Embed Size (px)
Citation preview

1
L’ossigeno, la sua storia e i suoi legami con la vita sulla Terra Matteo Cortesi – 845852 Tutti noi abbiamo bisogno di precise condizioni per poter sopravvivere. La presenza di ossigeno è una di queste e ci sembra scontato che ce ne sia in abbondanza nell’aria che respiriamo e che non venga mai a mancare. Tuttavia questa molecola non è sempre stata così diffusa nell’atmosfera come ai giorni nostri. Qui di seguito verranno riassunte le attuali conoscenze sull’andamento dei livelli di ossigeno nel tempo e verranno esaminate le relazioni tra l’ossigeno e la storia dei viventi.
Introduzione
Gli organismi viventi, per poter svolgere le
normali funzioni vitali, necessitano di energia.
Essa è ottenibile attraverso una gran varietà di vie
metaboliche, durante le quali il consumo di una
molecola presente nell’ambiente che funga da
accettore di elettroni è indispensabile. I batteri
denitrificanti, per esempio, si servono dello ione
nitrato (NO3-) riducendolo ad azoto molecolare
(N2). Altri organismi, invece, i cosiddetti batteri
metanogeni, sono in grado di ridurre anidride
carbonica (CO2) a metano (CH4). Analogamente,
altri composti o ioni sono impiegati dagli altri
esseri viventi allo stesso scopo.
Tra tutti, è però l’ossigeno molecolare (O2)
l’accettore di elettroni che permette il massimo
guadagno energetico a parità di tutti gli altri
fattori, nonché quello utilizzato dalla maggior
parte degli organismi attualmente viventi.
L’importanza dell’ossigeno come accettore di
elettroni è sottolineata dalla netta distinzione che
fanno gli ecologi definendo aerobi gli organismi il
cui metabolismo è basato sul consumo di questa
molecola e anaerobi tutti gli altri. I taxa di
Eucarioti attualmente viventi sono quasi tutti
aerobi, così come la maggior parte dei Procarioti.
Questo non sorprende alla luce di due
considerazioni. La prima, precedentemente
enunciata, è costituita dall’efficienza assoluta
dell’O2 in termini di guadagno energetico in
seguito al suo impiego nel metabolismo. La
seconda risiede nell’abbondanza di questa
molecola sul nostro pianeta.
L’ossigeno molecolare, essendo gassoso, si trova
prevalentemente in atmosfera e di essa ne
occupa più di un quinto del volume (21%),
secondo solo all’azoto molecolare (78%). Inoltre,
la sua solubilità in acqua fa sì che l’ossigeno si
trovi anche in idrosfera come gas disciolto, con
concentrazioni massime nelle acque superficiali e
fredde. Nel Mar Glaciale Artico la concentrazione
media annuale in superficie può arrivare a circa
9,5mL/L (World Ocean Atlas 2013). Ambienti
anossici, completamente privi di ossigeno,
costituiscono l’eccezione sulla Terra. Si trovano
ad esempio in ridotti bacini eutrofici, oppure
nelle profondità oceaniche qualora si verifichino
determinate condizioni, tra le quali un’assenza di

2
mescolamento con le acque ossigenate
superficiali e un’accentuata decomposizione nei
fondali (Diaz 2001).
L’ossigeno è quindi presente in modo diffuso sulla
Terra. Ma nella storia del pianeta non è sempre
stato così.
L’era pre-GOE
Nella prima metà circa della sua storia
l’atmosfera terrestre era dominata da anidride
carbonica, azoto molecolare e metano -
quest’ultimo accumulatosi massicciamente in
seguito all’evoluzione dei batteri metanogeni,
avvenuta nel primo Archeano (Kasting 2014). I
livelli di ossigeno rimasero al di sotto dello
0,001% rispetto al livello attuale o PAL (present
atmospheric level; Kump 2008, Lyons et al. 2014)
fino a subire una rapida crescita tra 2,5 e 2,3
miliardi di anni fa (Bya) nota come Great
Oxydation Event o GOE (Fig.1). Le evidenze di
questo evento sono parecchie e ormai tutti gli
scienziati sembrano riconoscerlo (fa eccezione:
Ohmoto 1996). Tra di essi vi è invece discordanza
sul momento in cui comparve la fotosintesi
ossigenica, oggi operata dalle piante e dai
cianobatteri per convertire l’energia solare in
energia chimica. Esplorare la questione è
importante in quanto la produzione di O2
mediante meccanismi diversi da questo - come la
fotolisi del vapore acqueo innescata dai raggi UV
(Buick 2008) - è quantitativamente irrilevante e
dunque la fotosintesi ossigenica può essere
considerata l’unica fonte significativa di O2 sulla
Terra (Buick 2008, Lyons et al. 2014).
La spiegazione che appare più intuitiva è che la
fotosintesi ossigenica si sia evoluta in
corrispondenza - e sia stata quindi una diretta
causa - del GOE (Kopp et al. 2005). Tuttavia molti
indizi contrastano con questa visione e fanno sì
che l’ipotesi più supportata sia quella di una
fotosintesi apparsa molto tempo prima. Secondo
questa ipotesi l’ossigeno non si sarebbe potuto
accumulare in atmosfera prima del GOE per il suo
continuo sequestro messo in atto dall’ossidazione
dei composti riducenti che fino a quel momento
dominavano il pianeta (Catling & Claire 2005,
Kump & Barley 2007, Gaillard et al. 2011).
Le stime sulla comparsa della fotosintesi
ossigenica spaziano tra i 3,8 e i 2,35 Bya e la
difficoltà sta soprattutto nello stabilire se i
fenomeni osservabili nel record geologico
richiedano indiscutibilmente la presenza o
l’assenza di O2. Con ogni probabilità, i primi
organismi fotosintetici erano unicellulari e simili
agli attuali cianobatteri. Ma i microfossili risalenti
all’Archeano (4,0-2,5 Bya) hanno una forma
troppo semplice, che rende difficile determinarne
il tipo di metabolismo (Lyons et al. 2014).
Similmente, al giorno d’oggi le stromatoliti
archeane non sono imputabili in modo certo a
una genesi biogenica, perciò non costituiscono
una prova inconfutabile della presenza di
fotosintesi ossigenica (Bosak et al. 2013).
