe differing physical habitat conditions afforded by each combination of style and condition. However,River Styles and condition categories are geographically clustered in the Bega basin, rather than beingdistributed at random. This raises the possibility that these relationships are indirect } a consequence ofconfounding by other factors that vary spatially in a similar manner to River Styles and conditioncategories.
Biological dispersal and migration are potential confounding factors because movement is generallyeasier between sites that are close together than sites that are farther apart. Long-range movement requiresthe expenditure of more energy and time than short-range movement and may be prevented by hostileenvironments or biogeographic barriers. Short-range movements could result in a partial homogenizationof the flora and fauna of nearby sites, which would also tend to be of the same style and condition.Alternatively, confounding could have arisen if biological distributions are controlled by physical factorssuch as water temperature and stream size rather than those factors that are embodied in the River Stylesand condition classifications (principally lateral confinement, channel planform, geomorphic units and bedmaterials). Stream sites of the same style and condition, by often being close together, can also be similar instream size and temperature regime.
The ANCOVA attempted to separate the roles of these factors by incorporating geographic proximity (asan easily measured surrogate for biotic mixing), altitude (as a surrogate for temperature regime) andcatchment area (as a surrogate for stream size). This analysis suggested that even when these factors weretaken into account, a significant effect of River Style and condition remained for macrophytes andmacroinvertebrates. However, the effect was lost for diatoms and fish.
If geomorphic style directly affects macrophyte and macroinvertebrate assemblages, it may do so becauseof differences in geomorphic units and bed composition among styles. It is well known that medium tosmall-scale morphological features and substratum type in streams are correlated with assemblagecomposition of both macrophytes (e.g. Bornette et al., 1994; Willby et al., 2000; Bernez et al., 2004) andmacroinvertebrates (e.g. Allan, 1975; Brussock and Brown, 1991; Vinson and Hawkins, 1998). However,assemblage composition has seldom been related explicitly to reach-scale classification of fluvialgeomorphology. Thomson et al. (2004) compared macroinvertebrate assemblages (at the taxonomic levelof family) between pool and run geomorphic units in three River Styles in northern New South Wales. Theyfound statistically significant differences among styles for pool macroinvertebrates (although not for thoseassociated with runs), but did not address the role of geographic separation and proximity. Parsons et al.(2003) assessed broad-scale variation in assemblages of riffle-dwelling macroinvertebrates in south-easternAustralia, mostly identified to genus and species level. They tailored their study to a set of nested spatialscales, from catchments and zones within catchments down to reaches and riffles within reaches. Thesespatial units were defined partly on hydrological criteria (e.g. tributary inflows), but also on geomorphic
B.C. CHESSMAN ET AL.282
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
criteria. In particular, Parsons et al. (2003) defined three zones according to the degree of channelconfinement: confined, unconfined and broad. They found that riffle assemblages were generally mostsimilar if they were spatially close, within the same reach, rather than being grouped by zone type, althoughone group comprised mainly riffles within confined zones. Although the geomorphic classification used byParsons et al. (2003) is less detailed than the River Styles classification, being based only on confinement,their results agree with those of the present study in revealing a dominant pattern associated with spatialproximity and a residual pattern associated with geomorphic category. Thus, geomorphic river typologycan account for a portion of broad-scale variation in macroinvertebrate assemblages, but most must beattributed to spatial patterning of other environmental factors, and to biogeographic and evolutionaryfactors such as dispersal, colonization and local extinction.
Diatoms in the Bega River basin showed significant associations in the ANCOVA with geographicdistance, altitude and catchment area, but not with River Style or geomorphic condition. Reach-scalegeomorphic character and behaviour may be of little relevance to diatoms because of their small size.Spatial patterns of assemblages of benthic diatoms and other algae in rivers tend to be strongly associatedwith variations in water quality that occur both naturally and as a result of land-use practices (Chessman,1986; Leland, 1995; Pan et al., 1996, 1999, 2000; Chessman et al., 1999; Leland and Porter, 2000; Lelandet al., 2001). In the Bega basin, the smaller upland streams tend to lie in forested terrain, whereas the largerlowland rivers are mostly in agricultural settings. The relationships of diatom assemblages to altitude andcatchment area may be, at least in part, a consequence of this geographical pattern of land use, with itsattendant effects on spatial variation in water quality.
