Embed Size (px)
Citation preview

LOCAL R E D U C T I O N OF S P I N D L E F I B E R B I R E F R I N G E N C E
I N L I V I N G N E P H R O T O M A S U T U R A L I S (LOEW)
S P E R M A T O C Y T E S I N D U C E D BY U L T R A V I O L E T
M I C R O B E A M I R R A D I A T I O N
A R T H U R F O R E R , Ph.D.
From the Department of Cytology, Dartmouth Medical School, Hanover, New Hampshire. Dr. Forer's present address is Biological Institute of the Carlsberg Foundation, Copenhagen, Denmark
A B S T R A C T
Irradiation of the mitotic spindle in living Nephrotoma suturalis (Loew) spermatocytes with an ultraviolet microbeam of controlled dose produced a localized area of reduced bire- fringence in the spindle fibers. The birefringence was reduced only at the site irradiated, and only on the spindle fibers irradiated. Areas of reduced birefringence, whether produced during metaphase or during anaphase, immediately began to move toward the pole in the direction of the chromosomal fiber, even though the associated chromosomes did not necessarily move poleward. Both the poleward and the chromosomal sides of the area of reduced birefringence on each chromosomal fiber moved poleward with about the same, constant, velocity. On the average, the areas of reduced birefringence moved poleward with about the same velocities as did the chromosomes during anaphase. The area of reduced birefringence was interpreted as a region in which most, though not necessarily all, of the previously oriented material was disoriented by the irradiation. The poleward movement of the areas of reduced birefringence indicates that the spindle fibers are not static, non- changeable structures. The poleward movement possibly represents the manner in which the birefringent spindle fibers normally become organized. All the experiments reported were on primary spermatocytes which completed the second meiotic division subsequent to the experimentation. Since both the irradiated and the control cells completed the two meiotic divisions, the movement and irradiation effects studied in the first division were non- degenerative.
I N T R O D U C T I O N
Though there has been much cytological work done on spindles (see E. B. Wilson, 1928; Schrader, 1953), the achromatic spindle fiber components were not conclusively shown to exist in living cells until 1953 (Inou6, 1953; see Schrader, 1953, and Mazia, 1961, for reviews). Using a highly sensitive polarizing microscope, Inou6 showed that the chromosomal fibers (from the chromosomes to the
poles), continuous fibers (from pole-to-pole), and asters are present in living cells, the birefringent fibers corresponding exactly to the appearance of the fibers seen in the best fixed and stained prepa- rations (Inou6, 1953; Schrader, 1953). Though spindle fibers do exist in living cells, and though many theories attribute to these fibers a major role in the movement of chromosomes during
95

mitosis (Schrader, 1953; Dietz, 1958; C)stergren et al., 1960; Mazia, 1961; Inou6, 1964; Roth, 1964), there is no direct evidence that the spindle fibers have such a role, and there is only scant knowledge of the physical and chemical nature of the fibers themselves. Most of the evidence for their role in chromosome movement is circumstantial, relying on the facts that kinetochores are necessary for normal anaphase movement to occur (Corn- man, 1944; Ris, 1949; Schrader, 1953; Mazia, 1961), and that during prometaphase movement kinetochores are often stretched in the direction of the movement (Hughes-Schrader, 1943, 1947; Cooper, 1951; Dietz, 1956; Bajer and Mol~-Bajer, 1963; Nicklas, 1963); also, various experimental agents which destroy spindle structure concomi- tantly stop chromosome movement (Pease, 1946; Cornman and Cornman, 1951; Inou~, 1964; Zimmerman and Marsland, 1964).
The best evidence that spindle fibers have an im- portant role in chromosome movement is of the last kind. Inou~ (1964) showed that the spindle fiber birefringence disappeared when the tem- perature was lowered during anaphase, and that the chromosomes stopped moving when the birefringence disappeared. Since the chromosomes did not resume movement until the birefringence reappeared, he concluded that apparently there is a direct relationship between birefringence and movement. Also, Inou~ (1952) showed that the chromosomes moved toward the periphery of the cell during colchicine-induced shortening of the spindle fibers, and that such movement stopped when the birefringence disappeared, and therefore that a shortening of the spindle fibers could cause the chromosomes to move. While these evidences are suggestive of the interpretations outlined, those interpretations are not the only ones possible, because the experimental agents do not affect the spindle alone. Temperature changes would affect all the cellular processes. And, since col- chicine is applied to the entire cell, colchicine too could affect components of the cell other than the birefringent spindle fibers. For example, colchicine is known to inhibit some dehydrogenases (Gal, 1938), and to alter chromosome structure (Eigsti, 1940), nucleolar structure (Herich, 1963), lyso- some and Golgi apparatus structure (Robbins and Gonatas, 1964), DNA synthesis (LaCour and Pelc, 1959; Hell and Cox, 1963; Sriramula, 1963), RNA synthesis (Creasy and Markiw, 1964), cell nucleotide content (Wang, Greenbaum, and Harkness, 1963), and muscle excitability (Le-
comte, 1949). Thus, while circumstantial evi- dences do implicate the spindle fibers, the pos- sibility of non-specific action by the experimental agents prevents unambiguous interpretation of the data.
The objection of non-specific action can be over- come if one uses an experimental tool which affects only the spindle, or a part of the spindle, without affecting other cellular processes. The ultraviolet microbeam is such a tool (see Zirkle, 1957; Smith, 1964; for reviews). When the ultraviolet micro- beam is focused to a small part of the spindle, ultraviolet light passes through only that part of the spindle, the cell membrane, and the cytoplasm above and below the spindle. The irradiation of the cell membrane and the cytoplasm near the spindle is unavoidable, but extra-spindle irradiation serves as a control for effects due to cytoplasmic and membrane irradiation. Thus, parts of the spindle can be selectively irradiated, and changes in func- tion specifically due to irradiation of those spindle parts can be measured. Direct information regard- ing the role of the spindle fibers in chromosome movement can be obtained, therefore, by irradi- ating spindle fibers with a microbeam, while following the spindle fiber birefringence with a sensitive polarizing microscope, by following chro- mosome movement before and after irradiation, and by comparing chromosome movement in these irradiated cells with that in non-irradiated cells, and with that in cells irradiated in extra-spindle areas. Such experiments have been performed (Forer, 1964), and this paper is the first of two papers reporting the results.
This paper describes the behavior of the areas of reduced birefringence which are produced by ultraviolet microbeam irradiation; some implica- tions of this behavior to the nature of normal, non-irradiated spindle fibers are considered in the discussion. Chromosome movements in these ir- radiated cells will be described in detail in a sub- sequent report (Forer, 1964, and manuscript in preparation).
M A T E R I A L S AND METHODS
Crane flies (Nephrotoma suturalis, Loew) 1 of all stages were maintained in the laboratory using the method
1 I would like to thank Dr. George W. Byers of the University of Kansas, Lawrence, Kansas, for identi- fying the species. The stocks derive from a single female caught by Dr. P. R. Dietz in Durham, North Carolina in the spring of 1961.
96 THE JOURNAL OF CELL BIOLOGY • VOLUME ~5, 1965