Analisi su scisti archeani e neogenico-quaternari
(23-0 Mya) mostrano dati sul contenuto totale di
carbonio organico (TOC, total organic carbon)
pressoché identici nei due periodi, suggerendo
l’esistenza anche prima del GOE di un
metabolismo particolarmente efficiente, come un

3
qualche tipo di fotosintesi. Varie considerazioni
sfavoriscono l’utilizzo di donatori di elettroni
come H2S, Fe2+ o H2 (rispettivamente: acido
solfidrico, ferro bivalente e idrogeno molecolare)
legati a fotosintesi diverse: la fotosintesi
ossigenica, che invece ricorre all’acqua (H2O), è
quella più probabile per giustificare i dati sul TOC
(Lyons et al. 2014).
Entrando più nel dettaglio, altri studi hanno
ricercato informazioni dai biomarcatori, fossili
molecolari derivanti da composti organici primari
che a volte sono legati a uno specifico produttore
biologico. L’individuazione di sterani eucarioti-
dipendenti in scisti australiani di Pilbara Craton
(Brocks et al. 1999) fornirebbe non solo una
prova dell’esistenza di cellule eucariotiche circa
2,7 Bya (ben 800-900 milioni di anni prima dei più
antichi fossili eucariotici; Xiao et al. 2013) ma
anche un sostegno a favore della presenza di O2,
in quanto richiesto, sebbene a bassissime
concentrazioni, per la sintesi di tali idrocarburi
(Lyons et al. 2014). Anche risultati più recenti
concordano con quelli di Brocks et al. (Waldbauer
et al. 2011). Tuttavia l’integrità dei dati ricavati
dai biomarcatori è molto dibattuta (Kopp et al.
2005, Xiao 2013, Lyons et al. 2014) e vari altri tipi
di analisi (si veda ad esempio: Parfrey et al. 2011)
fanno risalire la comparsa degli Eucarioti a GOE
ormai avvenuto, permettendo di dubitare dei dati
di Brocks et al. (1999) e del relativo sostegno a
favore di una fotosintesi ossigenica pre-GOE. Il
campo di battaglia sui biomarcatori rimane
comunque più che aperto.
Alcuni metalli (RSM, redox-sensitive metals)
variano la propria reattività e solubilità in
relazione alla valenza. Molibdeno (Mo) e renio
(Rh) sono ricorrenti nei minerali come i solfuri e
quando si ossidano in ambiente subaereo
vengono rilasciati nei fiumi e raggiungono gli
oceani, dove poi si possono depositare (Lyons et
al. 2014). Arricchimenti di tali metalli in sedimenti
marini depositati anteriormente al GOE sono stati
messi in luce da alcune indagini (Anbar et al.
2007, Duan et al. 2010). I ricercatori hanno
fornito varie possibili spiegazioni ma quella più
parsimoniosa riguarda proprio l’esistenza di O2 in
atmosfera, in grado di provocare l’alterazione
ossidativa di questi elementi persino a
concentrazioni molto basse (Duan et al. 2010,
Lyons et al. 2014), compatibili con le
quantificazioni riportate in precedenza (< 0,001%
PAL).
Rocce meta-sedimentarie della Isua Greenstone
Belt (Groenlandia) sono state sottoposte ad
analisi isotopiche sul carbonio (C) e sul sistema
uranio-torio-piombo (U-Th-Pb) (Rosing & Frei
2004). In particolare, è stato calcolato un elevato
rapporto U/Th. U è solubile solo in fluidi
ossidanti, mentre la mobilità di Th è indipendente
dallo stato ossidativo dell’ambiente; ciò rende
plausibile che almeno le acque superficiali
fossero relativamente ossidate per la presenza di
organismi fotosintetizzanti (Rosing & Frei 2004,
Buick 2008). Questa visione è naturalmente
contrastata da altre (Kopp et al. 2005), ma se è
corretta fornisce la stima più antica sulla
comparsa della fotosintesi ossigenica, datandola
a più di 3,7 Bya.
Tra le formulazioni di possibili scenari, alcune
ipotizzano temporanei “soffi di ossigeno” (whiffs
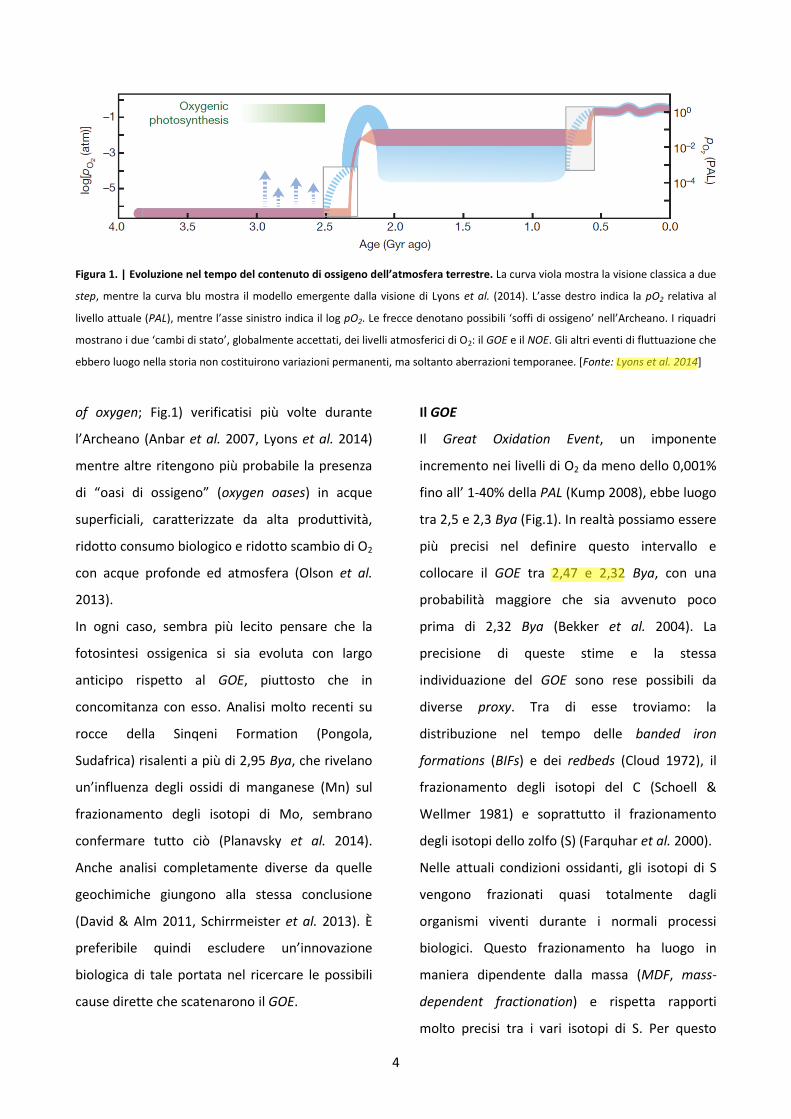
4
Figura 1. | Evoluzione nel tempo del contenuto di ossigeno dell’atmosfera terrestre. La curva viola mostra la visione classica a due
step, mentre la curva blu mostra il modello emergente dalla visione di Lyons et al. (2014). L’asse destro indica la pO2 relativa al
livello attuale (PAL), mentre l’asse sinistro indica il log pO2. Le frecce denotano possibili ‘soffi di ossigeno’ nell’Archeano. I riquadri
mostrano i due ‘cambi di stato’, globalmente accettati, dei livelli atmosferici di O2: il GOE e il NOE. Gli altri eventi di fluttuazione che
ebbero luogo nella storia non costituirono variazioni permanenti, ma soltanto aberrazioni temporanee. [Fonte: Lyons et al. 2014]
of oxygen; Fig.1) verificatisi più volte durante
l’Archeano (Anbar et al. 2007, Lyons et al. 2014)
mentre altre ritengono più probabile la presenza
di “oasi di ossigeno” (oxygen oases) in acque
superficiali, caratterizzate da alta produttività,
ridotto consumo biologico e ridotto scambio di O2
con acque profonde ed atmosfera (Olson et al.
