Embed Size (px)
Citation preview

Long-Term Deficits on a Foraging Task After BilateralVestibular Deafferentation in Rats
Yiwen Zheng,* Matthew Goddard, Cynthia L. Darlington, and Paul F. Smith
ABSTRACT: Animal studies have shown that bilateral vestibular deaf-ferentation (BVD) causes deficits in spatial memory that may be relatedto electrophysiological and neurochemical changes in the hippocampus.Recently, human studies have also indicated that human patients can ex-hibit spatial memory impairment and hippocampal atrophy even 8–10yr following BVD. Our previous studies have shown that rats withunilateral vestibular deafferentation (UVD) showed an impairment at 3months after the surgery on a food foraging task that relies on hippo-campal integration of egocentric cues, such as vestibular information;however, by 6 months postop, they showed a recovery of function. Bycontrast, the long-term effects of BVD on spatial navigation have neverbeen well studied. In this study, we tested BVD or sham rats on a foodforaging task at 5 months postop. Under light conditions, BVD ratswere able to use visual cues to guide themselves home, but did so witha significantly longer homing time. However, in darkness, BVD ratswere severely impaired in the foraging task, as indicated by a signifi-cantly longer homing distance and homing time, with more errors andlarger heading angles when compared with sham rats. These results sug-gest that, unlike UVD, BVD causes long-term deficits in spatial navi-gation that are unlikely to recover, even with repeated T-maze training.VVC 2008 Wiley-Liss, Inc.
KEY WORDS: vestibular system; hippocampus; food carrying; homingaccuracy; egocentric cues
INTRODUCTION
Traditionally, the vestibular inner ear has been considered importantmainly for the initiation of ocular motor and postural reflexes. However,in recent years, its contribution to cognitive function has been increas-ingly recognized. Activation (Matthews et al., 1989; Semenov and Bures,1989) or inactivation of the vestibular system (Peruch et al., 1999;Stackman and Herbert, 2002; Wallace et al., 2002; Russell et al., 2003a;Schautzer et al., 2003) has been demonstrated to disrupt spatial learningand memory in radial arm maze and foraging tasks in rats and guineapigs. In humans, lesions to the vestibular system have been reported toresult in deficits in attention and spatial memory (Grimm et al., 1989;Peruch et al., 1999; Redfern et al., 2004; Brandt et al., 2005).
The cognitive effects of vestibular damage in ani-mals and humans are likely to be at least partlybecause of their effects on the hippocampus, since ves-tibular nucleus neurons relay vestibular informationpolysynaptically to the hippocampus (Vitte et al.,1996; Cuthbert et al., 2000; de Waele et al., 2001;Hicks et al., 2004; Horii et al., 2004), and bilateralinactivation of the vestibular system has been shownto disrupt the response of place cells (Stackman et al.,2002; Russell et al., 2003b) and theta activity in CA1(Russell et al., 2006).
There seems, however, to be a substantial differencein the impact of bilateral and unilateral peripheral ves-tibular lesions on the hippocampus. Although bilateralvestibular deafferentation (BVD) was found to resultin spatial memory deficits and bilateral hippocampalatrophy (of �17%) in human patients (Brandt et al.,2005), unilateral vestibular deafferentation (UVD)produced neither of these effects (Hufner et al.,2007). It appears that the transmission of vestibularinformation from the vestibular nucleus to the hippo-campus must be bilateral, since in vitro brain slicestudies following UVD have demonstrated a bilateraldecrease in electrical excitability in CA1 (Zheng et al.,2003) and unilateral stimulation of the vestibular lab-yrinth or the vestibular nucleus increases hippocampalcell firing bilaterally (Cuthbert et al., 2000; Horiiet al., 2004). Nonetheless, it may be the case that acomplete bilateral loss of vestibular information isnecessary in order for hippocampal function andstructure in vivo, to be altered in a major way.
Although the effects of bilateral inactivation of thevestibular system on spatial memory have been investi-gated using injections of sodium arsinilate and tetrodo-toxin into the middle ear of rats (Ossenkopp andHargreaves, 1993; Stackman et al., 2002; Wallaceet al., 2002), few studies have been conducted on theeffects of permanent bilateral surgical lesions of the ves-tibular system, as used in human surgery. Russell et al.(2003a) used surgical BVD in rats and found impairedacquisition of a radial arm maze task up to 6 weekspostop. Zheng et al. (2006) found that rats with UVDwere impaired in a foraging task in darkness at 3months but recovered by 6 months postop; on theother hand, BVD rats were reticent to engage in thetask even at 6 months postop. Nonetheless, this obser-vation was based on a small sample only (n 5 4) andtherefore there has been a need for a larger and more
Department of Pharmacology and Toxicology, School of Medical Scien-ces, University of Otago Medical School, Dunedin, New ZealandGrant sponsor: Health Research Council of New Zealand; Grant number:07/047; Grant sponsor: Neurological Foundation of New Zealand; Grantnumber: 0014/PG.*Correspondence to: Yiwen Zheng, Department of Pharmacology andToxicology, School of Medical Sciences, University of Otago MedicalSchool, P.O. Box 913, Dunedin, New Zealand.E-mail: [email protected] for publication 7 October 2008DOI 10.1002/hipo.20533Published online 12 December 2008 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 19:480–486 (2009)
VVC 2008 WILEY-LISS, INC.