For fish assemblages in the Bega River basin, the association with altitude overwhelmed all other sourcesof variation, although the relationship with catchment area was also significant. The Bega basin containsmany fish species with an estuarine or marine phase in their life-cycles, which favour streams at loweraltitudes, closer to the coast. Examples include the lamprey, Mordacia sp. and the tupong Pseudaphritisurvillii. The association of these species with the River Style known as ‘low sinuosity sand bed’ is probably aresult of the proximity to the ocean of sites belonging to this style. In contrast, rainbow trout, Oncorhynchusmykiss, were most frequent in sites in the ‘steep headwater’ River Style, which occurs far from the coast.These more elevated sites support cold-water species such as rainbow trout because of their greater altitudeand consequent lower water temperatures.
Broad-scale surveys of riverine fish assemblages in various parts of the world have found most spatialvariation to be associated with geographical location and large-scale environmental gradients, rather thanwith local habitat structure (e.g. Angermeier and Winston, 1999; Magalhaes et al., 2002; Jowett andRichardson, 2003; Townsend et al., 2003; Williams et al., 2003). Nevertheless, site-scale physical structurehas been correlated with fish diversity within individual streams (e.g. Gorman and Karr, 1978; Schlosser,1982) and with assemblage composition at a range of spatial scales (e.g. Sutherland et al., 2002; Walterset al., 2003; Wang et al., 2003). Moreover, fish assemblages do seem to respond to reach-scale geomorphicrehabilitation (e.g. Brooks et al., 2004; Moerke et al., 2004). The relative importance assumed by different
Table 6. F-statistics from analysis of covariance of differences among sites in geomorphic River Style, geomorphic condition, altitudeand catchment area, together with geographic distance, for each group of biota. L, variable log-transformed before analysis; ***,
P50:001; **, 0:0015P50:01; *, 0:015P50:05; ns, not significant
Factor or covariate Diatoms Macrophytes Macroinvertebrates Fish
Style difference 3.2 ns 18.9*** 24.6*** 0.1 nsCondition difference 3.0 ns 48.0*** 14.3*** 3.3 nsGeographic distance 39.3*** 39.4*** 70.1*** 0.8 nsAltitudinal difference (L) 6.4* 12.4*** 7.0*** 78.8***Catchment area difference (L) 37.3*** 1.2 ns 0.1 ns 6.6**
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 283
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

factors in structuring fish assemblages depends on the scale of the study (Lamoroux et al., 1999; Jacksonet al., 2001), and so it is inevitable that large-scale studies will emphasize biogeographic factors. Thesefactors are even more important for fish than for macroinvertebrates, since many freshwater invertebratescan disperse by non-aquatic pathways (Bilton et al., 2001), whereas fish are restricted to dispersal throughthe channel network. Such dispersal can be blocked by physical barriers (e.g. waterfalls) or reaches ofunfavourable habitat. Studies at a smaller spatial extent than the present one (e.g. within narrow bands ofaltitude), and at a finer spatial grain (e.g. within in-channel geomorphic units rather than across a 200-mreach) would elucidate the relationships between fluvial geomorphology and fish assemblages in the BegaRiver basin (cf. Bond and Lake, 2003). Such studies would also provide further insights for macrophytesand macroinvertebrates, by allowing the relationships with geomorphology to stand out from the large-scale biogeographic background.