i
i
Strain-free ~ k ) condenser ~
To irradiate • ' To focus •
iI
I i
i
i i !
! !
I !
<
Image
Analyzer
Compensator
Rectified objective
Stage Reflecting condenser
Cell containing hexylene glycol and water
UV mirror
Polarizer
Filters
!
!
t !
t I
Lamp housing I
t ~ _ _ _ HBO- 200 light source
of Dietz, (1956 ; and personal communication, 1961 ).2 Last-instar larvae of the proper stage were chosen, surface-sterilized with 70 per cent ethanol, and then covered with Kel-F 10 fluorocarbon oil (Minnesota Mining and Mfg. Co., St. Paul, Minnesota). Each testis was dissected out under KeI-F 10 and smeared onto a clean quartz coverslip (A. D. Jones, Cam- bridge, Massachusetts). 70 # thick Fluorglas (Com- mercial Plastics, New York) spacers were placed on the quartz, a clean glass coverslip was placed on top of that, and the preparation was partially sealed with dentist's wax (Conger, 1960). Using this method, the cells completed the 2 meiotic divisions (prophase I to telophase II) in over /24 of the preparations.
Chromosome behavior in normal cells will be described elsewhere (Forer, manuscript in prepara- tion); it is similar to that described by Dietz (1956, 1959, 1963) for other species of crane flies. The three autosomal bivalents (Fig. 2 A1 and Fig. 2 B2) divide at anaphase, and the daughter dyads move to the poles (Fig. 13 A1) while the 2 unpaired sex chromosome univalents (Fig. 13 A2) remain at the equator. The
2 The method for maintaining a laboratory culture of N. suturalis is described in detail elsewhere (Forer, 1964). Other methods are given in Laughlin (1958) and Stich (1963). A detailed description of the tech- nique for making living cell preparations is given elsewhere (Forer, 1964).
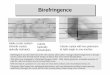
FIGURE 1 A schematic diagram of the polariz- ing microscope-ultraviolet microbeam ar- rangement. For simplicity, the substage front- surface mirror between the focusing cell and the reflecting condenser was omitted from the diagram.
univalents do not move poleward until the autosomes have neared the pole. The individual chromosomal spindle fibers are clearly visible in metaphase and anaphase with a sensitive polarizing microscope (e.g., Fig. 4 A2 and Fig. 13 A 1).
Phase contrast microscope observations were made with a Carl Zeiss phase contrast microscope (Model KF124-202), using the 40 X, 0.65 na ob- jective.
Polarizing microscope observations were made with an American Optical Company (New York) Super- Bin Polarizing Microscope with rectified optics (Inou6 and Hyde, 1957; Shurcliff, 1962, pp. 154- 155), 3 or with a Model P-42 polarizing microscope 4 modified such that a 20 X, 0.5 na straln-free objec- tive (Swann and Mitchison, 1950; Inou6, 1961) was used as a condenser, a 43 X, 0.66 na rectified objec- tive was used as the objective, a 17 m# retardation compensator (E. Leitz, New York; see Inou6, 1961) was inserted between the objective and the analyzer, and Polaroid sheets (HN-22 from the Polaroid Cor- poration, Cambridge, Massachusetts) were used as analyzer and polarizer. An Osram HBO-200 high-
3 Made available through the courtesy of the Marine Biological Laboratories, Woods Hole, Massachusetts, and Dr. Shinya Inou6. 4 Loaned by the American Optical Company to Dr. Shinya Inou6.
ARTHUR FORER Spindle Fiber Birefringence 97

pressure mercury arc was used as the light source for observation, and wavelengths other than the mercury green line (546 m/z) were removed from the beam by a combination of Corning glass filters (No. 4600 and No. 3387, from Corning Glass Works, Corning, New York), and an interference filter with peak transmission of 70 per cent at 546 m# (Baird- Atomic, Cambridge, Massachusetts).
Photographs were taken with an AO Spencer No. 668 35 mm camera back with compensator lens and shutter, using KB-17 film ( A D O X Fotowerke, Frankfurt, Germany). Photographs of living ceils were taken at various time intervals, distance measure- ments were made from positive prints at a final magnification of 1000, and these measurements were used to make graphs of position versus time. Distances were measured from one pole chosen as a reference point, and were measured in the pole-to-pole direc- tion.
The microbeam irradiations were from the con- denser side, using one microscope for both observa- tion and irradiation. (A similar system is described by Inou~, (1964).) For the irradiations the substage condenser was replaced by an American Optical 50 X, 0.56 na reflecting lens, and a small a luminum front-surface mirror (2.0 m m X 0.2 ram) was inserted into the system such that the light from the ultra- violet source was reflected from it and into the reflect- ing lens (Fig. 1)) Portions of the small mirror were masked by painting with India ink; in Fig. 4 A3, for example, the bright area (UV) is visible light re- flecting from the unpainted portion of the mirror and the dark area surrounding it is the painted por- tion.
The ultraviolet source was a General Electric
5 Such a system has been suggested by Zamenhof (1943) and used by Inou5 (1964).
Explanation of Figures
B, bivalent U, univalent CF, chromosomal fiber UV, image of the ultraviolet D, dyad irradiation source
Figs. ~ A1, ~ C1, l0 B~, and 10 B4 were photographed through a phase contrast micro- scope. The chromosomes appear dark against the non-granular spindle area (Fig. ~ A1- arrows).
The other photographs (Figs. ~, 4, 6, 8, 10, l l , 13, 15) were taken through a polarizing microscope. The birefringent spindle fibers (e.g., the chromosomal fibers labeled CF in Fig. 4 A~, and Fig. 13 A1) appear bright or dark against the background, depending on whether there is additive or subtractive compensation, respectively (Swann and Mitchi- son, 1950; Inou~ and Dan, 1951; Inou~, 1961). In the polarizing microscope the bivalents and dyads have low contrast (Figs. ~ Be; 4 A~2; 13 A1; labeled), and they are easiest seen as the termination of the chromosomal fibers. (The kinetochores are, by definition, the chromosomal position at which the chromosomal spindle fibers terminate.)
All photographs are printed at X 1000. The scale in the lower right corner is: 9.5~ in Figs. 13 and 15; 10~ in Figs. ~, 6, 8, 10 and 11; and 10.5~ in Fig. ~.
Figs. 3, 5, 7, 9, 1~, 14, and 16 are graphs of distance from the pole on the irradiated side (ordinate), versus time (abscissa), the times being plotted with respect to the time at which tile cell was irradiated (UV). In the schematic diagram at the top of each graph, the solid black area represents the area of reduced birefringence, a black line represents a chromosomal fiber, an ellipse represents a bivalent, the univalents are not shown, a half- ellipse represents a dyad, the pole on the irradiated side is labeled Pb and the areas of reduced birefringence, the kinetochores, and the opposite pole are labeled with the geo- metrical objects by which they are represented in the graphs. The geometrical objects which represent the kinetochores are closed (circles, and triangles), and those which repre- sent the areas of reduced birefringence and the pole on the non-irradiated side are open (circles, and squares). In the graphs, pole P1 is a horizontal straight line. The points repre- senting the area of reduced birefringence distances from/)1 are connected by a solid curve, as are the points representing the distances from P1 of the other pole; the points represent- ing the distances of the kinetochores from P1 are connected by a dashed curve.
The vertical arrow in Figs. 3, 5, 9, 1~, 14, and 16 represents the time of dyad separation. The position of the focused ultraviolet source measured from a picture taken 1.0 to 0.5
minutes before the irradiation is plotted on the graphs at the time of irradiation (UV).
98 THE JOURNAL OF CELL BIOLOGY • VOLUME ~5, 1965

AH-4 lamp with its glass cover removed. The meas- ured relative line energies were as follows:
254 m/z 265 mg 280 m# 297 m/,t 303 m/z 1.5 2 1 2 3.5
(See also Baum and Dunkelman, 1950.) The rationale and the method used to focus the
mierobeam are discussed in detail in the Appendix. Describing the procedure briefly, the ultraviolet wavelengths were removed with a filter, a hexylene glyeol~ontaining cell was inserted between the small mirror and the reflecting lens, the reflected visible image of the mirror was focused onto the
FIGURE ~, Cell 68j~7,C. Irradiated during metaphase. A1 : each of the three autosomal bivalents is labeled with an arrow. A~, AS: the position to be irradiated (on the chromosomal fiber of the left bivalent) is indicated by a bracket. A4: The ultraviolet source is labeled UV. B1, B~, BS, C~: The position of the area of reduced birefringence (on the chromosomal fiber of the left bivalent) is indicated by a bracket. There is weak birefringence inside the area of reduced birefringence. B~: the three autosomal bivalents are labeled with arrows, which correspond to the arrows in A1.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, --11 A~, --7 AS, --5 A4, --0.5 B1, +~ B~, +~.5 BS, +6 B4, +7 C1, +10 C~, +14.5 C8, +18.5 C4, +19.5
The area of reduced birefringence moved to the pole, and did not displaoe the pole when it reached the pole.
ARTHUR FORER Spindle Fiber Birefrlngence 99