2013).
In ogni caso, sembra più lecito pensare che la
fotosintesi ossigenica si sia evoluta con largo
anticipo rispetto al GOE, piuttosto che in
concomitanza con esso. Analisi molto recenti su
rocce della Sinqeni Formation (Pongola,
Sudafrica) risalenti a più di 2,95 Bya, che rivelano
un’influenza degli ossidi di manganese (Mn) sul
frazionamento degli isotopi di Mo, sembrano
confermare tutto ciò (Planavsky et al. 2014).
Anche analisi completamente diverse da quelle
geochimiche giungono alla stessa conclusione
(David & Alm 2011, Schirrmeister et al. 2013). È
preferibile quindi escludere un’innovazione
biologica di tale portata nel ricercare le possibili
cause dirette che scatenarono il GOE.
Il GOE
Il Great Oxidation Event, un imponente
incremento nei livelli di O2 da meno dello 0,001%
fino all’ 1-40% della PAL (Kump 2008), ebbe luogo
tra 2,5 e 2,3 Bya (Fig.1). In realtà possiamo essere
più precisi nel definire questo intervallo e
collocare il GOE tra 2,47 e 2,32 Bya, con una
probabilità maggiore che sia avvenuto poco
prima di 2,32 Bya (Bekker et al. 2004). La
precisione di queste stime e la stessa
individuazione del GOE sono rese possibili da
diverse proxy. Tra di esse troviamo: la
distribuzione nel tempo delle banded iron
formations (BIFs) e dei redbeds (Cloud 1972), il
frazionamento degli isotopi del C (Schoell &
Wellmer 1981) e soprattutto il frazionamento
degli isotopi dello zolfo (S) (Farquhar et al. 2000).
Nelle attuali condizioni ossidanti, gli isotopi di S
vengono frazionati quasi totalmente dagli
organismi viventi durante i normali processi
biologici. Questo frazionamento ha luogo in
maniera dipendente dalla massa (MDF, mass-
dependent fractionation) e rispetta rapporti
molto precisi tra i vari isotopi di S. Per questo

5
motivo la MDF è distinguibile dalla MIF (mass-
independent fractionation), che al contrario non
separa gli isotopi secondo regole fisse. La MIF si
attua principalmente per processi abiotici come
la fotodissociazione dell’anidride solforosa (SO2)
presente in atmosfera. La scissione di questa
molecola è praticata dai raggi UV, perciò può
verificarsi in modo tangibile solo in assenza di una
schermatura a questi raggi da parte di uno strato
di ozono (O3), a sua volta esistente in presenza di
O2 in atmosfera. In altre parole, in assenza di O2
la fotodissociazione della SO2 può avvenire ed
avviene in maniera preponderante, mentre in
presenza di O2 essa sarà irrisoria rispetto al
frazionamento biologico. Riscontrare MIF in rocce
sedimentarie deposte in un determinato periodo
indica dunque assenza di O2 in atmosfera in quel
periodo, mentre osservare MDF indica la sua
presenza. Proprio la scomparsa di MIF tra circa
2,4 e 2,3 Bya, fino a quel momento prevalente, e
il coincidente affermarsi di MDF fino ai giorni
nostri ci permettono di datare il GOE con relativa
precisione (Fig.2; Farquhar et al. 2000, Bekker et
al. 2004).
Una complicazione deriva da risultati molto
recenti (Reinhard, Planavsky & Lyons 2013). Essi
rendono verosimile una sopravvivenza di segnali
di MIF nel record sedimentario fino a 10-100
milioni di anni dopo la crescita della pO2. Gli
autori sostengono che i segnali di MIF in pirite e
altri solfuri generati in assenza di O2 atmosferico,
una volta che tali minerali furono riaffiorati,
alterati, dilavati e riseppelliti, si sarebbero
conservati a lungo nonostante i livelli atmosferici
di O2 fossero già sensibilmente alti; sarebbero
occorsi quindi ripetuti cicli di riaffioramento,
alterazione, diluizione e seppellimento sotto
un’atmosfera ossidativa per eliminare il segnale
di MIF dalle rocce sedimentarie (Reinhard
Planavsky & Lyons 2013, Lyons et al. 2014).
Questo cosiddetto ‘crustal memory effect’ non
solo complicherebbe la stima dei livelli di O2
archeani e la datazione di inizio del GOE (che
risulterebbe anticipata di qualche decina di
milioni di anni; Fig.1), ma renderebbe lecito
pensare al GOE come a un processo prolungato
piuttosto che a un evento discreto marcato dalla
perdita di MIF nel record geologico (Lyons et al.