systematic analysis of the behavior of BVD rats in spatial mem-ory tasks such as the foraging task. In this study, we investigatedthe effects of BVD on spatial memory in a larger sample of ratsat 5 months postop, a time when the animals have recoveredfrom the acute postural reflex deficits caused by BVD and arehyperactive (Goddard et al., 2008). We also determined whethertraining in a spatial forced alternation task in a T-maze couldimprove BVD animals’ performance in the foraging task.
MATERIALS AND METHODS
Animals and Surgery
Data were obtained from 35 male Wistar rats, randomlyallocated to the BVD (n 5 18) and sham surgery (n 5 17)conditions and tested at 5 months postop. As part of a largerseries of experiments (e.g., Zheng et al., 2007), half of theBVD and sham animals were given training on a spatial forcedalternation task in a T maze at 3 weeks, 3 months, and 5months postop, in order to determine whether this wouldimprove their spatial working memory, and the other half wereexposed simultaneously to a T maze of the same dimensionswithout spatial alternation training. At each of the three timepoints, rats received eight trials per day for a total of 20 days[the details of the spatial alternation procedure are provided inZheng et al. (2007)]. The groups are referred to here as theBVD and sham (i.e., no training, n 5 9 and n 5 8, respec-tively) and BVD-T and sham-T (i.e., training, n 5 9 and n 5
9, respectively) groups. Animals were maintained on a 12:12 hlight:dark cycle at 228C and housed two per cage.
The BVD surgery was performed under microscopic controlas described in detail previously (Zheng et al., 2006, 2007).Briefly, animals were anesthetized with 300 lg/kg fentanyl citrate(i.p.) and 300 lg/kg medetomidine hydrochloride (i.p.). Thetympanic bulla was exposed using a retroauricular approach, andthe tympanic membrane, malleus, and incus were removed. Thestapedial artery was cauterized, and the horizontal and anteriorsemicircular canal ampullae drilled open. The contents of thecanal ampullae and the utricle and saccule were aspirated andthe temporal bone sealed with dental cement. Carprofen (5 mg/kg, s.c.) was used for postoperative analgesia. Sham surgery con-sisted of exposing the temporal bone and removing the tympanicmembrane without producing a vestibular lesion. Using tempo-ral bone histology, our previous studies have shown that theBVD surgical procedure produces a complete and permanentlesion of the vestibular labyrinth with no damage beyond thetemporal bone (Zheng et al., 2006). All experimental procedureswere approved by the University of Otago Committee on Ethicsin the Care and Use of Laboratory Animals.
Food Deprivation
The animals were food deprived during the course of behav-ioral testing, and their body weight was maintained at 85% oftheir normal feeding weight. Large oat and wheat honey cereal
loops were used during behavioral testing. After testing eachday, the rats were supplementally fed with normal laboratoryrodent pellets in their home cage to maintain body weight.
Apparatus
The apparatus was similar to that used by other researchersand in our previous study (Whishaw and Tomie, 1997; Maas-winkel et al., 1999; Wallace et al., 2002; Zheng et al., 2006)with slight modifications. It consisted of a large circular table(140 cm in diameter) with eight holes (10 cm in diameter)equally distributed around the edge and centered 10.5 cm fromit. In our previous study, it was noticed that BVD rats oftenaccidentally fell through the holes and off the edge of the table,which prevented them from foraging. Therefore, in the presentstudy, a metal sheet was inserted at the bottom of each hole(except the home base) to prevent falls, so that the eight holesstill looked the same to the rats on the table. A 10-cm high,clear transparent perspex edging was also fitted to the edge ofthe table to prevent falls from the edge. The table was locatedin a room (4.9 3 3.5 m2) with plenty of visual cues on thewall and mounted on a center bearing 105 cm above the floorso that the table could be rotated between animals. Twenty-three food cups (4 cm in diameter, 1 cm in height) wereattached to the table. A cage that served as the rat’s home basecould be placed beneath one of the eight holes. An opaque cur-tain was hung from the ceiling so that the table could beenclosed for dark conditions. For light conditions, the roomwas illuminated by two fluorescent lights located on the ceiling;while for dark conditions, the fluorescent lights were turned offand an infrared light source was placed inside the curtain. Aninfrared camera was located above the center of the table sothat the behavior of the animals could be video recorded underboth light and dark conditions. A speaker continually playingwhite noise was also mounted above the center of the table.
Behavioral Training and Testing
Pretraining
For the pretraining, the rats were required to retrieve fourpellets per session over a maximum period of 10 min for 10consecutive days. All the food cups were baited for the first fewdays, and the number of cups baited was gradually reducedover time. By the end of the pretraining, only one cup wasbaited per trial. After the rat found the food and carried itback home, a new cup was baited while the rat was eating. Thefood was baited in different cups for different trials, but thesame cup was baited for all of the animals on each trial forthe purpose of comparison. The home cage was located underthe same hole in relation to the room throughout the pretrain-ing phase. The table was cleaned and randomly rotated 1358clockwise or anticlockwise between the animals. The room lightswere on and the curtain surrounding the table was drawn back.