specimen, and the filter and the hexylene glycol were removed before the irradiation (Fig. 1). The focus was corrected for 275 m#.
The ultraviolet output from the AH-4 lamp was monitored with a General Electric PV-10 ultraviolet sensitive photovoltaic cell (Jagger, 1961) used in conjunction with filters and a microvoltmeter (Leeds and Northrup, Philadelphia, Pennsylvania). The irradiation times were controlled, and were adjusted (between 14 and 20 seconds) to give a constant ir- radiation dose (energy/area). (The AH-4 output did not vary much in the first 100 hours of use.)
The total ultraviolet energy incident upon the cell was measured by placing the circular photovoltaic cell on the stage of the microscope and defoeusing the reflecting lens such that the ultraviolet beam covered 0.9 of the surface of the photovoltaic cell (Uretz, 1962). The measurements indicate that the ultraviolet energies at the focus point in air were of the order of 10 ergs//~ 2. This heterochromatic ultra- violet dose was used in all irradiation experiments.
The effective wavelengths for the irradiation effects described are less than 320 mu. This was determined by prolonged irradiation through a filter which had zero transmission for wavelengths less than 320 m#.
R E S U L T S
Ul t raviole t mic robeam i r radia t ion of spindle fibers in living Nephrotoma suturalis (Loew) pr imary spermatocytes produced areas of reduced bire- fringence. Each discrete area of reduced bire- fringence was abou t the same size and shape as the image of the i r radia t ion source aper ture (Figs. 4, 6, 8, I0, 13, and 15), and the chromosomal fiber birefringence on bo th the poleward and chromo- somal sides of the area remained essentially un- changed by the i r radia t ion (Figs. 2, 6, 8, 10, 13, and 15). Each area of reduced birefr ingence re- mained localized on the i r radia ted fibers; the affected area did not expand to include non-ir- rad ia ted regions. I r rad ia t ion with the same dose outside the spindle region had no effect on spindle birefringence. The cont inuous fiber hirefr ingence is very weak and ha rd to detect at the meiotic stages when spindle fibers were i rradiated (Forer, manu- script in preparat ion) , so the experiments are concerned mainly with areas of reduced bire- fringence on chromosomal fibers; however, when
::L c:
o .u'+ E5
I0o
20-
50-
40 -20
PI
............. ~ ~ I O0 i •
e-e+e ' e e . . . . . . . . . . ~ . . e -e . , e . . . . . . . . . . ~ . . . . . v e 4 . . . . e . . . . , o
D
n i I i ;0 -I0 +I0 +20 + UV
Time in minutes I~GURE 3 The graph is for tile cell photographed in Fig. 2. The distances from pole Px of both sides of the area of reduced birefringence are plotted. Both sides of the area moved poleward with about the same, constant, velocity while the associated chromosome remained at the equator. (For simplicity, the kinetochore positions of only the left bivalent are plotted; the other two bivalents acted in the same manner.)
100 TIlE JOURNAL OF CELL BIOLOGY • VOLUME 25, 1965

II 1
::L I0 .=_
o o
FIGURE 4 Cell 63h~ft,1. Irradiated during metaphase. Aft: The position to be irradiated (on the chromo- somal fiber labeled CF) is indicated by a bracket. A3: The ultraviolet source is labeled UV. A4, A5, A6: The position of the area of reduced birefringence (on the middle and left chromosomal fibers) is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, --8 Aft, --5.5 AS, -0 .5 A4, "1-~ A5, +5.5 A6, +10
The area of reduced birefringence moved to the pole, and did not displace the pole.
°w'Q qP v~l q~Oo'°'tlJ~e° • " Q i . . . . . . •
E O
-~ 2 0 - a
30" t ! +1~) "1-~0 - ] 0 I +~'0
UV T i m e in m inu tes
FIGVRE 5 The graph is for the cell photo- graphed in Fig. 4. The area of reduced bire- fringence distances from /)1 which are shown are of the chromosomal side of the area only. The area of reduced birefringence moved pole- ward with a constant velocity, while the as- sociated chromosome remained at the equator. (The other bivalents acted in the same manner.)
cont inuous fibers were discernible at the t ime of i rradiat ion, they too had reduced birefringence in the area i r radiated (Figs. 2, 6, 8, and 15). T h e amoun t of birefringence remain ing in the irradi- ated areas was not the same in the different ex- per iments (e.g., Figs. 2, 6, and 8), even though the incident i r radiat ion dose (energy/area) was the same in all experiments. The area of reduced birefringence on any given chromosomal fiber was not necessarily perpendicular to the long axis of tha t fiber; its angle (with respect to the fiber axis) depended on the angle of the focused i r radiat ion source aperture, which was different in different experiments (Figs. 2, 6, 10, and 13).
The area of reduced birefringence could be seen in the polarizing microscope, bu t could not be dist inguished in the phase contrast microscope as being ei ther l ighter or darker than its surroundings (Figs. 2 and 10).
The first post i r radiat ion observations were made at 20 seconds after the i r radia t ion was finished; at this t ime the area of reduced birefr ingence was visible (Forer, 1964). Therefore, if there is a t ime lag between i r radia t ion and format ion of the area of reduced birefringence, this figure (20 seconds) is the upper l imit for such a t ime lag.
Each area of reduced birefr ingence moved toward the pole immedia te ly after it was formed,
ARTHUR FoRErt Spindle Fiber Birefringence 101

F m u a ~ 6 Cell 68j8,C. Irradiated during metaphase. A1, A~: The position to be irradiated (on the chromosomal fibers of the two left bivalents) is indicated by a bracket. AS: The ultraviolet source is labeled UV. A4, B1, B~: The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, - 5 . 5 A~, -4 .5 AS, -0 .5 A4, +5~ B1, +2.5 B~2, +8.5 BS, +9.5 B4, +18.5
The area of reduced birefringence moved to the pole, and did not displace the pole or tim adjacent not- affected fibers.
and it cont inued to move unt i l it reached the pole. 6 This poleward movemen t occurred wi thout excep- t ion in each of the chromosomal fiber areas of re- duced birefr ingence studied, even though the chromosomes associated with the same chromo- somal fibers did not necessarily move poleward at tha t t ime (Figs. 2, 4, 6, 8, and 10), and even though the cell was in metaphase at the t ime of i r radia t ion (Figs. 2, 4, 6, and 8). The relat ion between move- men t of the areas of reduced birefringence and movement of the chromosomes will be discussed in detail in a subsequent paper.
6"Movement" is defined operationally: as time proceeds, the distance from the area of reduced bire- fringence to the pole changes. The area "moves" toward the poles.
Both sides of the area of reduced birefr ingence on each chromosomal spindle fiber moved pole- ward in the direct ion of the chromosomal fiber, and bo th sides of the area moved toward the pole with about the same velocity (Figs. 2, 8, 13, and 15). In some cells the reduced birefringence on different chromosomal fibers moved poleward with different velocities, and when this happened , the area of reduced birefringence changed in shape or in angle relative to the spindle axis as the area moved toward the pole (Figs. 10, 11, and 15).
The areas did not deform the adjacent unaf- fected fibers as they moved toward the pole (Figs. 2, 6, 8, 13, and 15), nor did they push away the asters once they reached the pole (Figs. 2, 6, 8, 11, 13, and 15). Rather , the size of each area gradual ly
102 TIlE JOURNAL OF CELL BIOLOGY • VOLT3ME 25, 1965