2014). Visto in quest’ottica, il GOE fu un intervallo
di transizione durante il quale i livelli di ossigeno
salirono e scesero ripetutamente, riflettendo un
equilibrio dinamico tra la sua produzione e il suo
sequestro. Inizialmente la produzione prevalse a
intermittenza, poi permanentemente (Lyons et
al. 2014).
Nella ricerca del fattore che innescò il GOE, dopo
aver scartato la fotosintesi ossigenica ed essendo
essa l’unica fonte significativa di O2 (source), non
resterebbe che individuare quale fu il
meccanismo di perquisizione di O2 (sink) che
smise di funzionare o diminuì drammaticamente
la sua efficienza. Uno dei sink di O2 è dato dai
processi di respirazione anaerobica e di
decomposizione, che insieme all’ossigeno
consumano materia organica. L’efficienza di
questo sink potrebbe essere crollata in seguito a
un gran seppellimento di materia organica, che
sarebbe stata così sottratta dall’azione degli
eterotrofi e dei decompositori. Tuttavia i dati sul
δ13C non evidenziano nessuna coincidenza tra il
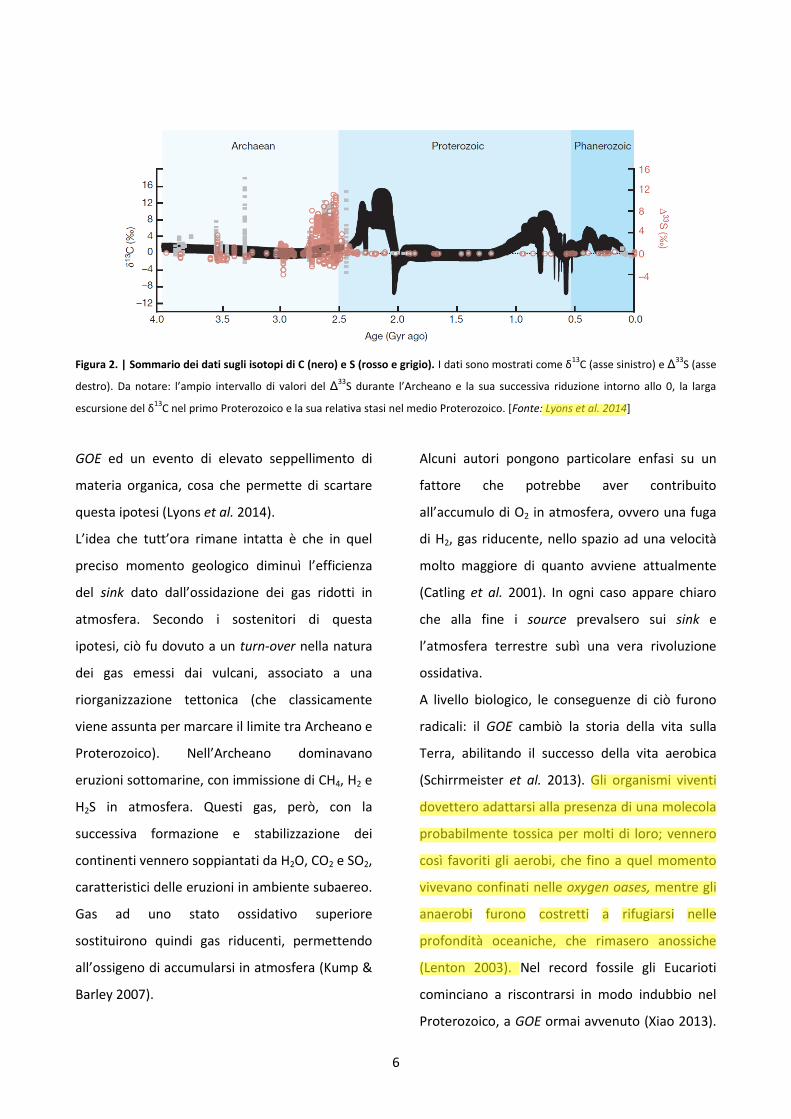
6
Figura 2. | Sommario dei dati sugli isotopi di C (nero) e S (rosso e grigio). I dati sono mostrati come δ13
C (asse sinistro) e Δ33
S (asse
destro). Da notare: l’ampio intervallo di valori del Δ33S durante l’Archeano e la sua successiva riduzione intorno allo 0, la larga
escursione del δ13
C nel primo Proterozoico e la sua relativa stasi nel medio Proterozoico. [Fonte: Lyons et al. 2014]
GOE ed un evento di elevato seppellimento di
materia organica, cosa che permette di scartare
questa ipotesi (Lyons et al. 2014).
L’idea che tutt’ora rimane intatta è che in quel
preciso momento geologico diminuì l’efficienza
del sink dato dall’ossidazione dei gas ridotti in
atmosfera. Secondo i sostenitori di questa
ipotesi, ciò fu dovuto a un turn-over nella natura
dei gas emessi dai vulcani, associato a una
riorganizzazione tettonica (che classicamente
viene assunta per marcare il limite tra Archeano e
Proterozoico). Nell’Archeano dominavano
eruzioni sottomarine, con immissione di CH4, H2 e
H2S in atmosfera. Questi gas, però, con la
successiva formazione e stabilizzazione dei
continenti vennero soppiantati da H2O, CO2 e SO2,
caratteristici delle eruzioni in ambiente subaereo.
Gas ad uno stato ossidativo superiore
sostituirono quindi gas riducenti, permettendo
all’ossigeno di accumularsi in atmosfera (Kump &
Barley 2007).
Alcuni autori pongono particolare enfasi su un
fattore che potrebbe aver contribuito
all’accumulo di O2 in atmosfera, ovvero una fuga
di H2, gas riducente, nello spazio ad una velocità
molto maggiore di quanto avviene attualmente
(Catling et al. 2001). In ogni caso appare chiaro
che alla fine i source prevalsero sui sink e
l’atmosfera terrestre subì una vera rivoluzione
ossidativa.
A livello biologico, le conseguenze di ciò furono
radicali: il GOE cambiò la storia della vita sulla
Terra, abilitando il successo della vita aerobica
(Schirrmeister et al. 2013). Gli organismi viventi
dovettero adattarsi alla presenza di una molecola
probabilmente tossica per molti di loro; vennero
così favoriti gli aerobi, che fino a quel momento
vivevano confinati nelle oxygen oases, mentre gli
anaerobi furono costretti a rifugiarsi nelle
profondità oceaniche, che rimasero anossiche
(Lenton 2003). Nel record fossile gli Eucarioti
cominciano a riscontrarsi in modo indubbio nel
Proterozoico, a GOE ormai avvenuto (Xiao 2013).