Relationships between biological assemblages and geomorphic condition are crucial from the perspectiveof river rehabilitation. The value of the River Styles framework for rehabilitation is that remedial methodscan be tailored to each style. This is possible because each style has characteristic behaviour in its naturaland disturbed states, and capacity to adjust (Brierley and Fryirs, 2005). Understanding these style-specificattributes enables management interventions to be designed to ‘work with nature’ (Brierley et al., 2002).These interventions aim to improve geomorphic condition by reinstating natural physical processes. Forexample, the appropriate placement of woody debris can induce a mixture of small-scale aggradation andscouring that leads to morphological diversity rather than uniformity (Brooks et al., 2004).
However, if one of the goals of rehabilitation is to enhance indigenous biodiversity, clear linkagesbetween geomorphic and biological condition need to be established. The significant relationships betweengeomorphic condition and both macrophytes and macroinvertebrates, and the partial segregation of sites ingood condition in ordination space, suggest that such linkages do occur. The taxa that differed significantlyin absolute or relative abundance among condition categories were usually most abundant in sites of goodcondition. However, alien taxa were strongly associated with sites in poor condition, perhaps reflecting apreference of invasive species for disturbed environments (Mack et al., 2000). Thus, reaches in goodgeomorphic condition are not merely biologically different from reaches in poor condition, but sustainnative biodiversity.
The influence of geomorphic condition on biodiversity can be explained by differences in physical habitatamong condition categories of the same style. In the Bega valley, deterioration in geomorphic condition isoften associated with increased physical instability through channel incision and widening, and habitatsimplification through the smothering of complex stony substrata with mobile sand sheets (Fryirs,2001, 2003). Instability is likely to be detrimental to sensitive macrophyte and macroinvertebrate species(Death and Winterbourn, 1995; Biggs et al., 2001), and simplification is likely to have an adverse effecton many macroinvertebrate species that favour complex stony substrata (Blyth et al., 1984; Hogg andNorris, 1991).
The observed differences in macrophyte and macroinvertebrate assemblages were mainly between sites ingood and moderate condition rather than between sites in moderate and poor condition. This suggests thatmacrophytes and macroinvertebrates are sensitive to geomorphic deterioration from good to moderatecondition, but that further deterioration from moderate to poor condition may have little additional impacton biodiversity. Therefore, protection of reaches that are in good geomorphic condition is likely to becritical to maintenance of indigenous biodiversity. Once the process of deterioration in geomorphiccondition begins, it is often difficult and costly to arrest. For reaches that are already in moderate or poorgeomorphic condition, any rehabilitation works should aim to achieve good geomorphic condition in orderto engender a high level of biological recovery.
Capabilities for predicting the biological effects of physical rehabilitation works would benefit from aschema that integrates geomorphic controls with other environmental drivers of the composition andabundance of riverine biological assemblages. Works aimed at physical improvement may not lead to
B.C. CHESSMAN ET AL.284
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

biological recovery if other factors such as altered flow regimes or poor water quality are limiting. Althoughthe River Styles classification has relationships to the hydrological regime, geomorphic structure isassociated mainly with large flow events that are competent to move bed materials and reshape riverchannels (e.g. Church, 1995). The frequencies of such flows are often altered downstream of largeimpoundments, with major and interlinked consequences for river geomorphology and biology (e.g. Petts,1980; Ligon et al., 1995; Power et al., 1996; Murle et al., 2003). However, biological condition may also beinfluenced substantially by artificial hydrological alterations that are unlikely to induce geomorphic change} for example altered seasonal timing of flows and abstraction of water at times of low flow (Bunn andArthington, 2002).
Water quality changes may be associated indirectly with geomorphic changes, since both water qualityand geomorphology can respond to human alterations of catchments such as widespread forest clearingand agricultural or urban development. However, geomorphic rehabilitation often proceeds by in-channeland riparian works that do not alter catchment land-use and therefore will have little effect on waterquality. In many other cases, water quality is affected by diffuse or point inflows in ways that are unrelatedto geomorphic condition.