Light probe test
After the rats had learned to successfully retrieve four foodpellets each day, they were given one probe trial with the home
SPATIAL NAVIGATION IMPAIRED AFTER LABYRINTHECTOMY 481
Hippocampus
cage located at a novel position under the light condition, withthe room setting exactly the same as in the pretraining stage. Ifa rat could not return back home on the probe trial, it wasremoved from the table after 120 s had elapsed.
Training in the dark
Following the probe trial in light, the animals were givenone day (four trials) of normal training, with the home cagelocated at the same place as on the pretraining trials. A further14 days of training was subsequently carried out in the darkwith the curtain surrounding the table. The training in thedark consisted of one trial per day, with the home cage locatedat a different position each day. The table was cleaned androtated between the animals.
Analysis and Measurements
Animal path traces were analyzed offline using VCR play-back displayed on a PC and custom-coded path-tracing soft-ware. Searching distance (mm), searching time (sec), homingdistance (mm), homing time (sec), the number of choicesmade before reaching the home, the first home choice, the sec-ond home choice, and the initial heading angle (8) wereobtained from each animal’s trials. The sampling rate of thehardware was 100 ms, and the frame-rate for manual path trac-ing was 10 frames/s. The initial heading angle was obtainedusing an onscreen protractor (Iconico Screen Protractor 4.0)overlaid on the path traces. Zero degrees was defined as themost direct angle possible to the correct home.
Foraging and homing distance (mm) and time (sec) and thenumber of choices required to reach home were analyzed usingtwo- or three -way repeated measures ANOVAs in Minitab.Anderson-Darling and Levene’s tests were used to check thatthe assumptions of normality and equality of variances (respec-tively) were upheld. Where assumptions were violated, a trans-formation (e.g., log) was applied and the data retested for nor-mality and equal variance (Rice, 2007). In cases where normal-ity could not be achieved, a nonparametric Kruskal-WallisANOVA was used (Rice, 2007). The analysis of the initialheading angles was conducted using circular statistics and theOriana 2 statistics package (Rice, 2007). A significant Rayleigh’stest (P < 0.05) indicated that the data were not uniform andwere concentrated around a mean vector. To test whether themean vector was in the direction of the home location, speci-fied as a mean angle of 08, a variant of the Rayleigh’s test, theV-test, was used with a significance level of 0.05. Significant V-test statistics indicated that the initial heading angle was in thedirection of the home location (Rice, 2007).
RESULTS
In no case did T-maze training have any significant effect onany of the variables studied (data not shown); therefore, thedata within the sham and BVD groups were pooled.
Pretraining
While the sham rats quickly learned to come out of thehome base searching for food and carry the food back home toeat, the BVD rats were hesitant to leave the home base initially.Once they came out, they either ate the food on the table orfailed to carry the food home. However, by session 8, both thesham and BVD rats were able to reach the criterion, i.e., threeor more food retrievals per session. When homing time overthe last five pretraining sessions was compared, there was asignificant treatment effect (F(1,132) 5 23.64, P 5 0.0001) anda significant subject effect (F(33,132) 5 41.68, P < 0.0001).There was neither a session effect (F(4,132) 5 0.99, P 5 0.42)nor treatment 3 session 3 subject interaction (F(4,132) 5 0.47,P 5 0.76) (Fig. 1).
Light Probe Test
During the light probe trial when the home base was movedto a novel location, it took both the sham and BVD animalssignificantly longer to find the home base compared with thepretraining days (data not shown). Most of the animals alsomade their first home choice the old home location (Fig. 2).Circular statistical analysis on the first home choice revealed asignificant preference for the old home location for both sham(V test: V 5 0.579, U 5 3.374, P < 0.001) and BVD animals(V test: V 5 0.295, U 5 1.769, P 5 0.038). However, theirsecond home choice was randomly distributed (Rayleigh test: R5 0.43, P 5 0.07 for sham and R 5 0.43, P 5 0.09 forBVD). There was also no difference between sham and BVDanimals in the number of errors they made before reaching thecorrect home and the number of visits to the old home (datanot shown).