7
Non è escluso che essi siano comparsi in
precedenza, vivendo in ambienti anossici (con
metabolismi anaerobi) o nelle oasi di ossigeno
(Lenton 2003), ma sicuramente con la grande
ossidazione per gli Eucarioti ci fu la possibilità di
prosperare. La nascita del nostro regno, gli
Animali, dovette però attendere ancora a lungo,
tutta la cosiddetta età di mezzo della Terra, il
Proterozoico.
Il Proterozoico
Il Proterozoico (2,5-0,55 Bya) fu per la maggior
parte caratterizzato da una grande stabilità dei
livelli di O2. Probabilmente però, nella fase
immediatamente successiva al GOE, lo scenario
fu molto dinamico, con un ulteriore incremento
iniziale e un successivo sprofondamento dei livelli
di ossigeno (Fig.1; Lyons et al. 2014). Queste
affermazioni derivano in parte da analisi su RSM
negli oceani (Frei et al. 2009, Partin et al. 2013),
ma non solo.
Si può infatti constatare grazie a studi condotti su
rocce di varie località del mondo, che tra circa
2,22 e 2,06 Bya si verificò in assoluto il più grande
scostamento positivo del δ13C nella storia della
Terra (Fig.2; Karhu & Holland 1996, Martin et al.
2013), la cosiddetta Lomagundi excursion, dal
nome della località dello Zimbabwe dove fu per la
prima volta registrata (Martin et al. 2013).
Questo picco si può spiegare con un tasso di
seppellimento di carbonio organico
eccezionalmente alto, che avrebbe condotto
quindi a un incremento straordinario della
concentrazione atmosferica di O2 durante questo
periodo (Karhu & Holland 1996). Le stime
mettono in luce livelli di O2 addirittura al di sopra
dell’attuale inventario (Karhu & Holland 1996).
Da questa interpretazione della Lomagundi
excursion, che resta la più parsimoniosa (Lyons et
al. 2014), si deduce che il nuovo accumulo di
ossigeno fu dovuto a una perdita di efficienza del
sink dato dalla decomposizione e dalla
respirazione aerobica: in tempi relativamente
rapidi, molta materia organica fu sottratta
dall’azione di eterotrofi e decompositori, che
consumarono perciò una quantità di O2
nettamente inferiore.
Secondo un’altra ipotesi, invece, è più probabile
che l’incremento massiccio di O2 sia avvenuto a
causa di una produzione straordinaria da parte
dei cianobatteri attraverso la fotosintesi
ossigenica, resa possibile da una maggiore
disponibilità di fosfati, dovuta a sua volta a tassi
di alterazione accelerati, eventi tettonici, o
condizioni greenhouse post-glaciali (Papineau et
al. 2012) – nel Paleoproterozoico (2,5-1,6 Bya)
difatti si verificarono alcuni eventi di glaciazione
globale.
Molti aspetti di questa crescita e della successiva
caduta dei livelli di ossigeno rimangono pertanto
questioni aperte. Oltre che sulle cause,
permangono interrogativi sulle conseguenze
biologiche di queste fluttuazioni; finora nessun
evento significativo è stato riscontrato a riguardo
(Lyons et al. 2014). Indipendentemente dai
meccanismi che lo determinarono, comunque,
questo andamento, inserito nella curva storica
della pO2, contrasta con i modelli classici, che
solitamente invocano un aumento unidirezionale

8
dall’Archeano fino ai giorni nostri (Lyons et al.
2014), dando luogo a un modello più dinamico
(Fig.1).
A cavallo del Mesoproterozoico (1,6-1,0 Bya), per
circa un miliardo di anni, la concentrazione
atmosferica di ossigeno rimase a una quota
stabile e molto bassa, forse minore dello 0,1%
della PAL (Fig.1; Lyons et al. 2014). A suffragare
tale affermazione, analisi su Mn in paleosuoli e su
Cr (cromo) in sedimenti mesoproterozoici
indicano una scarsa ossidazione terrestre del Mn,
che normalmente si effettua perfino a
concentrazioni di O2 molto basse (< 0,001% PAL;
Zbinden et al. 1988, Frei et al. 2009).
Naturalmente non fu solo l’atmosfera ad essere
contraddistinta da queste condizioni, ma anche
l’idrosfera. Inizialmente vennero ipotizzate
condizioni di euxinia (mancanza di O2 accoppiata
a una sovrabbondanza di H2S) globalmente
diffuse negli oceani da 1,8 a 0,8 Bya circa
(Canfield 1998). Uno stato scarsamente
ossidativo e una ricchezza di H2S sono entrambi
fattori che agiscono negativamente sulla
presenza di Mo nelle acque (Lyons et al. 2014).
Ciò potrebbe aver avuto un ruolo chiave nel
determinare la scarsa efficienza fotosintetica dei
cianobatteri e la scarsa diversità eucariotica
proprie dell’età di mezzo della Terra. I
cianobatteri infatti necessitano di Mo per la
sintesi della nitrogenasi, l’enzima che permette
loro di fissare l’azoto atmosferico. Una carenza di
Mo nelle acque avrebbe dunque provocato per i
cianobatteri una diminuzione dell’efficienza
dell’azotofissazione, che si sarebbe ripercossa su
una scarsa efficienza del loro metabolismo in
toto, compresa la fotosintesi ossigenica. Livelli
bassi di O2 avrebbero poi sfavorito la
diversificazione degli Eucarioti, chiudendo il
circolo vizioso mesoproterozoico (Anbar & Knoll
2002).
Analisi sui livelli di Mo in questo periodo
mostrano valori molto bassi, eppure non
abbastanza da giustificare condizioni euxiniche
globali (Lyons et al. 2014). Nuovi modelli, infatti,
mettono in luce che in questo periodo il 30-40%
degli oceani era anossico e che solo una parte
minoritaria di questo (1-10%) era anche euxinica
(Reinhard et al. 2013). Le rimanenti porzioni
dell’oceano presentavano comunque livelli di O2
bassissimi, situazioni dette subossiche o
ipossiche. Ad ogni modo le condizioni non erano
favorevoli per un apporto di metalli bioessenziali
(come il Mo) sufficiente a soddisfare gli standard
ottimali richiesti dagli organismi viventi (Lyons et
al. 2014). Una serie di fattori tra loro concatenati
negarono quindi ai viventi la possibilità di
diversificarsi ed evolversi in modo palpabile nel
Proterozoico. Per quanto riguarda la pO2, si
dovette attendere fino alla fine dell’Eone per
assistere a un’ulteriore definitiva spinta, nota
come Neoproterozoic Oxidation Event o NOE.