An integrated approach to the prediction of biological recovery might proceed by the development ofhydrological and water quality classifications and condition ratings to complement the River Stylesframework (Thomson et al., 2004). Chessman and Royal (2004) developed a preliminary method thatjointly considers physical structure, hydrology and water quality, in order to predict spatial patterns ofaquatic macroinvertebrate assemblages within biogeographic regions. As yet the structural component ofthis scheme is limited to a simple dichotomy of stony and soft-bottom streams. However, it is intended toexpand this component to consider the more detailed classification of morphologies in the River Stylesframework. An integrated prediction system would allow the identification of any factors that are limitingto biological recovery in a particular river, so that management can be targeted on removal of theseconstraints.
ACKNOWLEDGEMENTS
We thank Jim Thomson and Mark Taylor for discussions and contributions to the geomorphological fieldwork as partof a project funded by Land and Water Australia (MQU6). Jim Armstrong, Andrew Brooks, Matthew Dasey, TimHaeusler, Bronwen Jones, Shelley Nancarrow, Ivars Reinfelds and Jenny Wood are gratefully acknowledged for theirparticipation in the biological fieldwork. Fish were surveyed by NSW Fisheries, diatoms were identified by Peter Gell’sresearch group at the University of Adelaide, and macroinvertebrates were identified by WSL Consultants. We thankDavid Outhet for comments on a draft manuscript.
REFERENCES
Allan JD. 1975. The distributional ecology and diversity of benthic insects in Cement Creek, Colorado. Ecology 56:1040–1053.
Angermeier PL, Winston MR. 1999. Characterizing fish community diversity across Virginia landscapes: prerequisitefor conservation. Ecological Applications 9: 335–349.
Bernez I, Daniel H, Haury J, Ferreira MT. 2004. Combined effects of environmental factors and regulation onmacrophyte vegetation along three rivers in western France. River Research and Applications 20: 43–59.
Biggs BJF, Duncan MJ, Suren AM, Holomuzki JR. 2001. The importance of bed sediment stability to benthicecosystems of streams. In Gravel-bed Rivers V, Mosley MP (ed.). Water Resources Publications: LLC, HighlandsRanch, CO; 423–449.
Bilton DT, Freeland JR, Okamura B. 2001. Dispersal in freshwater invertebrates. Annual Review of Ecology andSystematics 32: 159–181.
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 285
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

Blyth JD, Doeg TJ, StClair RM. 1984. Response of the macroinvertebrate fauna of the Mitta Mitta River, Victoria, tothe construction and operation of Dartmouth Dam. 1. Construction and initial filling period. Occasional Papers fromthe Museum of Victoria 1: 83–100.
Bond NR, Lake PS. 2003. Characterizing fish-habitat associations in streams as the first step in ecological restoration.Austral Ecology 28: 611–621.
Bornette G, Henry C, Barrat M, Amoros C. 1994. Theoretical habitat templates, species traits and species richness:aquatic macrophytes in the upper Rhone River and its floodplain. Freshwater Biology 31: 487–505.
Brierley GJ, Fryirs K. 1998. A fluvial sediment budget for upper Wolumla Creek, south coast, New South Wales,Australia. Australian Geographer 29: 107–124.
Brierley GJ, Fryirs K. 1999. Tributary-trunk stream relations in a cut-and-fill landscape, New South Wales, Australia.Geomorphology 28: 61–73.
Brierley GJ, Fryirs K. 2000. River Styles, a geomorphic approach to catchment characterization: implications for riverrehabilitation in Bega catchment, New South Wales, Australia. Environmental Management 25: 661–679.
Brierley GJ, Fryirs K. 2005. Geomorphology and River Management: Applications of the River Styles Framework.Blackwell Publishing: Oxford.
Brierley GJ, Cohen T, Fryirs K, Brooks A. 1999. Post-European changes to the fluvial geomorphology of Begacatchment, Australia: implications for river ecology. Freshwater Biology 41: 839–848.
Brierley GJ, Fryirs K, Outhet D, Massey C. 2002. Application of the River Styles framework as a basis for rivermanagement in New South Wales, Australia. Applied Geography 22: 91–122.