C
q~
P1
I0
20-
50
• ~ , ~ 0 P1
• ,.,-.,-.-o~,~ n
44 ~ ~ . . . . . at ... . . om --'*°
ao~.O"" llo "'' '°e ..... * ..... "o-, . . . . . . ~ . . . . . . . . - - " " 0
40 l ,u02 I -I0 I +I0 +50
UV Time in minutes
FIGURE 7 The graph is for the cell photo- graphed in Fig. 6. The area of reduced bire- fringence distances from Px which are shown are of the chromosomal side of the area associated with the left bivalent. The area of reduced blre- fringence moved poleward with a constant velocity, while the associated chromosome re- mained at the equator. (The other chromosomes, and area of reduced birefringence, acted in the same manner.)
FIGURE 8 Cell 68h13,B. Irradiated during metaphase. A1, A~: The position to be irradiated (on tile chromosomal fibers of the two left bivalents) is indicated by a bracket. A3: The ultraviolet source is labeled UV. A4, B1, B~, B3, B4 : The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, --6.5 AS, --5 AS, -0 .5 A4, +2 A5, +3 B1, +4.5 B~, +6 BS, +11 B4, +16 B5, +17
The area of reduced birefringence moved to the pole, and did not displace the pole.
decreased after the area reached the pole until such an area was no longer distinguishable.
The primary interest was in effects on the chro- mosomal fibers of the autosomes. Areas of reduced
birefringence on 99 such chromosomal fibers were studied; 39 were in cells in metaphase at the time of irradiation, and 60 were in cells in anaphase at the time of irradiation. (Figs. 2, 4, 6, and 8 show
ARTrma FOREa Spindle Fiber Birefrlngenee 103

P I
I0
::L c
20 - c o
~5
5 0 -
4 0 -
- 2 0
o--QO . . . . . . . " FIGURE 9 The graph is for the cell photographed in Fig. 8. The area of re-
. . .o . . .o . .m. o . . o . . . u , .o . , ~ - . . . . . , m , o . . . ~ i i .
I l I I - i o i + 1 o +zo
UV
I +30
duced birefringence distances from P1 which are plotted are of the chromo- somal side of the area on the left bi- valent's chromosomal fiber. The area of reduced birefringence moved to the pole with a constant velocity while the as- sociated chromosome remained at the equator. (The other chromosomes, and area of reduced birefringence, acted in the s a m e m a n n e r . )
Time in m inu tes
areas of reduced birefr ingence in metaphase cells, and Figs. 10, 11, 13, and 15 show areas of reduced birefr ingence in anaphase cells.) Areas of reduced birefr ingence on chromosomal fibers of univalents also moved toward the pole immedia te ly after being formed.
There was a change in the birefr ingence of bo th the areas of reduced birefringence, and the chro- mosomal fibers immedia te ly adjacent to the areas of reduced birefringence, as the areas moved to- ward the pole. T he birefringence inside each area of reduced birefringence often increased in mag- ni tude as the area moved to the poles (Figs. 2, 4, and 13). The birefringence of the chromosomal fiber immedia te ly adjacent to each area decreased in magni tude as the area moved to the pole. (Tha t the last s ta tement must be true can be seen from the following considerations. Since an area of re- duced birefringence moves toward the pole, wi th time, the position of the chromosomal fiber ad- j acen t to the area moves toward the pole, wi th time. After the area of reduced birefringence has reached the pole, the consecutive positions along the fiber, from the initial position of the area of reduced birefringence to the pole, represent the t ime course of the position of the fiber adjacent to the area of reduced birefringence. Since in these
cells the spindle fiber birefringence is strong near the kinetochores and weaker toward the poles (Figs. 2, 6, and 13), and since after the area reached the pole the i rradiated chromosomal fiber was indist inguishable f rom non- i r radia ted fibers, the birefringence of the chromosomal fiber adja- cent to the area of reduced birefringence decreased in magni tude as the area moved to the pole.)
The distance between the area of reduced bire- fr ingence and the spindle pole on the i r radia ted side was measured on each pho tograph in the series of photographs of each cell. In favorable cases the distances f rom the pole of bo th sides of the area (of reduced birefringence) were measured (Figs. 3 and 16), bu t in general the distances from the pole of only the side of the area closest to the chromo- somes were measured. The chromosomal side of an area could be followed unt i l it was 3 to 4 /~ from the pole, bu t because of the increasing birefrin- gence inside the area, because the chromosomal fibers have less birefr ingence near the poles, and because the chromosomal fibers converge toward the poles, it was difficult to dist inguish the area after this point.
The poleward velocity of the area of reduced birefr ingence on each chromosomal fiber was cal- culated from the distance measurements . The dis-
104 THE JOURNAL OF CELL BIOLOGY ' VOLUME ~5, 1965

tance of each area from the pole was plotted with respect to the time elapsed since the fiber was irradiated, and the slope of this curve is the pole- ward velocity.
Each area of reduced birefringence moved pole- ward with a constant velocity (Figs. 3, 5, 7, 9, 12, 14, and 16), but different areas did not necessarily move with the same velocity, even in the same cell (Figs. 10, 11, and 15). The distribution of the pole- ward velocities of the areas (of reduced birefrin- gence) is shown in Fig. 17 A, shaded, for cells ir- radiated before anaphase, and in Fig. 17 B, shaded, for cells irradiated during anaphase. The anaphase poleward velocities of chromosomes in non-ir- radiated (control) cells adjacent to the irradiated cells is the non-shaded area of the same figures. (The chromosome velocities were measured in the
same way as were the area of reduced birefringence velocities.) These comparisons show that after irradiation either in metaphase or in anaphase, the poleward velocities of the areas of reduced bire- fringence are, on the average, about the same as the anaphase poleward velocities of chromo-
somes in non-irradiated control cells. Before mak-
ing statements regarding the equivalence of area
of reduced birefringence velocities and chromo-
some velocities, the velocities of individual areas of
reduced birefringence should be compared directly
with the velocities of the chromosomes associated
with the same fiber, or with the velocities of indi-
vidual control chromosomes. When this is done
(Forer, manuscript in preparation), it is seen that
the velocities of the areas are often different from
FIGURE 10 Cell 63L7,4. Irradiated during anaphase. A1, A~: The position to be irradiated (across the entire half-spindle) is indicated by a bracket. A3: The ultraviolet source is labeled UV. A4, BI: The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, - 9 A~, --7 A3, --0.5 A4, +1.5 B1, +3.5 B~, +4.5 B3, +16.5 B4, +19
The area of reduced birefringence moved to the pole and did not displace the pole. In A4 the chromo- somal side of the area of reduced birefringence makes a straight line; in B1 the chromosomal side of the area of reduced birefringence does not make a straight line, because the areas on different fibers moved poleward at different rates.
ARTHUR FORER Spindle Fiber Birefringenee 105

FIGURE 11 Cell 63hS,A. Irradiated during anaphase. A1, A~: The position to be irradiated (across the entire half-spindle) is indicated by a bracket. A4: The ultraviolet source is labeled UV. B1, B% B4: The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, --8 A~, --5.5 .4.8, --4.5 A4, - 0 .5 AS, + 4 B1, +4.5 B~, +5.5 B8, +13 B4, +16.5 B5, +~1
The area of reduced birefringencc moved to the pole and did not displace the pole. In A5 and B1, the chromosomal side of the area of reduced birefringence makes a straight line; in B3, and B4 the area of reduced birefringencc does not make a straight line because the areas on different fibers moved poleward at different rates.
the velocities of the chromosomes, by as much as a factor of 3. These results will be given in detail in a subsequent report.
In the experiments reported here, both the ir- radia ted cells and the non- i r radia ted control cells completed the two meiotic divisions. ( In five cases the da ta include i r radiated cells which did not complete division II . In these cases there was indicat ion of prepara t ion for division II such as aster formation, or spindle formation, and the data were not in variance with other cases where the i r radiated cells completed the second division.) I t is necessary to follow the cells th rough the com- pleted second division to ensure tha t the move- ments observed in non- i r radia ted cells were normal and not degenerat ive, and to ensure tha t the differ- ences between the i r radiated and the control cells
were not due to lethal or semilethal effects of the i r radia t ion (e.g., Uretz and Zirkle, 1955; Izutsu, 1961; Bajer and Mol~-Bajer, 1961). Since the ceils completed a second division, the movements and i r radiat ion effects studied in the first division were non-degenerat ive.
D I S C U S S I O N
The birefringence of the i r radiated par t of a spindle can be greatly reduced without changing the bire- fringence of other parts of the spindle. Though this is not the first t ime such results have been reported (see Inou~, 1964), most ul traviolet mic robeam workers report tha t the entire spindle structure disappears after i rradiat ion, even after i r radiat ion of the cytoplasm (Zirkle, 1957; Smith, 1964). The difference between the findings of those workers
106 THE JOUnNAL OF CELL BIOLOGY • VOLUME ~5, 1965