Il NOE
Purtroppo, nonostante l’età relativamente
giovane, molto rimane sconosciuto sull’atmosfera
del Neoproterozoico (1,0-0,55 Bya) e sulle sue
relazioni con le forme di vita (Lyons et al. 2014).
Quel che si sa riguardo all’ossigeno è che in
questo periodo - più verosimilmente nella fase

9
finale - la sua concentrazione crebbe fino a
raggiungere un ordine di grandezza paragonabile
a quello odierno (Fig.1). A testimoniare il NOE
troviamo ancora una volta diverse proxy tra cui
analisi su C e S (Fike et al. 2006) e su RSM come
Mo, V (vanadio) e U (Scott et al. 2008, Sahoo et
al. 2012), effettuate su rocce di varie parti del
mondo.
Non è ben chiaro quanto questo trend fu
protratto nel tempo: a tal proposito restano
aperte possibilità che si sia manifestato con
continue crescite e decrescite durante tutto
l’Ediacarano (635-541 Mya) oppure che sia
cominciato prima dell’evento di Snowball Earth
Marinoano (650-635 Mya), e in questo caso abbia
contribuito a scatenare i cambiamenti climatici
che seguirono (Lyons et al. 2014).
Le ipotesi sulle cause che determinarono il NOE
sono numerose. L’innalzamento dei livelli di O2
potrebbe essere stato generato da un’alterazione
dei cicli dei nutrienti scatenata dalla glaciazione
marinoana, che avrebbe favorito una maggiore
produttività organica o un maggior seppellimento
organico (Sahoo et al. 2012). È altrettanto
probabile che tali cambiamenti furono prodotti
da rinnovamenti tettonici globali durante il
Neoproterozoico o da un tasso di diversificazione
algale superiore alla norma (Lyons et al. 2014).
Ulteriori ricerche sono quindi necessarie per fare
più chiarezza.
Con il NOE le profondità oceaniche vennero
ossigenate forse per la prima volta (Canfield
1998). È inoltre noto che la seconda grande
crescita della pO2 avvenne in concomitanza con
l’emergenza dei metazoi. Ciò viene
comunemente interpretato attribuendo alle
condizioni scarsamente ossidative del
Mesoproterozoico una inadeguatezza nel
supportare l’elevata domanda energetica di una
vita pluricellulare, condizioni che si sarebbero poi
verificate solo dopo il NOE (Lyons et al. 2014).
Pochi spiegano invece il ritardo della comparsa
degli animali come un vincolo dell’evoluzione
biologica (Butterfield 2009). Per questa ipotesi si
sarebbe trattato soltanto di una questione di
tempo: una complessità genica così elevata si
sarebbe potuta raggiungere solo dopo miliardi di
anni dalla nascita della vita.
Forse livelli così alti di ossigeno non furono
indispensabili per la nascita della pluricellularità,
o per sostenere il metabolismo dei primi animali
viventi (Mills et al. 2014), ma quasi
inevitabilmente sono richiesti per spiegare
l’ingente diversità animale, l’avvento della
biomineralizzazione, la straordinaria crescita di
dimensioni corporee, la complessità dei rapporti
prede-predatori e gli altri grandi passi storici
dell’evoluzione biologica (Lenton 2003, Lyons et
al. 2014).
Allo stesso modo, non si può negare che una così
grande innovazione nella storia della vita abbia
influenzato a sua volta caratteristiche ambientali
globali come i cicli dei nutrienti e la composizione
atmosferica, compresa quindi la quota di
ossigeno. Perciò, con la nascita degli animali, la
rete di feedback reciproci tra vita e ambiente si
infittì rispetto al passato: variazioni nell’uno
generalmente scatenano mutamenti nell’altro e
viceversa (Erwin et al. 2011, Lyons et al. 2014).

10
Il Fanerozoico
L’ultimo Eone, il Fanerozoico (541-0 Mya) è
caratterizzato da una relativa stabilità dei livelli di
ossigeno, che si mantennero sempre al di sopra
del 10% del volume atmosferico (Berner 2006).
Tuttavia, analisi dettagliate – principalmente su C
e S – permettono di osservare le fluttuazioni della
curva della pO2 occorse in questo periodo (Fig.3;
Berner 2006).
Figura 3. | Evoluzione nel tempo del contenuto atmosferico
di ossigeno durante il Fanerozoico. I valori osservabili nel
grafico sono quelli stimati da Berner (2006). In ascissa è
riportato il tempo in milioni di anni dal presente, in ordinata
la percentuale del volume di atmosfera occupata
dall’ossigeno molecolare. [Fonte: Berner 2006]
In particolar modo colpisce il picco a cavallo tra il
Carbonifero (359-299 Mya) e il Permiano (299-
251 Mya), durante il quale la concentrazione
atmosferica di O2 raggiunse il 30%. A causare
l’aberrazione fu probabilmente un elevato tasso
di seppellimento di materia organica dovuto
all’impiego da parte delle piante di un nuovo tipo
di sostanza resistente alla degradazione, la
lignina, che tra l’altro fu essenziale per
l’evoluzione di piante di grandi dimensioni
(Berner et al. 2007). Durante questo intervallo di
tempo, nel regno animale si verificò un generale
aumento di dimensioni corporee, che in alcuni
taxa di insetti condusse al gigantismo (Lenton
2003, Berner et al. 2007). Esso viene
generalmente attribuito a un miglioramento
dell’efficienza degli apparati respiratori degli
animali legato a un aumento della pressione di O2
(Berner et al. 2007) ma è anche possibile che tali
organismi possedessero speciali adattamenti per
concentrare l’ossigeno (Lenton 2003) o che
l’abbondanza di tale gas non fu il fattore
principale nel determinare il gigantismo
(Butterfield 2009).