Brooks AP, Brierley GJ. 1997. Geomorphic responses of lower Bega River to catchment disturbance, 1851–1926.Geomorphology 18: 291–304.
Brooks AP, Brierley GJ. 2000. The role of European disturbance in the metamorphosis of the lower Bega River.In River Management: The Australasian Experience, Brizga S, Finlayson B (eds). John Wiley: New York;221–246.
Brooks AP, Gehrke PC, Jansen JD, Abbe T. 2004. Experimental reintroduction of woody debris on the Williams River,NSW: geomorphic and ecological responses. River Research and Applications 20: 513–536.
Brussock PP, Brown AV. 1991. Riffle-pool geomorphology disrupts longitudinal patterns of stream benthos.Hydrobiologia 220: 109–117.
Brussock PP, Brown AV, Dixon JC. 1985. Channel form and stream ecosystem models. Water Resources Bulletin21: 859–866.
Bunn SE, Arthington AH. 2002. Basic principles and ecological consequences of altered flow regimes for aquaticbiodiversity. Environmental Management 30: 492–507.
Chessman BC. 1986. Diatom flora of an Australian river system: spatial patterns and environmental relationships.Freshwater Biology 16: 805–819.
Chessman BC. 1995. Rapid assessment of rivers using macroinvertebrates: a procedure based on habitat-specificsampling, family level identification, and a biotic index. Australian Journal of Ecology 20: 122–129.
Chessman BC, Royal MJ. 2004. Bioassessment without reference sites: use of environmental filters to predict naturalassemblages of river macroinvertebrates. Journal of the North American Benthological Society 23: 599–615.
Chessman B, Growns I, Currey J, Plunkett-Cole N. 1999. Prediction of diatom communities at the genus level for therapid biological assessment of rivers. Freshwater Biology 41: 317–331.
Church M. 1995. Geomorphic response to river flow regulation: case studies and time-scales. Regulated Rivers:Research and Management 11: 3–22.
Clarke SJ, Bruce-Burgess L, Wharton G. 2003. Linking form and function: towards an eco-hydromorphic approach tosustainable river restoration. Aquatic Conservation: Marine and Freshwater Ecosystems 13: 439–450.
Death RG, Winterbourn MJ. 1995. Diversity patterns in stream benthic invertebrate communities: the influence ofhabitat stability. Ecology 76: 1446–1460.
Dufrene M, Legendre P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach.Ecological Monographs 67: 345–366.
Ebersole JL, Liss WJ, Frissell CA. 1997. Restoration of stream habitats in the western United States: restoration asreexpression of habitat capacity. Environmental Management 21: 1–14.
Frissell CA, Liss WJ, Warren CE, Hurley MD. 1986. A hierarchical framework for stream habitat classification:viewing streams in a watershed context. Environmental Management 10: 199–214.
Frothingham KM, Rhoads BL, Herricks EE. 2002. A multiscale conceptual framework for integrated ecogeomor-phological research to support stream naturalization in the agricultural Midwest. Environmental Management29: 16–33.
Fryirs KA. 2001. A geomorphic approach for assessing the condition and recovery potential of rivers: application inBega Catchment, South Coast, New South Wales, Australia. PhD thesis, Macquarie University, Sydney.
B.C. CHESSMAN ET AL.286
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

Fryirs K. 2003. Guiding principles for assessing geomorphic river condition: application of a framework in the Begacatchment, South Coast, New South Wales, Australia. Catena 53: 17–52.
Fryirs K, Brierley GJ. 1998. The character and age structure of valley fills in upper Wolumla Creek catchment,south coast, New South Wales, Australia. Earth Surface Processes and Landforms 23: 271–287.
Fryirs K, Brierley GJ. 1999. Slope-channel decoupling in Wolumla catchment, New South Wales, Australia: thechanging nature of sediment sources following European settlement. Catena 35: 41–63.