Training in the Dark
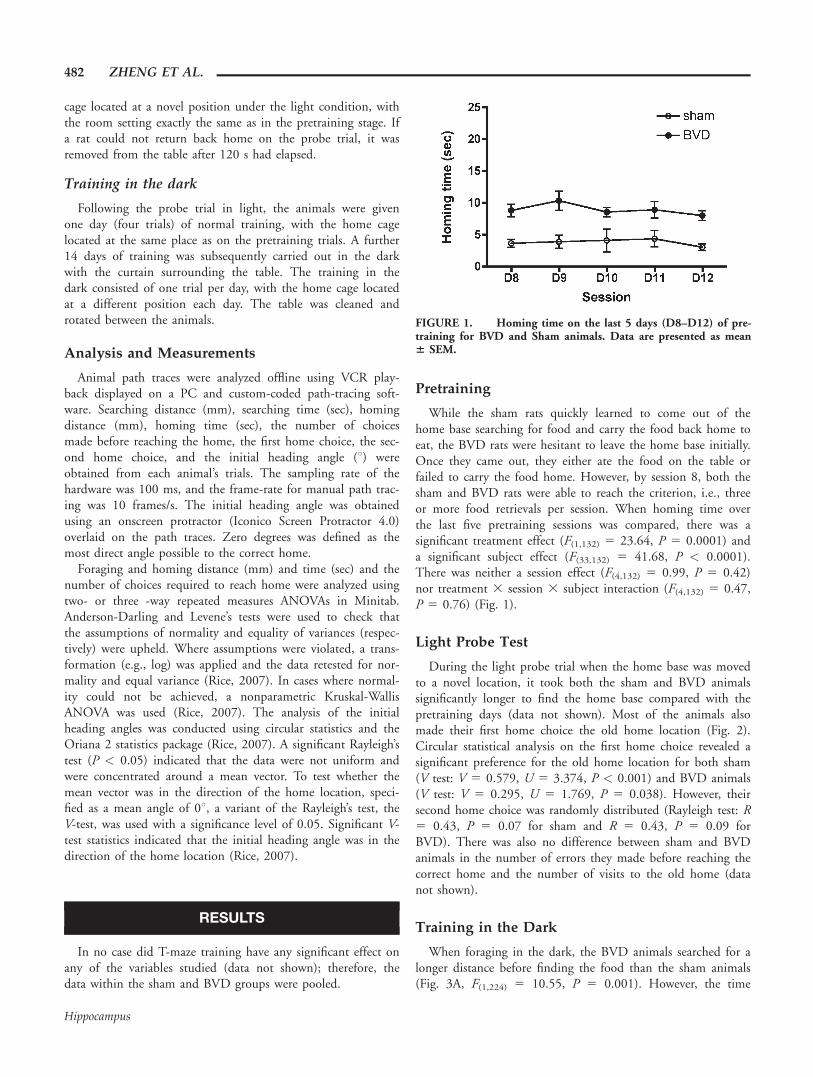
When foraging in the dark, the BVD animals searched for alonger distance before finding the food than the sham animals(Fig. 3A, F(1,224) 5 10.55, P 5 0.001). However, the time
FIGURE 1. Homing time on the last 5 days (D8–D12) of pre-training for BVD and Sham animals. Data are presented as mean6 SEM.
482 ZHENG ET AL.
Hippocampus

spent searching for food was not significantly different betweenthe BVD and sham groups (Fig. 3B, F(1,224) 5 0.02, P 5
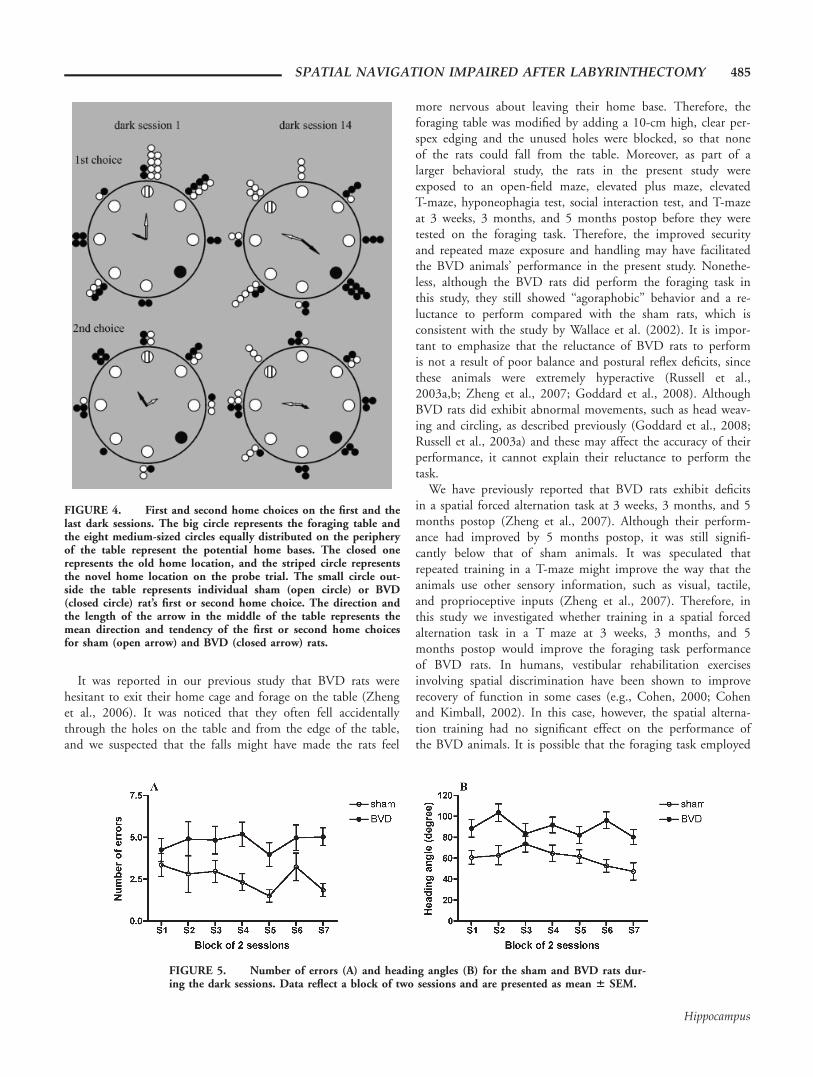
0.884). Upon finding the food, sham animals took a moredirect route home, as evidenced by a significantly shorter hom-ing distance (Fig. 3C, F(1,224) 5 53.47, P < 0.0001) and fasterhoming time (Fig. 3D, F(1,224) 5 20.74, P < 0.0001) whencompared with BVD animals. The sham animals’ first homechoices were clustered around the correct home base on everytraining session in the dark. Figure 4 shows examples of firsthome choices on the first and the last training session (V test:session 1, V 5 0.611, U 5 3.562, P < 0.0001; session 14, V5 0.473, U 5 2.755, P 5 0.002). However, the BVD rats’first home choices over the 14 sessions in darkness showed no
preference for the correct home base (Fig. 4, session 1, V 5
0.114, U 5 0.684, P 5 0.249; session 14, V 5 20.56, U 5
23.362, P 5 1). Once the animals made the incorrect firsthome choice, their second home choices were randomly distrib-uted for both sham and BVD animals on the first dark trainingsession (Raleigh test: sham, R 5 0.32, P 5 0.41; BVD, R 5
0.43, P 5 0.06). However, on the last session, sham animals,but not the BVD animals, showed a significant preferencetoward the correct home base on their second home choices(V test: V 5 0.377, U 5 1.849, P 5 0.032).