and the localized disappearance reported here is most likely due to a difference in dose rather than a peculiarity of crane fly spermatocytes, for the following reasons: (a) While cytoplasmic irradi- ation of Tradescantia stamen hair cells could cause the entire spindle structure to disappear, Otrosh- chenko and Sakharov (1964) showed that this did not happen at lower doses, and that depending on the dose used the spindle might or might not disappear. (b) Inou~'s results on Haemanthus endo- sperm (Inou6, 1964) are similar to those on Nephro- toma, yet it had been reported by other workers that cytoplasmic irradiation destroyed the entire Haemanthus spindle (Zirkle, Uretz, and Haynes, 1960). (c) The entire Nephrotoma spindle birefrin- gence will disappear if the dose (energy/area) used for irradiation of spindle fibers is three times that used here (Forer, 1964).
Since the birefringence was changed only at the site of the microbeam irradiation, experiments were performed to test the role of the birefringent spindle fibers in chromosome movement. These results will be given in detail elsewhere. In this paper we will consider only the movement be-
havior of the areas of reduced birefringence, and the implications of this behavior toward the nature of normal (non-irradiated) birefringent spindle fibers. Since such inferences depend greatly on the exact nature of the area of reduced birefringence, I will first discuss the nature of the areas of reduced birefringence, and then discuss some implications of the behavior of these areas to the properties of non-irradiated spindle fibers.
Nature of the Areas of Reduced Birefringence
Material is birefringent when the refractive index for light whose electric vector is polarized in a certain direction is different from the refractive index for light polarized in a perpendicular di- rection. In an area of reduced birefringence the refractive indices are roughly the same for all ori- entations of the electric vector. The ultraviolet mi- crobeam could cause the birefringence to be re- duced (a) by causing the molecules which were previously oriented to become disoriented, or (b) by changing the intrinsic birefringence of the oriented molecules without greatly disorienting them, either through cross-linking or denaturation,
P1
IO-
c
2o-
50 -
[]
I .... *"i~* ......... 6""
---*---*-----~--°"°- ~--Q • . . % ° • " * ~ ~ ' o " ~ " ' • .......
• 0 " ' " - . . • .
-10 - 2 0 + I0 + 0 +50 +40 UV
Time in minutes
FIGURE le The graph is for the cell photographed in Fig. l l . The area of reduced birefringence distances from /)1 which are plotted are of the chromosomal side of the area on the right dyad's chromosomal fiber. The area of reduced birefringence moved to the pole with a constant velocity even though the two associated dyads temporarily stopped moving poleward.
ARTHUR FORER Spindle Fiber Birefringence 107

FmURE 13 Cell 63j10,~. Irradiated during anaphase. A~: The position to be irradiated (on the chromo- somal fibers of the two right dyads and the univalents, U) is indicated by a bracket. A3: The ultraviolet source is labeled UV. A4, B3: The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to tile time at wtfich the celt was irradiated:
A1, --6 A~, --5 A3, --0.5 A4, "4"~ AS, -4-3 B1, -4.4.5.5 B~, -4.8 B3, ,4.10.5 B4, +1~.5 B5, .4.15.5
Tile area of reduced bircfringence moved to the pole, and did not displace the pole.
or (c) by both denaturation and disorientation, or (d) if the spindle birefringence is mostly form- birefringence, by changing the relative volumes of oriented versus non-oriented material. Informa- tion to distinguish between these possibilities might be obtained by light microscope and electron microscope studies of fixed and stained areas of reduced birefringence, by micromanipulator stud- ies of such areas compared with non-irradiated spindle fibers, by radioactive labeling of spindles
and determining if areas of reduced birefringence
lose label, or by following the movement of gran-
ules found in such areas; but such information is not available at the moment.
However, since the amount of birefringence in- side the area increased as the area moved toward
the pole, and since the area deformed neither the adjacent non-irradiated fibers nor the pole, the area of reduced birefringence is more likely ma- terial which is disoriented rather than material
cross-linked or denatured, for I would not expect cross-linked or denatured material to gain bire- fringence as it moved toward the pole, and I would expect cross-linked or denatured material to push
the pole away rather than be "disassembled" when it reached the pole.
Such an interpretation does not imply that all the structure or orientation is absent from an area of reduced birefringence, (a) because weak bire- fringence is often present inside the area after ir- radiation (Figs. 2 and 13), implying some remain- ing oriented material, and (b) because there could be oriented material in such an area which would not be detected in the polarizing microscope. This interpretation does imply that much or most of the
oriented structure was destroyed by the irradi- ation. Thus, I interpret the area of reduced bire-
fringence to be an area in which most, though not necessarily all, of the previously oriented material has been disoriented by the irradiation.
Interpretations of the Behavior of the Areas of
Reduced Birefringenee
In Nephrotoma spermatocytes the birefringence poleward from the area of reduced birefringence was unaffected by the irradiation which produced the area of reduced birefringence (this report). Inou~ (1964) reported that the birefringence pole-
108 THE JOURNAL OF CELL BIOLOGY • VOLUME ~5, 1965

ward (distal) from the irradiation site disappeared after ultraviolet microbeam irradiation of chromo- somal spindle fibers in Haemanthus endosperm. He attributed the poleward disappearance of bire- fringence to the absence of an "organizing center" poleward from the irradiated site. If the poleward birefringence is unaffected in Nephrotoma because of the influence of the centriole acting as an "or i - enting center," the poleward side of the area of reduced birefringence must be given "disorient- ing" attributes as well, for the fiber between the area of reduced birefringence and the pole de- creases in length as the area moves toward the pole. It is surprising that the centriole would have an influence in maintaining the birefringence, for both chromosome movement and spindle fiber birefringence are normal even when there is no centriole at the pole of Nephrotoma suturalis sperma- tocytes (Forer, 1964; this was originally demon-
::k , c
c o
C3
I0
20-
50-
: . - : - : . . . . . . . . . . . . . . -
: : iL~: : : : : : : : - - tw- .m . - L * - ~ -_.,
leA..
- , , [ : ~ j -
4.0 I t I - I 0 | + 1 0 -I-20
UV
T i m e in minutes
FmuRB 14 Tile graph is for tile cell photographed in Fig. 13. The area of reduced birefringence distances from P1 which are plotted are of the chromosomal side of the area on the middle dyad's chromosomal fiber. The kinetochore positions of the members of both the middle dyad pair and the left (not-irradiated) dyad pair are plotted. The area of reduced birefringence moved to the pole with constant velocity even though the poleward movement of the two associated dyads temporarily stopped. Both dyads in the left (not-ir- radiated) dyad pair moved normally.
strated by Dietz, 1959, 1963 for spermatocytes of the crane fly, Pales crocata). The role of the centriole in maintaining the birefringence poleward from the irradiated site could be directly tested by microbeam irradiation of chromosomal fibers in spermatocytes in which the centriole has been ex- perimentally dissociated from the pole.
The poleward movement of the area of reduced birefringence is defined operationally: as time proceeds, the distance from the area to the pole becomes smaller. Such behavior might mean that there is actual movement of the fiber material to the pole; i.e. that the birefringent material, and the non-birefringent material in the area of re- duced birefringence, both move to the pole, are broken down at the pole (i.e., are disoriented and made non-birefringent), and are then re-cycled, and that the area of reduced birefringence is just a marker in this continuously moving system. This is not the only possibility, however. For example, it is possible that without some accessory com- ponent the oriented spindle fiber material will re- vert to the disoriented state, and that it is this component and not the birefringent fiber material which moves to the pole; with this interpretation, the ultraviolet inactivates this accessory compo- nent, the ultraviolet inactivated component is un- able to cause orientation, disorientation ensues, and an area of reduced birefringence is formed. The area of reduced birefringence moves not when non-birefringent fiber material moves poleward, as in the previous interpretation, but rather when the non-functional accessory component moves poleward and displaces the functional component. As another alternative, it is also possible that no material moves poleward at all, but rather the area
of reduced birefringence movement indicates a
wave of organization which moves to the pole. Pease (1946) found that spindle fibers grew
from the chromosomes after removal from the high
pressure which had caused spindle breakdown.
While this might suggest that the area of reduced birefringence movement is an actual movement of
fiber material, the data presented in this paper do not rule out the other possibilities. However, if the poleward movement of the area of reduced
birefringence does indicate movement of the fiber material, this fiber material must change state, or
orientation, as it moves, for as described in the Results section, there are changes in the bire- fringence of both the area of reduced birefringence, and the chromosomal fiber adjacent to the area,
as the area moves to the pole.
AnTlt~J~, FORER Spindle Fiber Birefringence 109