Fryirs K, Brierley GJ. 2000. A geomorphic approach to identification of river recovery potential. Physical Geography21: 244–277.
Fryirs K, Brierley GJ. 2001. Variability in sediment delivery and storage along river courses in Bega catchment, NSW,Australia: implications for geomorphic river recovery. Geomorphology 38: 237–265.
Gooderham J, Tsyrlin E. 2002. The Waterbug Book. A Guide to the Freshwater Invertebrates of Temperate Australia.CSIRO Publishing: Collingwood, Australia.
Gorman OT, Karr JR. 1978. Habitat structure and stream fish communities. Ecology 59: 507–515.Harper D, Everard M. 1998. Why should the habitat-level approach underpin holistic river survey and management?Aquatic Conservation: Marine and Freshwater Ecosystems 8: 395–413.
Hawkins CP, Hogue JN, Decker LM, Feminella JW. 1997. Channel morphology, water temperature, and assemblagestructure of stream insects. Journal of the North American Benthological Society 16: 728–749.
Hildrew AG. 1996. Whole river ecology: spatial scale and heterogeneity in the ecology of running waters. Archiv furHydrobiologie Supplement 113: 25–43.
Hogg ID, Norris RH. 1991. Effects of runoff from land clearing and urban development on the distributionand abundance of macroinvertebrates in pool areas of a river. Australian Journal of Marine and Freshwater Research42: 507–518.
Jackson DA, Peres-Neto PR, Olden JD. 2001. What controls who is where in freshwater fish communities }the roles of biotic, abiotic, and spatial factors. Canadian Journal of Fisheries and Aquatic Sciences 58:157–170.
Jowett IG, Richardson J. 2003. Fish communities in New Zealand rivers and their relationships to environmentalvariables. New Zealand Journal of Marine and Freshwater Research 37: 347–366.
Lamoroux N, Olivier J-M, Persat H, Pouilly M, Souchon Y, Statzner B. 1999. Predicting community characteristicsfrom habitat conditions: fluvial fish and hydraulics. Freshwater Biology 42: 275–299.
Leland HV. 1995. Distribution of phytobenthos in the Yakima River basin, Washington, in relation to geology, landuse, and other environmental factors. Canadian Journal of Fisheries and Aquatic Sciences 52: 1108–1129.
Leland HV, Porter SD. 2000. Distribution of benthic algae in the upper Illinois River basin in relation to geology andland use. Freshwater Biology 44: 279–301.
Leland HV, Brown LR, Mueller DK. 2001. Distribution of algae in the San Joaquin River, California, in relation tonutrient supply, salinity and other environmental factors. Freshwater Biology 46: 1139–1167.
Ligon FK, Dietrich WE, Trush WJ. 1995. Downstream ecological effects of dams. A geomorphic perspective.BioScience 45: 183–192.
Lockwood LJ, Pimm SL. 1999. When does restoration succeed? In Ecological Assembly Rules: Perspectives, Advances,Retreats, Weiher E, Kelly PA (eds). Cambridge University Press: Cambridge; 363–392.
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA. 2000. Biotic invasions: causes, epidemiology,global consequences, and control. Ecological Applications 10: 689–710.
Magalhaes MF, Batalha DC, Collares-Pereira MJ. 2002. Gradients in stream fish assemblages across theMediterranean landscape: contributions of environmental factors and spatial structure. Freshwater Biology47: 1015–1031.
McCune B, Mefford MJ. 1999. PC-ORD. Multivariate Analysis of Ecological Data, Version 4. MjM Software Design:Gleneden Beach, OR, USA.
Moerke AH, Gerard KJ, Latimore JA, Hellenthal RA, Lamberti GA. 2004. Restoration of an Indiana, USA, stream:bridging the gap between basic and applied lotic ecology. Journal of the North American Benthological Society23: 647–660.