When the number of incorrect holes visited before reachingthe correct home base (number of errors) was analyzed, BVDanimals made significantly more errors than the sham animals(Fig. 5A, H(1) 5 45.06, P < 0.0001). The animals’ perform-ance was further analyzed by comparing the initial headingangle on the homeward path. As shown in Figure 5B, whilesham animals showed a nonsignificant trend toward improve-ment in the heading angle over the dark training sessions, theBVD animals’ heading angles were consistently larger than thatof sham animals throughout the sessions (F(1,224) 5 41.75, P< 0.0001). Circular statistics further revealed that sham ani-mals’ heading angles showed a significant preference toward thecorrect home location throughout the dark sessions (e.g., Vtest: session 1, V 5 0.445, U 5 2.597, P 5 0.004), whileBVD animals’ heading angles showed no preference for the cor-rect home location (e.g., V test: session 1, V 5 0.029, U 5
0.175, P 5 1).
DISCUSSION
The results of the present study demonstrate that, at 5months postop, a permanent surgical BVD does not affect arat’s ability to find the correct home in a spatial foraging taskwhen visual cues are available, but results in increases in hom-ing time. However, in darkness, BVD causes a significantimpairment in spatial memory, at a time following the surgerywhen rats with UVD exhibit some recovery of function (Zhenget al., 2006).
The food foraging task is designed to test the animal’s abilityto use different strategies to return home by manipulating theavailability of different cues and has been used to investigatethe contribution of the hippocampus and vestibular informa-tion to spatial navigation (Whishaw and Maaswinkel, 1998;Maaswinkel et al., 1999; Whishaw et al., 2001a,b; Wallaceet al., 2002; Zheng et al., 2006). In the present study, BVDrats were able to carry the food back home under the lightcondition after a few days of training, but they did so with asignificantly longer homing time than the sham rats. However,when they emerged from a novel location under the light con-dition, their first home choice showed a significant preferencefor the old home location, which suggests that BVD rats wereable to use distal visual cues. It has been reported that BVDrats were hyperkinetic with significantly greater movement ve-locity and time spent in movement than sham rats (Goddard
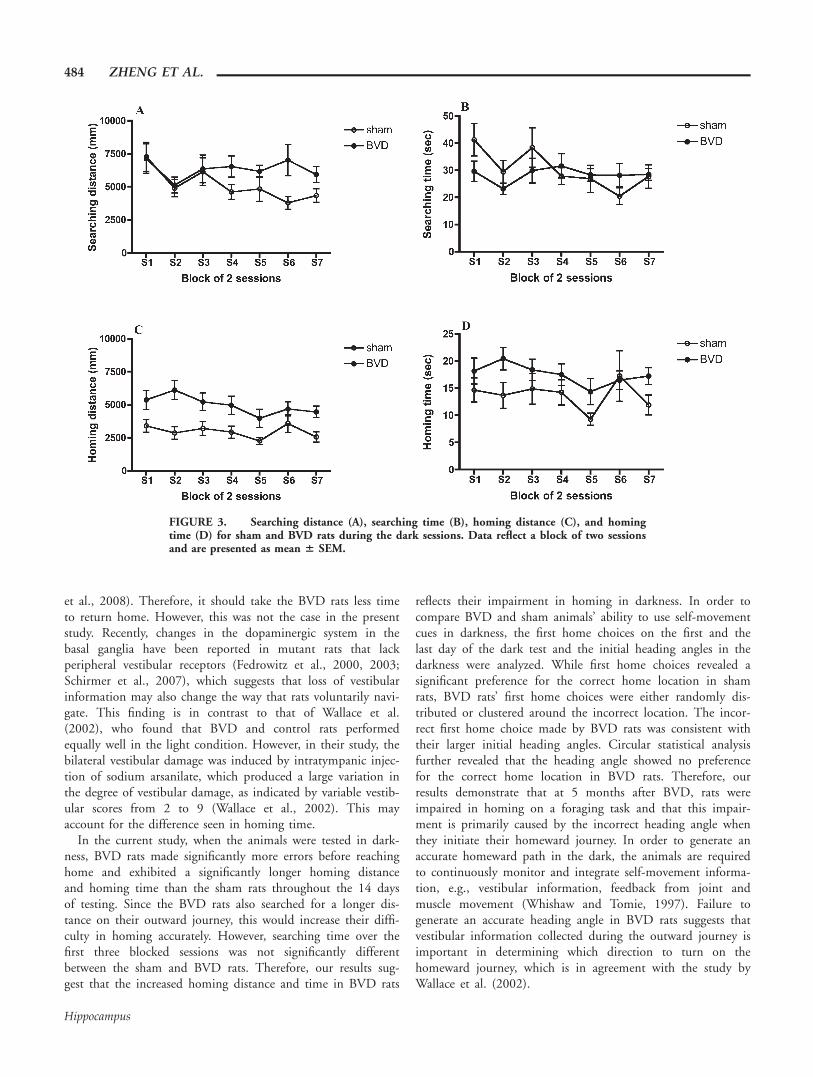
FIGURE 2. First and second home choices on the light probetrial. The big circle represents the foraging table and the eight me-dium-sized circles equally distributed on the periphery of the tablerepresent the potential home bases. The closed one represents theold home location, and the striped circle represents the novelhome location on the probe trial. The small circle outside the tablerepresents individual sham (open circle) or BVD (closed circle)rat’s first or second home choice. The direction and the length ofthe arrow in the middle of the table represents the mean directionand tendency of the first or second home choices for sham (openarrow) and BVD (closed arrow) rats.
SPATIAL NAVIGATION IMPAIRED AFTER LABYRINTHECTOMY 483
Hippocampus