FIGURE 15 Cell 63h~,1. Irradiated during anaphase. Ae: The position to be irradiated (on the chromo- somal fibers of the two left dyads) is indicated by a bracket. A3: The ultraviolet source is labeled UV. A4, B1, B3: The position of the area of reduced birefringence is indicated by a bracket.
The times of the photographs follow; they are given in minutes with respect to the time at which the cell was irradiated:
A1, --8 A~, --5.5 A3, --0.5 A4, -{-~ B1, +3 B~, +5 B3, -{-6 B4, -t-16
The area of reduced birefringence moved to the pole and did not displace the pole. The chromosomal side of the area of reduced birefringence is a straight line in A4, but is not straight in B1, B~, and B3 because the areas of reduced birefringence on the different chromosomal fibers moved poleward at different rates.
The poleward movement of the areas of reduced birefringence, then, represents a poleward move- men t of some material , or a wave of organization. Regardless of which of the interpretat ions is true, this indicates tha t even when the chromosomes do not move, the spindle fibers are not static, non- changeable structures, for they can drastically change their organizat ion (see Inoufi, 1959, 1964, for reviews). One question which does arise, how- ever, is whether this poleward movement , or wave, indicates a dynamic organizat ion which was pres- ent before the irradiat ion, or whe ther the fibers are really basically static, and tha t in changing the birefringence the i r radia t ion set in mot ion a proc- ess whereby the changed region is propagated to the pole. At present there are no da ta to distinguish between these two possibilities. I t is clear tha t in Nephrotoma the spindle fiber organizat ion changes even wi thout i rradiat ion, for the chromosomal
fibers gradually increase in width and birefrin- gence between nuclear m e m b r a n e breakdown and anaphase (Forer, 1964, and manuscr ip t in prepa- rat ion; also Dietz, 1963; and Inoufi's evidence for dynamic equil ibr ium, 1964), and thus tha t even before anaphase the organizat ion of the spindle fibers is not static. But it is not known whether the normal increase in width and birefringence is at all related to the movement of the area of re- duced birefringence, nor if the area movement is indeed induced by the irradiation. The existence of movement might be ascertained from the fol- lowing experiment: Ostergren, Mol~-Bajer, and Bajer (1960) hypothesized tha t the poleward movement of spindle inclusions (such as nucleoli, acentric chromosomes, g r a n u l e s , . . . ) was due to extra-spindle-fiber "pumps , " which acted on spindle fibers in the same way as on the inclusions. According to the hypothesis, the poleward move-
l l 0 THE JOURNAL OF CELL BIOLOGY • VOLUME ~5, 1965

PI
i O -
::t.. c :
2 o -
o
¢o
5 0 -
4 0
°.~-~----®
ar -0 -0 ,4PO 09"~ i l t ~ O. ~ • "0
• t P" I w • ~ $ ' Q " -0 ~ • •
O
~ 0 a [ i i i - - I 0 ÷ I 0 + 2 0 + 5 0
UV
Time in minutes
FmURE 16 The graph is for the cell photographed in Fig. 15. Tim distances from pole P1 of both sides of the area of reduced birefringence are plotted for the area on the middle dyad's chromosomal fiber. Both sides of the area of reduced birefringence moved poleward with about the same, constant velocity, even though the two associated dyads temporarily stopped moving.
men t of the area of reduced birefringence would be an actual movement of material , and would be due to the same force which causes the spindle in- clusions to move poleward. This can be directly tested by compar ing the movement of an area of reduced birefringence with movement of spindle inclusions in the same cell. I f the two always moved with the same, or a related velocity, this would imply tha t the poleward movement of the area of reduced birefringence was due to the same mechanism as the poleward movement of spindle inclusions. Since the spindle inclusions move pole- ward in all stages, such a result would lend cre- dence to the idea tha t the movement of the area indicates a phenomenon which occurs in all stages, and not one which is induced by the irradiation. (If the two did not agree, however, it would mean only tha t the reduced birefringence movement and the inclusion movement did not occur by the same mechanism, and with this result the exper iment would not give information whether or not a flow or wave in the spindle fibers always occurred.)
I in terpre t the poleward movement of the area of reduced birefr ingence to indicate a process
which occurs even wi thout induct ion by the irradi- ation, and which represents the way in which the birefr ingent spindle fibers are organized from disoriented material , in conjunct ion with an as yet morphological ly unidentif ied t ract ion element (Forer, 1964; and manuscr ip t in preparat ion) . Fur ther evidence in support of this hypothesis will be considered in a subsequent publication.
In conclusion, the results reported here confirm the findings of Inou~ (1964) tha t the birefringence of local parts of a spindle can be greatly reduced by ul traviolet mic robeam i r radia t ion wi thout affecting the birefringence of non- i r radia ted parts. The area of reduced birefr ingence is in terpre ted as an area in which most, though not necessarily all, of the previously oriented mater ia l has been disoriented by the i rradiat ion. O the r al ternat ives are not ruled out by the data, however.
The poleward movement of the areas of reduced birefringence which occurs in bo th metaphase and anaphase p robab ly represents a movement of material , e i ther of or iented molecules themselves, or of molecules necessary for orientat ion. I t is also possible tha t no such molecules move, bu t tha t a
AI~THUR FoaELt Spindle Fiber Birefringence l l l

60,
50
4O
E
z 30
20
I0
O i
0.0 0.5
70 A
H |
1.0 o.0
Veloc i ty / .dmin.
0.5 1.0
FIGURE 17 The shaded areas are the num- ber of autosomal chromosomal fibers on which an area of reduced birefringence was formed (ordinate), versus the poleward velocity of each such area (abscissa). Fig. 17 A, shaded, is for areas of reduced birefringence in cells ir- radiated before dyad separation (during late prometaphase and early metaphase), and Fig. 17 B, shaded, is for areas of reduced birefrin- gence in cells irradiated after dyad separation (during anaphase). The non-shaded areas are the number of dyads (ordinate) versus the poleward velocity of each such dyad (abscissa) for dyads in the ~lls used as controls for the Irradiation ,expernnents.
wave of organization is propagated along the
fiber. With either alternative the results indicate that the chromosomal fibers are not static, non- changeable structures.
The poleward movement of the areas of reduced birefringence is interpreted to indicate a movement which occurs all the time, and which indicates the manner in which the birefringent spindle fibers are
organized. An alternative interpretation is possible, in which such movement or propagation is in- duced by the irradiation.
I acknowledge with gratitude the counsel of Dr. Shinya Inour, under whose sponsorship this work was performed, and I wish to thank him for his generous loans of equipment, and his critical com- ments on this manuscript in various stages of its
preparation. I would also like to thank Dr. Roland Dietz for instruction on the laboratory care of crane flies, and for the gift of some Nephrotoma suturalis (Loew) larvae, from which my laboratory supply derives and Richard Markley for help with the photographs.
This investigation was supported in part by grants to Dr. Inou6 from the National Science Foundation (G-19487) and the National Cancer Institute, United States Public Health Service (CA 04552). Some of this work was carried out during the tenure of a Predoctoral Research Fellowship from the Division of General Medical Sciences, United States Public Health Service.
Portions of this paper were presented to Dartmouth College in partial fulfillment of the requirements for the degree of Doctor of Philosophy.
Received for publication, September 9, 1964.
B I B L I O G R A P H Y
BAJER, A., and MOL~;-BAjER, J., 1961, UV micro- beam irradiation of chromosomes during mitosis in endosperm, Exp. Cell Research, 25, 251.
BAJER, A., and MOL~-BAjER, J., 1963, Cine analysis of some aspects of mitosis in endosperm, in Cine-
micrography in Cell Biology, (G. G. Rose, editor), New York, Academic Press, Inc., 357-410.
BAUM, W. A., and DUNr,~LMAN, L., 1950, Ultraviolet radiation of the high pressure Xenon arc, J. Opt. Soc. Am., 40, 782.
112 THE JOURN2,1~ OF CELL BIOLOGY . VOLUME ~5, 1965