Murle U, Ortlepp J, Zahner M. 2003. Effects of experimental flooding on riverine morphology, structure and riparianvegetation: the River Spol, Swiss National Park. Aquatic Sciences 65: 191–198.
Newson MD. 2002. Geomorphological concepts and tools for sustainable river ecosystem management. AquaticConservation: Marine and Freshwater Ecosystems 12: 365–379.
Pan Y, Stevenson RJ, Hill BH, Herlihy AT, Collins GB. 1996. Using diatoms as indicators of ecological conditions inlotic systems: a regional assessment. Journal of the North American Benthological Society 15: 481–495.
Pan Y, Stevenson RJ, Hill BH, Kaufmann PR, Herlihy AT. 1999. Spatial patterns and ecological determinants ofbenthic algal assemblages in mid-Atlantic streams, USA. Journal of Phycology 35: 460–468.
LINKING GEOMORPHIC CHARACTER TO FLUVIAL BIODIVERSITY 287
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)

Pan Y, Stevenson RJ, Hill BH, Herlihy AT. 2000. Ecoregions and benthic diatom assemblages in Mid-AtlanticHighlands streams, USA. Journal of the North American Benthological Society 19: 518–540.
Parsons M, Thoms MC, Norris RH. 2003. Scales of macroinvertebrate distribution in relation to the hierarchicalorganization of river systems. Journal of the North American Benthological Society 22: 105–122.
Petts GE. 1980. Long-term consequences of upstream impoundment. Environmental Conservation 7: 325–332.Petts GE. 2000. A perspective on the abiotic processes sustaining the ecological integrity of running waters.Hydrobiologia 422/423: 15–27.
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC. 1997. The natural flowregime. A paradigm for river conservation and restoration. BioScience 47: 769–784.
Power ME, Dietrich WE, Finlay JC. 1996. Dams and downstream aquatic biodiversity: potential food webconsequences of hydrologic and geomorphic change. Environmental Management 20: 887–895.
Rhoads BL, Wilson D, Urban M, Herricks EE. 1999. Interaction between scientists and nonscientists in community-based watershed management: emergence of the concept of stream naturalization. Environmental Management24: 297–308.
Robinson CT, Uehlinger U. 2003. Using artificial floods for restoring river integrity. Aquatic Sciences 65: 181–182.Rosgen DL. 1994. A classification of natural rivers. Catena 22: 169–199.Schlosser IJ. 1982. Fish community structure and function along two habitat gradients in a headwater stream.Ecological Monographs 52: 395–414.
Sear DA. 1994. River restoration and geomorphology. Aquatic Conservation: Marine and Freshwater Ecosystems4: 169–177.
Stanford JA, Ward JV, Liss WJ, Frissell CA, Williams RN, Lichatowich JA, Coutant CC. 1996. A general protocol forrestoration of regulated rivers. Regulated Rivers: Research and Management 12: 391–413.
Sutherland AB, Meyer JL. Gardiner EP. 2002. Effects of land cover on sediment regime and fish assemblage structure infour southern Appalachian streams. Freshwater Biology 47: 1791–1805.
Thomson JR, Taylor MP, Fryirs KA, Brierley GJ. 2001. A geomorphological framework for river characterisation andhabitat assessment. Aquatic Conservation: Marine and Freshwater Ecosystems 11: 373–389.
Thomson JR, Taylor MP, Brierley GJ. 2004. Are River Styles ecologically meaningful? A test of the ecologicalsignificance of a geomorphic river characterization scheme. Aquatic Conservation: Marine and Freshwater Ecosystems14: 25–48.
Townsend CR, Doledec S, Norris R, Peacock K, Arbuckle C. 2003. The influence of scale and geography onrelationships between stream community composition and landscape variables: description and prediction.Freshwater Biology 48: 768–785.
Vinson MR, Hawkins CP. 1998. Biodiversity of stream insects: variation at local, basin, and regional scales. AnnualReview of Entomology 43: 271–293.