Anche l’accrescimento post-cretacico (da 65 Mya)
dei mammiferi placentati è stato ascritto a un
corrispondente aumento dei livelli di ossigeno
(Falkowski et al. 2005) ma il meccanismo e le
eventuali prove di un diretto rapporto causa-
effetto rimangono ignoti (Berner et al. 2007).
L’origine del volo nei vertebrati ebbe luogo tra
fine Giurassico e inizio Cretaceo (tra 150 e 100
Mya circa; Lenton 2003). Sebbene in precedenza
quest’innovazione venne utilizzata come prova di
un nuovo picco positivo di pO2, non ci sono
evidenze geologiche che questo sia realmente
avvenuto (Lenton 2003). Al contrario, sembra che
al momento del primo involo dei vertebrati, i
livelli di O2 fossero relativamente bassi (Fig.3).
Perciò l’evoluzione del volo fu più realisticamente
dovuta a spinte evolutive, come un vantaggio
durante la fuga da un predatore (Lenton 2003).
Infine, tre fra i più incisivi episodi di estinzione di
massa della storia sembrano essere avvenuti in
fasi di declino della pO2: nel tardo evoniano

11
( 364 Mya), al limite Permo-Triassico ( 251 Mya) e
al limite Triassico- iurassico ( 201 Mya) (Berner
et al. 2007). Anche in questo caso troviamo le più
svariate interpretazioni. Alcuni tendono ad
assegnare alla caduta di ossigeno la
responsabilità principale delle estinzioni (Berner
et al. 2007). All’estremo opposto, altri
concludono che non fu questo fattore a guidare
le repentine perdite di biodiversità (Glasspool &
Scott 2010). Altri ancora invece pongono il
declino di ossigeno all’interno del complesso di
fattori che essi sostengono occorrano per
spiegare tali catastrofi (Shen et al. 2011).
La storia dell’ossigeno e quella vita sembrano
comunque in stretto contatto reciproco e
l’ossigeno resta tuttora un elemento
fondamentale per l’evoluzione della vita
complessa. Nuovi lavori aiuteranno ad aggiungere
un numero sempre maggiore di dettagli a questa
affascinante storia.
Bibliografia
Anbar A.D. & Knoll A.H. (2002) Proterozoic ocean chemistry and evolution: a bioinorganic bridge? Science; 297, 1137-1142. Anbar A.D., Duan Y., Lyons T.W., Arnold G.L., Kendall B., Creaser R.A., Kaufman A.J., Gordon G.W., Scott C., Garvin J., Buick R. (2007) A Whiff of Oxygen Before the Great Oxidation Event? Science; 317, 1903-1906. Bekker A., Holland H.D., Wang P.-L., Rumble D.III, Stein H.J., Hannah J.L., Coetzee L.L., Beukes N.J. (2004) Dating the rise of atmospheric oxygen. Nature; 427, 117-120. Berner R.A. (2006) GEOCARBSULF: A combined model for Phanerozoic atmospheric O2 and CO2. Geochimica et Cosmochimica Acta; 70, 5653-5664. Berner R.A., VandenBrooks J.M., Ward P.D. (2007) Oxygen and Evolution. Science; 316, 557-558.
Bosak T., Knoll A.H., Petroff A.P. (2013) The meaning of stromatolites. Annual Reviews of Earth and Planetary Sciences; 41, 21-44. Brocks J.J., Logan G.A., Buick R., Summons R.E. (1999) Archean molecular fossils and the early rise of eukaryotes. Science; 285, 1033-1036. Buick R. (2008) When did oxygenic photosynthesis evolve? Philosophical Transactions of the Royal Society – B; 363, 2731-2743. Butterfield N.J. (2009) Oxygen, animals and oceanic ventilation: an alternative view. Geobiology; 7, 1-7. Canfield D.E. (1998) A new model for Proterozoic ocean chemistry. Nature; 396, 450-453. Catling D.C., Zahnle K.J., McKay C.P. (2001) Biogenic methane, hydrogen escape, and the irreversible oxidation of early Earth. Science; 293, 839-843. Catling D.C. & Claire M.W. (2005) How Earth’s atmosphere evolved to an oxic state: a status report. Earth and Planetary Science Letters; 237, 1-20. Cloud P. (1972) A working model of the primitive Earth. American Journal of Science; 272, 537-548. David L.A. & Alm E.J. (2011) Rapid evolutionary innovation during an Archaean genetic expansion. Nature; 469, 93-96. Diaz R.J. (2001) Overview of Hypoxia around the World. Journal of Environmental Quality; 30, 275-281. Duan Y., Anbar A.D., Arnold G.L., Lyons T.W., Gordon G.W., Kendall B. (2010) Molybdenum isotope evidence for mild environmental oxygenation before the Great Oxidation Event. Geochimica et Cosmochimica Acta; 74, 6655-6668. Erwin D.H., Laflamme M., Tweedt S.M., Sperling E.A., Pisani D., Peterson K.J. (2011). The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science; 334, 1091-1097. Falkowski P., Katz M., Milligan A., Fennel K., Cramer B., Aubry M.P., Berner R.A., Zapol W.M. (2005) The rise of atmospheric oxygen levels over the past 205 million years and the evolution of large placental mammals. Science; 309, 2202-2204. Farquhar J., Bao H., Thiemens M. (2000) Atmospheric Influence of Earth’s Earliest Sulfur Cycle. Science; 289, 756-758. Fike D.A., Grotzinger J.P., Pratt L.M., Summons R.E. (2006) Oxidation of the Ediacaran ocean. Nature; 444, 744-747. Frei R., Gaucher C., Poulton S.W., Canfield D.E. (2009) Fluctuations in Precambrian atmospheric oxygenation recorded by chromium isotopes. Nature; 461, 250-253. Gaillard F., Scaillet B., Arndt N.T. (2011) Atmospheric oxygenation caused by a change in volcanic degassing pressure. Nature; 478, 229-232. Glasspool I.J. & Scott A.C. (2010) Phanerozoic concentrations of atmospheric oxygen reconstructed from sedimentary charcoal. Nature geoscience; 3, 627-630.