et al., 2008). Therefore, it should take the BVD rats less timeto return home. However, this was not the case in the presentstudy. Recently, changes in the dopaminergic system in thebasal ganglia have been reported in mutant rats that lackperipheral vestibular receptors (Fedrowitz et al., 2000, 2003;Schirmer et al., 2007), which suggests that loss of vestibularinformation may also change the way that rats voluntarily navi-gate. This finding is in contrast to that of Wallace et al.(2002), who found that BVD and control rats performedequally well in the light condition. However, in their study, thebilateral vestibular damage was induced by intratympanic injec-tion of sodium arsanilate, which produced a large variation inthe degree of vestibular damage, as indicated by variable vestib-ular scores from 2 to 9 (Wallace et al., 2002). This mayaccount for the difference seen in homing time.
In the current study, when the animals were tested in dark-ness, BVD rats made significantly more errors before reachinghome and exhibited a significantly longer homing distanceand homing time than the sham rats throughout the 14 daysof testing. Since the BVD rats also searched for a longer dis-tance on their outward journey, this would increase their diffi-culty in homing accurately. However, searching time over thefirst three blocked sessions was not significantly differentbetween the sham and BVD rats. Therefore, our results sug-gest that the increased homing distance and time in BVD rats
reflects their impairment in homing in darkness. In order tocompare BVD and sham animals’ ability to use self-movementcues in darkness, the first home choices on the first and thelast day of the dark test and the initial heading angles in thedarkness were analyzed. While first home choices revealed asignificant preference for the correct home location in shamrats, BVD rats’ first home choices were either randomly dis-tributed or clustered around the incorrect location. The incor-rect first home choice made by BVD rats was consistent withtheir larger initial heading angles. Circular statistical analysisfurther revealed that the heading angle showed no preferencefor the correct home location in BVD rats. Therefore, ourresults demonstrate that at 5 months after BVD, rats wereimpaired in homing on a foraging task and that this impair-ment is primarily caused by the incorrect heading angle whenthey initiate their homeward journey. In order to generate anaccurate homeward path in the dark, the animals are requiredto continuously monitor and integrate self-movement informa-tion, e.g., vestibular information, feedback from joint andmuscle movement (Whishaw and Tomie, 1997). Failure togenerate an accurate heading angle in BVD rats suggests thatvestibular information collected during the outward journey isimportant in determining which direction to turn on thehomeward journey, which is in agreement with the study byWallace et al. (2002).
FIGURE 3. Searching distance (A), searching time (B), homing distance (C), and homingtime (D) for sham and BVD rats during the dark sessions. Data reflect a block of two sessionsand are presented as mean 6 SEM.