BRUblBERG, E. M., 1943, Colour microscopy in ultra- violet rays, Nature 152, 357.
CONGER, A. D., 1960, Dentists' sticky wax: a cover- sealing compound for temporary slides, Stain Technol., 35, 225.
COOPER, K. W., 1951, Compound sex chromosomes with anaphasic precocity in the male mecopteran, Boreus brumalis Fitch, J. Morphol., 89, 37.
CORNUAN, I., 1944, A summary of evidence in favor of the traction fiber in mitosis, Am. Naturalist, 78, 410.
CORNMAN, I., and CORNMAN, M. E., 1951, The action of podophyllin and its fractions on marine eggs, Ann. New York Acad. Sc., 51, 1443.
CREASEr, W. A., and M. E. MARKIW, 1964, Bio- chemical effects of the vinca alkaloids. II. A com- parison of the effects of colchicine, vinblastine and vincristine on the synthesis of ribonucleic acids in Ehrlich ascites carcinoma cells, Biochim. et Biophysica Acta 87, 601.
DmTZ, R., 1956, Die Spermatocytenteilungen der Tipuliden. II. Graphische Analyse der Chromo- somenbewegung wahrend der Prometaphase I i m Leben, Chromosoma, 8, 183.
Dmwz, R., 1958, Multiple Geschlechtschromosomen bei den Cypriden Ostracoden, ihre Evolution und ihr Teilungsverhalten, Chromosoma, 9, 359.
DmTZ, R., 1959, Centrosomenfreie Spindelpole in Tipuliden-Spermatocyten, Z. Naturforsch., 14 b, 749.
DIETZ, R., 1963, Polarisationsmikroskopische Befunde zur Chromosomeninduzierten Spindelbildung bei der Tipulide Pales crocata (Nematocera), Zool. Anz., 26, suppl., 131.
ErnsT1, O. J., 1940, Effects of colchicine upon the nuclear and cytoplasmic phases of cell division in the pollen tube, Genetics, 25, 116.
FORER, A., 1964, Evidence for two spindle fiber com- ponents: a study of chromosome movement in living crane fly (Nephrotoma suturalis) spermatocytes, using polarization microscopy and an ultraviolet microbeam, Doctoral Thesis, Dartmouth College.
GAL, M. E., 1938, Etude de l'~tction du cacodylate de soude et de la colchlcine sur diff~rentes dtshy- drogenases, Bull. Soc. Chim. Biol., 20, 1188.
HALL, C. E., 1953, Introduction to Electron Micro- scopy, New York, McGraw-Hill Book Co.
HARDY, A. C., and PERRXN, F. H., 1932, The Prin- ciples of Optics, New York, McGraw-Hill Book Co.
HELL, E., and Cox, D. G., 1963, Effects of colchicine and colchemid on synthesis of deoxyribonucleic acid in the skin of the guinea pig's ear in vitro, Nature, 197, 287.
HERICrt, R., 1963, Influence of colchicine on nucleoli, Caryologia, 16, 521.
HUGHEs-SeHRAD~R, S., 1943, Polarization, kineto- chore movements, and bivalent structure in the meiosis of male mantids, Biol. Bull., 85, 265.
HUGHEs-ScHRADER, S., 1947, The "pre-metaphase"
stretch and kinetochore orientation in phasmids, Chromosoma, 3, 1.
INov~, S., 1952, The effect of colchicine on the microscopic and submicroscopic structure of the mitotic spindle. Exp. Cell Research, Suppl., 2, 305.
INOU~, S., 1953, Polarization optical studies of the mitotic spindle. The demonstration of spindle fibers in living cells, Chromosoma, 5, 487.
INOU~, S., 1959, Motility of cilia and the mechanism of mitosis, Rev. Mod. Physics, 31,402.
INotr~, S., 1961, Polarizing microscope, in The Encyclopedia of Microscopy, (G. I. Clark, editor), New York, Reinhold Publishing Corp., 480-485.
INOU~, S., 1964, Organization and function of the mitotic spindle, in Primitive Motile Systems in Cell Biology, (R. Allen and N. Kamiya, editors), New York, Academic Press, Inc., 549-598.
INOU~, S., and DAN, K., 1951, Birefringence of the dividing cell, J. Morphol., 89, 423.
INolJ~, S., and HYDE, W, L., 1957, Studies on de- polarization of light at microscope lens surfaces. II. The simultaneous realization of high resolution and high sensitivity with the polarizing microscope, J. Biophysic. and Biochem. Cytol., 3, 831.
Izua~p, K., 1961, Effects of ultraviolet microbeam irradiation upon division in grasshopper spermato- cytes. II. Results of irradiation during metaphase and anaphase I, Mie Med. J., 11, 213.
JAO~ER, J., 1961, A small and inexpensive ultraviolet dose-rate meter useful in biological experiments, Radiation Research, 14, 394.
LACouR, L. F., and PELt, S. R., 1959, Effect of colchicine on the utilization of thymidine labelled with tritium during chromosomal reproduction, Nature, 183, 1455.
LAUOHLIN, R., 1958, The rearing of crane flies (Ti- pulidae), Entomol. Exp. et Appl., l , 241.
LECOMa~, J. , 1949, Action de la colchicine et des poisons radiomim&iques sur le muscle de gre- nouille, Arch. Internat. Pharmacodyn., 78, 440.
MAZIA, D., 1961, Mitosis and the physiology of cell division, in The Cell, (J. Brachet and A. E. Mirsky, editors), New York, Academic Press, Inc., 3, 77-412.
NmKLAS, R. B., 1963, A quantitative study of chro- mosomal elasticity and its influence on chromo- some movement, Chromosoma, 14, 276.
NORRXS, K. P., SEEDS, W. E., and WmmNS, M. H. F., 1951, Reflecting microscopes with spherical mir- rors, J. Opt. Soc. Am., 41, 111.
(~STERGREN, G., MOL~-BAJER, J., and BAJER, A., 1960, An interpretation of transport phenomena at mitosis, Ann. New York Acad. Sc., 90, 381.
OTROSHCHENKO, V. A., and SAICHAROV, V. N., 1964, Local irradiation of the cytoplasm of the cell by an ultraviolet microbeam at various phases of mitosis, Dokl. Akad. Nauk. SSSR, 154, 1441.
PEASE, D. C., 1946, Hydrostatic pressure effects upon
ARTHUR FORER Spindle Fiber Birefringence 113