12
Karhu J.A. & Holland H.D. (1996) Carbon isotopes and the rise of atmospheric oxygen. Geology; 24, 867-870. Kasting J.F. (2014) Atmospheric composition of Hadean–early Archean Earth: The importance of CO. Geological Society of America Special Papers; 504, 19-28. Kopp R.E., Kirschvink J.L., Hilburn I.A., Nash, C.Z. (2005) The Paleoproterozoic snowball Earth: a climate disaster triggered by the evolution of oxygenic photosynthesis. Proceedings of the National Academy of Sciences of the USA; 102, 11131-11136. Kump L.R. & Barley M.E. (2007) Increased subaerial volcanism and the rise of atmospheric oxygen 2.5 billion years ago. Nature; 448, 1033-1036. Kump L.R. (2008) The rise of atmospheric oxygen. Nature; 451, 277-278. Lenton T.M. (2003) The coupled evolution of life and atmospheric oxygen. From: Academic Press; Evolution on Planet Earth. Impact of the Physical Environment (Rothschild L.J & Lister A.M.); chapter 3, 35-53. Lyons T.W., Reinhard C.T., Planavsky N.J. (2014) The rise of oxygen in Earth’s early ocean and atmosphere. Nature; 506, 307-315. Martin A.P., Condon D.J., Prave A.R., Melezhik V.A., Lepland A., Fallick A.E. (2013) Dating the termination of the Palaeoproterozoic Lomagundi-Jatuli carbon isotopic event in the North Transfennoscandian Greenstone Belt. Precambrian Research; 224, 160-168. Mills D.B., Ward L.M., Jones CA., Sweeten B., Forth M., Treusch A.H., Canfield D.E. (2014) Oxygen requirements of the earliest animals. Proceedings of the National Academy of Sciences of the USA; 111, 4168-4172. Ohmoto H. (1996) Evidence in pre-2.2 Ga paleosols for the early evolution of atmospheric oxygen and terrestrial biota. Geology; 24, 1135-1138. Olson S.L., Kump L.R., Kasting J.F. (2013) Quantifying the areal extent and dissolved oxygen concentrations of Archean oxygen oases. Chemical Geology; 362, 35-43. Papineau D., Purohit R., Fogel M.L., Shields-Zhou G.A. (2013) High phosphate availability as a possible cause for massive cyanobacterial production of oxygen in the Paleoproterozoic atmosphere. Earth and Planetary Science Letters; 362, 225-236. Parfrey L.W., Lahr D.J.G., Knoll A.H., Katz L.A. (2011) Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proceedings of the National Academy of Sciences of the USA; 108, 13624-13629. Partin C.A. Bekker A., Planavsky N.J., Scott C.T., Gill B.C., Li C., Podkovyrov V., Maslov A., Konhauser K.O., Lalonde S.V., Love G.D., Poulton S.W., Lyons T.W. (2013) Large-scale fluctuations in Precambrian atmospheric and oceanic oxygen levels from the record of U in shales. Earth and Planetary Science Letters; 369-370, 284-293. Planavsky N.J., Asael D., Hofmann A., Reinhard C.T., Lalonde S.V., Knudsen A., Wang X., Ossa Ossa F., Pecoits E., Smith A.J.B., Beukes N.J., Bekker A., Johnson T.M., Konhauser K.O., Lyons T.W., Rouxel O.J. (2014) Evidence for oxygenic
photosynthesis half a billion years before the Great Oxidation Event. Nature Geoscience; 7, 283-286. Reinhard C.T., Planavsky N.J., Robbins L.J., Partin C.A., Lalonde S.V., Bekker A., Konhauser K.O., Lyons T.W. (2013) Proterozoic ocean redox and biogeochemical stasis. Proceedings of the National Academy of Sciences of the USA; 110, 5357-5362. Reinhard C.T., Planavsky N.J., Lyons T.W. (2013) Long-term sedimentary recycling of rare sulphur isotope anomalies. Nature; 497, 100-103. Rosing M.T. & Frei R. (2004) U-rich Archaean sea-floor sediments from Greenland—indications of 3700 Ma oxygenic photosynthesis. Earth and Planetary Science Letters; 217, 237-244. Sahoo S.K., Planavsky N.J., Kendall B., Wang X., Shi X., Scott C., Anbar A.D., Lyons T.W., Jiang G. (2012) Ocean oxygenation in the wake of the Marinoan glaciation. Nature; 489, 546-549. Schirrmeister B.E., de Vos J.M., Antonelli A., Bagheri H.C. (2013) Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proceedings of the National Academy of Sciences of the USA; 110, 1791-1796. Schoell M. & Wellmer F.W. (1981) Anomalous 13C depletion in early Precambrian graphites from Superior Province, Canada. Nature; 290, 696-699. Scott C., Lyons T.W., Bekker A., Shen Y., Poulton S.W., Chu X., Anbar A.D. (2008) Tracing the stepwise oxygenation of the Proterozoic biosphere. Nature; 452, 456-459. Shen S.Z., Crowley J.L., Wang Y., Bowring S.A., Erwin D.H., Sadler P.M., Cao C.Q., Rothman D.H., Henderson C.M., Ramezani J., Zhang H., Shen Y.A., Wang X.D., Wang W., Mu L., Li W.Z., Tang Y.G., Liu X.L., Liu L.J., Zeng Y., Jiang Y.F., Jin Y. G. (2011) Calibrating the end-Permian mass extinction. Science; 334, 1367-1372. Waldbauer J.R., Newman D.K., Summons R.E. (2011) Microaerobic steroid biosynthesis and the molecular fossil record of Archean life. Proceedings of the National Academy of Sciences of the USA; 108, 13409-13414. Xiao S. (2013) Written in Stone: The Fossil Record of Early Eukaryotes. From: Springer; Evolution from the Galapagos; chapter 8, 107-124. Zbinden E.A., Holland H.D., Feakes C.R., Dobos S.K. (1988) The Sturgeon Falls paleosol and the composition of the atmosphere 1.1 Ga BP. Precambrian Research; 42, 141-163.
Sitografia World Ocean Atlas 2013 http://www.nodc.noaa.gov/cgi-bin/OC5/woa13f/woa13oxnuf.pl


























![INDEX [users.unimi.it]users.unimi.it/rdipp/indexes/2014.pdf · INDEX ARTICLES C. Campiglio, Italian Provisions on Assisted Procreation and International Pa- ... F. Salerno, Fundamental](https://img.pdfslide.net/doc/110x75/5bae888e09d3f2b5718cd94b/index-usersunimiitusersunimiitrdippindexes2014pdf-index-articles-c.jpg)