484 ZHENG ET AL.
Hippocampus
It was reported in our previous study that BVD rats werehesitant to exit their home cage and forage on the table (Zhenget al., 2006). It was noticed that they often fell accidentallythrough the holes on the table and from the edge of the table,and we suspected that the falls might have made the rats feel
more nervous about leaving their home base. Therefore, theforaging table was modified by adding a 10-cm high, clear per-spex edging and the unused holes were blocked, so that noneof the rats could fall from the table. Moreover, as part of alarger behavioral study, the rats in the present study wereexposed to an open-field maze, elevated plus maze, elevatedT-maze, hyponeophagia test, social interaction test, and T-mazeat 3 weeks, 3 months, and 5 months postop before they weretested on the foraging task. Therefore, the improved securityand repeated maze exposure and handling may have facilitatedthe BVD animals’ performance in the present study. Nonethe-less, although the BVD rats did perform the foraging task inthis study, they still showed ‘‘agoraphobic’’ behavior and a re-luctance to perform compared with the sham rats, which isconsistent with the study by Wallace et al. (2002). It is impor-tant to emphasize that the reluctance of BVD rats to performis not a result of poor balance and postural reflex deficits, sincethese animals were extremely hyperactive (Russell et al.,2003a,b; Zheng et al., 2007; Goddard et al., 2008). AlthoughBVD rats did exhibit abnormal movements, such as head weav-ing and circling, as described previously (Goddard et al., 2008;Russell et al., 2003a) and these may affect the accuracy of theirperformance, it cannot explain their reluctance to perform thetask.
We have previously reported that BVD rats exhibit deficitsin a spatial forced alternation task at 3 weeks, 3 months, and 5months postop (Zheng et al., 2007). Although their perform-ance had improved by 5 months postop, it was still signifi-cantly below that of sham animals. It was speculated thatrepeated training in a T-maze might improve the way that theanimals use other sensory information, such as visual, tactile,and proprioceptive inputs (Zheng et al., 2007). Therefore, inthis study we investigated whether training in a spatial forcedalternation task in a T maze at 3 weeks, 3 months, and 5months postop would improve the foraging task performanceof BVD rats. In humans, vestibular rehabilitation exercisesinvolving spatial discrimination have been shown to improverecovery of function in some cases (e.g., Cohen, 2000; Cohenand Kimball, 2002). In this case, however, the spatial alterna-tion training had no significant effect on the performance ofthe BVD animals. It is possible that the foraging task employed
FIGURE 4. First and second home choices on the first and thelast dark sessions. The big circle represents the foraging table andthe eight medium-sized circles equally distributed on the peripheryof the table represent the potential home bases. The closed onerepresents the old home location, and the striped circle representsthe novel home location on the probe trial. The small circle out-side the table represents individual sham (open circle) or BVD(closed circle) rat’s first or second home choice. The direction andthe length of the arrow in the middle of the table represents themean direction and tendency of the first or second home choicesfor sham (open arrow) and BVD (closed arrow) rats.
FIGURE 5. Number of errors (A) and heading angles (B) for the sham and BVD rats dur-ing the dark sessions. Data reflect a block of two sessions and are presented as mean 6 SEM.
SPATIAL NAVIGATION IMPAIRED AFTER LABYRINTHECTOMY 485
Hippocampus

here required continuously monitoring orientation in order togenerate the most direct homing path, which is different fromalternating in a T-maze.
The fact that BVD animals demonstrated such a degree ofimpairment in a foraging task in dark even at 5 monthspostop, raises the question of whether their performance defi-cits are permanent. It is impossible to answer this questionwithout longer term studies. However, spatial memory impair-ment has been demonstrated in humans even 8–10 years fol-lowing BVD, suggesting that at least some of the deficits maybe permanent (Brandt et al., 2005).
REFERENCES
Brandt T, Schautzer F, Hamilton D, Bruning R, Markowitsch HJ,Kalla R, Darlington CL, Smith PF, Strupp M. 2005. Vestibularloss causes hippocampal atrophy and impaired spatial memory inhumans. Brain 128:2732–2741.
Cohen HS. 2000. Vestibular disorders and impaired path integrationalong a linear trajectory. J Vestib Res 10(1):7–15.
Cohen HS, Kimball KT. 2002. Improvements in path integration aftervestibular rehabilitation. J Vestib Res 12:47–51.
Cuthbert PC, Gilchrist DPD, Hicks SL, MacDougall HG, CurthoysIS. 2000. Electrophysiological evidence for vestibular activation ofthe guinea pig hippocampus. NeuroReport 11:1443–1447.
de Waele C, Baudonniere P, Lepecq J, Tran Ba Huy P, Vidal P. 2001.Vestibular projections in the human cortex. Exp Brain Res141:541–551.
Fedrowitz M, Potschka H, Richter A, Loscher W. 2000. A microdialy-sis study of striatal dopamine release in the circling rat, a geneticmodel with spontaneous rotational behavior. Neuroscience 97:69–77.
Fedrowitz M, Lindemann S, Loscher W, Gernert M. 2003. Alteredspontaneous discharge rate and pattern of basal ganglia output neu-rons in the circling (ci2) rat mutant. Neuroscience 118:867–878.
Goddard M, Zheng Y, Darlington CL, Smith PF. 2008. Locomotorand exploratory behavior in the rat following bilateral vestibulardeafferentation. Behav Neurosci 122:448–459.
Grimm RJ, Hemenway WG, Lebray PR, Black FO. 1989. The peril-ymph fistula syndrome defined in mild head trauma. Acta Otolar-yngol (Stockh) 464:1–40.
Hicks SL, Gilchrist DPD, Cuthbert PC, Curthoys IS. 2004. Hippo-campal field responses to direct electrical stimulation of the vestib-ular system in awake or anesthetised guinea pigs. Proc Aust Neuro-sci Soci 15:87.