the spindle figure and chromosome movement. II. Experiments on the meiotic divisions of Trad- escantia pollen mother cells, Biol. Bull., 91, 145.
Rm, Iq., t949, The anaphase movement of chromo- somes in the spermatocytes of the grasshopper, Biol. Bull., 96, 90.
ROBBmS, E., and GONATAS, N. K., 1964, Histochem- ical and ultrastructural studies on HeLa cells ex- posed to spindle inhibitors with special reference to the interphase cell, J. Histochem. and Cytochem., 12, 704.
ROTH, L. E., 1964, Motile systems with continuous filaments, in Primitive Motile Systems in Cell Biology, (R. Allen and N. Kamiya, editors), New York, Academic Press, Inc., 527-546.
SCHRADER, F., 1953, Mitosis: The Movements of Chromosomes in Cell Division, New York, Co- lumbia University Press.
SHURCLIFI', W. A., 1962, Polarized Light, Cambridge, Massachusetts, Harvard University Press.
SMITH, C. L., 1964, Microbeam and partial cell irradiation, Internat. Rev. Cytol., 16, 133.
SRmAMULA, V., 1963, Effect of colchicine on the somatic chromosomes of Oryzias melastigrna McClel- land, La Cellule, 63, 369.
STroll, H. F., 1963, An experimental analysis of the courtship pattern of Tipula oleracea (Diptera), Canad. J. Zool., 41, 99.
SWANN, M. M., and MITCmSON, J. M., 1950, Refine-
A P P E N D I X
Focusing the Microbeam
The visible light image of the ultraviolet source is used to focus the ultraviolet microbeam into the specimen plane (see Zirkle, 1957). When a reflect- ing lens is used for this focusing, the ultraviolet and visible images should be focused to the same posi- tion. But since a refracting material is present be- tween the lens and the focus point, namely the quartz coverslip, light of different wavelengths will be focused at different positions because of the refractive index dispersion of this material. Thus, when the visible image of the ultraviolet source is in focus, other wavelengths are not in focus (Norris et al., 1951; Uretz and Perry, 1957). This focus shift is illustrated in Fig. 18 A and the calculated focus shift for various wavelengths with respect to the mercury green line (546 m/~) is shown in Fig. I8 B for different thickness coverslips. (The calcu- lation uses Snell's law, the refractive indices of fused quartz as given in the Chemical Rubber Publishing Company's Handbook of Chemistry and Physics, and the application of simple trigo-
ments in polarized light microscopy, J. Exp. Biol., 27, 226.
URETZ, R. B., 1962, cited in Nature, 196, 728. URETZ, R. B., and PERRY, R. P., t957, hnproved
ultraviolet microbeam apparatus, Rev. Scient. Instr., 28, 861.
URETZ, R. B., and ZmKLE, R. E., 1955, Disappear- ance of spindles in sand-dollar blastomeres after ultraviolet irradiation of cytoplasm, Biol. Bull., 109, 370.
WANG, D. Y., GREENBAUra, A. L., and HARKNESS, R. D., 1963, The action of colchicine on the acid soluble ribonucleotides of normal and regenerating rat liver, Biochem. J., 86, 62.
WiLsot% E. B., 1928, The Cell in Development and Heredity, New York, The Macmillan Co.
ZAMENHOF, S., 1943, Methods and instruments for the selective irradiation of chromatin, Rev. Scient. Instr. 14, 17.
ZIMMERMAN, A. M., and MARSLAND, D., 1964, Cell division: effects of pressure on the mitotic mecha- nisms of marine eggs (Arbacia punctulata), Exp. Cell Research, 35, 293.
ZmKLE, R. E., 1957, Partial-cell irradiation, Adv. Biol. and Med. Physics, 5, 103.
ZIRKL~, R. E., URETZ, R. B., and HAYNES, R. H., 1960, Disappearance of spindles and phragmo- plasts after microbeam irradiation of cytoplasm, Ann. New York Acad. Sc., 90, 435.
nometry.) The problem in focusing an ultraviolet microbeam, then, is to correct for this focus dis- placement between the visible and the ultraviolet images of the source.
Usual methods of refocusing require (a) the ultraviolet aperture on the optic axis, and (b) mo- tion of this aperture (or the reflecting lens) along the optic axis for (c) a measurable distance (e.g., see Uretz and Perry, 1957). This situation is illus- trated in Fig. 19 A: Ah is the displacement along the optic axis (oa) between the visible (v) and the ultraviolet (uv) images of the source (UV), which is caused by the quartz coverslip (qz); M is the lateral magnification and M 2 the longitudinal magnification of the lens which is represented in the thin lens approximation (Hall, 1953, chapter 3; Hardy and Perrin, 1932) as a straight line, L, with focal points at F. When the ultraviolet source aperture is moved a distance M~Ah along the axis away from the lens to a new position (UV'), the ultraviolet image (uv 1) for this new position is brought into focus in the specimen plane (sp) and the corresponding visible image (v') is in focus
114 TIlE JOURNAL OF CELL BEOLOGY • VOLUME ~5, 1965

A h (/z) 8 -
/ k - - T =0.170
7 -- ( ~ ) - - T - 0 . 1 7 5
• . . . . . . . T = O , 1 8 0
6 --
5
3
i
\ \ ~ " L~h -
I I I 3 0 0 4 0 0
B A
Z/t ,-o2, f J
ZXh
S n e l l ' s Law
N Sin e - constant
Sin e =Nx2 Sin e z, -NxI Sin e I,
t a n 8 2, - t a n O t, ] r 7o7 0
500 6 0 0 700
k (m F )
FIGURE 18
between the lens and the specimen plane. Re- focusing is accomplished by moving the aperture by M2Ah in a direction along the optic axis, or, equivalently, by moving the lens by a distance Ah. In Fig. 19 A the ultraviolet aperture is drawn very large, such that the correction which focuses the ultraviolet image in the specimen plane introduces a lateral shift in the image position for those points of the source which are off the optic axis (cf. uv and ug in Fig. 19 A). Since the ultraviolet apertures used as sources are less than a millimeter, the lateral shift is no problem, if the apertures are placed on the optic axis of the system.
An optical method for refocusing the micro- beam was devised which eliminates the need for accurate mechanical movement of the lens or aperture, and eliminates the need for the ultra- violet source to be on the optic axis. In this method, a fiat cell is inserted between the ultraviolet source aperture and the reflecting lens while the aperture
is focused onto the specimen with visible light; the cell is removed after focusing, prior to irradiation with ultraviolet. The effect of inserting and remov- ing the flat is illustrated in Fig. 19 B. Interposing the fiat makes the actual source aperture (UV ~) appear to come from a position, (UV), M2Ah closer to the lens. There is no lateral shift (i.e., in a direction perpendicular to the optic axis) be- tween either the two visible images (v, v ~) of the source aperture or the two ultraviolet images (uv, uv ~) of the source aperture when the surfaces of the flat are perpendicular to the line between the ultraviolet source aperture (UW) and the focal point F, even when the source aperture is not on the optic axis (Fig. 19 B). The flat position is adjusted such that by inserting and removing the flat the two visible images of the aperture occur at the same position on an ocular grid, the visible images being focused upon by changing the fine adjustment of the microscope. (This adjustment
ARTHUR FORER Spindle Fiber Birefringence 115

QZ
Sp
X
v 013 ]
i V - '" 4 - - - " |
t.
d 4 4 - -
UV UV
$p
OCI'
UV UV i
QZ
r
B
/ "~ A UV UV e
M ~ ~ h
FmURE 19
relies on the mechanical stability of the micro- scope fine adjustment, which, in our experience, is quite reliable, and much better than that of the substage lens-focusing system.) The fiat was a machined cylinder of Plexiglass (Commercial Plastics, New York) to which was cemented two microscope slides, and into which was added aqueous solutions of hexylene glycol. The exact concentration varied depending on the optical path desired. The optical path of such a flat is:
(N -q (N,_q ah '=\~-g I T s + \ N, ] T,
where T is thickness, and N is refractive index. Ngl .... Tgl .... and T~olutio., were measured, and
N~olution was adjusted (by changing the ratios of hexylene glycol and water) such that:
Ah' = M~Ah
where M is the lateral magnification of the reflect- ing lens, and Ah is the focal shift due to the quartz coverslip as shown in Fig. 18 B. M was measured for the aperture position in question, and Ah was chosen for the wavelength desired, usually 275 m/~.
The advantages of the refocusing system which used the ([at are (a) it can be used with common laboratory microscopes, since it does not rely on well-machined aperture movement or lens move- ment devices; (b) it can be used with a min imum
116 THE JOURNAL OF CELL BIOLOGY " VOLUME ~5, 1965

of expensive equipment, and is as reliable as the expensive equipment; (c) it does not require that the ultraviolet aperture be placed on the optic axis; and (d) it permits very quick change of the focus correction, for no aperture position changes need be made but only a new solution, or a new cell, be inserted between the aperture and the re-
flecting lens. Brumberg (1943) and Norris et al. (1951) had previously corrected for the dispersion of the coverslip by using a substage plano-convex lens made from the same material as the coverslip. The optical refocusing method described here is similar in principle to their correction, but is much simpler in fabrication.
ARTHUR FORER Spindle Fiber Birefringenee 117