Walters DM, Leigh DS, Freeman MC, Freeman BJ, Pringle CM. 2003. Geomorphology and fish assemblages in aPiedmont river basin, U.S.A. Freshwater Biology 48: 1950–1970.
Wang L, Lyons J, Rasmussen P, Seelbach P, Simon T, Wiley M, Kanehl P, Baker E, Niemela S, Stewart PM. 2003.Watershed, reach, and riparian influences on stream fish assemblages in the Northern Lakes and Forest Ecoregion,USA. Canadian Journal of Fisheries and Aquatic Sciences 60: 491–505.
Ward JV, Tockner K, Uehlinger U, Malard F. 2001. Understanding natural patterns and processes in river corridors asthe basis for effective river restoration. Regulated Rivers: Research and Management 17: 311–323.
Willby NJ, Abernethy VJ, Demars BOL. 2000. Attribute-based classification of European hydrophytes and itsrelationship to habitat utilization. Freshwater Biology 43: 43–74.
Williams LR, Taylor CM, Warren ML, Clingenpeel JA. 2003. Environmental variability, historical contingency, andthe structure of regional fish and macroinvertebrate faunas in Ouachita Mountain stream systems. EnvironmentalBiology of Fishes 67: 203–216.
B.C. CHESSMAN ET AL.288
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16: 267–288 (2006)
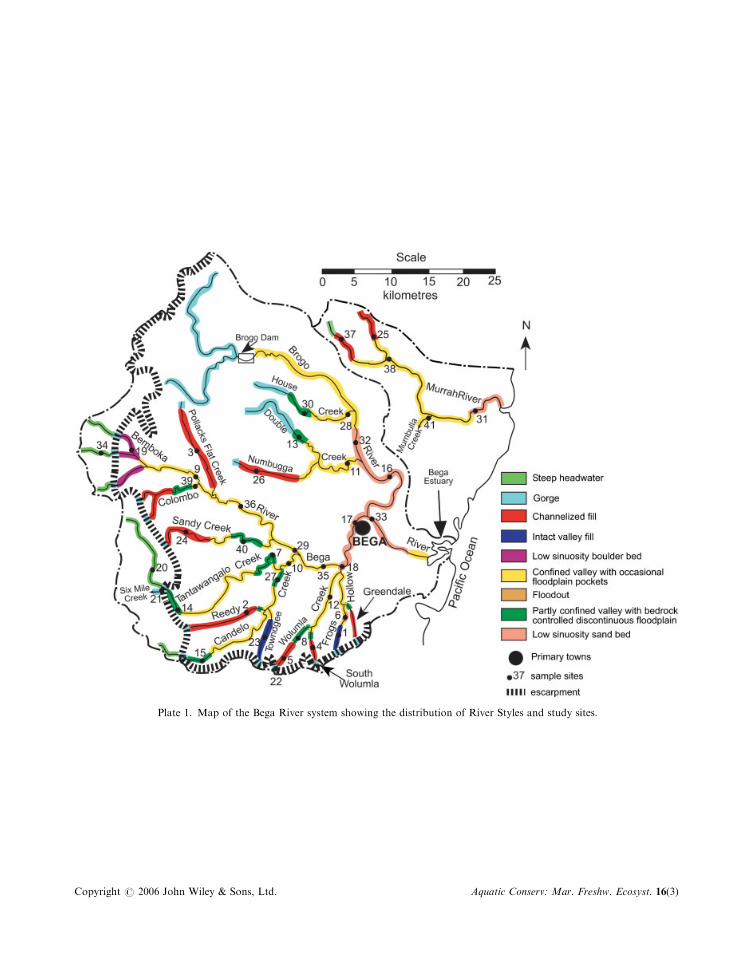
Plate 1. Map of the Bega River system showing the distribution of River Styles and study sites.
Copyright # 2006 John Wiley & Sons, Ltd. Aquatic Conserv: Mar. Freshw. Ecosyst. 16(3)





