Horii A, Russell NA, Smith PF, Darlington CL, Bilkey DK. 2004.Vestibular influences on CA1 neurons in the rat hippocampus: Anelectrophysiological study in vivo. Exp Brain Res 155:245–250.
Hufner K, Hamilton DA, Kalla R, Stephan T, Glasauer S, Ma J,Bruning R, Markowitsch HJ, Labudda K, Schichor C, Strupp M,Brandt T. 2007. Spatial memory and hippocampal volume inhumans with unilateral vestibular deafferentation. Hippocampus17:471–485.
Maaswinkel H, Jarrard LE, Whishaw IQ. 1999. Hippocampectomizedrats are impaired in homing by path integration. Hippocampus9:553–561.
Matthews BL, Ryu JH, Bockaneck C. 1989. Vestibular contribution tospatial orientation. Evidence of vestibular navigation in an animalmodel. Acta Otolaryngol (Stockh) 468:149–154.
Ossenkopp KP, Hargreaves EL. 1993. Spatial learning in an enclosedeight-arm maze in rats with sodium arsinilate-induced labyrinthec-tomies. Behav Neural Biol 59:253–257.
Peruch P, Borel L, Gaunet F, Thinus-Blanc G, Magnan J, Lacour M.1999. Spatial performance of unilateral vestibular defective patientsin nonvisual versus visual navigation. J Vestib Res 9:37–47.
Redfern M, Talkowski J, Jennings J, Furman J. 2004. Cognitive influ-ences in postural control of patients with unilateral vestibular loss.Gait Posture 19:105–114.
Rice J. 2007. Mathematical Statistics and Data Analysis. Duxbury Press,Belmont, California, MA.
Russell NA, Horii A, Smith PF, Darlington CL, Bilkey DK. 2003a.Bilateral peripheral vestibular lesions produce long-term changes inspatial learning in the rat. J Vestib Res 13(1):9–16.
Russell NA, Horii A, Smith PF, Darlington CL, Bilkey DK. 2003b.Long-term effects of permanent vestibular lesions on hippocampalspatial firing. J Neurosci 23:6490–6498.
Russell NA, Horii A, Smith PF, Darlington CL, Bilkey DK. 2006.Lesions of the vestibular system disrupt hippocampal theta rhythmin the rat. J Neurophysiol 96:4–14.
Schautzer F, Hamilton D, Kalla R, Strupp M, Brandt T. 2003. Spatialmemory deficits in patients with chronic bilateral vestibular failure.Ann NY Acad Sci 1004:316–324.
Schirmer M, Lessenich A, Lindemann S, Loscher W. 2007. Markeddifferences in response to dopamine receptor antagonism in two ratmutants, ci2 and ci3, with lateralized rotational behavior. BehavBrain Res 180:218–225.
Semenov LV, Bures J. 1989. Vestibular stimulation disrupts acquisitionof place navigation in the Morris water tank task. Behav NeuralBiol 51:346–363.
Stackman RW, Herbert AM. 2002. Rats with lesions of the vestibularsystem require a visual landmark for spatial navigation. BehavBrain Res 128:27–40.
Stackman RW, Clark AS, Taube JS. 2002. Hippocampal spatial repre-sentations require vestibular input. Hippocampus 12:291–303.
Vitte E, Derosier C, Caritu Y, Berthoz A, Hasboun D, Soulie D.1996. Activation of the hippocampal formation by vestibular stim-ulation: a functional magnetic resonance imaging study. Exp BrainRes 112:523–526.
Wallace DG, Hines DJ, Pellis SM, Whishaw IQ. 2002. Vestibularinformation is required for dead reckoning in the rat. J Neurosci22:10009–10017.
Whishaw IQ, Maaswinkel H. 1998. Rats with fimbria-fornix lesionsare impaired in path integration: a role for the hippocampus in‘‘sense of direction’’. J Neurosci 18:3050–3058.
Whishaw IQ, Tomie J. 1997. Piloting and dead reckoning dissociatedby fimbria-fornix lesions in a rat food carrying task. Behav BrainRes 89:87–97.
Whishaw IQ, Hines DJ, Wallace DG. 2001a. Dead reckoning (pathintegration) requires the hippocampal formation: Evidence fromspontaneous exploration and spatial learning tasks in light (allo-thetic) and dark (idiothetic) tests. Behav Brain Res 127:49–69.
Whishaw IQ, Maaswinkel H, Gonzalez CL, Kolb B. 2001b. Deficitsin allothetic and idiothetic spatial behavior in rats with posteriorcingulate cortex lesions. Behav Brain Res 118:67–76.
Zheng Y, Kerr DS, Darlington CL, Smith PF. 2003. Unilateral innerear damage results in lasting changes in hippocampal CA1 fieldpotentials in vitro. Hippocampus 13:873–878.
Zheng Y, Darlington CL, Smith PF. 2006. Impairment and recoveryon a food foraging task following unilateral vestibular deafferenta-tion in rats. Hippocampus 16:368–378.
Zheng Y, Goddard M, Darlington CL, Smith PF. 2007. Bilateral ves-tibular deafferentation impairs performance in a spatial forcedalternation task in rats. Hippocampus 17:253–256.
486 ZHENG ET AL.
Hippocampus